 Zhenhong Fu1*†
Zhenhong Fu1*† Yang Jiao1†
Yang Jiao1† Jihang Wang1Ying Zhang1Mingzhi Shen1
Jihang Wang1Ying Zhang1Mingzhi Shen1 Russel J. Reiter2,3Qing Xi4*Yundai Chen1*
Russel J. Reiter2,3Qing Xi4*Yundai Chen1*- 1Department of Cardiology, The First Medical Center, Chinese PLA General Hospital, Beijing, China
- 2Department of Cellular and Structural Biology, UT Health San Antonio, San Antonio, TX, United States
- 3San Antonio Cellular Therapeutics Institute, Department of Biology, College of Sciences, University of Texas at San Antonio, San Antonio, TX, United States
- 4The First Medical Center, Chinese PLA General Hospital, Beijing, China
Melatonin is a pleiotropic, indole secreted, and synthesized by the human pineal gland. Melatonin has biological effects including anti-apoptosis, protecting mitochondria, anti-oxidation, anti-inflammation, and stimulating target cells to secrete cytokines. Its protective effect on cardiomyocytes in acute myocardial infarction (AMI) has caused widespread interest in the actions of this molecule. The effects of melatonin against oxidative stress, promoting autophagic repair of cells, regulating immune and inflammatory responses, enhancing mitochondrial function, and relieving endoplasmic reticulum stress, play crucial roles in protecting cardiomyocytes from infarction. Mitochondrial apoptosis and dysfunction are common occurrence in cardiomyocyte injury after myocardial infarction. This review focuses on the targets of melatonin in protecting cardiomyocytes in AMI, the main molecular signaling pathways that melatonin influences in its endogenous protective role in myocardial infarction, and the developmental prospect of melatonin in myocardial infarction treatment.
Introduction
With the general improvement of the human living standard, the change of living habits and the prolongation of life span, the prevalence rate of cardiovascular diseases has risen sharply. According to China Cardiovascular Disease Report in 2018, the population with cardiovascular diseases in China has reached up to 290 million, and the number of patients with acute myocardial infarction (AMI) is about 2.5 million annually. AMI has become a disease seriously affecting people’s life span and quality. Despite great progress of modern medicine, science and technology, iterative new anticoagulants and antiplatelet drug, improvements in the management of AMI patients (Amanakis et al., 2019; Heusch, 2019), including the more frequent coronary reperfusion using fibrinolysis, primary percutaneous coronary intervention (PCI) or coronary artery bypass grafting (CABG), the prognosis of AMI has improved, yet, there are frequent occurrences of malignant arrhythmia events, a decline of cardiac function following AMI leading to the development of heart failure and a poor prognosis (Botker et al., 2018; Davidson et al., 2018; Joshi et al., 2019). Due to ischemia and hypoxia of the infarct area, a large number of inflammatory cells infiltrate the lesion, cardiomyocyte apoptosis is frequent and scar repair is common (Li et al., 2010; Galluzzi et al., 2018). Due to the non-regenerative feature of cardiomyocytes, irreversible cardiomyocyte apoptosis and infarction caused by acute ischemia are main factors for poor prognosis of AMI (Abukar et al., 2018; Beckendorf et al., 2018). After myocardial infarction, the myocardial tissue in the infarcted area and its adjacent non-infarcted areas undergoes apoptosis and necrosis due to ischemia, hypoxia and inflammatory response (Zhou et al., 2018c; Hu et al., 2019). Thus, the hemodynamic parameters and electrical nerve conduction of the myocardium are changed, and bring about cardiac systolic, diastolic dysfunction and dysrhythmia, and result in ventricular remodeling and electrocardiogram (ECG) changes (Zhang et al., 2014; Joshi et al., 2019). Mitochondria are organelles that generate ATP (Kowaltowski, 2019; Lee B.W.L. et al., 2019). They play a significant role in myocardial injury after myocardial infarction (Zhou et al., 2018h; Zhou et al., 2020). Stabilizing the structure and function of mitochondria effectively inhibits cardiomyocyte injury and necrosis (Audia et al., 2018; Lu J. et al., 2019). Melatonin, an indoleamine secreted by pineal gland, is a highly effective antioxidant and beneficial to many diseases including diabetes, infectious diseases, metabolic syndrome, depression, and neurodegenerative diseases (Zaouali et al., 2011; de Oliveira Junior et al., 2019). The value of melatonin in the treatment of myocardial infarction has drawn wide-spread attention in recent years. Highly compatible with mitochondrial membrane receptors, melatonin effectively reduces mitochondrial dysfunction, thus inhibiting post-myocardial infarction damage of cardiomyocytes. This review summarizes the effect of melatonin in protecting cardiomyocytes and improving prognosis after myocardial infarction by enhancing the adaptability of cardiomyocytes to ischemia and hypoxia via stabilizing mitochondrial function. We hope this introduction will be helpful to the research of melatonin in the treatment of myocardial infarction.
Pathophysiological Changes of Heart During Myocardial Infarction
Lipid Deposition and Atherosclerotic Plaque Formation
The direct cause of AMI is myocardial necrosis caused by prolonged ischemia and hypoxia resulting from coronary artery occlusion or spasm. The key factor leading to acute myocardial ischemia is the rupture of an atherosclerotic plaque in the coronary artery and the gradual formation of a thrombus (Zhou et al., 2018e; Bacmeister et al., 2019). The pathogenesis of atherosclerosis includes lipid deposition, inflammation, thrombosis, endothelial dysfunction, smooth muscle cell cloning and other processes (Nofe et al., 2010; Zhou et al., 2018b; Zhao J. et al., 2019). The most important risk factors are lipid metabolism disorder and endothelial cell injury. Under long-term lipid metabolism abnormality, low density lipoprotein cholesterol (LDL-C) enters intima through damaged arterial endothelium where it is converted to oxidized LDL cholesterol (Ox LDL-C), causing further damage to the intima of the coronary artery (Liu et al., 2019; Aimo et al., 2020). Ox LDL-C is engulfed by macrophages, gradually forming foam cells; there after, atherosclerotic plaques gradually develop along with the aggregation of foam cells and lipids (Abdelnaseer et al., 2016; Wu et al., 2019). If the intima of a coronary artery ruptures, the atheromatous plaque substance enters the lumen and becomes an embolus which completely occludes the blood vessel (Yuqi et al., 2015; Tian et al., 2018; Xiong et al., 2019). Thus, regulating the lipid metabolism balance, delaying the formation of atherosclerotic plaques and inhibiting plaque rupture are key measures in preventing myocardial infarction.
Platelet Activation Aggregation and Thrombosis
When atherosclerotic plaques are formed and protrude into the lumen, local arterial stenosis causes changes in blood flow turbulence and shear force, leading to an interruption of arterial intima continuity, contraction of endothelial cells, exposure of tissues such as subendothelial collagen, and activation of platelets; this causes further adhesion and aggregation of platelets on the arterial intima, and finally formation of atherosclerotic thrombotic disease (Cohen et al., 2019; Trindade et al., 2019). Reports show that the mural thrombosis plays a vital role in pathophysiological changes of myocardial infarction progression (Hu S.Y. et al., 2017; Zhou et al., 2017b). At present, there are three types of antiplatelet drugs used in the clinic: (1) cyclooxygenase-1 (COX-1) inhibitor: aspirin. (2) ADP receptor antagonists: clopidogrel, prasugrel, ticagrelor. (3) platelet glycoprotein IIb/IIIa (GP IIb/IIIa) receptor antagonists: tirofiban, etibadin, abciximab. Among these, aspirin is a first-line antiplatelet drug and can irreversibly inactivate COX-1, an enzyme expressed in platelets, which moderates the synthesis of thromboxane; thromboxane is a potent platelet aggregation agent (Davidson et al., 2018; Merz et al., 2018).
Inflammatory Cell Infiltration and Inflammatory Response
The microenvironment of the infarcted cardiomyocyte is a dynamic and complex area. Inflammation and immune responses play important roles in the initiation and progression of myocardial infarction. After myocardial infarction, inflammatory cells such as neutrophils, monocytes, and macrophages are activated, causing their number to increase sharply (Basalay et al., 2018; Wang Z. et al., 2019; Faghfouri et al., 2020). A large amount of inflammatory cytokines are released and move toward the injured area, and the permeability of endothelial cells increases, which results in the infiltration of inflammatory cells (Chen et al., 2016; Thieltges et al., 2018). Cytokine release and the inflammatory response are important conditions for tissue healing after myocardial infarction, but an excessive inflammatory response leads to myocardial tissue remodeling, activating apoptosis signals in cardiomyocytes with destruction of the integrity of extracellular matrix, which is not conducive to the survival of cardiomyocytes and the recovery of cardiac function (Nahrendorf et al., 2010; Krause et al., 2018). Apoptosis and the inflammatory response are initiated by myocardial tissue injury in the ischemic area. Specific cytokines and mediators including interleukin-1 (IL-1), IL-6, nuclear transcription factor kB (NF-κB) p65, NF-κB p50, Toll-like receptor 4(TLR4), Tenascin-C (TNC) and other inflammatory response regulatory proteins, as well as apoptosis regulatory proteins caspase-3, Bcl-2, etc. are produced. Excessive inflammatory response and fiber proliferation lead to ventricular remodeling (Nokik et al., 2017; Crist et al., 2018; Bocci et al., 2019; Chen M. et al., 2019). TLRs are a major reaction pathway of the inflammatory response after myocardial infarction. Recognition receptors are expressed by inflammatory cells and recognize the danger signal released by injured cells (Jager and Hoefer, 2019; Sircana et al., 2019; Yang M.Y. et al., 2019). Studies have shown that adjusting the activity of TLRs enhances the positive effect of the inflammatory response on myocardial tissue healing and limits damage to tissue, thus providing a new therapeutic process for promoting myocardial tissue recovery after myocardial infarction (Dominguez-Rodriguez et al., 2010b; Cao et al., 2019). Protein kinase D1 (PKD1) mediates the processes of myocardial remodeling, angiogenesis and myocardial contraction (Ren, 2016; Serocki et al., 2018), but the mechanism of PKD1 in the processes of the inflammatory response and cell injury in the microenvironment after myocardial infarction has not been determined.
Cardiomyocyte Death
Cardiomyocyte injury following AMI is a very complex with multi-linked pathophysiological processes, including not only an inflammatory response, immune response, and cell signal transduction, but also complex processes such as apoptosis, necrosis, autophagy, and mitochondrial dysfunction (Westman et al., 2016; Hockings et al., 2018; Heckmann et al., 2019). Immunological studies show that cardiomyocyte in the myocardial infarction region is seriously damaged, apoptosis is severe, the expressions of Bax and caspase-3 are significantly increased, while the expression of Bcl-2 is reduced (Jin et al., 2018). It is reported that, caspase-8 is involved in fatty acid synthase (Fas)/FasL-related death receptor pathway, caspase-9 is depended on mitochondrial damage, and caspase-12 is related to endoplasmic reticulum stress (Fuhrmann and Brune, 2017; Karwi et al., 2018; Jiang et al., 2019). These are associated with cardiomyocyte apoptosis following myocardial infarction. After AMI, the focal area mainly consists of a peripheral ischemic penumbra and inner infarct area. The ischemic penumbra is mainly composed of apoptotic cardiomyocytes, and the inner infarcted area contains numerous dead cardiomyocytes (Niccoli et al., 2016; Hadebe et al., 2018). Studies reveal that the number of necrotic cells is seven times that of apoptotic cells; thus apoptosis plays only a minor role in the progression of myocardial infarction (Adameova et al., 2016; Zhou et al., 2018a; Zhu P. et al., 2018). Mitochondrial reactive oxygen species (ROS) and xanthine oxidase activity in ischemic areas lead to oxidative stress, and the calcium-induced calcium release (CICR) leads to intracellular calcium overload (Zollbrecht et al., 2016; Espinosa-Diez et al., 2018; Graczyk-Jarzynka et al., 2019; Guidarelli et al., 2019). Oxidative stress and inflammatory response activate receptor interaction protein kinase 3 (Ripk3) via interaction with cell death receptors; the common receptors are the Fas receptor, TRAIL receptor and TNFR1 (Zhou et al., 2017e; Zhou and Toan, 2020). Ripk3 destroys cell membrane integrity by mediating cell membrane-related chromosome translocation, resulting in cell necrosis (Kohlstedt et al., 2018; Zhong et al., 2019).
Mitochondrial Dysfunction and Mitophagy
As organelles with important functions in eukaryotic cells, mitochondria mainly generate ATP, moderating apoptosis and oxidative stress reactions and exhibit dynamic changes. Their development and degradation, fusion and fission, and their quantity and morphology are precisely regulated and controlled based on changes in the environment where the cells are located (Zhou et al., 2017a,f; Chen S. et al., 2019; Mayorov et al., 2019; Simula et al., 2019). Mitochondrial dynamics include mitochondrial fission, fusion and mitophagy. Mitochondrial fission aggravates the damage to mitochondrial structure and function by inducing mitochondrial fragmentation. The fission process is considered as an initiation event for mitochondrial apoptosis (Zhou et al., 2017c; Xu Z. et al., 2019; Zhou et al., 2019). Mitochondrial fusion reduces mitochondrial damage and maintains function of these organelles by promoting the integration of mitochondrial fragments, enhancing the stability of mitochondrial genes, and promoting the exchange of mitochondrial contents (Filadi et al., 2018; Kanaan et al., 2018; Yao et al., 2019). As a process of organelle autophagy, mitophagy removes and digests damaged mitochondria fragment via mitochondrial degradation moderated by lysosomes (Zhou et al., 2018d, e; Breda et al., 2019; Mei et al., 2019). Mitophagy is specific and a main pathway of mitochondrial metabolism, and plays a key role in maintaining cell function stability (Liu et al., 2012; Bhandari et al., 2014; Kang et al., 2016).
When cellular damage causes mitochondrial structural dysfunction, cells firstly maintain their original structure through antioxidant factors, DNA repair, protein folding, and so on. If this first line of defense is breached, autophagy, fusion, and biogenesis of the mitochondria are activated, which is a more effective and extensive quality control system (Yang M. et al., 2019). The process of mitophagy is closely related to ubiquitin/proteasome system (UPS). First, the damaged mitochondria are ubiquitinated through modification and then recognized by ubiquitin receptors. The substrates are induced to mitophagy degradation by binding to LC3 on phages, and finally the components after degradation are released to the cytoplasm for recycling, providing nutrients and energy for cell survival (Ji and Kwon, 2017). Recent studies reveal that mitophagy participates in mitochondrial protection induced by mitochondrial fusion (Kornicka et al., 2019). Mitochondrial fusion initially promotes the integration of fragmented mitochondria, and then “cleans” mitochondria by proteolysis in lysosomes to moderately degrade damaged mitochondrial fragments, thus maintaining the stability of mitochondrial quality and quantity (Tahrir et al., 2019; Wang et al., 2020). In most cases, mitophagy can remove defective mitochondria from AMI injuries, playing a protective and adaptive role. The experimental results of Siddall et al. (2013) show that the enhancement of mitophagy activity protects cardiomyocytes by reducing the production of mitochondrial ROS and inhibiting calcium overload (Reddy et al., 2018). The role of mitophagy in myocardial infarction cell injury, however, is still controversial. However, experimental data of Yang et al. (2018) indicates that the rise of mitophagy reduces the energy supply of cells, thus aggravating cell injury (Koentges et al., 2018; Zhou et al., 2018g).
The heart has a very high demand for energy. There are a large number of mitochondria in cardiomyocytes. Their total volume of mitochondria in a cardiomyocyte accounts for approximately 22–37%. The energy is used to maintain the normal blood pumping function of the heart. The distribution and supply of energy are related closely to the functional state of mitochondria (Battogtokh et al., 2018; Joshi and Mochly-Rosen, 2018; Zhong et al., 2019). The mitochondrial membrane is a bilayer structure, with a non-specific ion channel in the inner membrane. This channel is known as the mitochondrial permeability transition pore (MPTP), which plays a critical role in maintaining Ca2+ dynamic balance and apoptosis (Meyer and Leuschner, 2018; Venugopal et al., 2018). The opening of MPTP leads to an increase of mitochondrial intimal permeability, resulting in imbalance of electrochemical driving force of ions inside and outside the membrane, a decrease of Na+-K+-ATP enzyme activity, and transmembrane transport barrier of sodium and potassium ions, which further leads to mitochondrial dysfunction, a drop of the mitochondrial inner membrane potential with the release of cytochrome c (Cyt c), and activation of the apoptosis program (Xiao et al., 2018; Xu T. et al., 2019). Under normal circumstances, MPTP is closed state and is activated when the concentration of oxygen-derived free radicals and Ca2+ is elevated (Yang et al., 2016; Xu S.F. et al., 2019). During AMI, cardiomyocytes are in a microenvironment of ischemia and hypoxia, with an extensive infiltration of inflammatory cells and oxygen-derived free radicals. Mitochondria then release apoptosis-inducing factors, Cyt c, pro-interleukin 1, and other mediators that induce apoptosis in this harsh microenvironment; this mediates the release of IL-1 through an inflammatory cascade reaction resulting in cardiomyocyte injury and necrosis (Teixeira et al., 2018; Lee E. et al., 2019). In addition, a large number of free radicals and inflammatory agents destroy the structure and function of mitochondria, causing inactivation of various enzymes, destruction of double membrane barrier and opening of the MPTP, causing mitochondrial edema, disintegration, and dysfunction of energy generation, and eventually leading to cardiomyocyte death and cardiac function damage. During the process of AMI, opening of MPTP plays an important role (Boengler et al., 2018; Schreiber et al., 2019). The cardiomyocyte injury in ischemia is reduced by inhibiting opening of the MPTP, lowering the release of Cyt c and limiting oxygen-derived free radicals and Ca2+ overload (Seidlmayer et al., 2015; Morell et al., 2018).
Target the Melatonin Protects Cardiomyocytes in Acute Myocardial Infarction
Melatonin Plays Anti-oxidative Stress Effect and Inhibits Inflammatory Response
Melatonin was originally discovered in the bovine pineal gland and was named after its ability to change pigmentation (melanin) in amphibian skin. Controlled by sympathetic nervous system, the synthesis and secretion of melatonin are in phase with the fluctuations in the light:dark cycle with little secretion during the day and high amounts of secretion at night (Perez-Gonzalez et al., 2019). Some of the biological effects of melatonin are related to its ability to effectively scavenge free radicals and enhance the activity of antioxidant enzymes; melatonin’s metabolites also exhibit high radical scavenging activity (Galano et al., 2013). Melatonin is highly potent free radical scavenger due to multiple mechanisms (Galano and Reiter, 2018). Free radicals that are eliminated include nitric oxide (NO⋅), superoxide anion radical (O2–⋅) and hydroxyl radical (OH⋅), etc. Its hydrophilicity and high lipophilicity, allows melatonin to pass through the cell membrane and the nuclear membrane easily, thus exerting strong antioxidant effects in cytoplasm and nucleus (Garcia et al., 2015). In addition to directly scavenging free radicals, melatonin also induces the expression of antioxidant enzymes to achieve indirect antioxidant effects (Kleszczynski et al., 2016). Nrf2 (NF-E2-related factor2), a transcription factor, plays a key role in cell oxidative stress response, and controls the expression of various antioxidant response genes after linking to the DNA antioxidant response element (ARE), while melatonin mainly influences the pathway through a nuclear retinoid-related orphan receptor (RZR/RORα) (Giudice et al., 2010).
Melatonin also plays an antioxidant role in coordination with reduced glutathione, nicotinamide adenine dinucleotide phosphate (NADPH), vitamin C, vitamin E, etc. (Yang C.H. et al., 2019). Its synergistic anti-inflammatory effects are mainly realized by up-regulating the activity of enzymes synthesizing such antioxidants so as to increase total content of antioxidants in the organism (Yoon et al., 2019). The anti-inflammatory mechanisms of melatonin include inhibiting the aggregation of inflammatory cells and the release of inflammatory cytokines including TNF-α, IL-1β, and IL-6 all of which are important inflammatory mediators of the inflammatory response. These inflammatory factors directly cause tissue injury and also stimulate other inflammatory cells to release inflammatory mediators, causing a chain reaction (Amin et al., 2019). Experiments show that melatonin increases the release of anti-inflammatory mediators such as IL-10 and while inhibiting the release of inflammatory mediators such as TNF-α, IL-1β, and IL-6, thus achieving an anti-inflammatory effect. In addition, animal experiments show that anti-inflammatory actives of melatonin are related to its inhibition of adhesion molecule related gene expression (Hu C. et al., 2017).
The TLRs mentioned above are the main response pathway of the inflammatory response after myocardial infarction. In a rat myocardial infarction model (Zhao Y. et al., 2019), the expression of the TLR4 signaling pathway is inhibited when melatonin is injected into the heart before ischemic injury. In addition, melatonin also blocks upstream signals (e.g., lipopolysaccharide binding protein CD14) of TLR4. This process can significantly reduce the release of inflammatory factors such as Granulocyte-Monocyte Colony-Stimulating Factor (GM-CSF), TNF-α, C-C Motif Chemokine Ligand 2 (CCL 2), IL-1β, IL-6, C-reactive protein (CRP), serum amyloid A, α-1 antitrypsin, while the content of Nrf2, IL-1α, heme oxygenase-1, and other anti-inflammatory cytokines rise significantly (Ter Horst et al., 2018; Ostjen et al., 2019; Tang et al., 2019).
Dyslipidemia is an independent risk factor for attack of coronary heart disease (CHD). It is reported that (Tengattini et al., 2008) melatonin regulates blood lipid, and reduces the Ox LDL-C, both of which are helpful for reducing the overall incidence of myocardial infarction injury. In terms of protecting vascular endothelial cells, melatonin reduces the degree of injury of endothelial cells in the process of myocardial infarction by inhibiting the activity of myosin light streptokinase, thus delaying the progression of atherosclerosis (Rezzani et al., 2013). In addition, several studies demonstrate that melatonin induces calcium overload and ROS generation in platelets, and activates caspase pathway and depolarizes mitochondrial membrane to mediate platelet inactivation, and this process also involves peroxisome proliferator-activated receptor γ(PPARγ)/ FUN14 domain containing 1 (FUNDC1) /mitophagy pathways (Zhou et al., 2017b; NaveenKumar et al., 2019). In addition to directly mediating platelet dysfunction, melatonin also down-regulates adhesion molecules and delays NO metabolism, thus indirectly inhibiting platelet aggregation (Girish et al., 2013). This antithrombotic effect reduces cardiomyocyte injury after myocardial infarction, and plays a role in protecting the myocardium (Dominguez-Rodriguez et al., 2010a). The circadian rhythm of melatonin release significantly reduces the activity of platelets at night, while the early morning with low melatonin levels is often the time for the occurrence of cardiovascular events. Therefore, supplementation of melatonin through external sources may effectively prevent cardiovascular events (Arushanian, 2013; Lansink et al., 2016).
Melatonin Mediates Myocardial Protection Through Receptor and Non-receptor Pathways
In rat models of myocardial infarction, melatonin concentrations in plasma and left ventricle tissue increase sharply within 1 day, and mRNA levels of the MT1, a member of melatonin receptor, rise significantly after 2 weeks, indicating that melatonin may play an endogenous protective role in myocardial infarction (Sallinen et al., 2007). Melatonin has a biological role mainly by being bound to receptors including both membrane and nuclear binding sites. Among them, membrane receptors include melatonin receptor, TNF receptor and Notch receptor. Melatonin nuclear receptors are members of the retinoic acid related orphan nuclear receptor/retinoic acid Z receptor (ROR/RZR) family, including three subtypes: α, β, γ. The subtype α is referred to as novel endogenous myocardial infarction injury defense agent in new development progress. In rats that lack RORα receptors, the size of the myocardial infarct and degree of cardiac dysfunction after cell injury increases significantly (He et al., 2016). The related pathways by which melatonin executes its cardiac protective role via the receptor pathway includes the reperfusion injury salvage kinase (RISK) pathway, SAFE pathway and Notch pathway, with a complex association among the downstream signaling molecules (Botker et al., 2018; Coverstone et al., 2018; Shanmugam et al., 2019; Yarana et al., 2019).
The RISK pathway has an intracellular biological role primarily through the best known melatonin receptors including MT1, MT2, and MT3, all of which belong to G-protein coupled receptor family (GPCR). MT1 and MT2 have a high affinity with melatonin, while MT3 has a low affinity (Cho et al., 2019). Researchers have discovered a large number of MT1 and MT2 in the heart of rats, ducks, and coronary arteries of chicken and human beings, indicating that the cardiovascular system is a major target organ of melatonin (Hukic et al., 2017). A non-specific melatonin receptor antagonist Luzindole, eliminates the protective action of melatonin on cardiomyocytes, thus confirming their role in mediating the protective effect of melatonin on the heart (Pan et al., 2015). There are three downstream signal pathways of MT1/2, namely MAPK-ERK signal pathway, AMP-dependent protein kinase (AMPK) signal pathway and PI3K-Akt signal pathway. The three routes transmit MT1/2 activation signals from extracellular to intracellular level and mediate intracellular second messenger transmission. The downstream signaling molecules of the three pathways are crossed and connected. (a) The MAPK-ERK signaling pathway is mediated by MT1/2, the activation of MAPK-ERK up-regulates of the level of antioxidant factor Nrf2, and Nrf2 couples with DNA antioxidant reaction elements (ARE) to up-regulate the expressions of HO-1, NADPH, quinone oxidoreductase 1 (NQO1), and glutathione s-transferase 1 (GST1), and reduces the expressions of apoptotic proteins, p21 and p38 (Audia et al., 2018; Canugovi et al., 2019; Kim C.Y. et al., 2019; Liu et al., 2019). The activity of the voltage dependent anion channel (VDAC) and the transcription factor of IP3R-cAMP response element binding protein (CREB) are inhibited by activated extracellular signal regulating kinase (ERK), while excessive activation of VDAC and CREB leads to intracellular calcium overload and then causes mitochondrial dysfunction, eventually bringing about cardiomyocyte necrosis (Li et al., 2018; Zhu H. et al., 2018). Activation of this pathway also leads to inactivation of glycogen synthase kinase-3β (GSK-3β) (Nduhirabandi et al., 2012). The downstream effects also involve the activation of endothelial nitric oxide synthase (eNOS), PKC, and p70 ribosomal protein S6 (p70S6), and down-regulation of the expression of apoptosis related factors such as Bax, Bad, and phosphorylation of caspases (Paradies et al., 2015; Eid et al., 2018; Wang S. et al., 2019; Xu N. et al., 2019). In addition, the activation of MAPK-ERK signaling pathway directly inhibits the opening of MPTP. (b) In the AMPK signaling pathway, Nrf2 is also a downstream signaling molecule of AMPK-PKG1α pathway (Wu et al., 2018b; Lu M. et al., 2019). The AMPK pathway and MAPK-ERK pathways are interrelated through Nrf2 and have a synergistic role in antioxidative stress processes and reducing apoptosis (Yu et al., 2018). In addition, the activation of AMPK inhibits the activity of mitochondrial motility related protein Drp1, which promotes mitochondrial fission, thereby activating VDAC-HK opening and ultimately promoting the MPTP opening (Singhanat et al., 2018). SIRT1 and SIRT3 are both important downstream signaling molecules that aid melatonin in its cardioprotective role. SIRT3 is the downstream target of peroxisome proliferator-activated receptor γ co-activator 1α (PGC-1α) which is stimulated by AMPK. The effect of this pathway is to reduce the transfer of Bax to mitochondria, promote the deacetylation of mitochondrial antioxidant enzyme GPX, boost the biosynthesis of mitochondria, and enhance the activity of superoxide dismutase (SOD) (Lombard and Zwaans, 2014; Lochner et al., 2018). (c) The primary downstream molecular effect of the PI3K-Akt signaling pathway is the reduction of cellular oxidative stress. The activation of Akt promotes phosphorylation of signal transducer and activator of transcription 3 (STAT3), thereby elevating TNFα release for cardiac protection (Yu et al., 2016; Kim E.H. et al., 2019). The enhanced activity of GPX and SOD raises the level of Nrf2, thus promoting the Akt signaling pathway (Song et al., 2017; Zhang H. et al., 2019). Melatonin regulates the activity of ERK through the Akt pathway. Signaling molecules in this pathway include Zrt/Irt-like protein 1 (Zip1), brain-derived neurotrophic factor (BDNF) and PPARγ. In the nuclear receptor signaling pathway, melatonin regulates autophagy and Cyt c release through ROR α, and also enhances the expression of the myocardial sarcoplasmic reticulum Ca2+-ATPase (SERCA)2α, sodium-calcium exchange 1 (NCX1), Ryanodine receptor 2 (RyR2), Ca2+-calmodulin-dependent kinase II (CAMKII) and other protein-related genes, thus enhancing the ability of cells to process calcium ions and reduce the stress injury to and apoptosis of the cardiomyocytes (Gebhard et al., 2018; Na et al., 2019).
In addition to the conventional melatonin receptor pathway, melatonin also binds to other receptors on the cell membrane for signal transduction, including the SAFE pathway and the Notch pathway. In the SAFE pathway, melatonin plays a role in phosphorylation of JAK2-STAT3 through TNF receptor on the cell membrane. Downstream molecular effects include the promotion of expression of BCL-2, antioxidant genes, TNFα, mcl 1, FAS and the inhibition of Bax, caspase-3, Cyt c, cyclin-dependent kinase (cyclin D1), P21 and GSK-3β (Yang et al., 2013). Melatonin also directly inhibits MPTP opening through this pathway. Phosphorylation of STAT3 activates the ERK and Akt pathways, which also promote phosphorylation of STAT3. In the Notch pathway, melatonin promotes the expression of Hairy and enhancer of split 1 (Hes 1) through Notch 1-Notch Intracellular area (NICD), while Hes1 inhibits the negative regulatory effect of chromosome 10 (PTEN) on phosphatidylinositol 3-kinase (PI3K). Notch pathway also reduces the effects of cardiomyocyte apoptosis by regulating mitophagy with mitochondrial fusion related protein (Mfn2) (Pei et al., 2016).
In addition to binding to receptors, melatonin also enters cells where it has direct biological effects (Mauriz et al., 2013). Melatonin enters the cytosol to promote the release of NO, enhances the activity of nitric oxide synthase (iNOS) and boosts the expression of SIRT3 via the activation of PKB-Akt. Activated by melatonin, SIRT1 regulates oxidative stress in cardiomyocytes, mitophagy and apoptosis by enhancing the expression of Bcl-2 and weakening Bax and caspase-3. Studies have shown that SIRT1 is an important upstream molecule of Nrf2 and can also be bound to the SIRT3 promoter to enhance the expression of SIRT3. However, whether melatonin regulates the expression of SIRT3 through the SIRT1-Nrf2 pathway requires verification (Figure 1).
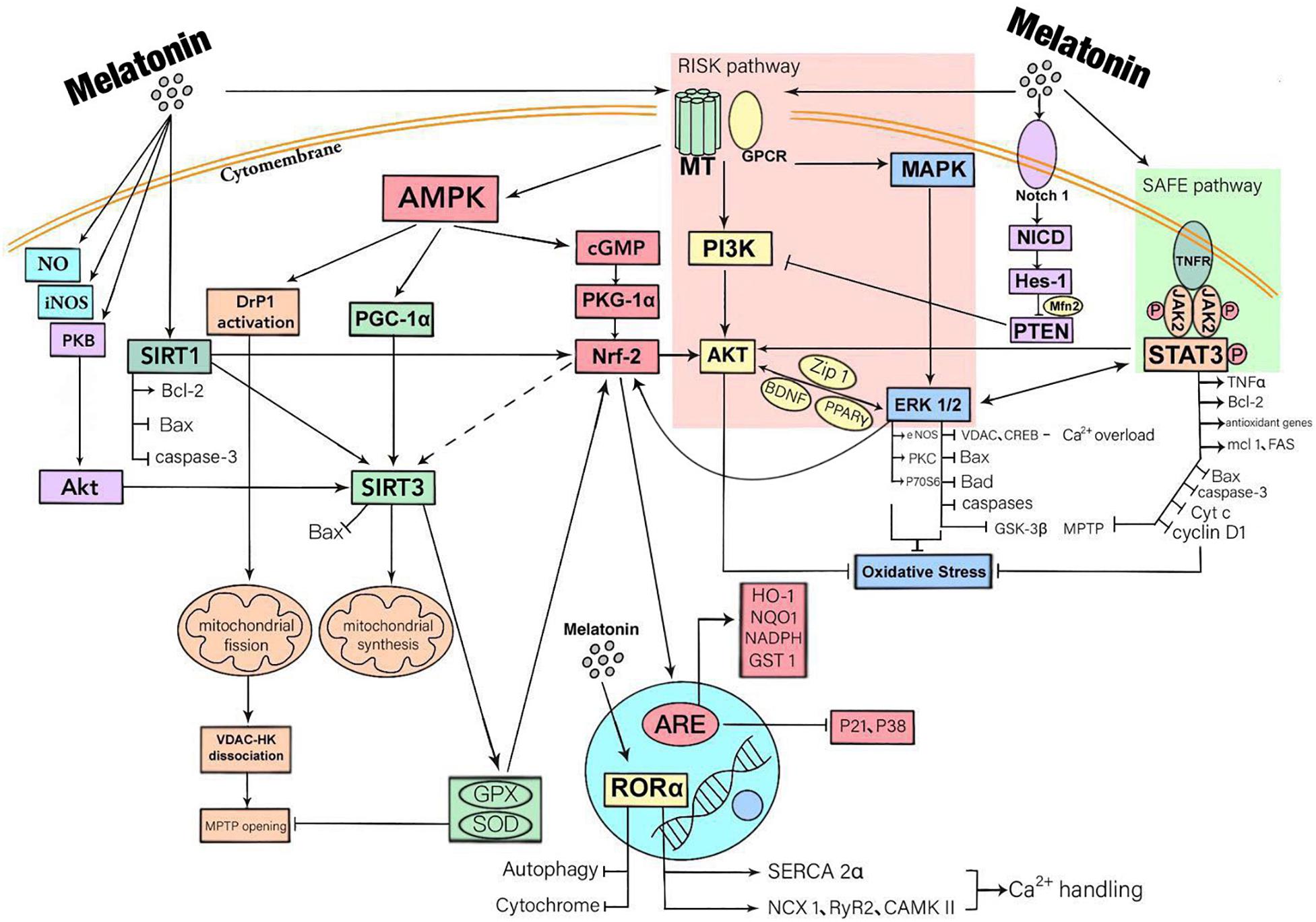
Figure 1. A summary of the mechanisms that myocardial protection mediated by melatonin through receptor and non-receptor pathways. Melatonin has a biological role mainly by being bound to receptors. There are three downstream signal pathways of MT1/2, namely MAPK-ERK signal pathway, AMP-dependent protein kinase (AMPK) signal pathway and PI3K-Akt signal pathway. Melatonin also activates STAT3, a signal transducer and activator of transcription factor for antioxidant enzymes, by activating SAFE pathway. Besides, melatonin activates the Notch1 pathway, thus inhibiting PIK3 function. These downstream signaling molecules are crossed and connected within the cell. In addition, melatonin also enters cells where it has direct biological effects. It promotes the release of NO, enhances the activity of iNOS and boosts the expression of SIRT3 via the activation of PKB-Akt. The ultimate effects of these responses are to reduce oxidative stress, inflammatory responses, and to protect mitochondrial function, thereby reducing cardiomyocyte apoptosis.
Melatonin Stabilizes the Structure and Function of Mitochondria After AMI
Mitochondria are the site of ATP and oxygen-derived free radical production, and the target of attack by various free radicals (Connolly et al., 2018; Boengler et al., 2019; Yuan et al., 2019). Melatonin protects cardiomyocytes by stabilizing the structure and function of mitochondria and regulates mitochondrial oxidative stress, raises mitochondrial antioxidant enzyme levels, restores mitochondrial energy metabolism (Soto-Heras et al., 2019), maintains mitochondrial membrane potential stability, reduces mitochondrial injury, and inhibits mitochondrial apoptosis (Yan et al., 2018) through receptors including MT1/2. In addition, mitochondrial biosynthesis, DNA homeostasis and regulation of SIRT3 system are closely related to the function of melatonin (Reiter et al., 2017; Zhou et al., 2017d). After myocardial infarction, injured tissue releases a large quantity of oxygen-derived free radicals, inflammatory mediators and other harmful substances, which cause direct injury to mitochondria (Kalkavan and Green, 2018; Riehle and Bauersachs, 2018; Nanadikar et al., 2019). Mitochondrial dysfunction aggravates cell injury, which then becomes a vicious cycle. Melatonin breaks this vicious cycle by virtue of its potent free radical scavenging ability and antioxidant effects, thus playing a protective role in myocardium such as in ischemia-reperfusion injury (Reina and Martínez, 2018). Studies show that melatonin, as a potent free radical scavenger, is abundant in mitochondria (Venegas et al., 2012). This shows that melatonin likely prevents mitochondrial injury during the oxidative stress response (Ma et al., 2016). Melatonin also up-regulates the activity of the four respiratory complexes, thus reducing electron leakage and the generation of oxygen-derived free radicals (Nair et al., 2011). Melatonin inhibits mitochondrion fission, prevents disintegration of VDAC1 and hexokinase 2 (HK2), inhibits MTPT opening, limits endothelial cell injury, improves endothelial barrier function, reduces inflammatory cell infiltration, restores eNOS content and blood flow, lowers infarct size, preserves myocardial microvasculature, and improve prognosis (Zhang and Zhang, 2014; Li et al., 2019). Based on published reports (Wang et al., 2016), activation of MT1/2 receptor strengthens AMPK signaling pathway, up-regulates optic atrophy 1 (OPA1) level and then modulates mitophagy during myocardial infarction. Mitophagy degrades injured mitochondria so as to maintain the structure and function of normal mitochondria, reduces cardiomyocyte apoptosis and lowers the degree of myocardial injury during infarction (Dominguez-Rodriguez et al., 2017a; Zhang Y. et al., 2019).
Clinical Application and Prospect of Melatonin Use in Acute Myocardial Infarction
Due to the non-renewability of cardiomyocytes, it is essential to reduce cardiac damage and improve long-term prognosis of patients with myocardial infarction by clarifying the mechanisms of injury and necrosis of cardiomyocytes after a cardiovascular episode (Gaspar et al., 2018; Wu et al., 2018a; Heusch, 2019). In addition to the mechanisms of inflammatory cell infiltration and oxidative stress injury which have been reported many times, mitochondrial injury and apoptosis are the key initiating processes of myocardial injury following myocardial infarction (Hofmann, 2018; Villalobos et al., 2018; Wang Y. et al., 2018; Boengler et al., 2019; Phan et al., 2019). There is an urgent need to find a method which can effectively reduce mitochondrial injury and to improve cardiomyocyte function following myocardial infarction and provide a new target for clinical treatment.
As an indole and age-related molecule secreted by the human pineal gland, melatonin has remarkable functions including antioxidant, anti-apoptosis, anti-fibrosis, direct free radical scavenging, and mitochondrial protection (Jumnongprakhon et al., 2016). Melatonin receptors also play a role in reducing cardiac damage (Liu et al., 2016). In models of myocardial reperfusion injury, melatonin promotes mitochondrial fusion through AMPK/OPA1 signaling pathway and OPA1 binds with lysine 70 residues of mitophagy receptor FUNDC1 to mediate mitophagy (Del Dotto et al., 2018; Wang B. et al., 2018). Recent studies reveal that mitochondria exhibit biological rhythms, which may be regulated by the melatonin cycle. However, studies have not been conducted to clarify the role of melatonin in the mitochondrial biological clock and the myocardial protective effects that involve these rhythms. As mentioned above, melatonin is secreted in large quantities at night according to its circadian rhythm. Studies illustrates that the nocturnal secretion of melatonin in patients with CHD decreases significantly compared with healthy people and it may be associated with various disease risk factors in these patients which can explain why AMI always happen (at a peak) in early morning when melatonin levels are adequate (Dominguez-Rodriguez et al., 2006, 2007a).
In terms of clinical disease treatment, most studies on melatonin-induced cardiovascular effect are in phase 2a of clinical trials, and according to literatures, melatonin has a beneficial therapeutic effect on hypertension, atherosclerosis, CHD, and other chronic cardiovascular diseases (Yang et al., 2014; Simko et al., 2016; Baltatu et al., 2019) (Table 1). Since it is an age-related molecule, it declines with age. Because of its high safety profile, it has important potential clinical applications. Clinical trials demonstrate that melatonin significantly reduce the area of myocardial infarction when used in the treatment of STEMI (ST-elevation myocardial infarction) patients after PCI (Dominguez-Rodriguez et al., 2017a). In addition, the use of melatonin before surgery significantly reduces CABG related oxidative stress and cardiac injury, and on the other hand increase the activity of Nrf2 in patients (Haghjooy Javanmard et al., 2013; Shafiei et al., 2018). Melatonin plays a role in reducing nocturnal hypertension in patients by affecting circadian cardiovascular rhythms of blood pressure (Grossman et al., 2011). The experimental results of Ma et al. (2018) show that melatonin eliminates ROS by regulating the mitophagy in macrophages, thus inhibiting the activation of NLRP3 (nucleotide-binding domain and leucine-rich repeat pyrin domain containing 3) inflammasome and ultimately inhibiting the progression of atherosclerosis, which is mediated at least in part through the Sirt3 signaling pathway.
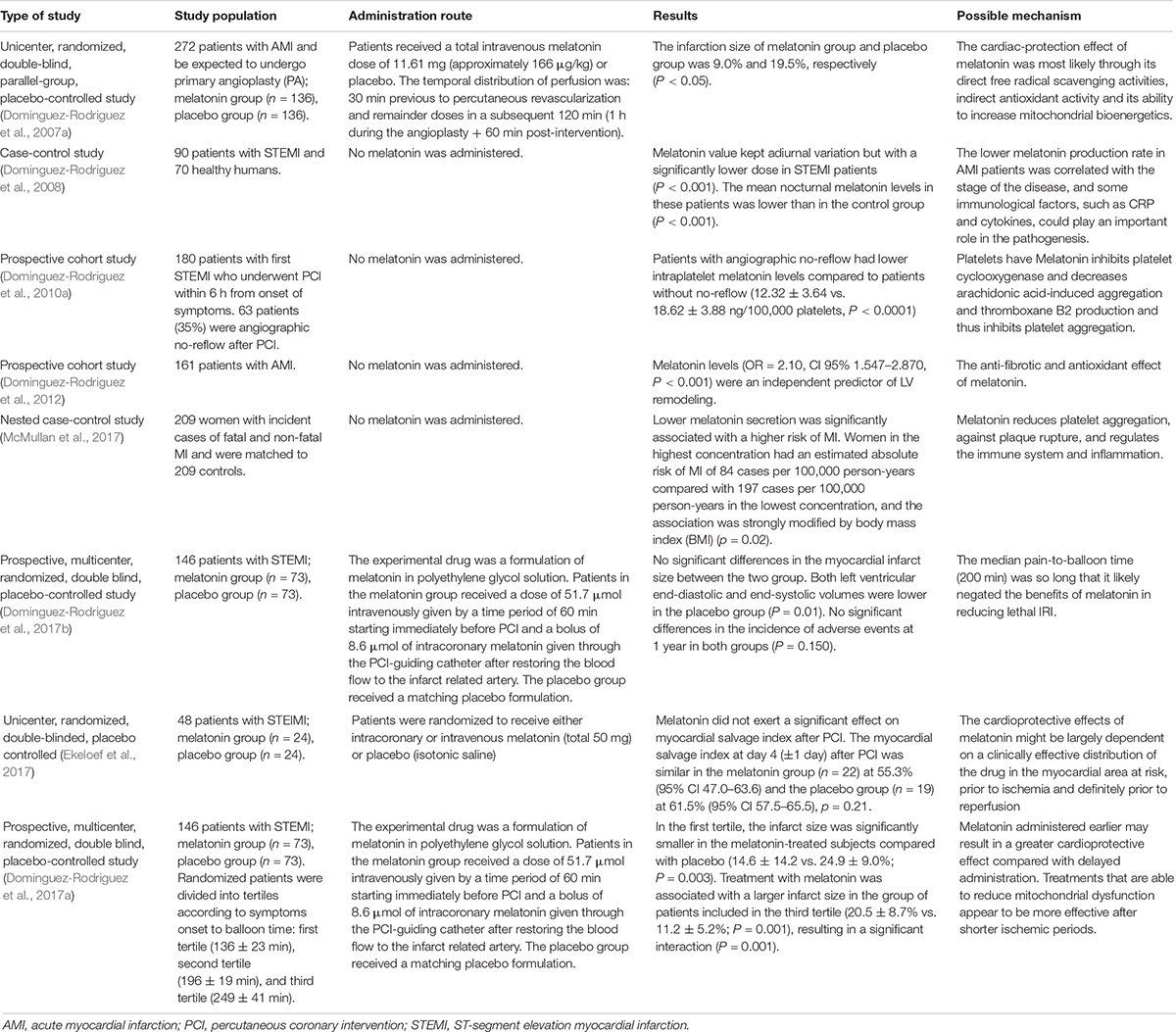
Table 1. A summary of the results of some clinical trials (there are many more), which illustrate the beneficial effects of melatonin in clinical acute myocardial infarction.
Studies reveal that (Dominguez-Rodriguez et al., 2017a), from the onset of myocardial infarction symptoms to the beginning of interventional therapy, the application of melatonin in a short time window can effectively reduce the size of a myocardial infarct. Repair after a myocardial infarction is a complex process involved with multiple factors, but whether melatonin is involved with these repair processes have not been investigated. The combination of melatonin with other myocardial protective drugs (e.g., antithrombotic drugs and new-type myocardial metabolic regulation drug GLP1) has not yet been reported, and further research is required to confirm the myocardial protective actions of melatonin treatment.
Author Contributions
ZF conceived and designed the review. ZF, YJ, JW, YZ, and MS collected the literatures. YJ and ZF wrote the manuscript. ZF, RR, and YC reviewed and edited the manuscript. QX revised the manuscript and the language. All authors read and approved the manuscript.
Funding
This study was supported by National Natural Science Foundation of China (fund numbers 81500269, 81500202, and 81570383).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Abdelnaseer, M., Elfayomi, N., Esmail, E. H., Kamal, M. M., Hamdy, A., Samie, R. M. A., et al. (2016). Relationship between matrix metalloproteinase-9 and common carotid artery intima media thickness. Neurol. Sci. 37, 117–122. doi: 10.1007/s10072-015-2358-z
Abukar, Y., Ramchandra, R., Hood, S. G., McKinley, M. J., Booth, L. C., Yao, S. T., et al. (2018). Increased cardiac sympathetic nerve activity in ovine heart failure is reduced by lesion of the area postrema, but not lamina terminalis. Basic Res. Cardiol. 113:35.
Adameova, A., Goncalvesova, E., Szobi, A., and Dhalla, N. S. (2016). Necroptotic cell death in failing heart: relevance and proposed mechanisms. Heart Fail. Rev. 21, 213–221. doi: 10.1007/s10741-016-9537-8
Aimo, A., Cerbai, E., Bartolucci, G., Adamo, L., Barison, A., Lo Surdo, G., et al. (2020). Pirfenidone is a cardioprotective drug: mechanisms of action and preclinical evidence. Pharmacol. Res. 155:104694. doi: 10.1016/j.phrs.2020.104694
Amanakis, G., Kleinbongard, P., Heusch, G., and Skyschally, A. (2019). Attenuation of ST-segment elevation after ischemic conditioning maneuvers reflects cardioprotection online. Basic Res. Cardiol. 114:22. doi: 10.1007/s00395-019-0732-3
Amin, N., Shafabakhsh, R., Reiter, R. J., and Asemi, Z. (2019). Melatonin is an appropriate candidate for breast cancer treatment: based on known molecular mechanisms. J. Cell. Biochem. 120, 12208–12215. doi: 10.1002/jcb.28832
Arushanian, E. B. (2013). Effect of melatonin on the thrombocyte hemostasis and its circadian organization. Eksp. Klin. Farmakol. 76, 32–36.
Audia, J. P., Yang, X. M., Crockett, E. S., Housley, N., Haq, E. U., O’Donnell, K., et al. (2018). Caspase-1 inhibition by VX-765 administered at reperfusion in P2Y12 receptor antagonist-treated rats provides long-term reduction in myocardial infarct size and preservation of ventricular function. Basic Res. Cardiol. 113:32. doi: 10.1007/s00395-018-0692-z
Bacmeister, L., Schwarzl, M., Warnke, S., Stoffers, B., Blankenberg, S., Westermann, D., et al. (2019). Inflammation and fibrosis in murine models of heart failure. Basic Res. Cardiol. 114:19.
Baltatu, O. C., Senar, S., Campos, L. A., and Cipolla-Neto, J. (2019). Cardioprotective melatonin: translating from proof-of-concept studies to therapeutic use. Int. J. Mol. Sci. 20:4342. doi: 10.3390/ijms20184342
Basalay, M. V., Davidson, S. M., Gourine, A. V., and Yellon, D. M. (2018). Neural mechanisms in remote ischaemic conditioning in the heart and brain: mechanistic and translational aspects. Basic Res. Cardiol. 113:25. doi: 10.1007/s00395-018-0684-z
Battogtokh, G., Choi, Y. S., Kang, D. S., Park, S. J., Shim, M. S., Huh, K. M., et al. (2018). Mitochondria-targeting drug conjugates for cytotoxic, anti-oxidizing and sensing purposes: current strategies and future perspectives. Acta Pharm. Sin. B 8, 862–880. doi: 10.1016/j.apsb.2018.05.006
Beckendorf, J., van den Hoogenhof, M. M. G., and Backs, J. (2018). Physiological and unappreciated roles of CaMKII in the heart. Basic Res. Cardiol. 113:29.
Bhandari, P., Song, M., Chen, Y., Burelle, Y., and Dorn, G. W. (2014). Mitochondrial contagion induced by Parkin deficiency in Drosophila hearts and its containment by suppressing mitofusin. Circ. Res. 114, 257–265. doi: 10.1161/CIRCRESAHA.114.302734
Bocci, M., Sjolund, J., Kurzejamska, E., Lindgren, D., Marzouka, N. A., Bartoschek, M., et al. (2019). Activin receptor-like kinase 1 is associated with immune cell infiltration and regulates CLEC14A transcription in cancer. Angiogenesis 22, 117–131. doi: 10.1007/s10456-018-9642-5
Boengler, K., Bornbaum, J., Schluter, K. D., and Schulz, R. (2019). P66shc and its role in ischemic cardiovascular diseases. Basic Res. Cardiol. 114:29. doi: 10.1007/s00395-019-0738-x
Boengler, K., Lochnit, G., and Schulz, R. (2018). Mitochondria “THE” target of myocardial conditioning. Am. J. Physiol. Heart Circ. Physiol. 315, H1215–H1231. doi: 10.1152/ajpheart.00124.2018
Botker, H. E., Hausenloy, D., Andreadou, I., Antonucci, S., Boengler, K., Davidson, S. M., et al. (2018). Practical guidelines for rigor and reproducibility in preclinical and clinical studies on cardioprotection. Basic Res. Cardiol. 113:39.
Breda, C. N. S., Davanzo, G. G., Basso, P. J., Saraiva Camara, N. O., and Moraes-Vieira, P. M. M. (2019). Mitochondria as central hub of the immune system. Redox Biol. 26:101255. doi: 10.1016/j.redox.2019.101255
Canugovi, C., Stevenson, M. D., Vendrov, A. E., Hayami, T., Robidoux, J., Xiao, H., et al. (2019). Increased mitochondrial NADPH oxidase 4 (NOX4) expression in aging is a causative factor in aortic stiffening. Redox Biol. 26:101288. doi: 10.1016/j.redox.2019.101288
Cao, T., Fan, S., Zheng, D., Wang, G., Yu, Y., Chen, R., et al. (2019). Increased calpain-1 in mitochondria induces dilated heart failure in mice: role of mitochondrial superoxide anion. Basic Res. Cardiol. 114:17.
Chen, M., Ye, K., Zhang, B., Xin, Q., Li, P., Kong, A. N., et al. (2019). Paris Saponin II inhibits colorectal carcinogenesis by regulating mitochondrial fission and NF-kappaB pathway. Pharmacol. Res. 139, 273–285. doi: 10.1016/j.phrs.2018.11.029
Chen, S., Dong, G., Wu, S., Liu, N., Zhang, W., and Sheng, C. (2019). Novel fluorescent probes of 10-hydroxyevodiamine: autophagy and apoptosis-inducing anticancer mechanisms. Acta Pharm. Sin. B 9, 144–156. doi: 10.1016/j.apsb.2018.08.003
Chen, W., Spitzl, A., Mathes, D., Nikolaev, V. O., Werner, F., Weirather, J., et al. (2016). Endothelial actions of ANP enhance myocardial inflammatory infiltration in the early phase after acute infarction. Circ. Res. 119, 237–248. doi: 10.1161/CIRCRESAHA.115.307196
Cho, J. H., Tae, H. J., Kim, I. S., Song, M., Kim, H., Lee, T. K., et al. (2019). Melatonin alleviates asphyxial cardiac arrest-induced cerebellar Purkinje cell death by attenuation of oxidative stress. Exp. Neurol. 320:112983. doi: 10.1016/j.expneurol.2019.112983
Cohen, A. T., Berger, S. E., Milenkovic, D., Hill, N. R., and Lister, S. (2019). Anticoagulant selection for patients with VTE-Evidence from a systematic literature review of network meta-analyses. Pharmacol. Res. 143, 166–177. doi: 10.1016/j.phrs.2019.03.017
Connolly, N. M. C., Theurey, P., Adam-Vizi, V., Bazan, N. G., Bernardi, P., Bolanos, J. P., et al. (2018). Guidelines on experimental methods to assess mitochondrial dysfunction in cellular models of neurodegenerative diseases. Cell Death Differ. 25, 542–572.
Coverstone, E. D., Bach, R. G., Chen, L., Bierut, L. J., Li, A. Y., Lenzini, P. A., et al. (2018). A novel genetic marker of decreased inflammation and improved survival after acute myocardial infarction. Basic Res. Cardiol. 113:38.
Crist, A. M., Lee, A. R., Patel, N. R., Westhoff, D. E., and Meadows, S. M. (2018). Vascular deficiency of Smad4 causes arteriovenous malformations: a mouse model of Hereditary Hemorrhagic Telangiectasia. Angiogenesis 21, 363–380. doi: 10.1007/s10456-018-9602-0
Davidson, S. M., Arjun, S., Basalay, M. V., Bell, R. M., Bromage, D. I., Botker, H. E., et al. (2018). The 10th biennial hatter cardiovascular institute workshop: cellular protection-evaluating new directions in the setting of myocardial infarction, ischaemic stroke, and cardio-oncology. Basic Res. Cardiol. 113:43. doi: 10.1007/s00395-018-0704-z
de Oliveira Junior, E. R., Nascimento, T. L., Salomao, M. A., da Silva, A. C. G., Valadares, M. C., and Lima, E. M. (2019). Increased nose-to-brain delivery of melatonin mediated by polycaprolactone nanoparticles for the treatment of glioblastoma. Pharm. Res. 36:131. doi: 10.1007/s11095-019-2662-z
Del Dotto, V., Fogazza, M., Lenaers, G., Rugolo, M., Carelli, V., and Zanna, C. (2018). OPA1: how much do we know to approach therapy? Pharmacol. Res. 131, 199–210. doi: 10.1016/j.phrs.2018.02.018
Dominguez-Rodriguez, A., Abreu-Gonzalez, P., Arroyo-Ucar, E., and Reiter, R. J. (2012). Decreased level of melatonin in serum predicts left ventricular remodelling after acute myocardial infarction. J. Pineal Res. 53, 319–323. doi: 10.1111/j.1600-079X.2012.01001.x
Dominguez-Rodriguez, A., Abreu-Gonzalez, P., de la Torre-Hernandez, J. M., Consuegra-Sanchez, L., Piccolo, R., Gonzalez-Gonzalez, J., et al. (2017a). Usefulness of early treatment with melatonin to reduce infarct size in patients with ST-segment elevation myocardial infarction receiving percutaneous coronary intervention (from the melatonin adjunct in the acute myocardial infarction treated with angioplasty trial). Am. J. Cardiol. 120, 522–526. doi: 10.1016/j.amjcard.2017.05.018
Dominguez-Rodriguez, A., Abreu-Gonzalez, P., de la Torre-Hernandez, J. M., Gonzalez-Gonzalez, J., Garcia-Camarero, T., Consuegra-Sanchez, L., et al. (2017b). Effect of intravenous and intracoronary melatonin as an adjunct to primary percutaneous coronary intervention for acute ST-elevation myocardial infarction: results of the Melatonin Adjunct in the acute myocaRdial Infarction treated with Angioplasty trial. J. Pineal Res. 62:e12374. doi: 10.1111/jpi.12374
Dominguez-Rodriguez, A., Abreu-Gonzalez, P., Garcia-Gonzalez, M., and Reiter, R. J. (2006). Prognostic value of nocturnal melatonin levels as a novel marker in patients with ST-segment elevation myocardial infarction. Am. J. Cardiol. 97, 1162–1164. doi: 10.1016/j.amjcard.2005.11.033
Dominguez-Rodriguez, A., Abreu-Gonzalez, P., Garcia-Gonzalez, M. J., Kaski, J. C., Reiter, R. J., and Jimenez-Sosa, A. (2007a). A unicenter, randomized, double-blind, parallel-group, placebo-controlled study of melatonin as an adjunct in patients with acute myocardial infarction undergoing primary angioplasty the melatonin adjunct in the acute myocardial infarction treated with angioplasty (MARIA) trial: study design and rationale. Contemp. Clin. Trials 28, 532–539. doi: 10.1016/j.cct.2006.10.007
Dominguez-Rodriguez, A., Abreu-Gonzalez, P., Garcia-Gonzalez, M. J., and Reiter, R. J. (2007b). Relation of nocturnal melatonin levels to serum matrix metalloproteinase-9 concentrations in patients with myocardial infarction. Thromb. Res. 120, 361–366. doi: 10.1016/j.thromres.2006.10.010
Dominguez-Rodriguez, A., Abreu-Gonzalez, P., Garcia-Gonzalez, M. J., Samimi-Fard, S., Kaski, J. C., and Reiter, R. J. (2008). Light/dark patterns of soluble vascular cell adhesion molecule-1 in relation to melatonin in patients with ST-segment elevation myocardial infarction. J. Pineal Res. 44, 65–69. doi: 10.1111/j.1600-079X.2007.00529.x
Dominguez-Rodriguez, A., Abreu-Gonzalez, P., Jimenez-Sosa, A., Avanzas, P., Bosa-Ojeda, F., and Kaski, J. C. (2010a). Usefulness of intraplatelet melatonin levels to predict angiographic no-reflow after primary percutaneous coronary intervention in patients with ST-segment elevation myocardial infarction. Am. J. Cardiol. 106, 1540–1544. doi: 10.1016/j.amjcard.2010.07.030
Dominguez-Rodriguez, A., Abreu-Gonzalez, P., Sanchez-Sanchez, J. J., Kaski, J. C., et al. (2010b). Melatonin and circadian biology in human cardiovascular disease. J. Pineal Res. 49, 14–22. doi: 10.1111/j.1600-079X.2010.00773.x
Eid, R. A., Alkhateeb, M. A., Eleawa, S., Al-Hashem, F. H., Al-Shraim, M., El-Kott, A. F., et al. (2018). Cardioprotective effect of ghrelin against myocardial infarction-induced left ventricular injury via inhibition of SOCS3 and activation of JAK2/STAT3 signaling. Basic Res. Cardiol. 113:13.
Ekeloef, S., Halladin, N., Fonnes, S., Jensen, S. E., Zaremba, T., Rosenberg, J., et al. (2017). Effect of intracoronary and intravenous melatonin on myocardial salvage index in patients with ST-elevation myocardial infarction: a randomized placebo controlled trial. J. Cardiovasc. Transl. Res. 10, 470–479. doi: 10.1007/s12265-017-9768-7
Espinosa-Diez, C., Miguel, V., Vallejo, S., Sanchez, F. J., Sandoval, E., Blanco, E., et al. (2018). Role of glutathione biosynthesis in endothelial dysfunction and fibrosis. Redox Biol. 14, 88–99. doi: 10.1016/j.redox.2017.08.019
Faghfouri, A. H., Zarrin, R., Maleki, V., Payahoo, L., and Khajebishak, Y. (2020). A comprehensive mechanistic review insight into the effects of micronutrients on toll-like receptors functions. Pharmacol. Res. 152:104619. doi: 10.1016/j.phrs.2019.104619
Filadi, R., Greotti, E., and Pizzo, P. (2018). Highlighting the endoplasmic reticulum-mitochondria connection: focus on mitofusin 2. Pharmacol. Res. 128, 42–51. doi: 10.1016/j.phrs.2018.01.003
Fuhrmann, D. C., and Brune, B. (2017). Mitochondrial composition and function under the control of hypoxia. Redox Biol. 12, 208–215. doi: 10.1016/j.redox.2017.02.012
Galano, A., and Reiter, R. J. (2018). Melatonin and its metabolites vs oxidative stress: from individual actions to collective protection. J. Pineal Res. 65:e12514. doi: 10.1111/jpi.12514
Galano, A., Tan, D. X., and Reiter, R. J. (2013). On the free radical scavenging activities of melatonin’s metabolites, AFMK and AMK. J. Pineal Res. 54, 245–257. doi: 10.1111/jpi.12010
Galluzzi, L., Vitale, I., Aaronson, S. A., Abrams, J. M., Adam, D., Agostinis, P., et al. (2018). Molecular mechanisms of cell death: recommendations of the nomenclature committee on cell death 2018. Cell Death Differ. 25, 486–541.
Garcia, J. A., Volt, H., Venegas, C., Doerrier, C., Escames, G., Lopez, L. C., et al. (2015). Disruption of the NF-kappaB/NLRP3 connection by melatonin requires retinoid-related orphan receptor-alpha and blocks the septic response in mice. FASEB J. 29, 3863–3875. doi: 10.1096/fj.15-273656
Gaspar, A., Lourenco, A. P., Pereira, M. A., Azevedo, P., and Roncon-Albuquerque, R. Jr., et al. (2018). Randomized controlled trial of remote ischaemic conditioning in ST-elevation myocardial infarction as adjuvant to primary angioplasty (RIC-STEMI). Basic Res. Cardiol. 113:14.
Gebhard, C., Maafi, F., Stahli, B. E., Dang, J., Nachar, W., de Oliveira Moraes, A. B., et al. (2018). Apolipoprotein A-I proteolysis in aortic valve stenosis: role of cathepsin S. Basic Res. Cardiol. 113:30. doi: 10.1007/s00395-018-0689-7
Girish, K. S., Paul, M., Thushara, R. M., Hemshekhar, M., Shanmuga Sundaram, M., Rangappa, K. S., et al. (2013). Melatonin elevates apoptosis in human platelets via ROS mediated mitochondrial damage. Biochem. Biophys. Res. Commun. 438, 198–204. doi: 10.1016/j.bbrc.2013.07.053
Giudice, A., Arra, C., and Turco, M. C. (2010). Review of molecular mechanisms involved in the activation of the Nrf2-ARE signaling pathway by chemopreventive agents. Methods Mol. Biol. 647, 37–74. doi: 10.1007/978-1-60761-738-9_3
Graczyk-Jarzynka, A., Goral, A., Muchowicz, A., Zagozdzon, R., Winiarska, M., Bajor, M., et al. (2019). Inhibition of thioredoxin-dependent H2O2 removal sensitizes malignant B-cells to pharmacological ascorbate. Redox Biol. 21:101062. doi: 10.1016/j.redox.2018.11.020
Grossman, E., Laudon, M., and Zisapel, N. (2011). Effect of melatonin on nocturnal blood pressure: meta-analysis of randomized controlled trials. Vasc. Health Risk Manag. 7, 577–584. doi: 10.2147/VHRM.S24603
Guidarelli, A., Fiorani, M., Cerioni, L., and Cantoni, O. (2019). Calcium signals between the ryanodine receptor- and mitochondria critically regulate the effects of arsenite on mitochondrial superoxide formation and on the ensuing survival vs apoptotic signaling. Redox Biol. 20, 285–295. doi: 10.1016/j.redox.2018.10.015
Hadebe, N., Cour, M., and Lecour, S. (2018). The SAFE pathway for cardioprotection: is this a promising target? Basic Res. Cardiol. 113:9. doi: 10.1007/s00395-018-0670-5
Haghjooy Javanmard, S., Ziaei, A., Ziaei, S., Ziaei, E., and Mirmohammad-Sadeghi, M. (2013). The effect of preoperative melatonin on nuclear erythroid 2-related factor 2 activation in patients undergoing coronary artery bypass grafting surgery. Oxid. Med. Cell. Longev. 2013:676829. doi: 10.1155/2013/676829
He, B., Zhao, Y. C., Xu, L. W., Gao, L. C., Su, Y. Y., Lin, N., et al. (2016). The nuclear melatonin receptor RORα is a novel endogenous defender against myocardial ischemia/reperfusion injury. J. Pineal Res. 60, 313–326. doi: 10.1111/jpi.12312
Heckmann, B. L., Tummers, B., and Green, D. R. (2019). Crashing the computer: apoptosis vs. necroptosis in neuroinflammation. Cell Death Differ. 26, 41–52. doi: 10.1038/s41418-018-0195-3
Heusch, G. (2019). Coronary microvascular obstruction: the new frontier in cardioprotection. Basic Res. Cardiol. 114:45. doi: 10.1007/s00395-019-0756-8
Hockings, C., Alsop, A. E., Fennell, S. C., Lee, E. F., Fairlie, W. D., Dewson, G., et al. (2018). Mcl-1 and Bcl-xL sequestration of Bak confers differential resistance to BH3-only proteins. Cell Death Differ. 25, 719–732. doi: 10.1038/s41418-017-0010-6
Hofmann, F. (2018). A concise discussion of the regulatory role of cGMP kinase I in cardiac physiology and pathology. Basic Res. Cardiol. 113:31. doi: 10.1007/s00395-018-0690-1
Hu, C., Wang, P., Zhang, S., Ren, L., Lv, Y., Yin, R., et al. (2017). Neuroprotective effect of melatonin on soluble Abeta1-42-induced cortical neurodegeneration via Reelin-Dab1 signaling pathway. Neurol. Res. 39, 621–631. doi: 10.1080/01616412.2017.1312805
Hu, S. Y., Zhang, Y., Zhu, P. J., Zhou, H., and Chen, Y. D. (2017). Liraglutide directly protects cardiomyocytes against reperfusion injury possibly via modulation of intracellular calcium homeostasis. Geriatr. Cardiol. 14, 57–66. doi: 10.11909/j.issn.1671-5411.2017.01.008
Hu, C., Zhang, X., Wei, W., Zhang, N., Wu, H., Ma, Z., et al. (2019). Matrine attenuates oxidative stress and cardiomyocyte apoptosis in doxorubicin-induced cardiotoxicity via maintaining AMPKalpha/UCP2 pathway. Acta Pharm. Sin. B 9, 690–701. doi: 10.1016/j.apsb.2019.03.003
Hukic, D. S., Lavebratt, C., Olsson, E., Ostenson, C. G., Eriksson, S. V., Erlinge, D., et al. (2017). Troponin T levels associated with genetic variants in NOTCH2 and MTNR1B in women with psychosis. Psychiatry Res. 250, 217–220. doi: 10.1016/j.psychres.2017.01.030
Jager, S. C. A. D., and Hoefer, I. E. (2019). Local inflammatory responses take their toll on the heart. Int. J. Cardiol. 293, 254–255. doi: 10.1016/j.ijcard.2019.07.055
Ji, C. H., and Kwon, Y. T. (2017). Crosstalk and interplay between the ubiquitin-proteasome system and autophagy. Mol. Cells 40, 441–449. doi: 10.14348/molcells.2017.0115
Jiang, Q., Xiang, B., Wang, H., Huang, K., Kong, H., and Hu, S. (2019). Remote ischaemic preconditioning ameliorates sinus rhythm restoration rate through Cox maze radiofrequency procedure associated with inflammation reaction reduction. Basic Res. Cardiol. 114:14. doi: 10.1007/s00395-019-0723-4
Jin, Q., Li, R., Hu, N., Xin, T., Zhu, P., Hu, S., et al. (2018). DUSP1 alleviates cardiac ischemia/reperfusion injury by suppressing the Mff-required mitochondrial fission and Bnip3-related mitophagy via the JNK pathways. Redox Biol. 14, 576–587. doi: 10.1016/j.redox.2017.11.004
Joshi, A. U., and Mochly-Rosen, D. (2018). Mortal engines: mitochondrial bioenergetics and dysfunction in neurodegenerative diseases. Pharmacol. Res. 138, 2–15. doi: 10.1016/j.phrs.2018.08.010
Joshi, C., Bapat, R., Anderson, W., Dawson, D., Hijazi, K., and Cherukara, G. (2019). Detection of periodontal microorganisms in coronary atheromatous plaque specimens of myocardial infarction patients: a systematic review and meta-analysis. Trends Cardiovasc. Med. [Epub ahead of print].
Jumnongprakhon, P., Govitrapong, P., Tocharus, C., and Tocharus, J. (2016). Inhibitory effect of melatonin on cerebral endothelial cells dysfunction induced by methamphetamine via NADPH oxidase-2. Brain Res. 1650, 84–92. doi: 10.1016/j.brainres.2016.08.045
Kalkavan, H., and Green, D. R. (2018). MOMP, cell suicide as a BCL-2 family business. Cell Death Differ. 25, 46–55. doi: 10.1038/cdd.2017.179
Kanaan, G. N., Ichim, B., Gharibeh, L., Maharsy, W., Patten, D. A., Xuan, J. Y., et al. (2018). Glutaredoxin-2 controls cardiac mitochondrial dynamics and energetics in mice, and protects against human cardiac pathologies. Redox Biol. 14, 509–521. doi: 10.1016/j.redox.2017.10.019
Kang, J. W., Hong, J. M., and Lee, S. M. (2016). Melatonin enhances mitophagy and mitochondrial biogenesis in rats with carbon tetrachloride-induced liver fibrosis. J. Pineal Res. 60, 383–393. doi: 10.1111/jpi.12319
Karwi, Q. G., Bice, J. S., and Baxter, G. F. (2018). Pre- and postconditioning the heart with hydrogen sulfide (H2S) against ischemia/reperfusion injury in vivo: a systematic review and meta-analysis. Basic Res. Cardiol. 113:6. doi: 10.1007/s00395-017-0664-8
Kim, C. Y., Kang, B., Suh, H. J., and Choi, H. S. (2019). Parthenolide, a feverfew-derived phytochemical, ameliorates obesity and obesity-induced inflammatory responses via the Nrf2/Keap1 pathway. Pharmacol. Res. 145:104259. doi: 10.1016/j.phrs.2019.104259
Kim, E. H., Wong, S. W., and Martinez, J. (2019). Programmed necrosis and disease:we interrupt your regular programming to bring you necroinflammation. Cell Death Differ. 26, 25–40. doi: 10.1038/s41418-018-0179-3
Kleszczynski, K., Zillikens, D., and Fischer, T. W. (2016). Melatonin enhances mitochondrial ATP synthesis, reduces reactive oxygen species formation, and mediates translocation of the nuclear erythroid 2-related factor 2 resulting in activation of phase-2 antioxidant enzymes (gamma-GCS, HO-1, NQO1) in ultraviolet radiation-treated normal human epidermal keratinocytes (NHEK). J. Pineal Res. 61, 187–197. doi: 10.1111/jpi.12338
Koentges, C., Pepin, M. E., Musse, C., Pfeil, K., Alvarez, S. V. V., Hoppe, N., et al. (2018). Gene expression analysis to identify mechanisms underlying heart failure susceptibility in mice and humans. Basic Res. Cardiol. 113:8. doi: 10.1007/s00395-017-0666-6
Kohlstedt, K., Trouvain, C., Fromel, T., Mudersbach, T., Henschler, R., and Fleming, I. (2018). Role of the angiotensin-converting enzyme in the G-CSF-induced mobilization of progenitor cells. Basic Res. Cardiol. 113:18. doi: 10.1007/s00395-018-0677-y
Kornicka, K., Szlapka-Kosarzewska, J., Smieszek, A., and Marycz, K. (2019). 5-Azacytydine and resveratrol reverse senescence and ageing of adipose stem cells via modulation of mitochondrial dynamics and autophagy. J. Cell. Mol. Med. 23, 237–259. doi: 10.1111/jcmm.13914
Kowaltowski, A. J. (2019). Strategies to detect mitochondrial oxidants. Redox Biol. 21:101065. doi: 10.1016/j.redox.2018.101065
Krause, J., Loser, A., Lemoine, M. D., Christ, T., Scherschel, K., Meyer, C., et al. (2018). Rat atrial engineered heart tissue: a new in vitro model to study atrial biology. Basic Res. Cardiol. 113:41. doi: 10.1007/s00395-018-0701-2
Lansink, M. O., Görlinger, K., Hartmann, M., de Groot, H., and Effenberger-Neidnicht, K. (2016). Melatonin does not affect disseminated intravascular coagulation but diminishes decreases in platelet count during subacute endotoxaemia in rats. Thromb. Res. 139, 38–43. doi: 10.1016/j.thromres.2015.10.025
Lee, B. W. L., Ghode, P., and Ong, D. S. T. (2019). Redox regulation of cell state and fate. Redox Biol. 25:101056. doi: 10.1016/j.redox.2018.11.014
Lee, E., Hwang, I., Park, S., Hong, S., Hwang, B., Cho, Y., et al. (2019). MPTP-driven NLRP3 inflammasome activation in microglia plays a central role in dopaminergic neurodegeneration. Cell Death Differ. 26, 213–228. doi: 10.1038/s41418-018-0124-5
Li, J. Y., Zheng, X. Y., Ma, X. Y., Xu, X. Y., Du, Y., Lv, Q. J., et al. (2019). Melatonin protects against chromium(VI)-induced cardiac injury via activating the AMPK/Nrf2 pathway. J. Inorg. Biochem. 197:110698. doi: 10.1016/j.jinorgbio.2019.110698
Li, Q., Turdi, S., Thomas, D. P., Zhou, T., and Ren, J. (2010). Intra-myocardial delivery of mesenchymal stem cells ameliorates left ventricular and cardiomyocyte contractile dysfunction following myocardial infarction. Toxicol. Lett. 195, 119–126. doi: 10.1016/j.toxlet.2010.03.009
Li, R., Xin, T., Li, D., Wang, C., Zhu, H., and Zhou, H. (2018). Therapeutic effect of Sirtuin 3 on ameliorating nonalcoholic fatty liver disease: the role of the ERK-CREB pathway and Bnip3-mediated mitophagy. Redox Biol. 18, 229–243. doi: 10.1016/j.redox.2018.07.011
Liu, J. B., Clough, S. J., Hutchinson, A. J., Adamah-Biassi, E. B., Popovska-Gorevski, M., and Dubocovich, M. L. (2016). MT1 and MT2 melatonin receptors: a therapeutic perspective. Annu. Rev. Pharmacol. Toxicol. 56, 361–383. doi: 10.1146/annurev-pharmtox-010814-124742
Liu, L., Feng, D., Chen, G., Chen, M., Zheng, Q., Song, P., et al. (2012). Mitochondrial outer-membrane protein FUNDC1 mediates hypoxia-induced mitophagy in mammalian cells. Nat. Cell Biol. 14, 177–185. doi: 10.1038/ncb2422
Liu, Y., Xu, W., Zhai, T., You, J., and Chen, Y. (2019). Silibinin ameliorates hepatic lipid accumulation and oxidative stress in mice with non-alcoholic steatohepatitis by regulating CFLAR-JNK pathway. Acta Pharm. Sin. B 9, 745–757. doi: 10.1016/j.apsb.2019.02.006
Lochner, A., Marais, E., and Huisamen, B. (2018). Melatonin and cardioprotection against ischaemia/reperfusion injury: what’s new? A review. J. Pineal Res. 65:e12490. doi: 10.1111/jpi.12490
Lombard, D. B., and Zwaans, B. M. M. (2014). SIRT3: as simple as it seems? Gerontology 60, 56–64. doi: 10.1159/000354382
Lu, J., Li, J., Hu, Y., Guo, Z., Sun, D., Wang, P., et al. (2019). Chrysophanol protects against doxorubicin-induced cardiotoxicity by suppressing cellular PARylation. Acta Pharm. Sin. B 9, 782–793. doi: 10.1016/j.apsb.2018.10.008
Lu, M., Wang, P., Qiao, Y., Jiang, C., Ge, Y., Flickinger, B., et al. (2019). GSK3beta-mediated Keap1-independent regulation of Nrf2 antioxidant response: a molecular rheostat of acute kidney injury to chronic kidney disease transition. Redox Biol. 26:101275. doi: 10.1016/j.redox.2019.101275
Ma, S., Chen, J., Feng, J., Zhang, R., Fan, M., Han, D., et al. (2018). Melatonin ameliorates the progression of atherosclerosis via mitophagy activation and NLRP3 inflammasome inhibition. Oxid. Med. Cell. Longev. 2018:9286458. doi: 10.1155/2018/9286458
Ma, Z. Q., Yang, Y., Fan, C. X., Han, J., Wang, D. J., Di, S. Y., et al. (2016). Melatonin as a potential anticarcinogen for non-small-cell lung cancer. Oncotarget 7, 46768–46784. doi: 10.18632/oncotarget.8776
Mauriz, J. L., Collado, P. S., Veneroso, C., Reiter, R. J., and Gonzalez-Gallego, J. (2013). A review of the molecular aspects of melatonin’s anti-inflammatory actions: recent insights and new perspectives. J. Pineal Res. 54, 1–14. doi: 10.1111/j.1600-079X.2012.01014.x
Mayorov, V., Uchakin, P., Amarnath, V., Panov, A. V., Bridges, C. C., Uzhachenko, R., et al. (2019). Targeting of reactive isolevuglandins in mitochondrial dysfunction and inflammation. Redox Biol. 26:101300. doi: 10.1016/j.redox.2019.101300
McMullan, C. J., Rimm, E. B., Schernhammer, E. S., and Forman, J. P. (2017). A nested case-control study of the association between melatonin secretion and incident myocardial infarction. Heart 103, 694–701. doi: 10.1136/heartjnl-2016-310098
Mei, D., Chen, B., He, B., Liu, H., Lin, Z., Lin, J., et al. (2019). Actively priming autophagic cell death with novel transferrin receptor-targeted nanomedicine for synergistic chemotherapy against breast cancer. Acta Pharm. Sin. B 9, 1061–1077. doi: 10.1016/j.apsb.2019.03.006
Merz, J., Albrecht, P., von Garlen, S., Ahmed, I., Dimanski, D., Wolf, D., et al. (2018). Purinergic receptor Y2 (P2Y2)- dependent VCAM-1 expression promotes immune cell infiltration in metabolic syndrome. Basic Res. Cardiol. 113:45. doi: 10.1007/s00395-018-0702-1
Meyer, I. S., and Leuschner, F. (2018). The role of Wnt signaling in the healing myocardium: a focus on cell specificity. Basic Res. Cardiol. 113:44. doi: 10.1007/s00395-018-0705-y
Morell, M., Burgos, J. I., Gonano, L. A., and Vila Petroff, M. (2018). AMPK-dependent nitric oxide release provides contractile support during hyperosmotic stress. Basic Res. Cardiol. 113:7. doi: 10.1007/s00395-017-0665-7
Na, H. J., Yeum, C. E., Kim, H. S., Lee, J., Kim, J. Y., and Cho, Y. S. (2019). TSPYL5-mediated inhibition of p53 promotes human endothelial cell function. Angiogenesis 22, 281–293. doi: 10.1007/s10456-018-9656-z
Nahrendorf, M., Pittet, M. J., and Swirski, F. K. (2010). Monocytes: protagonists of infarct inflammation and repair after myocardial infarction. Circulation 121, 2437–2445. doi: 10.1161/CIRCULATIONAHA.109.916346
Nair, S. M., Rahman, R. M., Clarkson, A. N., Sutherland, B. A., Taurin, S., Sammut, I. A., et al. (2011). Melatonin treatment following stroke induction modulates L-arginine metabolism. J. Pineal Res. 51, 313–323. doi: 10.1111/j.1600-079X.2011.00891.x
Nanadikar, M. S., Vergel Leon, A. M., Borowik, S., Hillemann, A., Zieseniss, A., Belousov, V. V., et al. (2019). O2 affects mitochondrial functionality ex vivo. Redox Biol. 22:101152. doi: 10.1016/j.redox.2019.101152
NaveenKumar, S. K., Hemshekhar, M., Kemparaju, K., and Girish, K. S. (2019). Hemin-induced platelet activation and ferroptosis is mediated through ROS-driven proteasomal activity and inflammasome activation: protection by melatonin. Biochim. Biophys. Acta 1865, 2303–2316. doi: 10.1016/j.bbadis.2019.05.009
Nduhirabandi, F., du Toit, E. F., and Lochner, A. (2012). Melatonin and the metabolic syndrome: a tool for effective therapy in obesity-associated abnormalities? Acta Physiol. 205, 209–223. doi: 10.1111/j.1748-1716.2012.02410.x
Niccoli, G., Scalone, G., Lerman, A., and Crea, F. (2016). Coronary microvascular obstruction in acute myocardial infarction. Eur. Heart J. 37, 1024–1033. doi: 10.1093/eurheartj/ehv484
Nofe, J. R., Brodde, M. F., and Kehrel, B. E. (2010). High-density lipoproteins, platelets and the pathogenesis of atherosclerosis. Clin. Exp. Pharmacol. Physiol. 37, 726–735. doi: 10.1111/j.1440-1681.2010.05377.x
Nokik, S. E., Nobuyuki, M., Kazuko, T., Dongzhu, X., Taizo, K., Rujie, Q., et al. (2017). Rev-erb agonist improves adverse cardiac remodeling and survival in myocardial infarction through an anti-inflammatory mechanism. PLoS One 12:e0189330. doi: 10.1371/journal.pone.0189330
Ostjen, C. A., Rosa, C. G. S., Hartmann, R. M., Schemitt, E. G., Colares, J. R., and Marroni, N. P. (2019). Anti-inflammatory and antioxidant effect of melatonin on recovery from muscular trauma induced in rats. Exp. Mol. Pathol. 106, 52–59. doi: 10.1016/j.yexmp.2018.12.001
Pan, S., Wever-Pinzon, O., Rao, S., Levin, A., Garan, A., Takeda, K., et al. (2015). Myocardial recovery from short- and long-term cardiac support devices: results from the UNOS registry. J. Heart Lung Transplant. 34:S41.
Paradies, G., Paradies, V., Ruggiero, F. M., and Petrosillo, G. (2015). Protective role of melatonin in mitochondrial dysfunction and related disorders. Arch. Toxicol. 89, 923–939. doi: 10.1007/s00204-015-1475-z
Pei, H. F., Du, J., Song, X. F., He, L., Zhang, Y. F., Li, X. C., et al. (2016). Melatonin prevents adverse myocardial infarction remodeling via Notch1/Mfn2 pathway. Free Radic. Biol. Med. 97, 408–417. doi: 10.1016/j.freeradbiomed.2016.06.015
Perez-Gonzalez, A., Castaneda-Arriaga, R., Alvarez-Idaboy, J. R., Reiter, R. J., and Galano, A. (2019). Melatonin and its metabolites as chemical agents capable of directly repairing oxidized DNA. J. Pineal Res. 66:e12539. doi: 10.1111/jpi.12539
Phan, T. K., Williams, S. A., Bindra, G. K., Lay, F. T., Poon, I. K. H., and Hulett, M. D. (2019). Phosphoinositides: multipurpose cellular lipids with emerging roles in cell death. Cell Death Differ. 26, 781–793. doi: 10.1038/s41418-018-0269-2
Reddy, K. R. K., Dasari, C., Duscharla, D., Supriya, B., Ram, N. S., Surekha, M. V., et al. (2018). Dimethylarginine dimethylaminohydrolase-1 (DDAH1) is frequently upregulated in prostate cancer, and its overexpression conveys tumor growth and angiogenesis by metabolizing asymmetric dimethylarginine (ADMA). Angiogenesis 21, 79–94. doi: 10.1007/s10456-017-9587-0
Reina, M., and Martínez, A. (2018). A new free radical scavenging cascade involving melatonin and three of its metabolites (3OHM, AFMK and AMK). Comput. Theor. Chem. 1123, 111–118. doi: 10.1016/j.comptc.2017.11.017
Reiter, R. J., Rosales-Corral, S., Tan, D. X., Jou, M. J., Galano, A., and Xu, B. (2017). Melatonin as a mitochondria-targeted antioxidant: one of evolution’s best ideas. Cell. Mol. Life Sci. 74, 3863–3881. doi: 10.1007/s00018-017-2609-7
Ren, B. (2016). Protein kinase D1 signaling in angiogenic gene expression and VEGF-mediated angiogenesis. Front. Cell Dev. Biol. 4:37. doi: 10.3389/fcell.2016.00037
Rezzani, R., Favero, G., Stacchiotti, A., and Rodella, L. F. (2013). Endothelial and vascular smooth muscle cell dysfunction mediated by cyclophylin A and the atheroprotective effects of melatonin. Life Sci. 92, 875–882. doi: 10.1016/j.lfs.2012.11.011
Riehle, C., and Bauersachs, J. (2018). Of mice and men: models and mechanisms of diabetic cardiomyopathy. Basic Res. Cardiol. 114:2. doi: 10.1007/s00395-018-0711-0
Sallinen, P., Manttari, S., Leskinen, H., Ilves, M., Vakkuri, O., Ruskoaho, H., et al. (2007). The effect of myocardial infarction on the synthesis, concentration and receptor expression of endogenous melatonin. J. Pineal Res. 42, 254–260. doi: 10.1111/j.1600-079X.2006.00413.x
Schreiber, T., Salhofer, L., Quinting, T., and Fandrey, J. (2019). Things get broken: the hypoxia-inducible factor prolyl hydroxylases in ischemic heart disease. Basic Res. Cardiol. 114:16. doi: 10.1007/s00395-019-0725-2
Seidlmayer, L. K., Juettner, V. V., Kettlewell, S., Pavlov, E. V., Blatter, L. A., and Dedkova, E. N. (2015). Distinct mPTP activation mechanisms in ischaemia-reperfusion: contributions of Ca2+, ROS, pH, and inorganic polyphosphate. Cardiovasc. Res. 106, 237–248. doi: 10.1093/cvr/cvv097
Serocki, M., Bartoszewska, S., Janaszak-Jasiecka, A., Ochocka, R. J., Collawn, J. F., and Bartoszewski, R. (2018). miRNAs regulate the HIF switch during hypoxia: a novel therapeutic target. Angiogenesis 21, 183–202. doi: 10.1007/s10456-018-9600-2
Shafiei, E., Bahtoei, M., Raj, P., Ostovar, A., Iranpour, D., Akbarzadeh, S., et al. (2018). Effects of N-acetyl cysteine and melatonin on early reperfusion injury in patients undergoing coronary artery bypass grafting: a randomized, open-labeled, placebo-controlled trial. Medicine 97:e11383. doi: 10.1097/MD.0000000000011383
Shanmugam, G., Challa, A. K., Litovsky, S. H., Devarajan, A., Wang, D., Jones, D. P., et al. (2019). Enhanced Keap1-Nrf2 signaling protects the myocardium from isoproterenol-induced pathological remodeling in mice. Redox Biol. 27:101212. doi: 10.1016/j.redox.2019.101212
Siddall, H. K., Yellon, D. M., Ong, S. B., Mukherjee, U. A., Burke, N., Hall, A. R., et al. (2013). Loss of PINK1 increases the heart’s vulnerability to ischemia-reperfusion injury. PLoS One 8:e62400. doi: 10.1371/journal.pone.0062400
Simko, F., Baka, T., Paulis, L., and Reiter, R. J. (2016). Elevated heart rate and nondipping heart rate as potential targets for melatonin: a review. J. Pineal Res. 61, 127–137. doi: 10.1111/jpi.12348
Simula, L., Campanella, M., and Campello, S. (2019). Targeting Drp1 and mitochondrial fission for therapeutic immune modulation. Pharmacol. Res. 146:104317. doi: 10.1016/j.phrs.2019.104317
Singhanat, K., Apaijai, N., Chattipakorn, S. C., and Chattipakorn, N. (2018). Roles of melatonin and its receptors in cardiac ischemia-reperfusion injury. Cell. Mol. Life Sci. 75, 4125–4149. doi: 10.1007/s00018-018-2905-x
Sircana, A., De Michieli, F., Parente, R., Framarin, L., Leone, N., Berrutti, M., et al. (2019). Gut microbiota, hypertension and chronic kidney disease: recent advances. Pharmacol. Res. 144, 390–408. doi: 10.1016/j.phrs.2018.01.013
Song, C., Zhao, J., Fu, B., Li, D., Mao, T., Peng, W., et al. (2017). Melatonin-mediated upregulation of Sirt3 attenuates sodium fluoride-induced hepatotoxicity by activating the MT1-PI3K/AKT-PGC-1alpha signaling pathway. Free Radic. Biol. Med. 112, 616–630. doi: 10.1016/j.freeradbiomed.2017.09.005
Soto-Heras, S., Catala, M. G., Roura, M., Menendez-Blanco, I., Piras, A. R., Izquierdo, D., et al. (2019). Effects of melatonin on oocyte developmental competence and the role of melatonin receptor 1 in juvenile goats. Reprod. Domest. Anim. 54, 381–390. doi: 10.1111/rda.13378
Tahrir, F. G., Langford, D., Amini, S., Mohseni Ahooyi, T., and Khalili, K. (2019). Mitochondrial quality control in cardiac cells: mechanisms and role in cardiac cell injury and disease. J. Cell. Physiol. 234, 8122–8133. doi: 10.1002/jcp.27597
Tang, V., Fu, S., Rayner, B. S., and Hawkins, C. L. (2019). 8-Chloroadenosine induces apoptosis in human coronary artery endothelial cells through the activation of the unfolded protein response. Redox Biol. 26:101274. doi: 10.1016/j.redox.2019.101274
Teixeira, J., Basit, F., Swarts, H. G., Forkink, M., Oliveira, P. J., Willems, P., et al. (2018). Extracellular acidification induces ROS- and mPTP-mediated death in HEK293 cells. Redox Biol. 15, 394–404. doi: 10.1016/j.redox.2017.12.018
Tengattini, S., Reiter, R. J., Tan, D. X., Terron, M. P., Rodella, L. F., and Rezzani, R. (2008). Cardiovascular diseases: protective effects of melatonin. J. Pineal Res. 44, 16–25. doi: 10.1111/j.1600-079X.2007.00518.x
Ter Horst, E. N., Krijnen, P. A. J., Hakimzadeh, N., Robbers, L., Hirsch, A., Nijveldt, R., et al. (2018). Elevated monocyte-specific type I interferon signalling correlates positively with cardiac healing in myocardial infarct patients but interferon alpha application deteriorates myocardial healing in rats. Basic Res. Cardiol. 114:1. doi: 10.1007/s00395-018-0709-7
Thieltges, K. M., Avramovic, D., Piscitelli, C. L., Markovic-Mueller, S., Binz, H. K., and Ballmer-Hofer, K. (2018). Characterization of a drug-targetable allosteric site regulating vascular endothelial growth factor signaling. Angiogenesis 21, 533–543. doi: 10.1007/s10456-018-9606-9
Tian, L., Lu, L., Feng, J., and Melancon, M. P. (2018). Radiopaque nano and polymeric materials for atherosclerosis imaging, embolization and other catheterization procedures. Acta Pharm. Sin. B 8, 360–370. doi: 10.1016/j.apsb.2018.03.002
Trindade, F., Vitorino, R., Leite-Moreira, A., and Falcao-Pires, I. (2019). Pericardial fluid: an underrated molecular library of heart conditions and a potential vehicle for cardiac therapy. Basic Res. Cardiol. 114:10. doi: 10.1007/s00395-019-0716-3
Venegas, C., Garcia, J. A., Escames, G., Ortiz, F., Lopez, A., Doerrier, C., et al. (2012). Extrapineal melatonin: analysis of its subcellular distribution and daily fluctuations. J. Pineal Res. 52, 217–227. doi: 10.1111/j.1600-079X.2011.00931.x
Venugopal, S., Kao, C., Chandna, R., Sulochana, K. N., Subramanian, V., Chen, M., et al. (2018). Angio-3, a 10-residue peptide derived from human plasminogen kringle 3, suppresses tumor growth in mice via impeding both angiogenesis and vascular permeability. Angiogenesis 21, 653–665. doi: 10.1007/s10456-018-9616-7
Villalobos, C., Gutierrez, L. G., Hernandez-Morales, M., Del Bosque, D., and Nunez, L. (2018). Mitochondrial control of store-operated Ca(2+) channels in cancer: pharmacological implications. Pharmacol. Res. 135, 136–143. doi: 10.1016/j.phrs.2018.08.001
Wang, B., Nie, J. L., Wu, L. J., Hu, Y. Y., Wen, Z., Dong, L. L., et al. (2018). AMPKα2 protects against the development of heart failure by enhancing mitophagy via PINK1 phosphorylation. Circ. Res. 122, 712–729. doi: 10.1161/circresaha.117.312317
Wang, J., Zhu, P., Li, R., Ren, J., and Zhou, H. (2020). Fundc1-dependent mitophagy is obligatory to ischemic preconditioning-conferred renoprotection in ischemic AKI via suppression of Drp1-mediated mitochondrial fission. Redox Biol. 30:101415. doi: 10.1016/j.redox.2019.101415
Wang, S., Dong, G., and Sheng, C. (2019). Structural simplification: an efficient strategy in lead optimization. Acta Pharm. Sin. B 9, 880–901. doi: 10.1016/j.apsb.2019.05.004
Wang, X. N., Shu, H., Wang, M. W., Sun, Y. Y., Liu, C. F., Liu, W. L., et al. (2016). Melatonin alleviates lipopolysaccharide-compromised integrity of blood brain barrier by activation of AMP-activated protein kinase in old mice. Chin. J. Pharmacol. Toxicol. 30, 1021–1022. doi: 10.1111/acel.12572
Wang, Y., Lei, T., Yuan, J., Wu, Y., Shen, X., Gao, J., et al. (2018). GCN2 deficiency ameliorates doxorubicin-induced cardiotoxicity by decreasing cardiomyocyte apoptosis and myocardial oxidative stress. Redox Biol. 17, 25–34. doi: 10.1016/j.redox.2018.04.009
Wang, Z., Xu, G., Gao, Y., Zhan, X., Qin, N., Fu, S., et al. (2019). Cardamonin from a medicinal herb protects against LPS-induced septic shock by suppressing NLRP3 inflammasome. Acta Pharm. Sin. B 9, 734–744. doi: 10.1016/j.apsb.2019.02.003
Westman, P. C., Lipinski, M. J., Luger, D., Waksman, R., Bonow, R. O., Wu, E., et al. (2016). Inflammation as a driver of adverse left ventricular remodeling after acute myocardial infarction. J. Am. Coll. Cardiol. 67, 2050–2060. doi: 10.1016/j.jacc.2016.01.073
Wu, C., Xi, C., Tong, J., Zhao, J., Jiang, H., Wang, J., et al. (2019). Design, synthesis, and biological evaluation of novel tetrahydroprotoberberine derivatives (THPBs) as proprotein convertase subtilisin/kexin type 9 (PCSK9) modulators for the treatment of hyperlipidemia. Acta Pharm. Sin. B 9, 1216–1230. doi: 10.1016/j.apsb.2019.06.006
Wu, D., Hu, Q., Tan, B., Rose, P., Zhu, D., and Zhu, Y. Z. (2018a). Amelioration of mitochondrial dysfunction in heart failure through S-sulfhydration of Ca(2+)/calmodulin-dependent protein kinase II. Redox Biol. 19, 250–262. doi: 10.1016/j.redox.2018.08.008
Wu, D., Wang, H., Teng, T., Duan, S., Ji, A., and Li, Y. (2018b). Hydrogen sulfide and autophagy: a double edged sword. Pharmacol. Res. 131, 120–127. doi: 10.1016/j.phrs.2018.03.002
Xiao, Y., Karam, C., Yi, J., Zhang, L., Li, X., Yoon, D., et al. (2018). ROS-related mitochondrial dysfunction in skeletal muscle of an ALS mouse model during the disease progression. Pharmacol. Res. 138, 25–36. doi: 10.1016/j.phrs.2018.09.008
Xiong, Q., Wang, Z., Yu, Y., Wen, Y., Suguro, R., Mao, Y., et al. (2019). Hydrogen sulfide stabilizes atherosclerotic plaques in apolipoprotein E knockout mice. Pharmacol. Res. 144, 90–98. doi: 10.1016/j.phrs.2019.04.006
Xu, N., Wang, Q., Jiang, S., Wang, Q., Hu, W., Zhou, S., et al. (2019). Fenofibrate improves vascular endothelial function and contractility in diabetic mice. Redox Biol. 20, 87–97. doi: 10.1016/j.redox.2018.09.024
Xu, S. F., Zhang, Y. H., Wang, S., Pang, Z. Q., Fan, Y. G., Li, J. Y., et al. (2019). Lactoferrin ameliorates dopaminergic neurodegeneration and motor deficits in MPTP-treated mice. Redox Biol. 21:101090. doi: 10.1016/j.redox.2018.101090
Xu, T., Ding, W., Ao, X., Chu, X., Wan, Q., Wang, Y., et al. (2019). ARC regulates programmed necrosis and myocardial ischemia/reperfusion injury through the inhibition of mPTP opening. Redox Biol. 20, 414–426. doi: 10.1016/j.redox.2018.10.023
Xu, Z., Fu, T., Guo, Q., Sun, W., and Gan, Z. (2019). Mitochondrial quality orchestrates muscle-adipose dialog to alleviate dietary obesity. Pharmacol. Res. 141, 176–180. doi: 10.1016/j.phrs.2018.12.020
Yan, G. G., Yu, L. Z., Jiang, S. M., and Zhu, J. F. (2018). Melatonin antagonizes oxidative stress-induced mitochondrial dysfunction in retinal pigmented epithelium cells via melatonin receptor 1 (MT1). J. Toxicol. Sci. 43, 659–669. doi: 10.2131/jts.43.659
Yang, C. H., Xu, J. H., Ren, Q. C., Duan, T., Mo, F., and Zhang, W. (2019). Melatonin promotes secondary hair follicle development of early postnatal cashmere goat and improves cashmere quantity and quality by enhancing antioxidant capacity and suppressing apoptosis. J. Pineal Res. 67:e12569. doi: 10.1111/jpi.12569
Yang, M., Linn, B. S., Zhang, Y., and Ren, J. (2019). Mitophagy and mitochondrial integrity in cardiac ischemia-reperfusion injury. Biochim. Biophys. Acta 1865, 2293–2302. doi: 10.1016/j.bbadis.2019.05.007
Yang, M. Y., Yu, Q. L., Huang, Y. S., and Yang, G. (2019). Neuroprotective effects of andrographolide derivative CX-10 in transient focal ischemia in rat: involvement of Nrf2/AE and TLR/NF-kappaB signaling. Pharmacol. Res. 144, 227–234. doi: 10.1016/j.phrs.2019.04.023
Yang, Q., He, G. W., Underwood, M. J., and Yu, C. M. (2016). Cellular and molecular mechanisms of endothelial ischemia/reperfusion injury: perspectives and implications for postischemic myocardial protection. Am. J. Transl. Res. 8, 765–777.
Yang, X., Wang, H., Ni, H.-M., Xiong, A., Wang, Z., Sesaki, H., et al. (2017). Inhibition of Drp1 protects against senecionine-induced mitochondria-mediated apoptosis in primary hepatocytes and in mice. Redox Biol. 12, 264–273. doi: 10.1016/j.redox.2017.02.020
Yang, Y., Duan, W., Jin, Z., Yi, W., Yan, J., Zhang, S., et al. (2013). JAK2/STAT3 activation by melatonin attenuates the mitochondrial oxidative damage induced by myocardial ischemia/reperfusion injury. J. Pineal Res. 55, 275–286. doi: 10.1111/jpi.12070
Yang, Y., Sun, Y., Yi, W., Li, Y., Fan, C. X., Xin, Z. L., et al. (2014). A review of melatonin as a suitable antioxidant against myocardial ischemia–reperfusion injury and clinical heart diseases. J. Pineal Res. 57, 357–366. doi: 10.1111/jpi.12175
Yao, X., Zhang, J., Jing, X., Ye, Y., Guo, J., Sun, K., et al. (2019). Fibroblast growth factor 18 exerts anti-osteoarthritic effects through PI3K-AKT signaling and mitochondrial fusion and fission. Pharmacol. Res. 139, 314–324. doi: 10.1016/j.phrs.2018.09.026
Yarana, C., Thompson, H., Chaiswing, L., Butterfield, D. A., Weiss, H., Bondada, S., et al. (2019). Extracellular vesicle-mediated macrophage activation: an insight into the mechanism of thioredoxin-mediated immune activation. Redox Biol. 26:101237. doi: 10.1016/j.redox.2019.101237
Yoon, Y. M., Kim, H. J., Lee, J. H., and Lee, S. H. (2019). Melatonin enhances mitophagy by upregulating expression of heat shock 70 kDa protein 1L in human mesenchymal stem cells under oxidative stress. Int. J. Mol. Sci. 20:4545. doi: 10.3390/ijms20184545
Yu, L. M., Di, W. C., Dong, X., Li, Z., Zhang, Y., Xue, X. D., et al. (2018). Melatonin protects diabetic heart against ischemia-reperfusion injury, role of membrane receptor-dependent cGMP-PKG activation. Biochim. Biophys. Acta 1864, 563–578. doi: 10.1016/j.bbadis.2017.11.023
Yu, L. M., Li, B. Y., Zhang, M., Jin, Z. X., Duan, W. X., Zhao, G. L., et al. (2016). Melatonin reduces PERK-eIF2α-ATF4-mediated endoplasmic reticulum stress during myocardial ischemia-reperfusion injury: role of RISK and SAFE pathways interaction. Apoptosis 21, 809–824. doi: 10.1007/s10495-016-1246-1
Yuan, T., Yang, T., Chen, H., Fu, D., Hu, Y., Wang, J., et al. (2019). New insights into oxidative stress and inflammation during diabetes mellitus-accelerated atherosclerosis. Redox Biol. 20, 247–260. doi: 10.1016/j.redox.2018.09.025
Yuqi, L., Lei, G., Yanqiu, S., Qiao, X., Yu, W., and Yundai, C. (2015). Efficacy and safety of limus-eluting versus paclitaxel-eluting coronary artery stents in patients with diabetes mellitus: a meta-analysis. Int. J. Cardiol. 64, 680–691. doi: 10.1016/j.ijcard.2015.02.002
Zaouali, M. A., Reiter, R. J., Padrissa-Altes, S., Boncompagni, E., Garcia, J. J., Ben Abnennebi, H., et al. (2011). Melatonin protects steatotic and nonsteatotic liver grafts against cold ischemia and reperfusion injury. J. Pineal Res. 50, 213–221. doi: 10.1111/j.1600-079X.2010.00831.x
Zhang, H., Jin, B., and Faber, J. E. (2019). Mouse models of Alzheimer’s disease cause rarefaction of pial collaterals and increased severity of ischemic stroke. Angiogenesis 22, 263–279. doi: 10.1007/s10456-018-9655-0
Zhang, H. M., and Zhang, Y. (2014). Melatonin: a well-documented antioxidant with conditional pro-oxidant actions. J. Pineal Res. 57, 131–146. doi: 10.1111/jpi.12162
Zhang, Y., Mi, S. L., Hu, N., Doser, T. A., Sun, A., Ge, J., et al. (2014). Mitochondrial aldehyde dehydrogenase 2 accentuates aging-induced cardiac remodeling and contractile dysfunction: role of AMPK, Sirt1, and mitochondrial function. Free Radic. Biol. Med. 71, 208–220. doi: 10.1016/j.freeradbiomed.2014.03.018
Zhang, Y., Wang, Y., Xu, J. N., Tian, F., Hu, S. Y., Chen, Y. D., et al. (2019). Melatonin attenuates myocardial ischemia-reperfusion injury via improving mitochondrial fusion/mitophagy and activating the AMPK-OPA1 signaling pathways. J. Pineal Res. 66:e12542. doi: 10.1111/jpi.12542
Zhao, J., Gao, J. L., Zhu, J. X., Zhu, H. B., Peng, X., Jiang, M., et al. (2019). The different response of cardiomyocytes and cardiac fibroblasts to mitochondria inhibition and the underlying role of STAT3. Basic Res. Cardiol. 114:12. doi: 10.1007/s00395-019-0721-6
Zhao, Y., Wang, H. Y., Chen, W., Chen, L. F., Liu, D. M., Wang, X., et al. (2019). Melatonin attenuates white matter damage after focal brain ischemia in rats by regulating the TLR4/NF-kappaB pathway. Brain Res. Bull. 150, 168–178. doi: 10.1016/j.brainresbull.2019.05.019
Zhong, J., Tan, Y., Lu, J., Liu, J., Xiao, X., Zhu, P., et al. (2019). Therapeutic contribution of melatonin to the treatment of septic cardiomyopathy: a novel mechanism linking Ripk3-modified mitochondrial performance and endoplasmic reticulum function. Redox Biol. 26:101287. doi: 10.1016/j.redox.2019.101287
Zhou, H., Hu, S., Jin, Q., Shi, C., Zhang, Y., Zhu, P., et al. (2017a). Mff-dependent mitochondrial fission contributes to the pathogenesis of cardiac microvasculature ischemia/reperfusion injury via induction of mROS-mediated cardiolipin oxidation and HK2/VDAC1 disassociation-involved mPTP opening. J. Am. Heart Assoc. 6:e005328. doi: 10.1161/JAHA.116.005328
Zhou, H., Li, D., Zhu, P., Hu, S., Hu, N., Ma, S., et al. (2017b). Melatonin suppresses platelet activation and function against cardiac ischemia/reperfusion injury via PPARgamma/FUNDC1/mitophagy pathways. J. Pineal Res. 63:e12438. doi: 10.1111/jpi.12438
Zhou, H., Zhang, Y., Hu, S., Shi, C., Zhu, P., Ma, Q., et al. (2017c). Melatonin protects cardiac microvasculature against ischemia/reperfusion injury via suppression of mitochondrial fission-VDAC1-HK2-mPTP-mitophagy axis. J. Pineal Res. 63:e12413. doi: 10.1111/jpi.12413
Zhou, H., Zhang, Y., Hu, S. Y., Shi, C., Zhu, P. J., Ma, Q., et al. (2017d). Melatonin protects cardiac microvasculature against ischemia/reperfusion injury via suppression of mitochondrial fission-VDAC1-HK2-mPTP-mitophagy axis. J. Pineal Res. 63;e12413. doi: 10.1111/jpi.12413
Zhou, H., Zhu, P., Guo, J., Hu, N., Wang, S., Li, D., et al. (2017e). Ripk3 induces mitochondrial apoptosis via inhibition of FUNDC1 mitophagy in cardiac IR injury. Redox Biol. 13, 498–507. doi: 10.1016/j.redox.2017.07.007
Zhou, H., Li, D., Zhu, P., Ma, Q., Toan, S., Wang, J., et al. (2018a). Inhibitory effect of melatonin on necroptosis via repressing the Ripk3-PGAM5-CypD-mPTP pathway attenuates cardiac microvascular ischemia-reperfusion injury. J. Pineal Res. 65:e12503. doi: 10.1111/jpi.12503
Zhou, H., Li, N., Yuan, Y., Jin, Y. G., Guo, H., Deng, W., et al. (2018b). Activating transcription factor 3 in cardiovascular diseases: a potential therapeutic target. Basic Res. Cardiol. 113:37.
Zhou, H., Ma, Q., Zhu, P., Ren, J., Reiter, R. J., and Chen, Y. (2018c). Protective role of melatonin in cardiac ischemia-reperfusion injury: from pathogenesis to targeted therapy. J. Pineal Res. 64:e12471. doi: 10.1111/jpi.12471
Zhou, H., Shi, C., Hu, S., Zhu, H., Ren, J., and Chen, Y. (2018d). BI1 is associated with microvascular protection in cardiac ischemia reperfusion injury via repressing Syk-Nox2-Drp1-mitochondrial fission pathways. Angiogenesis 21, 599–615. doi: 10.1007/s10456-018-9611-z
Zhou, H., Wang, J., Zhu, P., Zhu, H., Toan, S., Hu, S., et al. (2018e). NR4A1 aggravates the cardiac microvascular ischemia reperfusion injury through suppressing FUNDC1-mediated mitophagy and promoting Mff-required mitochondrial fission by CK2alpha. Basic Res. Cardiol. 113:23. doi: 10.1007/s00395-018-0682-1
Zhou, H., Wang, S., Hu, S., Chen, Y., and Ren, J. (2018f). ER-mitochondria microdomains in cardiac ischemia-reperfusion injury: a fresh perspective. Front. Physiol. 9:755. doi: 10.3389/fphys.2018.00755
Zhou, H., Wang, S., Zhu, P., Hu, S., Chen, Y., and Ren, J. (2018g). Empagliflozin rescues diabetic myocardial microvascular injury via AMPK-mediated inhibition of mitochondrial fission. Redox Biol. 15, 335–346. doi: 10.1016/j.redox.2017.12.019
Zhou, H., Zhu, P., Wang, J., Zhu, H., Ren, J., and Chen, Y. (2018h). Pathogenesis of cardiac ischemia reperfusion injury is associated with CK2alpha-disturbed mitochondrial homeostasis via suppression of FUNDC1-related mitophagy. Cell Death Differ. 25, 1080–1093. doi: 10.1038/s41418-018-0086-7
Zhou, H., and Toan, S. (2020). Pathological roles of mitochondrial oxidative stress and mitochondrial dynamics in cardiac microvascular ischemia/reperfusion injury. Biomolecules 10:85. doi: 10.3390/biom10010085
Zhou, H., Toan, S., Zhu, P., Wang, J., Ren, J., and Zhang, Y. (2020). DNA-PKcs promotes cardiac ischemia reperfusion injury through mitigating BI-1-governed mitochondrial homeostasis. Basic Res. Cardiol. 115:11. doi: 10.1007/s00395-019-0773-7
Zhou, H., Wang, J., Hu, S., Zhu, H., Toanc, S., and Ren, J. (2019). BI1 alleviates cardiac microvascular ischemia-reperfusion injury via modifying mitochondrial fission and inhibiting XO/ROS/F-actin pathways. J. Cell Physiol. 234, 5056–5069. doi: 10.1002/jcp.27308
Zhu, H., Jin, Q., Li, Y., Ma, Q., Wang, J., Li, D., et al. (2018). Melatonin protected cardiac microvascular endothelial cells against oxidative stress injury via suppression of IP3R-[Ca(2+)]c/VDAC-[Ca(2+)]m axis by activation of MAPK/ERK signaling pathway. Cell Stress Chaperones 23, 101–113. doi: 10.1007/s12192-017-0827-4
Zhu, P., Hu, S., Jin, Q., Li, D., Tian, F., Toan, S., et al. (2018). Ripk3 promotes ER stress-induced necroptosis in cardiac IR injury: a mechanism involving calcium overload/XO/ROS/mPTP pathway. Redox Biol. 16, 157–168. doi: 10.1016/j.redox.2018.02.019
Zollbrecht, C., Persson, A. E. G., Lundberg, J. O., Weitzberg, E., and Carlström, M. (2016). Nitrite-mediated reduction of macrophage NADPH oxidase activity is dependent on xanthine oxidoreductase-derived nitric oxide but independent of S-nitrosation. Redox Biol. 10, 119–127. doi: 10.1016/j.redox.2016.09.015
Keywords: melatonin, cardioprotective, cardiomyocyte, myocardial infarction, mitochondrion
Citation: Fu Z, Jiao Y, Wang J, Zhang Y, Shen M, Reiter RJ, Xi Q and Chen Y (2020) Cardioprotective Role of Melatonin in Acute Myocardial Infarction. Front. Physiol. 11:366. doi: 10.3389/fphys.2020.00366
Received: 07 February 2020; Accepted: 30 March 2020;
Published: 29 April 2020.
Edited by:
Jun Ren, University of Washington, United StatesReviewed by:
Ying Tan, Southern Medical University, ChinaAna Beatriz Rodríguez, University of Extremadura, Spain
Jian Yang, Fourth Military Medical University, China
Copyright © 2020 Fu, Jiao, Wang, Zhang, Shen, Reiter, Xi and Chen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zhenhong Fu, ZnV6aGVuaEAxMjYuY29t; Qing Xi, eGlxaW5nMzAxQDE2My5jb20=; Yundai Chen, eXVuZGFpY0AxNjMuY29t
†These authors have contributed equally to this work