
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Physiol., 18 September 2019
Sec. Integrative Physiology
Volume 10 - 2019 | https://doi.org/10.3389/fphys.2019.01168
This article is part of the Research TopicInterventional Strategies for Enhancing Quality of Life and Health Span in Older AdultsView all 17 articles
 Jeanne Brenda Martins1,2
Jeanne Brenda Martins1,2 Vanessa Amaral Mendonça1,2,3
Vanessa Amaral Mendonça1,2,3 Grazielle Cordeiro Aguiar4
Grazielle Cordeiro Aguiar4 Sueli Ferreira da Fonseca1,2
Sueli Ferreira da Fonseca1,2 Jousielle Márcia dos Santos1,3
Jousielle Márcia dos Santos1,3 Rosalina Tossige-Gomes1,2
Rosalina Tossige-Gomes1,2 Dirceu de Sousa Melo1,2
Dirceu de Sousa Melo1,2 Murilo Xavier Oliveira1,3
Murilo Xavier Oliveira1,3 Hércules Ribeiro Leite1,2
Hércules Ribeiro Leite1,2 Ana Cristina Resende Camargos5
Ana Cristina Resende Camargos5 Anderson José Ferreira4
Anderson José Ferreira4 Cândido Celso Coimbra4
Cândido Celso Coimbra4 Jacques Poortmans6
Jacques Poortmans6 Vinícius Cunha Oliveira1,3
Vinícius Cunha Oliveira1,3 Sara Barros Silva1,3
Sara Barros Silva1,3 Talita Emanuela Domingues1,2
Talita Emanuela Domingues1,2 Mário Bernardo-Filho7
Mário Bernardo-Filho7 Ana Cristina Rodrigues Lacerda1,2,3*
Ana Cristina Rodrigues Lacerda1,2,3*Background: Knee osteoarthritis (kOA) is a common chronic disease that induces changes in redox status and inflammatory biomarkers, cell death, and motor impairment. Aerobic training can be a non-pharmacological alternative to prevent the progression of the disease.
Objective: To evaluate the effects of an 8 weeks moderate-intensity treadmill aerobic training program on redox status and inflammatory biomarkers and motor performance in kOA-like changes induced by monosodium iodoacetate (MIA) in rats.
Methods: Twenty-seven rats were randomly divided into three groups: SHAM; induced kOA (OA); and induced kOA + aerobic training (OAE). Motor performance was evaluated by the number of falls on rotarod test, the total time of displacement and the number of failures on a 100 cm footbridge. Data for cytokines and histology were investigated locally, whereas plasma was used for redox status biomarkers.
Results: The OA group, compared to the SHAM group, increased 1.13 times the total time of displacement, 6.05 times the number of failures, 2.40 times the number of falls. There was also an increase in cytokine and in thiobarbituric acid reactive substances (TBARS) (IL1β: 5.55-fold, TNF: 2.84-fold, IL10: 1.27-fold, IL6: 1.50-fold, TBARS: 1.14-fold), and a reduction of 6.83% in the total antioxidant capacity (FRAP), and of 35% in the number of chondrocytes. The aerobic training improved the motor performance in all joint function tests matching to SHAM scores. Also, it reduced inflammatory biomarkers and TBARS level at values close to those of the SHAM group, with no change in FRAP level. The number of falls was explained by IL1β and TNF (58%), and the number of failures and the total time of displacement were also explained by TNF (29 and 21%, respectively).
Conclusion: All findings indicate the efficacy of moderate-intensity aerobic training to regulate inflammatory biomarkers associated with improved motor performance in induced kOA-like changes, thus preventing the loss of chondrocytes.
Chondrocytes are responsible for tissue maintenance which impact on joint function and performance (Sophia Fox et al., 2009; Akkiraju and Nohe, 2015; Charlier et al., 2016). Current literature has reported exercise-induced chondroprotection in knee osteoarthritis (kOA) (Loeser, 2010; Golightly et al., 2012; Geneen et al., 2017). A potential explanation is the mechanical signal transduction (O’Hara et al., 1990; Urban, 1994), preserving the cartilage proteoglycans, and promoting chondrocytes modulation (Little and Ghosh, 1997; Loeser et al., 2012). Low-magnitude mechanical stress seems to suppress the pathway of interleukin-1 beta (IL1β) and tumor necrosis factor (TNF) release (Roman-Blas et al., 2009; Leong et al., 2010; Li et al., 2013; Assis et al., 2016). The IL1β and TNF are inflammatory mediators involved in joint degeneration caused by kOA (Blasioli and Kaplan, 2013; Mabey and Honsawek, 2015). The modulation of these cytokines in the joint would regulate the synthesis of proteoglycans and collagen, thereby attenuating the swelling process (Knobloch et al., 2008; Iijima et al., 2015).
The literature points out redox imbalance and the related increase on reactive oxygen species (ROS), swelling process and necrosis of chondrocytes as the pathophysiology of kOA (Altindag et al., 2007; Rose et al., 2012; Hui et al., 2016). Moreover, redox imbalance and the increased inflammatory biomarkers may cause cartilage damage, neuroinflammatory disease progression, and joint disability (Kim et al., 2010; Watari et al., 2011; Attur et al., 2013, 2015; Reed et al., 2014).
Several interventions are used to improve motor performance in patients with kOA (Golightly et al., 2012; Geneen et al., 2017), and a low-cost aerobic training may be an alternative (Hunter and Eckstein, 2009; Semanik et al., 2012). Aerobic training improves blood soluble TNF receptors level and brain-derived neurotrophic factor (BDNF) plasma level in people with kOA (Gomes et al., 2012; Simão et al., 2014). Moreover, BDNF seems to impact on chondrocyte differentiation, changing it from proliferative to differentiation program (Hutchison, 2012).
Beyond insufficient information about the modulatory effect of the aerobic training in the knee joint degeneration parameters and possible relationships between joint parameters and motor performance, the literature still presents gaps regarding the effect of aerobic training in the joint preservation on kOA-like changes. Many studies detail mechanotransduction mechanisms but still remain questions about joint function. Moreover, few studies address joint function tests to establish a new therapeutic approach in clinical practice. The current study aimed to investigate the effects of moderate-intensity aerobic training in the inflammatory and redox biomarkers modulation of kOA-like changes, and its possible link to motor performance in rats. Therefore, because experimental kOA induces joint swelling process, we hypothesized that moderate-intensity aerobic training would attenuate the swelling process, favoring the redox balance and preserving chondrocytes in rats with kOA -like changes. Improved modulation in joint biomarkers levels might explain an effect on motor performance.
Male Wistar rats were used in the current study that was part of a Masters in Physiology at the Universidade Federal dos Vales do Jequitinhonha e Mucuri (Martins, 2017). The project was approved by the local Ethics Committee (protocol 005/2015).
Twenty-seven rats were randomized into three groups: sham group (SHAM), n = 9; induced kOA group (OA), n = 9; and induced kOA + aerobic training (OAE), n = 9. The rats had available water and food (i.e., Nuvilab CR1, Nuvital Nutrientes S/A, Brazil) as they desire, and their environment was controlled (i.e., humidity of 60% and temperature of 22°C).
Twelve-week-old male Wistar rats were anesthetized with ketamine (80 mg/kg, i.p.) and xylazine (15 mg/kg, i.p.). Then, kOA was induced on the right knee joint at 90° flexion by direct infiltration of monosodium iodoacetate (MIA) (1.2 mg diluted in 50 μL saline solution). We used a 29G X 1/2 BD Ultra-FineTM insulin needle (Guzman et al., 2003; Cifuentes et al., 2010; Takahashi et al., 2018). The SHAM group received an infiltration containing 50 μL of saline solution (0.9% NaCl).
To perform the aerobic training, a motorized treadmill (Insight®, SP, Ribeirão Preto, Brazil) with six individual lanes and with no inclination was used in the study. OAE group started the familiarization to the treadmill (10 min/day for 5 days) 24 h after the kOA-induction procedure. Then, 24 h after the familiarization period, initiated the training program, which consisted of treadmill running at the velocity of 16 m/min, 3 days per week during 8 weeks (Cifuentes et al., 2010), and the duration of running sessions was increased from 30 to 50 min at the fourth week. The workload of all groups was analyzed at the end of the training program during a treadmill incremental test (initial speed of 10 m min–1, 1% slope, with no electrical stimulus, an increase of 2 m min–1 every 3 min) (Balthazar et al., 2009; Primola-Gomes et al., 2009). The workload (W; J) was calculated as: W = body weight (kg) × total time to fatigue (min) × treadmill speed (m min–1) × sin θ (treadmill inclination) × 10 (Lacerda et al., 2006).
In the rotarod test (Scienlabor, Brazil), the rats are stimulated to walk around a circle drum surface. The rotarod test measures balance, coordination, physical performance, and motor-planning by calculating the number of falls during determined speed. The time that a given rat stays on this circle rod represents the joint function. We used a protocol adapted from Piel et al. (2014) to quantify the number of falls of rats during 3-min period keeping a fixed speed of 8 revolutions per minute (rpm) (Piel et al., 2014).
Rats locomoted on a footbridge to investigate joint function when moving a short distance. The footbridge had a length of 100 cm delimited by 3 mm thick aluminum filets. Image and time to complete a single pass during animals’ locomotion were recorded. Records were later analyzed by a blinded investigator to quantify the total time of displacement and the number of times each rat stepped out of the space between the filets (number of failures).
Animals were euthanized individually by decapitation and their right knee joints were analyzed. Approximately 12 mL of blood was collected in tubes containing ethylenediaminetetraacetic acid (EDTA), then centrifuged at 500 × g for 10 min and the serum aliquoted and frozen in a −80°C freezer for further analysis.
For later analysis, the joint lavage (JAL) supernatants were stored at −80°C. Immediately after recovering JAL, we removed the joint capsule and stored it in a freezer at −80°C. Moreover, we homogenized the sample in phosphate buffer and also frozen it at −80°C for future analysis. IL1β, TNF, and interleukin-10 (IL10) knee joint biomarkers were analyzed according to the manufacturer’s instructions by ELISA kits (DuoSet, R&D Systems, United States).
The capsule was placed into a beaker with 750 μL of cytokine extraction solution, and a tissue homogenizer (Tecnal, TE-103) was used to obtain the macerate. Then, the capsule macerate was processed at 8 rpm speed for 2 min, the volume centrifuged at 3500 xg at 4°C for 10 min and stored in a freezer. BDNF and interleukin-6 (IL6) levels were analyzed according to the manufacturer’s instructions by ELISA.
The reaction of the thiobarbituric acid with malondialdehyde (MDA) was used to determine lipid peroxidation by thiobarbituric acid reactive substances (TBARS) plasma levels (Ohkawa et al., 1979). The ferric reducing ability of plasma (FRAP), i.e., the reduction of ferric-tripyridyltriazine [Fe(III)-TPTZ] complex to ferrous-tripyridyltriazine [Fe(II)-TPTZ] (Benzie and Strain, 1996) was used to determine the total antioxidant capacity. The Bradford method using bovine serum albumin was used as a standard to determine the samples protein levels (Bradford, 1976).
The right knee joints were placed in 4% neutral-buffered formalin for 24 h. After that, tissues were placed in 10% EDTA at pH 7.4 for decalcification (Jimson et al., 2014). Sagittal sections were prepared from knee joints. Two slides of the femur compromised by MIA induction or SHAM were prepared for histological analyses. Three cuts in each slide. Tissues were placed in formalin, dehydrated in a graded series of ethanol and xylol, embedded in paraffin, cut into 6 μm serial sections, and stained with hematoxylin-eosin.
To investigate the number of active cells, the middle third of the joint was used. For cell counting, the nuclei stained by hematoxylin present in the superficial, intermediate and transitional areas were considered. The analysis was performed on the articular facet of the right femur. The image was captured by a microscope with a 40 × magnification. The software Image J was used for cell counting. Two micrographs were taken in series and a blinded investigator analyzed them on different days (ICC = 0.99). For statistical analysis, the average number of cells was established and used.
We used the SPSS statistical package, version 22.0 (Inc., United States) and Graph Pad Prism, version 5.0 (Inc., United States). Data are expressed as mean ± standard error (S.E.M). Normality of data was assessed using the Shapiro–Wilk test. For comparisons, we used the one-way ANOVA with Tukey’s post hoc tests for parametric data (Body mass, IL6 and number of chondrocytes) and Kruskal-Wallis with Dunn’s post hoc test for non-parametric data (IL1β, TNF, IL10, TBARS, FRAP, number of falls, total time of displacement, and number of failures). Effect Size (d) was checked in G∗Power 3.1.9.2 program. Effect size conventions for test family (F tests) and one-way ANOVA: d = 0.10 (small); d = 0.25 (medium); d = 0.40 (large). The Spearman correlation investigated associations between two intra-articular biomarkers of joint damage (IL1β, TNF) and joint function tests: number of falls, total time of displacement and number of failures.
To determine intra-examiner reliability for the evaluated outcomes, the intraclass correlation coefficient (ICC) adopting a 95% confidence interval was determined. Multiple linear stepwise regression models confirmed the association between selected biomarkers and joint function, adjusting by Bonferroni at α = 0.017. Graphs were built using the GraphPad Prism 5 (GraphPad Software Inc., San Diego, CA, United States).
Twenty-seven animals were available for this study, recovered uneventfully from surgery and exercise procedure. By the end of the study, the rats were about 6 months old. No significant differences were observed in body weight between the three groups at the surgery and at the 8th-week post-surgery (Figure 1).
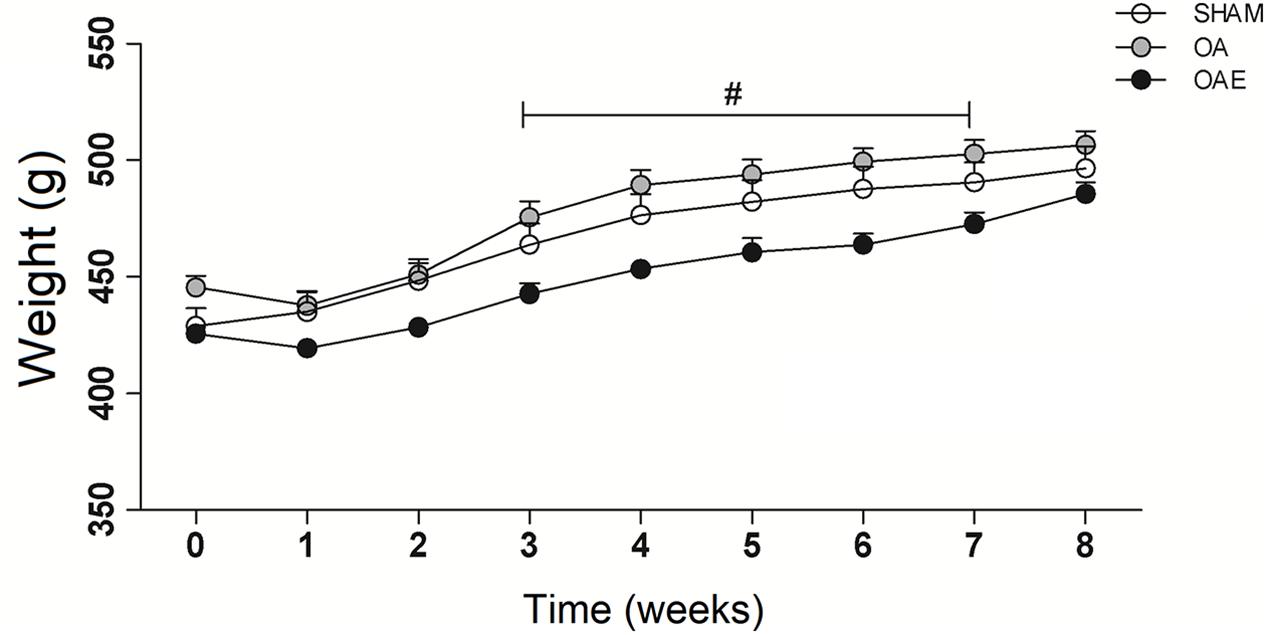
Figure 1. Body mass over the 8-week interventions. (White circle) SHAM group; (Gray circle) OA: knee osteoarthritis group; (Black circle) OAE: knee osteoarthritis plus aerobic training group. N = 9 per group. Data are reported as Mean ± S.E.M. #Difference between OA vs. OAE.
Aerobic training increased the total workload of OAE rats by 64% as compared to sham and OA groups (SHAM: 23.8 ± 8.0 J, OA: 23.7 ± 11.0 J, OAE: 69.2 ± 15.9 J, p = 0.0002, d = 0.85). Levels of IL1β, TNF, IL10 in the joint washed and the level of IL6 in the joint capsule increased respectively by 5.55, 2.84, 1.27, and 1.50-fold in the OA group. In OAE group compared with OA group, the aerobic training modulated levels of these joint cytokines close to those in the SHAM group (IL1β: p < 0.0001, d = 1.18; TNF: p = 0.0001, d = 1.00; IL10: p < 0.0001, d = 1.32; IL6: p = 0.0001, d = 1.68) (Figure 2).
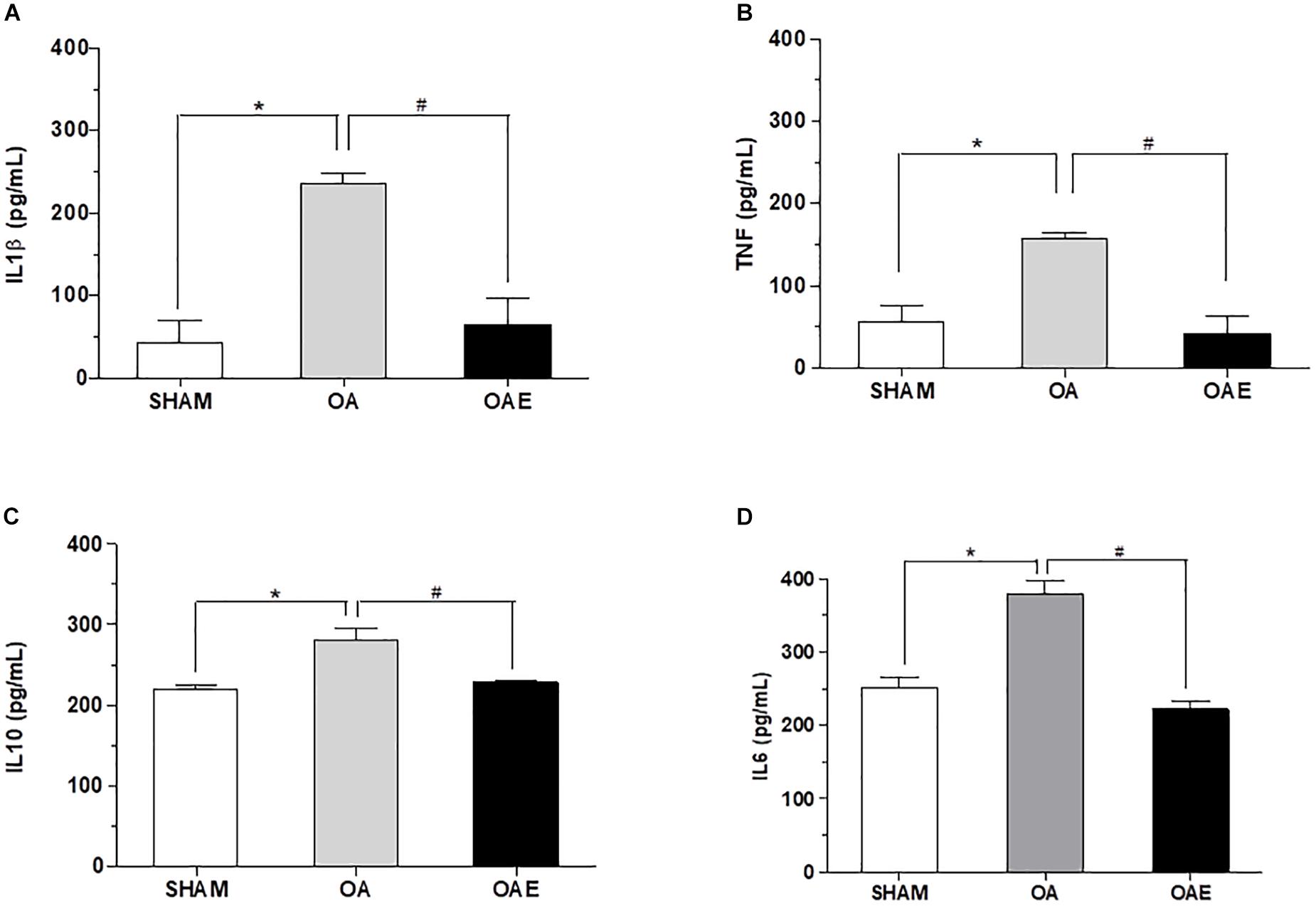
Figure 2. Effect of aerobic training on inflammatory biomarkers. (A) Interleukin-1beta (IL1β) measured in joint wash fluid; (B) Tumor necrose factor (TNF) measured in joint washed; (C) Interleukin-10 (IL10) measured in joint washed, and (D) Interleukin-6 (IL6) measured in the joint capsule for SHAM group; OA: knee osteoarthritis group; OAE: knee osteoarthritis plus aerobic training group. N = 9 per group. Data are reported as Mean ± S.E.M. #Difference between OA vs. OAE. ∗Difference between OA vs. SHAM.
The BDNF level in the OAE group was 41% higher than in the OA group (p = 0.05; d = 0.86). The induction of kOA-like changes increased by 1.14 times the TBARS plasma level and reduced the FRAP plasma level by 6.83% (p = 0.02) in the OA group compared with SHAM group. In OAE group compared with OA group, the aerobic training returned the plasma level of TBARS close to those in the SHAM group, without changing the total antioxidant capacity (TBARS: p = 0.001; d = 0.86; P = 0.95; FRAP: p = 0.05; d = 0.62) (Figure 3).
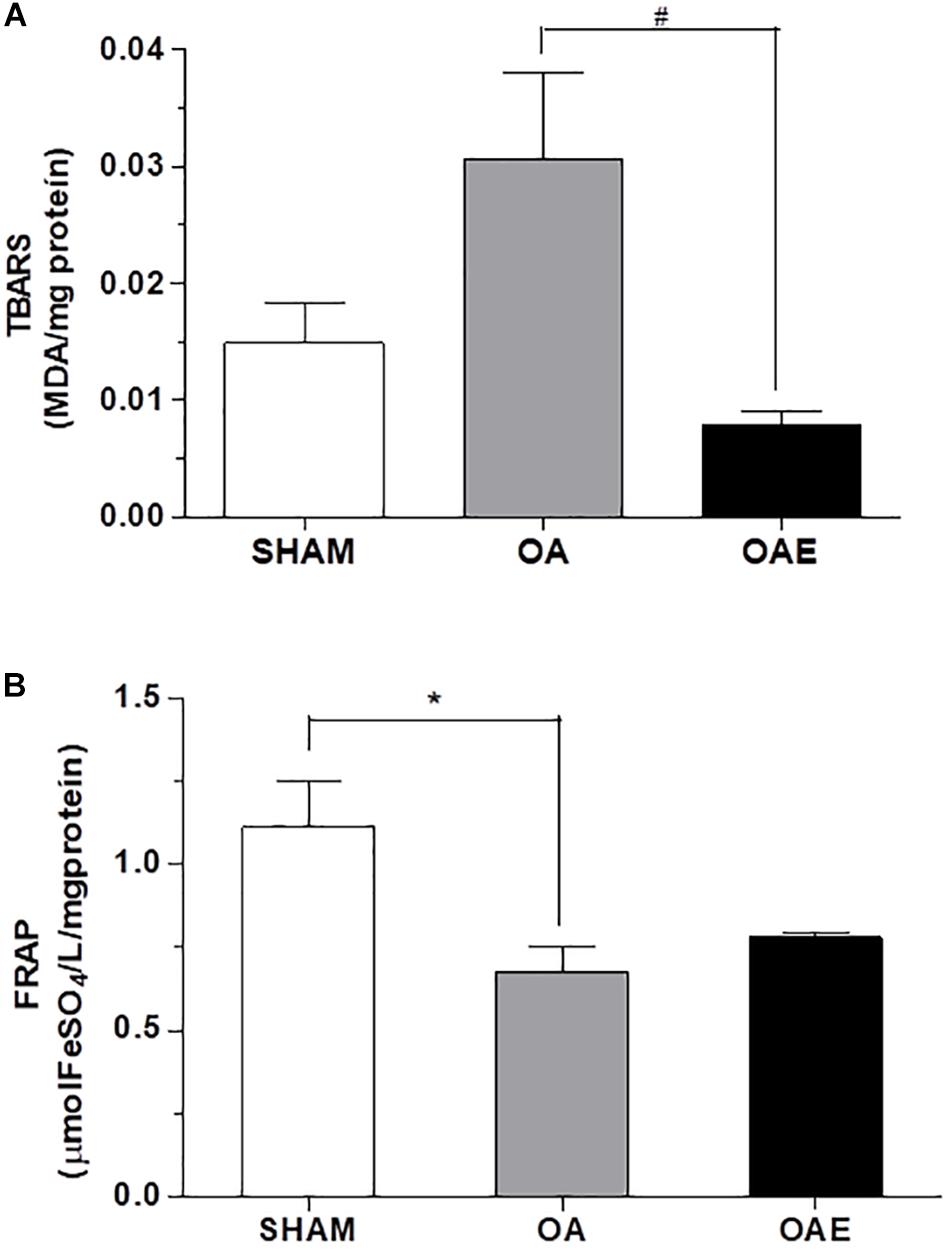
Figure 3. Effect of aerobic training on status redox biomarkers. (A) Thiobarbituric acid-reactive substance (TBARS) measured in plasma; (B) The ferric reducing ability of plasma (FRAP) measured in plasma for SHAM group, OA: knee osteoarthritis group; OAE: knee osteoarthritis plus aerobic training group. N = 9 per group. Data are reported as Mean ± S.E.M. #Difference between OA vs. OAE. ∗Difference between OA vs. SHAM.
The analysis of the joint function tests showed that the kOA-induction (OA group) increased by 1.13 times the total time of displacement on a 100 cm footbridge and increased by 6.05 times the number of failures in the course when compared with SHAM group. The number of falls during the rotarod test increased 2.40 times in the OA group compared with SHAM. Therefore, in OAE group compared with OA group, the aerobic training had a positive effect on the disease, since it improved the performance in the three parameters evaluated, matching the SHAM score (Number of falls: p = 0.002; d = 1.05; Number of failures: p = 0.0002 d = 0.26; Total time of displacement: p = 0.005; d = 0.72) (Figure 4).
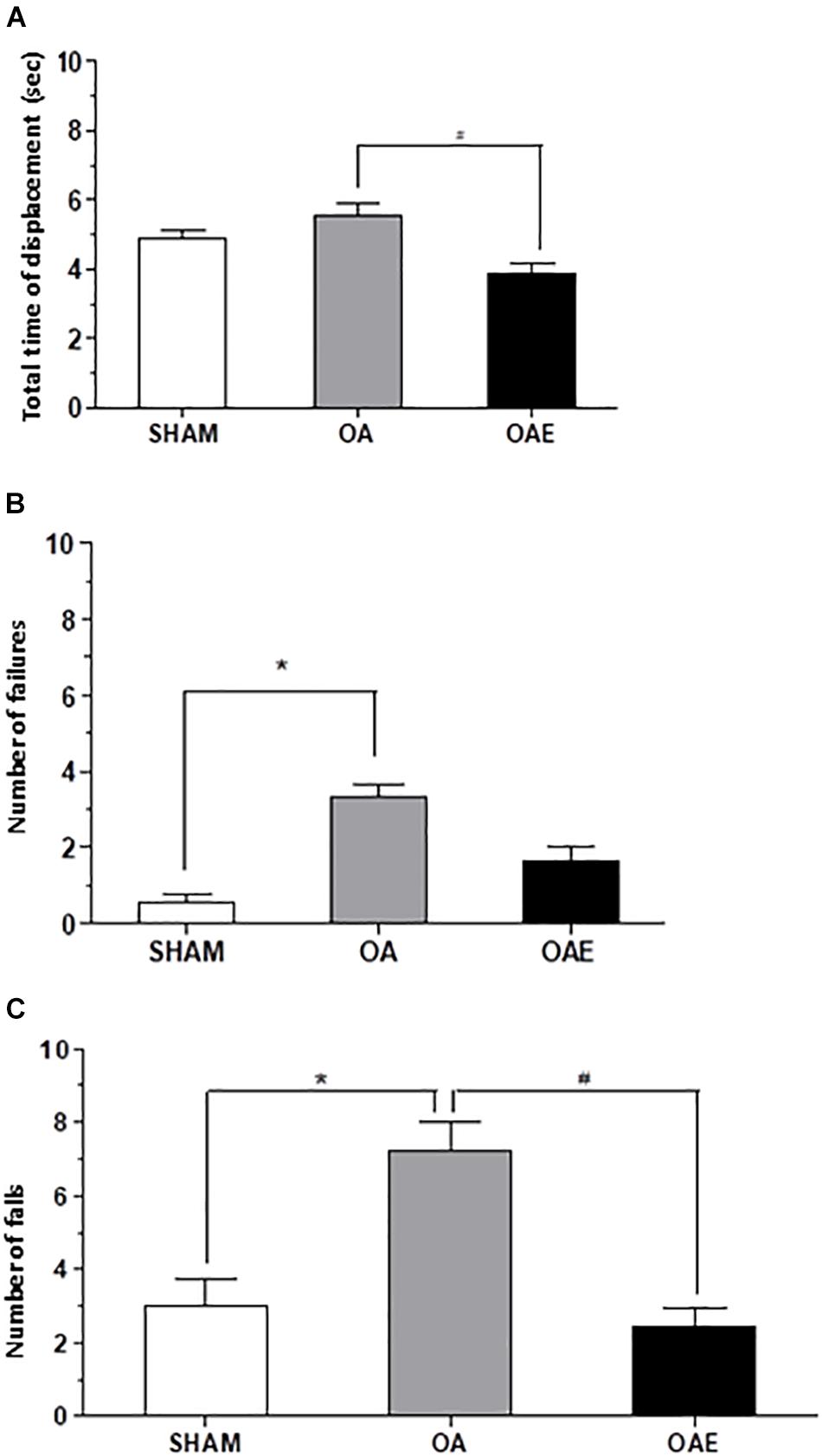
Figure 4. Effect of treadmill aerobic training on joint function tests. (A) Total time of displacement; (B) Number of failures, and (C) Number of falls in SHAM group, OA: knee osteoarthritis group; OAE: knee osteoarthritis plus aerobic training. N = 9 per group. Data are reported as Mean ± S.E.M. #Difference between OA vs OAE; ∗Difference between OA vs. SHAM.
Number of falls (rs = 0.69; p = 0.0003) and number of failures (rs = 0.66; p = 0.0002) were associated with the IL1β. Functional performance measures: number of falls in the rotarod test (rs = 0.73; p < 0.0001); total time of displacement (rs = 0.63; p = 0.0005); and number of failures (rs = 0.52; p = 0.005) were associated with the TNF. Multiple linear stepwise regression models of IL1β and TNF explained 58% of the variability in the number of falls (IL1 β: p = 0.001; TNF: p = 0.002). TNF only explained 29% of the variability of the total time of displacement (p = 0.02) and 21% of the variability of the number of failures (p = 0.02). The increase of 1 pg/mL in intra-articular IL1β level leads to an increase of 0.40 points in the number of falls. The increase of 1 pg/mL in the intra-articular TNF level leads to an increase of 0.51 points in the number of falls; 0.57 s in the total time of displacement; and 0.49 points in the number of failures (Table 1).
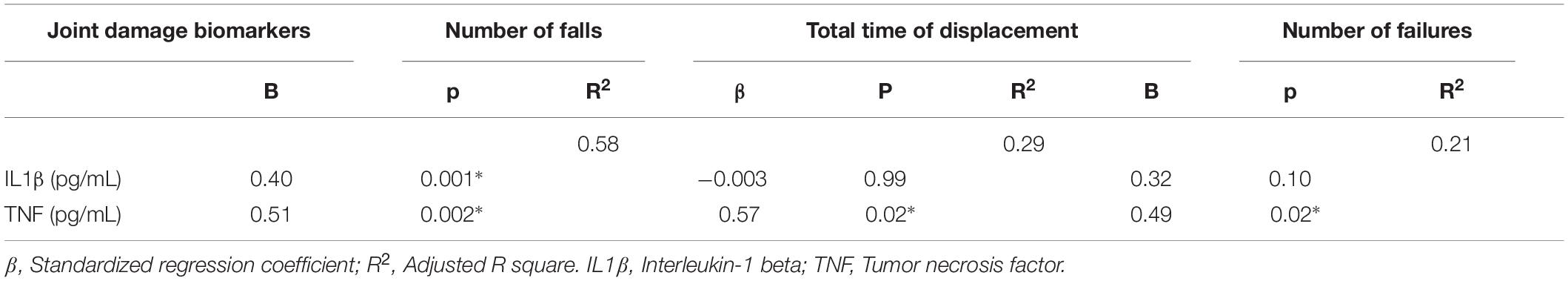
Table 1. Multiple linear stepwise regression analysis.
Quantification of hematoxylin-stained nuclei showed that the induction of kOA reduced the number of cells (chondrocytes) alive by approximately 35% compared with the SHAM group. Aerobic training (OAE group) prevented chondrocyte cell death since the number of active cells in the trained group was 51.35% higher than in the OA group (p < 0.0001) (Figure 5).
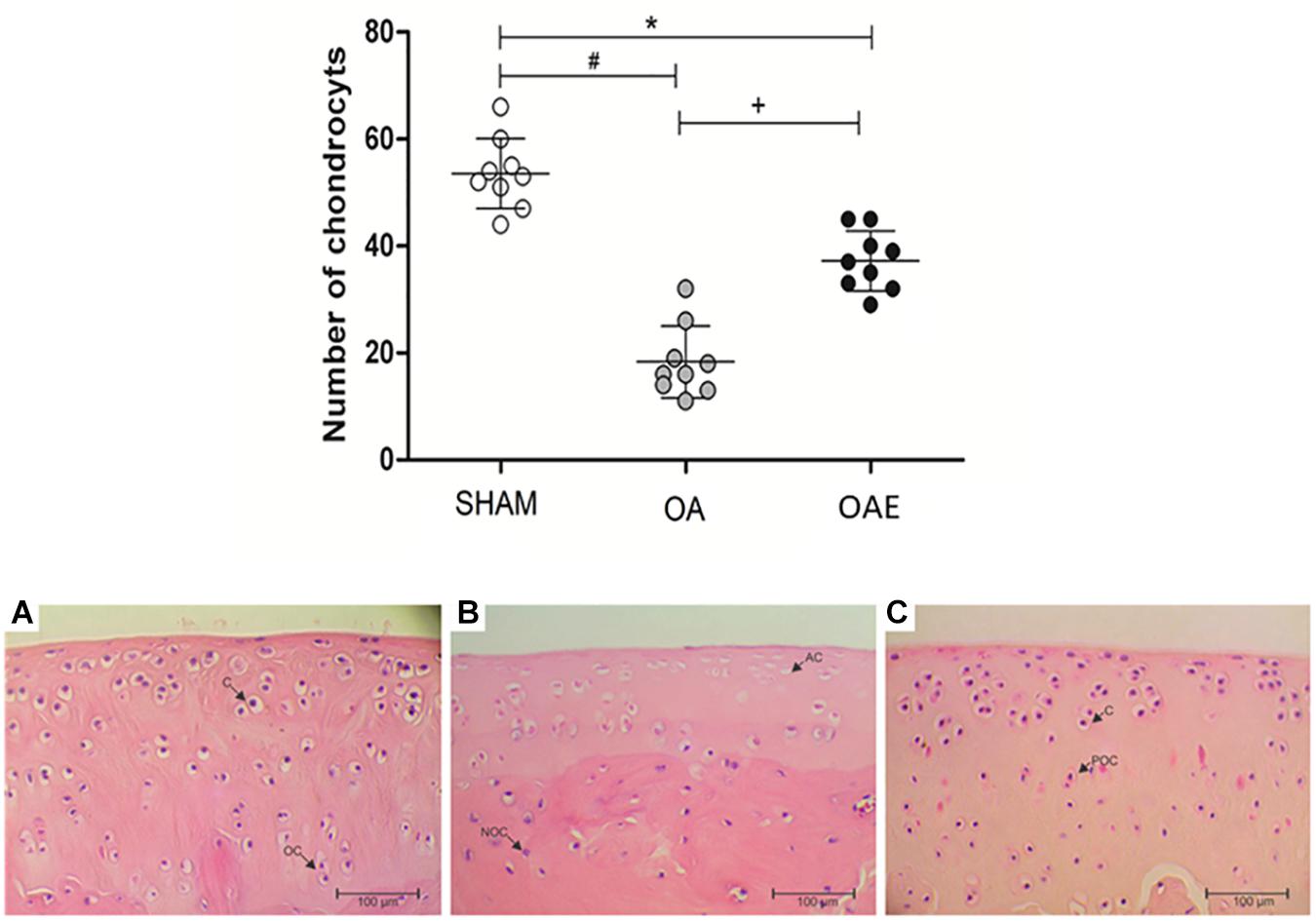
Figure 5. Effect of moderate-intensity aerobic training on the number of chondrocytes, and representation of the histological cell number (chondrocytes). SHAM group (SHAM): white circles and A; knee osteoarthritis group (OA): gray circles and B; knee osteoarthritis plus moderate-intensity aerobic training group (OAE): black circles and C. N = 9 per group. Data are reported as Mean ± S.E.M. +Difference between OA vs. OAE; ∗Difference between OAE vs. SHAM. #Difference between OA vs. SHAM. C: chondrocytes; OC: organized cells; AC: the absence of cells; NOC: no organized cells; POC: partially organized cells.
Overall, the current study showed an association between worse joint function and high levels of joint degeneration biomarkers, as indicated by high levels of TNF and IL1β. Moreover, aerobic training could reverse local inflammatory biomarkers and decrease systemic MDA level with an improvement in gait tasks, motor, and physical performance. Histological data of the femoral joint also confirmed the beneficial effect of the proposed exercise to the kOA-induced.
Scientific evidence suggests the involvement of inflammatory biomarkers and redox status parameters for the advancement and progression of kOA (Regan et al., 2005; Koike et al., 2015). In patients with kOA, chondrocytes and synovial cells stimulate the production of inflammatory cytokines, i.e., IL1β and TNF (Altindag et al., 2007; Kim et al., 2010; Wojdasiewicz et al., 2014; Kunisch et al., 2016). Kim et al. (2010) evidenced that IL1β and TNF are responsible for mitochondrial DNA damage in kOA, promoting the development of ROS and chondrocyte death (Kim et al., 2010).
The compression forces of low magnitude during physical exercise seem to promote physiological control. Such control modulates the synthesis of collagen and proteoglycans possibly inhibited in swelling joints (Quinn et al., 1998; Fehrenbacher et al., 2003; Park et al., 2004). This could indicate the greater maintenance of proteoglycan in the joint cartilage of rats exposed to moderate exercise (Galois et al., 2003; Cifuentes et al., 2010; Li et al., 2013; Mohammadi et al., 2013). Thus, the authors suggest that the positive effects are linked to the decrease of chondrocyte necrosis in the experimental group, lowering metabolites of cell death and induction of inflammatory factors expression (Galois et al., 2003). The high level of ROS would act on the expression of cytokines, making worse the swelling. The ROS may initiate the cartilage degeneration and advancement of lipid peroxidation in chondrocytes (Yudoh et al., 2005; Ostalowska et al., 2006; Kim et al., 2010; Watari et al., 2011).
In kOA, inflammatory cytokines alter the joint biochemical balance inducing chondrocyte necrosis. The production and secretion of catabolic cytokines augment ROS production inducing a redox imbalance. Free radicals interacting with chondrocyte DNA acts directly by altering cellular components, i.e., proteoglycans, collagens, and protein oxidation, favoring cartilage degeneration, which may compromise the thickness of the synovial fluid, and the synthesis of other components (Henrotin et al., 2003; Rose et al., 2012; Reed et al., 2014). Physical and functional damages occur due to tissue structural change. Catabolic cytokines (IL1β and TNF), and regulatory cytokine (IL6) are probably the main factors in this process, facilitating paths of degeneration by activating paths such as matrix metalloproteases (Rojas-Ortega et al., 2015; Assis et al., 2016). The augmented inflammatory profile marks the loss of extracellular matrix integrity, developing an oxidative injury, and, lastly, the chondrocytes death. The death of these chondrocytes seems related to the compromised joint function since this cellular type is responsible for the mobilization of essential components that assure the main functions of load distribution and reduction of friction during static or dynamic exercises that guarantee the joint function. Mohammadi et al. (2013) assessed histological data of depth ratio of lesions demonstrating that 4 weeks of moderate exercise almost treated kOA symptoms in rats (Mohammadi et al., 2013). Moreover, the level of MDA increased in induced kOA dogs. This intensification revealed degeneration of the type II collagen (Goranov, 2007), implying a relationship between redox imbalance and cartilage degeneration. Oxidative injury can result in cell death, triggering particles and oxidized molecules release, cellular degeneration, and increased inflammation.
kOA subjects have greater ROS plasma level and lower antioxidant supplies (Abruzzo et al., 2013; Germanou et al., 2013). The redox imbalance can play a critical role in the cartilage degeneration (Henrotin et al., 2003; Reed et al., 2014). Thus, in both human and animal, kOA transporters have a high level of systemic biomarkers which means cellular damage led by ROS. As a consequence, cell-matrix may be compromised. Inhibition of this course can successfully avoid degeneration of articular cartilage and neo-formation of type II collagen (Poole et al., 2002).
The transduction of mechanical signals of dynamic pressure in chondrocytes may favor the pathways that counteract tissue catabolism. Li et al. (2013) assessed anabolic responses on bovine cartilage in vitro cells culture inducing matrix biosynthesis with different compression ranges (10, 20, and 30%) and demonstrated that moderate dynamic compression can exert an “anti-catabolic” effect, and suppress the expression of TNF, IL6, and soluble IL6 receptors (Li et al., 2013). The catabolism control appears to relate to a range of compression frequency, amplitude, and to the low-to-moderate intensity load, showing the importance of a voltage amplitude threshold for the regulation of inflammatory paths and cell survival (Cifuentes et al., 2010; Beckett et al., 2012; Rojas-Ortega et al., 2015; Rios et al., 2018). Moreover, the exercise load influences BDNF production and release (Nofuji et al., 2008; Yarrow et al., 2010; Szuhany et al., 2015). BDNF is an important growth factor expressed in joint chondrocytes and in epiphyseal plaques of kOA subjects. BDNF causes the growth and, mainly, differentiation (Hutchison, 2012) of chondrocytes, inducing the proliferation pathway that can act as a restoration mediator. Furthermore, BDNF level has been also systemically increased in patients with kOA (Simão et al., 2014).
Because mild- to moderate-intensity exercises seem to play an anti-inflammatory role, we decided to perform a moderate-intensity aerobic training on a treadmill to promote mechanical biostimulation caused by joint compression (Galois et al., 2003; Cifuentes et al., 2010; Li et al., 2013; Assis et al., 2016; Rios et al., 2018). In the current study, we decided to use an aerobic training similar to the study of Cifuentes et al. (2010) that investigated the effects of impact exercise on the kOA-induced cartilage aspects in rats. This protocol was chosen because the data of that investigation demonstrated that aerobic training contributed to the preservation of some joint cartilage parameters in experimental kOA. Moreover, the overload to the intensity and inclination of exercise sessions were not applied, once it could directly affect the dynamic compression of knee joint cartilage cells, leading to misinterpretations of our results. Thus, active mechanotransduction induces changes in oxygen tension and subsequent positive effects on matrix synthesis and cell growth (Urban, 1994; Park et al., 2004). These effects are explained by the displacement of growth factors or cellular cytokines by shifting cellular metabolism (Tilwani et al., 2017). Finally, our results showed that the proposed training offered an inflammatory control, confirmed by the modulation effect in biomarkers levels (IL1β and TNF in the joint wash; IL10 and IL6 in the knee joint).
The positive effects of aerobic training are attributed to the ability of the suppression of signals transduction paths of inflammatory and catabolic mediators together with the stimulation of anabolic paths. In vitro studies have verified that mild to moderate mechanical stress inhibits swelling by suppression of IL1β, TNF and the transcription of various joint degeneration inflammatory biomarkers (Fehrenbacher et al., 2003; Rose et al., 2012; Yamabe et al., 2013). Experimental kOA studies showed a positive effect of aerobic training, whereas high-intensity training had a deleterious effect (Beckett et al., 2012; Ni et al., 2012; Rojas-Ortega et al., 2015; Li et al., 2017). These data seem to determine the role of aerobic exercise appropriate dose (intensity, frequency, and duration) in modulating chondrocyte response (Ni et al., 2013; Na et al., 2014; Hill et al., 2017). In rats without previous kOA induction, high-intensity exercises in many treadmill angles were not able to induce knee damage (Beckett et al., 2012; Rios et al., 2018). Thus, biochemical responses appear to be sensitive to the force only in the injury. Our data once again is in accordance to this premise since the proposed protocol was effective in dropping the joint IL1β and TNF in the trained group, as well as in reducing the TBARS systemic level, augmenting knee joint function and physical performance. In the present study, we identified an increased chondrocytes number in the OAE group. It is already known that chondrocytes are responsible for tissue maintenance which impact on joint function and motor performance (Sophia Fox et al., 2009; Akkiraju and Nohe, 2015). Thus, we speculate that the largest number of chondrocytes and the lower joint degradation cytokines level (IL1β and TNF) (Kim et al., 2010; Li et al., 2013, 2015; Rojas-Ortega et al., 2015) might have preserved the proteoglycans and collagen joint which lead to better motor performance of the OAE group compared to the OA group. Based on the results, exercise prevented the increase of inflammatory biomarkers, and, consequently, prevented the loss of chondrocytes (the only marker evaluated). We cannot extrapolate our results to the whole joint, because the number of chondrocytes was the only analysis. This theory needs to be better clarified ahead. Our study is innovative as it points out the benefits of a therapeutic approach to an experimental OA model. Assessing the effect on physical performance of the animals it was demonstrated that the dose-controlled impact is achieved by ways not fully elucidated in this study. The modulation of inflammation, due to the known effects of aerobic training, has affected the redox status balance, lowering oxidative damage, improving motor performance in functional tasks.
A limitation of this study was that we did not determine kOA-like changes according to the OARSI score. However, it was not possible because we used a single hematoxylin and eosin (H&E) staining for analysis. Thus, immunohistochemistry assessments on cartilage tissue would be useful in future studies.
To conclude, a moderate-intensity aerobic treadmill training appears to modulate chondrocytes via activation of anabolic paths, swelling control by IL1β and TNF levels modulation, systemic TBARS level lowering and positive regulation in joint BDNF level, resulting in physical and motor performance improvements. As a perspective, the efficacy of the training protocol used here should be investigated in older animals.
The datasets generated for this study are available on request to the corresponding author.
This animal study was reviewed and approved by the Commission on Ethics in Animal Use of the Universidade Federal dos Vales do Jequitinhonha e Mucuri (protocol 005/2015). Written informed consent was obtained from the owners for the participation of their animals in this study.
JM, VM, MO, HL, AC, CC, and AL conceived and designed the study. JM, VM, GA, SF, JS, RT-G, DS, MO, HL, AC, AF, CC, JP, and AL contributed to analysis and interpretation of the data. JM, VM, SF, SS, JS, RT-G, DS, TD, and AL drafted the article. JM, VM, SF, MO, HL, AC, AF, CC, JP, MB-F, and AL critically revised the article for important intellectual content. JM, VM, GA, SF, JS, RT-G, DS, MO, HL, AC, AF, CC, JP, VO, MB-F, and AL approved the final article. JM, SF, MO, HL, AC, JP, and AL statistically expertised the study. VM, MO, HL, AC, AF, CC, and AL provisioned the study materials. VM, GA, RT-G, DS, MO, AF, CC, and AL contributed to administrative, technical, or logistic support.
This study was supported in part by the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) (Numbers 408721/2016-2 and 303746/2018-1), the Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG) (Numbers APQ-02503-16, APQ-01239-14, and APQ-03119-18), and the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) (finance code 001).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We acknowledge Martins (2017) who provided data from her master’s degree conducted in the Universidade Federal dos Vales do Jequitinhonha e Mucuri. Besides, the assistance of the Brazilian Society of Physiology is gratefully acknowledged.
Abruzzo, P., Esposito, F., Marchionni, C., Di Tullio, S., Belia, S., Fulle, S., et al. (2013). Moderate exercise training induces ROS-related adaptations to skeletal muscles. Int. J. Sports Med. 34, 676–687. doi: 10.1055/s-0032-1323782
Akkiraju, H., and Nohe, A. (2015). Role of chondrocytes in cartilage formation, progression of osteoarthritis and cartilage regeneration. J. Dev. Biol. 3, 177–192. doi: 10.3390/jdb3040177
Altindag, O., Erel, O., Aksoy, N., Selek, S., Celik, H., and Karaoglanoglu, M. (2007). Increased oxidative stress and its relation with collagen metabolism in knee osteoarthritis. Rheumatol. Int. 27, 339–344. doi: 10.1007/s00296-006-0247-8
Assis, L., Milares, L., Almeida, T., Tim, C., Magri, A., Fernandes, K., et al. (2016). Aerobic exercise training and low-level laser therapy modulate inflammatory response and degenerative process in an experimental model of knee osteoarthritis in rats. Osteoarthr. Cartil. 24, 169–177. doi: 10.1016/j.joca.2015.07.020
Attur, M., Krasnokutsky, S., Statnikov, A., Samuels, J., Li, Z., Friese, O., et al. (2015). Low-grade inflammation in symptomatic knee osteoarthritis: prognostic value of inflammatory plasma lipids and peripheral blood leukocyte biomarkers. Arthritis Rheumatol. 67, 2905–2915. doi: 10.1002/art.39279
Attur, M., Krasnokutsky-Samuels, S., Samuels, J., and Abramson, S. B. (2013). Prognostic biomarkers in osteoarthritis. Curr. Opin. Rheumatol. 25:136. doi: 10.1097/BOR.0b013e32835a9381
Balthazar, C. H., Leite, L. H., Rodrigues, A. G., and Coimbra, C. C. (2009). Performance-enhancing and thermoregulatory effects of intracerebroventricular dopamine in running rats. Pharmacol. Biochem. Behav. 93, 465–469. doi: 10.1016/j.pbb.2009.06.009
Beckett, J., Jin, W., Schultz, M., Chen, A., Tolbert, D., Moed, B. R., et al. (2012). Excessive running induces cartilage degeneration in knee joints and alters gait of rats. J. Orthop. Res. 30, 1604–1610. doi: 10.1002/jor.22124
Benzie, I. F., and Strain, J. J. (1996). The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: the FRAP assay. Anal. Biochem. 239, 70–76. doi: 10.1006/abio.1996.0292
Blasioli, D. J., and Kaplan, D. L. (2013). The roles of catabolic factors in the development of osteoarthritis. Tissue Eng. Part B Rev. 20, 355–363. doi: 10.1089/ten.teb.2013.0377
Bradford, M. M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248–254. doi: 10.1016/0003-2697(76)90527-3
Charlier, E., Relic, B., Deroyer, C., Malaise, O., Neuville, S., Collée, J., et al. (2016). Insights on molecular mechanisms of chondrocytes death in osteoarthritis. Int. J. Mol. Sci. 17:2146. doi: 10.3390/ijms17122146
Cifuentes, D., Rocha, L., Silva, L., Brito, A., Rueff-Barroso, C., Porto, L., et al. (2010). Decrease in oxidative stress and histological changes induced by physical exercise calibrated in rats with osteoarthritis induced by monosodium iodoacetate. Osteoarthr. Cartil. 18, 1088–1095. doi: 10.1016/j.joca.2010.04.004
Fehrenbacher, A., Steck, E., Rickert, M., Roth, W., and Richter, W. (2003). Rapid regulation of collagen but not metalloproteinase 1, 3, 13, 14 and tissue inhibitor of metalloproteinase 1, 2, 3 expression in response to mechanical loading of cartilage explants in vitro. Arch. Biochem. Biophys. 410, 39–47. doi: 10.1016/s0003-9861(02)00658-6
Galois, L., Etienne, S., Grossin, L., Cournil, C., Pinzano, A., Netter, P., et al. (2003). Moderate-impact exercise is associated with decreased severity of experimental osteoarthritis in rats. Rheumatology 42, 692–693. doi: 10.1093/rheumatology/keg094
Geneen, L. J., Moore, R. A., Clarke, C., Martin, D., Colvin, L. A., and Smith, B. H. (2017). Physical activity and exercise for chronic pain in adults: an overview of cochrane reviews. Cochrane Database Syst. Rev. 1:CD011279.
Germanou, E. I., Chatzinikolaou, A., Malliou, P., Beneka, A., Jamurtas, A. Z., Bikos, C., et al. (2013). Oxidative stress and inflammatory responses following an acute bout of isokinetic exercise in obese women with knee osteoarthritis. Knee 20, 581–590. doi: 10.1016/j.knee.2012.10.020
Golightly, Y. M., Allen, K. D., and Caine, D. J. (2012). A comprehensive review of the effectiveness of different exercise programs for patients with osteoarthritis. Phys. Sports Med. 40, 52–65. doi: 10.3810/psm.2012.11.1988
Gomes, W. F., Lacerda, A. C. R., Mendonça, V. A., Arrieiro, A. N., Fonseca, S. F., Amorim, M. R., et al. (2012). Effect of aerobic training on plasma cytokines and soluble receptors in elderly women with knee osteoarthritis, in response to acute exercise. Clin. Rheumatol. 31, 759–766. doi: 10.1007/s10067-011-1927-7
Goranov, N. V. (2007). Serum markers of lipid peroxidation, antioxidant enzymatic defense, and collagen degradation in an experimental (Pond-Nuki) canine model of osteoarthritis. Vet. Clin. Pathol. 36, 192–195. doi: 10.1111/j.1939-165x.2007.tb00208.x
Guzman, R. E., Evans, M. G., Bove, S., Morenko, B., and Kilgore, K. (2003). Mono-iodoacetate-induced histologic changes in subchondral bone and articular cartilage of rat femorotibial joints: an animal model of osteoarthritis. Toxicol. Pathol. 31, 619–624. doi: 10.1080/714044694
Henrotin, Y., Bruckner, P., and Pujol, J.-P. (2003). The role of reactive oxygen species in homeostasis and degradation of cartilage. Osteoarthr. Cartil. 11, 747–755. doi: 10.1016/s1063-4584(03)00150-x
Hill, R. J., Mason, H. M., Yeip, G., Merchant, S. S., Olsen, A. L., Stott, R. D., et al. (2017). The influence of oblique angle forced exercise in surgically destabilized stifle joints is synergistic with bone, but antagonistic with cartilage in an ovine model of osteoarthritis. Arthritis 2017:7481619. doi: 10.1155/2017/7481619
Hui, W., Young, D. A., Rowan, A. D., Xu, X., Cawston, T. E., and Proctor, C. J. (2016). Oxidative changes and signalling pathways are pivotal in initiating age-related changes in articular cartilage. Ann. Rheum. Dis. 75, 449–458. doi: 10.1136/annrheumdis-2014-206295
Hutchison, M. R. (2012). BDNF alters ERK/p38 MAPK activity ratios to promote differentiation in growth plate chondrocytes. Mol. Endocrinol. 26, 1406–1416. doi: 10.1210/me.2012-1063
Iijima, H., Aoyama, T., Ito, A., Yamaguchi, S., Nagai, M., Tajino, J., et al. (2015). Effects of short-term gentle treadmill walking on subchondral bone in a rat model of instability-induced osteoarthritis. Osteoarthr. Cartil. 23, 1563–1574. doi: 10.1016/j.joca.2015.04.015
Jimson, S., Balachander, N., Masthan, K., and Elumalai, R. (2014). A comparative study in bone decalcification using different decalcifying agents. Int. J. Sci. Res. 3, 1226–1229.
Kim, J., Xu, M., Xo, R., Mates, A., Wilson, G., Pearsall, A., et al. (2010). Mitochondrial DNA damage is involved in apoptosis caused by pro-inflammatory cytokines in human OA chondrocytes. Osteoarthr. Cartil. 18, 424–432. doi: 10.1016/j.joca.2009.09.008
Knobloch, T. J., Madhavan, S., Nam, J., Agarwal, S. Jr., and Agarwal, S. (2008). Regulation of chondrocytic gene expression by biomechanical signals. Crit. Rev. Eukaryot. Gene Expr. 18:139. doi: 10.1615/critreveukargeneexpr.v18.i2.30
Koike, M., Nojiri, H., Ozawa, Y., Watanabe, K., Muramatsu, Y., Kaneko, H., et al. (2015). Mechanical overloading causes mitochondrial superoxide and SOD2 imbalance in chondrocytes resulting in cartilage degeneration. Sci. Rep. 5:11722. doi: 10.1038/srep11722
Kunisch, E., Kinne, R. W., Alsalameh, R. J., and Alsalameh, S. (2016). Pro-inflammatory IL-1beta and/or TNF-alpha up-regulate matrix metalloproteases-1 and-3 m RNA in chondrocyte subpopulations potentially pathogenic in osteoarthritis: in situ hybridization studies on a single cell level. Int. J. Rheum. Dis. 19, 557–566. doi: 10.1111/1756-185X.12431
Lacerda, A. C. R., Marubayashi, U., Balthazar, C. H., Leite, L. H., and Coimbra, C. C. (2006). Central nitric oxide inhibition modifies metabolic adjustments induced by exercise in rats. Neurosci. Lett. 410, 152–156. doi: 10.1016/j.neulet.2006.09.067
Leong, D. J., Gu, X. I., Li, Y., Lee, J. Y., Laudier, D. M., Majeska, R. J., et al. (2010). Matrix metalloproteinase-3 in articular cartilage is upregulated by joint immobilization and suppressed by passive joint motion. Matrix Biol. 29, 420–426. doi: 10.1016/j.matbio.2010.02.004
Li, Y., Frank, E. H., Wang, Y., Chubinskaya, S., Huang, H.-H., and Grodzinsky, A. J. (2013). Moderate dynamic compression inhibits pro-catabolic response of cartilage to mechanical injury, TNF-α and IL-6, but accentuates degradation above a strain threshold. Osteoarthr. Cartil. 21, 933–941.
Li, Z., Liu, S.-Y., Xu, L., Xu, S.-Y., and Ni, G.-X. (2017). Effects of treadmill running with different intensity on rat subchondral bone. Sci. Rep. 7:1977. doi: 10.1038/s41598-017-02126-z
Li, Z.-C., Han, N., Li, X., Li, G., Liu, Y.-Z., Sun, G.-X., et al. (2015). Decreased expression of microRNA-130a correlates with TNF-α in the development of osteoarthritis. Int. J. Clin. Exp. Pathol. 8:2555.
Little, C. B., and Ghosh, P. (1997). Variation in proteoglycan metabolism by articular chondrocytes in different joint regions is determined by post-natal mechanical loading. Osteoarthr. Cartil. 5, 49–62. doi: 10.1016/s1063-4584(97)80031-3
Loeser, R. F. (2010). Age-related changes in the musculoskeletal system and the development of osteoarthritis. Clin. Geriatr. Med. 26, 371–386. doi: 10.1016/j.cger.2010.03.002
Loeser, R. F., Goldring, S. R., Scanzello, C. R., and Goldring, M. B. (2012). Osteoarthritis: a disease of the joint as an organ. Arthritis Rheum. 64, 1697–1707. doi: 10.1002/art.34453
Mabey, T., and Honsawek, S. (2015). Cytokines as biochemical markers for knee osteoarthritis. World J. Orthop. 6:95. doi: 10.5312/wjo.v6.i1.95
Martins, J. B. (2017). Treinamento Físico Moderado Em Esteira Modula Biomarcadores Articulares E Melhora O Desempenho Funcional Em Ratos Com Osteoartrite De Joelho Induzida. Ph.D. thesis, Universidade Federal dos Vales do Jequitinhonha e Mucuri, Diamantina, MG.
Mohammadi, M. F., Moghaddam, A. H., and Mirkarimpur, H. (2013). The effects of a moderate exercise program on knee osteoarthritis in male wistar rats. Ir. J. Basic Med. Sci. 16:683.
Na, S. S., Kim, S. G., Yong, M. S., and Hwangbo, G. (2014). Study of treadmill exercise effect on rats with osteoarthritis using proteomic analysis. J. Phys. Ther. Sci. 26, 487–490. doi: 10.1589/jpts.26.487
Ni, G.-X., Lei, L., and Zhou, Y.-Z. (2012). Intensity-dependent effect of treadmill running on lubricin metabolism of rat articular cartilage. Arthritis Res. Ther. 14:R256. doi: 10.1186/ar4101
Ni, G.-X., Liu, S.-Y., Lei, L., Li, Z., Zhou, Y.-Z., and Zhan, L.-Q. (2013). Intensity-dependent effect of treadmill running on knee articular cartilage in a rat model. BioMed. Res. Int. 2013:172392. doi: 10.1155/2013/172392
Nofuji, Y., Suwa, M., Moriyama, Y., Nakano, H., Ichimiya, A., Nishichi, R., et al. (2008). Decreased serum brain-derived neurotrophic factor in trained men. Neurosci. Lett. 437, 29–32. doi: 10.1016/j.neulet.2008.03.057
O’Hara, B., Urban, J., and Maroudas, A. (1990). Influence of cyclic loading on the nutrition of articular cartilage. Ann. Rheum. Dis. 49, 536–539. doi: 10.1136/ard.49.7.536
Ohkawa, H., Ohishi, N., and Yagi, K. (1979). Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 95, 351–358. doi: 10.1016/0003-2697(79)90738-3
Ostalowska, A., Birkner, E., Wiecha, M., Kasperczyk, S., Kasperczyk, A., Kapolka, D., et al. (2006). Lipid peroxidation and antioxidant enzymes in synovial fluid of patients with primary and secondary osteoarthritis of the knee joint. Osteoarthr. Cartil. 14, 139–145. doi: 10.1016/j.joca.2005.08.009
Park, S., Hung, C., and Ateshian, G. (2004). Mechanical response of bovine articular cartilage under dynamic unconfined compression loading at physiological stress levels. Osteoarthr. Cartil. 12, 65–73. doi: 10.1016/j.joca.2003.08.005
Piel, M. J., Kroin, J. S., van Wijnen, A. J., Kc, R., and Im, H.-J. (2014). Pain assessment in animal models of osteoarthritis. Gene 537, 184–188. doi: 10.1016/j.gene.2013.11.091
Poole, A., Kobayashi, M., Yasuda, T., Laverty, S., Mwale, F., Kojima, T., et al. (2002). Type II collagen degradation and its regulation in articular cartilage in osteoarthritis. Ann. Rheum. Dis. 61(Suppl. 2), ii78–ii81.
Primola-Gomes, T. N., Campos, L. A., Lauton-Santos, S., Balthazar, C. H., Guatimosim, S., Capettini, L. S., et al. (2009). Exercise capacity is related to calcium transients in ventricular cardiomyocytes. J. Appl. Physiol. 107, 593–598. doi: 10.1152/japplphysiol.91218.2008
Quinn, T. M., Grodzinsky, A. J., Buschmann, M. D., Kim, Y.-J., and Hunziker, E. B. (1998). Mechanical compression alters proteoglycan deposition and matrix deformation around individual cells in cartilage explants. J. Cell Sci. 111, 573–583.
Reed, K. N., Wilson, G., Pearsall, A., and Grishko, V. I. (2014). The role of mitochondrial reactive oxygen species in cartilage matrix destruction. Mol. Cell. Biochem. 397, 195–201. doi: 10.1007/s11010-014-2187-z
Regan, E., Flannelly, J., Bowler, R., Tran, K., Nicks, M., Carbone, B. D., et al. (2005). Extracellular superoxide dismutase and oxidant damage in osteoarthritis. Arthritis Rheum. 52, 3479–3491. doi: 10.1002/art.21387
Rios, J. L., Boldt, K. R., Mather, J. W., Seerattan, R. A., Hart, D. A., and Herzog, W. (2018). Quantifying the effects of different treadmill training speeds and durations on the health of rat knee joints. Sports Med. Open 4:15. doi: 10.1186/s40798-018-0127-2
Rojas-Ortega, M., Cruz, R., Vega-López, M. A., Cabrera-González, M., Hernández-Hernández, J. M., Lavalle-Montalvo, C., et al. (2015). Exercise modulates the expression of IL-1β and IL-10 in the articular cartilage of normal and osteoarthritis-induced rats. Pathol. Res. Pract. 211, 435–443. doi: 10.1016/j.prp.2015.01.008
Roman-Blas, J. A., Contreras-Blasco, M. A., Largo, R., Álvarez-Soria, M. A., Castañeda, S., and Herrero-Beaumont, G. (2009). Differential effects of the antioxidant N-acetylcysteine on the production of catabolic mediators in IL-1β-stimulated human osteoarthritic synoviocytes and chondrocytes. Eur. J. Pharmacol. 623, 125–131. doi: 10.1016/j.ejphar.2009.09.016
Rose, J., Söder, S., Skhirtladze, C., Schmitz, N., Gebhard, P., Sesselmann, S., et al. (2012). DNA damage, discoordinated gene expression and cellular senescence in osteoarthritic chondrocytes. Osteoarthr. Cartil. 20, 1020–1028. doi: 10.1016/j.joca.2012.05.009
Semanik, P. A., Chang, R. W., and Dunlop, D. D. (2012). Aerobic activity in prevention and symptom control of osteoarthritis. PM R 4, S37–S44. doi: 10.1016/j.pmrj.2012.02.015
Simão, A. P., Mendonça, V. A., de Oliveira Almeida, T. M., Santos, S. A., Gomes, W. F., Coimbra, C. C., et al. (2014). Involvement of BDNF in knee osteoarthritis: the relationship with inflammation and clinical parameters. Rheumatol. Int. 34, 1153–1157. doi: 10.1007/s00296-013-2943-5
Sophia Fox, A. J., Bedi, A., and Rodeo, S. A. (2009). The basic science of articular cartilage: structure, composition, and function. Sports Health 1, 461–468. doi: 10.1177/1941738109350438
Szuhany, K. L., Bugatti, M., and Otto, M. W. (2015). A meta-analytic review of the effects of exercise on brain-derived neurotrophic factor. J. Psychiatr. Res. 60, 56–64. doi: 10.1016/j.jpsychires.2014.10.003
Takahashi, I., Matsuzaki, T., Kuroki, H., and Hoso, M. (2018). Induction of osteoarthritis by injecting monosodium iodoacetate into the patellofemoral joint of an experimental rat model. PLoS One 13:e0196625. doi: 10.1371/journal.pone.0196625
Tilwani, R., Vessillier, S., Pingguan-Murphy, B., Lee, D., Bader, D., and Chowdhury, T. (2017). Oxygen tension modulates the effects of TNFα in compressed chondrocytes. Inflamm. Res. 66, 49–58. doi: 10.1007/s00011-016-0991-5
Urban, J. (1994). The chondrocyte: a cell under pressure. Rheumatology 33, 901–908. doi: 10.1093/rheumatology/33.10.901
Watari, T., Naito, K., Sakamoto, K., Kurosawa, H., Nagaoka, I., and Kaneko, K. (2011). Evaluation of the effect of oxidative stress on articular cartilage in spontaneously osteoarthritic STR/OrtCrlj mice by measuring the biomarkers for oxidative stress and type II collagen degradation/synthesis. Exp. Ther. Med. 2, 245–250. doi: 10.3892/etm.2011.196
Wojdasiewicz, P., Poniatowski, ŁA., and Szukiewicz, D. (2014). The role of inflammatory and anti-inflammatory cytokines in the pathogenesis of osteoarthritis. Mediators Inflamm. 2014:561459. doi: 10.1155/2014/561459
Yamabe, S., Hirose, J., Uehara, Y., Okada, T., Okamoto, N., Oka, K., et al. (2013). Intracellular accumulation of advanced glycation end products induces apoptosis via endoplasmic reticulum stress in chondrocytes. FEBS J. 280, 1617–1629. doi: 10.1111/febs.12170
Yarrow, J. F., White, L. J., McCoy, S. C., and Borst, S. E. (2010). Training augments resistance exercise induced elevation of circulating brain derived neurotrophic factor (BDNF). Neurosci. Lett. 479, 161–165. doi: 10.1016/j.neulet.2010.05.058
Yudoh, K., van Trieu, N., Nakamura, H., Hongo-Masuko, K., Kato, T., and Nishioka, K. (2005). Potential involvement of oxidative stress in cartilage senescence and development of osteoarthritis: oxidative stress induces chondrocyte telomere instability and downregulation of chondrocyte function. Arthritis Res. Ther. 7:R380.
Keywords: osteoarthritis, aerobic training, exercise, biomarkers, joint function
Citation: Martins JB, Mendonça VA, Aguiar GC, Fonseca SF, Santos JM, Tossige-Gomes R, Melo DS, Oliveira MX, Leite HR, Camargos ACR, Ferreira AJ, Coimbra CC, Poortmans J, Oliveira VC, Silva SB, Domingues TE, Bernardo-Filho M and Lacerda ACR (2019) Effect of a Moderate-Intensity Aerobic Training on Joint Biomarkers and Functional Adaptations in Rats Subjected to Induced Knee Osteoarthritis. Front. Physiol. 10:1168. doi: 10.3389/fphys.2019.01168
Received: 14 June 2019; Accepted: 29 August 2019;
Published: 18 September 2019.
Edited by:
Leonardo Alexandre Peyré-Tartaruga, Federal University of Rio Grande do Sul, BrazilReviewed by:
Júlio Cezar De Oliveira, Federal University of Mato Grosso, BrazilCopyright © 2019 Martins, Mendonça, Aguiar, Fonseca, Santos, Tossige-Gomes, Melo, Oliveira, Leite, Camargos, Ferreira, Coimbra, Poortmans, Oliveira, Silva, Domingues, Bernardo-Filho and Lacerda. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ana Cristina Rodrigues Lacerda, bGFjZXJkYWFjckBnbWFpbC5jb20=; bGFjZXJkYS5hY3JAdWZ2am0uZWR1LmJy
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.