 Xiao-Jie Yang1,2Hai-Long Zheng1,2Ying-Yang Liu1,2Hong-Wei Li1,2Yu-Hang Jiang1,2Lian-Bing Lin1,2*Xian-Yu Deng1,2*
Xiao-Jie Yang1,2Hai-Long Zheng1,2Ying-Yang Liu1,2Hong-Wei Li1,2Yu-Hang Jiang1,2Lian-Bing Lin1,2*Xian-Yu Deng1,2* Qi-Lin Zhang1,2*
Qi-Lin Zhang1,2*- 1Faculty of Life Science and Technology, Kunming University of Science and Technology, Kunming, China
- 2Engineering Research Center for Replacement Technology of Feed Antibiotics of Yunnan College, Kunming, China
Aquatic fireflies are important indicators of the quality of freshwater environments and key models for research on insect adaptation to freshwater environments. For these investigations, gene expression analyses using quantitative real-time PCR are heavily dependent on reliable reference genes. In this study, based on a transcriptome assembly and annotation for the aquatic firefly Aquatica leii at the adult and larval stages, 10 candidate reference genes (α-tubulin, β-tubulin, β-actin, EF1A, SDHA, UBQ, GST, GAPDH, RPS31, and RPL13A) were identified for analyses of expression stability. Quantitative real-time PCR analyses for each candidate reference genes in A. leii was conducted for four developmental stages, four adult tissue types, two adult sexes, and two ecological stressors [adults exposed to five temperatures and larvae exposed to four concentrations of benzo(a)pyrene]. Results were evaluated by three independent algorithms (geNorm, NormFinder, and BestKeeper) and one comparative algorithm (RefFinder). The expression stability of candidate reference genes in A. leii differed under various conditions. Reference genes with the most stable expressions levels in different tissues, temperatures, sexes, developmental stages, and concentrations of benzo(a)pyrene were α-tubulin, GST, β-actin, β-tubulin, and α-tubulin, respectively. Furthermore, the optimal normalization factors (NFs) for the quantification of the expression levels of target genes by quantitative real-time PCR analyses of A. leii were identified for each experimental group. In particular, NF = 2 for different tissues (α-tubulin + β-tubulin), different sexes (β-actin + EF1A), and larvae exposed to different concentrations of benzo(a)pyrene (α-tubulin + EF1A); NF = 3 for developmental stages (GST + GAPDH + SDHA) and adults exposed to different temperatures (β-tubulin + EFA + GST). In addition, we surveyed the expression profiles of two target genes (CYP3A and CSP8) in larvae exposed to different concentrations of benzo(a)pyrene and in different adult tissues. The results further validated the reliability of the reference genes. The optimal reference genes for various experimental conditions identified in these analyses provide a useful tool for ecological studies of aquatic fireflies.
Introduction
Quantitative real-time PCR (qPCR) is a key technique for the quantification of expression levels of target genes under different experimental condition in genetic and evolutionary research (Zhang Q. L. et al., 2016). Useful applications include studies of ecological adaptation, growth and development, comparative tissues analyses, and stress resistance (Feng et al., 2016; Tang et al., 2017; Zhang et al., 2017; Jia et al., 2019). Raw qPCR data must be normalized by suitable reference genes (RGs), to avoid expression differences among samples (Zhang Q. L. et al., 2016; Zhang et al., 2017). Ideally, expression levels of optimal RGs should not be influenced by experimental conditions (Scharlaken et al., 2008; Chapman and Waldenström, 2015). However, expression profiles of commonly used RGs are not universally stable across different experimental conditions, because these RGs participate in mRNA translation and cell metabolism (Zhang Q. L. et al., 2016; Zhang et al., 2017). For example, actin (ACT) and ribosomal RNA 18S (18S) are widely used as RGs in insects, but their expression is not stable among Gynaephora moths at different altitudes (Zhang et al., 2017). In addition, the optimal number (i.e., normalization factor, NF) of RG combinations for experiments should be determined to improve the accuracy of qPCR results by a multi-RG strategy (Bansal et al., 2016; Zhang et al., 2017). Therefore, the determination of suitable RGs and optimal NFs for different experiments is a prerequisite for the normalization of expression levels of target genes in qPCR analyses.
Aquatica leii belongs to the order Coleoptera, family Lampyridae, subfamily Luciolinae, genus Aquatica (Fu and Meyer-Rochow, 2012; Jiao et al., 2013) and is mainly distributed in the middle and lower reaches of the Yangtze River in mainland China. A. leii is the most widely studied aquatic firefly species. All adult fireflies are terrestrial, however, according to the habitats of larvae, species can be divided into aquatic, semi-aquatic, and terrestrial types (Fu, 2005; Fu et al., 2012). A. leii is an important insect with a delight to watch and an ecological indicator owing to its flashing behavior and high sensitivity to deteriorating water quality caused by water pollutants (Fu, 2005; Zhang et al., 2019).
In recent years, increasing water pollution, artificial light source interference, commercial capture, climate change, and other human activities, have dramatically decreased the population size of aquatic fireflies (Fu, 2005; Zhang et al., 2019). Cold hardiness resulting from climate change is a crucial strategy for survival in insects. Thus, temperature largely determines the distribution, abundance, and rates of development of insects (Jing and Kang, 2002). Therefore, to protect aquatic firefly resources, it is essential to understand the mechanisms underlying ecological adaptation (e.g., water pollutants and temperature) at gene expression levels. Furthermore, gene expression across firefly lineage and its tissue specificity are important indicators for the study of the mechanisms underlying freshwater adaptation (Zhang et al., 2020). These genes can be identified via intraspecific gene expression difference comparisons, such as different developmental stages (terrestrial adults vs. aquatic larvae), sexes and tissues, as well as interspecific comparisons (e.g., aquatic vs. terrestrial fireflies) (Zhang et al., 2020). Therefore, it is important to identify suitable RGs for the calibration of target gene expression under different experimental conditions (e.g., developmental stages, tissues, sex, temperature, and particular water pollutants) in aquatic fireflies.
In this study, expression of 10 candidate RGs (Table 1) widely used for qPCR analyses of insects is not universally stable among different experimental conditions (Liu et al., 2014; Jia et al., 2019). Expression levels of these 10 candidate RGs were quantified in A. leii in four developmental stages, four adult tissues types, different sexes, and two ecological stressors (temperatures and water pollutants). Subsequently, the expression stability of RGs by qPCR was evaluated using the following four algorithms: geNorm, NormFinder, BestKeeper, and RefFinder. The optimal NFs for qPCR in five different experimental groups were determined using the geNorm algorithm. Finally, two target genes, CYP3A encoding cytochrome P450 3A and CSP8 encoding chemosensory protein 8, which have important detoxifying and olfactory functions, were chosen for validation.
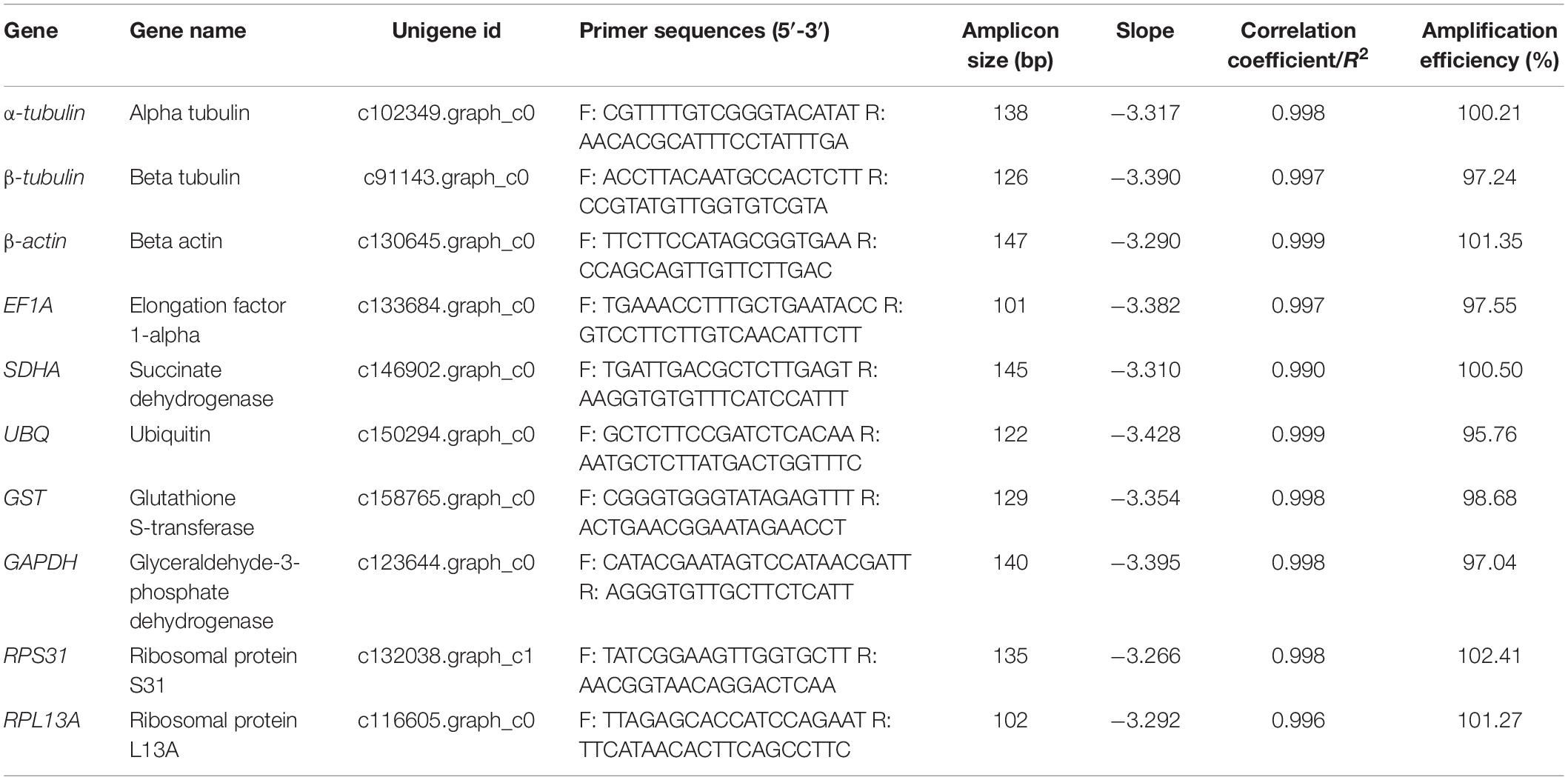
Table 1. Details of candidate RGs.
Materials and Methods
Animals
Aquatica leii was collected from the Culture Center of Fireflies, Ganzhou city, Jiangxi province, China. Before collecting or treating A. leii, larvae were maintained in a plastic box (40 cm × 30 cm × 10 cm) at 24−27°C under an 8-h light/16-h dark photoperiod, with oxygenators, heating rods (to increase the water temperature) and 5−6 cm of water (changed every 5 days) (detailed processes for the cultivation of fireflies are described in Zhang et al., 2020). A. leii adults were reared at 90 ± 5% relative humidity, under the same temperature and photoperiod cycle used for larvae. The expression of the reference genes was studied in animals and tissues which represented five experimental groups. First, different developmental stages; second, various types of adult tissues; third, adult sexes; fourth, adults exposed to different temperatures; and fifth, larvae exposed to a water contaminant. (1) Different developmental stage were collected from adult A. leii, such as 200 eggs, 20 larvae (4−6 instar mixture), 5 pupae, and 10 adults (a mixture of males and females). (2) Different tissues were dissected from adult A. leii, such as 30 heads, 30 thorax, 20 abdomen, and 100 antenna. (3) Ten individuals from each of adult female and male A. leii were collected. (4) Adult A. leii exposed to various temperatures for 2 h were collected, 10 individuals per different temperature, including 15°C, 20°C, 25°C, 30°C, and 35°C. (5) Due to ecological deterioration and intensifying human activity, polycyclic aromatic hydrocarbons (PAHs) can be easily detected in freshwater environments. Benzo(a)pyrene (BaP) is a PAH known as one of the most potent environmental carcinogens, and intensifying human activity is the main cause of its release into rivers and lakes (Xiao et al., 2018). Animals at the 4−6 larval instar stages were exposed to different concentrations of BaP for 24 h were collected as water contaminant-exposed groups, including control, 0.1, 0.01, and 0.001 mg/L, 20 individuals per concentration. Animals were collected three separate times, resulting in three biological replicates for each experimental group. All collected samples were immediately frozen in liquid nitrogen after labeling and stored at −80°C at the Faculty of Life Science and Technology, Kunming University of Science and Technology, Kunming, China.
RNA Extraction and cDNA Synthesis
Total RNA was extracted from each sample using TRIzol reagent (Ambion, Austin, TX, United States) according to the manufacturer’s instructions. RNase-free DNase (Qiagen, Hilden, Germany) was used to remove residual genomic DNA, following the manufacturer’s instructions. Three biological replicates were performed per sample. RNA quality and quantity were measured using a Nanodrop 1000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, United States), and RNA integrity was confirmed by 1.5% agarose gel electrophoresis. Only qualified RNA with an OD260/OD280 ratio between 1.8 and 2.0 and a concentration exceeding 300 ng/μL was retained for downstream analyses. Single-stranded cDNA was synthesized in a total volume of 20 μL, containing 4 μL of 5 × PrimeScript RT Master Mix (including oligo dT primers; TaKaRa, Kusatsu, Japan), 1 μL of total RNA (200 ng/μL), and 15 μL of RNase-free ddH2O at 37°C for 15 min and 85°C for 5 s, according to the manufacturer’s protocol with modifications. cDNA (100 ng/μL) was diluted with RNase-free water and used for subsequent experiments.
Design and Evaluation of Primers for Each Gene
According to previous studies (Liu et al., 2014; Nagy et al., 2017; Zhang et al., 2017; Jia et al., 2019), α-tubulin, β-tubulin, β-actin, EF1A, SDHA, UBQ, GST, GAPDH, RPS31, and RPL13A were identified as candidate RGs. These genes were identified from mixed assembled transcriptome (plus annotation) results for larval and adult A. leii (Zhang et al., 2020); original clean reads used for the assembly were downloaded from the NCBI Sequence Read Archive database, under accession no. PRJNA555399. All specific primers were designed using Beacon Designer 7.0. The specificity of the amplification product for each primer was checked by the appearance of a single band at the targeted length by 2% agarose gel electrophoresis. Detailed information for qPCR primers is presented in Table 1. Furthermore, all gene-specific amplified PCR products were confirmed by sequencing. The qPCR amplification efficiency (E) and correlation coefficient (R2) of each primer were calculated using a standard curve generated from a 10-fold dilution series of mixed cDNA samples at five dilutions (1/10, 1/100, 1/1000, 1/10000, and 1/100000). The corresponding qPCR amplification efficiency (E) was calculated according to the following formula: E(%) = (10(−1/slope)− 1)× 100 (Radoniæ et al., 2004; Zhang Q. L. et al., 2016).
Quantitative RT-PCR
Reactions were prepared in a total volume of 10 μL, including 1 μL of diluted cDNA (100 ng/μL), 5 μL of 2 × SYBR Premix ExTaq II (TaKaRa, Kusatsu, Japan), and 0.5 μL of each of the forward and reverse primers (10 ng/μL). RNase-free sterile water was added to obtain the final volume. The PCR program was as follows: 95°C for 35 s, 40 cycles of 95°C for 5 s, 55°C for 30 s, and 72°C for 30 s. A melting curve analysis was performed to confirm the amplification specificity for each reaction. A reaction solution without cDNA template was used as negative control to confirm template-specific amplification. PCRs were conducted for three biological replicates, and the detection of each gene was performed in an independent sample with three technical replicates. All qPCRs were conducted using an ABI 7300 real-time PCR system (Applied Biosystems, Waltham, MA, United States).
Determining the Expression Stability of Candidate RGs
Data for each of the five experimental groups were analyzed independently. Average Ct values for each candidate RG were calculated based on data for three biological and three technical replicates (n = 9) in each sample. BestKeeper (an Excel-based algorithm1) was employed to evaluate the expression stability of candidate RGs according to the correlation coefficient (r), standard deviation (SD), and coefficient of variation (CV) generated by pairwise comparison between candidate RGs. Smaller SD and CV-values indicated more stable expression levels of RGs (Pfaffl et al., 2004). geNorm2 sorted expression stabilities according to the M-values of candidate RGs (Vandesompele et al., 2002). A lower M-value indicated higher expression stability (Vandesompele et al., 2002). geNorm also calculates pairwise variation (V) between each RG and all other RGs to determine the NFs of RGs (Vandesompele et al., 2002). The value of Vn/(n+1) for two sequential NFs was used to determine the optimal number of RGs required for normalization. Values below a threshold of 0.15 suggest that no additional RGs are needed for normalization (Vandesompele et al., 2002). NormFinder3 assessed the expression stability of candidate RGs based on the stability value (SV) and SD of Ct values. A lower SV in NormFinder indicated more stable expression levels of RGs (Andersen et al., 2004). Finally, these candidate RGs were ranked using the web-based analytical tool RefFinder4, a comprehensive ranking tool that integrates different programs (Silver et al., 2006).
Validation of RGs
CYP3A, a detoxifying protein involved in the response to oxidative stress in insects, is up-regulated in A. leii in response to BaP exposure (Zhang et al., 2019). Chemosensory proteins, including olfaction-related genes in insect antennae, play important roles in olfaction (Zhang L. W. et al., 2016; Wang et al., 2017). Previous studies have reported the antenna-specific expression of CSPs in insects (Zhao et al., 2018). To confirm the reliability of the selected RGs, the expression profiles of the target gene CYP3A and CSP8 were determined in different concentrations of BaP and adult tissues, respectively, and independently normalized with the two most stable RGs as well as the least stable RGs. The PCR system and program were the same as those for the qPCR analysis of RGs. In qPCR analyses, three technical and three biological replicates were conducted for each treatment. Normalization of expression levels of target genes was performed according to the 2–ΔΔCT method (Livak and Schmittgen, 2001). Differences in target gene expression levels among concentrations and tissues were valuated by one-way ANOVA plus Bonferroni post hoc tests implemented in IBM SPSS 22.0.
Results
Expression Profiles of Candidate RGs
The specificity of gene amplification of all candidate RGs was confirmed by a single band with expected molecular mass using 1.5% agarose (Supplementary Figure 1). A standard curve was generated for each gene based on ten-fold serial dilution of the pooled cDNA at five dilutions. The amplification efficiencies of all candidate RGs were 95.76−102.41%, with correlation coefficients (R2) exceeding 0.995 (Table 1). The Ct value was the only parameter used to quantify the expression level of candidate RGs. Changes in Ct values of each candidate RG under five different experimental conditions are shown in Figure 1 and Supplementary Table 1. There was a wide range of expression levels and high variation across samples in expression patterns for the ten candidate RGs in each experimental group (Supplementary Figure 2). EF1A had the lowest Ct values, while GAPDH had the highest Ct values; the median Ct values for the 10 candidate RGs ranged from 15 to 25. Variance in Ct values decreased in the following order: GST > α-tubulin > β-tubulin > SDHA > GAPDH > UBQ > EF1A > β-actin > RPL13A > RPL31.
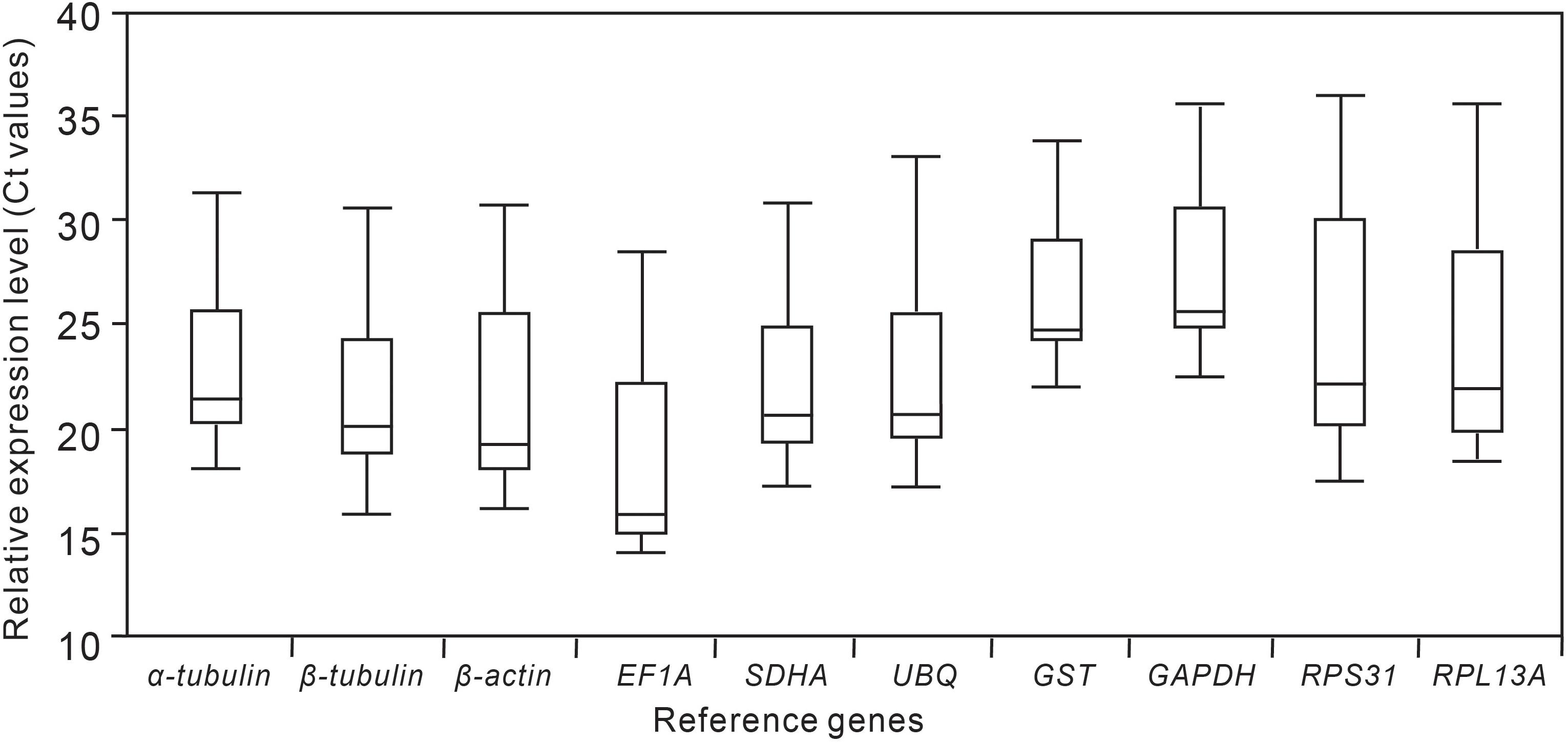
Figure 1. Expression levels of candidate reference genes in different samples. The expression levels are presented in terms of the raw cycle threshold number (Ct values) of the candidate reference genes in different samples. The line in the box indicates the median. The box ranges between the 25th and the 75th percentiles, and the maximum and minimum values are represented by upper and lower caps.
Expression Stability of Candidate RGs Determined Using BestKeeper
In different tissue types of adult A. leii, the SD values for α-tubulin and β-tubulin were less than 1, and values for other candidate RGs exceeded 1. α-Tubulin and β-tubulin showed high expression stability. In adult A. leii exposed to different temperatures, only the SD value of GST was less than 1; suggesting that GST has the most stable expression. Among other candidate RGs, RPS31 expression was most unstable. In adults of different sexes, the SD values of UBQ and GAPDH exceeded 1, and showed the least stable expression. β-Tubulin, EF1A, and β-actin had the most stable expression, followed by RPL13A, α-tubulin, GST, SDHA, and RPS31. At different developmental stages, the expression of GAPDH was most stable. In larvae exposed to different concentrations of BaP, the SD values for all 10 candidate RGs were below 1, and the expression stability of these candidates could be ordered from high to low as follows: α-tubulin, EF1A, RPS31, β-tubulin, GST, β-actin, UBQ, SDHA, GAPDH, and RPL13A (Table 2 and Supplementary Table 2).
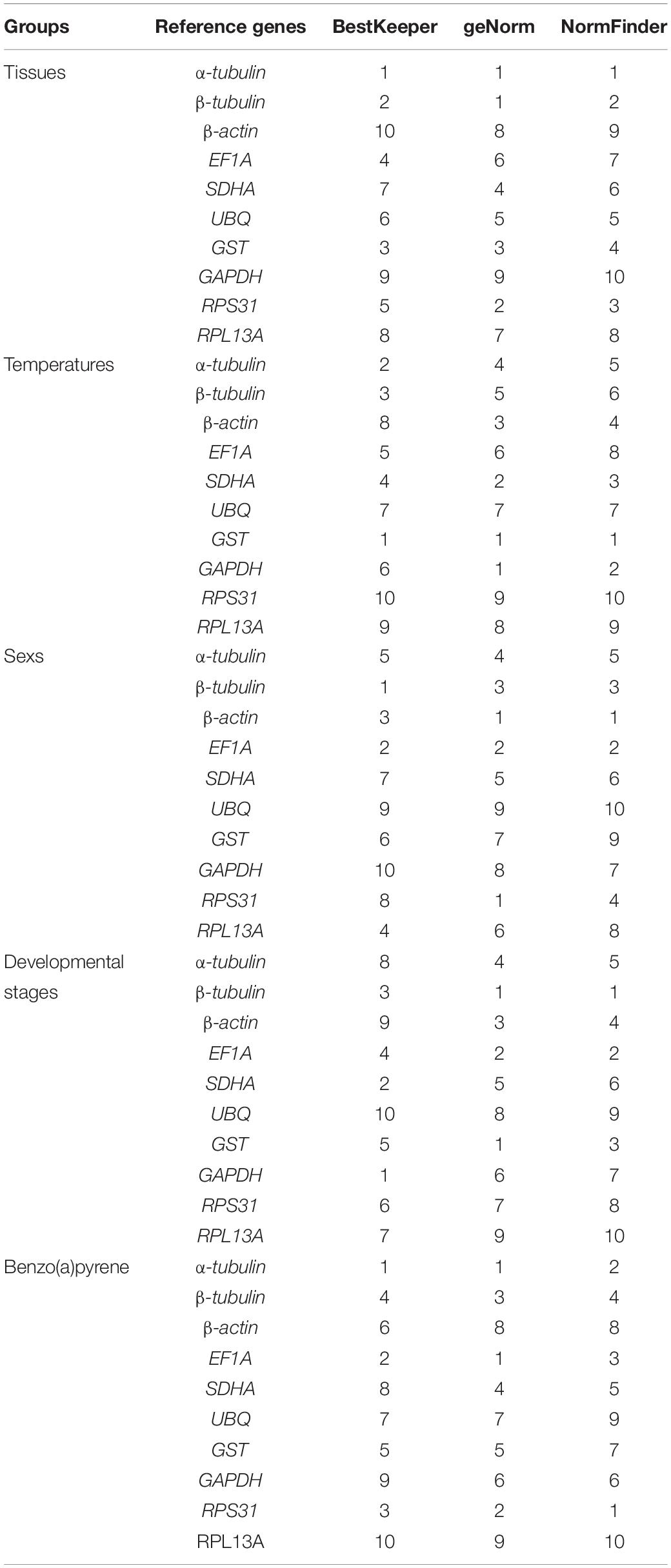
Table 2. Expression stability ranking of candidate reference genes in Aquatica leii under five different experimental groups.
Expression Stability of Candidate RGs Determined Using geNorm
In different adult tissues of A. leii, the three candidate RGs with the highest expression stabilities were α-tubulin, β-tubulin, and RPS31. The three candidate RGs with the most unstable expression levels were GAPDH, β-actin, and RPL13A. In adults exposed to different temperatures, the three candidate RGs with the highest expression stabilities were GST, GAPDH, and SDHA, and the three candidate RGs with the most unstable expression levels were RPS31, RPL13A, and UBQ. In adults of different sexes, the three candidate RGs with the highest expression stabilities were β-actin, RPS31, and EF1A, and three candidates with the worst expression stabilities were UBQ, GAPDH, and GST. At different developmental stages, the three most stable candidate RGs were β-tubulin, GST, EF1A, RPL13A, and UBQ, while RPS31 showed the most unstable expression. In larvae exposed to different concentrations of BaP, the three most stable candidates were α-tubulin, EF1A, and RPS31, while the most unstable candidates were RPL13A, β-actin, and UBQ (Table 2 and Supplementary Table 3).
Expression Stability of Candidate RGs Determined Using NormFinder
The NormFinder analysis showed that the expression stability of candidate RGs in different adult tissues was in the following order: α-tubulin > β-tubulin > RPS31 > GST > UBQ > SDHA > EF1A > RPL13A > β-actin > GAPDH. In adults exposed to different temperatures, the expression stability of candidate RGs decreased in the following order: GST > GAPDH > SDHA > β-actin > α-tubulin > β-tubulin > UBQ > EF1A > RPL13A > RPS31. In adults of different sexes, the expression stabilities of candidate RGs ranked as follows: β-actin > EF1A > β-tubulin > RPS31 > α-tubulin > SDHA > GAPDH > RPL13A > GST > UBQ. At different developmental stages, the expression stabilities of candidate RGs decreased in the following order: β-tubulin > EF1A > GST > β-actin > α-tubulin > SDHA > GAPDH > RPS31 > UBQ > RPL13A. In larvae exposed to different concentrations of BaP, the expression stabilities of candidate RGs decreased in the following order: RPS31 > α-tubulin > EF1A > β-tubulin > SDHA > GAPDH > GST > β-actin > UBQ > RPL13A (Table 2 and Supplementary Table 4).
Expression Stability of Candidate RGs Determined Using RefFinder
The expression stabilities of candidate RGs in different adult tissues decreased in the following order: α-tubulin > β-tubulin > RPS31 > GST > UBQ > SDHA > EF1A > RPL13A > β-actin > GAPDH. In adults exposed to different temperatures, the expression stabilities of candidate RGs decreased in the following order: GST > GAPDH > SDHA > α-tubulin > β-tubulin > β-actin > EF1A > UBQ > RPL13A > RPS31. In adults of different sexes, the expression stabilities of candidate RGs decreased in the following order: β-actin > EF1A > β-tubulin > RPS31 > α-tubulin > SDHA > RPL13A > GST > GAPDH > UBQ. At different developmental stages, the expression stabilities of candidate RGs decreased in the following order: β-tubulin > EF1A > GST > SDHA > GAPDH > β-actin > α-tubulin > RPS31 > RPL13A > UBQ. In larvae exposed to different concentrations of BaP, the expression stabilities of candidate RGs decreased in the following order: α-tubulin > EF1A > RPS31 > β-tubulin > SDHA > GST > GAPDH > β-actin > UBQ > RPL13A (Table 3).
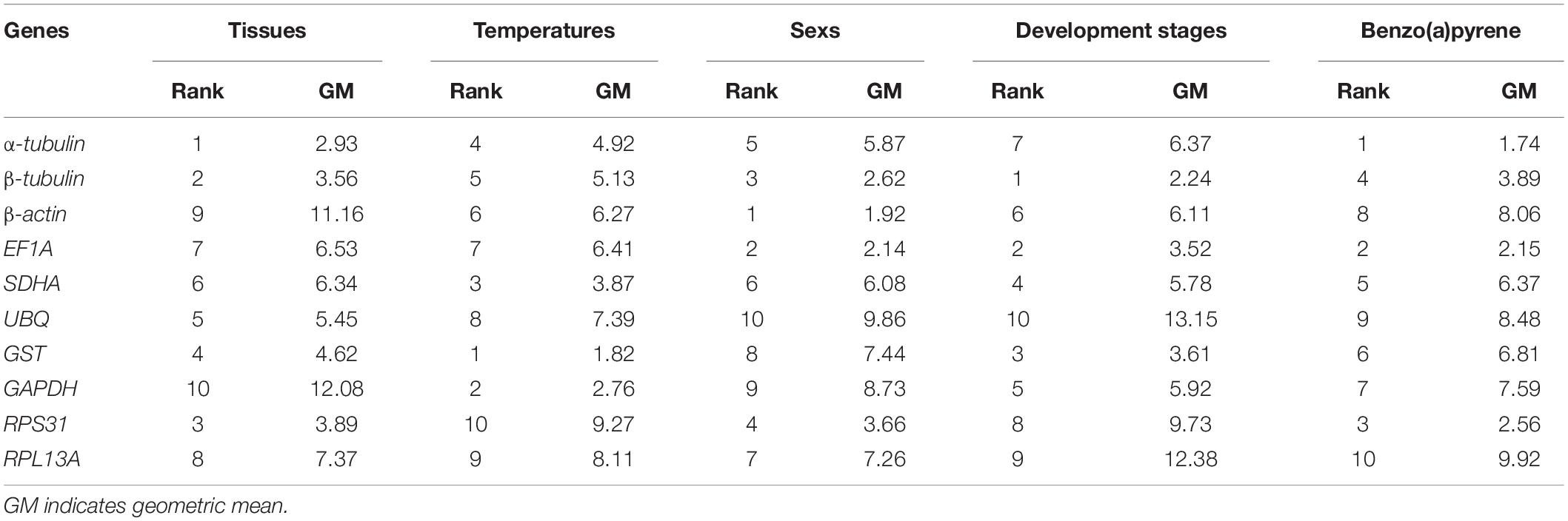
Table 3. Expression stability of candidate reference genes in Aquatica leii under five different experimental groups calculated by RefFinder.
Determination of Optimal NFs
If V-value is initially lower than the default value 0.15 in all pairwise variation, or V-value is lowest when all the V-values are more than 0.15, the number of the corresponding gene pairings will be good enough for normalization (Zhang et al., 2017). The V2/3 value was first lower than 0.15 in all pairwise variation in adult tissues, sexes, and larval BaP concentrations (Figure 2), indicating that the optimal number of RGs for normalization was two for the above three experimental groups, and the combinations were α-tubulin + β-tubulin, β-actin + EF1A, and α-tubulin + EF1A, respectively (Table 4). At different developmental stages and temperatures, all Vn/(n+1) values exceeded 0.15, and the lowest value was more than V4/5 and V7/8, respectively. According to the geNorm recommendation, the optimal number of NFs was three, and the optimal combinations of RGs were GST + GAPDH + SDHA and β-tubulin + EFA + GST, respectively.
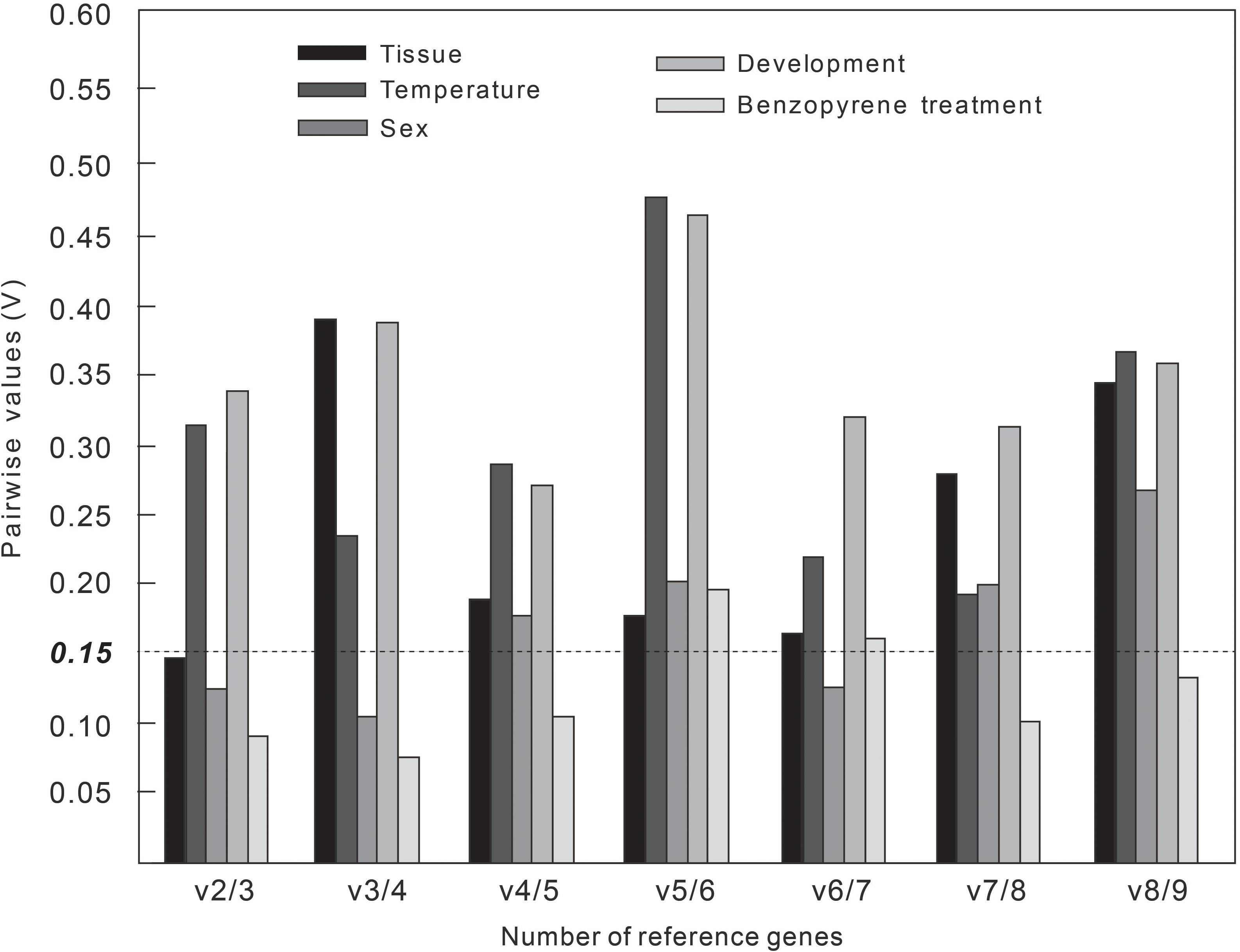
Figure 2. Number of optimal combinations of reference genes under five different experimental groups.

Table 4. Optimal combination of reference genes in Aquatica leii under five different experimental groups.
Validation of RGs
CYP3A and CSP8 were selected as target genes to validate the applicability of the selected RGs. Under the most stable RGs (α-tubulin and/or EF1A) at different BaP concentrations, the expression patterns of CYP3A were consistent with down-regulation at low BaP concentration period and up-regulation at higher concentration period. This indicates that we have shown a reliable expression pattern of CYP3A using the most stable RGs screened in this study [higher expression level of CYP3A under higher BaP concentrations (Zhang et al., 2019)]. Nevertheless, when the most unstable RG (RPL13A) was employed to normalize expression levels of target genes, CYP3A failed to present a consistent expression pattern (Figure 3). In different adult tissues, when the most stable RGs (α-tubulin and/or β-tubulin) were used as normalization factors, the expression of the target gene CSP8 was highest in the antenna. However, when the most unstable RG (GAPDH) was employed, the expression pattern of CSP8 significantly differed with normalization by α-tubulin and α-tubulin + β-tubulin (Figure 3). It is generally true that the accurate expression of target genes could be obtained, when the most stable RGs are employed as normalization factors.
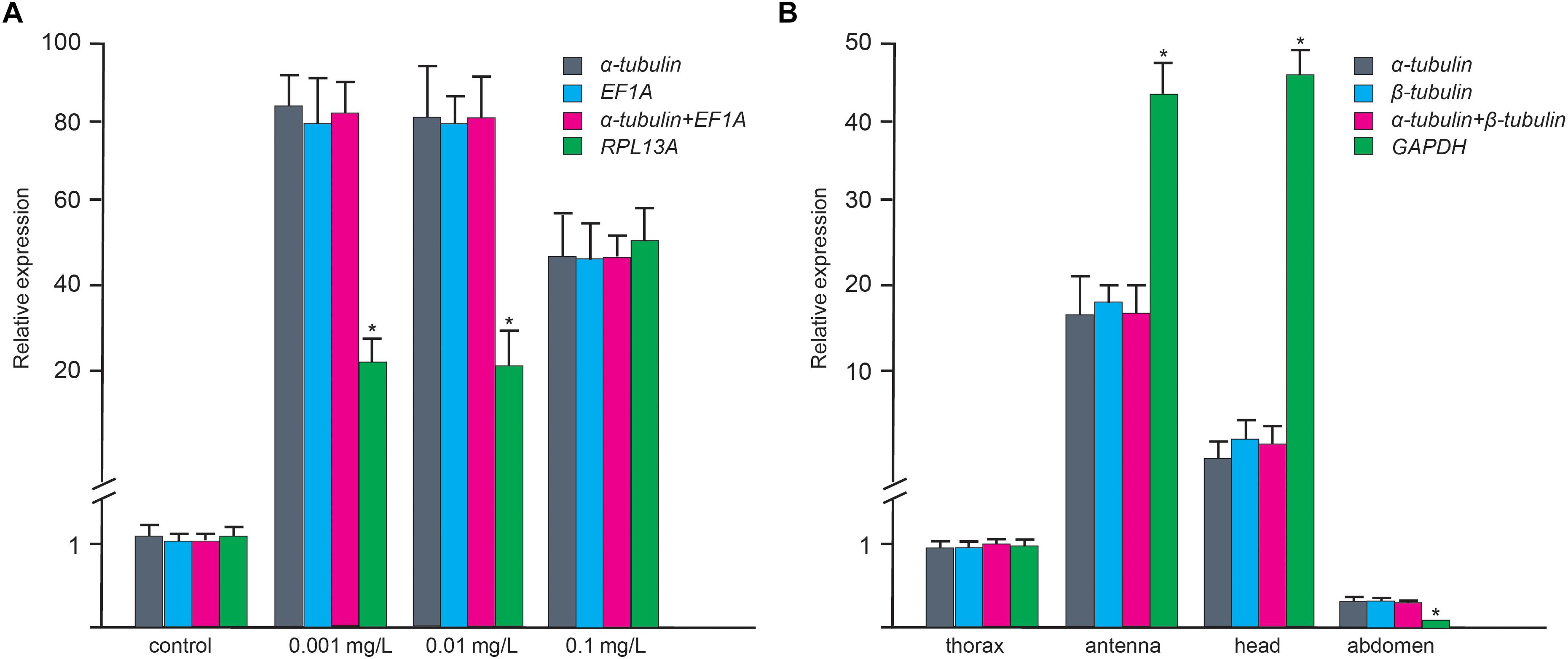
Figure 3. Validation of selected reference genes in larval A. leii under different BaP concentrations (A) and adult tissues (B) in A. leii. Relative expression level of the target genes (CYP3A and CSP8) in different samples using different normalization factors (the most and least stable genes). Asterisks indicate significant differences in the expression levels of the target genes.
Discussion
In this study, the results obtained by different independent algorithms were highly consistent. For example, the stability ranking of candidate RGs in geNorm was highly similar to that of NormFinder. However, there were also differences among algorithms with respect to the expression stability of candidate RGs, as reported in many similar studies (e.g., Ibanez and Tamborindeguy, 2016; Koramutla et al., 2016; Zhang et al., 2017). A comparison of the three algorithms used here showed that BestKeeper exhibited the most discrepancies, consistent with previous reports (Lu et al., 2015; Zhang et al., 2017). This result indicated that it is essential to select optimal RGs under specific conditions using multiple independent algorithms.
In the current study, we evaluated common genes used for normalization. α-Tubulin and β-tubulin showed high expression stability in different adult tissues and in larvae exposed to different BaP concentrations in A. leii. Even β-tubulin retained a highly stable expression level in different adult sexes and developmental stages, indicating that β-tubulin is a suitable RG for A. leii under various experimental conditions. Therefore, when several limitations (e.g., limited samples, time, and funds) prevented experiments that screened the most suitable RGs, α-tubulin and β-tubulin can be considered as optimal RGs for qPCR analyses of target genes in fireflies. β-Actin, a traditional insect RG, showed high expression stability only in the sex group and showed either low or the lowest expression stability in the other four experimental conditions evaluated. Similar results were reported in investigations of RGs in insects belonging to Hemiptera and Lepidoptera (Zhang et al., 2017; Jia et al., 2019). Accordingly, traditional and widely used RGs may not be stably expressed in many experimental conditions and thus should be used cautiously for qPCR analyses. EF1A showed extreme expression stability between sexes, developmental stages, and larvae exposed to different concentrations of BaP in A. leii. This was consistent with the results of RG selection for qPCR analyses of grassland caterpillars (genus Gynaephora) along an altitudinal gradient (Zhang et al., 2017). However, the expression stability of EF1A was low among different adult tissues and temperature treatments. These results provided a useful insight into the appropriate conditions for the application of EF1A in A. leii. GST showed the highest expression stability in A. leii adults exposed to different temperatures; thus, this gene could be considered a suitable RG for qPCR analyses of the temperature response of A. leii. RPS31 and RPL13A are ribosomal protein-coding genes. Several studies have reported that these two candidate RGs are stably expressed in Lygus pratensis, and participate in protein translation and synthesis (Jia et al., 2019). Our results showed that the expression stability of RPS31 was higher than that of RPL13A in five experimental conditions in A. leii. This indicates that candidate RGs with similar biological functions exhibit differences in expression stability. Both SDHA and GAPDH showed high expression stability in several experimental conditions in A. leii; thus, these two genes should also be considered candidates for studies of other firefly groups. UBQ showed moderate or low expression stability in all five experimental conditions evaluated; therefore, UBQ can be excluded from candidate RGs in the future.
Many studies have proposed that multiple RGs should be used in combination to improve normalization (i.e., biased normalization can be removed via mutiple-RG correction) (Kumar et al., 2012; Fu et al., 2013; Dzaki and Azzam, 2018). The optimal numbers of NFs were seven and four for A. leii at different temperatures and developmental stages, respectively. However, owing to limited experimental resources and time, it is often unrealistic to use many RGs in combination in practical applications. Moreover, an excessive number of RGs could introduce more uncertainty and error, decreasing the accuracy of expression normalization of target genes, particularly for NFs exceeding three (Fu et al., 2013; Zhu et al., 2014; Jia et al., 2019). Therefore, geNorm suggests that the optimal number of NFs should not exceed three (Vandesompele et al., 2002). In addition, the validation results for the RGs suggested that it is essential to use different RGs depending on experimental conditions, and the selection and validation of the RGs for aquatic firefly species was effective in this study.
Conclusion
In this study, suitable RGs were identified for investigations of ecological adaptation in fireflies. As a case study, this study represents first key step toward establishing a standardized qPCR analysis procedure for firefly species. Overall, α-tubulin, GST, β-actin and β-tubulin were effective RGs in five different experimental conditions. As additional genomes and transcriptomes of various firefly species are obtained, the results of this study lay a foundation for subsequent research in genomics and functional genomics. Notably, for other experimental conditions, it will be essential to evaluate the stability of candidate RGs by standard processes.
Data Availability Statement
All datasets generated for this study are included in the article/Supplementary Material.
Author Contributions
Q-LZ and X-YD conceived and designed the study. X-JY, H-LZ, Y-YL, H-WL, and Y-HJ performed the experiments. Q-LZ, X-JY, H-LZ, Y-YL, H-WL, and Y-HJ analyzed the data. Q-LZ and X-JY carried out the statistics. Q-LZ, X-JY, H-WL, and Y-HJ drafted the manuscript. L-BL, X-YD, and Q-LZ revised the manuscript. All authors have read, commented on, and approved the manuscript.
Funding
This work was supported by the Natural Science Foundation of China (31960286), and Yunnan Fundamental Research Projects, and the Undergraduate Innovation and Entrepreneurship Training Program of Kunming University of Science and Technology (201910674321).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphys.2020.555233/full#supplementary-material
Supplementary Figure 1 | Specificity and product length of PCR amplification for ten candidate RGs, including α-tubulin (a), β-tubulin (b), β-actin (c), EFA (d), SDHA (e), UBQ (f), GST (g), GAPDH (h), RPS31 (i), RPL13A (j).
Supplementary Figure 2 | Ct values of ten candidate reference genes among samples for each experimental group. A, different normal tissues; B, different temperature; C, different sex; D, different developmental stages; E, different dose of benzopyrene (mg/L). The line in the box indicates median. The box describes between 25th and 75th percentiles, the maximum and minimum values and were represented by up and low caps.
Supplementary Table 1 | Ct values (mean ± SD) of candidate reference genes of Aquatica leii under five different experimental groups.
Supplementary Table 2 | Expression stability (SD ± [CP]) of candidate reference genes in Aquatica leii under different experimental groups calculated by BestKeeper.
Supplementary Table 3 | Expression stability (M) of candidate reference genes in Aquatica leii under different experimental groups calculated by geNorm.
Supplementary Table 4 | Expression stability of candidate reference genes in Aquatica leii under different experimental groups calculated by NormFinder.
Abbreviations
BaP, benzo(a)pyrene; CV, coefficient of variation; NFs, normalization factors; PAHs, polycyclic aromatic hydrocarbons; qPCR, quantitative real-time PCR; RGs, reference genes; SD, standard deviation.
Footnotes
- ^ http://www.gene-quantification.de/bestkeeper.html
- ^ https://genorm.cmgg.be/
- ^ http://moma.dk/normfinder-software
- ^ https://omictools.com/reffinder-tool
References
Andersen, C. L., Jensen, J. L., and Ørntoft, T. F. (2004). Normalization of real-time quantitative reverse transcription-PCR data: a model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 64, 5245–5250. doi: 10.1158/0008-5472.CAN-04-0496
Bansal, R., Mittapelly, P., Chen, Y., Mamidala, P., Zhao, C., and Michel, A. (2016). Quantitative RT-PCR gene evaluation and RNA interference in the brown marmorated stink bug. PLoS One 11:e0152730. doi: 10.1371/journal.pone.0152730
Chapman, J. R., and Waldenström, J. (2015). With reference to reference genes: a systematic review of endogenous controls in gene expression studies. PLoS One 10:e0141853. doi: 10.1371/journal.pone.0141853
Dzaki, N., and Azzam, G. (2018). Assessment of Aedes albopictus reference genes for quantitative PCR at different stages of development. PLoS One 13:e0194664. doi: 10.1371/journal.pone.0194664
Feng, B., Guo, Q. S., Mao, B. P., and Du, Y. J. (2016). Identification and selection of valid reference genes for assaying gene expression in the chemosensory tissues of Monochamus alternatus (Coleoptera: Cerambycidae) by RT-qPCR. Acta Entomol. Sin. 59, 427–437. doi: 10.16380/j.kcxb.2016.04.008
Fu, W., Xie, W., Zhang, Z., Wang, S., Wu, Q., Liu, Y., et al. (2013). Exploring valid reference genes for quantitative real-time PCR analysis in Plutella xylostella (Lepidoptera: Plutellidae). Int. J. Biol. Sci. 9, 792–802. doi: 10.7150/ijbs.5862
Fu, X. H. (2005). The Biology and Behavior of Two Species of Aquatic Fireflies in Mainland China. Doctoral dissertation, Huazhong Agricultural University, Wuhan.
Fu, X. H., and Meyer-Rochow, V. B. (2012). An investigation into the morphological and behavioral adaptations of the aquatic larvae of Aquatica leii (Coleoptera: Lampyridae) to prey upon freshwater snails that serve as intermediate hosts for the liver fluke. Biol. Control. 62, 127–134. doi: 10.1016/j.biocontrol.2011.12.007
Fu, X. H., Lesley, B., and Christine, L. (2012). The external larval morphology of aquatic and terrestrial Luciolinae fireflies (Coleoptera: Lampyridae). Zootaxa 3405, 1–34. doi: 10.1111/j.1600-0684.2012.00549.x
Ibanez, F., and Tamborindeguy, C. (2016). Selection of reference genes for expression analysis in the potato psyllid, Bactericera cockerelli. Insect Mol. Biol. 25, 227–238. doi: 10.1111/imb.12219
Jia, B., Ma, Y., Pang, B. P., Shan, Y. M., Bao, Q. L., Han, H. B., et al. (2019). Screening of reference genes for quantitative real-time PCR in Lygus pratensis (Hemiptera: Miridae). Acta Entomol. Sin. 62, 1379–1391. doi: 10.16380/j.kcxb.2019.12.004
Jiao, H., Ding, M., and Zhao, H. (2013). Sequence and organization of complete mitochondrial genome of the firefly, Aquatica leii (Coleoptera: Lampyridae). Mitochondr. DNA 26, 775–776. doi: 10.3109/19401736.2013.855746
Jing, X. H., and Kang, L. (2002). Research progress in insect cold hardiness. Acta Ecol. Sin. 22, 2202–2207. doi: 10.3321/j.issn:1000-0933.2002.12.026
Koramutla, M. K., Aminedi, R., and Bhattacharya, R. (2016). Comprehensive evaluation of candidate reference genes for qRT-PCR studies of gene expression in mustard aphid, Lipaphis erysimi (Kalt). Sci. Rep. 6:25883. doi: 10.1038/srep25883
Kumar, P., Yadav, P., Verma, A., Singh, D., De, S., and Datta, T. K. (2012). Identification of stable reference genes for gene expression studies using quantitative real time PCR in buffalo oocytes and embryos. Reprod. Domest. Anim. 47, e88–e91. doi: 10.1111/j.1439-0531.2012.01998.x
Liu, J. P., Ou, J., Yao, F. J., He, L., Xi, G. S., and Wei, C. M. (2014). Identification of appropriate reference genes for gene expression studies by quantitative real-time PCR in Tribolium castaneum after exposure to phosphine. J. Agric. Biotechnol. 22, 257–264. doi: 10.3969/j.issn.1674-7968.2014.02.016
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Lu, Y. H., Zheng, X. S., Liang, Q., Xu, H. X., Yang, Y. J., Tian, J., et al. (2015). Evaluation and validation of reference genes for SYBR Green qRT-PCR normalization in Sesamia inferens (Lepidoptera: Noctuidae). J. Asia Pac. Entomol. 18, 669–675. doi: 10.1016/j.aspen.2015.08.002
Nagy, N. A., Németh, Z., Juhász, E., Póliska, S., Rácz, R., Kosztolányi, A., et al. (2017). Evaluation of potential reference genes for real-time qPCR analysis in a biparental beetle, Lethrus apterus (Coleoptera: Geotrupidae). PeerJ 5:e4047. doi: 10.7717/peerj.4047
Pfaffl, M. W., Tichopad, A., Prgomet, C., and Neuvians, T. P. (2004). Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: bestKeeper – Excel-based tool using pair-wise correlations. Biotechnol. Lett. 26, 509–515. doi: 10.1023/b:bile.0000019559.84305.47
Radoniæ, A., Thulke, S., Mackay, I. M., Landt, O., Siegert, W., and Nitsche, A. (2004). Guideline to reference gene selection for quantitative real-time PCR. Biochem. Biophys. Res. Commun. 313, 856–862. doi: 10.1016/j.bbrc.2003.11.177
Scharlaken, B., de Graaf, D. C., Goossens, K., Brunain, M., Peelman, L. J., and Jacobs, F. J. (2008). Reference gene selection for insect expression studies using quantitative real-time PCR: the head of the honeybee, after a bacterial challenge. J. Insect Sci. 8:33. doi: 10.1673/031.008.3301
Silver, N., Best, S., Jiang, J., and Thein, S. (2006). Selection of housekeeping genes for gene expression studies in human reticulocytes using real-time PCR. BMC Mol. Biol. 7:33. doi: 10.1186/1471-2199-7-33
Tang, P. A., Wu, H. J., Xue, H., Ju, X. R., Song, W., Zhang, Q. L., et al. (2017). Characterization of transcriptome in the Indian meal moth Plodia interpunctella (Lepidoptera: Pyralidae) and gene expression analysis during developmental stages. Gene 622, 29–41. doi: 10.1016/j.gene.2017.04.018
Vandesompele, J., De Preter, K., Pattyn, F., Poppe, B., Van Roy, N., De Paepe, A., et al. (2002). Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 3:research0034. doi: 10.1186/gb-2002-3-7-research0034
Wang, J., Hu, P., Gao, P., Tao, J., and Luo, Y. (2017). Antennal transcriptome analysis and expression profiles of olfactory genes in Anoplophora chinensis. Sci. Rep. 7:15470. doi: 10.1038/s41598-017-15425-2
Xiao, R., Zhou, H., Chen, C. M., Cheng, H., Li, H., Xie, J., et al. (2018). Transcriptional responses of Acropora hyacinthus embryo under the benzo(a)pyrene stress by deep sequencing. Chemosphere 206, 387–397. doi: 10.1016/j.chemosphere.2018.04.149
Zhang, L., Zhang, Q. L., Wang, X. T., Yang, X. Z., Li, X. P., and Yuan, M. L. (2017). Selection of reference genes for qRT-PCR and expression analysis of high-altitude-related genes in grassland caterpillars (Lepidoptera: Erebidae: Gynaephora) along an altitude gradient. Ecol. Evol. 7, 9054–9065. doi: 10.1002/ece3.3431
Zhang, Q. L., Guo, J., Deng, X. Y., Wang, F., Chen, J. Y., and Lin, L. B. (2019). Comparative transcriptomic analysis provides insights into the response to the benzo(a)pyrene stress in aquatic firefly (Luciola leii). Sci. Total Environ. 661, 226–234. doi: 10.1016/j.scitotenv.2019.01.156
Zhang, Q. L., Li, H. W., Dong, Z. X., Yang, X. J., Lin, L. B., Chen, J. Y., et al. (2020). Comparative transcriptomic analysis of fireflies (Coleoptera: Lampyridae) to explore the molecular adaptations to fresh water. Mol. Ecol. 29, 2676–2691. doi: 10.1111/mec.15504
Zhang, L. W., Kang, K., Jiang, S. C., Zhang, Y. N., Wang, T. T., Zhang, J., et al. (2016). Analysis. (of) the antennal transcriptome and insights into olfactory genes in Hyphantria cunea (Drury). PLoS One 11:e0164729. doi: 10.1371/journal.pone.0164729
Zhang, Q. L., Zhu, Q. H., Liao, X., Wang, X. Q., Chen, T., Xu, H. T., et al. (2016). Selection. (of) reliable reference genes for normalization of quantitative RT-PCR from different developmental stages and tissues in amphioxus. Sci. Rep. 6:37549. doi: 10.1038/srep37549
Zhao, Y., Ding, J., Zhang, Z., Liu, F., Zhou, C., and Mu, W. (2018). Sex- and tissue-specific expression profiles of odorant binding protein and chemosensory protein genes in Bradysia odoriphaga (Diptera: Sciaridae). Front. Physiol. 9:107. doi: 10.3389/fphys.2018.00107
Keywords: Aquatica leii, qPCR, reference gene, gene expression, stability evaluation
Citation: Yang X-J, Zheng H-L, Liu Y-Y, Li H-W, Jiang Y-H, Lin L-B, Deng X-Y and Zhang Q-L (2020) Selection of Reference Genes for Quantitative Real-Time PCR in Aquatica leii (Coleoptera: Lampyridae) Under Five Different Experimental Conditions. Front. Physiol. 11:555233. doi: 10.3389/fphys.2020.555233
Received: 13 May 2020; Accepted: 08 September 2020;
Published: 06 October 2020.
Edited by:
Xanthe Vafopoulou, York University, CanadaReviewed by:
Linkai Huang Monika Sodhi, Linkai Huang Monika Sodhi, Sichuan Agricultural University, ChinaCopyright © 2020 Yang, Zheng, Liu, Li, Jiang, Lin, Deng and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Qi-Lin Zhang, emhhbmdxbEBrdXN0LmVkdS5jbg==; Xian-Yu Deng, ZGVuZ3h5MTAwOEAxMjYuY29t; Lian-Bing Lin, bGlubGJAa3VzdC5lZHUuY24=