
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Physiol. , 11 February 2020
Sec. Fractal Physiology
Volume 11 - 2020 | https://doi.org/10.3389/fphys.2020.00066
This article is part of the Research Topic The New Frontier of Network Physiology: From Temporal Dynamics to the Synchronization and Principles of Integration in Networks of Physiological Systems View all 65 articles
 Thomas A. W. Bolton1,2*†
Thomas A. W. Bolton1,2*† Diana Wotruba3,4†
Diana Wotruba3,4† Roman Buechler4,5
Roman Buechler4,5 Anastasia Theodoridou4,6
Anastasia Theodoridou4,6 Lars Michels5
Lars Michels5 Spyros Kollias5
Spyros Kollias5 Wulf Rössler3,4,6,7
Wulf Rössler3,4,6,7 Karsten Heekeren4,6
Karsten Heekeren4,6 Dimitri Van De Ville1,2†
Dimitri Van De Ville1,2†Emerging evidence has attributed altered network coordination between the default mode, central executive, and salience networks (DMN/CEN/SAL) to disturbances seen in schizophrenia, but little is known for at-risk psychosis stages. Moreover, pinpointing impairments in specific network-to-network interactions, although essential to resolve possibly distinct harbingers of conversion to clinically diagnosed schizophrenia, remains particularly challenging. We addressed this by a dynamic approach to functional connectivity, where right anterior insula brain interactions were examined through co-activation pattern (CAP) analysis. We utilized resting-state fMRI in 19 subjects suffering from subthreshold delusions and hallucinations (UHR), 28 at-risk for psychosis with basic symptoms describing only self-experienced subclinical disturbances (BS), and 29 healthy controls (CTR) matched for age, gender, handedness, and intelligence. We extracted the most recurring CAPs, compared their relative occurrence and average dwell time to probe their temporal expression, and quantified occurrence balance to assess the putative loss of competing relationships. Our findings substantiate the pivotal role of the right anterior insula in governing CEN-to-DMN transitions, which appear dysfunctional prior to the onset of psychosis, especially when first attenuated psychotic symptoms occur. In UHR subjects, it is longer active in concert with the DMN and there is a loss of competition between a SAL/DMN state, and a state with insula/CEN activation paralleled by DMN deactivation. These features suggest that abnormal network switching disrupts one's capacity to distinguish between the internal world and external environment, which is accompanied by inflexibility and an excessive awareness to internal processes reflected by prolonged expression of the right anterior insula-default mode co-activation pattern.
A fundamental feature of the healthy human brain is its intrinsic organization into coupled functional networks (Fox et al., 2005), which crucially includes coordinated default mode network (DMN) and central executive network (CEN) activity for cognitive and executive functions (Bressler and Kelso, 2001; Buckner et al., 2008). Their antagonistic activity putatively reflects competing modes of information processing (Fox et al., 2005; Fransson, 2005). The DMN, with its posterior cingulate cortex and medial prefrontal cortex hubs, serves untargeted inner thought (Andrews-Hanna, 2012); the CEN, particularly within the dorsolateral prefrontal and posterior parietal cortices, contributes to focused stimulus-dependent attention (Fox et al., 2005; Seeley et al., 2007; Menon and Uddin, 2010). The right anterior insula, the main salience network (SAL) driver, causally regulates this competing inter-network activity and thus facilitates both bottom-up perception and reorienting of attention, contributing as such to appropriate behavioral responses to salient stimuli (Menon and Uddin, 2010).
Disturbed DMN/CEN/SAL coordination has been related to the confusion of internally and externally focused attention and to the disturbance of cognition, as seen in psychotic disorders (Carhart-Harris and Friston, 2010; Nekovarova et al., 2014). In support of this, emerging evidence has attributed cognitive deficits in schizophrenia to dysfunctions in proper DMN/CEN coordination (Whitfield-Gabrieli et al., 2009; Chai et al., 2011; Whitfield-Gabrieli and Ford, 2012), whereas SAL anomalies have been mostly linked to reality distortion (Palaniyappan and Liddle, 2012; Pu et al., 2012), and thus posited to play a cardinal role in the development of psychotic symptoms (Palaniyappan et al., 2012a,b). Along the same line, we have recently demonstrated a loss of CEN-DMN anticorrelation, accompanied by DMN, CEN, and SAL anatomical overlap, in spontaneous brain activity of subjects at-risk for psychosis (Wotruba et al., 2014b). Importantly, those measures were associated differentially with cognitive (DMN/CEN) and psychopathological (SAL) symptoms.
A better understanding of the neurobiological underpinnings of different at-risk criteria is key to resolve possibly distinct harbingers of conversion to clinically diagnosed schizophrenia, and thus enable early intervention and lead to improved patient outcomes (Satterthwaite and Baker, 2015). Thus, different at-risk criteria have been conceptualized, including basic (BS) or ultra-high risk (UHR) symptoms. The former have been suggested to describe subtle, subclinical disturbances in mental processes, and to be the most direct self-experienced expression of the underlying neurobiological aberrations of schizophrenia; the latter, including subthreshold delusions and hallucinations, were conceptualized as secondary phenomena occurring at a later stage (Schultze-Lutter et al., 2016).
So far, few neuroimaging studies explore the relevance of these different conceptualizations, although divergent structural (Hurlemann et al., 2008; Koutsouleris et al., 2009a,b; Harrisberger et al., 2016) and functional (Ebisch et al., 2013, 2014; Wotruba et al., 2014a,b) brain abnormalities could be associated with different at-risk criteria.
Most resting-state studies in schizophrenia hypothesize stationary connectivity between brain regions, an oversimplification: for example, although the DMN and CEN are anticorrelated most of the time, they also exhibit some temporal intervals of correlated activity (Chang and Glover, 2010). Complex facets of brain activity, such as its non-stationary (de Pasquale et al., 2018; O'Neill et al., 2018) or scale-free (Van Den Heuvel et al., 2008; Van De Ville et al., 2010) nature, should thus be taken into account.
The growing field of network physiology—see Ivanov and Bartsch (2014) and Ivanov et al. (2016) for reviews—precisely attempts to quantify these complex properties. In the context of electroencephalography, approaches such as time-delay stability (Bashan et al., 2012; Bartsch et al., 2015) or delay-correction landscape (Lin et al., 2016) analyses characterize the nature and stability of the interactions between different network nodes, which jointly consist in different frequency rhythms at which functional processing occurs, and in organs under neural regulation (e.g., the eyes or the heart).
This sophisticated dynamics of brain function is modulated by the physiological state, as seen in the case of sleep stages (Bashan et al., 2012) or concomitant physical exercise and cognitive processing (Ciria et al., 2019). In addition, alterations are also seen in disease (Goldberger et al., 2002), which positions the study of resting-state functional brain dynamics as a question of particular interest regarding the emergence of pre-psychotic symptoms.
Recent electroencephalography work has shown the merits of probing the non-linear relationships between not just frequency rhythms, but also spatially remote brain centers (Liu et al., 2015). Functional magnetic resonance imaging (fMRI) is a particularly fitting imaging modality for this purpose, owing to its finer spatial resolution. Dedicated analytical strategies must be established, and accordingly, dynamic functional connectivity (Hutchison et al., 2013; Preti et al., 2017) analyses of schizophrenia (Damaraju et al., 2014; Du et al., 2016; Miller et al., 2016), and time-resolved fMRI investigations (Karahanoğlu and Van De Ville, 2017), have been gaining popularity.
However, the consideration of impairments at the level of specific network-to-network interactions, and so, possible maladaptive dynamics of the particular DMN/CEN/SAL relationship, remains subject to debate. Further, the analysis of connectivity, a second-order statistic, lowers the effective temporal resolution of the data, justifying recent developments toward frame-wise techniques to extract brain networks and analyse their interplays (Caballero-Gaudes et al., 2013; Karahanoğlu and Van De Ville, 2015).
Co-activation pattern (CAP) analysis (Liu and Duyn, 2013) is one such approach, where the sets of regions co-activating with a seed region of interest at different time points can be disentangled. To probe network-specific dynamic impairments, spatial or temporal features of the extracted CAPs can then be analyzed (Amico et al., 2014; Chen et al., 2015).
To improve the early recognition of psychosis and clarify the underlying role of the right anterior insula and co-activating regions, we carried resting-state fMRI on 25 subjects at-risk for psychosis with BS criteria, with full insight into their abnormal nature (Klosterkötter et al., 2001; Schultze-Lutter et al., 2016); 18 individuals with attenuated and/or brief intermittent psychotic symptoms (Yung and Mcgorry, 2007)—UHR criteria; and 29 healthy controls (CTR), matched on premorbid intelligence, age, handedness, and gender. We extracted six recurring CAPs, and given our specific interest in the interplay between the SAL, DMN, and CEN, focused the analysis on related ones, comparing their temporal expression across groups. We considered normalized counts, informing on the relative occurrence of each CAP, and duration, characterizing the average time for which a CAP is sustained. To assess the putative loss of competing relationship between CEN/DMN/SAL-containing CAPs in the at-risk stages for psychosis, we also quantified the balance in occurrence between these patterns of interest.
The present study included 76 participants (29 CTR, 28 BS, 19 UHR), and was approved by the local ethics committee of Zürich. Risk groups were recruited in the Swiss region of Zürich within the context of a larger study on early psychosis recognition (Theodoridou et al., 2014). Following an initial screening, in-person diagnostic interviews were administered, and a complete project description was provided. All subjects provided their written, informed consent. The procedure and imaging data acquisition were identical to those used in a previous study (Wotruba et al., 2014b).
Participants reporting at least one cognitive-perceptive basic symptom or at least two cognitive disturbances, as assessed by the Schizophrenia Proneness Interview (Schultze-Lutter et al., 2007) or, for adolescents (age < 18), by the Schizophrenia Proneness Interview “Child and Youth” version (Fux et al., 2013), fulfilled the inclusion criteria for the BS status. Those describing at least one attenuated psychotic symptom or brief, limited, intermittent psychotic symptoms, as assessed by the Structured Interview for Prodromal Syndromes (Miller et al., 2003), fulfilled the criterion for UHR status. Five BS and four UHR subjects were taking second-generation (atypical) antipsychotic medication at the time of scanning. Chlorpromazine equivalents were calculated for them (Andreasen et al., 2010). Five BS and three UHR subjects were being treated with an antidepressant. Our healthy CTRs were screened with the Mini-International Neuropsychiatric Interview (Sheehan et al., 1998) to ensure that none had any current or prior history of psychiatric illness. Those receiving any medication were excluded.
Exclusion criteria were contraindications against MRI, pregnancy, a history of neurological illness, as well as drug or alcohol dependence. Structural MRI scans were checked by an experienced neuroradiologist, excluding participants with structural brain abnormalities. The groups had a mean estimated premorbid intelligence slightly above average, as assessed using a German test for estimating premorbid, verbal intelligence—Mehrfachwortschatz Test, Version B (Lehrl, 1999); adolescent subjects (age < 20) adopted the Leistungsprüfsystem-3 German test for fluid, nonverbal intelligence (Horn, 1983). Handedness was examined by the Edinburgh Handedness questionnaire (Oldfield, 1971).
A list of all 76 subjects, including their taken antipsychotic drugs/antidepressants and exclusion criteria—as further detailed below—is provided in Table S1.
Resting-state fMRI data were acquired at the University Hospital of Psychiatry Zürich, Switzerland, using a Philips Achieva TX 3 T whole-body MR unit with an eight-channel head coil. Functional scans (6 min runs) involved a sensitivity-encoded single-shot echo-planar (factor 1) -weighted echo-planar imaging sequence [repetition time = 2,000 ms; echo time = 35 ms; field of view = 220 × 220 mm2; acquisition matrix = 88 × 87, interpolated to 128 × 128; 32 contiguous slices with a spatial resolution of 2.5 × 2.5 × 4 mm3 (reconstructed 1.72 × 1.72 × 4 mm3); flip angle θ = 78°; and sensitivity-encoded acceleration factor R = 1.8]. We also acquired three-dimensional T1-weighted anatomical images [160 slices; TR = 1,900 ms; TE = 2.2 ms; inversion time = 900 ms; θ = 78°; spatial resolution of 1 × 1 × 1 mm3 (reconstructed 0.94 × 0.94 × 1 mm3); field of view = 240 × 240 mm2]. Echo-planar imaging sequences were conducted under darkness; participants were asked to keep their eyes closed, lying quietly without falling asleep, as confirmed after scanning. To minimize arousal and anxiety effects, acquisition started 10 min after moving the subjects to their final MR bore positioning.
Preprocessing was conducted using SPM8 (Wellcome Department of Imaging Neuroscience, London, UK; http://www.fil.ion.ucl.ac.uk/spm), running in Matlab (Mathworks Inc., Sherbon, MA, USA), and the conn toolbox pipeline (version 15a). We performed realignment, slice timing correction, co-registration to structural T1 scan, spatial normalization to Montreal Neurological Institute space, and spatial smoothing (6 mm full width at half maximum). Spurious sources of noise were estimated by the anatomical component-based noise reduction strategy (Behzadi et al., 2007), and included with the six movement parameters as first level nuisance co-variates in a General Linear Model. Temporal band-pass filtering (0.01–0.10 Hz) and z-scoring were applied, and framewise displacement was quantified using Power's criterion (Power et al., 2012). We excluded three subjects (2 BS and 1 UHR) for whom more than 20% of volumes were censored.
Figure 1 illustrates the key methodological steps underlying CAP analysis. Based on previous analyses (Wotruba et al., 2014b) and recently proposed theoretical models for disturbance in the triple network (Menon, 2011), we selected a right anterior insula seed. A spherical region (8 mm radius, Montreal Neurological Institute coordinates: x = 38, y = 22, z = −10) based on previous studies (Seeley et al., 2007; Woodward et al., 2011) was created with the MARSBAR toolbox (http://marsbar.sourceforge.net/), and had its activity thresholded to solely retain frames with signal larger than T = 0.5 (seed activation). To rule out biases due to a different number of selected frames across groups, we also computed the amount of baseline, unselected time points, and included it as a co-variate in our analyses.
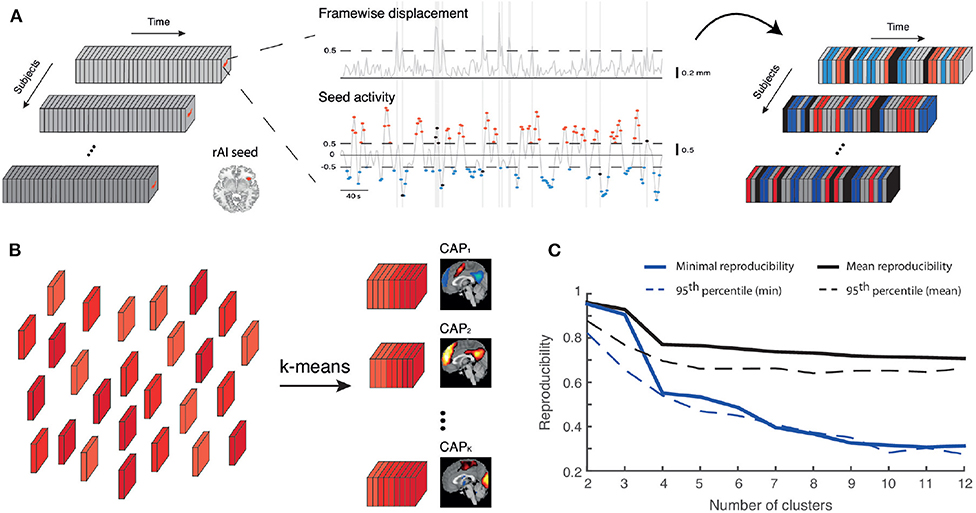
Figure 1. Co-activation pattern analysis. (A) A right anterior insula (rAI) activity time course (middle panel, bottom trace) is computed for each subject, and the frames for which it exceeds a threshold T = 0.5 positively or negatively (respectively red or blue) are tagged. Frames corrupted by a framewise displacement (middle panel, top trace) larger than 0.5 mm are also tagged in black. Here, only non-corrupted activation frames are analyzed. (B) Retained frames across subjects (depicted by different shades of red) undergo k-means clustering to be separated into K different co-activation patterns (CAPs), each the arithmetic mean between a subset of frames denoting one particular set of regions with which the seed was strongly co-active at some time points. (C) Mean (black) and minimal (blue) reproducibility for cluster number ranging from 2 to 12, computed as the average over 100 separate trials (plain lines), and for the same measures, 95th percentiles of the related null distributions (dashed lines). The selected value of K = 6 shows mean and minimal reproducibility values above this null data threshold.
K-means clustering was applied to the retained frames. If C(i, j) is Pearson's spatial correlation coefficient between fMRI volumes i and j, then d(i, j) = 1 − C(i, j) was used as a distance measure. The algorithm was run 20 separate times, to avoid being trapped in a local optimum. Each run was performed with a maximal limit of 100 iterations, using randomly selected data points for initialization. Similarly to previous CAP studies (Liu and Duyn, 2013; Liu et al., 2013), only the 15% most active and 5% most deactive voxels in each fMRI volume were considered for clustering, in order to damp the impact of noisy dimensions on the clustering process, and all remaining non-null clusters with <6 neighboring elements were also discarded to further attenuate noise.
Through k-means clustering, each of the retained frames is assigned to an output cluster from the algorithm. Denoting a vectorized fMRI volume i by Fi, the set of frames assigned to cluster k by , and the number of frames assigned to cluster k by Nk, a CAP is defined as the arithmetic mean between all the frames attributed to the same cluster:
Finally, in order to be able to delineate spatial territories showing significant seed co-(de)activation, each CAP map was individually subjected to spatial z-scoring. A threshold of 1.5 was chosen, in accordance with previous work (Karahanoğlu and Van De Ville, 2015), and also used for visualization (see Figure 2A).
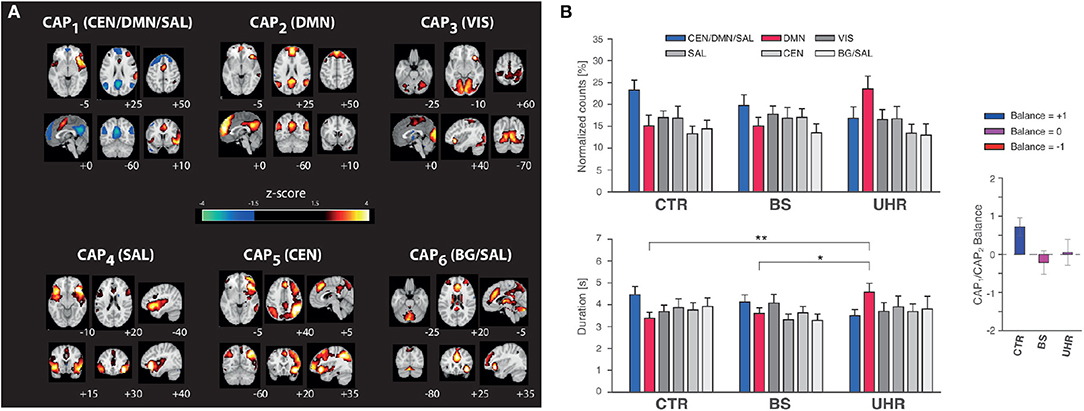
Figure 2. Altered co-activation pattern temporal features in subjects at risk for psychosis. (A) The six co-activation patterns (CAPs) found for a right anterior insula seed, with Montreal Neurological Institute slice coordinates (bottom right of each slice). Left on the figure stands for the left side of the brain. CEN, central executive network; DMN, default mode network; SAL, salience network; VIS, visual network; BG, basal ganglia. (B) Normalized counts (top left graph), duration (bottom left graph), and balance (right graph) metrics for all six co-activation patterns in healthy controls (CTR), subjects with basic symptoms of psychosis (BS), and subjects at ultra-high risk for psychosis (UHR). Normalized counts for CAP1 (p = 0.06) and CAP2 (p = 0.02) trended toward significance for a group difference. Duration trended toward significance for CAP1 (p = 0.06), and was significant for CAP2 (p = 0.002), with post-hoc tests revealing CTR vs. UHR (p = 0.001) and BS vs. UHR (p = 0.02) differences. Balance was significantly different across groups (p < 0.05). Error bars indicate standard error of the means. *p < 0.05, **p < 0.005.
To estimate an optimal cluster number K, we randomly split our subject population into two equally-sized groups 100 times; each time, clustering was performed separately on each half, for K = 2–12. Best-match pairs between each group's CAPs were established (Kuhn, 1955), and lowest/mean correlation values (averaged over the 100 trials) were considered as two reproducibility measures.
To generate reproducibility null distributions, we created 1,000 sets of surrogate data under a stationarity assumption: white noise with the same variance as the real data was added to a subject-specific stationary activity pattern estimated from the first eigenvector of its spatial covariance matrix, and randomly scaled over time. Following temporal z-scoring of this surrogate data, null distributions for both assessed reproducibility measures were generated as described above.
For each CAP, we computed two dynamically informative subject-specific metrics: (1) normalized count, equal to 0/1 if the considered CAP never occurs/occurs every time the right anterior insula seed is active; (2) duration [s] (average time during which the CAP remains active). To assess a possible CAP1/CAP2 competing relationship, a balance metric was also computed for each subject as log2(CAP1 counts/CAP2 counts). Positive/negative values reflect a dominance of CAP1/CAP2.
A one-way multivariate analysis of variance (MANOVA) was conducted, with and without baseline counts/chlorpromazine equivalents as co-variates, to determine group differences in CAP1 and CAP2 normalized counts and duration. We identified one BS univariate outlier, as assessed by inspection of a box plot for values >3 box-lengths from the edge. This subject, who reported intake of relaxane prior to scanning, was taken out of the analysis, because MANOVA is sensitive to the effect of outliers. This resulted in a total of 72 subjects (following scrubbing-related exclusion) matched across groups for handedness, gender, age, and IQ (Table 1).
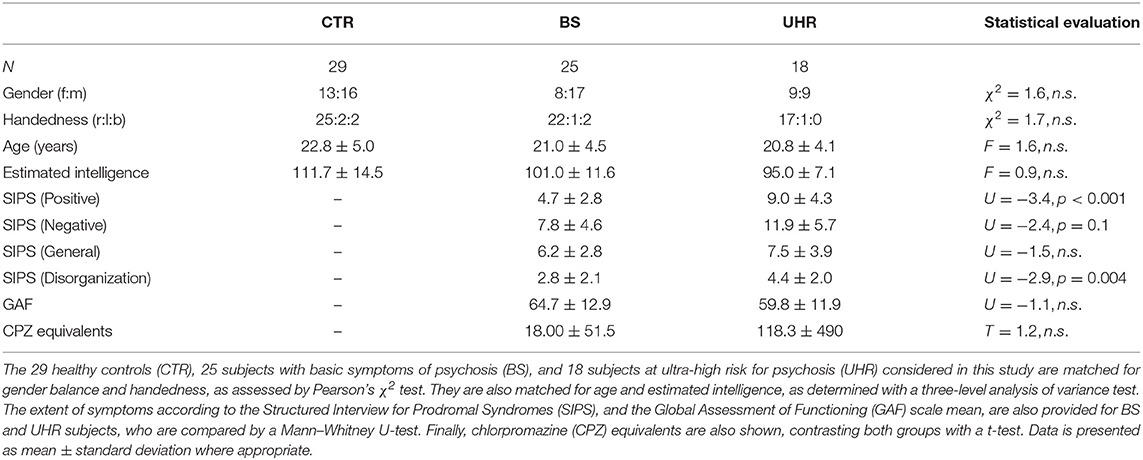
Table 1. Demographic characteristics and rating of symptoms for the analyzed subjects.
Baseline, balance, duration, and counts data were normally distributed (Shapiro-Wilk test, p > 0.05), except for CTR CAP1 duration (p = 0.01) and CAP2 counts (p = 0.02), both positively skewed. This was tolerated, as MANOVA is considered robust to non-normality, with only a small effect on Type I error (Weinfurt, 1995). MANOVA analysis was judged appropriate, as there were no multivariate outliers seen by box plot and Mahalanobis distance (p > 0.001), no multi-collinearity (r < 0.65), and homogeneity of variance-covariance matrices (Box's M test, p > 0.001). Linear relationships were always found as assessed by scatter plot, except for BS CAP1 duration and counts, which may have resulted in loss of power.
The CAP metrics differing significantly between groups were used to determine the relation to clinical symptom sub-scores, as assessed by the Schizophrenia Proneness Interview and Structured Interview for Prodromal Syndromes. We avoid reporting any significance values and thus take into account the issue of circular analysis (Kriegeskorte et al., 2009), as the BS and UHR groups were selected based on such criteria.
Data is presented as mean ± standard error of the mean, unless otherwise stated.
The right anterior insula was subjected to CAP analysis with K = 6 (see Figure 1C) to identify co-activating regions. 20.1% of retained frames were attributed to CAP1, 17.8% to CAP2, 17.0% to CAP3, 16.9% to CAP4, 14.7% to CAP5, and 12.5% to CAP6. CAP1 displayed typical CEN (right dorsolateral prefrontal cortex, posterior parietal cortex) and SAL hubs, as well as negative DMN signal. In contrast, CAP2 predominantly displayed positive signal from DMN areas. CAP3 included a visual network, CAP4 featured SAL areas, CAP5 contained CEN regions, and CAP6 was a mix between SAL and basal ganglia nodes. Due to their frequent occurrence and specific link to the CEN/DMN/SAL relationship, CAP1 and CAP2 dynamic features were further analyzed.
With one-way MANOVA, we found statistically significant group differences on the combined dependent variables (F = 2.2, p = 0.02; Pillai-trace = 2.4; partial η2 = 0.13). Follow-up univariate ANOVAs showed that CAP2 duration (F = 7.2, p = 0.002, partial η2 = 0.18) differed significantly between groups (Bonferroni-adjusted α-level 0.0125), while all other variables trended toward significance (CAP1 duration: F = 2.9, p = 0.06, partial η2 = 0.08; CAP1 counts: F = 2.9, p = 0.06, partial η2 = 0.08; CAP2 counts: F = 4.1, p = 0.02, partial η2 = 0.11).
After adjustment for baseline counts and chlorpromazine equivalents, we similarly found CAP2 duration significance (F = 6.7, p = 0.002, partial η2 = 0.18), while other variables trended toward significance (CAP1 duration: F = 2.8, p = 0.07, partial η2 = 0.08; CAP1 counts: F = 3.2, p = 0.04, partial η2 = 0.09; CAP2 counts: F = 4.1, p < 0.02, partial η2 = 0.12).
Tukey-Kramer post-hoc tests revealed a significant difference in CAP2 duration between CTR (3.4 ± 1.2 s) and UHR (4.6 ± 0.3 s) subjects (p = 0.001), and between BS (3.6 ± 0.2 s) and UHR individuals (p = 0.02), but no difference between CTR and BS groups (p = 0.4; see Figure 2B, bottom left graph).
The CAP1/CAP2 balance metric was positive for CTR (0.72 ± 0.23), negative for BS (−0.22 ± 0.3), and close to zero for UHR subjects (0.05 ± 0.34; see Figure 2B, right graph). A one-way ANOVA revealed statistically significant group differences (F = 3.3, p < 0.05, partial η2 = 0.09), which remained after adjusting for chlorpromazine equivalents (F = 3.4, p < 0.05, partial η2 = 0.09).
CAP2 duration was related to the severity of psychopathological symptoms in at-risk subjects. For the Structured Interview for Prodromal Syndromes ratings, we found associations to disorganization symptoms (ρ = 0.3) such as odd behavior or trouble with focus, and to positive symptoms (ρ = 0.3) such as unusual thought content, persecutory delusions, perceptional abnormalities, or disorganized communication. We also found associations with the Schizophrenia Proneness Interview sub-score for disturbances in experiencing the self and surroundings (ρ = 0.3), including decreased emotional discrimination abilities or increased emotional reactivity in response to routine social interactions.
An aberrant orchestration within the triple network (DMN/CEN/SAL) has been suggested as a backbone for features of various psychiatric disorders (Menon, 2011). Viewing the brain as a dynamic system flexibly adapting to changes in internal and external states (Dixon et al., 2016), we analyzed the dynamic interplay of right anterior insula-driven networks, characterizing and quantifying these states in subjects reporting different at-risk for psychosis criteria.
Our analysis revealed that across intrinsic states, the insular seed activates over the entire resting-state scan, across subjects, in concert with well-known networks: in agreement with prior studies (Nomi et al., 2016), CAP3 and CAP6 displayed visual and basal ganglia networks. Thus, the right anterior insula may be working together with those to coordinate sensory information, with CAP6 supporting the view that dopaminergic paths are involved in insular activity (Menon and Uddin, 2010; Shine et al., 2013). Most frequently, we found that our insular seed fluctuated between a state of antagonistic relationship to the DMN in CAP1, which also included SAL-TPN co-activation, and a state of co-activation with the DMN in CAP2. This is in line with recent dynamic studies (Karahanoğlu and Van De Ville, 2015; Nomi et al., 2016), but contradicts the findings of Ryali et al. (2016), who showed segregation of the SAL from a DMN-CEN state, possibly owing to a different computational approach. In any case, we confirm the pivotal role of the SAL to guarantee the balance between the DMN and CEN by integrating information (Menon, 2011; Nekovarova et al., 2014; Uddin, 2015), as hypothesized by the triple network model, which implies a fundamental neural constraint on cognition, assuming that this constitution of antagonistic network relationships reflects the competition between external and internal information processing (Smallwood et al., 2013).
Due to our specific interest in the networks involving the CEN/DMN/SAL, we focused on the related co-activation patterns, CAP1 and CAP2. UHR subjects exhibited longer durations of excursions in the CAP2 state (right anterior insula/DMN co-activation), and also, with a trend toward significance, larger counts. This is somewhat in line with a report on young subjects with sub-threshold psychosis-spectrum symptoms (Satterthwaite and Baker, 2015), who showed diminished connectivity within the SAL, but enhanced connectivity within the DMN. The right anterior insula has been ascribed a central role for bottom-up processing through interoceptive paths, possibly through the basal ganglia, assisting target brain regions in generating appropriate behavioral responses to salient stimuli via the CEN (Menon and Uddin, 2010), while the DMN is a key structure for awareness and source attribution (Northoff et al., 2006; Leech and Sharp, 2013), and reflects internally guided, self-referential thoughts (Zabelina and Andrews-Hanna, 2016). Thus, the overly potent role of the right anterior insula in engaging with the DMN in UHR may reflect an increasing shift to the internal focus of attention. According to the proximal salience hypothesis, as well as the source monitoring model (Menon, 2011; Damaraju et al., 2014; Robinson et al., 2016), this process might lead to the externalization of thoughts at a later stage of symptom progress and distort the attribution of agency, explaining a number of the positive symptoms of schizophrenia.
The significant group difference seen between BS and UHR in terms of CAP2 duration may relate to the neurodevelopmental etiology of psychosis, as basic symptoms are presumed to characterize the early, and UHR, the late, prodromal phase (Fusar-Poli et al., 2013). Thus, this finding could account for the symptomatic worsening and cognitive decline on the path to a psychotic outbreak, as paralleled by the finding of UHR samples exhibiting more frequent and more pronounced neurocognitive impairments compared to samples with BS symptoms (Schultze-Lutter et al., 2016). Moreover, this network feature then reflects an increased risk toward a psychotic outbreak, rather than a trait or an endophenotype.
Another important difference to draw for those two at-risk groups is further substantiated by the analysis of the balance metric. Here, we show that in contrast to the controls and BS, the UHR do not exhibit a competitive relationship between CAP1 and CAP2. Similarly, children also revealed longer persistence in individual SAL, CEN, and DMN states, and reduced state switching probability (Ryali et al., 2016), pointing to an immature brain network organization in UHR (Satterthwaite and Baker, 2015), perhaps reflecting a state of inflexibility to adapt in a constantly changing environment (Dajani and Uddin, 2015). The loss of CAP1 dominance may be caused by weakened top-down regulation by CEN regions, as supported by the observation of failure of reciprocal influence between the right anterior insula and the dorsolateral prefrontal cortex in schizophrenia (Palaniyappan et al., 2013). Additionally, a takeover of the CEN in favor of voluntary process in a destabilized triple network in hallucinating patients was followed by discontinuation of hallucinations (Lefebvre et al., 2016). Albeit speculative, an initial loss in dominance of CAP1 (frontal and salience-related regions under active inhibition of the DMN) in the early at-risk stage (BS) might be followed by further disintegration of the triple network, as ego disturbances and other related psychotic symptoms (Lebedev et al., 2015) become more advanced in UHR. Future longitudinal studies leveraging the present analytical approach to a finer prospective cohort design will be important to confirm whether a failure in dynamic direct connection, as suggested, is accompanied by worsening of psychotic symptoms (Satterthwaite and Baker, 2015).
The loss of network competition could reflect the risk of a breakdown of separateness of one's internal world and external environment (Carhart-Harris et al., 2013; Nelson et al., 2014). Corroborating this, we found an association of CAP2 duration to positive (psychotic) and disorganized symptoms (such as odd appearance, bizarre thinking, unusual ideas, or attentional troubles), but also to sub-scores for disturbances in experiencing the self and surroundings.
Our findings remained when controlling for baseline right anterior insula activity, demonstrating that they involve the engagement of this seed within different network constellations, rather than mediation of insular activity per se. In addition, as some of the analyzed subjects were medicated and given the vivid debate on the extent to which antipsychotic medication contributes to changes in resting-state fMRI (Sambataro et al., 2010), we also controlled for chlorpromazine equivalents; all observed group differences were still present. While other medications (e.g., antidepressants) may also exert a modulatory influence on resting-state functional properties (McCabe and Mishor, 2011; Arnone et al., 2018), they were taken by only a minor fraction of the analyzed subjects (see Table S1), and thus, this factor also likely played a minimal role on our results.
In future work, a bigger sample will be necessary to learn more about the neurobiological underpinnings of different stages within the disease trajectory. In accordance with this, we did find a trend toward significant group differences in both CAP1 metrics, and also for CAP2 counts (applying a conservative Bonferroni-adjusted significance threshold). Thus, it is unclear whether a bigger sample size would also reveal BS-specific features, as nonlinear relationships for CAP1 duration and counts in this group may have resulted in loss of statistical power.
In summary, our findings substantiate the pivotal role of the right anterior insula in governing transitions between CEN and DMN, which appear dysfunctional prior to the onset of psychosis, especially when first attenuated psychotic symptoms occur. In the UHR stage, this is in at least two ways: firstly, the right anterior insula is longer active in concert with the DMN, and secondly, there is a loss of competition between a SAL/DMN state, and a state with insular/CEN activation paralleled by DMN deactivation, denoting decreased state switching. Considered jointly with our recent stationary analysis of the triple network (Wotruba et al., 2014b), these features suggest that abnormal network switching disrupts one's capacity to distinguish between the internal world and external environment, which is accompanied by inflexibility and an excessive awareness to internal processes (reflected by prolonged expression of the right anterior insula/DMN co-activation pattern). This may, in turn, contribute to the rise in psychotic perceptions. More generally, our work demonstrates the mechanistic relevance of dynamic transitions between temporal network states to support healthy cognition.
The datasets analyzed in this article are not publicly available. Requests to access the datasets should be directed to Thomas Bolton, dGhvbWFzLmJvbHRvbkBlcGZsLmNo.
The studies involving human participants were reviewed and approved by the local ethics committee of Zürich. The patients/participants provided their written informed consent to participate in this study.
TB and DW jointly designed the research, performed the analyses, and wrote the manuscript. RB, AT, LM, SK, WR, and KH were involved in the follow-up of the patients and acquired the analyzed data. DV surveyed the work, contributed to the analyses, and interpretation of the results.
This work was funded by the Bertarelli Foundation (to TB and DV).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphys.2020.00066/full#supplementary-material
Amico, E., Gomez, F., Di Perri, C., Vanhaudenhuyse, A., Lesenfants, D., Boveroux, P., et al. (2014). Posterior cingulate cortex-related co-activation patterns: a resting state fMRI study in propofol-induced loss of consciousness. PLoS ONE 9:e100012. doi: 10.1371/journal.pone.0100012
Andreasen, N. C., Pressler, M., Nopoulos, P., Miller, D., and Ho, B. C. (2010). Antipsychotic dose equivalents and dose-years: a standardized method for comparing exposure to different drugs. Biol. Psychiatry 67, 255–262. doi: 10.1016/j.biopsych.2009.08.040
Andrews-Hanna, J. R. (2012). The brain's default network and its adaptive role in internal mentation. Neuroscientist 18, 251–270. doi: 10.1177/1073858411403316
Arnone, D., Wise, T., Walker, C., Cowen, P. J., Howes, O., and Selvaraj, S. (2018). The effects of serotonin modulation on medial prefrontal connectivity strength and stability: a pharmacological fMRI study with citalopram. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 84, 152–159. doi: 10.1016/j.pnpbp.2018.01.021
Bartsch, R. P., Liu, K. K. L., Bashan, A., and Ivanov, P. C. (2015). Network physiology: how organ systems dynamically interact. PLoS ONE 10:e0142143. doi: 10.1371/journal.pone.0142143
Bashan, A., Bartsch, R. P., Kantelhardt, J. W., Havlin, S., and Ivanov, P. C. (2012). Network physiology reveals relations between network topology and physiological function. Nat. Commun. 3:702. doi: 10.1038/ncomms1705
Behzadi, Y., Restom, K., Liau, J., and Liu, T. (2007). A component based noise correction method (CompCor) for BOLD and perfusion based fMRI. NeuroImage 37, 90–101. doi: 10.1016/j.neuroimage.2007.04.042
Bressler, S. L., and Kelso, J. A. S. (2001). Cortical coordination dynamics and cognition. Trends Cogn. Sci. 5, 26–36. doi: 10.1016/S1364-6613(00)01564-3
Buckner, R. L., Andrews-Hanna, J. R., and Schacter, D. L. (2008). The brain's default network. Ann. N.Y. Acad. Sci. 1124, 1–38. doi: 10.1196/annals.1440.011
Caballero-Gaudes, C., Petridou, N., Francis, S. T., Dryden, I. L., and Gowland, P. A. (2013). Paradigm free mapping with sparse regression automatically detects single-trial functional magnetic resonance imaging blood oxygenation level dependent responses. Hum. Brain Mapp. 34, 501–518. doi: 10.1002/hbm.21452
Carhart-Harris, R. L., and Friston, K. J. (2010). The default-mode, ego-functions and free-energy: a neurobiological account of Freudian ideas. Brain 133, 1265–1283. doi: 10.1093/brain/awq010
Carhart-Harris, R. L., Leech, R., Erritzoe, D., Williams, T. M., Stone, J. M., Evans, J., et al. (2013). Functional connectivity measures after psilocybin inform a novel hypothesis of early psychosis. Schizophr. Bull. 39, 1343–1351. doi: 10.1093/schbul/sbs117
Chai, X. J., Whitfield-Gabrieli, S., Shinn, A. K., Gabrieli, J. D., Nieto Castañón, A., McCarthy, J. M., et al. (2011). Abnormal medial prefrontal cortex resting-state connectivity in bipolar disorder and schizophrenia. Neuropsychopharmacology 36, 2009–2017. doi: 10.1038/npp.2011.88
Chang, C., and Glover, G. H. (2010). Time-frequency dynamics of resting-state brain connectivity measured with fMRI. Neuroimage 50, 81–98. doi: 10.1016/j.neuroimage.2009.12.011
Chen, J. E., Chang, C., Greicius, M. D., and Glover, G. H. (2015). Introducing co-activation pattern metrics to quantify spontaneous brain network dynamics. Neuroimage 111, 476–488. doi: 10.1016/j.neuroimage.2015.01.057
Ciria, L. F., Luque-Casado, A., Sanabria, D., Holgado, D., Ivanov, P. C., and Perakakis, P. (2019). Oscillatory brain activity during acute exercise: tonic and transient neural response to an oddball task. Psychophysiology 56:e13326. doi: 10.1111/psyp.13326
Dajani, D. R., and Uddin, L. Q. (2015). Demystifying cognitive flexibility: implications for clinical and developmental neuroscience. Trends Neurosci. 38, 571–578. doi: 10.1016/j.tins.2015.07.003
Damaraju, E., Allen, E. A., Belger, A., Ford, J. M., McEwen, S., Mathalon, D. H., et al. (2014). Dynamic functional connectivity analysis reveals transient states of dysconnectivity in schizophrenia. Neuroimage 5, 298–308. doi: 10.1016/j.nicl.2014.07.003
de Pasquale, F., Corbetta, M., Betti, V., and Della Penna, S. (2018). Cortical cores in network dynamics. Neuroimage 180, 370–382. doi: 10.1016/j.neuroimage.2017.09.063
Dixon, M. L., Andrews-Hanna, J. R., Spreng, R. N., Irving, Z. C., Mills, C., Girn, M., et al. (2016). Interactions between the default network and dorsal attention network vary across default subsystems, time, and cognitive states. Neuroimage 147, 632–649. doi: 10.1016/j.neuroimage.2016.12.073
Du, Y., Pearlson, G. D., Yu, Q., He, H., Lin, D., Sui, J., et al. (2016). Interaction among subsystems within default mode network diminished in schizophrenia patients: a dynamic connectivity approach. Schizophr. Res. 170, 55–65. doi: 10.1016/j.schres.2015.11.021
Ebisch, S., Salone, A., Ferri, F., De Berardis, D., Romani, G.-L., Ferro, F. M., et al. (2013). Out of touch with reality? Social perception in first-episode schizophrenia. Soc. Cogn. Affect. Neurosci. 8, 394–403. doi: 10.1093/scan/nss012
Ebisch, S. J. H., Mantini, D., Northoff, G., Salone, A., De Berardis, D., Ferri, F., et al. (2014). Altered brain long-range functional interactions underlying the link between aberrant self-experience and self-other relationship in first-episode schizophrenia. Schizophr. Bull. 40, 1072–1082. doi: 10.1093/schbul/sbt153
Fox, M. D., Snyder, A. Z., Vincent, J. L., Corbetta, M., Van Essen, D. C., and Raichle, M. E. (2005). The human brain is intrinsically organized into dynamic, anticorrelated functional networks. Proc. Natl. Acad. Sci. U.S.A. 102, 9673–9678. doi: 10.1073/pnas.0504136102
Fransson, P. (2005). Spontaneous low-frequency BOLD signal fluctuations: an fMRI investigation of the resting-state default mode of brain function hypothesis. Hum. Brain Mapp. 26, 15–29. doi: 10.1002/hbm.20113
Fusar-Poli, P., Borgwardt, S., Bechdolf, A., Addington, J., Riecher-Rössler, A., Schultze-Lutter, F., et al. (2013). The psychosis high-risk state: a comprehensive state-of-the-art review. JAMA Psychiatry 70, 107–120. doi: 10.1001/jamapsychiatry.2013.269
Fux, L., Walger, P., Schimmelmann, B., and Schultze-Lutter, F. (2013). The schizophrenia proneness instrument, child and youth version (SPI-CY): practicability and discriminative validity. Schizophr. Res. 146, 69–78. doi: 10.1016/j.schres.2013.02.014
Goldberger, A. L., Amaral, L. A. N., Hausdorff, J. M., Ivanov, P. C., Peng, C., and Stanley, H. E. (2002). Fractal dynamics in physiology: alterations with disease and aging. Proc. Natl. Acad. Sci. U.S.A. 99, 2466–2472. doi: 10.1073/pnas.012579499
Harrisberger, F., Buechler, R., Smieskova, R., Lenz, C., Walter, A., Egloff, L., et al. (2016). Alterations in the hippocampus and talamus in individuals at high risk for psychosis. NPJ Schizophr. 2:16033. doi: 10.1038/npjschz.2016.33
Hurlemann, R., Jessen, F., Wagner, M., Frommann, I., Ruhrmann, S., Brockhaus, A., et al. (2008). Interrelated neuropsychological and anatomical evidence of hippocampal pathology in the at-risk mental state. Psychol. Med. 38, 843–851. doi: 10.1017/S0033291708003279
Hutchison, R. M., Womelsdorf, T., Allen, E. A., Bandettini, P. A., Calhoun, V. D., Corbetta, M., et al. (2013). Dynamic functional connectivity: Promise, issues, and interpretations. NeuroImage 80, 360–378. doi: 10.1016/j.neuroimage.2013.05.079
Ivanov, P. C., and Bartsch, R. P. (2014). “Network physiology: mapping interactions between networks of physiologic networks,” in Networks of Networks: The Last Frontier of Complexity, eds G. D'Agostino and A. Scala (Berlin: Springer), 203–222.
Ivanov, P. C., Liu, K. K. L., and Bartsch, R. P. (2016). Focus on the emerging new fields of network physiology and network medicine. New J. Phys. 18:100201. doi: 10.1088/1367-2630/18/10/100201
Karahanoğlu, F. I., and Van De Ville, D. (2015). Transient brain activity disentangles fMRI resting-state dynamics in terms of spatially and temporally overlapping networks. Nat. Commun. 6:7751. doi: 10.1038/ncomms8751
Karahanoğlu, F. I., and Van De Ville, D. (2017). Dynamics of large-scale fMRI networks: Deconstruct brain activity to build better models of brain function. Curr. Opin. Biomed. Eng. 3, 28–36. doi: 10.1016/j.cobme.2017.09.008
Klosterkötter, J., Hellmich, M., Steinmeyer, E., and Schultze-Lutter, F. (2001). Diagnosing schizophrenia in the initial prodromal phase. Arch. Gen. Psychiatry 58, 158–164. doi: 10.1001/archpsyc.58.2.158
Koutsouleris, N., Meisenzahl, E. M., Davatzikos, C., Bottlender, R., Frodl, T., Scheuerecker, J., et al. (2009a). Use of neuroanatomical pattern classification to identify subjects in at-risk mental states of psychosis and predict disease transition. Arch. Gen. Psychiatry 66, 700–712. doi: 10.1001/archgenpsychiatry.2009.62
Koutsouleris, N., Schmitt, G., Gaser, C., Bottlender, R., Scheuerecker, J., McGuire, P., et al. (2009b). Neuroanatomical correlates of different vulnerability states for psychosis and their clinical outcomes. Br. J. Psychiatry 195, 218–226. doi: 10.1192/bjp.bp.108.052068
Kriegeskorte, N., Simmons, W. K., Bellgowan, P. S. F., and Baker, C. I. (2009). Circular analysis in systems neuroscience: the dangers of double dipping. Nat. Neurosci. 12, 535–540. doi: 10.1038/nn.2303
Kuhn, H. W. (1955). The hungarian method for the assignment problem. Naval Res. Logist. Q. 2, 83–97. doi: 10.1002/nav.3800020109
Lebedev, A. V., Lövdén, M., Rosenthal, G., Feilding, A., Nutt, D. J., and Carhart-Harris, R. L. (2015). Finding the self by losing the self: neural correlates of ego-dissolution under psilocybin. Hum. Brain Mapp. 36, 3137–3153. doi: 10.1002/hbm.22833
Leech, R., and Sharp, D. J. (2013). The role of the posterior cingulate cortex in cognition and disease. Brain 137, 12–32. doi: 10.1093/brain/awt162
Lefebvre, S., Demeulemeester, M., Leroy, A., Delmaire, C., Lopes, R., Pins, D., et al. (2016). Network dynamics during the different stages of hallucinations in schizophrenia. Hum. Brain Mapp. 37, 2571–2586. doi: 10.1002/hbm.23197
Lin, A., Liu, K. K. L., Bartsch, R. P., and Ivanov, P. C. (2016). Delay-correlation landscape reveals characteristic time delays of brain rhythms and heart interactions. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 374:20150182. doi: 10.1098/rsta.2015.0182
Liu, K. K. L., Bartsch, R. P., Lin, A., Mantegna, R. N., and Ivanov, P. C. (2015). Plasticity of brain wave network interactions and evolution across physiologic states. Front. Neural Circ. 9:62. doi: 10.3389/fncir.2015.00062
Liu, X., Chang, C., and Duyn, J. H. (2013). Decomposition of spontaneous brain activity into distinct fMRI co-activation patterns. Front. Syst. Neurosci. 7:101. doi: 10.3389/fnsys.2013.00101
Liu, X., and Duyn, J. H. (2013). Time-varying functional network information extracted from brief instances of spontaneous brain activity. Proc. Natl. Acad. Sci. U.S.A. 110, 4392–4397. doi: 10.1073/pnas.1216856110
McCabe, C., and Mishor, Z. (2011). Antidepressant medications reduce subcortical–cortical resting-state functional connectivity in healthy volunteers. Neuroimage 57, 1317–1323. doi: 10.1016/j.neuroimage.2011.05.051
Menon, V. (2011). Large-scale brain networks and psychopathology: a unifying triple network model. Trends Cogn. Sci. 15, 483–506. doi: 10.1016/j.tics.2011.08.003
Menon, V., and Uddin, L. Q. (2010). Saliency, switching, attention and control: a network model of insula function. Brain Struct. Funct. 214, 655–667. doi: 10.1007/s00429-010-0262-0
Miller, R. L., Yaesoubi, M., Turner, J. A., Mathalon, D., Preda, A., Pearlson, G., et al. (2016). Higher dimensional meta-state analysis reveals reduced resting fMRI connectivity dynamism in schizophrenia patients. PLoS ONE 11:0149849. doi: 10.1371/journal.pone.0149849
Miller, T. J., McGlashan, T. H., Rosen, J. L., Cadenhead, K., Cannon, T., Ventura, J., et al. (2003). Prodromal assessment with the structured interview for prodromal syndromes and the scale of prodromal symptoms: predictive validity, interrater reliability, and training to reliability. Schizophr. Bull. 29, 703–715. doi: 10.1093/oxfordjournals.schbul.a007040
Nekovarova, T., Fajnerova, I., Horacek, J., and Spaniel, F. (2014). Bridging disparate symptoms of schizophrenia: a triple network dysfunction theory. Front. Behav. Neurosci. 8:171. doi: 10.3389/fnbeh.2014.00171
Nelson, B., Whitford, T., Lavoie, S., and Sass, L. (2014). What are the neurocognitive correlates of basic self-disturbance in schizophrenia? Integrating phenomenology and neurocognition: part 2 (aberrant salience). Schizophr. Res. 152, 20–27. doi: 10.1016/j.schres.2013.06.033
Nomi, J. S., Farrant, K., Damaraju, E., Rachakonda, S., Calhoun, V. D., and Uddin, L. Q. (2016). Dynamic functional network connectivity reveals unique and overlapping profiles of insula subdivisions. Hum. Brain Mapp. 37, 1770–1787. doi: 10.1002/hbm.23135
Northoff, G., Heinzel, A., de Greck, M., Bermpohl, F., Dobrowolny, H., and Panksepp, J. (2006). Self-referential processing in our brain–a meta-analysis of imaging studies on the self. NeuroImage 31, 440–457. doi: 10.1016/j.neuroimage.2005.12.002
Oldfield, R. C. (1971). The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia 9, 97–113. doi: 10.1016/0028-3932(71)90067-4
O'Neill, G. C., Tewarie, P., Vidaurre, D., Liuzzi, L., Woolrich, M. W., and Brookes, M. J. (2018). Dynamics of large–scale electrophysiological networks: a technical review. Neuroimage 180, 559–576. doi: 10.1016/j.neuroimage.2017.10.003
Palaniyappan, L., Balain, V., and Liddle, P. F. (2012a). The neuroanatomy of psychotic diathesis: a meta-analytic review. J. Psychiatr. Res. 46, 1249–1256. doi: 10.1016/j.jpsychires.2012.06.007
Palaniyappan, L., and Liddle, P. F. (2012). Does the salience network play a cardinal role in psychosis? An emerging hypothesis of insular dysfunction. J. Psychiatry Neurosci. 37, 17–27. doi: 10.1503/jpn.100176
Palaniyappan, L., Simmonite, M., White, T., Liddle, E., and Liddle, P. (2013). Neural primacy of the salience processing system in schizophrenia. Neuron 79, 814–828. doi: 10.1016/j.neuron.2013.06.027
Palaniyappan, L., White, T. P., and Liddle, P. F. (2012b). The concept of salience network dysfunction in schizophrenia: from neuroimaging observations to therapeutic opportunities. Curr. Top. Med. Chem. 12, 2324–2338. doi: 10.2174/156802612805289881
Power, J. D., Barnes, K. A., Snyder, A. Z., Schlaggar, B. L., and Petersen, S. E. (2012). Spurious but systematic correlations in functional connectivity MRI networks arise from subject motion. Neuroimage 59, 2142–2154. doi: 10.1016/j.neuroimage.2011.10.018
Preti, M. G., Bolton, T. A. W., and Van De Ville, D. (2017). The dynamic functional connectome: state-of-the-art and perspectives. Neuroimage 160, 41–54. doi: 10.1016/j.neuroimage.2016.12.061
Pu, W., Li, L., Zhang, H., Ouyang, X., Liu, H., Zhao, J., et al. (2012). Morphological and functional abnormalities of salience network in the early-stage of paranoid schizophrenia. Schizophr. Res. 141, 15–21. doi: 10.1016/j.schres.2012.07.017
Robinson, J. D., Wagner, N., and Northoff, G. (2016). Is the sense of agency in schizophrenia influenced by resting-state variation in self-referential regions of the brain? Schizophr. Bull. 42, 270–276. doi: 10.1093/schbul/sbv102
Ryali, S., Supekar, K., Chen, T., Kochalka, J., Cai, W., Nicholas, J., et al. (2016). Temporal dynamics and developmental maturation of salience, default and central-executive network interactions revealed by variational Bayes hidden Markov modeling. PLoS Comput. Biol. 12:1005138. doi: 10.1371/journal.pcbi.1005138
Sambataro, F., Blasi, G., Fazio, L., Caforio, G., Taurisano, P., Romano, R., et al. (2010). Treatment with olanzapine is associated with modulation of the default mode network in patients with schizophrenia. Neuropsychopharmacology 35, 904–912. doi: 10.1038/npp.2009.192
Satterthwaite, T. D., and Baker, J. T. (2015). How can studies of resting-state functional connectivity help us understand psychosis as a disorder of brain development? Curr. Opin. Neurobiol. 30, 85–91. doi: 10.1016/j.conb.2014.10.005
Schultze-Lutter, F., Addington, J., Ruhrmann, S., and Klosterkötter, J. (2007). Schizophrenia Proneness Instrument, Adult Version (Spi-A). Rome: Giovanni Fiority Editore.
Schultze-Lutter, F., Debbané, M., Theodoridou, A., Wood, S. J., Raballo, A., Michel, C., et al. (2016). Revisiting the basic symptom concept: toward translating risk symptoms for psychosis into neurobiological targets. Front. Psychiatry 7:9. doi: 10.3389/fpsyt.2016.00009
Seeley, W. W., Menon, V., Schatzberg, A. F., Keller, J., Glover, G. H., Kenna, H., et al. (2007). Dissociable intrinsic connectivity networks for salience processing and executive control. J. Neurosci. 27, 2349–2356. doi: 10.1523/JNEUROSCI.5587-06.2007
Sheehan, D. V., Lecrubier, Y., Sheehan, K. H., Amorim, P., Janavs, J., Weiller, E., et al. (1998). The mini-international neuropsychiatric interview (M.I.N.I.): the development and validation of a structured diagnostic psychiatric interview for DSM-IV and ICD-10. J. Clin. Psychiatry 59, 22–33.
Shine, J. M., Matar, E., Ward, P. B., Frank, M. J., Moustafa, A. A., Pearson, M., et al. (2013). Freezing of gait in Parkinson's disease is associated with functional decoupling between the cognitive control network and the basal ganglia. Brain 136, 3671–3681. doi: 10.1093/brain/awt272
Smallwood, J., Tipper, C., Brown, K., Baird, B., Engen, H., Michaels, J. R., et al. (2013). Escaping the here and now: evidence for a role of the default mode network in perceptually decoupled thought. NeuroImage 69, 120–125. doi: 10.1016/j.neuroimage.2012.12.012
Theodoridou, A., Heekeren, K., Dvorsky, D., Metzler, S., Franscini, M., Haker, H., et al. (2014). Early recognition of high risk of bipolar disorder and psychosis: an overview of the zinep “early recognition” study. Front. Public Health 2:166. doi: 10.3389/fpubh.2014.00166
Uddin, L. Q. (2015). Salience processing and insular cortical function and dysfunction. Nat. Rev. Neurosci. 16, 55–61. doi: 10.1038/nrn3857
Van De Ville, D., Britz, J., and Michel, C. M. (2010). EEG microstate sequences in healthy humans at rest reveal scale-free dynamics. Proc. Natl. Acad. Sci. U.S.A. 107, 18179–18184. doi: 10.1073/pnas.1007841107
Van Den Heuvel, M. P., Stam, C. J., Boersma, M., and Pol, H. E. H. (2008). Small-world and scale-free organization of voxel-based resting-state functional connectivity in the human brain. Neuroimage 43, 528–539. doi: 10.1016/j.neuroimage.2008.08.010
Weinfurt, K. P. (1995). “Multivariate analysis of variance,” in Reading and Understanding Multivariate Statistics, eds L. G. Grimm and P. R. Yarnolds (Washington, DC: American Psychological Association).
Whitfield-Gabrieli, S., and Ford, J. (2012). Default mode network activity and connectivity in psychopathology. Annu. Rev. Clin. Psychol. 8, 49–76. doi: 10.1146/annurev-clinpsy-032511-143049
Whitfield-Gabrieli, S., Thermenos, H. W., Milanovic, S., Tsuang, M. T., Faraone, S. V., McCarley, R. W., et al. (2009). Hyperactivity and hyperconnectivity of the default network in schizophrenia and in first-degree relatives of persons with schizophrenia. Proc. Natl. Acad. Sci. U.S.A. 106, 1279–1284. doi: 10.1073/pnas.0809141106
Woodward, N. D., Rogers, B., and Heckers, S. (2011). Functional resting-state networks are differentially affected in schizophrenia. Schizophr. Res. 130, 86–93. doi: 10.1016/j.schres.2011.03.010
Wotruba, D., Heekeren, K., Michels, L., Buechler, R., Simon, J. J., Theodoridou, A., et al. (2014a). Symptom dimensions are associated with reward processing in unmedicated persons at risk for psychosis. Front. Behav. Neurosci. 8:382. doi: 10.3389/fnbeh.2014.00382
Wotruba, D., Michels, L., Buechler, R., Metzler, S., Theodoridou, A., Gerstenberg, M., et al. (2014b). Aberrant coupling within and across the default mode, task-positive, and salience network in subjects at risk for psychosis. Schizophr. Bull. 40, 1095–1104.
Yung, A. R., and Mcgorry, P. D. (2007). Prediction of psychosis: setting the stage. Br. J. Psychiatry 191, 1–8. doi: 10.1192/bjp.191.51.s1
Keywords: pre-psychotic, co-activation patterns, functional magnetic resonance imaging—fMRI, dynamic functional connectivity, default mode network (DMN), central executive network (CEN), salience network
Citation: Bolton TAW, Wotruba D, Buechler R, Theodoridou A, Michels L, Kollias S, Rössler W, Heekeren K and Van De Ville D (2020) Triple Network Model Dynamically Revisited: Lower Salience Network State Switching in Pre-psychosis. Front. Physiol. 11:66. doi: 10.3389/fphys.2020.00066
Received: 10 October 2019; Accepted: 21 January 2020;
Published: 11 February 2020.
Edited by:
Plamen Ch. Ivanov, Boston University, United StatesReviewed by:
Rossella Rizzo, University of Calabria, ItalyCopyright © 2020 Bolton, Wotruba, Buechler, Theodoridou, Michels, Kollias, Rössler, Heekeren and Van De Ville. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Thomas A. W. Bolton, dGhvbWFzLmJvbHRvbkBlcGZsLmNo
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.