 Li Su
Li Su Jian-Yu Meng
Jian-Yu Meng Hong Yang1
Hong Yang1- 1Provincial Key Laboratory for Agricultural Pest Management of Mountainous Regions, Institute of Entomology, Guizhou University, Guiyang, China
- 2Guizhou Tobacco Science Research Institute, Guiyang, China
Ostrinia furnacalis, an important pest of corn, has substantial detrimental effects on corn production. The mitogen-activated protein kinase (MAPK) signaling pathway plays a pivotal role in an insect’s resistance to environmental stress. The expression levels of JNK and p38 have been well recorded in several insects under different environmental stressors, at different developmental stages, and in various tissue types; however, there is limited information on JNK and p38 in agricultural insects. To clarify the mechanism whereby O. furnacalis responds to environmental stress, we cloned JNK and p38 from O. furnacalis and subsequently named them OfJNK and Ofp38, respectively. Further, we examined the expression levels of OfJNK and Ofp38 under different environmental stressors. In this study, we obtained full-length sequences of OfJNK and Ofp38, and RT-qPCR results showed that these genes were expressed at all developmental stages, in various tissues (head, chest, abdomen, leg, wing, antennae, compound eye, midgut, and ovary) and under different environmental stressors (4°C and ultraviolet A treatment for 0, 30, 60, 90, and 120 min). The expression levels of OfJNK and Ofp38 were relatively higher in eggs and 3-day-old adult females than in other developmental stages. Moreover, the expression level of OfJNK was higher in the wings than in other tissues, whereas that of Ofp38 was significantly higher in the ovaries than in other tissues. OfJNK and Ofp38 showed high expression 90 min after being subjected to treatment at 4°C and ultraviolet A irradiation; the expression of Ofp38 peaked at 30 min, whereas that of OfJNK peaked at 60 min. These results indicate that O. furnacalis differs in terms of its response under different environmental stressors. In summary, our results will provide a foundation for additional research needed to determine the role of the MAPK signaling pathway and the underlying mechanisms by which it shows resistance to environmental stresses in insects.
Introduction
Ostrinia furnacalis (Guenée) (Lepidoptera: Pyralidae) is an important global agricultural pest that has caused substantial economic losses in corn, sorghum, cotton, and millet production (Afidchao et al., 2013). Its first and second instar larvae easily damage the whorl leaves of corn and then attack its stalk, ears, and cobs. In addition, the larvae feed on silk and grains, leading to ear rot, increased mycotoxin contamination, and declined corn quality (He et al., 2003). It is estimated that O. furnacalis can cause yield losses of 10–30% and might lead to absence of harvest in an outbreak year (Wang et al., 2000). The global harm to crops by O. furnacalis is related to its high underlying capacity to survive under various environmental stresses (Zhang et al., 2013; Liu et al., 2014). Several studies have demonstrated that O. furnacalis exhibits physiological adaptations that are induced by low temperature and ultraviolet (UV) stress. For instance, Feng et al. (2007) reported that low temperature treatment could increase the cold tolerance in O. furnacalis. Shang et al. (2015) demonstrated that the accumulation of glutathione transferase in larvae is most probably involved in a low temperature stress response. Recently, a comparative proteomic study has demonstrated that the response patterns of O. furnacalis to UV-A stress are complex because the differentially expressed proteins were involved in diverse biological processes including signal transduction, transport processing, cellular stress response, metabolism, and cytoskeleton organization (Zhang and Meng, 2018). Moreover, our previous study found that UV-A irradiation can enhance the fecundity of O. furnacalis and prolong the development of F1 generation (Liu et al., in press). However, the molecular mechanisms via which O. furnacalis exhibits the remarkable potential to adapt to these stresses remain unclear.
Insects adapt to fluctuant and unfavorable environments (Grubor-Lajsic et al., 1992; Bale and Hayward, 2010). Low temperature stress is a common environmental stimulus for insects (Fujita, 1999). A previous study has shown that the mitogen-activated protein kinase (MAPK) is cold responsive and that in Microdera punctipennis exposed to low temperatures, MAPK may play a role in the signal transduction (Meng et al., 2015). Hu et al. (2017) reported that in O. furnacalis, the relative expressions of Hsc70 and Hsp90 were induced upon exposure to low temperature. The UV-A have been widely used as light sources in light traps for the management of agriculture and forest insect pests which exhibit phototaxis. However, increasing evidence indicates that UV-A irradiation is an important stress factor that impacts all living creatures including insects (Meyer-Rochow et al., 2002; Meng et al., 2009). Ali et al. (2017) found that UV-A could induce oxidative stress and alter antioxidant enzyme activity, thereby affecting the normal physiological functions of Mythimna separata adults. Meng et al. (2010) reported that MAPK was upregulated following UV light irradiation in Helicoverpa armigera. Insects can apparently perceive signals using sensors and transmit them to the cellular machinery via signal transduction to regulate gene expression. The MAPK signaling pathway is one of the major signal transduction pathways (Meng et al., 2010).
MAPK is a serine/threonine protein that is implicated in the regulation of multiple cellular activities, such as differentiation, proliferation, death, and adaptive and immune responses (Zhang and Liu, 2002; Huang et al., 2009). Multicellular organisms have two well-characterized subfamilies of MAPKs, c-Jun N-terminal kinase (JNK) and p38 (Johnson and Lapadat, 2002), each of which plays a vital role in the physiological processes of cells, including growth and development, apoptosis, and responses to external environmental stresses (Cowan and Storey, 2003). JNK and p38 are pivotal for the stress-related signaling pathways that transport signals from the cell epidermis into the nucleus to initiate gene expression (Rahaus et al., 2004). JNK is mainly implicated in the transcriptional regulation of cells, and the activation of p38 is generally believed to be a response of cells to inflammatory cytokines and environmental stresses [e.g. UV irradiation, temperature, ischemia, and hypoxia] (Ono and Han, 2000; Hohmann, 2002). Several studies have suggested that JNK and p38 participate in cellular responses to environmental stresses (Xia et al., 1995; Ludwig et al., 2003; Vogelweith et al., 2011).
Several important roles of the MAPK signaling pathway have been reported in insects. For example, Fujiwara and Denlinger (2007) showed that p38 might be part of the signaling pathway that causes rapid frozen stiffening in the flesh fly Sarcophaga crassipalpis. Furthermore, their studies on Bombyx mori revealed an underlying function of MAPKs of initiating and terminating embryonic diapause (Fujiwara et al., 2006). Moreover, it has recently been demonstrated that the JNK and p38 MAPK signaling pathways play significant roles during the responses of Bemisia tabaci to low temperature stresses (Li et al., 2012; Wang et al., 2013). Nevertheless, until now, information about the function of MAPKs in Lepidoptera under various environmental stresses is lacking. The aim of this study was to investigate whether the MAPK signaling pathway is involved in responses of O. furnacalis to different environmental stresses. To determine this, we cloned the JNK and p38 of O. furnacalis (subsequently named OfJNK and Ofp38, respectively), examined their expression levels at various developmental stages and in different tissues, and investigated the expression levels of OfJNK and Ofp38 under low temperature and UVA irradiation. Our results could contribute to identifying the molecular mechanism of MAPKs in O. furnacalis under various environmental stresses and to designing cultural management strategies that could be used to control this pest.
Materials and Methods
Insect Rearing
O. furnacalis were reared in climate-controlled cabinets under a 14:10-h light–dark photoperiod at 26 ± 1°C and 70–80% relative humidity. The larvae were fed an artificial diet as previously described (Feng et al., 2011). Three-day-old adult insects were fed a 10% honey–water solution that was kept in a 100-mL transparent plastic container.
Insect Treatment and Sample Collection
The experimental insect treatments are as follows: (1) low temperature treatment: female adults aged 3 days old were exposed to a temperature of 4°C for 0 (control), 30, 60, 90, and 120 min, and ten female adults were collected per biological replicate. (2) UVA treatment: female adults aged 3 days old were irradiated with 315–400 nm UVA light (NanJing HuaQiang Electronic Engineering Co., Ltd., Nanjing, China) at a frequency of ∼300 μW/cm1. After adapting to a 2-h scotophase period at 26 ± 1°C, female adults aged 3 days old were exposed to UVA for 0 (control), 30, 60, 90, and 120 min, and ten female adults were collected per biological replicate. (3) Samples were collected in different developmental stages: eggs, the first to fifth instar larvae, 3-day-old pupae, and 3-day-old adults (females and males). A total of 60 eggs, 40 first instar, 20 second instar, 15 third instar, 10 fourth to fifth instar larvae, 3-day-old pupae, and 3-day-old adults were collected per biological replicate. (4) nine different tissues, including those from the head (without antennae and compound eye), chest, abdomen, leg, wing (from 10 adults), antennae, compound eye (from 30 adults), midgut and ovary (from 20 adults), were collected from O. furnacalis female adults aged 3 days old and were individually dissected on ice in phosphate-buffered saline solution with RNase inhibitor (TaKaRa, Dalian, China) using the Olympus binocular microscope (Germany). O. furnacalis female adults aged 3 days old (n = 30) were maintained in an incubator at 26°C and used as control (0 min). All samples were quickly frozen in liquid nitrogen and stored at −80°C for subsequent RNA extraction. Each treatment group had three biological replicates.
Total RNA Extraction and cDNA Synthesis
RNA was extracted from the O. furnacalis adults using the Trizol reagent (Invitrogen, Carlsbad, CA, United States) following the manufacturer’s protocol. The 260/280 ratio was measured using the NanoDrop 2000 (Thermo Fisher Scientific, Waltham, MA, United States) to evaluate the purity of the total RNA, and the integrity of the isolated RNA was verified using 1% agarose gel electrophoresis. Reverse transcription was conducted using the RevertAid First-Strand cDNA Synthesis Kit (Thermo Fisher) according to the manufacturer’s instructions. The synthesized first-strand cDNAs were stored at −20°C and used as templates.
Cloning OfJNK and Ofp38 From O. furnacalis
Based on conserved JNK and p38 sequences from Spodoptera litura, Dichocrocis punctiferalis, and H. armigera, degenerate primers were designed to amplify OfJNK and Ofp38 fragments (Table 1). Polymerase chain reaction (PCR) was conducted using the following amplification conditions: initial cycle at 94°C for 3 min, 35 amplification cycles (denaturation at 94°C for 30 s, annealing at 53–57°C for 30 s, and elongation at 72°C for 1 min), and a final elongation step at 72°C for 10 min. Electrophoresis of the PCR products was run on 1% agarose gel. The expected fragments were purified using the SanPrep DNA Extraction Kit [Sangon Biotech (Shanghai) Co., Ltd., Shanghai, China] and cloned into the pMD-18T vector (TaKaRa). The ligation products were then transformed into the Escherichia coli DH5α cell line (TaKaRa), and the cloned fragments of OfJNK and Ofp38 were sequenced by Sangon Biotech.

Table 1. Primers used in this study.
Based on the information obtained for the partially cloned OfJNK and Ofp38 fragments, gene-specific primers for 5′ and 3′ rapid amplification of cDNA ends (RACE) were designed using Primer Premier 6.0 (Table 1). The 5′ and 3′ RACE-ready cDNAs were synthesized using the SMARTTM RACE cDNA amplification kit (TaKaRa) following the manufacturer’s instructions. RACE PCR was conducted under the following conditions: 25 amplification cycles at 94°C for 30 s and annealing at 60–70°C for 30 s (depending on the primer) and 72°C for 3 min. RACE PCR products were cloned into a linearized pRACE vector (a SMARTer® RACE 5′/3′ Kit component) before sequencing. The complete OfJNK and Ofp38 cDNA sequences were assembled using DNAMAN 6.0.
Analysis of OfJNK and Ofp38 Sequences
Using the open reading frame (ORF) finder2, we identified the ORFs of OfJNK and Ofp38 and determined the amino acid sequences of the encoded proteins. The homology of JNK and p38 was analyzed using the National Center for Biotechnology Information/Basic Local Alignment Search Tool database with other sequences available in GeneBank. The molecular weights and isoelectric points (pH[I]s) of the proteins were obtained using ProtParam2. Multiple sequence alignment was performed using DNAMAN 6.0, and a phylogenetic tree was created in MEGA 6.0 using the neighbor-joining method (Tamura et al., 2013) with a bootstrap test of 1,000 repetitions. Phosphorylation sites were predicted using NetPhos 2.03, and protein domains or motifs were identified using SMART4.
Quantitative Real-Time PCR
To analyze OfJNK and Ofp38 expression levels, total RNA from different developmental stages, from various tissues, and under different environmental stresses of O. furnacalis was extracted and used to prepare cDNA templates using the PrimeScript RT Reagent Kit with gDNA Eraser (perfect real-time) (TaKaRa). The quantitative primers JNK-qF/JNK-qR and p38-qF/p38-qR were designed based on the JNK and p38 sequences in O. furnacalis, and the internal reference gene primers actin-qF/actin-qR and GAPDH-qF/GAPDH-qR were designed according to the reported β-actin and GAPDH in the O. furnacalis DNA sequences (Table 1). Each quantitative real-time PCR (qRT-PCR) was conducted in a 20 μL reaction volume that comprised the following components: 10 μL ilaqTM SYBR Green Supermix (2×), 1 μL cDNA template, 1 μL each primer (10 μM) (Table 1), and 7 μL ddH2O. RT-PCR was conducted using the Applied Biosystems 7500 RT-PCR system (Thermo Fisher) under the following conditions: denaturing at 95°C for 3 min followed by 39 cycles at 95°C for 30 s and 55°C for 30 s. To check the reproducibility of the experiment, qPCR for each treatment comprised three biological replications and three technical replications. The relative expression levels were quantified using the 2–ΔΔCT method (Livak and Schmittgen, 2001).
Statistical Analysis
All data were statistically analyzed using SPSS v. 20.0 (IBM Corporation, Armonk, NY, United States) and were compared using analysis of variance followed by the Tukey’s test. The data are presented as mean ± SEM. P < 0.05 was considered a statistically significant difference, which is clearly illustrated using different letters.
Results
Cloning OfJNK and Ofp38 and Sequence Analysis
OfJNK and Ofp38 were cloned using RT-PCR and RACE from O. furnacalis adults. We obtained 1,657 base pairs (bp) of the OfJNK cDNA (GenBank accession number MK779706) that contained a 472-bp 5′ untranslated region (UTR), 39-bp 3′ UTR, and 1,143-bp ORF encoding 381 amino acid residues; the molecular weight was calculated to be 43.44 kDa and theoretical pH(I) to be 6.06. Multiple sequence alignment showed that the OfJNK protein includes an active site, an activation loop structure, a substrate-binding site, and a threonine–proline–tyrosine motif (Figure 1A). The predicted serine/threonine protein kinase (S_TKc) domain was observed at positions 20–316. The identity of OfJNK compared with JNK in S. litura, B. mori, H. armigera, and Bombus terrestris was 90.7, 90.2, 81.16, and 88%, respectively (Figure 1A). Phylogenetic analysis showed that OfJNK was clustered with the JNK of other Lepidoptera and was most closely related to that of H. armigera and S. litura (Figure 1B).
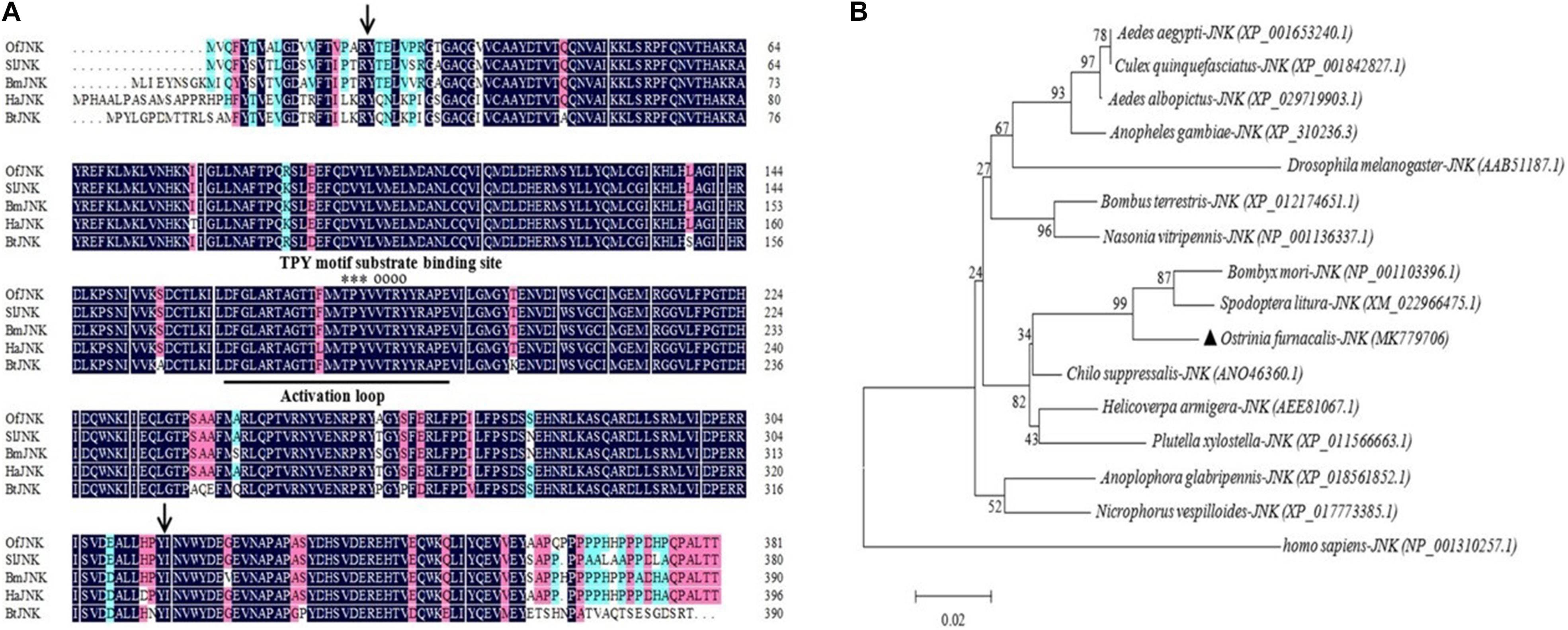
Figure 1. (A) Multiple sequence alignment of the amino acid sequences of Ostrinia furnacalis JNK with the JNK of other insects. In the representative JNK structure, the activation loop is underlined, substrate-binding site is indicated by empty circles, and threonine–proline–tyrosine (TPY) motif is indicated by asterisks found at positions 20–310 in the serine/threonine protein kinase (S_TKc) domain (black arrows). (B) The phylogenetic tree of OfJNK and JNK in various species. The location of OfJNK is indicated by a black-filled triangle. Abbreviations: Of, Ostrinia furnacalis (MK779706); Sl, Spodoptera litura (XM_022966475.1); Bm, Bombyx mori (NP_001103396.1); Ha, Helicoverpa armigera (AEE81067.1); Bt, Bombus terrestris (XP_012174651.1).
The 1,253 bp Ofp38 cDNA (GenBank accession No.: MK779707) we obtained contains a 156-bp 5′ UTR, 17-bp 3′ UTR, and 1,080 bp ORF encoding 360 amino acid residues with a calculated molecular weight of 41.72 kDa and theoretical pH(I) of 5.84. Pairwise and multiple sequence alignments showed that Ofp38 includes an active site, an activation loop structure, a specific threonine–glycine–tyrosine motif, a substrate-binding site ATRW, and an extracellular signal-regulated kinase docking (ED) site. The predicted S_TKc domain was observed at positions 20–304 (Figure 2A). The Ofp38 protein showed a 94.75% similarity with B. mori p38, followed by that of Nasonia vitripennis (85.12%), Aedes aegypti (82.77%), and Myzus persicae (79.34%). Phylogenetic analysis revealed that the Ofp38 protein was most homologous to that of S. litura, B. mori, and Amyelois transitella, which jointly constituted a correspondingly distinct clade (Figure 2B).
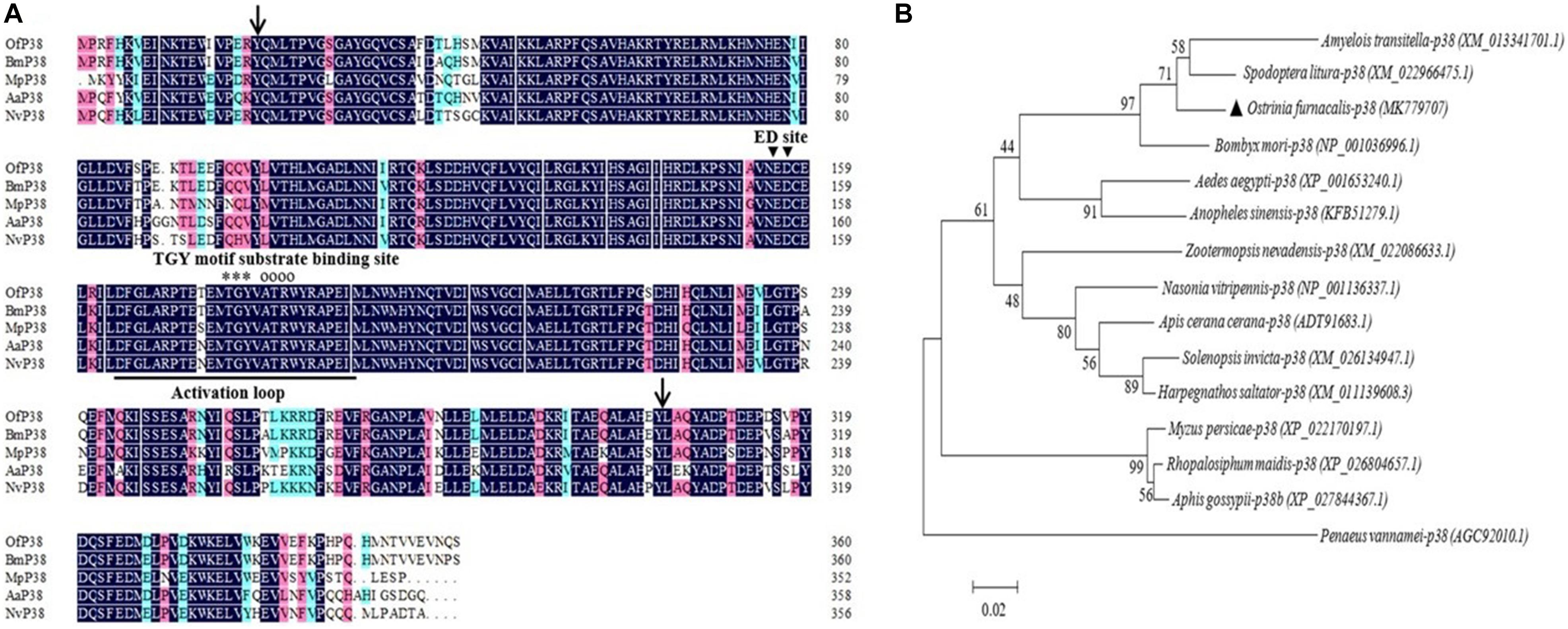
Figure 2. (A) Multiple sequence alignment of the amino acid sequences of Ofp38 with the p38 from other insects. The predicted serine/threonine protein kinase (S_TKc) domain was found at positions 20–304 (black arrows). In the typical p38 structure, the activation loop is underlined, substrate-binding site is indicated by empty circles, extracellular signal-regulated kinase (ERK) and docking (ED) site is indicated by the black-filled inverted triangle, and threonine–glycine–tyrosine (TGY) motif is indicated by asterisks. (B) The phylogenetic tree of Ofp38 and p38 in various species. The location of Ofp38 is indicated by a black-filled triangle. Abbreviations: Of, Ostrinia furnacalis (MK779707); Bm, Bombyx mori (NP_001036996.1); Mp, Myzus persicae (XP_022170197.1); Aa, Aedes aegypti (XP_001653240.1); Nv, Nasonia vitripenn (NP_001136337.1).
Developmental Expression Profiles of Tissue-Specific OfJNK and Ofp38
The developmental expression profiles of OfJNK and Ofp38 were quantified in the insect’s eggs, first to fifth instar larvae, 3-day-old pupae, and 3-day-old adults (females and males). The results showed that OfJNK and Ofp38 were expressed at all examined developmental stages (Figure 3). However, OfJNK was more highly expressed in the eggs than in the other developmental stages (Figure 3A), and the relative expression levels of Ofp38 in 3-day-old adult females and eggs were significantly higher than those in the other developmental stages (Figure 3B). The results of tissue-specific expression showed that OfJNK was more highly expressed in the wings than in the other tissues (Figure 4A) and that the relative expression levels of Ofp38 in the ovary were observably higher than those in the other tissues (Figure 4B).
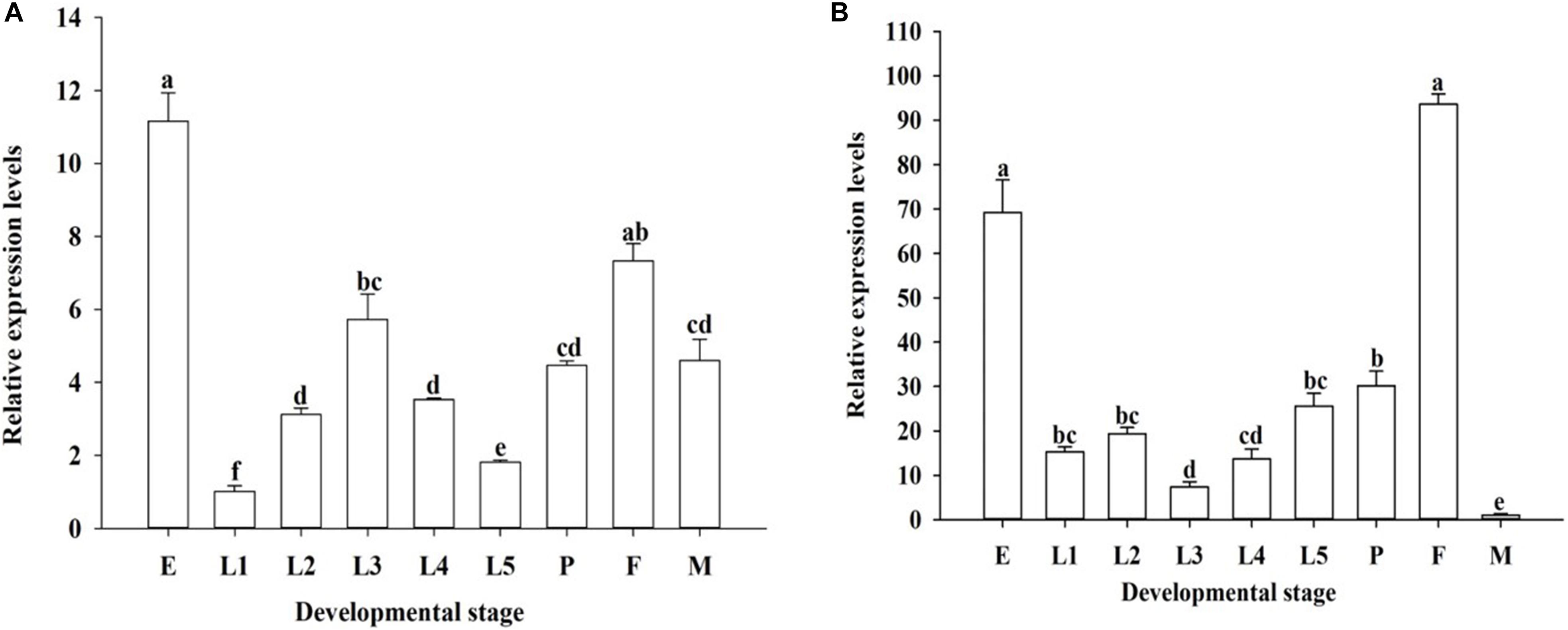
Figure 3. Relative expression levels of OfJNK (A) and Ofp38 (B) in eggs (E), first to fifth instar nymphs (L1–L5), 3-day-old pupae (P), and 3-day-old adult female (F) and male (M). Data represent mean ± SEM (N = 3). The OfJNK expression level in the first instar (L1) was used for baseline; the Ofp38 expression level in male (M) was used for baseline. Different letters above bars represent significant difference in relative expression levels (Tukey’s test; P < 0.05).
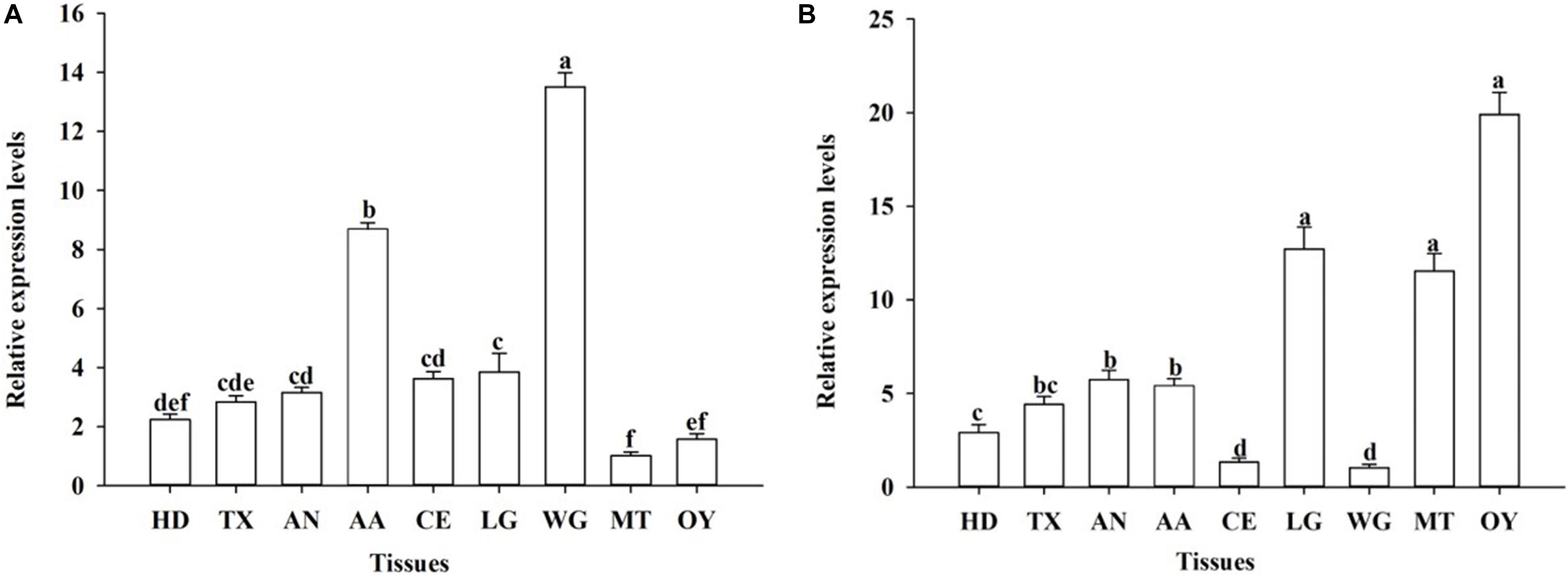
Figure 4. Relative expression levels of OfJNK (A) and Ofp38 (B) in the head (HD) (without antennae and compound eye), thorax (TX), abdomen (AN), antenna (AA), compound eye (CE), leg (LG), wing (WG), midgut (MT), and ovary (OY) of mixed 10–30 adult females. Data represent mean ± SEM (N = 3). The OfJNK expression level in midgut (MT) was used for baseline; the Ofp38 expression level in wing (WG) was used for baseline. Different letters above bars represent significant difference in relative expression levels (Tukey’s test; P < 0.05).
Expression of OfJNK and Ofp38 After Exposure to Low Temperature
To investigate whether the MAPKs are involved in the response of O. furnacalis to environmental stress, we measured the expression levels of OfJNK and Ofp38 under low temperature stress. After exposing 3-day-old adults to 4°C for different durations, the relative expression levels of OfJNK and Ofp38 were examined. The results showed that the expression levels first increased and eventually decreased with an increase in the treatment duration and that both OfJNK (Figure 5A) and Ofp38 (Figure 5B) were highly expressed at 90 min.
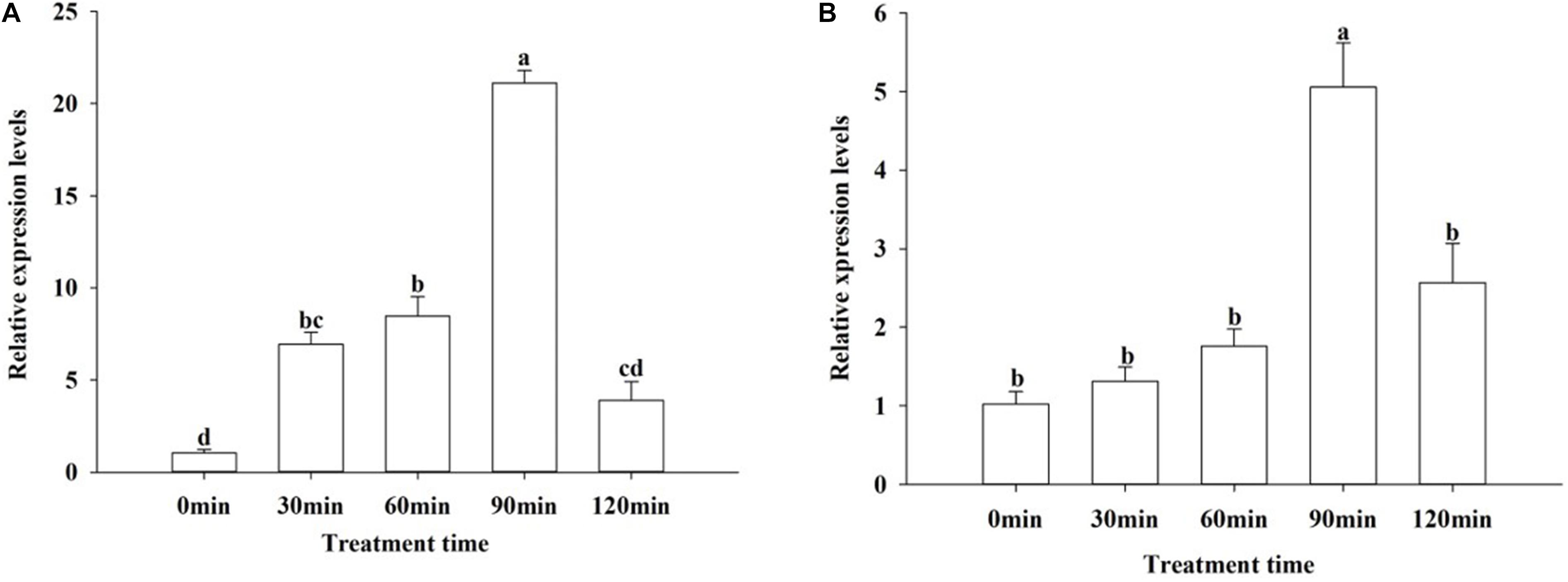
Figure 5. Relative expression levels of OfJNK (A) and Ofp38 (B) at low temperature. Data represent mean ± SEM (N = 3). The OfJNK and Ofp38 expression levels at 4°C treatment for 0 min (CK) were used for baseline. Different letters above bars represent significant difference in relative expression levels (Tukey’s test; P < 0.05).
Effect of UV Irradiation on OfJNK and Ofp38 Expression
To examine the effect of UV irradiation on OfJNK and Ofp38 expression, the insects were subjected to UVA exposure for different durations. Our results showed that the mRNA levels of OfJNK and Ofp38 first increased and then decreased as the duration of UVA exposure increased and that Ofp38 (Figure 6B) showed the highest expression levels at 30 min and OfJNK (Figure 6A) at 60 min.
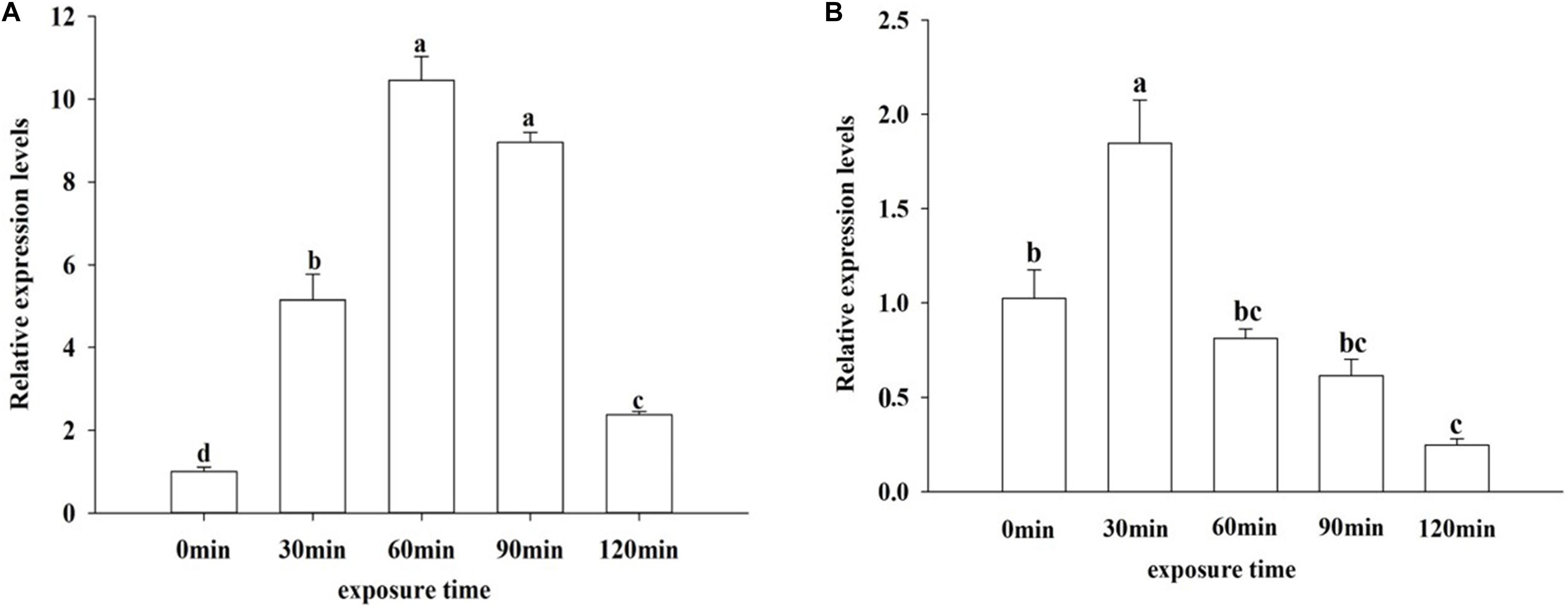
Figure 6. Relative expression levels of OfJNK (A) and Ofp38 (B) after treatment with ultraviolet irradiation for different durations. Data represent mean ± SEM (N = 3). The OfJNK and Ofp38 expression levels under UVA exposure for 0 min (CK) were used for baseline. Different letters above bars represent significant difference in relative expression levels (Tukey’s test; P < 0.05).
Discussion
Mitogen-activated protein kinases play vital roles in transmitting stress signals. Environmental stresses, such as cold, heat shock, UV irradiation, and viral and bacterial infections trigger these stress signals (Mizutani et al., 2003; Li et al., 2012). Previous studies on insects have shown that as important members of cell signaling pathways, JNK and p38 can be activated by various extracellular stresses such as UV irradiation, heat shock, pro-inflammatory factors, and specific antigens. Currently, the expression levels of JNK and p38 have been well recorded under different environmental stressors, at different developmental stages, and in various tissue types in several insects. However, the information on JNK and p38 in agricultural insects is limited. In our study, these two MAPK genes were cloned from O. furnacalis and were subsequently named OfJNK and Ofp38, respectively. Moreover, their molecular properties and expression levels were characterized under different environmental stimuli. The sequence and phylogenetic analyses showed a high sequence homology of the OfJNK and Ofp38 proteins with the JNK and p38 proteins of S. litura, B. mori, H. armigera, and N. vitripennis (Figures 1A, 2A). Our study results suggested that the JNK and p38 signaling cascade systems may play an important role in the response of O. furnacalis to various environmental stresses (Xia et al., 1995; Ludwig et al., 2003).
Using qPCR, we observed that OfJNK and Ofp38 were expressed in the developmental stages of O. furnacalis but that their expression levels were significantly different depending on the specific stage. OfJNK was highly expressed in the eggs compared with those in the other developmental stages. This is similar to previous study results that the JNK signaling pathway was involved in egg formation in Drosophila (Suzanne et al., 2001). The relative expression levels of Ofp38 in eggs and 3-day-old adult females were significantly higher than those in the other developmental stages. This difference might be related to the changes made by the p38 signaling pathway to the expression levels of this gene through the phosphorylation of transcription factors at different developmental stages of the insect, thereby allowing it to participate in cell growth and proliferation (Li et al., 2003). Tissue-specific quantitative results showed that OfJNK and Ofp38 were expressed in various O. furnacalis tissues, which is consistent with the study results on mammalian JNK and p38 reported by Widmann et al. (1999). The expression levels of OfJNK were higher in the wings than in the other tissues. This corresponds to the results of previous studies in which the JNK signaling pathway reportedly participated in wing formation and development of the insect (Zhang et al., 2019). Similarly, we found that Ofp38 was significantly higher in the ovary than in the other tissues, indicating that Ofp38 may be involved in the process of ovarian cell growth and development in O. furnacalis (Lu et al., 2007; Sheng et al., 2015).
Furthermore, we detected the expression levels of OfJNK and Ofp38 in O. furnacalis under low temperature and UVA irradiation. Low temperature and UV irradiation are important stress factors that impact every living creature including insects. Some studies have reported that JNK and p38 could respond to various environmental stresses, such as cold stress, heat shock, oxidative stress, immunostimulants, serum starvation, and UV irradiation, in cells (Hanks and Hunter, 1995; Han et al., 1998). In the present study, the expression levels of OfJNK and Ofp38 significantly increased when O. furnacalis adults were exposed to 4°C for 90 min, which indicated that JNK and p38 might be involved in responses to low temperatures. Insects can feel signals and transmit them to the cellular machinery through signal transduction to regulate gene expression. We suspected that JNK/P38 pathway might be activated by low temperatures, which could promote DNA damage repair and cell growth in O. furnacalis to resist environmental stress. This result is consistent with a report that JNK is activated to enhance DNA repair and promote cell survival in DNA damage response and the activated p38 pathway mediates cell growth and proliferation (Johansson et al., 2000; Chock et al., 2010). However, as time elapsed, the JNK and p38 signals were attenuated. It may be due to the fact that prolonged stress beyond the maximum tolerance of the JNK/P38 pathway leads to cell damage. It is reported that short-term activation of JNK and p38 can promote cell survival, while sustained activation can cause cell apoptosis (Harper and Lograsso, 2001). This has been founded in S. crassipalpis and the liver and kidneys of the turtles (Greenway and Storey, 2000; Fujiwara and Denlinger, 2007). UV-A light used as light sources in light traps is regarded as an environmental stress factor to insects (Zhang and Meng, 2018). In our study, when O. furnacalis adults were exposed to UVA, the expression levels of Ofp38 and OfJNK significantly increased at 30 and 60 min, respectively, which indicated that JNK and p38 might be involved in responses to UVA exposure. We suspected that JNK/P38 pathways could be activated UV-A irradiation, which in turn helps tolerate UV stress and improves the ability of O. furnacalis to adapt to stress. However, with exposure to UV light for longer time (120 min), the expression levels of OfJNK and Ofp38 decreased. This result agrees with findings of Liu et al. (2019) who reported that UVA stress induced mRNA expression of JNK in H. armigera adult females and that the highest expression level was at 60 min, which further decreased with the exposure time.
Conclusion
In conclusion, the results of this study suggested that OfJNK and Ofp38 responded to low temperature and UV irradiation, and that the MAPK signaling pathway plays critical roles in response to environmental stresses in O. furnacalis. We believe these results will broaden our knowledge regarding the MAPK signaling pathway in insects.
Data Availability Statement
Publicly available datasets were analyzed in this study. This data can be found in GenBank under the accession numbers MK779706 and MK779707.
Author Contributions
C-YZ conceived and designed the experiments. LS cloned and analyzed OfJNK and Ofp38, examined OfJNK and Ofp38 expression levels, and prepared the manuscript. All authors finalized the manuscript, and have read and approved the final manuscript.
Funding
This research was supported by the National Key R&D Program of China (2017YFD0200900), National Natural Science Foundation of China (31401754), and Provincial Key Project for Agricultural Science and Technology of Guizhou (NY20123007).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Footnotes
- ^ http://kr.expasy.org/tools/protparam.html
- ^ http://www.ncbi.nlm.nih.gov/orffinder/
- ^ http://www.cbs.dtu.dk/services/NetPhos/
- ^ http://smart.embl-heidelberg.de
References
Afidchao, M. M., Musters, C., and Snoo, G. R. D. (2013). Asian corn borer (ACB) and non-ACB pests in GM corn (Zea mays L.) in the Philippines. Pest Manag. Sci. 69, 792–801. doi: 10.1002/ps.3471
Ali, A., Rashid, M. A., Huang, Q. Y., and Lei, C. L. (2017). Influence of UV-A radiation on oxidative stress and antioxidant enzymes in Mythimna separata (Lepidoptera: Noctuidae). Environ. Sci. Pollut. Res. 24, 1–7. doi: 10.1007/s11356-017-8514-7
Bale, J. S., and Hayward, S. A. L. (2010). Insect overwintering in a changing climate. J. Exp. Biol. 213, 980–994. doi: 10.1242/jeb.037911
Chock, K., Allison, J. M., and Elshamy, W. M. (2010). BRCA1-IRIS overexpression abrogates UV-induced p38MAPK/p53 and promotes proliferation of damaged cells. Oncogene 29, 5274–5285. doi: 10.1038/onc.2010.262
Cowan, K. J., and Storey, K. B. (2003). Mitogen-activated protein kinases: new signaling pathways functioning in cellular responses to environmental stress. J. Exp. Biol. 206, 1107–1115. doi: 10.1242/jeb.00220
Feng, C. J., Huang, J. H., Song, Q. S., Stanley, D., Lü, W. J., Zhang, Y., et al. (2011). Parasitization by Macrocentrus cingulum (Hymenoptera: Braconidae) influences expression of prophenoloxidase in Asian corn borer Ostrinia furnacalis. Arch. Insect Biochem. Physiol. 77, 99–117. doi: 10.1002/arch.20425
Feng, C. J., Lu, W. J., An, Dong Q., Chen, Jun, and Fu, W. J. (2007). Effect of low temperature treatment on larvae of the Asian corn borer, Ostrinia furnacalis (Guenée) (Lepidoptera: Pyralidae). Acta Entomol. Sin. 50, 1–6.
Fujita, J. (1999). Cold shock response in mammalian cells. J. Mol. Microbiol. Biotechnol. 1, 243–255.
Fujiwara, Y., and Denlinger, D. L. (2007). P38 mapk is a likely component of the signal transduction pathway triggering rapid cold hardening in the flesh fly Sarcophaga crassipalpis. J. Exp. Biol. 210, 3295–3300. doi: 10.1242/jeb.006536
Fujiwara, Y., Shindome, C., Takeda, M., and Shiomi, K. (2006). The roles of ERK and p38 MAPK signaling cascades onembryonic diapause initiation and termination of thesilkworm, Bombyx mori. Insect Biochem. Mol. Biol. 36, 47–53. doi: 10.1016/j.ibmb.2005.10.005
Greenway, S. C., and Storey, K. B. (2000). Mitogen-activated protein kinases and anoxia tolerance in turtles. J. Exp. Zool. 287, 477–484. doi: 10.1002/1097-010x(20001201)287:7<477::aid-jez3>3.0.co;2-4
Grubor-Lajsic, G., Block, W., and Worland, R. (1992). Comparison of the cold hardiness of two larval Lepidoptera (Noctuidae). Physiol. Entomol. 17, 148–152. doi: 10.1111/j.1365-3032.1992.tb01192.x
Han, Z. S., Enslen, H., Hu, X., Meng, X., Wu, I. H., Barrett, T., et al. (1998). A conserved p38 mitogen-activated protein kinase pathway regulates Drosophila immunity gene expression. Mol. Cell Biol. 18, 3527–3539.
Hanks, S. K., and Hunter, T. (1995). Protein kinases 6. the eukaryotic protein kinase superfamily: kinase (catalytic) domain structure and classification. FASEB J. 9, 576–596. doi: 10.1096/fasebj.9.8.7768349
Harper, S. J., and Lograsso, P. (2001). Signalling for survival and death in neurones: the role of stress-activated kinases, jnk and p38. Cell. Signal. 13, 299–310. doi: 10.1016/s0898-6568(01)00148-6
He, K., Wang, Z., Zhou, D., Wen, L. P., Song, Y. Y., and Yao, Z. Y. (2003). Evaluation of transgenic Bt corn for resistance to the Asian corn borer (Lepidoptera: Pyralidae). J. Econ. Entomol. 96, 935–940. doi: 10.1093/jee/96.3.935
Hohmann, S. (2002). Osmotic stress signaling and osmoadaptation in yeasts. Microbiol. Mol. Biol. Rev. 66, 300–372. doi: 10.1128/mmbr.66.2.300-372.2002
Hu, Z. F., Xing, Y., Zhang, T. S., Dong, H., and Cong, B. (2017). Expression analysis of heat shock protein gene (HSPs) of Ostrinia furnacalis (Guenée) under alternating temperature stress. J. Shenyang Agric. Univ. 48, 213–218.
Huang, G. H., Shi, L. Z., and Chi, H. (2009). Regulation of JNK and p38 MAPK in the immune system: signal integration, propagation and termination. Cytokine 48, 161–169. doi: 10.1016/j.cyto.2009.08.002
Johansson, N., Ala-aho, R., Uitto, V., Grénman, R., Fusenig, N. E., López-Otín, C., et al. (2000). Expression of collagenase-3 (MMP-13) and collagenase-1 (MMP-1) by transformed kerati-nocytes is dependent on the activity of p38 mitogen-activated protein kinase. J. Cell Sci. 113, 227–235.
Johnson, G. L., and Lapadat, R. (2002). Mitogen-activated protein kinase pathways mediated by ERK, JNK, and p38 protein kinases. Science 298, 1911–1912. doi: 10.1126/science.1072682
Li, F. F., Xia, J., Li, J. M., Liu, S. S., and Wang, X. W. (2012). p38/MAPK is a component of the signal transduction pathway triggering cold stress response in the MED cryptic species of Bemisia tabaci. J. Integr. Agric. 11, 303–311. doi: 10.1016/s2095-3119(12)60014-8
Li, X., Udagawa, N., Takami, M., Sato, N., Kobayashi, Y., and Takahashi, N. (2003). p38 mitogen-activated protein kinase is crucially involved in osteoclast differentiation but not in cytokine production, phagocytosis, or dendritic cell differentiation of bone marrow macrophages. Endocrinology 144, 4999–5005. doi: 10.1210/en.2003-0166
Liu, X. F., Meng, J. Y., Zhao, X. C., and Zhang, C. Y. (2019). cDNA cloning and expression profiling of the c-Jun N-terminal kinase gene and its response to UV-A stress in Helicoverpa armigera (Lepidoptera: Noctuidae). Acta Entomol. Sin. 62, 407–417.
Liu, Y., Shen, D., Zhou, F., Wang, G., and An, C. (2014). Identification of immunity-related genes in Ostrinia furnacalis against entomopathogenic fungi by RNA-seq analysis. PLoS One 9:e86436. doi: 10.1371/journal.pone.0086436
Liu, F., Meng, J. Y., Yang, C. L., and Zhang, C. Y. (in press). Cloning and(expression)profile of vitellogenin gene and its response to UV-A stress in Ostrinia furnacalis (Guenée). Acta Entomol. Sin.
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C (T)) method. Methods 25, 402–408.
Lu, M. S., Deng, S., Xiao, L., Liang, M. Z., and Wang, Z. H. (2007). Study on the relationship between P38 MAPK activity and apoptosis in drug-resistant cell line of ovarian carcinoma. J. Harbin. Med. Univ. 41, 125–128.
Ludwig, S., Planz, O., Pleschka, S., and Wolff, T. (2003). Influenza-virus-induced signaling cascades: targets for antiviral therapy? Trends Mol. Med. 9, 46–52. doi: 10.1016/s1471-4914(02)00010-2
Meng, J. Y., Zhang, C. Y., and Lei, C. L. (2010). A proteomic analysis of Helicoverpa armigera adults after exposure to UV light irradiation. J. Insect Physiol. 56, 405–411. doi: 10.1016/j.jinsphys.2009.11.015
Meng, J. Y., Zhang, C. Y., Zhu, F., Wang, X. P., and Lei, C. L. (2009). Ultraviolet light-induced oxidative stress: effects on antioxidant response of Helicoverpa armigera adults. J. Insect Physiol. 55, 588–592. doi: 10.1016/j.jinsphys.2009.03.003
Meng, S. S., Li, J. Q., Ruan, M. G., Lu, X. Y., and Ma, J. (2015). Cloning and expression analysis of extrac-ellular signal-regulated kinase ERK gene from the desert beetle Microdera punctipennis. J. Environ. Entomol. 37, 987–995.
Meyer-Rochow, V. B., Kashiwagi, T., and Eguchi, E. (2002). Selective photoreceptor damage in four species of insects induced by experimental exposures to UV-irradiation. Micron 33, 23–31. doi: 10.1016/s0968-4328(00)00073-1
Mizutani, T., Kobayashi, M., Eshita, Y., Inanami, O., Yamamori, T., Goto, A., et al. (2003). Characte- rization of JNK-like protein derived from a mosquito cell line, C6/36. Insect Mol. Biol. 129, 61–66. doi: 10.1046/j.1365-2583.2003.00387.x
Ono, K., and Han, J. (2000). The p38 signal transduction pathway activation and function. Cell Signal. 12, 1–13. doi: 10.1016/s0898-6568(99)00071-6
Rahaus, M., Desloges, N., and Wolff, M. H. (2004). Replication of varicella-zoster virus is influenced by the levels of JNK/SAPK and p38/MAPK activation. J. Gen. Virol. 85, 3529–3540. doi: 10.1099/vir.0.80347-0
Shang, Q. L., Pan, Y. O., Peng, T. F., Yang, S., Lu, X., Wang, Z. Y., et al. (2015). Proteomics analysis of overexpressed plasma proteins in response to cold acclimation in Ostrinia furnacalis. Arch. Insect Biochem. Physiol. 90, 195–208. doi: 10.1002/arch.21302
Sheng, M., Zhang, H. Y., Song, D. D., and Zou, C. H. (2015). P38 MAPK signaling pathway is involved in expression regulation of uPA protein in ovarian cancer cell and tissues. Cancer Res. Prev. Treat. 42, 789–793.
Suzanne, M., Perrimon, N., and Noselli, S. (2001). The Drosophila jnk pathway controls the morphog- enesis of the egg dorsal appendages and micropyle. Dev. Biol. 237, 282–294. doi: 10.1006/dbio.2001.0384
Tamura, K., Stecher, G., Peterson, D., Filipski, A., and Kumar, S. (2013). MEGA6: molecular evoluti- onary genetics analysis version 6.0. Mol. Biol. Evol. 30, 2725–2729. doi: 10.1093/molbev/mst197
Vogelweith, F., Thiéry, D., Quaglietti, B., Moret, Y., and Moreau, J. (2011). Host plant variation plastically impacts different traits of the immune system of a phytophagous insect. Funct. Ecol. 25, 1241–1247. doi: 10.1111/j.1365-2435.2011.01911.x
Wang, L. L., Huang, H., Zhang, C. R., Xia, J., Liu, S. S., and Wang, X. W. (2013). Cloning and functional characterization of c-jun NH2-terminal kinase from the mediterranean species, of the whitefly Bemisia tabaci complex. Int. J. Mol. Sci. 14, 13433–13446. doi: 10.3390/ijms140713433
Wang, Z. Y., Lu, X., He, K. L., and Zhou, D. R. (2000). Review of history, present situation and prospect of the Asian maize borer research in China. Shenyang Agric. Univ. 31, 402–412.
Widmann, C., Gibson, S., Jarpe, M. B., and Johnson, G. L. (1999). Mitogen-activated protein kinase: conservation of a three-kinase module from yeast to human. Physiol. Rev. 79, 143–180. doi: 10.1152/physrev.1999.79.1.143
Xia, Z., Dickens, M., Raingeaud, J., Davis, R. J., and Greenberg, M. E. (1995). Opposing effects of ERK and JNK-p38 MAP kinases on apoptosis. Science 270, 1326–1331. doi: 10.1126/science.270.5240.1326
Zhang, C. Y., and Meng, J. Y. (2018). Identification of differentially expressed proteins in Ostrinia furnacalis adults after exposure to ultraviolet A. Environ. Sci. Pollut. Res. 25, 25071–25079. doi: 10.1007/s11356-018-2580-3
Zhang, M. M., Zhou, F., Chu, Y., Zhao, Z. W., and An, C. J. (2013). Identification and expression profile analysis of antimicrobial peptide/protein in Asian corn borer, Ostrinia furnacalis (Guenée). Int. J. Biol. Sci. 9, 1004–1012. doi: 10.7150/ijbs.6813
Zhang, W., and Liu, H. T. (2002). MAPK signal pathways in the regulation of cell proliferation in mammalian cells. Cell Res. 12, 9–18. doi: 10.1038/sj.cr.7290105
Keywords: Ostrinia furnacalis, c-Jun N-terminal kinase, p38 mitogen-activated protein kinase, gene cloning, expression analysis, environmental stressors
Citation: Su L, Meng J-Y, Yang H and Zhang C-Y (2020) Molecular Characterization and Expression of OfJNK and Ofp38 in Ostrinia furnacalis (Guenée) Under Different Environmental Stressors. Front. Physiol. 11:125. doi: 10.3389/fphys.2020.00125
Received: 25 June 2019; Accepted: 04 February 2020;
Published: 25 February 2020.
Edited by:
Shigehiro Namiki, The University of Tokyo, JapanReviewed by:
Yuan-Xi Li, Nanjing Agricultural University, ChinaQingjun Wu, Chinese Academy of Agricultural Sciences, China
Copyright © 2020 Su, Meng, Yang and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Chang-Yu Zhang, emN5MTEyMUBhbGl5dW4uY29t