 Hannah Chew
Hannah Chew Victoria A. Solomon3
Victoria A. Solomon3 Alfred N. Fonteh
Alfred N. Fonteh- 1Huntington Medical Research Institutes, Pasadena, CA, United States
- 2University of California, Los Angeles, Los Angeles, CA, United States
- 3University of Southern California, Los Angeles, CA, United States
Lipids constitute the bulk of the dry mass of the brain and have been associated with healthy function as well as the most common pathological conditions of the brain. Demographic factors, genetics, and lifestyles are the major factors that influence lipid metabolism and are also the key components of lipid disruption in Alzheimer’s disease (AD). Additionally, the most common genetic risk factor of AD, APOE ϵ4 genotype, is involved in lipid transport and metabolism. We propose that lipids are at the center of Alzheimer’s disease pathology based on their involvement in the blood-brain barrier function, amyloid precursor protein (APP) processing, myelination, membrane remodeling, receptor signaling, inflammation, oxidation, and energy balance. Under healthy conditions, lipid homeostasis bestows a balanced cellular environment that enables the proper functioning of brain cells. However, under pathological conditions, dyshomeostasis of brain lipid composition can result in disturbed BBB, abnormal processing of APP, dysfunction in endocytosis/exocytosis/autophagocytosis, altered myelination, disturbed signaling, unbalanced energy metabolism, and enhanced inflammation. These lipid disturbances may contribute to abnormalities in brain function that are the hallmark of AD. The wide variance of lipid disturbances associated with brain function suggest that AD pathology may present as a complex interaction between several metabolic pathways that are augmented by risk factors such as age, genetics, and lifestyles. Herewith, we examine factors that influence brain lipid composition, review the association of lipids with all known facets of AD pathology, and offer pointers for potential therapies that target lipid pathways.
Background
The Importance of Cellular Lipid Membranes
Cell membranes are composed of several lipid classes and membrane-bound proteins/receptors that interface cellular organelles, and cells with their environment. It is now recognized that these membrane lipids are important in maintaining cellular functions. Several studies show that perturbation of membrane lipids can have devastating consequences on the brain. These changes underlie Alzheimer’s disease (AD) pathology depicted in Figure 1. We will examine factors that affect lipid metabolism, describe the functions of brain lipids, and examine the consequences and contributions of lipid dyshomeostasis on AD pathology.
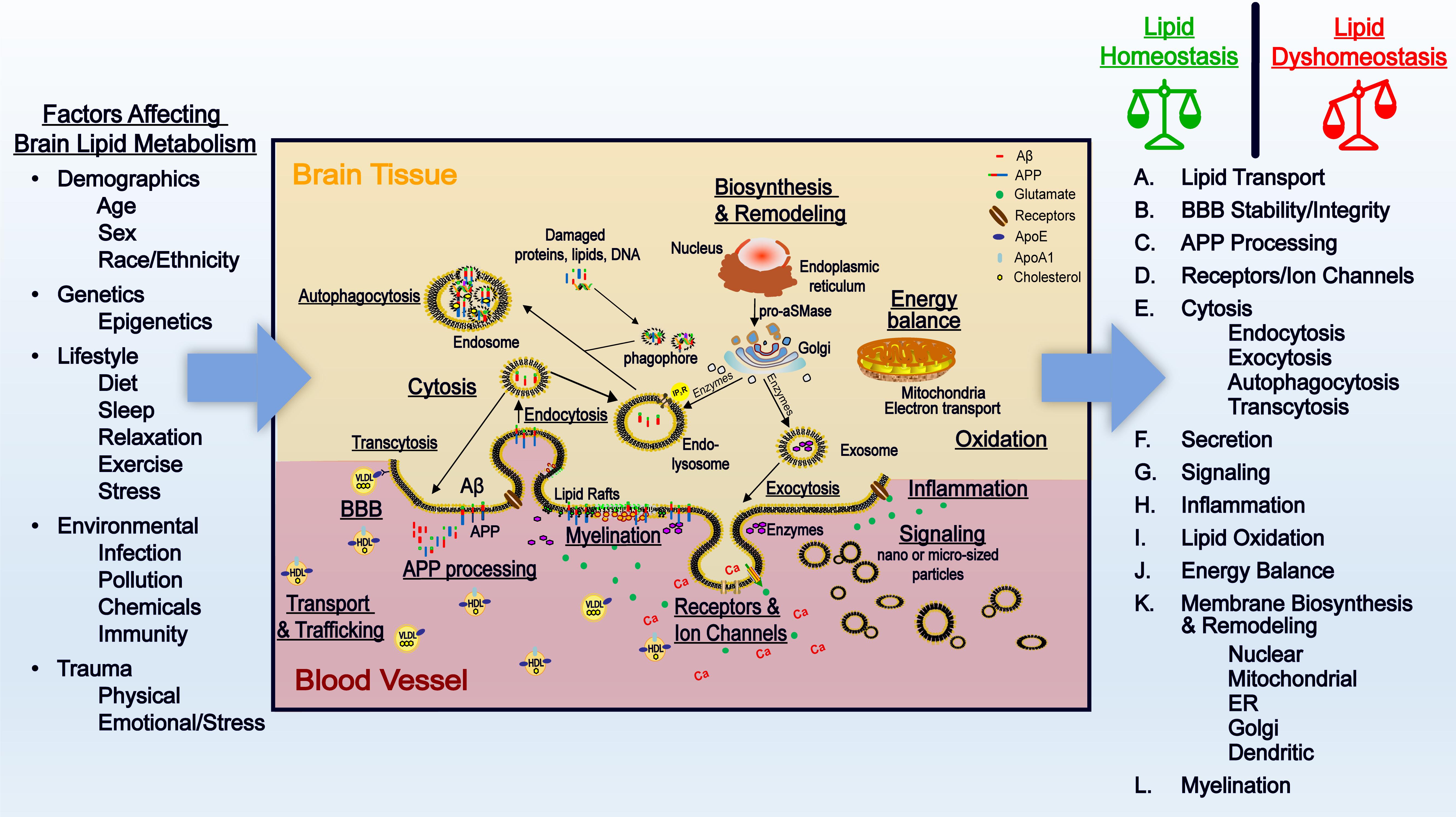
Figure 1. Factorsthat affect brain lipid metabolism and the importance of lipids in healthy aging and AD. Factors that affect brain lipid metabolism – Demographic factors, genetics, lifestyle, the environment, and trauma can influence lipid metabolism in the brain. Interestingly, these factors that influence lipid metabolism are also recognized risk factors of AD. Abnormalities in lipid metabolism can contribute to dysfunctional brain networks that associate with AD pathology. Importance of lipid metabolism in brain function and AD pathology – In healthy aging, normal transport of lipids through apolipoproteins contribute to the function of the brain. Homeostatic control of the brain lipid environment is responsible for sustaining a normal BBB, providing the right environment for normal APP processing, the right composition for ion channels and receptors, cytosis, vesicle formation, and secretion, signaling, inflammation, oxidation, energy balance, and membrane biosynthesis and remodeling. Dyshomeostasis in lipid delivery into the brain and its metabolism attributes to disturbed BBB, abnormal APP processing, disturbance in cytosis, signaling, energy balance, and enhanced/sustained inflammation and oxidation. Over time, these processes lead to neuronal death that is the hallmark of AD pathology.
Brain Lipids in Healthy Aging and AD Pathology
Most of the brain is composed of lipids, which can be grouped as sphingolipids, glycerophospholipids, and cholesterol (Svennerholm et al., 1994; O’Brien and Sampson, 1965; Kishimoto et al., 1969). The brain consists of straight-chain monocarboxylic acids ranging from C12 to C26, and omega-3 (n-3) and omega-6 (n-6) fatty acids are most abundant (Kishimoto et al., 1969; Siegel, 1999). Docosahexaenoic acid (DHA) and eicosapentaenoic acid (EPA) are prominent polyunsaturated fatty acids (PUFA) in the brain that are derived from alpha-linolenic acid (ALA), an omega-3 fatty acid (Chappus-McCendie et al., 2019). Arachidonic acid (AA) and docosatetraenoic acid (DPA) constitute a large proportion of PUFA’s that are derived from linolenic acid (LNA), an omega 6 fatty acid (Leonard et al., 2000; Sinclair et al., 2007).
Factors That Affect Brain Lipids
Demographic Factors That Influence Brain Lipids
Brain Lipids Changes in Aging
These PUFA’s are incorporated into membrane phospholipids and therefore play a significant role in structural integrity and function of cell membranes. Lipid metabolism is changed during aging (Montanini et al., 1983; Yehuda et al., 2002; Whelan, 2008; Denis et al., 2015; Cutuli, 2017; Chappus-McCendie et al., 2019), as shown by a decline in omega-3 fatty acids and an increase in lipid peroxidation (Chen et al., 2017). Omega-3 fatty acids have antioxidant properties, and a lack of these fatty acids in one’s diet may accelerate neuronal degeneration (Yehuda et al., 2002; Janssen and Kiliaan, 2014). Susceptibility of lipids to peroxidation increases with age (Bourre, 1991; Spiteller, 2010; Denis et al., 2015; Chen et al., 2017), which supports using the level of oxidative stress as a critical determinant of neuronal health and longevity (Hulbert et al., 2006). Previous studies have suggested that DHA and EPA may protect against peroxidation and the effects of age-related brain pathology (Hasadsri et al., 2013; Chen et al., 2017). Lipids are involved in cellular signaling, energy balance, blood-brain barrier (BBB), and inflammation (Song et al., 2008; Willis et al., 2009), and such age-dependent lipidome changes that disrupt these functions may contribute to neurodegenerative diseases (Arnoldussen et al., 2016; Bos et al., 2016; Hooper et al., 2018; Luo et al., 2018; McNamara et al., 2018; Lepping et al., 2019), such as Alzheimer’s disease (AD) (Schmitt et al., 2014; Hussain et al., 2019).
Lipids and Race/Ethnicity
Race and ethnicity play a significant role in the risk of AD and related disorders. In 2014, nearly 5 million people over the age of 65 had been diagnosed with Alzheimer’s disease or related dementias (ADRD) (Matthews et al., 2019). African Americans and Hispanics had the highest prevalence of ADRD (13.8% and 12.2%, respectively), while ADRD was least common in Asian and Pacific Islanders (8.4%), followed by American Indian/Alaska Natives (9.1%), non-Hispanic whites (10.3%), and people with two or more races (11.5%) (Matthews et al., 2019). Ethnic and racial backgrounds impact many aspects of health, including diet, stress, access to medical treatment, and biological factors of disease. From past research, we can clearly see the ways in which ethnicity, race, and lipids overlap. Most clearly seen in the high incidence of both dyslipidemia, or abnormal amounts of lipids in the blood, and cardiovascular disease observed in minority populations (Frank et al., 2014), race/ethnic disparities affect the regulation of lipid metabolism. Increased concentrations of triglycerides (TG) and decreased levels of lipid carriers, such as HDL-C (high-density lipoprotein-cholesterol) in the blood of Mexican, Filipino, Indian, and Vietnamese people compared to whites may provide a possible explanation for higher risk of both ADRD and cardiovascular disease within these populations (Frank et al., 2014; Gazzola et al., 2017). HDL-C is often referred to as “good cholesterol,” has beneficial antioxidant and anti-inflammatory effects in the body, and has been observed to modulate ß-amyloid (Aß) production in the brain, a key biomarker of AD pathogenesis (Reitz, 2012; Hottman et al., 2014). Lowered levels of HDL-C have been associated with increased cognitive decline and poor cardiovascular health outcomes (Hottman et al., 2014). TG, which is increased in almost every minority population, except African Americans, has been shown to relate to central leptin- and insulin resistance in the brain and decreases in cognition (Sumner, 2009; Frank et al., 2014; Banks et al., 2018). In light of the less marked changes in lipid make-up and metabolism seen in African American populations at increased risk of ADRD, it has been suggested that African Americans are underdiagnosed with metabolic syndromes and vascular-cognitive disorders (Sumner, 2009). Furthermore, it has been observed that there is a differential expression of various molecular biomarkers of AD (phosphorylated tau and total tau) in African Americans compared to whites (Morris et al., 2019), suggesting even small, imperceptible changes in lipid distribution in this population may be sufficient to affect cognition negatively. It is important to note that despite the disproportionate impact ADRD has on minority populations, these individuals continue to be considerably underrepresented in ADRD research, contributing to large gaps in our understanding of brain lipid metabolism as it pertains to race and ethnicity (Gilmore-Bykovskyi et al., 2019).
Lipids and Sex
Sex continues to be one of the largest risk factors for developing AD. Females not only makeup two-thirds of all cases of AD diagnoses but also possess a greater lifetime risk of dementia compared to men due to longer life expectancy (Viña and Lloret, 2010; Mielke, 2018). Increased prevalence and risk of AD and other age-related disorders among females have been attributed to not only extended life expectancy but also to sudden decreases in estrogen post-menopause, among many other factors including education level and mental health status (Viña and Lloret, 2010; Mielke, 2018). Despite the many factors that may contribute to increased risk of AD in women, the contribution of sex-hormone levels and differential lipid distribution play evident roles in cognitive decline are not fully understood. Not only is fat in the form of TG distributed differently in the adipose tissue of male and females, which can be attributed in part to sex-hormone signaling, but concentrations of long-chain PUFAs (LC-PUFAs) have also been observed to be increased in women pre-menopause compared to men (Decsi and Kennedy, 2011; Lohner et al., 2013). Correspondingly, a positive association has been established between omega-3 LC-PUFA biosynthesis, i.e., the production of EPA and DHA, and circulating concentrations of estrogen and progesterone (Childs et al., 2008). Estrogen, an ovarian steroid hormone, is hypothesized to affect lipid metabolism at several points during biosynthesis, including playing a key role in lipid transport and exchange, increasing expression of metabolic enzymes, and reducing the oxidation of α-linoleic acid (ALA), the deriving fatty acid in n-3 LC-PUFA production (Childs et al., 2008; Decsi and Kennedy, 2011; Lohner et al., 2013; Palmisano et al., 2018). Estrogen has also been directly associated with inhibiting memory function impairment in premenopausal women following the surgical removal of their ovaries and loss of the ability to produce estrogen endogenously (Duka et al., 2000; Sherwin, 2012). In a study of trans-sexual subjects, those transitioning from male to female and receiving estrogen observed an increase in DHA plasma levels while those transitioning from female to male and receiving testosterone treatment experienced a marked decrease in plasma DHA (Giltay et al., 2004). The decrease in estrogen levels, as seen in post-menopausal women, has also been associated with increased TG content and lower HDL-C, both of which have been linked to cognitive decline (Derby et al., 2009; Anceline et al., 2014). This is to say, the increased prevalence and risk for AD among women can be explained in part by the abrupt decrease in estrogen production that accompanies the post-menopausal state. Not only does the lack of estrogen decrease concentrations of anti-inflammatory LC-PUFAs and HDL-C in the body, but it also increases TG levels, augmenting secretion of VLDL (very-low density lipoprotein), a lipid carrier known to induce neuroinflammation (Burgess et al., 2006; Chen et al., 2014; Nägga et al., 2018). Additionally, genetic factors, such as ApoE status, and social determinants, such as education, mental illness, and diet, interact with the post-menopausal state to amplify these detrimental effects, increasing risk of AD.
Lipids and Lifestyle
Diet
Dietary lipids play an integral part in physiological lipid metabolism and, consequently, in the risk of AD and cardiovascular disease. Essential fatty acids like DHA (n-3) and AA (n-6) are largely derived from the dietary consumption of their shorter-chained, slightly less-saturated counterparts ALA (n-3) and LA (n-6), respectively (Schmitz, 2008; Morris and Tangney, 2014). After consumption of these deriving fatty acids, the body is able to anabolize them, creating the LC-PUFAs that contribute to neural processes (Morris and Tangney, 2014). Early on in human existence, our diet consisted of an equal balance of n-6 to n-3 essential fatty acids, but as we have evolved, the n-3 to the n-6 ratio of dietary fatty acids has greatly shifted to one side (Simopoulos, 2006). Today, the Western diet has a ratio of about 17 to 1 n-6 to n-3 fatty acids, meaning most Americans have a lot more LA, AA, and DPA in their bodies, which are able to produce relatively large quantities of inflammatory and oxidative mediators (Simopoulos, 2006). Increased ratios of n-6 to n-3 dietary fatty acids have also been directly associated with increased cognitive decline and risk of AD (Loef and Walach, 2013; MacDonald-Wicks et al., 2019). DHA, on the other hand, an n-3 LC-PUFA usually found in fish and algae, is not largely found in the Western diet. Studies suggest, however, that DHA supplementation may work to combat neuroinflammation, oxidative stress, and cognitive decline. Fish oil supplements containing large amounts of DHA, given to older adults with varying levels of cognition, found that supplementation resulted in decreased brain atrophy and less cognitive decline compared to controls in an APOE allele-dependent manner (Daiello et al., 2015). Similarly, Morris et al. observed among subjects over the age of 65 that those who ate fish at least once a week had 60% less risk of AD than those who rarely or never ate fish (Morris et al., 2003). Dietary DHA has also been shown to improve cognition, memory, and brain development from the earliest stages of life through adulthood (Dunstan et al., 2008; McNamara et al., 2010; Muldoonm et al., 2010; Stonehouse et al., 2013; Weiser et al., 2016).
It is important to note that diet can be particularly impacted by race/ethnicity, as well as physical geography, helping to explain differences in AD risk among ethnic groups. According to a global survey of 298 studies, highest levels of DHA and EPA, another n-3 fatty acid, were observed among Japanese, Scandanavian, and indigenous populations, as well as in areas where the Westernized diet had not been fully adopted (Stark et al., 2016). Authors of this survey argue increased consumption of seafood, as dictated by culture or geographical location, greatly impact n-3 LC-PUFA levels in the bloodstream, which offer protective cognitive effects at every stage in life (Joffre et al., 2014; Stark et al., 2016; Weiser et al., 2016).
Genetical Evidence for the Importance of Lipid Metabolism in AD Pathology
Genetic Risk Factors of AD-Related to Lipid Metabolism
Genome-wide Association Studies GWAS and Transcriptome-Wide Association Studies (TWAS) associate AD pathology with several lipid genes (Shi et al., 2010; Hao et al., 2018). While the APOE4 allele carries the greatest risk for AD, other genes and gene-products commonly associated with AD pathology are linked to or interact with lipid metabolism. Several lipid genes associated with AD pathology have recently been reviewed (Tindale et al., 2017). Table 1 is the list of the major genes from GWAS that are linked with lipid metabolism (Jones et al., 2010).
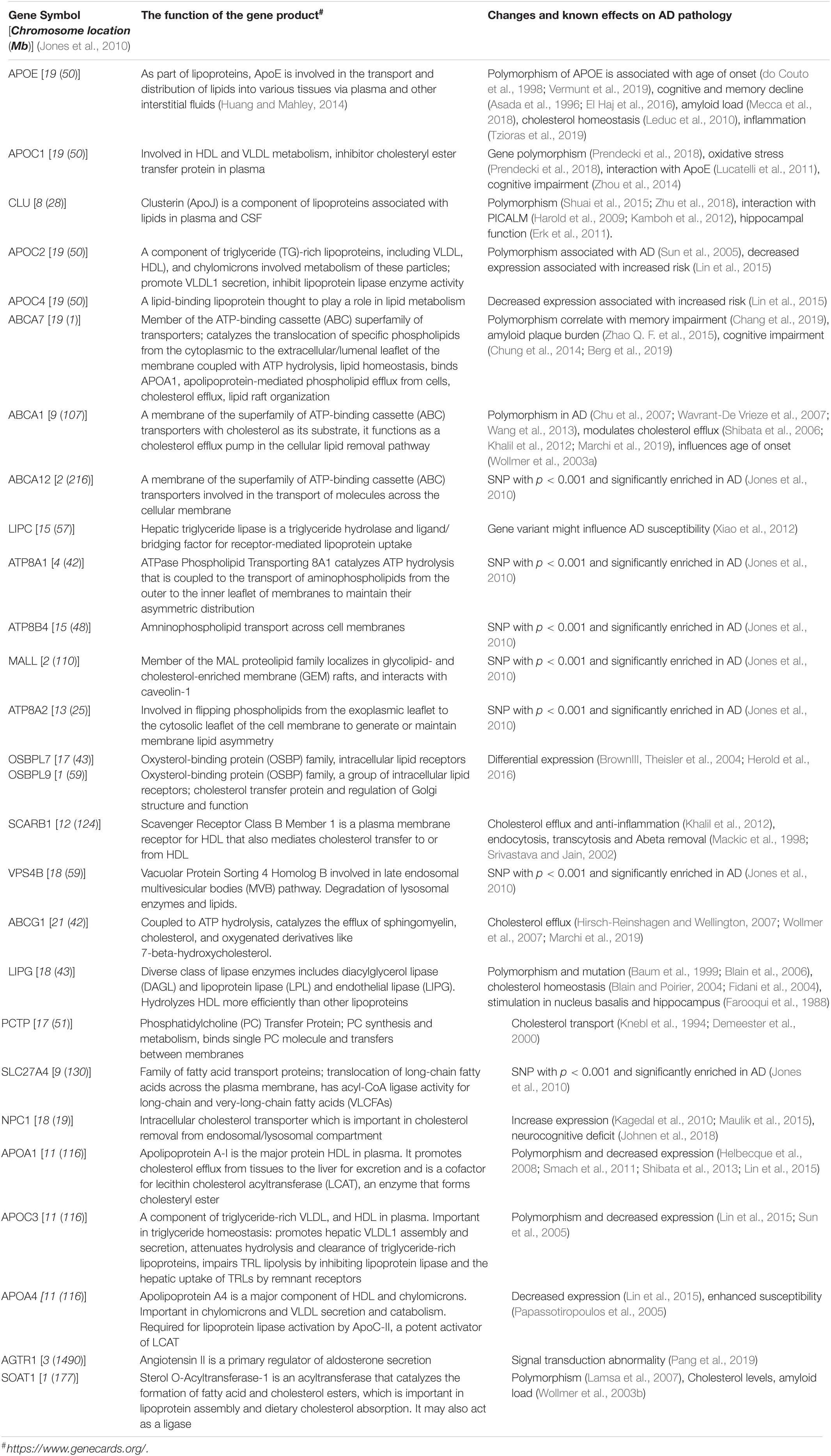
Table 1. Lipid metabolism-associated genes with SNP (<0.001) linked with AD from GWAS.
Genome-wide Association Studies suggest that age-related changes in brain lipid metabolism may be essential to healthy aging and longevity (Tindale et al., 2017). Identification of AD-related genes and how these interact with specific risk factors may provide the rationale for designing effective therapies.
The onset of age related disease can be accelerated with suppression of anti-aging genes, such as Sirtuin 1 (SIRT1). SIRT1 is a histone deacetylase involved with gene expression related to metabolic activity (Grabowska et al., 2017). SIRT1 interacts with lipid metabolism regulation and hepatic oxidative stress and inflammation (Ding et al., 2017). It also regulates circadian rhythms in the liver and brain, maintaining the body’s regulation of glucogenesis, fatty acid beta-oxidation, and cholesterol biosynthesis (Bellet et al., 2016). Its involvement in metabolism explains its effects on energy metabolism, neurogenesis, glucose and cholesterol metabolism, and amyloidosis. Sirt 1 also contributes to neuron apoptosis and survival. Downregulation of this anti-aging gene may lead to acceleration of neurodegenerative disease. Nutritional interventions, such as a reduction in overconsumption of carbohydrates, are recommended because they may be associated with preventing cell senescence and maintaining anti-aging gene activity (Martins et al., 2017). SIRT1 expression promotes APP processing on a non-amyloidogenic pathway and clearance of tau from the brain (Herskovits and Guarente, 2014). SIRT1’s deacetylase activity increases the activity of lysosome-related genes, facilitating Aβ degradation (Li et al., 2018). SIRT1 is a potential therapeutic target for AD because of its involvement in many amyloid beta and cholesterol pathways.
Contribution of Lipids to AD Pathology
Although the brain has a very high concentration of long-chain omega-3 and omega-6 fatty acids, there is no conclusive explanation for how these fatty acids participate in various signaling cascades and in AD (Torres et al., 2014; Mohaibes et al., 2017). However, lipodomic studies related to AD pathology have demonstrated a decrease in DHA levels within the brain, predominantly in the hippocampus (Belkouch et al., 2016). Damage to the hippocampus is associated with impaired learning and memory abilities, a symptom of AD onset (Sarrafpour et al., 2019). With growing evidence that AD is associated with dysregulation of fatty acid metabolism, fatty acid levels may be potential biomarkers of this disease (Fonteh et al., 2014; Wong et al., 2017). In addition to omega fatty acids, the levels of several lipids change with AD pathology (Table 2).
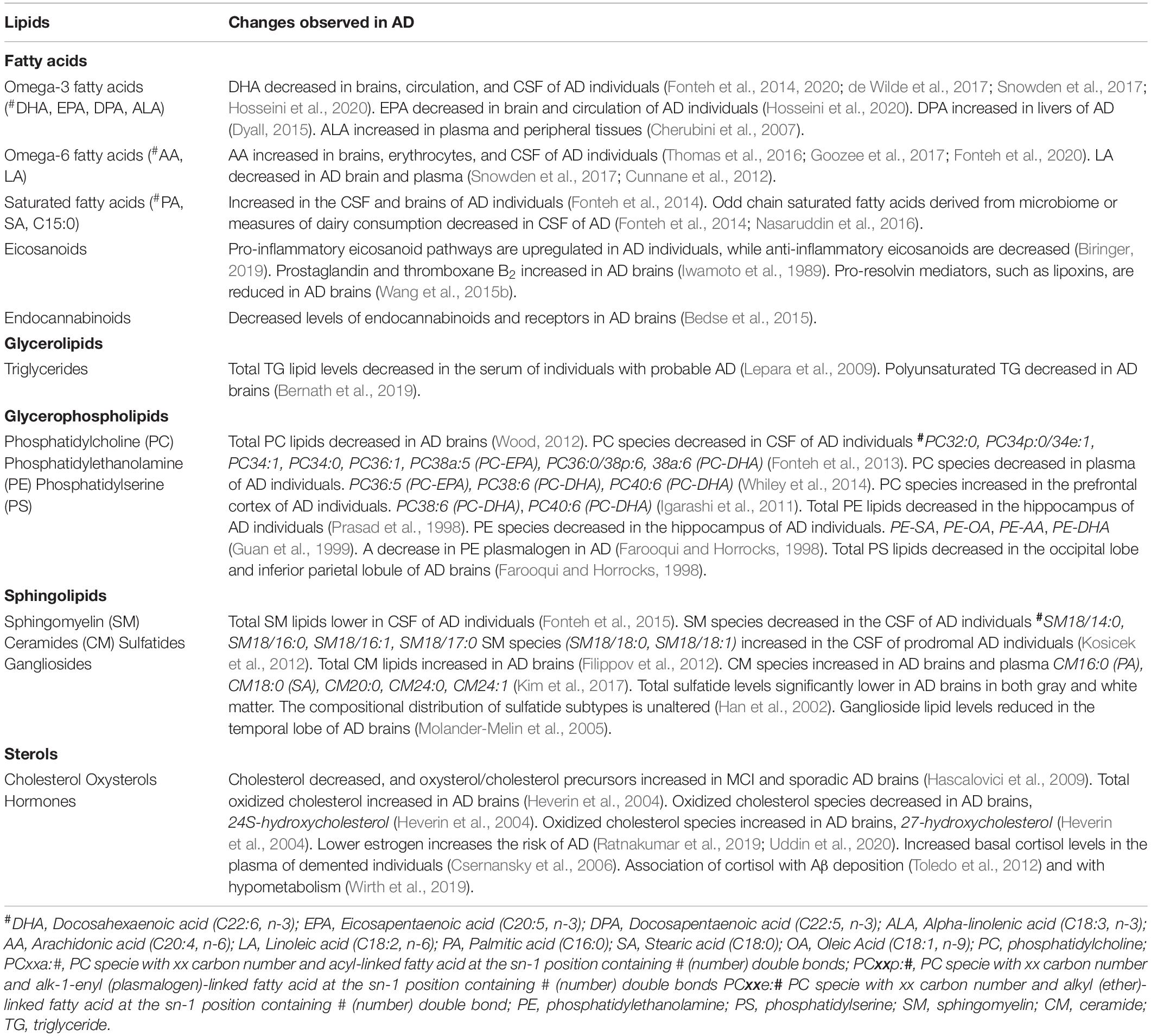
Table 2. Summary of lipids that change in AD.
Lipid Transport: Apolipoproteins
Brain Lipoproteins and Their Function
Lipoproteins are molecules with a hydrophobic lipid core composed of cholesterol, esters, and triglycerides and a hydrophilic exterior of phospholipids, apolipoproteins, and free cholesterol (Alaupovic, 1996; Hoofnagle and Heinecke, 2009; Braun and Hantke, 2019). Lipoproteins assist with the transport of lipids and amphipathic compounds throughout the body (Feingold and Grunfeld, 2000). However, circulating plasma lipoproteins differ from those within the CNS because only high-density lipoproteins (HDL) can cross the blood-brain barrier (Balazs et al., 2004). The most abundant apolipoproteins, apolipoprotein E (ApoE), and apolipoprotein J (ApoJ) are synthesized by astrocytes and serve as enzyme cofactors and receptor ligands on HDL (Pitas et al., 1987; Feingold and Grunfeld, 2000; Ito et al., 2014).
Apolipoproteins are greatly involved in metabolism, serving as both activators and inhibitors of metabolic enzymes, ligands for lipoprotein receptors, and providing structural support (Feingold and Grunfeld, 2000; Bolanos-Garcia and Miguel, 2003; Filou et al., 2016). They also regulate lipid transport by controlling interactions with receptors, enzymes, and lipid-transport proteins (Bolanos-Garcia and Miguel, 2003; Ramasamy, 2014). Apolipoproteins have receptor binding domains containing low-density lipoprotein (LDL) receptors that direct lipid and substrate delivery to specific brain cells (Clavey et al., 1995; Dehouck et al., 1997; Herz, 2001). Their amphipathic-helices facilitate lipid-binding and lipid transport (Clavey et al., 1995; Prevost and Kocher, 1999; Elliott et al., 2010). LDL receptors also facilitate the clearance of amyloid peptides through the BBB (Shibata et al., 2000).
Contribution of Lipoproteins to AD Pathology
Brain lipoproteins with ApoE are responsible for phospholipid and cholesterol transport (Growdon and Hyman, 2014; Wong et al., 2019). ApoE is mainly expressed in astrocytes and microglia and appears as three major isoforms, ApoE2, ApoE3, and ApoE4, of which ApoE4 is the strongest genetic risk factor for AD (Stone et al., 1997; Ito et al., 2005; Vance and Hayashi, 2010; Chung et al., 2016; Liu et al., 2017; Montoliu-Gaya et al., 2018; Tulloch et al., 2018). ApoE4 demonstrates a lower affinity for lipids than ApoE2 and ApoE3, limiting CNS transport of lipids needed for neuronal remodeling and repair (Bradley and Gianturco, 1986; Barbagallo et al., 1998; Li et al., 2002; Frieden et al., 2017). Furthermore, levels of ApoE LDL receptors directly correlate with Aβ clearance, and promoting the expression of these receptors are potential therapeutic targets for AD treatment (Zhao et al., 2018). ApoJ, also known as clusterin, is expressed in astrocytes, neurons, and ependymal cells (Nuutinen et al., 2005, 2007). This neuroprotectant initiates a defense response to neuronal damage and clears Aβ across the BBB via LDLR-2 (Merino-Zamorano et al., 2016; Nelson et al., 2017; Zandl-Lang et al., 2018). ApoJ’s role in Aβ accumulation and toxicity is still undetermined because variability under different contexts and environments confound results (Foster et al., 2019).
Lipids and the Blood-Brain Barrier
The Blood-Brain Barrier
The blood-brain barrier (BBB) is a semipermeable membrane that carefully regulates the exchange of solutes between blood and brain to maintain CNS homeostasis and block entry of toxins and pathogens into the CNS (Bradbury, 1984; Abbott et al., 2010; Betsholtz, 2014; Daneman and Prat, 2015; Ferreira, 2019; Moura et al., 2019). The integrity of the BBB is largely dependent on its tight junctions (Brown and Davis, 2002; Castro Dias et al., 2019), adherens junction proteins, and ability to control the vesicular movement of macromolecules through transcytosis and pinocytosis (Dehouck et al., 1997; Baldo et al., 2014). The BBB permits free diffusion of gases, such as oxygen and carbon dioxide, but small solutes such as lipophilic molecules and ions enter through receptor-mediated transcytosis or via channels (Fishman et al., 1987; Zlokovic, 2008; Preston et al., 2014; Andreone et al., 2017; Villasenor et al., 2017; Ayloo and Gu, 2019). The BBB is critical in linking multiple major organ systems, and any dysfunction in the lipid bilayer’s ability to act as a barrier may lead to neuronal degeneration (Zhao Z. et al., 2015; Halliday et al., 2016; Muszynski et al., 2017; Nation et al., 2019).
Importance of Lipids in BBB Function
In addition to composing the BBB lipid bilayer, lipids, including phospholipids, sphingolipids, and cholesterol, also compose the plasma membrane of vesicles involved with receptor-mediated transcytosis within the CNS (Kramer et al., 2002; Dodelet-Devillers et al., 2009; Campbell et al., 2014; Andreone et al., 2017). The formation and function of vesicles required to transport essential macromolecules across the BBB may be affected by the plasma membrane lipid composition (Lingwood et al., 2009; Lingwood and Simons, 2010; Kaiser et al., 2011). In particular, DHA disrupts the membrane domains necessary to form such transport vesicles and therefore contributes to BBB integrity and suppression of transcytosis (Ouellet et al., 2009; Freund Levi et al., 2014; Pan et al., 2015, 2016; Belayev et al., 2018). There is also recent evidence that the membrane transport protein, Mfsd2a, controls lipid exchange and plays a key role in the transport of DHA into the brain, though this pathway is largely undetermined (Segi-Nishida, 2014; Zhao and Zlokovic, 2014; Keaney and Campbell, 2015; Andreone et al., 2017). Loss of Msfd2a transport function resulted in decreased DHA transport and increased activity levels of transcytosis within CNS endothelial cells (Andreone et al., 2017). A leaky barrier increases the brain’s susceptibility to toxins and pathogens and homeostasis disruption, and ultimately, neuronal dysfunction (Abbott, 2000; Hutchinson, 2010; Ikeshima-Kataoka and Yasui, 2016; Block, 2019).
The Contribution of the BBB to AD Pathology
Loss of BBB function may contribute to neurodegenerative diseases, including AD (Banks, 1999; Gilgun-Sherki et al., 2001; Zlokovic, 2008; Carvey et al., 2009; Karamanos et al., 2014; Sweeney et al., 2018; Katt et al., 2019). According to multiple independent studies, BBB breakdown in AD is demonstrated by decreased integrity of BBB tight junctions, pericyte and endothelial degeneration, RBC extravasation, and brain capillary leakages (Zlokovic, 2008; Carvey et al., 2009; de Vries et al., 2012; Nelson et al., 2016; Sweeney et al., 2018). A buildup of blood proteins and macromolecules due to barrier leakiness may damage vasculature and brain parenchyma, which induces neuronal degeneration. Studies have also indicated that AD pathology includes reduced expression of glucose transporters in the BBB (Kalaria and Harik, 1989; Harik and Kalaria, 1991; Guo et al., 2005; Agrawal et al., 2017; Block, 2019). This may exacerbate AD cerebrovascular degeneration and cognitive function, considering that the brain requires a continuous supply of glucose and utilizes the most glucose of the major organs (Benton et al., 1996; Dienel et al., 1997; Benton, 2001; Gong et al., 2006). The BBB contains a wide variety of structural components to regulate the brain’s health and function, but a loss of function in any such component may lead to dyshomeostasis and a rapid cascade of dysfunctions in other structures within the brain.
Lipids Contribute to Amyloid Precursor Protein Processing
Amyloid Precursor Protein Processing
Amyloid precursor protein (APP) is a type I transmembrane protein that is cleaved into amyloid β-peptide (Aβ) by β- and γ-secretases (Nunan and Small, 2000; Hartmann, 2012). APP is synthesized in the endoplasmic reticulum and is found in the highest concentrations in neuron’s trans-Golgi-network, suggesting that APP is associated with secretory pathways (Palacios et al., 1992; Stephens and Austen, 1996; Kitazume et al., 2001; Tam et al., 2014; Toh et al., 2017; Liu et al., 2019). There are two accepted proteolytic pathways for APP processing − non-amyloidogenic and amyloidogenic (Ishiura, 1991; Kojima and Omori, 1992; Sisodia, 1992; Roberts et al., 1994; Mills and Reiner, 1999; Soriano et al., 2001; Irizarry et al., 2004; Song et al., 2004; Chow et al., 2010; Wang et al., 2010; Tomita and Wong, 2011). The non-amyloidogenic pathway involves cleavage of APP by α-secretase at the plasma membrane, releasing soluble APPα (sAPPα) fragments into the extracellular environment, and normalizes AG genes and memory (Volmar et al., 2017). The amyloidogenic pathway involves cleavage of APP by β-secretase in early endosomes, releasing sAPPβ fragments in the endosomal lumen, and increasing susceptibility to Aβ plaques that are relevant to AD pathology (Estus et al., 1992; Golde et al., 1992; Saftig et al., 1996; Ehehalt et al., 2003; Andrew et al., 2016; Grimm et al., 2016).
The Role of Lipids in APP Processing
The β-site APP-cleaving enzyme 1 (BACE-1) is the major β-secretase that targets endosomes with APP in transit to endocytosis sites on the plasma membrane (Shimokawa et al., 1993; O’Brien and Wong, 2011; Chun et al., 2015; Audagnotto et al., 2018). Both APP and BACE-1 are associated with lipid rafts, which are membrane domains enriched with cholesterol, sphingolipids, and gangliosides that are crucial to vesicle trafficking and intracellular transport (Ehehalt et al., 2003; Yoon et al., 2007; Marquer et al., 2011; Bhattacharyya et al., 2013). Recent studies have proposed that BACE-1 in cholesterol depleted environments displayed inhibited β-secretase activity, suggesting that cholesterol and lipid composition of the intracellular environment may be a large determinant of whether BACE-1 can access APP endosomes (Dash and Moore, 1993; Cheng et al., 2014; Mukadam et al., 2018). However, other studies suggest that both homeostasis of lipid composition and oxidation state of lipids, including DHA, are critical to APP processing (Grimm et al., 2012; Bhattacharyya et al., 2013; Figure 2). Under conditions with high concentrations of oxidized lipids, levels of sAPPα fragments decreased while sAPPβ levels increased (Grimm et al., 2016). A novel mechanism of proteolytic activity regulation of secretases involves a separating lipid boundary with their substrates, APP (Kaether and Haass, 2004). Lipid mediators of inflammation also interact with APP processing at the level of O-GlcNAcylation (Sastre et al., 2008; Jean-Louis et al., 2018). Thus, oxidized or inflammatory lipids may shift APP processing from the non-amyloidogenic to an amyloidogenic pathway (Figure 2).
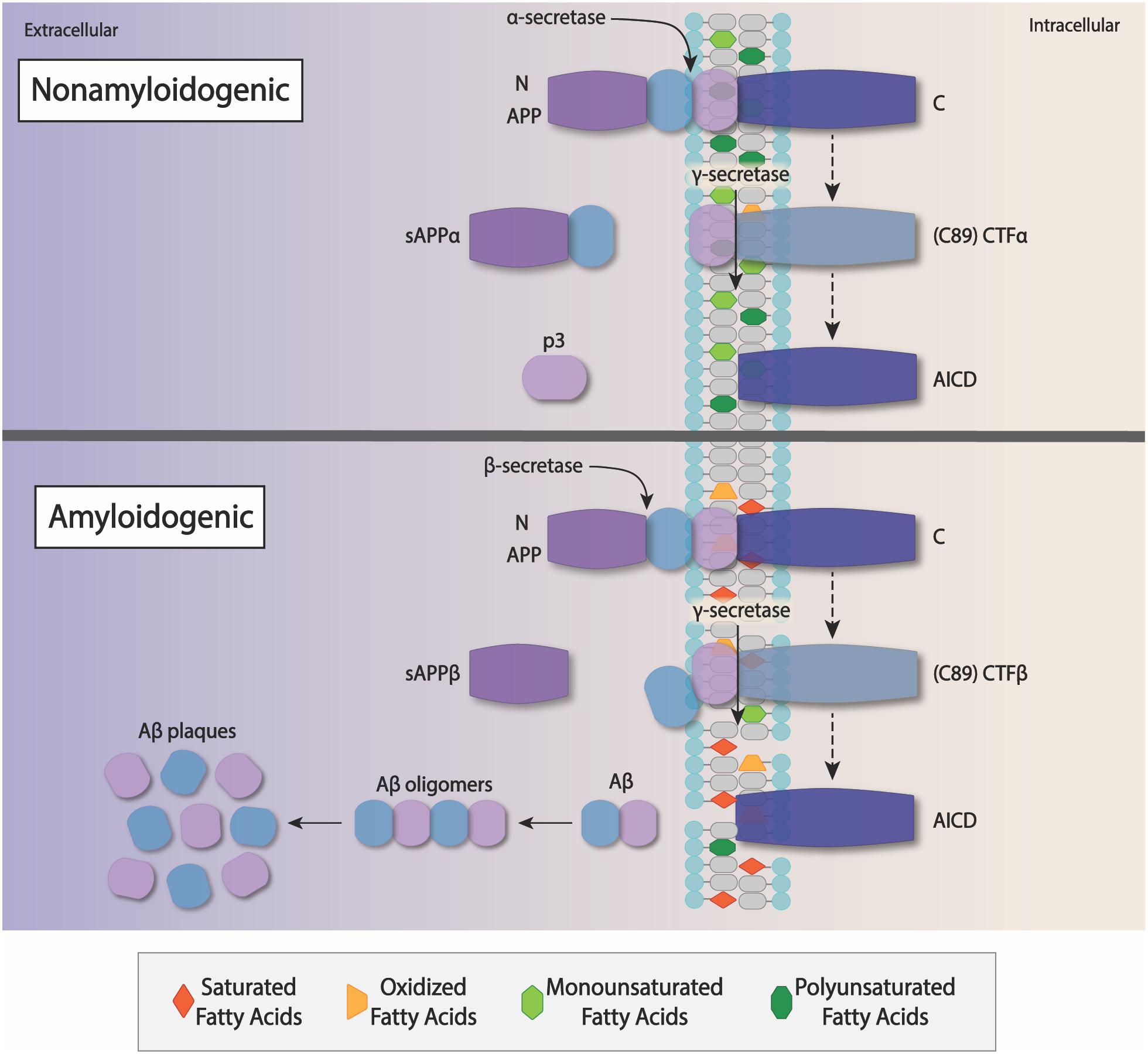
Figure 2. The importance of Lipids on APP processing – APP is a transmembrane protein that is cleaved by several proteases: α-secretase, β-secretases, and γ-secretases. Non-amyloidogenic processing of APP− In a cell with a membrane containing normal or high amounts of unsaturated fatty acids, especially DHA, preference is given to cleavage by α-secretase In this case, a well-structured membrane holds onto an intact APP as it is cleaved by the α-secretase and subsequently the α-secretase releasing the secreted ectodomain sAPPα, along with a small protein fragment, p3, and APP intracellular C-terminal domain (AICD) peptide in the extracellular space. sAPPα and p3 do not form neurotoxic fibrils and plaques, and so this process is referred to as non-amyloidogenic APP processing. Amyloidogenic processing of APP – In contrast, PUFA enriched structure of healthy neurons, the presence of saturated and oxidized fatty acids results in the disruption of the cell membrane structure, and this favors β-secretase activation. APP is cleaved at its’ N-terminus by β-secretase, releasing a soluble ectodomain sAPPβ into the extracellular space. γ-secretase subsequently cleaves the cell-associated C-terminus releasing and Aβ peptides of varying lengths into the extracellular space. Insoluble Aβ fibrils aggregate as oligomers that ultimately clump to form plaques within the brain. These plaques contribute to oxidative stress, neuroinflammation, and eventually decreased brain function.
The Intersection of Lipids, APP Processing, and AD Pathology
The Aβ fragments of APP is the major component of AD amyloid plaques, and such dysregulation of APP trafficking and processing are relevant to understanding AD pathology (Caporaso et al., 1994; Thinakaran and Koo, 2008; Zhang et al., 2011; Tan and Gleeson, 2019; Yuksel and Tacal, 2019). Intracellular Aβ accumulation in neurons of patients with AD and metabolic analysis of brain function indicate a possible dysfunction in Aβ transport exiting the brain (O’Brien and Wong, 2011; Yuksel and Tacal, 2019). Lipids rafts play important roles in APP trafficking (Yoon et al., 2007; Yang et al., 2013). Moreover, palmitoylation dictates how APP is processed (Bhattacharyya et al., 2013). Trans fatty acids influence amylogenic APP processing, while the level of fatty acid unsaturation determines the activity of secretases (Yang et al., 2011; Grimm et al., 2012). Future research relating to changes in brain lipid composition in pre-symptomatic AD may provide a link with early disease onset, dysregulation of lipid metabolism, and APP processing.
The Intersection of Lipid Rafts, APP Processing, and AD Pathology
Lipid rafts are dynamic clusters of membrane lipids that interact with protein complexes to promote intracellular signal transduction (Mesa-Herrera et al., 2019). Normal aging is associated with gradual reductions in cholesterol and polyunsaturated fatty acids (PUFAs) in lipid rafts. With age-related changes lipid rafts composition, alterations in intracellular communication may be associated with age-associated reductions in synaptic plasticity. In neurodegenerative diseases, the composition of lipid rafts changes more rapidly, most notably in n-3 and n-6 PUFAs (Li et al., 2018). Lipid raft aging appears to be exacerbated in Alzheimer’s Disease, which may serve as the underlying contribution to disrupted signal transduction, increased APP processing, and rapid formation of AB aggregates (Grassi et al., 2019). Normal APP signal transduction involves cleaving APP into AB into the extracellular environment. However, if APP interacts with ApoE and tau on a lipid raft with an atypical lipid composition, signal transduction may be disrupted, promoting the formation of AB aggregates. Other alterations include reductions in unsaturation of FA in AD patients, as compared to controls (Kao et al., 2020). Lipid raft aging also appears to exhibit gender differences, such that women had more severe changes in lipid raft composition as compared to men. This may serve as supportive evidence for the finding that postmenopausal women are more likely to progress from MCI to AD than age-matchd men (Herrera). Considering that lipid raft function is sensitive to aging, further characterization of composition changes in lipid rafts within the brain may be useful as a biomarker of neurodegenerative stages.
Lipids and Cellular Remodeling
Role of Lipid Remodeling in Synaptogenesis
Lipid bodies (LBs) are spherical lipid-rich organelles associated with lipid storage, metabolism, cell signaling, and inflammation (Schmitz and Muller, 1991; Melo et al., 2011). At regulated levels, LBs maintain lipid homeostasis and cellular function, but in response to brain inflammation and increased neuronal oxidative stress, these LBs grow in size and accumulate within microglial cells (Tremblay et al., 2016; Hu et al., 2017). Though the pathway is still largely undiscovered, LBs in microglia appear to communicate with organelles such as the mitochondria, which control cell-death mechanisms (Tyurina et al., 2014). When exposed to lipopolysaccharides, LBs contact to mitochondria was disrupted, but DHA treatment reduced such effects. DHA may be a key factor in preserving mitochondrial health and regulation of microglial activity (Tremblay et al., 2016; Maysinger et al., 2018). When regulated in rodent models of AD, microglia slows the accumulation of Aβ plaques, but a proliferation of microglia activity may result in brain inflammation and degradation of neuronal synapses (Lim et al., 2000; Stahl et al., 2006; McClean et al., 2015). Microglial dysfunction has been implicated as a contributor to AD pathogenesis (Hansen et al., 2018). Microglia cells in the brain contribute to the reorganization of neuronal circuits by phagocytosing dead neurons and their dendritic spines and axon terminals. These immune cells contribute to neural plasticity (Wu and Zhuo, 2008; Yates, 2014; Yang et al., 2019), which refers to the brain’s ability to maintain, modify, and strengthen these synapses in order to permit neuronal communication (Tremblay et al., 2011).
Importance of Lipid Remodeling/Synaptogenesis in AD Pathology
Synaptogenesis is the formation of nerve synapses involving the reorganization of cell structural components (Aoki et al., 2003; Kelsch et al., 2010). Several studies suggest that presynaptic and postsynaptic development is initiated by signaling pathways involving cholesterol (Mauch et al., 2001; Fester et al., 2009). Changes in fatty acid content occur prior to synaptogenesis in cones (Martin and Bazan, 1992). Studies have shown that neurons deprived of lipid rafts underwent a cascade of effects inhibiting synaptic growth and development (Bazan, 2005; Welberg, 2014; Mochel, 2018). Depletion of lipid rafts decreased dendritic density and increased the synapse, disrupting neuronal communication (Martin, 2000; Hering et al., 2003; Sebastiao et al., 2013; Wang, 2014). The transport protein, apolipoprotein E (apoE), monitors cholesterol transport from glial cells to neurons, and impaired ApoE is implicated in deficits in synaptic plasticity and cognitive function (Periyasamy et al., 2017). Of the three isoforms of ApoE, ApoE4 is a prevalent risk factor that is synergistic with obesity and age for AD (Butler, 1994; Riedel et al., 2016; Jones and Rebeck, 2018; O’Donoghue et al., 2018; Glorioso et al., 2019). ApoE4 binds fewer lipids and is most likely involved in changes in cholesterol flux and metabolism (de Chaves and Narayanaswami, 2008; van den Kommer et al., 2012; Mahley, 2016; Nunes et al., 2018), accounting for altered synaptogenesis and neural plasticity.
Lipids and Myelination
The Importance of Myelination
Action potentials propagate along axons through rapid saltatory conduction. Synthesized by oligodendrocytes in the CNS and Schwann glial cells in the PNS, myelin membranes act as electrical insulators, permitting higher nerve conduction velocities and greater neuronal communication efficiency (Almeida and Lyons, 2014; Almeida and Lyons, 2017). Without myelin, axons would require more energy to depolarize its membrane (Stassart et al., 2018). Myelin is composed of several lipids and protein layers that wrap around most of the axon, except at nodes of Ranvier, which are regions highly concentrated with sodium ion channels (Finean and Robertson, 1958; Davison, 1972; Burgisser et al., 1986; Wender et al., 1988; Ando et al., 2003; Schmitt et al., 2015; Montani and Suter, 2018). Myelination of axons is a dynamic process through development and adulthood, and this process, in addition to myelin sheath modification and myelin repair, contributes to synaptic remodeling and neural plasticity (Zatorre et al., 2012).
The Role of Lipids in Myelination
The myelin membrane consists of myelin-specific proteins and high-level synthesis of lipids representative of all major classes, such as cholesterol, glycosphingolipids, glycerophospholipids, and galactolipids (Chrast et al., 2011). Lipids comprise approximately 80% of myelin’s dry weight, accounting for glia’s high demand for fatty acids, which are fundamental building blocks of its lipid structure (Dimas et al., 2019). Myelin accounts for a majority of the white matter in the brain, which is consistent with reported reduced myelin density associated with AD white matter changes in the brain (Nasrabady et al., 2018).
Brain Myelination and AD Pathology
Reduced number and activity of oligodendrocytes and precursor cells can damage myelin integrity, contributing to AD pathology’s characteristic neuronal loss (Bartzokis, 2011). Oligodendrocytes support and regulate neurons, but they are primarily responsible for myelin production (Simons and Nave, 2015). Myelinating oligodendrocytes are sensitive to lipid peroxidation because oxidative stress inhibits expression of genes that promote oligodendrocyte differentiation (French et al., 2009). This implies that disruption of myelin synthesis may be a central feature of AD pathology, and can be exploited for therapy (Desai et al., 2010). Dysfunction in these processes may be linked to white matter abnormalities and cognitive impairment associated with AD due to damaged signal conductivity and synchronicity needed for information processing between neurons (Ihara et al., 2010; Alexander, 2017; Nasrabady et al., 2018). The causal relationship between myelination and AD has not been elucidated, but white matter changes arising from myelination dysfunctions have been described in AD brains (Kohama et al., 2012). Additional evidence for the contribution of myelin breakdown on AD pathology comes from studies showing that the rate and severity of myelin breakdown in healthy seniors are associated with APOE status, a major risk factor of AD (Bartzokis et al., 2006).
Lipids and Receptor-Mediated Signaling
Neuronal Receptor Signaling Pathways
Neurons communicate via electrochemical signals and neurotransmitters across gaps called synapses associated with several integrated networks (Mayer, 1993; Laughlin and Sejnowski, 2003; Salinas, 2009; Hahn et al., 2019). The presynaptic neuron releases neurotransmitters through exocytosis, and those chemicals bind to the postsynaptic neuron’s neurotransmitter receptors to alter postsynaptic neuronal activity (Kennedy, 2013). One class of neurotransmitter receptors, called ligand-gated ion channel receptors, opens an ion pore through the membrane upon ligand binding. Ions cannot travel through the hydrophobic lipid membrane and, therefore, can only pass through channels controlled by these receptors. Ions entering the ligand-gated channel can initiate excitatory or inhibitory signals, but both rapidly influence neuronal function (Cantor, 2018). Another class of neurotransmitter receptors, G-protein-coupled receptors (GPCRs), bind to the ligand and initiate an intracellular mechanism in which its G-proteins alter cAMP levels to stimulate or inhibit the neuron, and may involve lipid agonists (Hansen, 2015). Unlike ligand-gated ion channel receptors, GPCRs are slower but longer-lasting in affecting neuronal activity (Lovinger, 2008).
Role of Lipids in Neuronal Signaling
While cascades of protein kinases and phosphatases have been largely studied, there is an increasing interest in lipid-based pathways involving lipid kinases and phosphatases. Lipids are versatile in signal transduction pathways and act as hormones, ligands, substrates, and mediators (Eyster, 2007; Piomelli et al., 2007; Piomelli, 2012). Sphingolipids and cholesterol comprise lipid rafts, which are regions in the plasma membrane that organize signaling molecules, amplify intracellular signaling cascades, and regulate both neurotransmission and membrane protein trafficking (Levental and Veatch, 2016). Additionally, lipids are integral to GPCR signaling cascades. Following GPCR binding, phospholipase C (PLC) cleaves the polar phosphate head of phospholipids and forms diacylglycerol (DAG), a lipid second messenger (Black et al., 2016). Fatty acids (FAs), especially those belonging to the omega-3 and omega-6 classes, act as ligands for membrane receptors in a variety of pathways (Mobraten et al., 2013). The wide diversity of lipids and their structures contributes to AD, and their multiple roles in signal transduction may influence AD pathology.
Signaling Lipids Contribute to AD Pathology
Endocannabinoid signaling is responsible for inhibition and excitation in modulating synaptic strength, implicating its possible role in AD and associated inflammatory pathology (Skaper and Di Marzo, 2012). Although the mechanism has not been elucidated yet, free radicals and oxidative stress increase GPCR cannabinoid 2 receptors (CB2) expression in AD microglial cells, increasing neuroinflammation (Paloczi et al., 2018). Inflammation protects the brain against neurotoxins, but excessive inflammation may contribute to neurodegeneration. Another study suggested that monoacylglycerol lipase (MAGL) produces neuroinflammatory prostaglandins through the hydrolysis of endocannabinoids (Piro et al., 2012). Inhibiting MAGL activity is a potential AD therapeutic target because it is reported to prevent neuroinflammation, neurodegeneration, and impaired synaptic plasticity (Chen et al., 2012). Dysregulation in neuronal signaling cascades may contribute to increased susceptibility to neuronal dysfunction and are, therefore, important in studying its effects and relation to AD.
Lipids and Inflammation
The Importance of Inflammation
Inflammation is a defense mechanism initiated by the immune system in response to pathogens, injured cells, infections, and other toxic stimuli. A signaling cascade results in leukocyte migration to damaged sites, in which released cytokines recruit other immune cells to heal injured tissue (Robinson et al., 2018). Specifically, within the CNS, activation of microglia and its associated cytokine production are primarily responsible for the inflammatory responses (Frank et al., 2007; Ghosh et al., 2012; Zhu et al., 2019). However, unregulated inflammation, excessive cytokine production, and failure to resolve inflammatory responses all contribute to chronic neuroinflammation, a biomarker of many neurodegenerative diseases, including AD (Wang et al., 2015a).
Lipids and Inflammation
Several studies implicate the role of lipids and lipid metabolism in inflammatory responses (Janciauskiene and Wright, 1998; Kang and Rivest, 2012; Zhang et al., 2018; Ntambi, 2019). Eicosanoids are a class of lipid mediators inflammation produced by innate immune cells that contribute to acute inflammation, resulting in pain, loss of function, heat, and swelling (Higgs et al., 1984; Williams and Higgs, 1988; Hedqvist et al., 1991; Umamaheswaran et al., 2018). Following the elimination of toxic stimuli, innate immune cells cease the production of eicosanoids and begin production of specialized pro-resolving lipid mediators (SPMs) to resolve inflammation (Serhan, 2010; Chandrasekharan and Sharma-Walia, 2015; Chiurchiu et al., 2018; Maclean et al., 2018). Synthesized from omega-3 fatty acids, docosahexaenoic acid (DHA), and eicosapentaenoic acid (EPA), SPMs resolve inflammation by inhibiting polymorphonuclear leukocytes (PMN) and lowering vascular permeability This process may be impaired in AD (Whittington et al., 2017).
Inflammatory Lipids and AD Pathology
A disproportionate level of inflammation can disrupt the balance between eicosanoids and SPMs, overwhelming the brain’s ability to return to a non-inflammatory state. This suggests the brain’s dependence on SPMs and its omega-3 precursors, DHA, and EPA (Serhan et al., 2018). AD pathology includes decreased DHA levels (Fonteh et al., 2014; Yassine et al., 2017), which may account for heightened brain inflammation that leads to declining cognitive health. Moreover, many studies have reported alterations to the eicosanoid pathway in AD (Biringer, 2019), further heightening research interest in the balance between eicosanoids and SPMs (Serhan et al., 2015). AD is also associated with elevated microglia-induced neuroinflammation, increases in proinflammatory cytokines, and upregulated expression of phagocytic receptors in white matter microglia (Zheng et al., 2016). One receptor, CD36, promotes both pro-inflammatory and oxidative pathways upon binding to ligands, including lipids and Aβ (Doens et al., 2017). Overexpression may lead to dysregulated inflammation and increased oxidative stress, a biomarker of the inflammatory response, and AD (Park et al., 2014; Koizumi et al., 2016). White matter is critical to neuronal connectivity and processing speed, and such white matter inflammation may result in neurodegeneration and, therefore, the cognitive decline (Raj et al., 2017). Further studies aim to determine if inflammation contributes to the onset of AD or exacerbates already-existing neuropathology (Heppner et al., 2015).
Lipids and Oxidative Stress
Oxidative Stress
Oxidative stress is defined as a disruption in homeostasis between antioxidants and oxidants, and more specifically, an accumulation of reactive oxidative species (ROS) and reactive nitrogen species (RNS) (Apak et al., 2016; Hameister et al., 2020). ROS belongs to a family of compounds containing partially reduced oxygen species, such as O2– and HO-, that are generated primarily by the electron transport chain during aerobic respiration (Zhao et al., 2019). ROS are involved in many redox-dependent processes, including cell signaling, homeostasis, immune system responses, energy metabolism, and tissue remodeling. However, an excess of ROS or impaired control of the balance between antioxidants and oxidants leads to oxidative stress, which is implicated in the progression of neurodegenerative diseases (Cheignon et al., 2017). Because the brain consumes approximately 25% of the body’s glucose, its high energy consumption increases neurons’ susceptibility to oxidative stress and overproduction of ROS (Wezyk et al., 2018).
Membrane Lipids Are Damaged During Oxidative Stress
Excess ROS can lead to increased lipid peroxidation within the brain, altering membrane permeability and activity of membrane receptors and their associated enzymes (Birben et al., 2012). Lipid peroxidation produces reactive aldehydes, including malondialdehyde (MDA) and 4-hydroxynonenal (HNE), that modify and bind to proteins involved in metabolism, antioxidant defense systems, and axonal growth. By modifying Tau protein, 4-HNE can indirectly lead to an increase in neurofibrillary tangles, which is consistent with proteomic reports of increased 4-HNE in AD hippocampal tissue and neurofibrillary tangles (Cheignon et al., 2018). Moreover, low-density lipid lipoprotein receptor-related protein (LRP1) is involved in Aβ peptide removal. LRP1 is oxidized by Aβ, inhibiting its ability to clear Aβ and therefore leading to Aβ accumulation in the brain (Shinohara et al., 2017). LRP1 is another protein that is covalently modified by 4-HNE, further supporting that unrestrained lipid peroxidation produces excess reactive products that initiate a cascade of dysregulations within pathways necessary to neuronal health (Butterfield et al., 2002). Oxidant/antioxidant imbalance forms blood-based biomarkers that can be used for early, non-invasive diagnosis (Wojsiat et al., 2018), or for AD therapies (Yatin et al., 2000; Sultana et al., 2004).
Oxidative Stress and AD Pathology
Many trials seek to assess different antioxidant therapeutic approaches to alleviate oxidative stress, a key biomarker of AD. CoQ10, creatine, idebenone, latrepirdine, triterpenoids, omega-3 PUFAs, vitamin E, and vitamin C are just a few antioxidants that are extensively studied in their treatment of neurodegenerative diseases (Yatin et al., 2000; Kumar and Singh, 2015).
Lipids and Immune Response
The Immune System
The immune system, which is divided into the innate and adaptive immune system, is critical to defending the body against infectious and toxic stimuli (Simon et al., 2015). The innate immune system utilizes cytokine production and modulation to mount a quick but sufficient response to pathogens, including viruses, bacteria, and parasites. The innate immune system is also responsible for activating the adaptive immune system, which is slower due to the lengthy production of specific antibodies to the foreign antigen (Iwasaki and Medzhitov, 2015). Studies in the past 20 years have refuted the notion of the brain as being “immunologically privileged” in relying largely on innate immune system mechanisms within the CNS. While it was thought that the CNS and immune system were separate due to the blood-brain barrier, the detection of lymphatic vessels connecting T-cells in lymph nodes to cerebrospinal fluid (CSF) in the meninges provided evidence for the brain’s semi-dependence on the adaptive immune system (Louveau et al., 2015). Neuroimmune processes are activated by vagal nerve signaling, immune signals, and complement proteins, resulting in increased activity of microglia and astrocytes (Tchessalova et al., 2018).
Lipids and Immunity
Studies reported increased levels of platelets and vascular lesions in AD patients outside of the brain, contributing to cerebral amyloid angiopathy, a biomarker of AD that shows increased amyloid protein in the brain arteries (Kniewallner et al., 2015). Although platelets combat vascular injury, they are also involved in APP processing, and transitively, the formation of Aβ plaques (Evin et al., 2003; Evin and Li, 2012). The balance of omega-3 and omega-6 PUFAs may affect platelet levels, as membrane essential fatty acids (EFAs), primarily DHA and EPA, form prostaglandins PGE1, PGE2, and PGE3, all of which participate in a variety of immunological and signaling pathways in the brain (Chang et al., 2009). PGE1 has anti-inflammatory properties, and conversely, PGE2 strongly promotes inflammation by acting on different receptors (Iyu et al., 2011). PGE3 is responsible for regulating PGE2’s inflammatory effects by competing with its formation from precursor EFAs (Chang et al., 2009). Imbalances in the omega-6 to omega-3 PUFA ratios disrupt the formation of PGE3, which minimizes the regulation of PGE2 induced inflammation. Moreover, this imbalance of PUFAs is associated with changes in neuronal brain composition that, in combination with drug therapies, can reduce the risks and slow the progression of AD (Giulietti et al., 2016).
Immunity and AD Pathology
An impaired BBB is implicated with the onset of AD, which may increase the BBB’s permeability to pathogens and immune cells (Veerhuis et al., 2011). Levels of cytotoxic and helper T-cells are upregulated in brain parenchyma of AD patients (Oberstein et al., 2018). Helper T cells and pro-inflammatory cytokines target neurofibrillary tangles and plaques composed of Aβ and Tau and activate microglia at these sites, further exacerbating neuroinflammation (Gold et al., 2014; Martinez-Frailes et al., 2019). One class of cytokines, called chemokines, stimulates leukocyte migration from blood to tissues. CCL5 is a chemokine that is amplified in response to reactive oxygen species and oxidative stress within the brain’s endothelial cells, promoting even more T cell migration across the leaky BBB. These inflammatory mediators are elevated in the CSF and blood and are possible biomarkers for detecting AD and its progression (Mietelska-Porowska and Wojda, 2017).
Lipids and Energy Regulation
Sources of Brain Energy
Although the human brain comprises only 2% of the body weight, it consumes approximately 20% of glucose, demonstrating its disproportionately high energy demand (Mergenthaler et al., 2013). The majority of the energy utilized by the brain is dedicated to returning neurons to their resting states after depolarization, and the remaining 20−25% of energy is allocated toward synthesizing vesicles and neurotransmitters (Harris et al., 2012). The brain relies on a constant flow of glucose and oxygen, which are delivered through the blood. However, during fasting periods, when glucose levels are decreased, the liver can supply ketone bodies to support metabolism within the brain (Patel et al., 1975; Hawkins and Biebuyck, 1979; Nehlig, 2004). These delivered ketone bodies are primarily utilized by astrocytes, and upon arrival, ketolysis of the ketone bodies generates acetyl CoA, an important substrate for the tricarboxylic acid (TCA) cycle and therefore, ATP production. Although the brain has a large ATP requirement, it does not utilize these ketone bodies or fatty acids as a significant source of energy like in other organs, such as the liver. It is hypothesized that evolution selected against this pathway because it produces ROS and therefore, contributes to oxidative stress that contributes to neurodegeneration (Schonfeld and Reiser, 2017).
Role of Brain Energy Regulation in AD Pathology
Transport and utilization of glucose within the brain are tightly regulated, but mitochondrial dysfunction and decreased expression of glucose transporters (GLUT) are potential contributors to AD (Yin et al., 2016). Highly concentrated in the BBB, GLUT1 transports glucose across the endothelium and into astrocytes, whereas GLUT3 is predominantly found in axons and dendrites, underscoring its role in neuronal glucose transport and distribution (Vannucci et al., 1998). Reduced GLUTs expression at the BBB and within neurons is associated with AD, which may explain overall decreased glucose metabolism in AD pathology (Yin et al., 2016).
Mitochondrial Dysfunction and AD Pathology
Mitochondria are organelles central to brain energy processes, and altering glucose availability or dysregulating oxidative phosphorylation can have direct effects on neuronal function and cognitive health (Picard and McEwen, 2014; Anderson, 2018). Recent reports have hypothesized that Aβ may initiate mitochondrial dysfunction, and one theory proposes that Aβ raises cytosolic calcium levels, inhibiting oxidative phosphorylation and, therefore, ATP production (Cardoso et al., 2001; Eckert et al., 2010; Spuch et al., 2012; Kaminsky et al., 2015; Brewer et al., 2020). Moreover, mitochondria delivery to needed brain regions is dependent on tau, a protein related to microtubules (Quintanilla et al., 2012; Amadoro et al., 2014). Mitochondria are observed to be differentially localized in AD brains, suggesting that mitochondria trafficking is affected (Nicholls and Budd, 2000; Duchen, 2012; Devine and Kittler, 2018; Son and Han, 2018; Rangaraju et al., 2019), and provides further support for mitochondrial-based contributors to neurodegeneration.
Potential AD Therapies Targeting Lipid Metabolism
Dietary Modification Studies
With the realization that lipids are altered in AD pathology, several studies have identified specific lipids that may be used as dietary supplements to alleviate AD symptoms (Table 3). The major lipids include omega-3 fatty acids (DHA, EPA), choline-containing lipids, cholesterol, and lipids with antioxidant properties (CoQ10, Vitamin K).

Table 3. Lipid diets and their effects on AD.
Several dietary intervention studies using DHA have yielded mixed effects on AD symptoms. A likely reason for these mixed results is that different disease severity, different formulations, and variable endpoint and time of interventions were studied (Fonteh, 2018). Recent studies indicate that the best form of DHA delivery into the brain is through the Msf2a LPC receptors (Sugasini et al., 2019). A better understanding of the right formulation and optimal concentrations of DHA probably supplemented at the prodromal phase of AD will likely yield beneficial outcomes.
Modification of Lipid Metabolism
Metabolism of lipids can be altered to prevent depletion of their levels in the AD by targeting pathways that transport or catabolize these lipids in the brain.
Lipid Transport Into the Brain
Several lipoproteins and their receptor complexes are the major form by which lipids bypass the BBB to be delivered into the brain. Several of these lipoprotein genes are linked to AD pathology (Table 1). Some lipoproteins have protective effects, while others have AD enhancing properties. For example, HDL has been shown to be protective by improving Aβ clearance, delaying Aβ fibrillization, suppressing vascular inflammation, and inducing endothelial nitric oxide production (Button et al., 2019).
Cholesterol Metabolism
Since cholesterol metabolism altered at several stages of AD, modulation of its metabolism may have beneficial effects on disease progression. Modification of cholesterol homeostasis can be influenced during its consumption, at the level of its biosynthesis, and during its transport into the brain. The use of statin to alter cholesterol biosynthesis is proposed to be insightful in AD pathophysiology and therapy (Wolozin et al., 2004; Hoglund et al., 2005; Biondi, 2007; Evans et al., 2009). Gene therapy targeting cholesterol 24-hydroxylase reduces the amyloid pathology before or after the onset of amyloid plaques in mouse AD models (Hudry et al., 2010). Studies in mouse models show that blocking the conversion of cholesterol to cholesterol esters has beneficial effects on AD (Shibuya et al., 2015). The relationship between hypercholesterolemia, cholesterol-lowering therapies, and the role of oxysterols in AD pathology have led to the proposition that cholesterol metabolites are valuable targets for alternative AD treatments or prevention (Loera-Valencia et al., 2019). Neuroinflammatory pathways mediated by toll-like receptor 4 (TLR4)-mediated signaling can aggravate AD symptoms. In a rodent AD model, treatment with an anti-inflammatory steroid (atorvastatin) regulates this inflammatory process and results in the amelioration of cognitive deficits (Wang et al., 2018).
Lipolytic Enzymes
The activity or expression of several lipolytic enzymes are altered in AD. Phospholipase A2 (PLA2) is associated with amyloid plaques, and reduction of its activity and expression ameliorates AD. Plasmalogen selective PLA2 is also altered in AD. Our studies show an increase in PLA2 activity of CSF of AD participants accompanied by an increase in lysophosphatidylcholine (LPC). LPC is known to disrupt the BBB, and changes in PLA2 are associated with inflammation. The association of PLA2 with AD pathology suggests that inhibitors of PLA2 activity or expression may be an effective means of preventing AD. Ong et al. (2015) reviewed the importance of several natural and synthetic PLA2 inhibitors on the treatment of neurological disorders. Since PLA2 isoforms may have divergent effects on membrane remodeling and function, there is a need for isoform-specific inhibitors in order to avoid toxicity encountered with non-selective inhibitors. In addition to PLA2, phospholipase D (PLD) and phospholipase C (PLC) expression and activities are associated with AD pathology. These lipases that are linked with neurite growth and signaling, respectively, offer other avenues for exploring AD treatments.
Lipid Oxidation Inhibitors
There is convincing evidence for the importance of oxidative stress on AD pathology (Sun et al., 2001; Bassett and Montine, 2003; Bacchetti et al., 2015). The most important brain fatty acid, DHA, is a polyunsaturated fatty that is easily susceptible to oxidative damage. While HDL is protective against oxidative damage, VLDL is easily oxidized. Oxidatively damaged lipids contribute to AD pathology by their highly neurotoxic properties (Bassett et al., 1999). Approaches that reduce oxidation are expected to reduce AD progression. These include the use of natural antioxidants, carnosine, lipoic acid, Ginkgo biloba flavonoids, soybean isoflavones, vitamin K, homocysteine, curcumin (Rutten et al., 2002; Vina et al., 2004; Frank and Gupta, 2005; Mancuso et al., 2007; Zhao, 2009; Cankurtaran et al., 2013). A limitation of natural antioxidant is the lack of demonstration of efficacy. Given that oxidative stress destroys mitochondrial function, an objective measure of any antioxidant can be their ability to restore mitochondrial function (Kumar and Singh, 2015; Kwon et al., 2016; Yu et al., 2016). The role of endogenous lipids in oxidative stress can be exploited when there is an uncontrolled formation of ROS and RNS or when the antioxidants contribute to disease pathology (Leuti et al., 2019). Also, the source of ROS determines the effects on cellular physiology and manipulation of the ubiquinone redox state is proposed to be a viable approach of delaying aging and therapy (Scialo et al., 2016; Wojsiat et al., 2018).
Concluding Remarks
Biochemical, physiological, and genetic analyses show that lipid metabolism interphases with all the major facets of AD pathology (Figure 1). In normal aging, lipid metabolic homeostasis ensures that the basic functions of the brain are met. In AD, there is dyshomeostasis of lipid metabolism, and this results in abnormal functions of the brain that characterize disease progression. This underscores the need for detailed analyses of brain lipid homeostasis in order to unravel more comprehensive mechanisms, specific biomarkers, and novel therapies of AD.
Author Contributions
AF contributed to the conceptualization and study design, supervised the data, and carried out the project administration and funding acquisition. HC, VS, and AF contributed to writing of the original draft and manuscript preparation. HC and AF contributed to the manuscript review and editing. VS and AF prepared the figures and illustrations.
Funding
AF was supported by funds from the L. K. Whittier Foundation and HMRI. VS is supported by R01 AG054434 and R01 AG055770 to Dr. H. Yassine.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank HMRI and Dr. M. Harrington for providing support and a scholarly environment for the summer student program. Dr. H. Yassine provided mentorship to VS.
References
Abbott, N. J. (2000). Inflammatory mediators and modulation of blood-brain barrier permeability. Cell. Mol. Neurobiol. 20, 131–147.
Abbott, N. J., Patabendige, A. A., Dolman, D. E., Yusof, S. R., and Begley, D. J. (2010). Structure and function of the blood-brain barrier. Neurobiol. Dis. 37, 13–25.
Agrawal, M., Ajazuddin, Tripathi, D. K., Saraf, S., Saraf, S., Antimisiaris, S. G., et al. (2017). Recent advancements in liposomes targeting strategies to cross blood-brain barrier (BBB) for the treatment of Alzheimer’s disease. J. Control. Release 260, 61–77. doi: 10.1016/j.jconrel.2017.05.019
Alaupovic, P. (1996). Significance of apolipoproteins for structure, function, and classification of plasma lipoproteins. Methods Enzymol. 263, 32–60. doi: 10.1016/s0076-6879(96)63004-3
Alexander, G. E. (2017). An Emerging role for imaging white matter in the preclinical risk for Alzheimer disease: linking beta-amyloid to myelin. JAMA Neurol. 74, 17–19.
Almeida, R. G., and Lyons, D. A. (2014). On the resemblance of synapse formation and CNS myelination. Neuroscience 276, 98–108. doi: 10.1016/j.neuroscience.2013.08.062
Almeida, R. G., and Lyons, D. A. (2017). On myelinated axon plasticity and neuronal circuit formation and function. J. Neurosci. 37, 10023–10034. doi: 10.1523/jneurosci.3185-16.2017
Amadoro, G., Corsetti, V., Florenzano, F., Atlante, A., Ciotti, M. T., Mongiardi, M. P., et al. (2014). AD-linked, toxic NH2 human tau affects the quality control of mitochondria in neurons. Neurobiol. Dis. 62, 489–507. doi: 10.1016/j.nbd.2013.10.018
Anceline, M.-L., Ripoche, E., Dupuy, A.-M., Samieri, C., Rouaud, O., Berr, C., et al. (2014). Gender-specific associations between lipids and cognitive decline in the elderly. Eur. Neuropsychopharmacol. 24, 1056–1066. doi: 10.1016/j.euroneuro.2014.02.003
Anderson, G. (2018). Linking the biological underpinnings of depression: role of mitochondria interactions with melatonin, inflammation, sirtuins, tryptophan catabolites, DNA repair and oxidative and nitrosative stress, with consequences for classification and cognition. Prog. Neuropsychopharmacol. Biol. Psychiatry 80, 255–266. doi: 10.1016/j.pnpbp.2017.04.022
Ando, S., Tanaka, Y., Toyoda, Y., and Kon, K. (2003). Turnover of myelin lipids in aging brain. Neurochem. Res. 28, 5–13.
Andreone, B. J., Chow, B. W., Tata, A., Lacoste, B., Ben-Zvi, A., Bullock, K., et al. (2017). Blood-brain barrier permeability is regulated by lipid transport-dependent suppression of caveolae-mediated transcytosis. Neuron 94, 581.e5–594.e5. doi: 10.1016/j.neuron.2017.03.043
Andrew, R. J., Kellett, K. A., Thinakaran, G., and Hooper, N. M. (2016). A greek tragedy: the growing complexity of alzheimer amyloid precursor protein proteolysis. J. Biol. Chem. 291, 19235–19244. doi: 10.1074/jbc.r116.746032
Aoki, C., Fujisawa, S., Mahadomrongkul, V., Shah, P. J., Nader, K., and Erisir, A. (2003). NMDA receptor blockade in intact adult cortex increases trafficking of NR2A subunits into spines, postsynaptic densities, and axon terminals. Brain Res. 963, 139–149. doi: 10.1016/s0006-8993(02)03962-8
Apak, R., Ozyurek, M., Guclu, K., and Capanoglu, E. (2016). Antioxidant activity/capacity measurement. 3. reactive oxygen and nitrogen species (ROS/RNS) scavenging assays, oxidative stress biomarkers, and chromatographic/chemometric assays. J. Agric. Food Chem. 64, 1046–1070. doi: 10.1021/acs.jafc.5b04744
Arnoldussen, I. A., Zerbi, V., Wiesmann, M., Noordman, R. H., Bolijn, S., Mutsaers, M. P., et al. (2016). Early intake of long-chain polyunsaturated fatty acids preserves brain structure and function in diet-induced obesity. J. Nutr. Biochem. 30, 177–188. doi: 10.1016/j.jnutbio.2015.12.011
Asada, T., Kariya, T., Yamagata, Z., Kinoshita, T., and Asaka, A. (1996). ApoE epsilon 4 allele and cognitive decline in patients with Alzheimer’s disease. Neurology 47:603. doi: 10.1212/wnl.47.2.603
Audagnotto, M., Kengo Lorkowski, A., and Dal Peraro, M. (2018). Recruitment of the amyloid precursor protein by gamma-secretase at the synaptic plasma membrane. Biochem. Biophys. Res. Commun. 498, 334–341. doi: 10.1016/j.bbrc.2017.10.164
Ayloo, S., and Gu, C. (2019). Transcytosis at the blood-brain barrier. Curr. Opin. Neurobiol. 57, 32–38. doi: 10.1016/0006-8993(87)90236-8
Bacchetti, T., Vignini, A., Giulietti, A., Nanetti, L., Provinciali, L., Luzzi, S., et al. (2015). Higher levels of oxidized low density lipoproteins in Alzheimer’s disease patients: roles for platelet activating factor acetyl hydrolase and paraoxonase-1. J. Alzheimers Dis. 46, 179-186. doi: 10.3233/JAD-143096
Balazs, Z., Panzenboeck, U., Hammer, A., Sovic, A., Quehenberger, O., Malle, E., et al. (2004). Uptake and transport of high-density lipoprotein (HDL) and HDL-associated alpha-tocopherol by an in vitro blood-brain barrier model. J. Neurochem. 89, 939–950. doi: 10.1111/j.1471-4159.2004.02373.x
Baldo, G., Giugliani, R., and Matte, U. (2014). Lysosomal enzymes may cross the blood-brain-barrier by pinocytosis: implications for enzyme replacement therapy. Med. Hypotheses 82, 478–480. doi: 10.1016/j.mehy.2014.01.029
Banks, W. A. (1999). Physiology and pathology of the blood-brain barrier: implications for microbial pathogenesis, drug delivery and neurodegenerative disorders. J. Neurovirol. 5, 538–555. doi: 10.3109/13550289909021284
Banks, W. A., Farr, S., Salameh, T. S., Niehoff, M. L., Rhea, E. M., Morley, J. E., et al. (2018). Triglycerides cross the blood–brain barrier and induce central leptin and insulin receptor resistance. Int. J. Obes. 42, 391–397. doi: 10.1038/ijo.2017.231
Barbagallo, C. M., Levine, G. A., Blanche, P. J., Ishida, B. Y., and Krauss, R. M. (1998). Influence of apoE content on receptor binding of large, bouyant LDL in subjects with different LDL subclass phenotypes. Arterioscler. Thromb. Vasc. Biol. 18, 466–472. doi: 10.1161/01.atv.18.3.466
Bartzokis, G. (2011). Alzheimer’s disease as homeostatic responses to age-related myelin breakdown. Neurobiol. Aging 32, 1341–1371. doi: 10.1016/j.neurobiolaging.2009.08.007
Bartzokis, G., Lu, P. H., Geschwind, D. H., Edwards, N., Mintz, J., and Cummings, J. L. (2006). Apolipoprotein E genotype and age-related myelin breakdown in healthy individuals: implications for cognitive decline and dementia. Arch. Gen. Psychiatry 63, 63–72.
Bassett, C. N., and Montine, T. J. (2003). Lipoproteins and lipid peroxidation in Alzheimer’s disease. J. Nutr. Health Aging 7, 24–29.
Bassett, C. N., Neely, M. D., Sidell, K. R., Markesbery, W. R., Swift, L. L., and Montine, T. J. (1999). Cerebrospinal fluid lipoproteins are more vulnerable to oxidation in Alzheimer’s disease and are neurotoxic when oxidized ex vivo. Lipids 34, 1273–1280. doi: 10.1007/s11745-999-0478-1
Baum, L., Chen, L., Masliah, E., Chan, Y. S., Ng, H. K., and Pang, C. P. (1999). Lipoprotein lipase mutations and Alzheimer’s disease. Am. J. Med. Genet. 88, 136–139. doi: 10.1002/(sici)1096-8628(19990416)88:2<136::aid-ajmg8>3.0.co;2-d
Bazan, N. G. (2005). Synaptic signaling by lipids in the life and death of neurons. Mol. Neurobiol. 31, 219–230. doi: 10.1385/mn:31:1-3:219
Bedse, G., Romano, A., Lavecchia, A. M., Cassano, T., and Gaetani, S. (2015). The role of endocannabinoid signaling in the molecular mechanisms of neurodegeneration in Alzheimer’s disease. J. Alzheimers Dis. 43, 1115–1136. doi: 10.3233/jad-141635
Belayev, L., Hong, S. H., Menghani, H., Marcell, S. J., Obenaus, A., Freitas, R. S., et al. (2018). Docosanoids promote neurogenesis and angiogenesis, blood-brain barrier integrity, penumbra protection, and neurobehavioral recovery after experimental ischemic stroke. Mol. Neurobiol. 55, 7090–7106. doi: 10.1007/s12035-018-1136-3
Belkouch, M., Hachem, M., Elgot, A., Lo Van, A., Picq, M., Guichardant, M., et al. (2016). The pleiotropic effects of Omega-3 docosahexaenoic acid on the hallmarks of Alzheimer’s disease. J. Nutr. Biochem. 38, 1–11. doi: 10.1016/j.jnutbio.2016.03.002
Bellet, M. M., Masri, S., Astarita, G., Sassone-Corsi, P., Della Fazia, M. A., and Servillo, G. (2016). Histone deacetylase SIRT1 controls proliferation, circadian rhythm, and lipid metabolism during liver regeneration in mice. J. Biol. Chem. 291, 23318–23329. doi: 10.1074/jbc.m116.737114
Benton, D. (2001). The impact of the supply of glucose to the brain on mood and memory. Nutr. Rev. 59, S20–S21.
Benton, D., Parker, P. Y., and Donohoe, R. T. (1996). The supply of glucose to the brain and cognitive functioning. J. Biosoc. Sci. 28, 463–479. doi: 10.1017/s0021932000022537
Berg, C. N., Sinha, N., and Gluck, M. A. (2019). The effects of APOE and ABCA7 on cognitive function and Alzheimer’s disease risk in african americans: a focused mini review. Front. Hum. Neurosci. 13:387. doi: 10.3389/fnhum.2019.00387
Bernath, M. M., Bhattacharyya, S., Nho, K., Barupal, D. K., Fiehn, O., Baillie, R., et al. (2019). Serum triglycerides in Alzheimer’s disease: relation to neuroimaging and CSF biomarkers. bioRxiv [Preprint]. doi: 10.1101/441394
Betsholtz, C. (2014). Physiology: double function at the blood-brain barrier. Nature 509, 432–433. doi: 10.1038/nature13339
Bhattacharyya, R., Barren, C., and Kovacs, D. M. (2013). Palmitoylation of amyloid precursor protein regulates amyloidogenic processing in lipid rafts. J. Neurosci. 33, 11169–11183. doi: 10.1523/jneurosci.4704-12.2013
Biondi, E. (2007). Statin-like drugs for the treatment of brain cholesterol loss in Alzheimer’s disease. Curr. Drug. Saf. 2, 173–176. doi: 10.2174/157488607781668927
Birben, E., Sahiner, U. M., Sackesen, C., Erzurum, S., and Kalayci, O. (2012). Oxidative stress and antioxidant defense. World Allergy Organ. J. 5, 9–19. doi: 10.1097/WOX.0b013e3182439613
Biringer, R. G. (2019). The role of eicosanoids in Alzheimer’s disease. Int. J. Environ. Res. Public Health 16:2560. doi: 10.3390/ijerph16142560
Black, J. B., Premont, R. T., and Daaka, Y. (2016). Feedback regulation of G protein-coupled receptor signaling by GRKs and arrestins. Semin. Cell Dev. Biol. 50, 95–104. doi: 10.1016/j.semcdb.2015.12.015
Blain, J. F., and Poirier, J. (2004). Cholesterol homeostasis and the pathophysiology of Alzheimer’s disease. Expert. Rev. Neurother. 4, 823–829. doi: 10.1586/14737175.4.5.823
Blain, J. F., Aumont, N., Theroux, L., Dea, D., and Poirier, J. (2006). A polymorphism in lipoprotein lipase affects the severity of Alzheimer’s disease pathophysiology. Eur. J. Neurosci. 24, 1245–1251. doi: 10.1111/j.1460-9568.2006.05007.x
Block, J. (2019). Alzheimer’s disease might depend on enabling pathogens which do not necessarily cross the blood-brain barrier. Med. Hypotheses. 125, 129–136. doi: 10.1016/j.mehy.2019.02.044
Bolanos-Garcia, V. M., and Miguel, R. N. (2003). On the structure and function of apolipoproteins: more than a family of lipid-binding proteins. Prog. Biophys. Mol. Biol. 83, 47–68. doi: 10.1016/s0079-6107(03)00028-2
Bos, D. J., van Montfort, S. J., Oranje, B., Durston, S., and Smeets, P. A. (2016). Effects of Omega-3 polyunsaturated fatty acids on human brain morphology and function: what is the evidence? Eur. Neuropsychopharmacol. 26, 546–561. doi: 10.1016/j.euroneuro.2015.12.031
Bourre, J. M. (1991). [Vitamin E: protection of membrane polyunsaturated fatty acids against radical peroxidation in the course of cerebral aging, particularly in cerebral capillaries and microvessels]. Bull. Acad. Natl. Med. 175, 1305–1317.
Bradbury, M. W. (1984). The structure and function of the blood-brain barrier. Fed. Proc. 43, 186–190.
Bradley, W. A., and Gianturco, S. H. (1986). ApoE is necessary and sufficient for the binding of large triglyceride-rich lipoproteins to the LDL receptor; apoB is unnecessary. J. Lipid. Res. 27, 40–48.
Braun, V., and Hantke, K. (2019). Lipoproteins: structure, function, biosynthesis. Subcell. Biochem. 92, 39–77. doi: 10.1007/978-3-030-18768-2_3
Brewer, G. J., Herrera, R. A., Philipp, S., Sosna, J., Reyes-Ruiz, J. M., and Glabe, C. G. (2020). Age-related intraneuronal aggregation of amyloid-beta in endosomes, mitochondria, autophagosomes, and lysosomes. J. Alzheimers Dis. 73, 229–246. doi: 10.3233/jad-190835
Brown, J. III, Theisler, C., Silberman, S., Magnuson, D., Gottardi-Littell, N., Lee, J. M., et al. (2004). Differential expression of cholesterol hydroxylases in Alzheimer’s disease. J. Biol. Chem. 279, 34674–34681. doi: 10.1074/jbc.m402324200
Brown, R. C., and Davis, T. P. (2002). Calcium modulation of adherens and tight junction function: a potential mechanism for blood-brain barrier disruption after stroke. Stroke 33, 1706–1711. doi: 10.1161/01.str.0000016405.06729.83
Burgess, B. L., McIsaac, S., Naus, K. E., Chan, J. Y., Tansley, G. H., Yang, J., et al. (2006). Elevated plasma triglyceride levels precede amyloid deposition in Alzheimer’s disease mouse models with abundant A beta in plasma. Neurobiol. Dis. 24, 114–127. doi: 10.1016/j.nbd.2006.06.007
Burgisser, P., Matthieu, J. M., Jeserich, G., and Waehneldt, T. V. (1986). Myelin lipids: a phylogenetic study. Neurochem. Res. 11, 1261–1272. doi: 10.1007/bf00966121
Butterfield, D. A., Castegna, A., Lauderback, C. M., and Drake, J. (2002). Evidence that amyloid beta-peptide-induced lipid peroxidation and its sequelae in Alzheimer’s disease brain contribute to neuronal death. Neurobiol. Aging 23, 655–664. doi: 10.1016/s0197-4580(01)00340-2
Button, E. B., Gilmour, M., Cheema, H. K., Martin, E. M., Agbay, A., Robert, J., et al. (2019). Vasoprotective functions of high-density lipoproteins relevant to Alzheimer’s disease are partially conserved in apolipoprotein B-depleted Plasma. Int. J. Mol. Sci. 20:462. doi: 10.3390/ijms20030462
Campbell, S. D., Regina, K. J., and Kharasch, E. D. (2014). Significance of lipid composition in a blood-brain barrier-mimetic PAMPA assay. J. Biomol. Screen 19, 437–444. doi: 10.1177/1087057113497981
Cankurtaran, M., Yesil, Y., Kuyumcu, M. E., Ozturk, Z. A., Yavuz, B. B., Halil, M., et al. (2013). Altered levels of homocysteine and serum natural antioxidants links oxidative damage to Alzheimer’s disease. J. Alzheimers Dis. 33, 1051–1058. doi: 10.3233/jad-2012-121630
Cantor, R. S. (2018). Path to the desensitized state of ligand-gated ion channels: why are inhibitory and excitatory receptors different? J. Phys. Chem. B 122, 5368–5374. doi: 10.1021/acs.jpcb.7b10961
Caporaso, G. L., Takei, K., Gandy, S. E., Matteoli, M., Mundigl, O., Greengard, P., et al. (1994). Morphologic and biochemical analysis of the intracellular trafficking of the Alzheimer beta/A4 amyloid precursor protein. J. Neurosci. 14, 3122–3138. doi: 10.1523/jneurosci.14-05-03122.1994
Cardoso, S. M., Santos, S., Swerdlow, R. H., and Oliveira, C. R. (2001). Functional mitochondria are required for amyloid beta-mediated neurotoxicity. FASEB J. 15, 1439–1441. doi: 10.1096/fj.00-0561fje
Carvey, P. M., Hendey, B., and Monahan, A. J. (2009). The blood-brain barrier in neurodegenerative disease: a rhetorical perspective. J. Neurochem. 111, 291–314. doi: 10.1111/j.1471-4159.2009.06319.x
Castro Dias, M., Coisne, C., Baden, P., Enzmann, G., Garrett, L., Becker, L., et al. (2019). Claudin-12 is not required for blood-brain barrier tight junction function. Fluids Barriers CNS 16:30. doi: 10.1186/s12987-019-0150-9
Chandrasekharan, J. A., and Sharma-Walia, N. (2015). Lipoxins: nature’s way to resolve inflammation. J. Inflamm. Res. 8, 181–192.
Chang, C. Y., Ke, D. S., and Chen, J. Y. (2009). Essential fatty acids and human brain. Acta Neurol. Taiwan. 18, 231–241.
Chang, Y. T., Hsu, S. W., Huang, S. H., Huang, C. W., Chang, W. N., Lien, C. Y., et al. (2019). ABCA7 polymorphisms correlate with memory impairment and default mode network in patients with APOEepsilon4-associated Alzheimer’s disease. Alzheimers Res. Ther. 11:103. doi: 10.1186/s13195-019-0563-3
Chappus-McCendie, H., Chevalier, L., Roberge, C., and Plourde, M. (2019). Omega-3 PUFA metabolism and brain modifications during aging. Prog. Neuropsychopharmacol. Biol. Psychiatry 94:109662. doi: 10.1016/j.pnpbp.2019.109662
Cheignon, C., Jones, M., Atrian-Blasco, E., Kieffer, I., Faller, P., Collin, F., et al. (2017). Identification of key structural features of the elusive Cu-Abeta complex that generates ROS in Alzheimer’s disease. Chem. Sci. 8, 5107–5118. doi: 10.1039/c7sc00809k
Cheignon, C., Tomas, M., Bonnefont-Rousselot, D., Faller, P., Hureau, C., and Collin, F. (2018). Oxidative stress and the amyloid beta peptide in Alzheimer’s disease. Redox. Biol. 14, 450–464.
Chen, J., Wei, Y., Chen, X., Jiao, J., and Zhang, Y. (2017). Polyunsaturated fatty acids ameliorate aging via redox-telomere-antioncogene axis. Oncotarget 8, 7301–7314. doi: 10.18632/oncotarget.14236
Chen, R., Zhang, J., Wu, Y., Wang, D., Feng, G., Tang, Y. P., et al. (2012). Monoacylglycerol lipase is a therapeutic target for Alzheimer’s disease. Cell. Rep. 2, 1329–1339. doi: 10.1016/j.celrep.2012.09.030
Chen, X., Hui, L., and Geiger, J. D. (2014). Role of LDL cholesterol and endolysosomes in amyloidogenesis and Alzheimer’s disease. J. Neurol. Neurophysiol. 5:236. doi: 10.4172/2155-9562.1000236
Cheng, F., Cappai, R., Lidfeldt, J., Belting, M., Fransson, L. A., and Mani, K. (2014). Amyloid precursor protein (APP)/APP-like protein 2 (APLP2) expression is required to initiate endosome-nucleus-autophagosome trafficking of glypican-1-derived heparan sulfate. J. Biol. Chem. 289, 20871–20878. doi: 10.1074/jbc.m114.552810
Cherubini, A., Andres-Lacueva, C., Martin, A., Lauretani, F., Iorio, A. D., Bartali, B., et al. (2007). Low plasma N-3 fatty acids and dementia in older persons: the InCHIANTI study. J. Gerontol. A Biol. Sci. Med. Sci. 62, 1120–1126. doi: 10.1093/gerona/62.10.1120
Childs, C. E., Romeu-Nadal, M., Burdge, G. C., and Calder, P. C. (2008). Gender differences in the n-3 fatty acid content of tissues. Proc. Nutr. Soc. 67, 19–27. doi: 10.1017/s0029665108005983
Chiu, C. C., Su, K. P., Cheng, T. C., Liu, H. C., Chang, C. J., Dewey, M. E., et al. (2008). The effects of Omega-3 fatty acids monotherapy in Alzheimer’s disease and mild cognitive impairment: a preliminary randomized double-blind placebo-controlled study. Prog. Neuropsychopharmacol. Biol. Psychiatry 32, 1538–1544. doi: 10.1016/j.pnpbp.2008.05.015
Chiurchiu, V., Leuti, A., and Maccarrone, M. (2018). Bioactive lipids and chronic inflammation: managing the fire within. Front. Immunol. 9:38. doi: 10.3389/fimmu.2018.00038
Chow, V. W., Mattson, M. P., Wong, P. C., and Gleichmann, M. (2010). An overview of APP processing enzymes and products. Neuromol. Med. 12, 1–12. doi: 10.1007/s12017-009-8104-z
Chrast, R., Saher, G., Nave, K. A., and Verheijen, M. H. (2011). Lipid metabolism in myelinating glial cells: lessons from human inherited disorders and mouse models. J. Lipid Res. 52, 419–434. doi: 10.1194/jlr.r009761
Chu, L. W., Li, Y., Li, Z., Tang, A. Y., Cheung, B. M., Leung, R. Y., et al. (2007). A novel intronic polymorphism of ABCA1 gene reveals risk for sporadic Alzheimer’s disease in Chinese. Am. J. Med. Genet. B Neuropsychiatr. Genet. 144B, 1007–1013. doi: 10.1002/ajmg.b.30525
Chun, Y. S., Park, Y., Oh, H. G., Kim, T. W., Yang, H. O., Park, M. K., et al. (2015). O-GlcNAcylation promotes non-amyloidogenic processing of amyloid-beta protein precursor via inhibition of endocytosis from the plasma membrane. J. Alzheimers. Dis. 44, 261–275. doi: 10.3233/jad-140096
Chung, S. J., Kim, M. J., Kim, Y. J., Kim, J., You, S., Jang, E. H., et al. (2014). CR1, ABCA7, and APOE genes affect the features of cognitive impairment in Alzheimer’s disease. J. Neurol. Sci. 339, 91–96. doi: 10.1016/j.jns.2014.01.029
Chung, W. S., Verghese, P. B., Chakraborty, C., Joung, J., Hyman, B. T., Ulrich, J. D., et al. (2016). Novel allele-dependent role for APOE in controlling the rate of synapse pruning by astrocytes. Proc. Natl. Acad. Sci. U.S.A. 113, 10186–10191. doi: 10.1073/pnas.1609896113
Clavey, V., Lestavel-Delattre, S., Copin, C., Bard, J. M., and Fruchart, J. C. (1995). Modulation of lipoprotein B binding to the LDL receptor by exogenous lipids and apolipoproteins CI, CII, CIII, and E. Arterioscler. Thromb. Vasc. Biol. 15, 963–971. doi: 10.1161/01.atv.15.7.963
Csernansky, J. G., Dong, H., Fagan, A. M., Wang, L., Xiong, C., Holtzman, D. M., et al. (2006). Plasma cortisol and progression of dementia in subjects with Alzheimer-type dementia. Am. J. Psychiatry 163, 2164–2169. doi: 10.1176/ajp.2006.163.12.2164
Cunnane, S. C., Schneider, J. A., Tangney, C., Tremblay-Mercier, J., Fortier, M., Bennett, D. A., et al. (2012). Plasma and brain fatty acid profiles in mild cognitive impairment and Alzheimer’s disease. J. Alzheimers Dis. 29, 691–697. doi: 10.3233/jad-2012-110629
Cutuli, D. (2017). Functional and structural benefits induced by Omega-3 polyunsaturated fatty acids during aging. Curr. Neuropharmacol. 15, 534–542. doi: 10.2174/1570159x14666160614091311
Daiello, L. A., Gongvatana, A., Dunsiger, S., Cohen, R. A., and Ott, B. R. (2015). Association of fish oil supplement use with preservation of brain volume and cognitive function. Alzheimers Dement. 11, 226–235. doi: 10.1016/j.jalz.2014.02.005
Daneman, R., and Prat, A. (2015). The blood-brain barrier. Cold Spring Harb. Perspect. Biol. 7:a020412.
Dash, P. K., and Moore, A. N. (1993). Inhibitors of endocytosis, endosome fusion, and lysosomal processing inhibit the intracellular proteolysis of the amyloid precursor protein. Neurosci. Lett. 164, 183–186. doi: 10.1016/0304-3940(93)90887-q
de Chaves, E. P., and Narayanaswami, V. (2008). Apolipoprotein E and cholesterol in aging and disease in the brain. Future Lipidol. 3, 505–530. doi: 10.2217/17460875.3.5.505
de Vries, H. E., Kooij, G., Frenkel, D., Georgopoulos, S., Monsonego, A., and Janigro, D. (2012). Inflammatory events at blood-brain barrier in neuroinflammatory and neurodegenerative disorders: implications for clinical disease. Epilepsia 53(Suppl. 6), 45–52. doi: 10.1111/j.1528-1167.2012.03702.x
de Wilde, M. C., Vellas, B., Girault, E., Yavuz, A. C., and Sijben, J. W. (2017). Lower brain and blood nutrient status in Alzheimer’s disease: results from meta-analyses. Alzheimers Dement. 3, 416–431. doi: 10.1016/j.trci.2017.06.002
Decsi, T., and Kennedy, K. (2011). Sex-specific differences in essential fatty acid metabolism. Am. J. Clin. Nutr. 94(6 Suppl), 1914S-1919S. doi: 10.3945/ajcn.110.000893
Dehouck, B., Fenart, L., Dehouck, M. P., Pierce, A., Torpier, G., and Cecchelli, R. (1997). A new function for the LDL receptor: transcytosis of LDL across the blood-brain barrier. J. Cell Biol. 138, 877–889. doi: 10.1083/jcb.138.4.877
Demeester, N., Castro, G., Desrumaux, C., De Geitere, C., Fruchart, J. C., Santens, P., et al. (2000). Characterization and functional studies of lipoproteins, lipid transfer proteins, and lecithin:cholesterol acyltransferase in CSF of normal individuals and patients with Alzheimer’s disease. J. Lipid. Res. 41, 963–974.
Denis, I., Potier, B., Heberden, C., and Vancassel, S. (2015). Omega-3 polyunsaturated fatty acids and brain aging. Curr. Opin. Clin. Nutr. Metab. Care 18, 139–146.
Derby, C. A., Crawford, S., Pasternak, R. C., Sowers, M., Sternfeld, B., and Matthews, K. A. (2009). Lipid changes during the menopause transition in relation to age and weight. Am. J. Epidemiol. 169, 1352–1361. doi: 10.1093/aje/kwp043
Desai, M. K., Mastrangelo, M. A., Ryan, D. A., Sudol, K. L., Narrow, W. C., and Bowers, W. J. (2010). Early oligodendrocyte/myelin pathology in Alzheimer’s disease mice constitutes a novel therapeutic target. Am. J. Pathol. 177, 1422–1435. doi: 10.2353/ajpath.2010.100087
Devine, M. J., and Kittler, J. T. (2018). Mitochondria at the neuronal presynapse in health and disease. Nat. Rev. Neurosci. 19, 63–80. doi: 10.1038/nrn.2017.170
Dienel, G. A., Cruz, N. F., Adachi, K., Sokoloff, L., and Holden, J. E. (1997). Determination of local brain glucose level with [14C]methylglucose: effects of glucose supply and demand. Am. J. Physiol. 273, E839–E849.
Dimas, P., Montani, L., Pereira, J. A., Moreno, D., Trotzmuller, M., Gerber, J., et al. (2019). CNS myelination and remyelination depend on fatty acid synthesis by oligodendrocytes. eLife 8:e44702. doi: 10.7554/eLife.44702
Ding, R. B., Bao, J., and Deng, C. X. (2017). Emerging roles of SIRT1 in fatty liver diseases. Int. J. Biol. Sci. 13, 852–867. doi: 10.7150/ijbs.19370
do Couto, F. S., de Mendonca, A., Garcia, C., Rocha, L., and Lechner, M. C. (1998). Age of onset in patients with Alzheimer’s disease with different apoE genotypes. J. Neurol. Neurosurg. Psychiatry 64:817. doi: 10.1136/jnnp.64.6.817
Dodelet-Devillers, A., Cayrol, R., van Horssen, J., Haqqani, A. S., de Vries, H. E., Engelhardt, B., et al. (2009). Functions of lipid raft membrane microdomains at the blood-brain barrier. J. Mol. Med. 87, 765–774. doi: 10.1007/s00109-009-0488-6
Doens, D., Valiente, P. A., Mfuh, A. M., X T Vo, A., Tristan, A., Carreno, L., et al. (2017). Identification of inhibitors of CD36-amyloid beta binding as potential agents for Alzheimer’s disease. ACS Chem. Neurosci. 8, 1232–1241. doi: 10.1021/acschemneuro.6b00386
Duchen, M. R. (2012). Mitochondria, calcium-dependent neuronal death and neurodegenerative disease. Pflugers. Arch. 464, 111–121. doi: 10.1007/s00424-012-1112-0
Duka, T., Tasker, R., and McGowan, J. F. (2000). The effects of 3-week estrogen hormone replacement on cognition in elderly healthy females. Psychopharmacology 149, 129–139. doi: 10.1007/s002139900324
Dunstan, J. A., Simmer, K., Dixon, G., and Prescott, S. L. (2008). Cognitive assessment of children at age 2(1/2) years after maternal fish oil supplementation in pregnancy: a randomised controlled trial. Arch. Dis. Child. Fetal. Neonatal. Ed. 93, F45–F50.
Dyall, S. C. (2015). Long-chain Omega-3 fatty acids and the brain: a review of the independent and shared effects of EPA, DPA and DHA. Front. Aging Neurosci. 7:52. doi: 10.3389/fnagi.2015.00052
Eckert, A., Schulz, K. L., Rhein, V., and Gotz, J. (2010). Convergence of amyloid-beta and tau pathologies on mitochondria in vivo. Mol. Neurobiol. 41, 107–114. doi: 10.1007/s12035-010-8109-5
Ehehalt, R., Keller, P., Haass, C., Thiele, C., and Simons, K. (2003). Amyloidogenic processing of the Alzheimer beta-amyloid precursor protein depends on lipid rafts. J. Cell. Biol. 160, 113–123. doi: 10.1083/jcb.200207113
El Haj, M., Antoine, P., Amouyel, P., Lambert, J. C., Pasquier, F., and Kapogiannis, D. (2016). Apolipoprotein E (APOE) epsilon4 and episodic memory decline in Alzheimer’s disease: a review. Ageing Res. Rev. 27, 15–22. doi: 10.1016/j.arr.2016.02.002
Elliott, D. A., Weickert, C. S., and Garner, B. (2010). Apolipoproteins in the brain: implications for neurological and psychiatric disorders. Clin. Lipidol. 51, 555–573. doi: 10.2217/clp.10.37
Erk, S., Meyer-Lindenberg, A., Opitz von Boberfeld, C., Esslinger, C., Schnell, K., Kirsch, P., et al. (2011). Hippocampal function in healthy carriers of the CLU Alzheimer’s disease risk variant. J. Neurosci. 31, 18180–18184. doi: 10.1523/jneurosci.4960-11.2011
Estus, S., Golde, T. E., Kunishita, T., Blades, D., Lowery, D., Eisen, M., et al. (1992). Potentially amyloidogenic, carboxyl-terminal derivatives of the amyloid protein precursor. Science 255, 726–728. doi: 10.1126/science.1738846
Evans, B. A., Evans, J. E., Baker, S. P., Kane, K., Swearer, J., Hinerfeld, D., et al. (2009). Long-term statin therapy and CSF cholesterol levels: implications for Alzheimer’s disease. Dement. Geriatr. Cogn. Disord. 27, 519–524. doi: 10.1159/000221835
Evin, G., and Li, Q. X. (2012). Platelets and Alzheimer’s disease: potential of APP as a biomarker. World J. Psychiatry 2, 102–113.
Evin, G., Zhu, A., Holsinger, R. M., Masters, C. L., and Li, Q. X. (2003). Proteolytic processing of the Alzheimer’s disease amyloid precursor protein in brain and platelets. J. Neurosci. Res. 74, 386–392.
Eyster, K. M. (2007). The membrane and lipids as integral participants in signal transduction: lipid signal transduction for the non-lipid biochemist. Adv. Physiol. Educ. 31, 5–16. doi: 10.1152/advan.00088.2006
Farooqui, A. A., and Horrocks, L. A. (1998). Plasmalogen-selective phospholipase A2 and its involvement in Alzheimer’s disease. Biochem. Soc. Trans. 26, 243–246.
Farooqui, A. A., Liss, L., and Horrocks, L. A. (1988). Stimulation of lipolytic enzymes in Alzheimer’s disease. Ann. Neurol. 23, 306–308. doi: 10.1002/ana.410230317
Feingold, K. R., and Grunfeld, C. (2000). “Introduction to lipids and lipoproteins,” in Endotext, eds K. R. Feingold, B. Anawalt, A. Boyce, G. Chrousos, K. Dungan, A. Grossman, et al. (South Dartmouth, MA: MDText.com, Inc).
Ferreira, L. (2019). What human blood-brain barrier models can tell us about BBB function and drug discovery? Expert. Opin. Drug. Discov. 14, 1113–1123. doi: 10.1080/17460441.2019.1646722
Fester, L., Zhou, L., Butow, A., Huber, C., von Lossow, R., Prange-Kiel, J., et al. (2009). Cholesterol-promoted synaptogenesis requires the conversion of cholesterol to estradiol in the hippocampus. Hippocampus 19, 692–705. doi: 10.1002/hipo.20548
Fidani, L., Goulas, A., Crook, R., Petersen, R. C., Tangalos, E., Kotsis, A., et al. (2004). An association study of the cholesteryl ester transfer protein TaqI B polymorphism with late onset Alzheimer’s disease. Neurosci. Lett. 357, 152–154. doi: 10.1016/j.neulet.2003.11.071
Filippov, V., Song, M. A., Zhang, K., Vinters, H. V., Tung, S., Kirsch, W. M., et al. (2012). Increased ceramide in brains with Alzheimer’s and other neurodegenerative diseases. J. Alzheimers Dis. 29, 537–547. doi: 10.3233/jad-2011-111202
Filou, S., Lhomme, M., Karavia, E. A., Kalogeropoulou, C., Theodoropoulos, V., Zvintzou, E., et al. (2016). Distinct roles of apolipoproteins A1 and E in the modulation of high-density lipoprotein composition and function. Biochemistry 55, 3752–3762. doi: 10.1021/acs.biochem.6b00389
Finean, J. B., and Robertson, J. D. (1958). Lipids and the structure of myelin. Br. Med. Bull. 14, 267–273. doi: 10.1093/oxfordjournals.bmb.a069695
Fishman, J. B., Rubin, J. B., Handrahan, J. V., Connor, J. R., and Fine, R. E. (1987). Receptor-mediated transcytosis of transferrin across the blood-brain barrier. J. Neurosci. Res. 18, 299–304. doi: 10.1002/jnr.490180206
Fonteh, A. (2018). Reasons why Omega-3 polyunsaturated fatty acids produce mixed results in alzheimer’s disease. J. Glycom. Lipid. 7:1.
Fonteh, A. N., Chiang, J., Cipolla, M., Hale, J., Diallo, F., Chirino, A., et al. (2013). Alterations in cerebrospinal fluid glycerophospholipids and phospholipase A2 activity in Alzheimer’s disease. J. Lipid. Res. 54, 2884–2897. doi: 10.1194/jlr.m037622
Fonteh, A. N., Cipolla, M., Chiang, A. J., Edminster, S. P., Arakaki, X., and Harrington, M. G. (2020). Polyunsaturated fatty acid composition of cerebrospinal fluid fractions shows their contribution to cognitive resilience of a pre-symptomatic Alzheimer’s disease cohort. Front. Physiol. 11:83. doi: 10.3389/fphys.2020.00083
Fonteh, A. N., Cipolla, M., Chiang, J., Arakaki, X., and Harrington, M. G. (2014). Human cerebrospinal fluid fatty acid levels differ between supernatant fluid and brain-derived nanoparticle fractions, and are altered in Alzheimer’s disease. PLoS One 9:e100519. doi: 10.1371/journal.pone.0100519
Fonteh, A. N., Ormseth, C., Chiang, J., Cipolla, M., Arakaki, X., and Harrington, M. G. (2015). Sphingolipid metabolism correlates with cerebrospinal fluid Beta amyloid levels in Alzheimer’s disease. PLoS One 10:e0125597. doi: 10.1371/journal.pone.0125597
Foster, E. M., Dangla-Valls, A., Lovestone, S., Ribe, E. M., and Buckley, N. J. (2019). Clusterin in Alzheimer’s disease: mechanisms, genetics, and lessons from other pathologies. Front. Neurosci. 13:164. doi: 10.3389/fnins.2019.00164
Frank, A. T., Zhao, B., Jose, P. O., Azar, K. M., Fortmann, S. P., and Palaniappan, L. P. (2014). Racial/ethnic differences in dyslipidemia patterns. Circulation 129, 570–579. doi: 10.1161/circulationaha.113.005757
Frank, B., and Gupta, S. (2005). A review of antioxidants and Alzheimer’s disease. Ann. Clin. Psychiatry 17, 269–286.
Frank, M. G., Baratta, M. V., Sprunger, D. B., Watkins, L. R., and Maier, S. F. (2007). Microglia serve as a neuroimmune substrate for stress-induced potentiation of CNS pro-inflammatory cytokine responses. Brain Behav. Immun. 21, 47–59. doi: 10.1016/j.bbi.2006.03.005
French, H. M., Reid, M., Mamontov, P., Simmons, R. A., and Grinspan, J. B. (2009). Oxidative stress disrupts oligodendrocyte maturation. J. Neurosci. Res. 87, 3076–3087. doi: 10.1002/jnr.22139
Freund Levi, Y., Vedin, I., Cederholm, T., Basun, H., Faxen Irving, G., Eriksdotter, M., et al. (2014). Transfer of Omega-3 fatty acids across the blood-brain barrier after dietary supplementation with a docosahexaenoic acid-rich Omega-3 fatty acid preparation in patients with Alzheimer’s disease: the OmegAD study. J. Intern. Med. 275, 428–436. doi: 10.1111/joim.12166
Freund-Levi, Y., Basun, H., Cederholm, T., Faxen-Irving, G., Garlind, A., Grut, M., et al. (2008). Omega-3 supplementation in mild to moderate Alzheimer’s disease: effects on neuropsychiatric symptoms. Int. J. Geriatr. Psychiatry 23, 161–169. doi: 10.1002/gps.1857
Freund-Levi, Y., Eriksdotter-Jonhagen, M., Cederholm, T., Basun, H., Faxen-Irving, G., Garlind, A., et al. (2006). Omega-3 fatty acid treatment in 174 patients with mild to moderate Alzheimer disease: OmegAD study: a randomized double-blind trial. Arch. Neurol. 63, 1402–1408.
Frieden, C., Wang, H., and Ho, C. M. W. (2017). A mechanism for lipid binding to apoE and the role of intrinsically disordered regions coupled to domain-domain interactions. Proc. Natl. Acad. Sci. U.S.A. 114, 6292–6297. doi: 10.1073/pnas.1705080114
Gazzola, K., Reeskamp, L., and van den Born, B. J. (2017). Ethnicity, lipids and cardiovascular disease. Curr. Opin. Lipidol. 28, 225–230. doi: 10.1097/mol.0000000000000412
Ghosh, M., Garcia-Castillo, D., Aguirre, V., Golshani, R., Atkins, C. M., Bramlett, H. M., et al. (2012). Proinflammatory cytokine regulation of cyclic AMP-phosphodiesterase 4 signaling in microglia in vitro and following CNS injury. Glia 60, 1839–1859. doi: 10.1002/glia.22401
Gilgun-Sherki, Y., Melamed, E., and Offen, D. (2001). Oxidative stress induced-neurodegenerative diseases: the need for antioxidants that penetrate the blood brain barrier. Neuropharmacology 40, 959–975. doi: 10.1016/s0028-3908(01)00019-3
Gilmore-Bykovskyi, A. L., Jin, Y., Gleason, C., Flowers-Benton, S., Block, L. M., Dilworth-Anderson, P., et al. (2019). Recruitment and retention of underrepresented populations in Alzheimer’s disease research: a systematic review. Alzheimers Dement. 19, 751–770. doi: 10.1016/j.trci.2019.09.018
Giltay, E. J., Gooren, L., Toorians, A. W., Katan, M. B., and Zock, P. L. (2004). Docosahexaenoic acid concentrations are higher in women than in men because of estrogenic effects. Am. J. Clin. Nutr. 80, 1167–1174. doi: 10.1093/ajcn/80.5.1167
Giulietti, A., Vignini, A., Nanetti, L., Mazzanti, L., Di Primio, R., and Salvolini, E. (2016). Alzheimer’s disease risk and progression: the role of nutritional supplements and their effect on drug therapy outcome. Curr. Neuropharmacol. 14, 177–190. doi: 10.2174/1570159x13666150928155321
Glorioso, C. A., Pfenning, A. R., Lee, S. S., Bennett, D. A., Sibille, E. L., Kellis, M., et al. (2019). Rate of brain aging and APOE epsilon4 are synergistic risk factors for Alzheimer’s disease. Life Sci. Alliance 2, e201900303. doi: 10.26508/lsa.201900303
Gold, M., Dolga, A. M., Koepke, J., Mengel, D., Culmsee, C., Dodel, R., et al. (2014). alpha1-antitrypsin modulates microglial-mediated neuroinflammation and protects microglial cells from amyloid-beta-induced toxicity. J. Neuroinflammation 11:165. doi: 10.1186/s12974-014-0165-8
Golde, T. E., Estus, S., Younkin, L. H., Selkoe, D. J., and Younkin, S. G. (1992). Processing of the amyloid protein precursor to potentially amyloidogenic derivatives. Science 255, 728–730. doi: 10.1126/science.1738847
Gong, C. X., Liu, F., Grundke-Iqbal, I., and Iqbal, K. (2006). Impaired brain glucose metabolism leads to Alzheimer neurofibrillary degeneration through a decrease in tau O-GlcNAcylation. J. Alzheimers Dis. 9, 1–12. doi: 10.3233/jad-2006-9101
Goozee, K., Chatterjee, P., James, I., Shen, K., Sohrabi, H. R., Asih, P. R., et al. (2017). Alterations in erythrocyte fatty acid composition in preclinical Alzheimer’s disease. Sci. Rep. 7:676.
Grabowska, W., Sikora, E., and Bielak-Zmijewska, A. (2017). Sirtuins, a promising target in slowing down the ageing process. Biogerontology 18, 447–476. doi: 10.1007/s10522-017-9685-9
Grassi, S., Giussani, P., Mauri, L., Prioni, S., Sonnino, S., and Prinetti, A. (2019). Lipid rafts and neurodegeneration: structural and functional roles in physiologic aging and neurodegenerative diseases. J. Lipid. Res. 61, 636–654.
Grimm, M. O., Haupenthal, V. J., Mett, J., Stahlmann, C. P., Blumel, T., Mylonas, N. T., et al. (2016). Oxidized docosahexaenoic acid species and lipid peroxidation products increase amyloidogenic amyloid precursor protein processing. Neurodegener. Dis. 16, 44–54. doi: 10.1159/000440839
Grimm, M. O., Rothhaar, T. L., Grosgen, S., Burg, V. K., Hundsdorfer, B., Haupenthal, V. J., et al. (2012). Trans fatty acids enhance amyloidogenic processing of the Alzheimer amyloid precursor protein (APP). J. Nutr. Biochem. 23, 1214–1223. doi: 10.1016/j.jnutbio.2011.06.015
Guan, Z., Wang, Y., Cairns, N. J., Lantos, P. L., Dallner, G., and Sindelar, P. J. (1999). Decrease and structural modifications of phosphatidylethanolamine plasmalogen in the brain with Alzheimer disease. J. Neuropathol. Exp. Neurol. 58, 740–747. doi: 10.1097/00005072-199907000-00008
Guo, X., Geng, M., and Du, G. (2005). Glucose transporter 1, distribution in the brain and in neural disorders: its relationship with transport of neuroactive drugs through the blood-brain barrier. Biochem. Genet. 43, 175–187. doi: 10.1007/s10528-005-1510-5
Hahn, G., Ponce-Alvarez, A., Deco, G., Aertsen, A., and Kumar, A. (2019). Portraits of communication in neuronal networks. Nat. Rev. Neurosci. 20, 117–127. doi: 10.1038/s41583-018-0094-0
Halliday, M. R., Rege, S. V., Ma, Q., Zhao, Z., Miller, C. A., Winkler, E. A., et al. (2016). Accelerated pericyte degeneration and blood-brain barrier breakdown in apolipoprotein E4 carriers with Alzheimer’s disease. J. Cereb. Blood Flow Metab. 36, 216–227. doi: 10.1038/jcbfm.2015.44
Hameister, R., Kaur, C., Dheen, S. T., Lohmann, C. H., and Singh, G. (2020). Reactive oxygen/nitrogen species (ROS/RNS) and oxidative stress in arthroplasty. J. Biomed. Mater. Res. B Appl. Biomater. 108, 2073–2087. doi: 10.1002/jbm.b.34546
Han, X., M Holtzman, D., McKeel, D. W. Jr., Kelley, J., and Morris, J. C. (2002). Substantial sulfatide deficiency and ceramide elevation in very early Alzheimer’s disease: potential role in disease pathogenesis. J. Neurochem. 82, 809–818. doi: 10.1046/j.1471-4159.2002.00997.x
Hansen, D. V., Hanson, J. E., and Sheng, M. (2018). Microglia in Alzheimer’s disease. J. Cell Biol. 217, 459–472.
Hansen, S. B. (2015). Lipid agonism: the PIP2 paradigm of ligand-gated ion channels. Biochim. Biophys. Acta 1851, 620–628. doi: 10.1016/j.bbalip.2015.01.011
Hao, S., Wang, R., Zhang, Y., and Zhan, H. (2018). Prediction of Alzheimer’s disease-associated genes by integration of gwas summary data and expression data. Front. Genet. 9:653. doi: 10.3389/fgene.2018.00653
Harik, S. I., and Kalaria, R. N. (1991). Blood-brain barrier abnormalities in Alzheimer’s disease. Ann. N. Y. Acad. Sci. 640, 47–52.
Harold, D., Abraham, R., Hollingworth, P., Sims, R., Gerrish, A., Hamshere, M. L., et al. (2009). Genome-wide association study identifies variants at CLU and PICALM associated with Alzheimer’s disease. Nat. Genet. 41, 1088–1093.
Harris, J. J., Jolivet, R., and Attwell, D. (2012). Synaptic energy use and supply. Neuron 75, 762–777. doi: 10.1016/j.neuron.2012.08.019
Hartmann, D. (2012). A brief history of APP secretases, their substrates and their functions. Curr. Alzheimer Res. 9, 138–139. doi: 10.2174/156720512799361628
Hasadsri, L., Wang, B. H., Lee, J. V., Erdman, J. W., Llano, D. A., Barbey, A. K., et al. (2013). Omega-3 fatty acids as a putative treatment for traumatic brain injury. J. Neurotrauma. 30, 897–906. doi: 10.1089/neu.2012.2672
Hascalovici, J. R., Vaya, J., Khatib, S., Holcroft, C. A., Zukor, H., Song, W., et al. (2009). Brain sterol dysregulation in sporadic AD and MCI: relationship to heme oxygenase-1. J. Neurochem. 110, 1241–1253. doi: 10.1111/j.1471-4159.2009.06213.x
Hawkins, R. A., and Biebuyck, J. F. (1979). Ketone bodies are selectively used by individual brain regions. Science 205, 325–327. doi: 10.1126/science.451608
Hedqvist, P., Raud, J., Palmertz, U., Kumlin, M., and Dahlen, S. E. (1991). Eicosanoids as mediators and modulators of inflammation. Adv. Prostaglandin. Thromboxane. Leukot. Res. 21B, 537–543.
Helbecque, N., Codron, V., Cottel, D., and Amouyel, P. (2008). An apolipoprotein A-I gene promoter polymorphism associated with cognitive decline, but not with Alzheimer’s disease. Dement. Geriatr. Cogn. Disord. 25, 97–102. doi: 10.1159/000112176
Heppner, F. L., Ransohoff, R. M., and Becher, B. (2015). Immune attack: the role of inflammation in Alzheimer disease. Nat. Rev. Neurosci. 16, 358–372. doi: 10.1038/nrn3880
Hering, H., Lin, C. C., and Sheng, M. (2003). Lipid rafts in the maintenance of synapses, dendritic spines, and surface AMPA receptor stability. J. Neurosci. 23, 3262–3271. doi: 10.1523/jneurosci.23-08-03262.2003
Herold, C., Hooli, B. V., Mullin, K., Liu, T., Roehr, J. T., Mattheisen, M., et al. (2016). Family-based association analyses of imputed genotypes reveal genome-wide significant association of Alzheimer’s disease with OSBPL6, PTPRG, and PDCL3. Mol. Psychiatry 21, 1608–1612. doi: 10.1038/mp.2015.218
Herskovits, A. Z., and Guarente, L. (2014). SIRT1 in neurodevelopment and brain senescence. Neuron 81, 471–483. doi: 10.1016/j.neuron.2014.01.028
Herz, J. (2001). The LDL receptor gene family: (un)expected signal transducers in the brain. Neuron 29, 571–581. doi: 10.1016/s0896-6273(01)00234-3
Heverin, M., Bogdanovic, N., Lutjohann, D., Bayer, T., Pikuleva, I., Bretillon, L., et al. (2004). Changes in the levels of cerebral and extracerebral sterols in the brain of patients with Alzheimer’s disease. J. Lipid Res. 45, 186–193. doi: 10.1194/jlr.m300320-jlr200
Higgs, G. A., Moncada, S., and Vane, J. R. (1984). Eicosanoids in inflammation. Ann. Clin. Res. 16, 287–299.
Hirsch-Reinshagen, V., and Wellington, C. L. (2007). Cholesterol metabolism, apolipoprotein E, adenosine triphosphate-binding cassette transporters, and Alzheimer’s disease. Curr. Opin. Lipidol. 18, 325–332. doi: 10.1097/mol.0b013e32813aeabf
Hoglund, K., Thelen, K. M., Syversen, S., Sjogren, M., von Bergmann, K., Wallin, A., et al. (2005). The effect of simvastatin treatment on the amyloid precursor protein and brain cholesterol metabolism in patients with Alzheimer’s disease. Dement. Geriatr. Cogn. Disord. 19, 256–265. doi: 10.1159/000084550
Hoofnagle, A. N., and Heinecke, J. W. (2009). Lipoproteomics: using mass spectrometry-based proteomics to explore the assembly, structure, and function of lipoproteins. J. Lipid Res. 50, 1967–1975. doi: 10.1194/jlr.r900015-jlr200
Hooper, C., De Souto Barreto, P., Pahor, M., Weiner, M., and Vellas, B. (2018). The relationship of Omega 3 polyunsaturated fatty acids in red blood cell membranes with cognitive function and brain structure: a review focussed on alzheimer’s disease. J. Prev. Alzheimers Dis. 5, 78–84.
Hosseini, M., Poljak, A., Braidy, N., Crawford, J., and Sachdev, P. (2020). Blood fatty acids in Alzheimer’s disease and mild cognitive impairment: a meta-analysis and systematic review. Ageing Res. Rev. 60, 101043. doi: 10.1016/j.arr.2020.101043
Hottman, D. A., Chernick, D., Cheng, S., Wang, Z., and Li, L. (2014). HDL and cognition in neurodegenerative disorders. Neurobiol. Dis. 72(Pt.A), 22–36. doi: 10.1016/j.nbd.2014.07.015
Hu, X., Xu, B., and Ge, W. (2017). The role of lipid bodies in the microglial aging process and related diseases. Neurochem. Res. 42, 3140–3148. doi: 10.1007/s11064-017-2351-4
Huang, Y., and Mahley, R. W. (2014). Apolipoprotein E: structure and function in lipid metabolism, neurobiology, and Alzheimer’s diseases. Neurobiol. Dis. 72(Pt. A), 3–12. doi: 10.1016/j.nbd.2014.08.025
Hudry, E., Van Dam, D., Kulik, W., De Deyn, P. P., Stet, F. S., Ahouansou, O., et al. (2010). Adeno-associated virus gene therapy with cholesterol 24-hydroxylase reduces the amyloid pathology before or after the onset of amyloid plaques in mouse models of Alzheimer’s disease. Mol. Ther. 18, 44–53. doi: 10.1038/mt.2009.175
Hulbert, A. J., Faulks, S. C., Harper, J. M., Miller, R. A., and Buffenstein, R. (2006). Extended longevity of wild-derived mice is associated with peroxidation-resistant membranes. Mech. Ageing Dev. 127, 653–657. doi: 10.1016/j.mad.2006.03.002
Hussain, G., Wang, J., Rasul, A., Anwar, H., Imran, A., Qasim, M., et al. (2019). Role of cholesterol and sphingolipids in brain development and neurological diseases. Lipids Health Dis. 18:26. doi: 10.1186/s12944-019-0965-z
Igarashi, M., Ma, K., Gao, F., Kim, H. W., Rapoport, S. I., and Rao, J. S. (2011). Disturbed choline plasmalogen and phospholipid fatty acid concentrations in Alzheimer’s disease prefrontal cortex. J. Alzheimers Dis. 24, 507–517. doi: 10.3233/jad-2011-101608
Ihara, M., Polvikoski, T. M., Hall, R., Slade, J. Y., Perry, R. H., Oakley, A. E., et al. (2010). Quantification of myelin loss in frontal lobe white matter in vascular dementia, Alzheimer’s disease, and dementia with Lewy bodies. Acta Neuropathol. 119, 579–589. doi: 10.1007/s00401-009-0635-8
Ikeshima-Kataoka, H., and Yasui, M. (2016). Correlation between astrocyte activity and recovery from blood-brain barrier breakdown caused by brain injury. Neuroreport 27, 894–900. doi: 10.1097/wnr.0000000000000619
Irizarry, M. C., Deng, A., Lleo, A., Berezovska, O., Von Arnim, C. A., Martin-Rehrmann, M., et al. (2004). Apolipoprotein E modulates gamma-secretase cleavage of the amyloid precursor protein. J. Neurochem. 90, 1132–1143.
Ishiura, S. (1991). Proteolytic cleavage of the Alzheimer’s disease amyloid A4 precursor protein. J. Neurochem. 56, 363–369. doi: 10.1111/j.1471-4159.1991.tb08160.x
Ito, J., Nagayasu, Y., Lu, R., Kheirollah, A., Hayashi, M., and Yokoyama, S. (2005). Astrocytes produce and secrete FGF-1, which promotes the production of apoE-HDL in a manner of autocrine action. J. Lipid Res. 46, 679–686. doi: 10.1194/jlr.m400313-jlr200
Ito, J., Nagayasu, Y., Miura, Y., Yokoyama, S., and Michikawa, M. (2014). Astrocytes endogenous apoE generates HDL-like lipoproteins using previously synthesized cholesterol through interaction with ABCA1. Brain Res. 1570, 1–12. doi: 10.1016/j.brainres.2014.04.037
Iwamoto, N., Kobayashi, K., and Kosaka, K. (1989). The formation of prostaglandins in the postmortem cerebral cortex of Alzheimer-type dementia patients. J. Neurol. 236, 80–84. doi: 10.1007/bf00314401
Iwasaki, A., and Medzhitov, R. (2015). Control of adaptive immunity by the innate immune system. Nat. Immunol. 16, 343–353. doi: 10.1038/ni.3123
Iyu, D., Juttner, M., Glenn, J. R., White, A. E., Johnson, A. J., Fox, S. C., et al. (2011). PGE1 and PGE2 modify platelet function through different prostanoid receptors. Prostaglandins Other Lipid Mediat. 94, 9–16. doi: 10.1016/j.prostaglandins.2010.11.001
Janciauskiene, S., and Wright, H. T. (1998). Inflammation, antichymotrypsin, and lipid metabolism: autogenic etiology of Alzheimer’s disease. Bioessays 20, 1039–1046. doi: 10.1002/(sici)1521-1878(199812)20:12<1039::aid-bies10>3.0.co;2-z
Janssen, C. I., and Kiliaan, A. J. (2014). Long-chain polyunsaturated fatty acids (LCPUFA) from genesis to senescence: the influence of LCPUFA on neural development, aging, and neurodegeneration. Prog. Lipid Res. 53, 1–17. doi: 10.1016/j.plipres.2013.10.002
Jean-Louis, T., Rockwell, P., and Figueiredo-Pereira, M. E. (2018). Prostaglandin J2 promotes O-GlcNAcylation raising APP processing by alpha- and beta-secretases: relevance to Alzheimer’s disease. Neurobiol. Aging 62, 130–145. doi: 10.1016/j.neurobiolaging.2017.10.009
Joffre, C., Nadjar, A., Lebbadi, M., Calon, F., and Laye, S. (2014). n-3 LCPUFA improves cognition: the young, the old and the sick. Prostaglandins Leukot. Essent. Fatty Acids 91, 1–20. doi: 10.1016/j.plefa.2014.05.001
Johnen, A., Pawlowski, M., and Duning, T. (2018). Distinguishing neurocognitive deficits in adult patients with NP-C from early onset Alzheimer’s dementia. Orphanet. J. Rare Dis. 13, 91.
Jones, L., Holmans, P. A., Hamshere, M. L., Harold, D., Moskvina, V., Ivanov, D., et al. (2010). Genetic evidence implicates the immune system and cholesterol metabolism in the aetiology of Alzheimer’s disease. PLoS One 5:e13950. doi: 10.1371/journal.pone.0013950
Jones, N. S., and Rebeck, G. W. (2018). The synergistic effects of APOE genotype and obesity on Alzheimer’s disease risk. Int. J. Mol. Sci. 20:63. doi: 10.3390/ijms20010063
Kaether, C., and Haass, C. (2004). A lipid boundary separates APP and secretases and limits amyloid beta-peptide generation. J. Cell Biol. 167, 809–812. doi: 10.1083/jcb.200410090
Kagedal, K., Kim, W. S., Appelqvist, H., Chan, S., Cheng, D., Agholme, L., et al. (2010). Increased expression of the lysosomal cholesterol transporter NPC1 in Alzheimer’s disease. Biochim. Biophys. Acta 1801, 831–838. doi: 10.1016/j.bbalip.2010.05.005
Kaiser, H. J., Orlowski, A., Rog, T., Nyholm, T. K., Chai, W., Feizi, T., et al. (2011). Lateral sorting in model membranes by cholesterol-mediated hydrophobic matching. Proc. Natl. Acad. Sci. U.S.A. 108, 16628–16633. doi: 10.1073/pnas.1103742108
Kalaria, R. N., and Harik, S. I. (1989). Abnormalities of the glucose transporter at the blood-brain barrier and in brain in Alzheimer’s disease. Prog. Clin. Biol. Res. 317, 415–421.
Kamboh, M. I., Minster, R. L., Demirci, F. Y., Ganguli, M., Dekosky, S. T., Lopez, O. L., et al. (2012). Association of CLU and PICALM variants with Alzheimer’s disease. Neurobiol. Aging 33, 518–521. doi: 10.1016/j.neurobiolaging.2010.04.015
Kaminsky, Y. G., Tikhonova, L. A., and Kosenko, E. A. (2015). Critical analysis of Alzheimer’s amyloid-beta toxicity to mitochondria. Front Biosci 20:173–197. doi: 10.2741/4304
Kang, J., and Rivest, S. (2012). Lipid metabolism and neuroinflammation in Alzheimer’s disease: a role for liver X receptors. Endocr. Rev. 33, 715–746. doi: 10.1210/er.2011-1049
Kao, Y. C., Ho, P. C., Tu, Y. K., Jou, I. M., and Tsai, K. J. (2020). Lipids and Alzheimer’s Disease. Int. J. Mol. Sci. 21:1505 doi: 10.3390/ijms21041505
Karamanos, Y., Gosselet, F., Dehouck, M. P., and Cecchelli, R. (2014). Blood-brain barrier proteomics: towards the understanding of neurodegenerative diseases. Arch. Med. Res. 45, 730–737. doi: 10.1016/j.arcmed.2014.11.008
Katt, M. E., Mayo, L. N., Ellis, S. E., Mahairaki, V., Rothstein, J. D., Cheng, L., et al. (2019). The role of mutations associated with familial neurodegenerative disorders on blood-brain barrier function in an iPSC model. Fluids Barriers CNS 16:20. doi: 10.1186/s12987-019-0139-4
Kelsch, W., Sim, S., and Lois, C. (2010). Watching synaptogenesis in the adult brain. Annu. Rev. Neurosci. 33, 131–149. doi: 10.1146/annurev-neuro-060909-153252
Kennedy, M. B. (2013). Synaptic signaling in learning and memory. Cold Spring Harb. Perspect. Biol. 8:a016824. doi: 10.1101/cshperspect.a016824
Khalil, A., Berrougui, H., Pawelec, G., and Fulop, T. (2012). Impairment of the ABCA1 and SR-BI-mediated cholesterol efflux pathways and HDL anti-inflammatory activity in Alzheimer’s disease. Mech. Ageing Dev. 133, 20–29. doi: 10.1016/j.mad.2011.11.008
Kim, M., Nevado-Holgado, A., Whiley, L., Snowden, S. G., Soininen, H., Kloszewska, I., et al. (2017). Association between plasma ceramides and phosphatidylcholines and hippocampal brain volume in late onset Alzheimer’s disease. J. Alzheimers Dis. 60, 809–817. doi: 10.3233/jad-160645
Kishimoto, Y., Agranoff, B. W., Radin, N. S., and Burton, R. M. (1969). Comparison of the fatty acids of lipids of subcellular brain fractions. J. Neurochem. 16, 397–404. doi: 10.1111/j.1471-4159.1969.tb10380.x
Kitazume, S., Tachida, Y., Oka, R., Shirotani, K., Saido, T. C., and Hashimoto, Y. (2001). Alzheimer’s beta-secretase, beta-site amyloid precursor protein-cleaving enzyme, is responsible for cleavage secretion of a Golgi-resident sialyltransferase. Proc. Natl. Acad. Sci. U.S.A. 98, 13554–13559. doi: 10.1073/pnas.241509198
Knebl, J., DeFazio, P., Clearfield, M. B., Little, L., McConathy, W. J., McPherson, R., et al. (1994). Plasma lipids and cholesterol esterification in Alzheimer’s disease. Mech. Ageing Dev. 73, 69–77. doi: 10.1016/0047-6374(94)90039-6
Kniewallner, K. M., Ehrlich, D., Kiefer, A., Marksteiner, J., and Humpel, C. (2015). Platelets in the Alzheimer’s disease brain: do they play a role in cerebral amyloid angiopathy? Curr. Neurovasc. Res. 12, 4–14. doi: 10.2174/1567202612666150102124703
Kohama, S. G., Rosene, D. L., and Sherman, L. S. (2012). Age-related changes in human and non-human primate white matter: from myelination disturbances to cognitive decline. Age 34, 1093–1110. doi: 10.1007/s11357-011-9357-7
Koizumi, K., Wang, G., and Park, L. (2016). Endothelial dysfunction and amyloid-beta-induced neurovascular alterations. Cell Mol. Neurobiol. 36, 155–165. doi: 10.1007/s10571-015-0256-9
Kojima, S., and Omori, M. (1992). Two-way cleavage of beta-amyloid protein precursor by multicatalytic proteinase. FEBS Lett. 304, 57–60. doi: 10.1016/0014-5793(92)80588-8
Kosicek, M., Zetterberg, H., Andreasen, N., Peter-Katalinic, J., and Hecimovic, S. (2012). Elevated cerebrospinal fluid sphingomyelin levels in prodromal Alzheimer’s disease. Neurosci. Lett. 516, 302–305. doi: 10.1016/j.neulet.2012.04.019
Kotani, S., Sakaguchi, E., Warashina, S., Matsukawa, N., Ishikura, Y., Kiso, Y., et al. (2006). Dietary supplementation of arachidonic and docosahexaenoic acids improves cognitive dysfunction. Neurosci. Res. 56, 159–164. doi: 10.1016/j.neures.2006.06.010
Kramer, S. D., Schutz, Y. B., Wunderli-Allenspach, H., Abbott, N. J., and Begley, D. J. (2002). Lipids in blood-brain barrier models in vitro II: influence of glial cells on lipid classes and lipid fatty acids. In Vitro Cell. Dev. Biol. Anim. 38, 566–571.
Kumar, A., and Singh, A. (2015). A review on mitochondrial restorative mechanism of antioxidants in Alzheimer’s disease and other neurological conditions. Front. Pharmacol. 6:206. doi: 10.3389/fphar.2015.00206
Kwon, H. J., Cha, M. Y., Kim, D., Kim, D. K., Soh, M., Shin, K., et al. (2016). Mitochondria-targeting ceria nanoparticles as antioxidants for Alzheimer’s disease. ACS Nano 10, 2860–2870. doi: 10.1021/acsnano.5b08045
Lamsa, R., Helisalmi, S., Herukka, S. K., Tapiola, T., Pirttila, T., Vepsalainen, S., et al. (2007). Study on the association between SOAT1 polymorphisms, Alzheimer’s disease risk and the level of CSF biomarkers. Dement. Geriatr. Cogn. Disord. 24, 146–150. doi: 10.1159/000105164
Laughlin, S. B., and Sejnowski, T. J. (2003). Communication in neuronal networks. Science 301, 1870–1874. doi: 10.1126/science.1089662
Leduc, V., Jasmin-Belanger, S., and Poirier, J. (2010). APOE and cholesterol homeostasis in Alzheimer’s disease. Trends Mol. Med. 16, 469–477. doi: 10.1016/j.molmed.2010.07.008
Lee, L. K., Shahar, S., Chin, A. V., and Yusoff, N. A. (2013). Docosahexaenoic acid-concentrated fish oil supplementation in subjects with mild cognitive impairment (MCI): a 12-month randomised, double-blind, placebo-controlled trial. Psychopharmacology 225, 605–612. doi: 10.1007/s00213-012-2848-0
Leonard, A. E., Kelder, B., Bobik, E. G., Chuang, L. T., Parker-Barnes, J. M., Thurmond, J. M., et al. (2000). cDNA cloning and characterization of human Delta5-desaturase involved in the biosynthesis of arachidonic acid. Biochem J. 347(Pt. 3), 719–724. doi: 10.1042/bj3470719
Lepara, O., Valjevac, A., Alajbegovic, A., Zaciragic, A., and Nakas-Icindic, E. (2009). Decreased serum lipids in patients with probable Alzheimer’s disease. Bosn. J. Basic Med. Sci. 9, 215–220. doi: 10.17305/bjbms.2009.2809
Lepping, R. J., Honea, R. A., Martin, L. E., Liao, K., Choi, I. Y., Lee, P., et al. (2019). Long-chain polyunsaturated fatty acid supplementation in the first year of life affects brain function, structure, and metabolism at age nine years. Dev. Psychobiol. 61, 5–16. doi: 10.1002/dev.21780
Leuti, A., Maccarrone, M., and Chiurchiu, V. (2019). Proresolving lipid mediators: endogenous modulators of oxidative stress. Oxid. Med. Cell. Longev. 2019:8107265. doi: 10.1155/2019/8107265
Levental, I., and Veatch, S. (2016). The continuing mystery of lipid rafts. J. Mol. Biol. 428, 4749–4764. doi: 10.1016/j.jmb.2016.08.022
Li, M. Z., Zheng, L. J., Shen, J., Li, X. Y., Zhang, Q., Bai, X., et al. (2018). SIRT1 facilitates amyloid beta peptide degradation by upregulating lysosome number in primary astrocytes. Neural Regen. Res. 13, 2005–2013. doi: 10.4103/1673-5374.239449
Li, X., Kan, H. Y., Lavrentiadou, S., Krieger, M., and Zannis, V. (2002). Reconstituted discoidal ApoE-phospholipid particles are ligands for the scavenger receptor BI. The amino-terminal 1-165 domain of ApoE suffices for receptor binding. J. Biol. Chem. 277, 21149–21157. doi: 10.1074/jbc.m200658200
Lim, G. P., Yang, F., Chu, T., Chen, P., Beech, W., Teter, B., et al. (2000). Ibuprofen suppresses plaque pathology and inflammation in a mouse model for Alzheimer’s disease. J. Neurosci. 20, 5709–5714. doi: 10.1523/jneurosci.20-15-05709.2000
Lin, Q., Cao, Y., and Gao, J. (2015). Decreased expression of the APOA1-APOC3-APOA4 gene cluster is associated with risk of Alzheimer’s disease. Drug. Des. Devel. Ther. 9, 5421–5431.
Lingwood, D., and Simons, K. (2010). Lipid rafts as a membrane-organizing principle. Science 327, 46–50. doi: 10.1126/science.1174621
Lingwood, D., Kaiser, H. J., Levental, I., and Simons, K. (2009). Lipid rafts as functional heterogeneity in cell membranes. Biochem. Soc. Trans. 37, 955–960. doi: 10.1042/bst0370955
Liu, K., Liu, Y., Xu, Y., Nandakumar, K. S., Shen, X., Lin, J., et al. (2019). Regulatory role of Golgi brefeldin a resistance factor-1 in amyloid precursor protein trafficking, cleavage and Abeta formation. J. Cell Biochem. 120, 15604–15615. doi: 10.1002/jcb.28827
Liu, L., MacKenzie, K. R., Putluri, N., Maletic-Savatic, M., and Bellen, H. J. (2017). The glia-neuron lactate shuttle and elevated ROS promote lipid synthesis in neurons and lipid droplet accumulation in glia via APOE/D. Cell Metab. 26, 719–737e716.
Loef, M., and Walach, H. (2013). The Omega-6/Omega-3 ratio and dementia or cognitive decline: a systematic review on human studies and biological evidence. J. Nutr. Gerontol. Geriatr. 32, 1–23. doi: 10.1080/21551197.2012.752335
Loera-Valencia, R., Goikolea, J., Parrado-Fernandez, C., Merino-Serrais, P., and Maioli, S. (2019). Alterations in cholesterol metabolism as a risk factor for developing Alzheimer’s disease: potential novel targets for treatment. J. Steroid. Biochem. Mol. Biol. 190, 104–114. doi: 10.1016/j.jsbmb.2019.03.003
Lohner, S., Fekete, K., Marosvölgyi, T., and Decsi, T. (2013). Gender differences in the long-chain polyunsaturated fatty acid status: systematic review of 51 publications. Ann. Nutr. Metabol. 62, 98–112. doi: 10.1159/000345599
Lopez, L. B., Kritz-Silverstein, D., and Barrett Connor, E. (2011). High dietary and plasma levels of the omega-3 fatty acid docosahexaenoic acid are associated with decreased dementia risk: the rancho bernardo study. J. Nutr. Health Aging 15, 25–31. doi: 10.1007/s12603-011-0009-5
Louveau, A., Smirnov, I., Keyes, T. J., Eccles, J. D., Rouhani, S. J., Peske, J. D., et al. (2015). Structural and functional features of central nervous system lymphatic vessels. Nature 523, 337–341. doi: 10.1038/nature14432
Lovinger, D. M. (2008). Communication networks in the brain: neurons, receptors, neurotransmitters, and alcohol. Alcohol. Res. Health 31, 196–214.
Lucatelli, J. F., Barros, A. C., Silva, V. K., Machado Fda, S., Constantin, P. C., Dias, A. A., et al. (2011). Genetic influences on Alzheimer’s disease: evidence of interactions between the genes APOE, APOC1 and ACE in a sample population from the South of Brazil. Neurochem. Res. 36, 1533–1539. doi: 10.1007/s11064-011-0481-7
Luo, C., Ren, H., Yao, X., Shi, Z., Liang, F., Kang, J. X., et al. (2018). Enriched brain Omega-3 polyunsaturated fatty acids confer neuroprotection against microinfarction. EBioMedicine 32, 50–61. doi: 10.1016/j.ebiom.2018.05.028
MacDonald-Wicks, L., McEvoy, M., Magennis, E., Schofield, P. W., Patterson, A. J., and Zacharia, K. (2019). Dietary long-chain fatty acids and cognitive performance in older australian adults. Nutrients 11:711. doi: 10.3390/nu11040711
Mackic, J. B., Stins, M., McComb, J. G., Calero, M., Ghiso, J., Kim, K. S., et al. (1998). Human blood-brain barrier receptors for Alzheimer’s amyloid-beta 1- 40. asymmetrical binding, endocytosis, and transcytosis at the apical side of brain microvascular endothelial cell monolayer. J. Clin. Invest. 102, 734–743. doi: 10.1172/jci2029
Maclean, F. L., Horne, M. K., Williams, R. J., and Nisbet, D. R. (2018). Review: biomaterial systems to resolve brain inflammation after traumatic injury. APL Bioeng. 2:021502. doi: 10.1063/1.5023709
Mahley, R. W. (2016). Central nervous system lipoproteins: ApoE and regulation of cholesterol metabolism. Arterioscler. Thromb. Vasc. Biol. 36, 1305–1315. doi: 10.1161/atvbaha.116.307023
Mancuso, C., Bates, T. E., Butterfield, D. A., Calafato, S., Cornelius, C., De Lorenzo, A., et al. (2007). Natural antioxidants in Alzheimer’s disease. Expert. Opin. Investig. Drugs 16, 1921–1931.
Marchi, C., Adorni, M. P., Caffarra, P., Ronda, N., Spallazzi, M., Barocco, F., et al. (2019). ABCA1- and ABCG1-mediated cholesterol efflux capacity of cerebrospinal fluid is impaired in Alzheimer’s disease. J. Lipid Res. 60, 1449–1456. doi: 10.1194/jlr.p091033
Marquer, C., Devauges, V., Cossec, J. C., Liot, G., Lecart, S., Saudou, F., et al. (2011). Local cholesterol increase triggers amyloid precursor protein-Bace1 clustering in lipid rafts and rapid endocytosis. FASEB J. 25, 1295–1305. doi: 10.1096/fj.10-168633
Martin, R. E., and Bazan, N. G. (1992). Changing fatty acid content of growth cone lipids prior to synaptogenesis. J. Neurochem. 59, 318–325. doi: 10.1111/j.1471-4159.1992.tb08906.x
Martin, T. F. (2000). Racing lipid rafts for synaptic-vesicle formation. Nat. Cell. Biol. 2, E9–E11.
Martinez-Frailes, C., Di Lauro, C., Bianchi, C., de Diego-Garcia, L., Sebastian-Serrano, A., Bosca, L., et al. (2019). Amyloid peptide induced neuroinflammation increases the p2x7 receptor expression in microglial cells, impacting on its functionality. Front. Cell Neurosci. 13:143. doi: 10.3389/fncel.2019.00143
Martins, M. J., Constancia, M., Neves, D., and Simm, A. (2017). Biomarkers of aging: from cellular senescence to age-associated diseases. Oxid. Med. Cell Longev. 2017:7280690. doi: 10.1155/2017/7280690
Matthews, K. A., Xu, W., Gaglioti, A. H., Holt, J. B., Croft, J. B., Mack, D., et al. (2019). Racial and ethnic estimates of Alzheimer’s disease and related dementias in the United States (2015–2060) in adults aged =65 years. Alzheimers Dement. 15, 17–24. doi: 10.1016/j.jalz.2018.06.3063
Mauch, D. H., Nagler, K., Schumacher, S., Goritz, C., Muller, E. C., Otto, A., et al. (2001). CNS synaptogenesis promoted by glia-derived cholesterol. Science 294, 1354–1357. doi: 10.1126/science.294.5545.1354
Maulik, M., Peake, K., Chung, J., Wang, Y., Vance, J. E., and Kar, S. (2015). APP overexpression in the absence of NPC1 exacerbates metabolism of amyloidogenic proteins of Alzheimer’s disease. Hum. Mol. Genet. 24, 7132–7150.
Maysinger, D., Ji, J., Moquin, A., Hossain, S., Hancock, M. A., Zhang, I., et al. (2018). Dendritic polyglycerol sulfates in the prevention of synaptic loss and mechanism of action on glia. ACS Chem. Neurosci. 9, 260–271. doi: 10.1021/acschemneuro.7b00301
McClean, P. L., Jalewa, J., and Holscher, C. (2015). Prophylactic liraglutide treatment prevents amyloid plaque deposition, chronic inflammation and memory impairment in APP/PS1 mice. Behav. Brain. Res. 293, 96–106. doi: 10.1016/j.bbr.2015.07.024
McNamara, R. K., Able, J., Jandacek, R., Rider, T., Tso, P., Eliassen, J. C., et al. (2010). Docosahexaenoic acid supplementation increases prefrontal cortex activation during sustained attention in healthy boys: a placebo-controlled, dose-ranging, functional magnetic resonance imaging study. Am. J. Clin. Nutr. 91, 1060–1067. doi: 10.3945/ajcn.2009.28549
McNamara, R. K., Asch, R. H., Lindquist, D. M., and Krikorian, R. (2018). Role of polyunsaturated fatty acids in human brain structure and function across the lifespan: an update on neuroimaging findings. Prostaglandins Leukot. Essent. Fatty Acids 136, 23–34. doi: 10.1016/j.plefa.2017.05.001
Mecca, A. P., Barcelos, N. M., Wang, S., Bruck, A., Nabulsi, N., Planeta-Wilson, B., et al. (2018). Cortical beta-amyloid burden, gray matter, and memory in adults at varying APOE epsilon4 risk for Alzheimer’s disease. Neurobiol. Aging 61, 207–214. doi: 10.1016/j.neurobiolaging.2017.09.027
Melo, R. C., D’Avila, H., Wan, H. C., Bozza, P. T., Dvorak, A. M., and Weller, P. F. (2011). Lipid bodies in inflammatory cells: structure, function, and current imaging techniques. J. Histochem. Cytochem. 59, 540–556. doi: 10.1369/0022155411404073
Mergenthaler, P., Lindauer, U., Dienel, G. A., and Meisel, A. (2013). Sugar for the brain: the role of glucose in physiological and pathological brain function. Trends Neurosci. 36, 587–597. doi: 10.1016/j.tins.2013.07.001
Merino-Zamorano, C., Fernandez-de Retana, S., Montanola, A., Batlle, A., Saint-Pol, J., Mysiorek, C., et al. (2016). Modulation of amyloid-beta1-40 transport by ApoA1 and ApoJ across an in vitro model of the blood-brain barrier. J. Alzheimers Dis. 53, 677–691. doi: 10.3233/jad-150976
Mesa-Herrera, F., Taoro-Gonzalez, L., Valdes-Baizabal, C., Diaz, M., and Marin, R. (2019). Lipid and lipid raft alteration in aging and neurodegenerative diseases: a window for the development of new biomarkers. Int. J. Mol. Sci. 20:3810. doi: 10.3390/ijms20153810
Mielke, M. (2018). Sex and gender differences in alzheimer’s disease dementia. Psychiatric. Time 35, 14–17.
Mietelska-Porowska, A., and Wojda, U. (2017). T lymphocytes and inflammatory mediators in the interplay between brain and blood in Alzheimer’s disease: potential pools of new biomarkers. J. Immunol. Res. 2017, 4626540. doi: 10.1155/2017/4626540
Mills, J., and Reiner, P. B. (1999). Regulation of amyloid precursor protein cleavage. J. Neurochem. 72, 443–460. doi: 10.1046/j.1471-4159.1999.0720443.x
Mobraten, K., Haug, T. M., Kleiveland, C. R., and Lea, T. (2013). Omega-3 and omega-6 PUFAs induce the same GPR120-mediated signalling events, but with different kinetics and intensity in Caco-2 cells. Lipids Health. Dis. 12:101. doi: 10.1186/1476-511X-12-101
Mochel, F. (2018). Lipids and synaptic functions. J. Inherit. Metab. Dis. 41, 1117–1122. doi: 10.1007/s10545-018-0204-1
Mohaibes, R. J., Fiol-deRoque, M. A., Torres, M., Ordinas, M., Lopez, D. J., Castro, J. A., et al. (2017). The hydroxylated form of docosahexaenoic acid (DHA-H) modifies the brain lipid composition in a model of Alzheimer’s disease, improving behavioral motor function and survival. Biochim. Biophys. Acta Biomembr. 1859, 1596–1603. doi: 10.1016/j.bbamem.2017.02.020
Molander-Melin, M., Blennow, K., Bogdanovic, N., Dellheden, B., Mansson, J. E., and Fredman, P. (2005). Structural membrane alterations in Alzheimer brains found to be associated with regional disease development; increased density of gangliosides GM1 and GM2 and loss of cholesterol in detergent-resistant membrane domains. J. Neurochem. 92, 171–182. doi: 10.1111/j.1471-4159.2004.02849.x
Montani, L., and Suter, U. (2018). Building lipids for myelin. Aging 10, 861–862. doi: 10.18632/aging.101458
Montanini, I., Gatti, C., Woelk, H., and Porcellati, S. (1983). The influence of polyunsaturated phosphatidylcholine on brain lipid synthesis during aging. Farmaco. Sci. 38, 376–382.
Montoliu-Gaya, L., Mulder, S. D., Herrebout, M. A. C., Baayen, J. C., Villegas, S., and Veerhuis, R. (2018). Abeta-oligomer uptake and the resulting inflammatory response in adult human astrocytes are precluded by an anti-Abeta single chain variable fragment in combination with an apoE mimetic peptide. Mol. Cell Neurosci. 89, 49–59. doi: 10.1016/j.mcn.2018.03.015
Morris, J. C., Schindler, S., and McCue, L. M. (2019). Assessment of racial disparities in biomarkers for Alzheimer disease. JAMA Neurol. 76, 264–273.
Morris, M. C., and Tangney, C. C. (2014). Dietary fat composition and dementia risk. Neurobiol. Aging 35(Suppl. 2), S59–S64.
Morris, M. C., Evans, D. A., Bienias, J. L., Tangney, C. C., Bennett, D. A., Wilson, R. S., et al. (2003). Consumption of fish and n-3 fatty acids and risk of incident Alzheimer disease. Arch. Neurol. 60, 940–946.
Moura, R. P., Martins, C., Pinto, S., Sousa, F., and Sarmento, B. (2019). Blood-brain barrier receptors and transporters: an insight on their function and how to exploit them through nanotechnology. Expert. Opin. Drug. Deliv. 16, 271–285. doi: 10.1080/17425247.2019.1583205
Mukadam, A. S., Breusegem, S. Y., and Seaman, M. N. J. (2018). Analysis of novel endosome-to-golgi retrieval genes reveals a role for PLD3 in regulating endosomal protein sorting and amyloid precursor protein processing. Cell Mol. Life Sci. 75, 2613–2625. doi: 10.1007/s00018-018-2752-9
Muldoonm, M. F., Ryan, C. M., Sheu, L., Yao, J. K., Conklin, S. M., and Manuck, S. B. (2010). Serum phospholipid docosahexaenonic acid is associated with cognitive functioning during middle adulthood. J. Nutr. 140, 848–853. doi: 10.3945/jn.109.119578
Muszynski, P., Kulczynska-Przybik, A., Borawska, R., Litman-Zawadzka, A., Slowik, A., Klimkowicz-Mrowiec, A., et al. (2017). The relationship between markers of inflammation and degeneration in the central nervous system and the blood-brain barrier impairment in Alzheimer’s disease. J. Alzheimers Dis. 59, 903–912. doi: 10.3233/jad-170220
Nägga, K., Gustavsson, A.-M., Stomrud, E., Lindqvist, D., van Westen, D., Blennow, K., et al. (2018). Increased midlife triglycerides predict brain β-amyloid and tau pathology 20 years later. Neurology 90, e73–e81. doi: 10.1212/WNL.0000000000004749
Nasaruddin, M. L., Holscher, C., Kehoe, P., Graham, S. F., and Green, B. D. (2016). Wide-ranging alterations in the brain fatty acid complement of subjects with late Alzheimer’s disease as detected by GC-MS. Am. J. Transl. Res. 8, 154–165.
Nasrabady, S. E., Rizvi, B., Goldman, J. E., and Brickman, A. M. (2018). White matter changes in Alzheimer’s disease: a focus on myelin and oligodendrocytes. Acta Neuropathol. Commun. 6:22. doi: 10.1186/s40478-018-0515-3
Nation, D. A., Sweeney, M. D., Montagne, A., Sagare, A. P., D’Orazio, L. M., Pachicano, M., et al. (2019). Blood-brain barrier breakdown is an early biomarker of human cognitive dysfunction. Nat. Med. 25, 270–276.
Nehlig, A. (2004). Brain uptake and metabolism of ketone bodies in animal models. Prostaglandins Leukot. Essent. Fatty Acids 70, 265–275. doi: 10.1016/j.plefa.2003.07.006
Nelson, A. R., Sagare, A. P., and Zlokovic, B. V. (2017). Role of clusterin in the brain vascular clearance of amyloid-beta. Proc. Natl. Acad. Sci. U.S.A. 114, 8681–8682. doi: 10.1073/pnas.1711357114
Nelson, A. R., Sweeney, M. D., Sagare, A. P., and Zlokovic, B. V. (2016). Neurovascular dysfunction and neurodegeneration in dementia and Alzheimer’s disease. Biochim. Biophys. Acta 1862, 887–900. doi: 10.1016/j.bbadis.2015.12.016
Nicholls, D. G., and Budd, S. L. (2000). Mitochondria and neuronal survival. Physiol. Rev. 80, 315–360. doi: 10.1152/physrev.2000.80.1.315
Ntambi, J. M. (2019). Highlighting inflammation and lipid metabolism. Biochem. Biophys. Res. Commun. 520, 688–689. doi: 10.1016/j.bbrc.2019.10.014
Nunan, J., and Small, D. H. (2000). Regulation of APP cleavage by alpha-, beta- and gamma-secretases. FEBS Lett. 483, 6–10. doi: 10.1016/s0014-5793(00)02076-7
Nunes, V. S., Cazita, P. M., Catanozi, S., Nakandakare, E. R., and Quintao, E. C. R. (2018). Decreased content, rate of synthesis and export of cholesterol in the brain of apoE knockout mice. J. Bioenerg. Biomembr. 50, 283–287. doi: 10.1007/s10863-018-9757-9
Nuutinen, T., Huuskonen, J., Suuronen, T., Ojala, J., Miettinen, R., and Salminen, A. (2007). Amyloid-beta 1-42 induced endocytosis and clusterin/apoJ protein accumulation in cultured human astrocytes. Neurochem. Int. 50, 540–547. doi: 10.1016/j.neuint.2006.11.002
Nuutinen, T., Suuronen, T., Kyrylenko, S., Huuskonen, J., and Salminen, A. (2005). Induction of clusterin/apoJ expression by histone deacetylase inhibitors in neural cells. Neurochem. Int. 47, 528–538. doi: 10.1016/j.neuint.2005.07.007
Oberstein, T. J., Taha, L., Spitzer, P., Hellstern, J., Herrmann, M., Kornhuber, J., et al. (2018). Imbalance of circulating Th17 and regulatory T cells in Alzheimer’s disease: a case control study. Front. Immunol. 9:1213. doi: 10.3389/fimmu.2018.01213
O’Brien, J. S., and Sampson, E. L. (1965). Lipid composition of the normal human brain: gray matter, white matter, and myelin. J. Lipid. Res. 6, 537–544.
O’Brien, R. J., and Wong, P. C. (2011). Amyloid precursor protein processing and Alzheimer’s disease. Annu. Rev. Neurosci. 34, 185–204.
O’Donoghue, M. C., Murphy, S. E., Zamboni, G., Nobre, A. C., and Mackay, C. E. (2018). APOE genotype and cognition in healthy individuals at risk of Alzheimer’s disease: a review. Cortex 104, 103–123. doi: 10.1016/j.cortex.2018.03.025
Ong, W. Y., Farooqui, T., Kokotos, G., and Farooqui, A. A. (2015). Synthetic and natural inhibitors of phospholipases A2: their importance for understanding and treatment of neurological disorders. ACS Chem. Neurosci. 6, 814–831.
Ouellet, M., Emond, V., Chen, C. T., Julien, C., Bourasset, F., Oddo, S., et al. (2009). Diffusion of docosahexaenoic and eicosapentaenoic acids through the blood-brain barrier: an in situ cerebral perfusion study. Neurochem. Int. 55, 476–482. doi: 10.1016/j.neuint.2009.04.018
Palacios, G., Palacios, J. M., Mengod, G., and Frey, P. (1992). Beta-amyloid precursor protein localization in the Golgi apparatus in neurons and oligodendrocytes. An immunocytochemical structural and ultrastructural study in normal and axotomized neurons. Brain Res. Mol. Brain Res. 15, 195–206. doi: 10.1016/0169-328x(92)90109-o
Palmisano, B. T., Zhu, L., Eckel, R. H., and Staffor, J. M. (2018). Sex differences in lipid and lipoprotein metabolism. Mol. Metab. 15, 45–55. doi: 10.1016/j.molmet.2018.05.008
Paloczi, J., Varga, Z. V., Hasko, G., and Pacher, P. (2018). Neuroprotection in oxidative stress-related neurodegenerative diseases: role of endocannabinoid system modulation. Antioxid. Redox. Signal. 29, 75–108. doi: 10.1089/ars.2017.7144
Pan, Y., Khalil, H., and Nicolazzo, J. A. (2015). The impact of docosahexaenoic acid on Alzheimer’s disease: is there a role of the blood-brain barrier? Curr. Clin. Pharmacol. 10, 222–241. doi: 10.2174/157488471003150820151532
Pan, Y., Short, J. L., Choy, K. H., Zeng, A. X., Marriott, P. J., Owada, Y., et al. (2016). Fatty acid-binding protein 5 at the blood-brain barrier regulates endogenous brain docosahexaenoic acid levels and cognitive function. J. Neurosci. 36, 11755–11767. doi: 10.1523/jneurosci.1583-16.2016
Pang, C., Yang, H., Hu, B., Wang, S., Chen, M., Cohen, D. S., et al. (2019). Identification and Analysis of Alzheimer’s Candidate Genes by an Amplitude Deviation Algorithm. J. Alzheimers Dis. Parkinsonism 9:460. doi: 10.4172/2161-0460.1000460
Papassotiropoulos, A., Wollmer, M. A., Tsolaki, M., Brunner, F., Molyva, D., Lutjohann, D., et al. (2005). A cluster of cholesterol-related genes confers susceptibility for Alzheimer’s disease. J. Clin. Psychiatry 66, 940–947. doi: 10.4088/jcp.v66n0720
Park, L., Wang, G., Moore, J., Girouard, H., Zhou, P., Anrather, J., et al. (2014). The key role of transient receptor potential melastatin-2 channels in amyloid-beta-induced neurovascular dysfunction. Nat. Commun. 5:5318. doi: 10.1038/ncomms6318
Patel, M. S., Johnson, C. A., Rajan, R., and Owen, O. E. (1975). The metabolism of ketone bodies in developing human brain: development of ketone-body-utilizing enzymes and ketone bodies as precursors for lipid synthesis. J. Neurochem. 25, 905–908. doi: 10.1111/j.1471-4159.1975.tb04428.x
Periyasamy, S., Sathya, M., Karthick, C., Kandasamy, M., Shanmugaapriya, S., Tamilselvan, J., et al. (2017). Association studies of specific cholesterol related genes (APOE, LPL, and CETP) with lipid profile and memory function: a correlative study among rural and tribal population of dharmapuri district, India. J. Alzheimers Dis. 60, S195–S207.
Phillips, M. A., Childs, C. E., Calder, P. C., and Rogers, P. J. (2015). No effect of Omega-3 fatty acid supplementation on cognition and mood in individuals with cognitive impairment and probable alzheimer’s disease: a randomised controlled trial. Int. J. Mol. Sci. 16, 24600–24613. doi: 10.3390/ijms161024600
Picard, M., and McEwen, B. S. (2014). Mitochondria impact brain function and cognition. Proc. Natl. Acad. Sci. U.S.A. 111, 7–8. doi: 10.1073/pnas.1321881111
Piomelli, D., Astarita, G., and Rapaka, R. (2007). A neuroscientist’s guide to lipidomics. Nat. Rev. Neurosci. 8, 743–754.
Piro, J. R., Benjamin, D. I., Duerr, J. M., Pi, Y., Gonzales, C., Wood, K. M., et al. (2012). A dysregulated endocannabinoid-eicosanoid network supports pathogenesis in a mouse model of Alzheimer’s disease. Cell Rep. 1, 617–623. doi: 10.1016/j.celrep.2012.05.001
Pitas, R. E., Boyles, J. K., Lee, S. H., Foss, D., and Mahley, R. W. (1987). Astrocytes synthesize apolipoprotein E and metabolize apolipoprotein E-containing lipoproteins. Biochim. Biophys. Acta 917, 148–161. doi: 10.1016/0005-2760(87)90295-5
Prasad, M. R., Lovell, M. A., Yatin, M., Dhillon, H., and Markesbery, W. R. (1998). Regional membrane phospholipid alterations in Alzheimer’s disease. Neurochem. Res. 23, 81–88.
Prendecki, M., Florczak-Wyspianska, J., Kowalska, M., Ilkowski, J., Grzelak, T., Bialas, K., et al. (2018). Biothiols and oxidative stress markers and polymorphisms of TOMM40 and APOC1 genes in Alzheimer’s disease patients. Oncotarget 9, 35207–35225. doi: 10.18632/oncotarget.26184
Preston, J. E., Joan Abbott, N., and Begley, D. J. (2014). Transcytosis of macromolecules at the blood-brain barrier. Adv. Pharmacol. 71, 147–163. doi: 10.1016/bs.apha.2014.06.001
Prevost, M., and Kocher, J. P. (1999). Structural characterization by computer experiments of the lipid-free LDL-receptor-binding domain of apolipoprotein E. Protein Eng. 12, 475–483. doi: 10.1093/protein/12.6.475
Quinn, J. F., Raman, R., Thomas, R. G., Yurko-Mauro, K., Nelson, E. B., Van Dyck, C., et al. (2010). Docosahexaenoic acid supplementation and cognitive decline in Alzheimer disease: a randomized trial. JAMA 304, 1903–1911.
Quintanilla, R. A., Dolan, P. J., Jin, Y. N., and Johnson, G. V. (2012). Truncated tau and Abeta cooperatively impair mitochondria in primary neurons. Neurobiol. Aging 33, e625–e635.
Raj, D., Yin, Z., Breur, M., Doorduin, J., Holtman, I. R., Olah, M., et al. (2017). Increased white matter inflammation in aging- and Alzheimer’s disease brain. Front. Mol. Neurosci. 10:206. doi: 10.3389/fnmol.2017.00206
Ramasamy, I. (2014). Recent advances in physiological lipoprotein metabolism. Clin. Chem. Lab. Med. 52, 1695–1727.
Rangaraju, V., Lewis, T. L. Jr., Hirabayashi, Y., Bergami, M., Motori, E., Cartoni, R., et al. (2019). Pleiotropic mitochondria: the influence of mitochondria on neuronal development and Disease. J. Neurosci. 39, 8200–8208. doi: 10.1523/jneurosci.1157-19.2019
Ratnakumar, A., Zimmerman, S. E., Jordan, B. A., and Mar, J. C. (2019). Estrogen activates Alzheimer’s disease genes. Alzheimers Dement 5, 906–917. doi: 10.1016/j.trci.2019.09.004
Reitz, C. (2012). Dyslipidemia and dementia: current epidemiology, genetic evidence and mechanisms behind the associations. J. Alzheimers Dis. 30, S127–S145.
Riedel, B. C., Thompson, P. M., and Brinton, R. D. (2016). Age, APOE and sex: triad of risk of Alzheimer’s disease. J. Steroid. Biochem. Mol. Biol. 160, 134–147. doi: 10.1016/j.jsbmb.2016.03.012
Roberts, S. B., Ripellino, J. A., Ingalls, K. M., Robakis, N. K., and Felsenstein, K. M. (1994). Non-amyloidogenic cleavage of the beta-amyloid precursor protein by an integral membrane metalloendopeptidase. J. Biol. Chem. 269, 3111–3116.
Robinson, J. L., Lee, E. B., Xie, S. X., Rennert, L., Suh, E., Bredenberg, C., et al. (2018). Neurodegenerative disease concomitant proteinopathies are prevalent, age-related and APOE4-associated. Brain 141, 2181–2193. doi: 10.1093/brain/awy146
Rosenberg, A., Mangialasche, F., Ngandu, T., Solomon, A., and Kivipelto, M. (2020). Multidomain interventions to prevent cognitive impairment, Alzheimer’s disease, and dementia: from FINGER to world-wide FINGERS. J. Prev. Alzheimers Dis. 7, 29–36.
Rutten, B. P., Steinbusch, H. W., Korr, H., and Schmitz, C. (2002). Antioxidants and Alzheimer’s disease: from bench to bedside (and back again). Curr. Opin. Clin. Nutr. Metab. Care 5, 645–651.
Saftig, P., Peters, C., von Figura, K., Craessaerts, K., Van Leuven, F., and De Strooper, B. (1996). Amyloidogenic processing of human amyloid precursor protein in hippocampal neurons devoid of cathepsin D. J. Biol. Chem. 271, 27241–27244. doi: 10.1074/jbc.271.44.27241
Salinas, E. (2009). Neuronal communication: a detailed balancing act. Nat. Neurosci. 12, 372–374. doi: 10.1038/nn0409-372
Sarrafpour, S., Ormseth, C., Chiang, A., Arakaki, X., Harrington, M., and Fonteh, A. (2019). Lipid metabolism in late-onset Alzheimer’s disease differs from patients presenting with other dementia phenotypes. Int. J. Environ. Res. Public Health 16:1995. doi: 10.3390/ijerph16111995
Sastre, M., Walter, J., and Gentleman, S. M. (2008). Interactions between APP secretases and inflammatory mediators. J. Neuroinflammation 5:25. doi: 10.1186/1742-2094-5-25
Schmitt, F., Hussain, G., Dupuis, L., Loeffler, J. P., and Henriques, A. (2014). A plural role for lipids in motor neuron diseases: energy, signaling and structure. Front. Cell Neurosci. 8:25. doi: 10.3389/fncel.2014.00025
Schmitt, S., Castelvetri, L. C., and Simons, M. (2015). Metabolism and functions of lipids in myelin. Biochim. Biophys. Acta 1851, 999–1005. doi: 10.1016/j.bbalip.2014.12.016
Schmitz, G. E. J. (2008). The opposing effects of n-3 and n-6 fatty acids. Prog. Lipid Res. 47, 147–155. doi: 10.1016/j.plipres.2007.12.004
Schmitz, G., and Muller, G. (1991). Structure and function of lamellar bodies, lipid-protein complexes involved in storage and secretion of cellular lipids. J. Lipid Res. 32, 1539–1570.
Schonfeld, P., and Reiser, G. (2017). Brain energy metabolism spurns fatty acids as fuel due to their inherent mitotoxicity and potential capacity to unleash neurodegeneration. Neurochem. Int. 109, 68–77. doi: 10.1016/j.neuint.2017.03.018
Scialo, F., Sriram, A., Fernandez-Ayala, D., Gubina, N., Lohmus, M., Nelson, G., et al. (2016). Mitochondrial ROS produced via reverse electron transport extend animal lifespan. Cell Metab. 23, 725–734. doi: 10.1016/j.cmet.2016.03.009
Sebastiao, A. M., Colino-Oliveira, M., Assaife-Lopes, N., Dias, R. B., and Ribeiro, J. A. (2013). Lipid rafts, synaptic transmission and plasticity: impact in age-related neurodegenerative diseases. Neuropharmacology 64, 97–107. doi: 10.1016/j.neuropharm.2012.06.053
Segi-Nishida, E. (2014). Double function of MFSD2A transporter at the blood-brain barrier. Nihon Yakurigaku Zasshi 144:253. doi: 10.1254/fpj.144.253
Serhan, C. N. (2010). Novel lipid mediators and resolution mechanisms in acute inflammation: to resolve or not? Am. J. Pathol. 177, 1576–1591. doi: 10.2353/ajpath.2010.100322
Serhan, C. N., Chiang, N., and Dalli, J. (2015). The resolution code of acute inflammation: novel pro-resolving lipid mediators in resolution. Semin. Immunol. 27, 200–215. doi: 10.1016/j.smim.2015.03.004
Serhan, C. N., Chiang, N., and Dalli, J. (2018). New pro-resolving n-3 mediators bridge resolution of infectious inflammation to tissue regeneration. Mol. Aspects. Med. 64, 1–17. doi: 10.1016/j.mam.2017.08.002
Sherwin, B. (2012). Estrogen and cognitive functioning in women: lessons we have learned. Behav. Neurosci. 126, 123–127. doi: 10.1037/a0025539
Shi, H., Medway, C., Bullock, J., Brown, K., Kalsheker, N., and Morgan, K. (2010). Analysis of genome-wide association study (GWAS) data looking for replicating signals in Alzheimer’s disease (AD). Int. J. Mol. Epidemiol. Genet. 1, 53–66.
Shibata, M., Yamada, S., Kumar, S. R., Calero, M., Bading, J., Frangione, B., et al. (2000). Clearance of Alzheimer’s amyloid-ss(1-40) peptide from brain by LDL receptor-related protein-1 at the blood-brain barrier. J. Clin. Invest. 106, 1489–1499. doi: 10.1172/jci10498
Shibata, N., Kawarai, T., Lee, J. H., Lee, H. S., Shibata, E., Sato, C., et al. (2006). Association studies of cholesterol metabolism genes (CH25H, ABCA1 and CH24H) in Alzheimer’s disease. Neurosci. Lett. 391, 142–146. doi: 10.1016/j.neulet.2005.08.048
Shibata, N., Nagata, T., Shinagawa, S., Ohnuma, T., Shimazaki, H., Komatsu, M., et al. (2013). Genetic association between APOA1 and APOD polymorphisms and Alzheimer’s disease in a Japanese population. J. Neural. Transm. 120, 1599–1603. doi: 10.1007/s00702-013-1036-7
Shibuya, Y., Chang, C. C., and Chang, T. Y. (2015). ACAT1/SOAT1 as a therapeutic target for Alzheimer’s disease. Future Med. Chem. 7, 2451–2467. doi: 10.4155/fmc.15.161
Shimokawa, M., Yanagisawa, K., Nishiye, H., and Miyatake, T. (1993). Identification of amyloid precursor protein in synaptic plasma membrane. Biochem. Biophys. Res. Commun. 196, 240–244. doi: 10.1006/bbrc.1993.2240
Shinohara, M., Tachibana, M., Kanekiyo, T., and Bu, G. (2017). Role of LRP1 in the pathogenesis of Alzheimer’s disease: evidence from clinical and preclinical studies. J. Lipid Res. 58, 1267–1281. doi: 10.1194/jlr.r075796
Shinto, L., Quinn, J., Montine, T., Dodge, H. H., Woodward, W., Baldauf-Wagner, S., et al. (2014). A randomized placebo-controlled pilot trial of omega-3 fatty acids and alpha lipoic acid in Alzheimer’s disease. J. Alzheimers Dis. 38, 111–120. doi: 10.3233/jad-130722
Shuai, P., Liu, Y., Lu, W., Liu, Q., Li, T., and Gong, B. (2015). Genetic associations of CLU rs9331888 polymorphism with Alzheimer’s disease: a meta-analysis. Neurosci. Lett. 591, 160–165. doi: 10.1016/j.neulet.2015.02.040
Siegel, G. J. (1999). Basic Neurochemistry : Molecular, Cellular, and Medical Aspects, 6th Edn. Philadelphia: Lippincott Williams & Wilkins.
Simon, A. K., Hollander, G. A., and McMichael, A. (2015). Evolution of the immune system in humans from infancy to old age. Proc. Biol. Sci. 282:20143085. doi: 10.1098/rspb.2014.3085
Simons, M., and Nave, K. A. (2015). Oligodendrocytes: myelination and axonal support. Cold Spring Harb. Perspect. Biol. 8:a020479. doi: 10.1101/cshperspect.a020479
Simopoulos, A. P. (2006). Evolutionary aspects of diet, the omega-6/omega-3 ratio and genetic variation: nutritional implications for chronic diseases. Biomed. Pharmacother. 60, 502–507. doi: 10.1016/j.biopha.2006.07.080
Sinclair, A. J., Begg, D., Mathai, M., and Weisinger, R. S. (2007). Omega 3 fatty acids and the brain: review of studies in depression. Asia Pac. J. Clin. Nutr. 16(Suppl. 1), 391–397.
Sisodia, S. S. (1992). Beta-amyloid precursor protein cleavage by a membrane-bound protease. Proc. Natl. Acad. Sci. U.S.A. 89, 6075–6079. doi: 10.1073/pnas.89.13.6075
Skaper, S. D., and Di Marzo, V. (2012). Endocannabinoids in nervous system health and disease: the big picture in a nutshell. Philos. Trans. R. Soc. Lond. B Biol. Sci. 367, 3193–3200. doi: 10.1098/rstb.2012.0313
Smach, M. A., Edziri, H., Charfeddine, B., Ben Othman, L., Lammouchi, T., Ltaief, A., et al. (2011). Polymorphism in apoA1 influences high-density lipoprotein cholesterol levels but is not a major risk factor of Alzheimer’s disease. Dement. Geriatr. Cogn. Dis. Extra. 1, 249–257. doi: 10.1159/000329910
Snowden, S. G., Ebshiana, A. A., Hye, A., An, Y., Pletnikova, O., O’Brien, R., et al. (2017). Association between fatty acid metabolism in the brain and Alzheimer disease neuropathology and cognitive performance: a nontargeted metabolomic study. PLoS Med 14:e1002266. doi: 10.1371/journal.pmed.1002266
Soininen, H., Solomon, A., Visser, P. J., Hendrix, S. B., Blennow, K., Kivipelto, M., et al. (2017). LipiDiDiet clinical study g: 24-month intervention with a specific multinutrient in people with prodromal Alzheimer’s disease (LipiDiDiet): a randomised, double-blind, controlled trial. Lancet Neurol. 16, 965–975. doi: 10.1016/s1474-4422(17)30332-0
Son, G., and Han, J. (2018). Roles of mitochondria in neuronal development. BMB Rep. 51, 549–556. doi: 10.5483/bmbrep.2018.51.11.226
Song, C., Manku, M. S., and Horrobin, D. F. (2008). Long-chain polyunsaturated fatty acids modulate interleukin-1beta-induced changes in behavior, monoaminergic neurotransmitters, and brain inflammation in rats. J. Nutr. 138, 954–963. doi: 10.1093/jn/138.5.954
Song, H., Hecimovic, S., Goate, A., Hsu, F. F., Bao, S., Vidavsky, I., et al. (2004). Characterization of N-terminal processing of group VIA phospholipase A2 and of potential cleavage sites of amyloid precursor protein constructs by automated identification of signature peptides in LC/MS/MS analyses of proteolytic digests. J. Am. Soc. Mass Spectrom. 15, 1780–1793. doi: 10.1016/j.jasms.2004.08.012
Soriano, S., Lu, D. C., Chandra, S., Pietrzik, C. U., and Koo, E. H. (2001). The amyloidogenic pathway of amyloid precursor protein (APP) is independent of its cleavage by caspases. J. Biol. Chem. 276, 29045–29050. doi: 10.1074/jbc.m102456200
Spiteller, G. (2010). Is lipid peroxidation of polyunsaturated acids the only source of free radicals that induce aging and age-related diseases? Rejuvenation Res. 13, 91–103. doi: 10.1089/rej.2009.0934
Spuch, C., Ortolano, S., and Navarro, C. (2012). New insights in the amyloid-Beta interaction with mitochondria. J. Aging Res. 2012, 324968.
Srivastava, R. A., and Jain, J. C. (2002). Scavenger receptor class B type I expression and elemental analysis in cerebellum and parietal cortex regions of the Alzheimer’s disease brain. J. Neurol. Sci. 196, 45–52. doi: 10.1016/s0022-510x(02)00026-6
Stahl, T., Reimers, C., Johne, R., Schliebs, R., and Seeger, J. (2006). Viral-induced inflammation is accompanied by beta-amyloid plaque reduction in brains of amyloid precursor protein transgenic Tg2576 mice. Eur. J. Neurosci. 24, 1923–1934. doi: 10.1111/j.1460-9568.2006.05069.x
Stark, K. D., Van Elswyk, M. E., Higgins, M. R., Weatherford, C. A., and Salem, N. Jr. (2016). Global survey of the omega-3 fatty acids, docosahexaenoic acid andeicosapentaenoic acid in the blood stream of healthy adults. Prog. Lipid Res. 63, 132–152. doi: 10.1016/j.plipres.2016.05.001
Stassart, R. M., Mobius, W., Nave, K. A., and Edgar, J. M. (2018). The axon-myelin unit in development and degenerative disease. Front Neurosci 12:467. doi: 10.3389/fnins.2018.00467
Stephens, D. J., and Austen, B. M. (1996). Metabolites of the beta-amyloid precursor protein generated by beta-secretase localise to the trans-Golgi network and late endosome in 293 cells. J. Neurosci. Res. 46, 211–225. doi: 10.1002/(sici)1097-4547(19961015)46:2<211::aid-jnr9>3.0.co;2-k
Stone, D. J., Rozovsky, I., Morgan, T. E., Anderson, C. P., Hajian, H., and Finch, C. E. (1997). Astrocytes and microglia respond to estrogen with increased apoE mRNA in vivo and in vitro. Exp. Neurol. 143, 313–318. doi: 10.1006/exnr.1996.6360
Stonehouse, W., Conlon, C. A., Podd, J., Hill, S. R., Minihane, A. M., Haskell, C., et al. (2013). DHA supplementation improved both memory and reaction time in healthy young adults: a randomized controlled trial. Am. J. Clin. Nutr. 97, 1134–1143. doi: 10.3945/ajcn.112.053371
Strike, S. C., Carlisle, A., Gibson, E. L., and Dyall, S. C. (2016). A high Omega-3 fatty acid multinutrient supplement benefits cognition and mobility in older women: a randomized, double-blind, placebo-controlled pilot study. J. Gerontol. A Biol. Sci. Med. Sci. 71, 236–242. doi: 10.1093/gerona/glv109
Sugasini, D., Yalagala, P. C. R., Goggin, A., Tai, L. M., and Subbaiah, P. V. (2019). Enrichment of brain docosahexaenoic acid (DHA) is highly dependent upon the molecular carrier of dietary DHA: lysophosphatidylcholine is more efficient than either phosphatidylcholine or triacylglycerol. J. Nutr. Biochem. 74, 108231. doi: 10.1016/j.jnutbio.2019.108231
Sultana, R., Newman, S., Mohmmad-Abdul, H., Keller, J. N., and Butterfield, D. A. (2004). Protective effect of the xanthate, D609, on Alzheimer’s amyloid beta-peptide (1-42)-induced oxidative stress in primary neuronal cells. Free Radic. Res. 38, 449–458. doi: 10.1080/1071576042000206478
Sumner, A. (2009). Ethnic differences in triglyceride levels and high-density lipoprotein lead to underdiagnosis of the metabolic syndrome in black children and adults. J. Pediatr. 155, S7.e7–S7.e11. doi: 10.1016/j.jpeds.2009.04.049
Sun, A. Y., Draczynska-Lusiak, B., and Sun, G. Y. (2001). Oxidized lipoproteins, beta amyloid peptides and Alzheimer’s disease. Neurotox Res. 3, 167–178. doi: 10.1007/bf03033189
Sun, Y., Shi, J., Zhang, S., Tang, M., Han, H., Guo, Y., et al. (2005). The APOC3 SstI polymorphism is weakly associated with sporadic Alzheimer’s disease in a Chinese population. Neurosci. Lett. 380, 219–222. doi: 10.1016/j.neulet.2005.01.038
Svennerholm, L., Bostrom, K., Jungbjer, B., and Olsson, L. (1994). Membrane lipids of adult human brain: lipid composition of frontal and temporal lobe in subjects of age 20 to 100 years. J. Neurochem. 63, 1802–1811. doi: 10.1046/j.1471-4159.1994.63051802.x
Sweeney, M. D., Sagare, A. P., and Zlokovic, B. V. (2018). Blood-brain barrier breakdown in Alzheimer disease and other neurodegenerative disorders. Nat. Rev. Neurol. 14, 133–150. doi: 10.1038/nrneurol.2017.188
Tam, J. H., Seah, C., and Pasternak, S. H. (2014). The amyloid precursor protein is rapidly transported from the golgi apparatus to the lysosome and where it is processed into beta-amyloid. Mol. Brain 7:54.
Tan, J. Z. A., and Gleeson, P. A. (2019). The role of membrane trafficking in the processing of amyloid precursor protein and production of amyloid peptides in Alzheimer’s disease. Biochim Biophys Acta Biomembr 1861, 697–712. doi: 10.1016/j.bbamem.2018.11.013
Tchessalova, D., Posillico, C. K., and Tronson, N. C. (2018). Neuroimmune activation drives multiple brain states. Front. Syst. Neurosci. 12:39. doi: 10.3389/fnsys.2018.00039
Thinakaran, G., and Koo, E. H. (2008). Amyloid precursor protein trafficking, processing, and function. J. Biol. Chem. 283, 29615–29619. doi: 10.1074/jbc.r800019200
Thomas, M. H., Pelleieux, S., Vitale, N., and Olivier, J. L. (2016). Dietary arachidonic acid as a risk factor for age-associated neurodegenerative diseases: potential mechanisms. Biochimie 130, 168–177. doi: 10.1016/j.biochi.2016.07.013
Tindale, L. C., Leach, S., Spinelli, J. J., and Brooks-Wilson, A. R. (2017). Lipid and Alzheimer’s disease genes associated with healthy aging and longevity in healthy oldest-old. Oncotarget 8, 20612–20621. doi: 10.18632/oncotarget.15296
Toh, W. H., Tan, J. Z., Zulkefli, K. L., Houghton, F. J., and Gleeson, P. A. (2017). Amyloid precursor protein traffics from the Golgi directly to early endosomes in an Arl5b- and AP4-dependent pathway. Traffic 18, 159–175. doi: 10.1111/tra.12465
Toledo, J. B., Toledo, E., Weiner, M. W., Jack, C. R. Jr., Jagust, W., Lee, V. M., et al. (2012). Alzheimer’s disease neuroimaging I: cardiovascular risk factors, cortisol, and amyloid-beta deposition in Alzheimer’s Disease Neuroimaging Initiative. Alzheimers Dement 8, 483–489. doi: 10.1016/j.jalz.2011.08.008
Tomita, T., and Wong, P. C. (2011). Selectivity to amyloid-beta precursor protein cleavage provides hope against Alzheimer’s. Alzheimers Res. Ther. 3:7. doi: 10.1186/alzrt66
Torres, M., Price, S. L., Fiol-Deroque, M. A., Marcilla-Etxenike, A., Ahyayauch, H., Barcelo-Coblijn, G., et al. (2014). Membrane lipid modifications and therapeutic effects mediated by hydroxydocosahexaenoic acid on Alzheimer’s disease. Biochim. Biophys. Acta 1838, 1680–1692. doi: 10.1016/j.bbamem.2013.12.016
Tremblay, M. E., Stevens, B., Sierra, A., Wake, H., Bessis, A., and Nimmerjahn, A. (2011). The role of microglia in the healthy brain. J. Neurosci. 31, 16064–16069. doi: 10.1523/jneurosci.4158-11.2011
Tremblay, M. E., Zhang, I., Bisht, K., Savage, J. C., Lecours, C., Parent, M., et al. (2016). Remodeling of lipid bodies by docosahexaenoic acid in activated microglial cells. J. Neuroinflammation 13:116.
Tulloch, J., Leong, L., Thomson, Z., Chen, S., Lee, E. G., Keene, C. D., et al. (2018). Glia-specific APOE epigenetic changes in the Alzheimer’s disease brain. Brain Res. 1698, 179–186. doi: 10.1016/j.brainres.2018.08.006
Tyurina, Y. Y., Poloyac, S. M., Tyurin, V. A., Kapralov, A. A., Jiang, J., Anthonymuthu, T. S., et al. (2014). A mitochondrial pathway for biosynthesis of lipid mediators. Nat. Chem. 6, 542–552.
Tzioras, M., Davies, C., Newman, A., Jackson, R., and Spires-Jones, T. (2019). Invited Review: APOE at the interface of inflammation, neurodegeneration and pathological protein spread in Alzheimer’s disease. Neuropathol. Appl. Neurobiol. 45, 327–346. doi: 10.1111/nan.12529
Uddin, M. S., Rahman, M. M., Jakaria, M., Rahman, M. S., Hossain, M. S., Islam, A., et al. (2020). Estrogen signaling in Alzheimer’s Disease: molecular insights and therapeutic targets for Alzheimer’s dementia. Mol. Neurobiol. doi: 10.1007/s12035-020-01911-8 [Epub ahead of print].
Umamaheswaran, S., Dasari, S. K., Yang, P., Lutgendorf, S. K., and Sood, A. K. (2018). Stress, inflammation, and eicosanoids: an emerging perspective. Cancer Metastasis Rev. 37, 203–211. doi: 10.1007/s10555-018-9741-1
van de Rest, O., Geleijnse, J. M., Kok, F. J., van Staveren, W. A., Dullemeijer, C., Olderikkert, M. G., et al. (2008). Effect of fish oil on cognitive performance in older subjects: a randomized, controlled trial. Neurology 71, 430–438. doi: 10.1212/01.wnl.0000324268.45138.86
van den Kommer, T. N., Dik, M. G., Comijs, H. C., Lutjohann, D., Lips, P., Jonker, C., et al. (2012). The role of extracerebral cholesterol homeostasis and ApoE e4 in cognitive decline. Neurobiol. Aging 33, e617–e628.
Vance, J. E., and Hayashi, H. (2010). Formation and function of apolipoprotein E-containing lipoproteins in the nervous system. Biochim. Biophys. Acta 1801, 806–818. doi: 10.1016/j.bbalip.2010.02.007
Vannucci, S. J., Clark, R. R., Koehler-Stec, E., Li, K., Smith, C. B., Davies, P., et al. (1998). Glucose transporter expression in brain: relationship to cerebral glucose utilization. Dev. Neurosci. 20, 369–379. doi: 10.1159/000017333
Veerhuis, R., Nielsen, H. M., and Tenner, A. J. (2011). Complement in the brain. Mol. Immunol. 48, 1592–1603.
Vermunt, L., Sikkes, S. A. M., van den Hout, A., Handels, R., Bos, I., van der Flier, W. M., et al. (2019). Duration of preclinical, prodromal, and dementia stages of Alzheimer’s disease in relation to age, sex, and APOE genotype. Alzheimers Dement. 15, 888–898. doi: 10.1016/j.jalz.2019.04.001
Villasenor, R., Schilling, M., Sundaresan, J., Lutz, Y., and Collin, L. (2017). Sorting tubules regulate blood-brain barrier transcytosis. Cell Rep. 21, 3256–3270. doi: 10.1016/j.celrep.2017.11.055
Viña, J., and Lloret, A. (2010). Why women have more Alzheimer’s disease than men: gender and mitochondrial toxicity of amyloid-β peptide. J. Alzheimers Dis. 20, S527–S533.
Vina, J., Lloret, A., Orti, R., and Alonso, D. (2004). Molecular bases of the treatment of Alzheimer’s disease with antioxidants: prevention of oxidative stress. Mol. Aspects. Med. 25, 117–123. doi: 10.1016/j.mam.2004.02.013
Volmar, C. H., Salah-Uddin, H., Janczura, K. J., Halley, P., Lambert, G., Wodrich, A., et al. (2017). M344 promotes nonamyloidogenic amyloid precursor protein processing while normalizing Alzheimer’s disease genes and improving memory. Proc. Natl. Acad. Sci. U.S.A. 114, E9135–E9144.
Wang, H. (2014). Lipid rafts: a signaling platform linking cholesterol metabolism to synaptic deficits in autism spectrum disorders. Front. Behav. Neurosci. 8:104. doi: 10.3389/fnbeh.2014.00104
Wang, J. F., Lu, R., and Wang, Y. Z. (2010). Regulation of beta cleavage of amyloid precursor protein. Neurosci. Bull. 26, 417–427.
Wang, S., Zhang, X., Zhai, L., Sheng, X., Zheng, W., Chu, H., et al. (2018). Atorvastatin attenuates cognitive deficits and neuroinflammation induced by Abeta1-42 involving modulation of TLR4/TRAF6/NF-kappaB pathway. J. Mol. Neurosci. 64, 363–373. doi: 10.1007/s12031-018-1032-3
Wang, W. Y., Tan, M. S., Yu, J. T., and Tan, L. (2015a). Role of pro-inflammatory cytokines released from microglia in Alzheimer’s disease. Ann. Transl. Med. 3:136. doi: 10.3978/j.issn.2305-5839.2015.03.49
Wang, X., Zhu, M., Hjorth, E., Cortes-Toro, V., Eyjolfsdottir, H., Graff, C., et al. (2015b). Resolution of inflammation is altered in Alzheimer’s disease. Alzheimers Dement 11, e41–e42.
Wang, X. F., Cao, Y. W., Feng, Z. Z., Fu, D., Ma, Y. S., Zhang, F., et al. (2013). Quantitative assessment of the effect of ABCA1 gene polymorphism on the risk of Alzheimer’s disease. Mol. Biol. Rep. 40, 779–785. doi: 10.1007/s11033-012-2115-9
Wavrant-De Vrieze, F., Compton, D., Womick, M., Arepalli, S., Adighibe, O., Li, L., et al. (2007). ABCA1 polymorphisms and Alzheimer’s disease. Neurosci. Lett. 416, 180–183.
Weiser, M. J., Butt, C. M., and Mohajeri, M. H. (2016). Docosahexaenoic acid and cognition throughout the Lifespan. Nutrients 8:99. doi: 10.3390/nu8020099
Welberg, L. (2014). Synaptic transmission: membrane lipids channel a message. Nat. Rev. Neurosci. 15:135. doi: 10.1038/nrn3701
Wender, M., Adamczewska-Goncerzewicz, Z., Szczech, J., and Godlewski, A. (1988). Myelin lipids in aging human brain. Neurochem. Pathol. 8, 121–130.
Wezyk, M., Szybinska, A., Wojsiat, J., Szczerba, M., Day, K., Ronnholm, H., et al. (2018). Overactive BRCA1 affects presenilin 1 in induced pluripotent stem cell-derived neurons in Alzheimer’s disease. J. Alzheimers Dis. 62, 175–202. doi: 10.3233/jad-170830
Whelan, J. (2008). (n-6) and (n-3) Polyunsaturated fatty acids and the aging brain: food for thought. J. Nutr. 138, 2521–2522. doi: 10.3945/jn.108.095943
Whiley, L., Sen, A., Heaton, J., Proitsi, P., Garcia-Gomez, D., Leung, R., et al. (2014). Evidence of altered phosphatidylcholine metabolism in Alzheimer’s disease. Neurobiol. Aging 35, 271–278.
Whittington, R. A., Planel, E., and Terrando, N. (2017). Impaired resolution of inflammation in alzheimer’s disease: a review. Front. Immunol. 8:1464. doi: 10.3389/fimmu.2017.01464
Willis, L. M., Shukitt-Hale, B., and Joseph, J. A. (2009). Dietary polyunsaturated fatty acids improve cholinergic transmission in the aged brain. Genes Nutr. 4, 309–314. doi: 10.1007/s12263-009-0141-6
Wirth, M., Lange, C., and Huijbers, W. (2019). Alzheimer’s disease neuroimaging I: plasma cortisol is associated with cerebral hypometabolism across the Alzheimer’s disease spectrum. Neurobiol. Aging 84, 80–89. doi: 10.1016/j.neurobiolaging.2019.08.003
Wojsiat, J., Zoltowska, K. M., Laskowska-Kaszub, K., and Wojda, U. (2018). Oxidant/antioxidant imbalance in Alzheimer’s disease: therapeutic and diagnostic prospects. Oxid. Med. Cell Longev. 2018:6435861.
Wollmer, M. A., Sleegers, K., Ingelsson, M., Zekanowski, C., Brouwers, N., Maruszak, A., et al. (2007). Association study of cholesterol-related genes in Alzheimer’s disease. Neurogenetics 8, 179–188.
Wollmer, M. A., Streffer, J. R., Lutjohann, D., Tsolaki, M., Iakovidou, V., Hegi, T., et al. (2003a). ABCA1 modulates CSF cholesterol levels and influences the age at onset of Alzheimer’s disease. Neurobiol. Aging 24, 421–426. doi: 10.1016/s0197-4580(02)00094-5
Wollmer, M. A., Streffer, J. R., Tsolaki, M., Grimaldi, L. M., Lutjohann, D., Thal, D., et al. (2003b). Genetic association of acyl-coenzyme A: cholesterol acyltransferase with cerebrospinal fluid cholesterol levels, brain amyloid load, and risk for Alzheimer’s disease. Mol. Psychiatry 8, 635–638. doi: 10.1038/sj.mp.4001296
Wolozin, B., Brown, J. III, Theisler, C., and Silberman, S. (2004). The cellular biochemistry of cholesterol and statins: insights into the pathophysiology and therapy of Alzheimer’s disease. CNS Drug. Rev. 10, 127–146. doi: 10.1111/j.1527-3458.2004.tb00009.x
Wong, M. W. K., Braidy, N., Crawford, J., Pickford, R., Song, F., Mather, K. A., et al. (2019). APOE genotype differentially modulates plasma lipids in healthy older individuals, with relevance to brain health. J. Alzheimers Dis. 72, 703–716. doi: 10.3233/jad-190524
Wong, M. W., Braidy, N., Poljak, A., Pickford, R., Thambisetty, M., and Sachdev, P. S. (2017). Dysregulation of lipids in Alzheimer’s disease and their role as potential biomarkers. Alzheimers Dement 13, 810–827. doi: 10.1016/j.jalz.2017.01.008
Wood, P. L. (2012). Lipidomics of Alzheimer’s disease: current status. Alzheimers Res. Ther. 4:5. doi: 10.1186/alzrt103
Wu, L. J., and Zhuo, M. (2008). Resting microglial motility is independent of synaptic plasticity in mammalian brain. J. Neurophysiol. 99, 2026–2032. doi: 10.1152/jn.01210.2007
Xiao, Z., Wang, J., Chen, W., Wang, P., Zeng, H., and Chen, W. (2012). Association studies of several cholesterol-related genes (ABCA1, CETP and LIPC) with serum lipids and risk of Alzheimer’s disease. Lipids Health Dis. 11:163. doi: 10.1186/1476-511x-11-163
Yang, M., Virassamy, B., Vijayaraj, S. L., Lim, Y., Saadipour, K., Wang, Y. J., et al. (2013). The intracellular domain of sortilin interacts with amyloid precursor protein and regulates its lysosomal and lipid raft trafficking. PLoS One 8:e63049. doi: 10.1371/journal.pone.0063049
Yang, R., Wang, H., Wen, J., Ma, K., Chen, D., Chen, Z., et al. (2019). Regulation of microglial process elongation, a featured characteristic of microglial plasticity. Pharmacol. Res. 139, 286–297. doi: 10.1016/j.phrs.2018.11.028
Yang, X., Sheng, W., Sun, G. Y., and Lee, J. C. (2011). Effects of fatty acid unsaturation numbers on membrane fluidity and alpha-secretase-dependent amyloid precursor protein processing. Neurochem. Int. 58, 321–329. doi: 10.1016/j.neuint.2010.12.004
Yassine, H. N., Braskie, M. N., Mack, W. J., Castor, K. J., Fonteh, A. N., Schneider, L. S., et al. (2017). Association of docosahexaenoic acid supplementation with alzheimer disease stage in apolipoprotein E epsilon4 carriers: a review. JAMA Neurol. 74, 339–347.
Yates, D. (2014). Synaptic plasticity: Microglial cell-mediated depression. Nat. Rev. Neurosci. 15:280.
Yatin, S. M., Varadarajan, S., and Butterfield, D. A. (2000). Vitamin E prevents Alzheimer’s Amyloid beta-peptide (1-42)-induced neuronal protein oxidation and reactive oxygen species production. J. Alzheimers Dis. 2, 123–131. doi: 10.3233/jad-2000-2212
Yehuda, S., Rabinovitz, S., Carasso, R. L., and Mostofsky, D. I. (2002). The role of polyunsaturated fatty acids in restoring the aging neuronal membrane. Neurobiol. Aging 23, 843–853. doi: 10.1016/s0197-4580(02)00074-x
Yin, F., Sancheti, H., Patil, I., and Cadenas, E. (2016). Energy metabolism and inflammation in brain aging and Alzheimer’s disease. Free Radic. Biol. Med. 100, 108–122. doi: 10.1016/j.freeradbiomed.2016.04.200
Yoon, I. S., Chen, E., Busse, T., Repetto, E., Lakshmana, M. K., Koo, E. H., et al. (2007). Low-density lipoprotein receptor-related protein promotes amyloid precursor protein trafficking to lipid rafts in the endocytic pathway. FASEB J. 21, 2742–2752. doi: 10.1096/fj.07-8114com
Yu, Q., Fang, D., Swerdlow, R. H., Yu, H., Chen, J. X., and Yan, S. S. (2016). Antioxidants rescue mitochondrial transport in differentiated Alzheimer’s disease trans-mitochondrial cybrid cells. J. Alzheimers Dis. 54, 679–690. doi: 10.3233/jad-160532
Yuksel, M., and Tacal, O. (2019). Trafficking and proteolytic processing of amyloid precursor protein and secretases in Alzheimer’s disease development: an up-to-date review. Eur. J. Pharmacol. 856:172415. doi: 10.1016/j.ejphar.2019.172415
Zandl-Lang, M., Fanaee-Danesh, E., Sun, Y., Albrecher, N. M., Gali, C. C., Cancar, I., et al. (2018). Regulatory effects of simvastatin and apoJ on APP processing and amyloid-beta clearance in blood-brain barrier endothelial cells. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 1863, 40–60. doi: 10.1016/j.bbalip.2017.09.008
Zatorre, R. J., Fields, R. D., and Johansen-Berg, H. (2012). Plasticity in gray and white: neuroimaging changes in brain structure during learning. Nat. Neurosci. 15, 528–536. doi: 10.1038/nn.3045
Zhang, C., Wang, K., Yang, L., Liu, R., Chu, Y., Qin, X., et al. (2018). Lipid metabolism in inflammation-related diseases. Analyst 143, 4526–4536. doi: 10.1039/c8an01046c
Zhang, Y. W., Thompson, R., Zhang, H., and Xu, H. (2011). APP processing in Alzheimer’s disease. Mol. Brain 4:3. doi: 10.1186/1756-6606-4-3
Zhao, B. (2009). Natural antioxidants protect neurons in Alzheimer’s disease and Parkinson’s disease. Neurochem. Res. 34, 630–638. doi: 10.1007/s11064-008-9900-9
Zhao, N., Liu, C. C., Qiao, W., and Bu, G. (2018). Apolipoprotein E, receptors, and modulation of Alzheimer’s disease. Biol. Psychiatry 83, 347–357.
Zhao, Q. F., Yu, J. T., Tan, M. S., and Tan, L. (2015). ABCA7 in Alzheimer’s disease. Mol. Neurobiol. 51, 1008–1016.
Zhao, R. Z., Jiang, S., Zhang, L., and Yu, Z. B. (2019). Mitochondrial electron transport chain, ROS generation and uncoupling (review). Int. J. Mol. Med. 44, 3–15.
Zhao, Z., and Zlokovic, B. V. (2014). Blood-brain barrier: a dual life of MFSD2A? Neuron 82, 728–730. doi: 10.1016/j.neuron.2014.05.012
Zhao, Z., Nelson, A. R., Betsholtz, C., and Zlokovic, B. V. (2015). Establishment and dysfunction of the blood-brain barrier. Cell 163, 1064–1078. doi: 10.1016/j.cell.2015.10.067
Zheng, C., Zhou, X. W., and Wang, J. Z. (2016). The dual roles of cytokines in Alzheimer’s disease: update on interleukins, TNF-alpha, TGF-beta and IFN-gamma. Transl. Neurodegener. 5:7.
Zhou, Q., Peng, D., Yuan, X., Lv, Z., Pang, S., Jiang, W., et al. (2014). APOE and APOC1 gene polymorphisms are associated with cognitive impairment progression in Chinese patients with late-onset Alzheimer’s disease. Neural. Regen. Res. 9, 653–660.
Zhu, L., Liu, X., Nemeth, D. P., DiSabato, D. J., Witcher, K. G., McKim, D. B., et al. (2019). Interleukin-1 causes CNS inflammatory cytokine expression via endothelia-microglia bi-cellular signaling. Brain Behav. Immun. 81, 292–304. doi: 10.1016/j.bbi.2019.06.026
Zhu, R., Liu, X., and He, Z. (2018). Association between CLU gene rs11136000 polymorphism and Alzheimer’s disease: an updated meta-analysis. Neurol. Sci. 39, 679–689. doi: 10.1007/s10072-018-3259-8
Keywords: amyloid precursor protein, apolipoproteins, blood-brain barrier, energy metabolism, inflammation, late-onset Alzheimer’s disease, mitochondria, myelination
Citation: Chew H, Solomon VA and Fonteh AN (2020) Involvement of Lipids in Alzheimer’s Disease Pathology and Potential Therapies. Front. Physiol. 11:598. doi: 10.3389/fphys.2020.00598
Received: 28 February 2020; Accepted: 14 May 2020;
Published: 09 June 2020.
Edited by:
Jue Wang, The University of Texas Health Science Center at Tyler, United StatesReviewed by:
Mario Diaz, University of La Laguna, SpainMarcus O. Grimm, Saarland University, Germany
Ian James Martins, The University of Western Australia, Australia
Copyright © 2020 Chew, Solomon and Fonteh. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alfred N. Fonteh, YWxmcmVkLmZvbnRlaEBobXJpLm9yZw==