 Naymã Dias1*
Naymã Dias1* Deise Cagliari1
Deise Cagliari1 Frederico Schmitt Kremer2
Frederico Schmitt Kremer2 Leticia Neutzling Rickes1
Leticia Neutzling Rickes1 Dori Edson Nava3
Dori Edson Nava3 Guy Smagghe4*
Guy Smagghe4* Moisés Zotti1*
Moisés Zotti1*- 1Molecular Entomology and Applied Bioinformatics Laboratory, Faculty of Agronomy, Department of Crop Protection, Federal University of Pelotas, Pelotas, Brazil
- 2Bioinformatics and Proteomics Laboratory, Technological Development Center, Federal University of Pelotas, Pelotas, Brazil
- 3Entomology Laboratory, Embrapa Clima Temperado, Pelotas, Brazil
- 4Faculty of Bioscience Engineering, Department of Plants and Crops, Ghent University, Ghent, Belgium
RNA interference (RNAi) technology has been used in the development of approaches for pest control. The presence of some essential genes, the so-called “core genes,” in the RNAi machinery is crucial for its efficiency and robust response in gene silencing. Thus, our study was designed to examine whether the RNAi machinery is functional in the South American (SA) fruit fly Anastrepha fraterculus (Diptera: Tephritidae) and whether the sensitivity to the uptake of double-stranded RNA (dsRNA) could generate an RNAi response in this fruit fly species. To prepare a transcriptome database of the SA fruit fly, total RNA was extracted from all the life stages for later cDNA synthesis and Illumina sequencing. After the de novo transcriptome assembly and gene annotation, the transcriptome was screened for RNAi pathway genes, as well as the duplication or loss of genes and novel target genes to dsRNA delivery bioassays. The dsRNA delivery assay by soaking was performed in larvae to evaluate the gene-silencing of V-ATPase, and the upregulation of Dicer-2 and Argonaute-2 after dsRNA delivery was analyzed to verify the activation of siRNAi machinery. We tested the stability of dsRNA using dsGFP with an in vitro incubation of larvae body fluid (hemolymph). We identified 55 genes related to the RNAi machinery with duplication and loss for some genes and selected 143 different target genes related to biological processes involved in post-embryonic growth/development and reproduction of A. fraterculus. Larvae soaked in dsRNA (dsV-ATPase) solution showed a strong knockdown of V-ATPase after 48 h, and the expression of Dicer-2 and Argonaute-2 responded with an increase upon the exposure to dsRNA. Our data demonstrated the existence of a functional RNAi machinery in the SA fruit fly, and we present an easy and robust physiological bioassay with the larval stages that can further be used for screening of target genes at in vivo organisms’ level for RNAi-based control of fruit fly pests. This is the first study that provides evidence of a functional siRNA machinery in the SA fruit fly.
Introduction
The South American fruit fly (SA fruit fly), Anastrepha fraterculus, is a major polyphagous pest of fruit crops. This fruit fly species occurs from the Southern United States (Texas) and Mexico to Argentina and is associated with 116 plant species in Brazil alone (Zucchi, 2008). Oviposition and larval feeding of A. fraterculus cause the damage that leads to accelerated ripening and premature fruit dropping (Aluja, 1994). Importantly, its presence limits access to markets because of quarantine limitations imposed by fruit-fly free countries (Lanzavecchia et al., 2014; Dias et al., 2018). The global losses caused by fruit flies can range 2 billion USD annually, and in Brazil, the economic losses are between $120 and 200 million USD per year (Macedo et al., 2017).
Currently, the control tactic available for A. fraterculus is the application of bait sprays (Dias et al., 2018). However, the control of SA fruit fly by chemical tactics is becoming increasingly restricted, because the effective but broad-spectrum neurotoxic and systemic-acting insecticides have been banned for commercialization (Böckmann et al., 2014). Fruit growers are also seeking new economic fruit fly control options, especially environmentally sustainable tactics (Sarles et al., 2015). Thus, RNA interference (RNAi) is a promising strategy for pest control used to suppress the expression of key genes (Katoch et al., 2013; Andrade and Hunter, 2017). Molecules of double-stranded RNA (dsRNA) are RNAi triggers that promote the post-transcriptional down-regulation of a target-gene (Elbashir et al., 2001). Some features, such as high target gene specificity and lack of environmental persistence, make RNAi technique desirable for crop protection (Huvenne and Smagghe, 2010; Zotti et al., 2018).
Efficient gene silencing by RNAi in insects requires some essential elements, such as dsRNA processing by RNAi enzymes, dsRNA uptake into cells, and expression of the RNAi core genes (Huvenne and Smagghe, 2010; Wang et al., 2016; Christiaens et al., 2018; Niu et al., 2018). Drosophila species have been used as a model for RNAi studies in Diptera. However, this species shows low sensibility to dsRNA uptake by cells, and consequently it is necessary to use transfection agents for delivery of dsRNA molecules (Taning et al., 2016). Soaking of Drosophila melanogaster larvae for a period of 1 h with naked dsRNA resulted in only 5–8% of knockdown for b-glucuronidase (gus) (Whyard et al., 2009). In Drosophila suzukii larvae, the RNAi efficiency varied between 20 and 40% in a study using dsRNA formulated with transfection reagent (Taning et al., 2016). For Bactrocera dorsalis, Shi et al. (2017) found knockdown around 50% in larval stages. This fact raises the question about the variability in the uptake routes and uptake mechanisms between different Diptera species (Whyard et al., 2009).
Thus, the understanding of the RNAi pathway in the target organism can provide information about use of this tool for pest control (Vélez et al., 2016). However, for the SA fruit fly, we first need the molecular information on RNAi core genes, in addition to insights into the gene-silencing mechanism by RNAi.
This paper is the first report of RNAi bioassays in the SA fruit fly together with a transcriptome analysis over the life stages of eggs, larvae, pupae, and female and male adults. We aimed to provide a genetic database to better understand this important pest insect and to screen for genes related to the RNAi machinery. We also aimed to identify possible gene duplication, gene loss, and novel target genes for dsRNA bioassays. Hence, we focus also on genes related to insect-specific biological processes involved in post-embryonic growth/development and reproduction as potential future insecticidal target genes.
In addition, we developed a straightforward experimental RNAi setup by soaking the SA fruit fly larvae. If successful, it is an easy and robust bioassay for the larval stages that can be used to screen target genes in vivo at organism level. In order to validate the RNAi response, we first investigated the expression of Dicer-2 and Argonaute-2. Next, we investigated the silencing of V-ATPase and the insect mortality. Finally, we measured the stability of dsRNA with an in vitro incubation together with hemolymph. Overall, this study will be the first to provide evidence of a functional siRNA machinery in the SA fruit fly.
Materials and Methods
SA Fruit Fly Colony and Maintenance
A colony of A. fraterculus was originally field-collected in 2015 from an orchard of strawberry guava (Psidium cattleianum) in Pelotas, Rio Grande do Sul, Brazil (31°40′47″ S e 52°26′24″ W) and was reared for thirteen generations before use in the experiments. SA fruit fly stages were maintained under standard conditions (temperature: 25 ± 1°C; RH: 70 ± 10% and 14L:10D photoperiod). The rearing methods were the same as described by Gonçalves et al. (2013).
RNA Extraction, cDNA Library, and RNA-Seq
Total RNA was extracted from eggs, larvae (first-, second-, and third-instar), pupae, and adults (female and male) of SA fruit fly using the RNAzol (GeneCopoeia, Rockville, MD) and treated with DNase I (Invitrogen, Carlsbad, CA), following the manufacturer’s instructions. The RNA samples were pooled prior to cDNA synthesis. The RNA quality and concentration were examined on the Agilent 2100 Bioanalyzer and cDNA library was performed using the TruSeq RNA Sample Prep kit (Illumina, San Diego, CA) protocol. The library was paired-end (2x125bp) sequenced (RNA-Seq) using the Illumina HiSeq2500 platform. Raw sequence data were submitted to the Short Read Archive (SRA) of the NCBI database (accession number SRP157027).
Reads Quality Control and de novo Assembly
All reads were trimmed for quality and length using the software Trimmomatic1 and the quality was checked using the software FastQC2. Phred score over 30 across more than 70% of the bases was used as a high-quality threshold. The high-quality reads were de novo assembled using Trinity software3 since there is no reference genome sequence for A. fraterculus. This software uses a De Bruijn graph algorithm and was executed using default settings, a k-mer length of 25.
Transcriptome Analysis and Target-Genes Database
The contigs generated by Trinity were aligned to the UniProt database using Diamond algorithm (Buchfink et al., 2015) and only those with hits on insects (E-value of 1e-10) were selected for analysis. For functional categorization by gene ontology (GO), a similarity search was performed to predict the contigs generated by searching the UniProt database with the Diamond. The predicted genes were used as query in QuickGo from EBI4 and to calculate GO terms. A database was generated for novel target genes related to post-embryonic growth and development of the SA fruit fly larvae and the reproduction events in adults. The target genes for SA fruit fly database were searched in QuickGo using the GO terms related to biological processes: larval development (GO:0002164), imaginal disk morphogenesis (GO:0007560), post-embryonic development (GO:0009791), female sex differentiation (GO:0046660), sexual reproduction (GO:0019953), genital disk anterior/posterior pattern formation (GO:0035224) and oviposition (GO:0018991). The D. melanogaster sequences corresponding to the genes found were recovered in UniProt database and were used as an input to search the transcriptome from A. fraterculus using the tblastn tool with a threshold bit score ≥ 150 and E-value ≤ 1e-5 (Supplementary Material S1).
Identification of RNAi Machinery Genes
A list of RNAi-related genes, as employed by Swevers et al. (2013), Prentice et al. (2015), and Yoon et al. (2016), was selected covering the RNAi core machinery, auxiliary factors (from RISC), dsRNA uptake, nucleases, antiviral RNAi, intracellular transport, and lipid metabolism. Homologous sequences from D. melanogaster corresponding to RNAi-related genes were obtained in UniProt database and were used as an input to search the transcriptome from SA fruit fly (Supplementary Material S2). Alternatively, sequences of Drosophila and Tephritidae species were used in the absence of sequences of D. melanogaster (Supplementary Material S2). The ORF Finder tool from NCBI was used to detect open reading frames. The protein domains were predicted by NCBI Conserved Domains using the Conserved Domain Database (CDD) (Supplementary Material S2). A similarity search was performed using the BLASTp against the NCBI database to confirm the identity of the RNAi-related genes (Supplementary Material S4).
Potential Loss and Duplication of RNAi-Related Genes
We screened the SA fruit fly transcriptome for the copy number of the 10 RNAi pathway genes using tblastn tool. The number of copies was based in the number of genes obtained by Trinity assembly. The distribution of these genes was compared to related insects, following the results showed by Dowling et al. (2016). We also searched for genes for a systemic RNAi response, as SID-1 found in cells of Caenorhabditis elegans (Winston et al., 2002).
Phylogenetic Analysis
A phylogenetic analysis was constructed to provide additional confirmation of the main siRNA machinery genes (Dicer-2 and Argonaute-2) and the candidate gene for silencing (Vacuolar-proton-ATPase) from the A. fraterculus transcriptome. Phylogenetic trees were constructed through the Neighbor-Joining method in the MEGA X software using the bootstrapping reconstructions (1000 replicates). The selected species and accession numbers of the sequences used for analysis are shown in Supplementary Table S4.
dsRNA Synthesis
The A. fraterculus transcriptome was searched for the Vacuolar-proton-ATPase V0-domain (V-ATPase V0) using D. melanogaster sequence as a query. Primers were designed from the A. fraterculus transcriptome sequences using Primer35. The V-ATPase V0 fragment (483 pb) was amplified by PCR using cDNA from second-instar larvae of A. fraterculus as a template, synthesized with SuperScript First-Strand Synthesis System for RT-PCR (Invitrogen, Carlsbad, CA). For dsRNA synthesis of green fluorescent protein (GFP), a 560 bp GFP fragment was amplified by PCR using plasmid pIG1783f. The GFP amplicon was confirmed by Sanger sequencing. The primers used for the PCR are listed in Supplementary Table S1.
The dsRNA templates were generated by PCR using primers with a T7 promoter region at the 5′ end of each primer (Supplementary Table S1). The PCR products were used for in vitro transcription and purification using MEGAscript kit (Ambion, Austin, TX) according to the manufacturer’s instructions. Synthesized dsRNA products were quantitated by a Nanovue spectrophotometer (GE Healthcare, Little Chalfont, United Kingdom) at 260 nm and the integrity was confirmed by electrophoresis on 1% agarose gel.
RNAi by Soaking of Larval Stages
The soaking treatment was performed using second-instar larvae of A. fraterculus. The dsRNA of V-ATPase V0 (dsVTP) was diluted with RNase-free water to yield 500 ng/μl, considering the data reported by Whyard et al. (2009). The dsGFP in the same concentration was used as control for the soaking assays. The insects were starved for 1 h and each larva was soaked in a 200 μl-tube with 25 μl of dsRNA solution for 30 min. After soaking, the treated larvae were transferred to artificial diet (Nunes et al., 2013). The mortality of the insects was monitored over a 7-day period.
Larvae of A. fraterculus were stored at -80°C at 24, 48, and 72 h after soaking with dsRNA for the RNAi silencing efficiency assay. The RNA was extracted with three biological replicates at each time point, using RNAzol (GeneCopoeia, Rockville, MD) following the manufacturer’s instructions. After, the RNA samples were incubated with 10 U DNase I (Invitrogen, Carlsbad, CA) at 37°C for 30 min. The RNA was quantified using a Nanovue spectrophotometer (GE Healthcare, Little Chalfont, United Kingdom) and verified by 2% agarose gel electrophoresis. The cDNA was produced from 2 μg RNA using the SuperScript First-Strand Synthesis System for RT-PCR (Invitrogen, Carlsbad, CA).
Measurement of RNAi Efficacy
Real-time Quantitative PCR analysis (qPCR) was performed to evaluated RNAi efficacy using a LightCycler 480 (Roche Life Science, Switzerland). A standard curve based on a serial dilution (1:1, 1:5, 1:25, and 1:125) of cDNA was performed to validate the primers used in the analysis (Supplementary Table S1). The reactions included 5 μl of EvaGreen 2X qPCR MasterMix (ABM, Canada), 0.3 μl (10 μM) of forward primer, 0.3 μl (10 μM) of reverse primer, 3.4 μl of nuclease-free water and 1 μl of cDNA, in a total volume of 10 μl. The qPCR conditions included 10 min at 95°C followed by 40 cycles of 30 s at 95°C, 45 s at 59°C, and 30 s at 77°C, interrupted by the dissociation curve with denaturation at 95°C (5 s), cooling at 70°C (1 min) and gradually heating at 0.11°C steps up to 95°C and cooling at 40°C (30 s), as described by Benemann et al. (2017). The reactions were set-up in 96-well microtiter plates (Roche Life Science, Indianapolis, IN), using the cDNA dilution of 1:25, with three technical replicates and no-template controls. Relative mRNA expression of the V-ATPase gene was normalized to the housekeeping genes α-tubulin and actin by the equation ratio 2-ΔΔCt (Livak and Schmittgen, 2001). The data were analyzed using analysis of variance (one-way ANOVA) and t-test (p ≤ 0.05).
Expression of siRNA Genes Dcr-2 and Ago-2 Upon Exposure to dsRNA
The regulation of siRNA pathway genes during the SA fruit fly RNAi bioassay was determined by the expression of Dicer-2 (Dcr-2) and Argonaute-2 (Ago-2) in response to soaking with dsGFP. The Dcr-2 and Ago-2 sequences found in the A. fraterculus transcriptome were used for primers design using the Primer3. The primers used for the qPCR are listed in Supplementary Table S1. The qPCR analysis was performed as described above, and the expression responses were evaluated at 24, 48, and 72 h after larvae soaking with dsGPF.
dsRNA Degradation Assay
Body fluid (hemolymph) was collected in pre-chilled 1.5 ml-tubes by centrifugation at 13,000 rpm for 10 min at 4°C from 5 second-instar larvae. For the degradation assay, 20 μl of dsGFP solution (500 ng/μl dsRNA) was mixed with 2 μl of body fluid and incubated at 25°C. We collected aliquots of 5 μl at 0, 1, 2, and 4 h after incubation and the same volume of EDTA (10 mM) was added to stop the enzymatic reaction. The samples were stocked at -80°C until the analysis. The results were verified by 1.5% agarose gel electrophoresis and the bands were analyzed using the Gel Analyzer software.
Results
SA Fruit Fly Transcriptome Analysis
The RNA sequencing generated a total of 103,808,135 reads of 125 bp length. The assembled transcriptome consisted of 163,359 transcripts, which accounted for 84,105 contigs (Supplementary Table S2). Of all contigs, 72,388 are eukaryotic. The length distribution of Eukaryote contigs in A. fraterculus transcriptome is shown in Supplementary Figure S1.
The Diamond analysis produced 73,193 hits, representing 45% of the total contigs (Supplementary Figure S2). For the sequences with significant hits, 72% of the contigs were similar to sequences from fruit fly species: 17% to Ceratitis capitata, 16% to Zeugodacus cucurbitae, 15% to B. dorsalis and Bactrocera latifrons, 9% to Bactrocera tryoni, and 28% to other organisms. The species distribution of the top 30 hits is shown in Supplementary Table S3. Considering the insect genera, the contigs were similar to sequences from Diptera, which featured for 55% to Bactrocera, 16% to Ceratitis, 3% to Drosophila, 1% to Tabanus, 0.9% to Glossina, 0.8% to Lucilia, and 20% to other insect genera.
The GO terms calculated starting the Diamond similarity search were grouped into three main categories: molecular function (48%), biological process (31%) and cellular component (20%). A total of 167,729 predicted GO terms was obtained. The major GO terms within the molecular function category were nucleic acid binding (11,734; 7%), for the biological processes it was RNA-dependent DNA biosynthetic process (4,070; 2%), and for the cellular component, it was the membrane (10,584; 6%) (Figure 1).
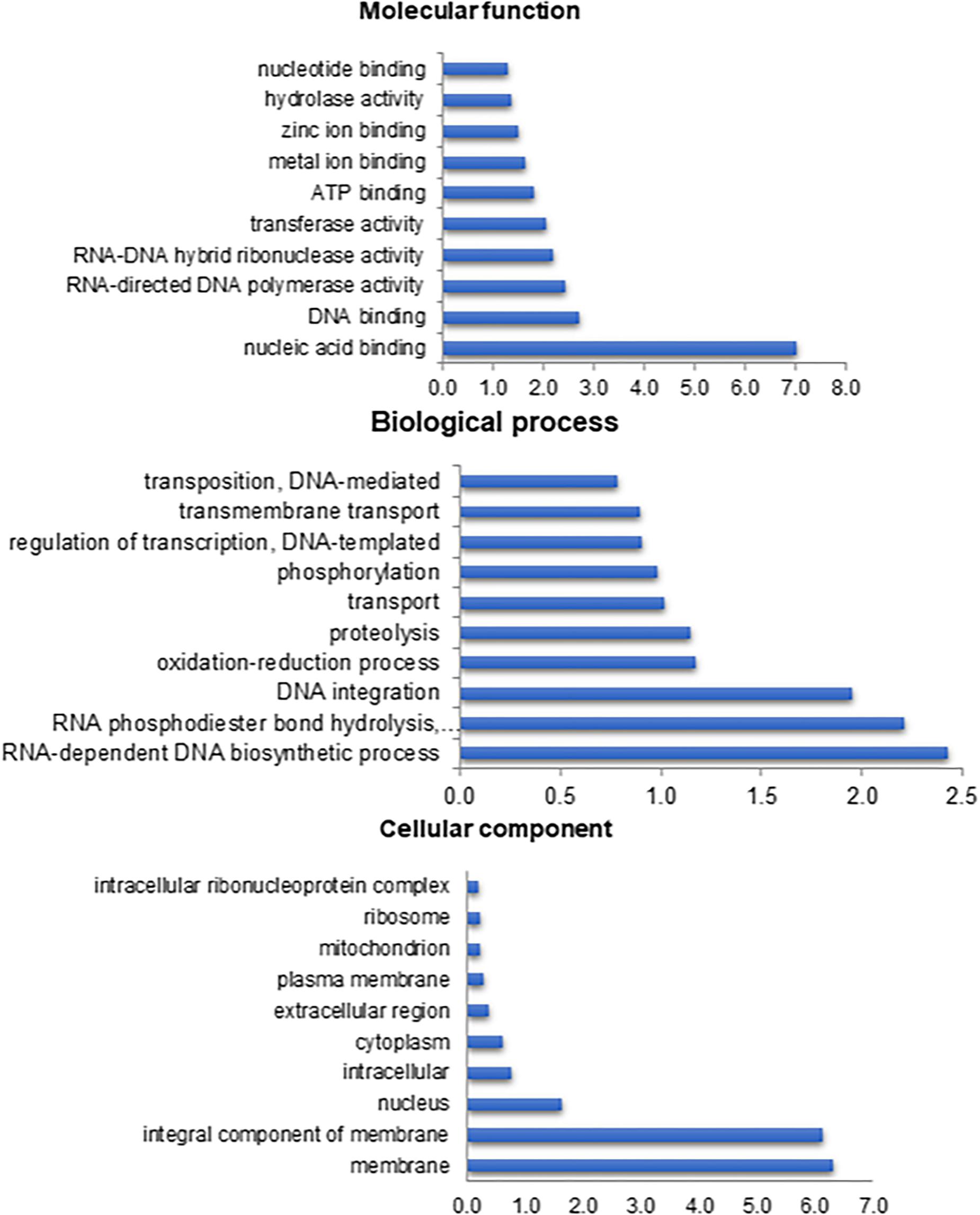
Figure 1. Percentage of Anastrepha fraterculus contigs assigned to a certain gene ontology term as predicted by QuickGO from EBI. Top 10 terms are shown.
Target Genes Related to Post-embryonic Growth/Development and Reproduction Events
We selected 143 different target genes related to biological processes involved in post-embryonic growth/development and reproduction of A. fraterculus, with preference for sequences with annotations reviewed by Swiss-Prot and with experimental evidence. The target genes selected are involved in five biological processes: larval development (54 genes), imaginal disk morphogenesis (22 genes), post-embryonic development (12 genes), sexual reproduction (44 genes), female sex differentiation (2), genital disk anterior/posterior pattern formation (2) and oviposition (7). The results are shown in Supplementary Material S1.
RNAi Machinery Genes Are Present in SA Fruit Fly
We identified 55 genes related to the RNAi machinery in the A. fraterculus transcriptome (Table 1). The sequences of the genes of the miRNA, siRNA, and piRNA pathways, auxiliary factors (from RISC), dsRNA uptake, intracellular transport, antiviral RNAi, nucleases, and lipid metabolism showed most structural and functional units conserved (protein domains) (Supplementary Material S2). The number of the copies at which these genes were found in A. fraterculus is shown in Figure 2.
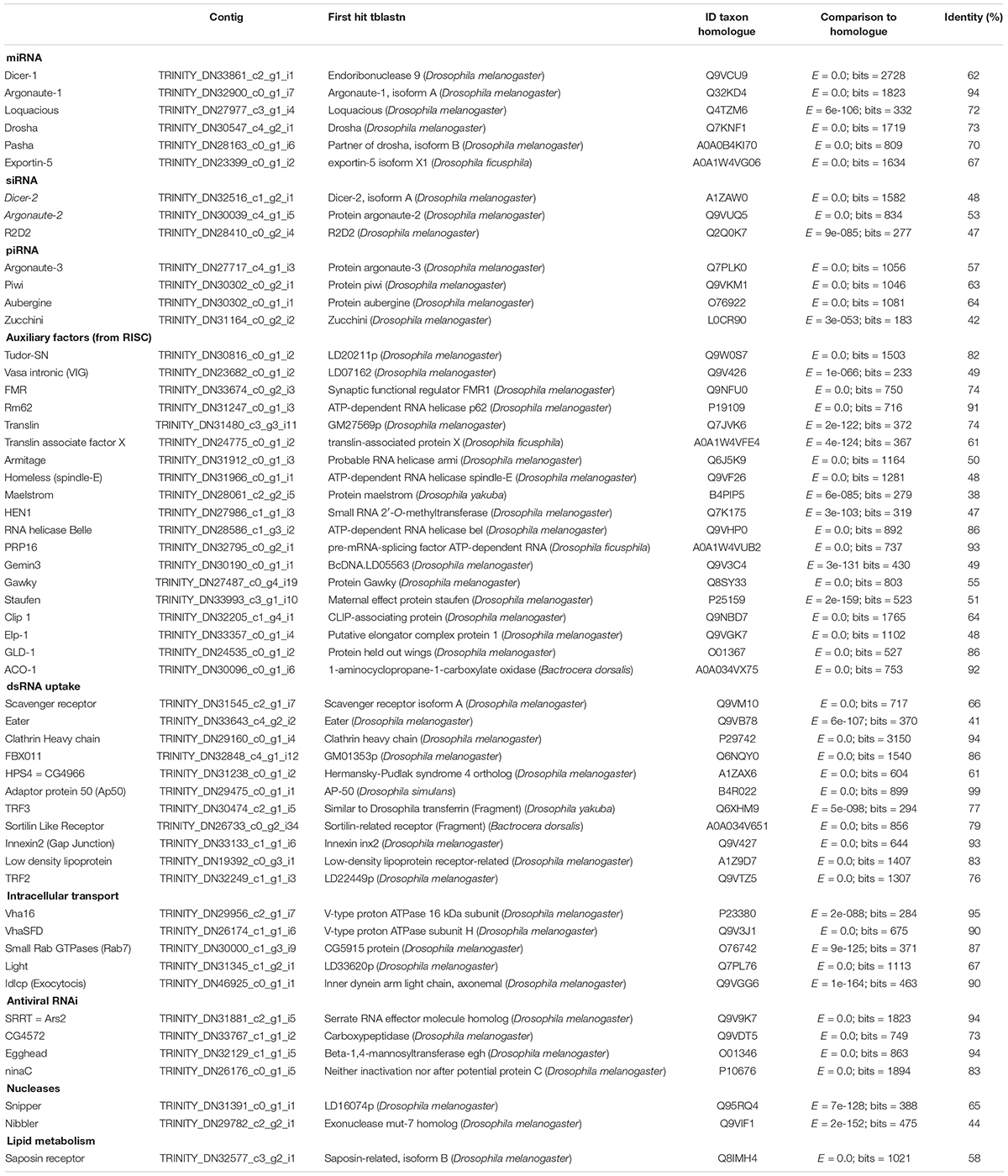
Table 1. Overview of the presence of genes related to the RNAi pathways in the Anastrepha fraterculus transcriptome.
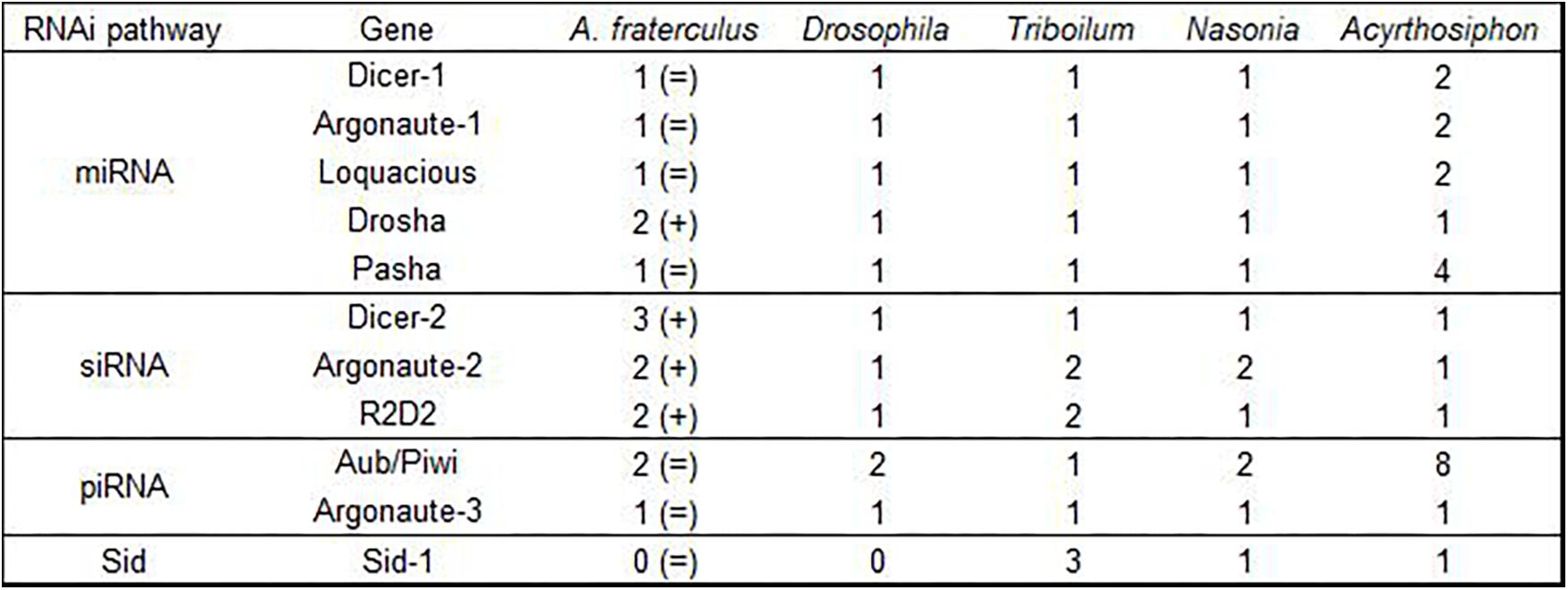
Figure 2. Copy number of the ten RNAi-related genes and SID-1 found in Anastrepha fraterculus transcriptome by Trinity and in other insect species (showed by Dowling et al., 2016). The number of copies showed in A. fraterculus is compared to Drosophila. (=) Same and (+) duplication.
The sequences of Rhagoletis zephyria, B. dorsalis, and C. capitata, from a BLASTp similarity search, showed the closest similarity to A. fraterculus (Supplementary Material S4). The phylogenetic analysis showed that the siRNA pathway gene sequences (Dcr-2 and Ago-2) from A. fraterculus transcriptome were classified in the same clade of D. melanogaster (Figure 3) and the V-ATPase sequence in the same of B. dorsalis clade (Figure 4). The V-ATPase sequence was grouped only with insect sequences, indicating the dsRNA sequence specificity.
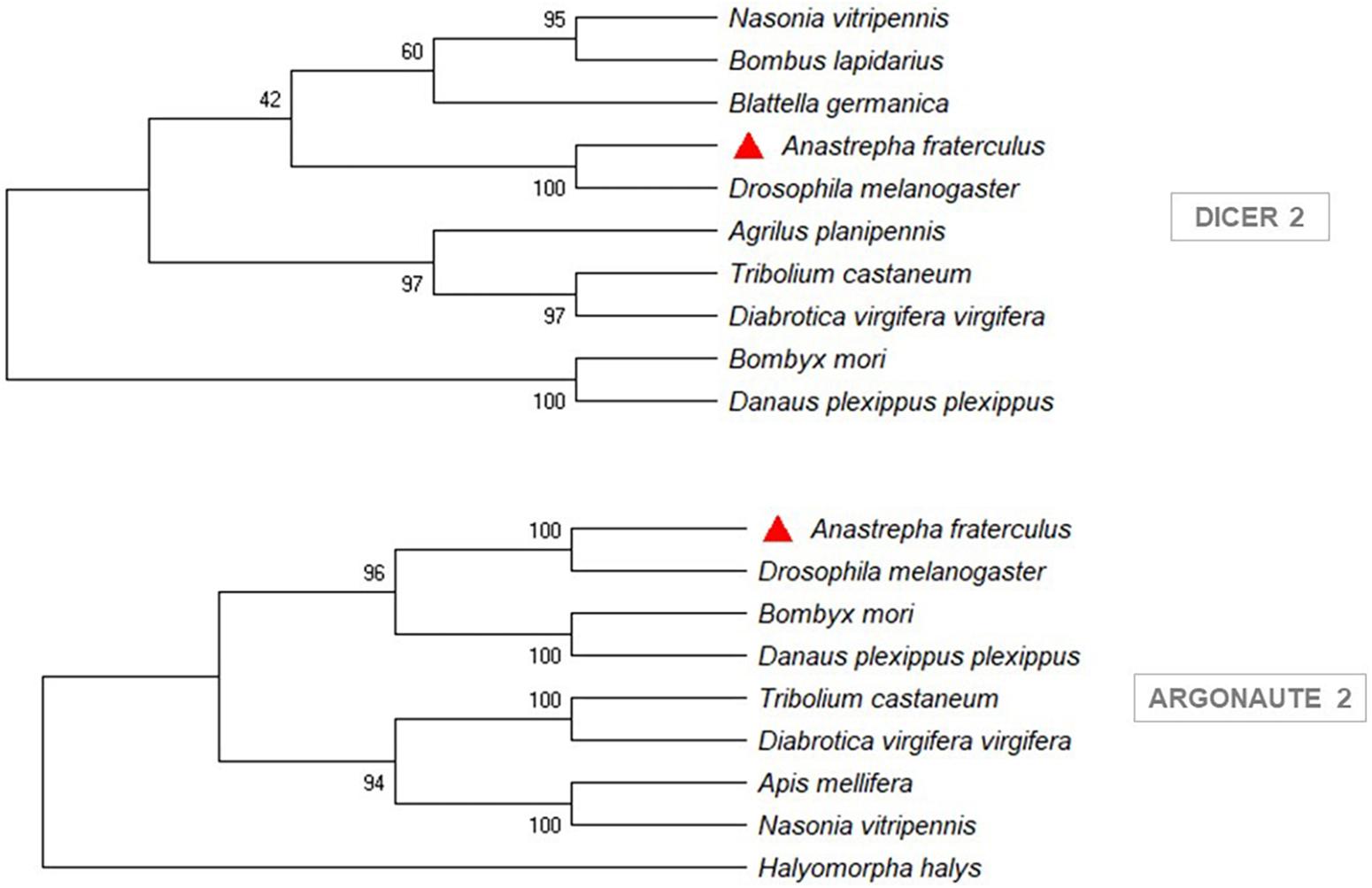
Figure 3. Phylogenetic trees of siRNA pathway genes, Dicer 2 (Dcr-2) and Argonaute 2 (Ago-2). MEGA X was used to construct the phylogenetic trees with Neighbor-Joining method. Anastrepha fraterculus sequence from transcriptome was marked with a red triangle. All accession numbers are shown in Supplementary Table S4.
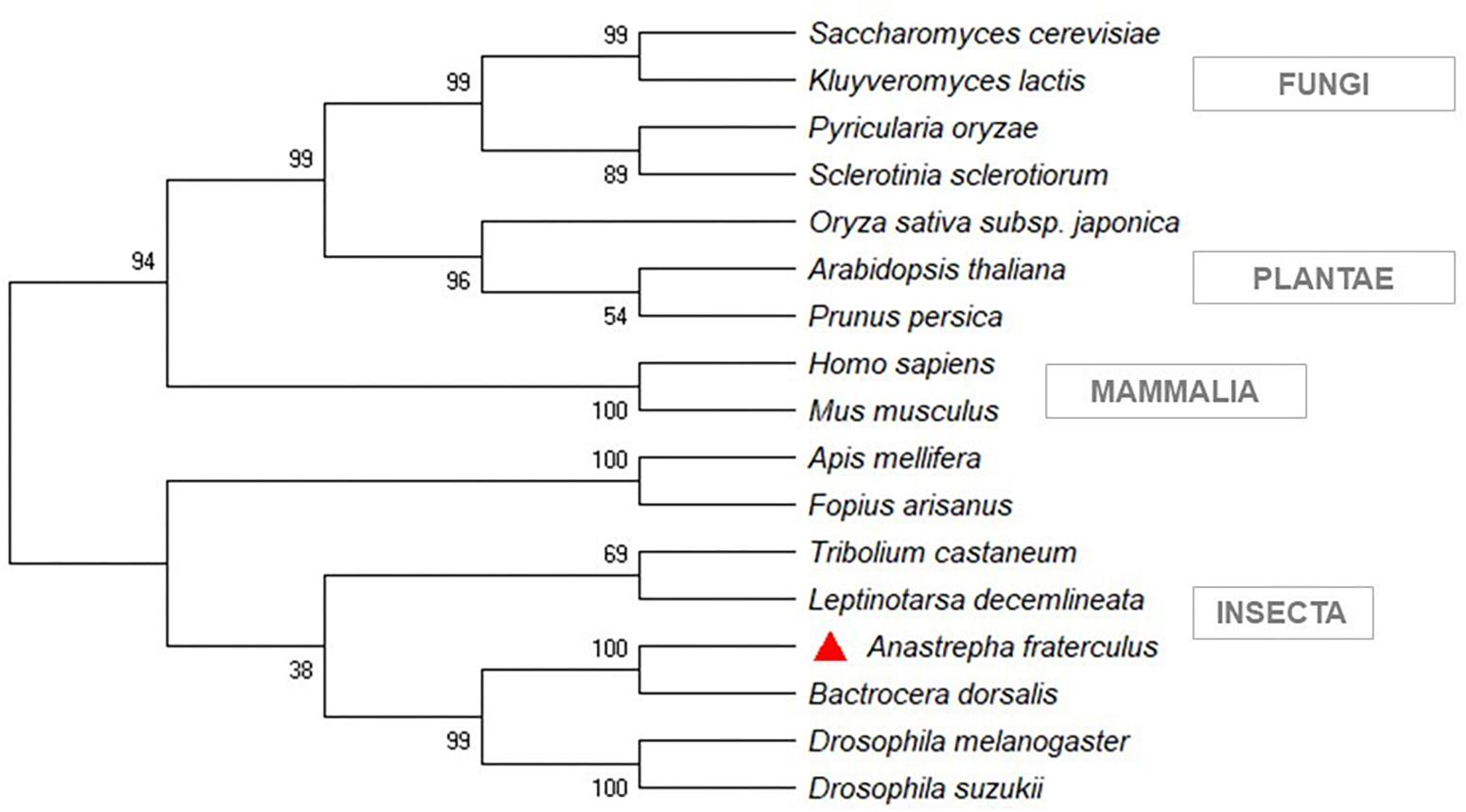
Figure 4. Phylogenetic tree of target gene of silencing, V-ATPase. MEGA X was used to construct the phylogenetic tree with Neighbor-Joining method. Anastrepha fraterculus sequence from transcriptome was marked with a red triangle. All accession numbers are shown in Supplementary Table S4.
Gene Silencing and Mortality in Larval Stages Induced by dsRNA Soaking
Larvae of A. fraterculus soaked in a concentration of 500 ng/μl of dsVTP showed a robust gene silencing as early as 24 h after exposure to dsRNA. The dsVTP soaking resulted in an 85% knockdown relative to dsGFP control and this increased to 100% after 48 h (Figure 5). The silencing effect persisted up to 72 h (p ≤ 0.05). The mortality of A. fraterculus was evaluated for a period of 7 days when larvae reached the pupal stage. Larval mortality started at one day post-soaking (dps), with 5% mortality in larvae soaked with dsVTP. The mortality induced by dsVTP became evident at 2 dps (19%) and rose further to 40% at 7 dps (Figure 6). While the mortality in larvae soaked with dsGFP (control) was 14% at 7 dps.
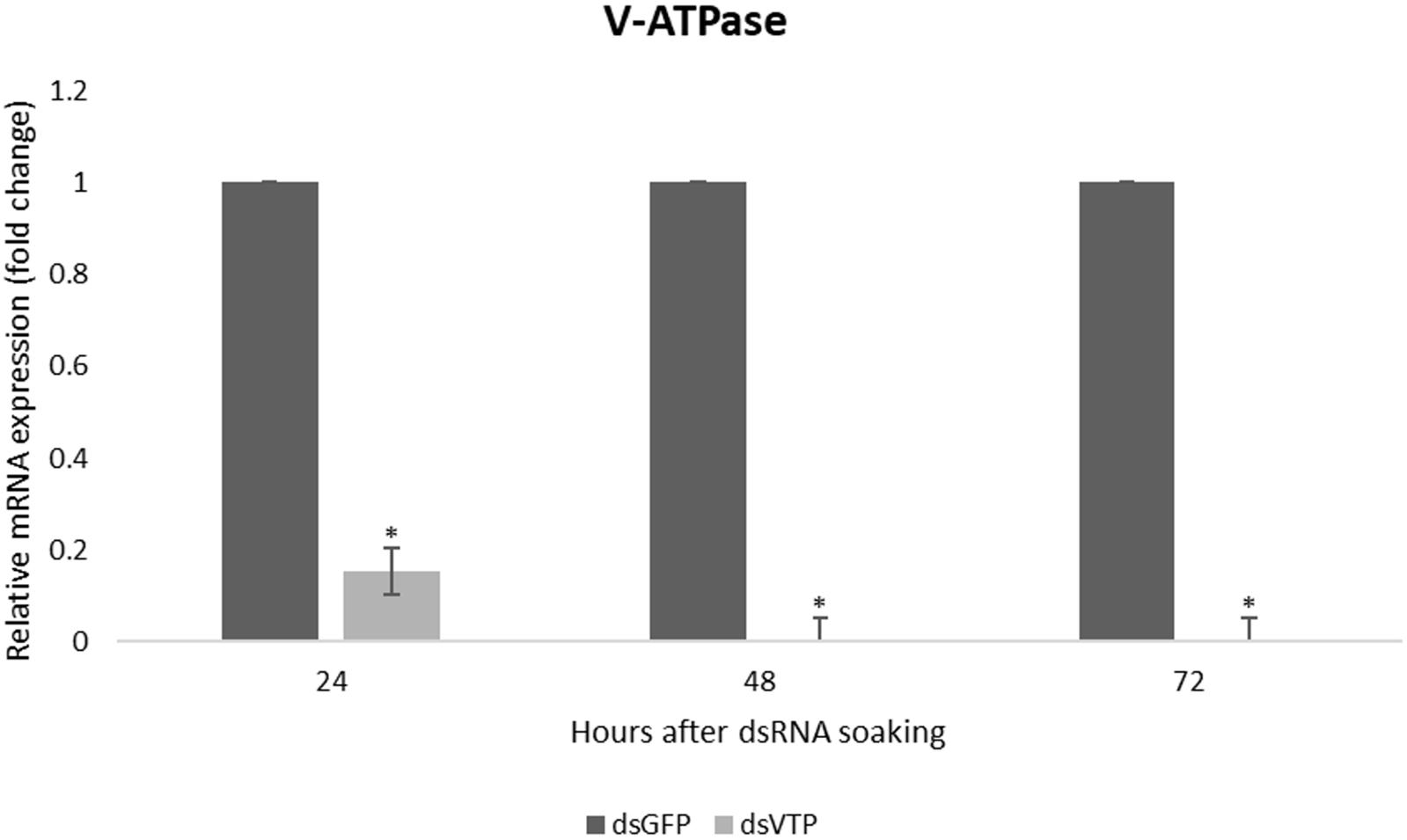
Figure 5. Relative mRNA expression of V-ATPase in Anastrepha fraterculus larvae after 24, 48, and 72 h of soaking in dsRNA (500 ng/μl). The mRNA levels were normalized using α-tubulin and actin as reference genes. The columns represent the mean ± SE (n = 3). *p ≤ 0.05 (t-test).
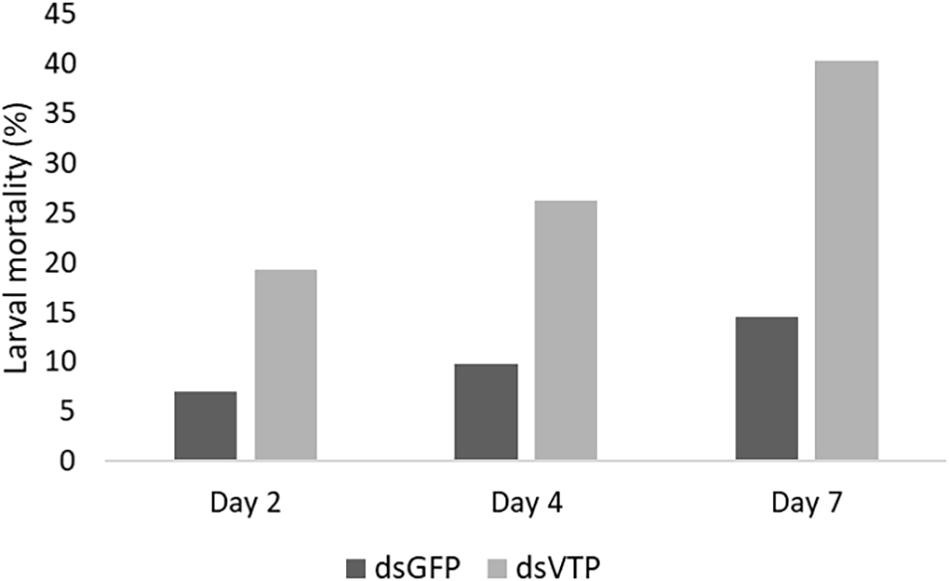
Figure 6. Mortality cumulative of Anastrepha fraterculus larvae (n = 57) after soaking in dsRNA solution (500 ng/μl) from V-ATPase (dsVTP) and GFP control (dsGFP) at 2, 4, and 7 days.
Expression of siRNA Pathway Genes Dcr-2 and Ago-2 dsRNA
The expression of the siRNA genes after the dsRNA soaking in the SA fruit fly larvae confirmed the robust response of the V-ATPase gene. The Dcr-2 mRNA levels were upregulated in the first 24 h after the dsRNA soaking and increased after 48 h; at that moment the V-ATPase mRNA levels were completely downregulated (Figure 7A). The Ago-2 mRNA levels needed a long time to show upregulation: the Ago-2 upregulation was significant at 72 h after soaking (Figure 7B).
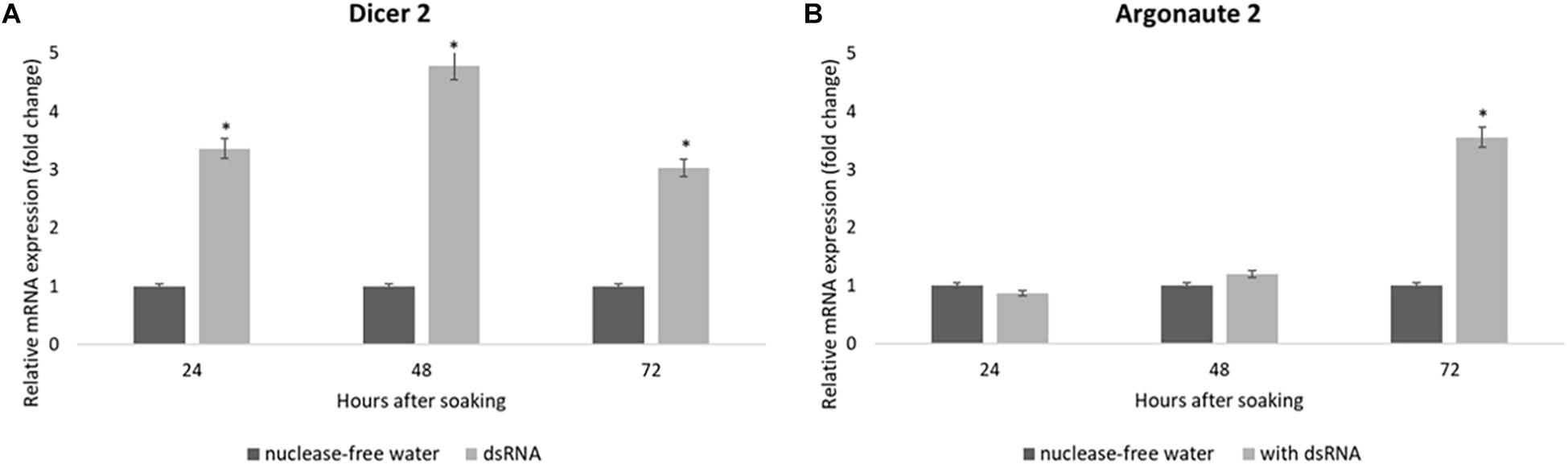
Figure 7. Relative mRNA expression of Dicer-2 (A) and Argonaute-2 (B) in Anastrepha fraterculus larvae in response to dsGFP soaking after 24, 48, and 72 h (500 ng/μl). Nuclease-free water was used as control. The mRNA levels were normalized using α-tubulin and actin as reference genes. The columns represent the mean ± SE (n = 3). *p ≤ 0.05 (t-test).
dsRNA Degradation in A. fraterculus Larvae
We checked the degradation of dsGFP by the dsRNases present in the body fluids (hemolymph) from A. fraterculus larvae. After 1 and 2 h of incubation, no significant degradation of dsRNA was observed (Supplementary Material S3). However, after a longer incubation of 4 h, approximately 40% of the body fluid band intensity was reduced when compared with the initial incubation (0 h).
Discussion
Although A. fraterculus is one of the main pests of fruit crops in the American continent, the lack of genetic information is still an obstacle to unraveling molecular mechanisms of this pest insect. The analysis of the insect transcriptome allows the identification of genes that can be used for pest control through different molecular approaches (Sagri et al., 2014; Garcia et al., 2017). Several studies in the context to develop RNAi for the control of fruit flies species were conducted so far, but only for Anastrepha suspensa (Schetelig et al., 2012), B. dorsalis (Chen et al., 2008, 2011; Suganya et al., 2010, 2011; Li et al., 2011, 2016; Zheng et al., 2012, 2015; Shen et al., 2013; Liu et al., 2015; Peng et al., 2015; Xie et al., 2017), Bactrocera minax (Xiong et al., 2016), and C. capitata (Gabrieli et al., 2016). With this project, more than 84,000 new entries related to A. fraterculus are now available. We also provide a database of 143 novel potential target genes.
The Diamond search analysis showed the greatest quantity of non-significant hits, which indicates that the A. fraterculus transcriptome contains unknown sequences that are not described in the protein sequences databases. Thus, the A. fraterculus transcriptome was screened for the presence of genes related to the RNAi machinery and for further exploration of essential genes to be silenced as target gene by RNAi technology. Similarity searches were performed using as reference the D. melanogaster sequences because it is the species with the complete genome sequenced and fully annotated (Adams et al., 2000) and phylogenetically more closely related to A. fraterculus. This is the first study showing evidence of a functional RNAi mechanism in the SA fruit fly.
Novel Target Genes Found in A. fraterculus Transcriptome
The target genes selected are involved in post-embryonic growth/development and reproduction. Fruit fly pests cause direct damage to production by the puncture for oviposition and the larval development inside the fruit (Aluja, 1994). Thus, the use of RNAi techniques in insect post-embryonic development is crucial for crops protection. Genes involved in the formation of posterior organs during the larval stage, for instance, the ovipositor, are very interesting for RNAi studies. Examples of genes involved in the formation of the posterior organs found in the SA fruit fly transcriptome are hedgehog (hh), homeobox protein abdominal-A (abd-A) and homeobox protein abdominal-B (abd-B). These genes are part of a developmental regulatory system and provide cells with specific positional identities on the anterior-posterior axis (Celniker et al., 1990).
Genes involved in reproductive events such as oviposition regulation can also be screened in the A. fraterculus database. The sex peptide receptor (spr), for example, is a gene involved in the suppression of mating receptivity and induces the egg laying (Yapici et al., 2008). These genes can be studied for dsRNA delivery sequentially or dsRNA-concatemerized.
Three Pathways of the RNAi in SA Fruit Fly
In insects, three RNAi pathways can be distinguished: miRNA, siRNA, and piRNA, based on the Dicers (Dcr) or Argonautes (Ago) enzymes and related small RNAs. Thus, the miRNA pathway includes nuclear Dicer (Drosha/Pasha), cytoplasmic Dicer (Dcr-1/Loquacious), and Ago-1 as core enzymes. The siRNA pathway is activated by exogenous dsRNA and involves Dcr-2/R2D2 and Ago-2. The piRNA is another pathway involved in the defense against transposable elements and includes Ago proteins (Aubergine/Ago-3), which is independent of Dcr (Taning et al., 2016). Sequences representing all core RNAi genes were identified in the A. fraterculus transcriptome with a bitscore ≥ 150 and E-value ≤ 1e-5. The domains of the Drosha and Dcr proteins were conserved in A. fraterculus (Supplementary Material S2). We found some Dcr domains as following: amino-terminal DExH-box helicase domains, PAZ domain, two RNaseIII domains, and carboxy-terminal dsRNA-binding domain (dsRBD) (Carmell and Hannon, 2004). However, some components of the Dcr family differ from this general arrangement; for instance, some of these lack a functional helicase domain or a PAZ domain, or the number of dsRBD is ranging from zero to two (Macrae et al., 2006). The sequence of Dcr-2 in A. fraterculus does not show a dsRBD domain.
Unlike Dcr, the PAZ and amino-terminal DExH-box helicase domains are not present in Drosha. Two cofactors with the conserved domains DSRM, Pasha, and Loquacious, were also identified in A. fraterculus. These proteins are needed to interact with the RNase III, Drosha, and Dcr-1, respectively (Carmell and Hannon, 2004). For R2D2, we found sequences inside the defined threshold, but without conserved domains. R2D2 can form the Dcr-2/R2D2 complex and bind to siRNA to enhance sequence-specific messenger RNA degradation mediated by the RNA-induced silencing complex (RISC).
The Ago superfamily is divided into Ago and Piwi clades. Drosophila species have two Ago members (Ago-1 and Ago-2) and three Piwi members (Piwi, Aubergine, and Ago-3) (Cerutti et al., 2000; Cox et al., 2000). In these insects, Ago-2 is involved in the siRNA-directed mRNA cleavage, and Ago-1 mainly mediates miRNA-directed translational inhibition. Some Argonaute proteins can cleave the target mRNA, while others affect their nucleic acid targets using alternative mechanisms (Ketting, 2011). Two domains characterize ago proteins: PAZ and C-terminal Piwi (Cerutti et al., 2000). In the A. fraterculus transcriptome, we have identified the five members of the Ago protein superfamily, with the PAZ and Piwi conserved domains.
The third pathway of RNAi, the piRNA, involves the proteins Aubergine, Ago-3, Piwi, and Zucchini (Hartig et al., 2007). Zucchini is an endoribonuclease that has a role in piRNA maturation. With the absence of this protein, the transposons are no longer repressed and no piRNAs can be detectable (Pane et al., 2007). In A. fraterculus we found sequences of Zucchini protein with the presence of conserved domains superfamily PLD (Phospholipase D).
Duplication and Loss of the RNAi-Related Genes in A. fraterculus
The biogenesis of the RNAi pathways and related-proteins is similar among eukaryotes, however, throughout evolution duplications and losses of genes have occurred in several insects. Duplications or loss of RNAi-related genes can lead to higher or lower functionalization of the RNAi mechanisms and could explain differences in the efficacy of RNAi in different insect groups (Dowling et al., 2016).
Our transcriptome analysis indicated gene duplication and gene loss events in A. fraterculus. Possible duplicates of Drosha, Ago-2 and R2D2 were found in the SA fruit fly transcriptome compared to D. melanogaster. Dowling et al. (2016) also found possible duplicates of Ago-2 in transcriptomes of other order insects, as Peruphasma schultei (Phasmatodea), Prorhinotermes simplex (Isoptera) and Pseudomallada prasinus (Neuroptera). These authors suggested that Ago-2 was present in two copies in the last common ancestor of insects. Is it possible that SA fruit fly has three copies to Dcr-2, while D. melanogaster has only one copy for Dcr-2? It is known that insects inherited a complete RNAi system from their common ancestor and diversified and expanded this original system (Dowling et al., 2016). One example of this is the piRNA pathway (Piwi/Aub) in insects that acts as a defense against transposons in the germ line. In the A. fraterculus transcriptome of this study, this gene is present with two copies, while Hemiptera species as Acyrthosiphon pisum has eight copies for this piRNA gene. Dowling et al. (2016) considered that homologs of both Piwi/Aub and Ago-3 could be present in the last common ancestor of insects in multiple copies. Although we have used a mix of all developmental stages of SA fruit fly to generate a comprehensive transcriptome, it must be remarked that the final conclusion that a gene is lost from a species cannot be made since the gene in question may not have been expressed or very lowly expressed (Dowling et al., 2016).
SA Fruit Fly Has Auxiliary Factors (From RISC)
We found 19 intracellular factors that are associated with the activity of the RISC. In the RISC assembly for exogenous dsRNA in D. melanogaster, the siRNA duplex is transferred from complex B to the RISC-loading complex (RLC), including Dcr-2 and R2D2. Next, C3PO (translin and TRAX) binds to the RLC and the RISC [consisting of the Dcr-1, Tudor-Staphylococcal nuclease (Tudor-SN), vasa intronic gene (VIG), FMR, and Ago-2 subunits] to produce the holoRISC (Jaendling and McFarlane, 2010). These sequences were found in our A. fraterculus transcriptome, holding conserved main domains and with an identity between 49 and 82% compared to D. melanogaster (Supplementary Material S2).
The nucleases involved in piRNA biogenesis, Armitage and Homeless (spindle-E), showed long sequences (>4,000 nc) in A. fraterculus, while small fragments represented Maelstrom. Genes that encode Gawky, an RNAi effector, Staufen, an RNA-binding protein, Elp-1, a component of the core elongator complex involved in the RNAi, and Clp-1, a kinase that can phosphorylate siRNAs, as well the RNA helicases Rm62 and Belle also showed long sequences (Findley, 2003; Vagin et al., 2006). The DEAD-box RNA helicase Belle has a function in the endo-siRNA pathway, interacting with Ago-2 and endo-siRNA-producing loci and is localized in condensing chromosomes in a Dcr-2- and Ago-2-dependent way (Cauchi et al., 2008). Another, the DEAD-box RNA helicase PRP16 has a key role in the pre-mRNA splicing and was found in the A. fraterculus transcriptome with an identity of 93% as compared to Drosophila sequences (Ansari and Schwer, 1995).
dsRNA Uptake Genes
With the exception of SID-1, all dsRNA uptake components were found in the A. fraterculus transcriptome. This confirms the idea that this gene is absent in Diptera. Unlike the typical model organism of C. elegans, which uses SID-1 to transport dsRNA into the cells, Drosophila species do not have SID-1 orthologues (Huvenne and Smagghe, 2010), therefore two scavenger receptors, namely SR-CI and Eater, were proven to undertake the transport function in Drosophila (Ulvila et al., 2006). Scavenger receptors play a role in phagocytosis and act for large molecules and microbes (Prentice et al., 2015). In A. fraterculus, scavenger receptors were found only for Eater and SR-CI sequences; this last one with conserved domains (Supplementary Material S2). Other genes coding for proteins involved in endocytosis were found in A. fraterculus, including HPS4 (Hermansky-Pudlak Syndrome 4), a factor involved in the regulation of the combination of late endosomes and RNA-processing GW bodies, FBX011 (F-box motif, Beta-helix motif), a regulator of endosome trafficking and the clathrin heavy chain (chc), which is needed for clathrin-mediated endocytosis (Swevers et al., 2013).
Nucleases in SA Fruit Fly Development Transcriptome
Nucleases sequences were identified only for Snipper, a histone involved in mRNA metabolism, siRNA degradation, and apoptosis, and for the Nibbler, a nuclease described in Drosophila and involved in the processing of 3′ends of miRNAs (Swevers et al., 2013). We identified the conserved domains ERI-1 3′ exoribonuclease for Snipper sequences in our A. fraterculus transcriptome (Supplementary Material S2).
Presence of Genes Involved in RNAi Efficacy
We found five intracellular transport components as classified by Yoon et al. (2016). The components Vha16 (Vacuolar H+ ATPase 16kD subunit 1) and VhaSFD (Vacuolar [+] ATPase SFD subunit) related to proton transport, Rab7 (Small Rab GTPases) involved in endocytosis process, Light involved in lysosomal transport, and Idlcp involved in exocytosis process.
Four antiviral RNAi were found in the A. fraterculus transcriptome, Ars2, a regulator involved in innate immunity via the siRNAs processing machinery by restricting the viral RNA production, CG4572, a protease implicated in systemic silencing and antiviral RNAi, Egghead (egh), a seven-transmembrane-domain glycosyltransferase with innate immunity against RNA virus, and ninaC, a protein involved in vesicle transport. All antiviral RNAi components were identified with conserved main domains (Supplementary Material S2).
The Saposin receptor, which is involved in lipid metabolism, was identified with Saposin A and Saposin B conserved domains in A. fraterculus (Supplementary Material S2). Saposin is a small lysosomal protein that serves as the activator of various lysosomal lipid-degrading enzymes (Darmoise et al., 2010).
Evidence for the Sensitivity of Larval Stages of A. fraterculus to RNAi
The functionality of the RNAi mechanism in A. fraterculus was demonstrated using a dsRNA targeting V-ATPase, evaluated by an in-house soaking bioassay. V-ATPases are ubiquitous holoenzymes among eukaryotes (Finbow and Harrison, 1997). These enzymes are composed of two subcomplexes, the cytosolic V1-domain, where ATP binding and hydrolysis take place, and a transmembranous V0-domain, through which protons are translocated (Vitavska et al., 2003). The V-ATPase sequence analyzed in A. fraterculus belongs to the V0-domain (Supplementary Material S2). The V-ATPases uses the energy produced from ATP hydrolysis to transport protons across intracellular and plasma membranes of eukaryotic cells (Nelson et al., 2000). Although the V0 complex plays a key role in protons translocation, just a few studies aiming V0-domain as the target were published with insects (Ahmed, 2016). Therefore, we synthesized a dsRNA fragment with 483 bp length targeting V-ATPase V0-domain gene.
The results presented here indicate that A. fraterculus has a sensitivity to RNAi. We demonstrated that a small dose of dsRNA (500 ng) administered by soaking for 30 min could produce significant RNAi responses (target-gene knockdown and mortality). We attributed the high knockdown efficiency in A. fraterculus to some factors confirmed in this study by transcriptome analysis and dsRNA delivery assay. These factors confirm the presence of RNAi machinery genes, the activation of siRNA pathway genes (especially Dcr-2 upregulation at 48 h after dsRNA delivery), few nucleases, and factors related to uptake, that need to be clarified.
The effective response of gene silencing as showed by A. fraterculus at 48 h after dsRNA soaking resulted in mortality of these larvae. The V-ATPase sequence from the A. fraterculus transcriptome contains the VMA21, a short domain that has two transmembrane helices (Supplementary Material S2). The product of the VMA21 is characterized by an 8.5 kDa integral membrane and contains a C-terminal di-lysine motif that is needed for retention in the endoplasmic reticulum, and disruption of the gene causes failure to assemble a stable V0, rapid turnover of Vph1p subunit (that contains charged residues that are essential for proton translocation) and consequent loss of V-ATPase function (Hill and Stevens, 1994). In other dipterans species, the V-ATPase knockdown responses were variable. In B. dorsalis, the ingestion of 2000 ng V-ATPase D (V1-domain) dsRNA through diet caused only 35% of gene silencing after 4 days (Li et al., 2011). The neonate larvae of D. melanogaster when soaked in 500 ng of V-ATPase E (V1-domain) dsRNA caused a decrease of 49% in gene expression and feeding larvae caused 56% knockdown and 70% mortality (Whyard et al., 2009). These studies suggest that the silencing of V-ATPase subunits demonstrate variable results related to subunit and target species.
Dcr-2 and Ago-2 Respond to dsRNA Exposure
The upregulation of the Dcr-2 at 24 h after the dsRNA soaking demonstrated that the RNAi response in A. fraterculus is active. The Dcr-2 is a specific ribonuclease that initiates RNAi by cleaving dsRNA substrates into small fragments (Macrae et al., 2006). The PAZ and RNase III domains from Dcr-2 found in our A. fraterculus transcriptome are shown in the Supplementary Material S2.
dsRNA Is Degraded in A. fraterculus Body Fluid
Degradation of dsGFP (0.5 mg/ml) was observed only after 4 h of incubation. Liu et al. (2012) verified dsGFP degradation only after 3 h of incubation using hemolymph of Bombyx mori larvae. On the other hand, the authors verified that dsGFP degradation in gut juice occurred at less than 10 min. Christiaens et al. (2014) demonstrated an intense dsRNA degradation shortly after 1 h in aphid hemolymph (A. pisum).
According to Singh et al. (2017), usually, a high concentration of body fluid from dipteran insects is required to degrade dsRNA. For A. suspensa, for example, Singh et al. (2017) showed that 4.44 mg/ml of body fluid was needed to degrade 50% of dsRNA, while for Spodoptera frugiperda a very low concentration of hemolymph (0.11 mg/ml) was enough to degrade dsRNA within an hour. Singh et al. (2017) also suggested that the expression of dsRNases could be lower in Diptera species when compared to other orders. This was noted in the present work in which only a nuclease (Snipper) involved in the siRNA degradation could be identified based on the lists previously reported (Swevers et al., 2013; Prentice et al., 2015; Yoon et al., 2016). Recently, Prentice et al. (2019) confirmed the impact of one specific ribonuclease in the gut of the African sweet potato weevil (SPW), Cylas puncticollis. Two nucleases were identified by transcriptome analysis and they were demonstrated to affect the dsRNA stability in the gut when dsRNA was delivered by oral feeding.
Conclusion
Our project made available more than 84,000 new entries related to the developmental of A. fraterculus and generated a database of 143 novel and different target genes to dsRNA bioassays. This transcriptome database is a handy tool for research on the SA fruit fly, especially in studies with a focus on RNAi. The identification of the RNAi machinery genes combined with dsRNA soaking, siRNA genes expression and dsRNA degradation bioassays demonstrated that an RNAi response is active in A. fraterculus. The presence of RNAi machinery and efficient genes for silencing confirms the sensitivity A. fraterculus to produce a robust RNAi response.
Interestingly, we demonstrated that soaking of the larval stages in dsV-ATPase lead to a strong gene-silencing and this concurred with strong mortality of 40%. This assay by soaking demonstrates that dsRNA delivery can also be effected via the cuticle of the insect (environmental RNAi) (Niu et al., 2018). Our data demonstrated the existence of a functional RNAi machinery in A. fraterculus and an easy and robust physiological bioassay with the larval stages that can be used for in vivo selection of target genes for RNAi-based control of fruit fly pests.
Data Availability
All datasets for this study are included in the manuscript and the Supplementary Files.
Author Contributions
ND, DC, GS, and MZ contributed the conception and design of the study. ND and FK organized the database. ND performed the statistical analysis and wrote the first draft of the manuscript. ND, LR, DN, GS, and MZ wrote the sections of the manuscript. All authors contributed to the manuscript revision, read and approved the final version of the manuscript.
Funding
We thank the support from the National Council for Scientific and Technological Development (CNPq), the Coordination of Improvement of Higher Education Personnel (CAPES) in Brazil, and the Foundation Research-Flanders (FWO-Vlaanderen) in Belgium. We also thank the support of COST (European Cooperation in Science and Technology) under grant agreement No. CA15223.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Alexandra Hart for the English editing.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphys.2019.00794/full#supplementary-material
Footnotes
- ^ http://www.usadellab.org/cms/?page=trimmomatic
- ^ http://www.bioinformatics.babraham.ac.uk/projects/fastqc
- ^ http://trinityrnaseq.sourceforge.net/
- ^ http://www.ebi.ac.uk/QuickGO/GAnnotation
- ^ http://primer3.ut.ee/
References
Adams, M. D., Celniker, S. E., Holt, R. A., Evans, C. A., Gocayne, J. D., Amanatides, P. G., et al. (2000). The genome sequence of Drosophila melanogaster. Science 287, 2185–2195. doi: 10.1126/science.287.5461.2185
Ahmed, M. A. M. (2016). RNAi-based silencing of genes encoding the vacuolar-ATPase subunits a and c in pink bollworm (Pectinophora gossypiella). Afr. J. Biotechnol. 15, 2547–2557. doi: 10.5897/AJB2016.15611
Aluja, M. (1994). Bionomics and management of Anastrepha. Annu. Rev. Entomol. 39, 155–178. doi: 10.1146/annurev.en.21.010176.001255
Andrade, E. C., and Hunter, W. B. (2017). RNAi feeding bioassay: development of a non-transgenic approach to control Asian citrus psyllid and other hemipterans. Entomol. Exp. Appl. 162, 389–396. doi: 10.1111/eea.12544
Ansari, A., and Schwer, B. (1995). SLU7 and a novel activity, SSF1, act during the PRP16-dependent step of yeast pre-mRNA splicing. EMBO J. 14, 4001–4009. doi: 10.1002/j.1460-2075.1995.tb00071.x
Benemann, D. P., Nohato, A. M., Vargas, L., Avila, L. A., and Agostinetto, D. (2017). Identification and validation of reference genes for the normalization in real-time RT-qPCR on rice and red rice in competition, under different nitrogen doses. Planta Daninha 35, 1–9. doi: 10.1590/s0100-83582017350100015
Böckmann, E., Köppler, K., Hummel, E., and Vogt, H. (2014). Bait spray for control of European cherry fruit fly: an appraisal based on semi-field and field studies. Pest Manag. Sci. 70, 502–509. doi: 10.1002/ps.3621
Buchfink, B., Xie, C., and Huson, D. H. (2015). Fast and sensitive protein alignment using DIAMOND. Nat. Methods 12, 59–60. doi: 10.1038/nmeth.3176
Carmell, M. A., and Hannon, G. J. (2004). RNase III enzymes and the initiation of gene silencing. Nat. Struct. Mol. Biol. 11, 214–218. doi: 10.1038/nsmb729
Cauchi, R. J., Davies, K. E., and Liu, J. L. (2008). A motor function for the DEAD-box RNA helicase, Gemin3, in Drosophila. PLoS Genet. 4:e1000265. doi: 10.1371/journal.pgen.1000265
Celniker, S. E., Sharma, S., Keelan, D. J., and Lewis, E. B. (1990). The molecular genetics of the bithorax complex of Drosophila: cis-regulation in the abdominal-B domain. EMBO J. 9, 4277–4786. doi: 10.1101/gad.3.9.1424
Cerutti, L., Mian, N., and Bateman, A. (2000). Domains in gene silencing and cell differentiation proteins: the novel PAZ domain and redefinition of the Piwi domain. Trends Biochem. Sci. 25, 481–482. doi: 10.1016/S0968-0004(00)01641-8
Chen, S., Dai, S., Lu, K., and Chang, C. (2008). Female-specific doublesex dsRNA interrupts yolk protein gene expression and reproductive ability in oriental fruit fly, Bactrocera dorsalis (Hendel). Insect Biochem. Mol. Biol. 38, 155–165. doi: 10.1016/j.ibmb.2007.10.003
Chen, S., Lu, K., Dai, S., Li, C., Shieh, C., and Chang, C. (2011). Display female-specific doublesex RNA interference in early generations of transformed oriental fruit fly, Bactrocera dorsalis (Hendel). Pest Manag. Sci. 67, 466–473. doi: 10.1002/ps.2088
Christiaens, O., Dzhambazova, T., Kostov, K., Salvatore Arpaia, S., Reddy Joga, M., Urru, I., et al. (2018). Literature review of baseline information on RNAi to support the environmental risk assessment of RNAi-based GM plants. EFSA Support. Public. 15:1424E. doi: 10.2903/sp.efsa.2018.EN-1424
Christiaens, O., Swevers, L., and Smagghe, G. (2014). DsRNA degradation in the pea aphid (Acyrthosiphon pisum) associated with lack of response in RNAi feeding and injection assay. Peptides 53, 307–314. doi: 10.1016/j.peptides.2013.12.014
Cox, D. N., Chao, A., and Lin, H. (2000). Piwi encodes a nucleoplasmic factor whose activity modulates the number and division rate of germline stem cells. Development 127, 503–514.
Darmoise, A., Maschmeyer, P., and Winau, F. (2010). The immunological functions of saposins. Adv. Immunol. 105, 25–62. doi: 10.1016/S0065-2776(10)05002-9
Dias, N. P., Zotti, M. J., Montoya, P., Carvalho, I. R., and Nava, D. E. (2018). Fruit fly management research: a systematic review of monitoring and control tactics in the world. Crop Prot. 112, 187–200. doi: 10.1016/j.cropro.2018.05.019
Dowling, D., Pauli, T., Donath, A., Meusemann, K., Podsiadlowski, L., Petersen, M., et al. (2016). Phylogenetic origin and diversification of RNAi pathway genes in insects. Genome Biol. Evol. 8, 3784–3793. doi: 10.1093/gbe/evw281
Elbashir, S. M., Harborth, J., Lendeckel, W., Yalcin, A., Weber, K., and Tuschl, T. (2001). Duplexes of 21 ± nucleotide RNAs mediate RNA interference in cultured mammalian cells. Nature 411, 494–498. doi: 10.1038/35078107
Finbow, M. E., and Harrison, M. A. (1997). The vacuolar H+-ATPase: a universal proton pump of eukaryotes. Biochem. J. 324, 697–712. doi: 10.1042/bj3240697
Findley, S. D. (2003). Maelstrom, a Drosophila spindle-class gene, encodes a protein that colocalizes with Vasa and RDE1/AGO-1 homolog, Aubergine, in nuage. Development 130, 859–871. doi: 10.1242/dev.00310
Gabrieli, P., Scolari, F., Di Cosimo, A., Savini, G., Fumagalli, M., Gomulski, L. M., et al. (2016). Sperm-less males modulate female behaviour in Ceratitis capitata (Diptera: Tephritidae). Insect Biochem. Mol. Biol. 79, 13–26. doi: 10.1016/j.ibmb.2016.10.002
Garcia, R. A., Pepino Macedo, L. L., Do Nascimento, D. C., Gillet, F. X., Moreira-Pinto, C. E., Faheem, M., et al. (2017). Nucleases as a barrier to gene silencing in the cotton boll weevil, Anthonomus grandis. PLoS One 12:e0189600. doi: 10.1371/journal.pone.0189600
Gonçalves, R. S., Nava, D. E., Pereira, H. C., Lisbôa, H., Grützmacher, A. D., and Valgas, R. A. (2013). Biology and fertility life table of Aganaspis pelleranoi (Hymenoptera: Figitidae) in larvae of Anastrepha fraterculus and Ceratitis capitata (Diptera: Tephritidae). Ann. Entomol. Soc. Am. 106, 791–798. doi: 10.1603/AN13044
Hartig, J. V., Tomari, Y., and Förstemann, K. (2007). piRNAs - The ancient hunters of genome invaders. Genes Dev. 21, 1707–1713. doi: 10.1101/gad.1567007
Hill, K. J., and Stevens, E. H. (1994). Vma21p is a yeast membrane protein retained in the endoplasmic reticulum by a di-lysine motif and is required for the assembly of the vacuolar H(+)-ATPase complex. Mol. Biol. Cell 5, 1039–1050. doi: 10.1091/mbc.5.9.1039
Huvenne, H., and Smagghe, G. (2010). Mechanisms of dsRNA uptake in insects and potential of RNAi for pest control: a review. J. Insect Physiol. 56, 227–235. doi: 10.1016/j.jinsphys.2009.10.004
Jaendling, A., and McFarlane, R. J. (2010). Biological roles of translin and translin-associated factor-X: RNA metabolism comes to the fore. Biochem. J. 429, 225–234. doi: 10.1042/BJ20100273
Katoch, R., Sethi, A., Thakur, N., and Murdock, L. L. (2013). RNAi for insect control: current perspective and future challenges. Appl. Biochem. Biotechnol. 171, 847–873. doi: 10.1007/s12010-013-0399-4
Ketting, R. F. (2011). The many faces of RNAi. Dev. Cell. 20, 148–161. doi: 10.1016/j.devcel.2011.01.012
Lanzavecchia, S. B., Juri, M., Bonomi, A., Gomulski, L., Scannapieco, A. C., Segura, D. F., et al. (2014). Microsatellite markers from the “South American fruit fly” Anastrepha fraterculus: a valuable tool for population genetic analysis and SIT applications. BMC Genet. 15(Suppl. 2):S13. doi: 10.1186/1471-2156-15-S2-S13
Li, X., Zhang, M., and Zhang, H. (2011). RNA interference of four genes in adult Bactrocera dorsalis by feeding their dsRNAs. PLoS One 6:e17788. doi: 10.1371/journal.pone.0017788
Li, Y.-L., Hou, M.-Z., Shen, G.-M., Lu, X.-P., Wang, Z., Jia, F.-X., et al. (2016). Functional analysis of five trypsin-like protease genes in the oriental fruit fly, Bactrocera dorsalis (Diptera: Tephritidae). Pestic. Biochem. Physiol. 136, 52–57. doi: 10.1016/j.pestbp.2016.08.004
Liu, G., Wu, Q., Li, J., Zhang, G., and Wan, F. (2015). RNAi-mediated knock-down of transformer and transformer 2 to generate male-only progeny in the oriental fruit fly, Bactrocera dorsalis (Hendel). PLoS One 10:e0128892. doi: 10.1371/journal.pone.0128892
Liu, J., Smagghe, G., and Swevers, L. (2012). Transcriptional response of BmToll9-1 and RNAi machinery genes to exogenous dsRNA in the midgut of Bombyx mori. J. Insect Physiol. 59, 646–654. doi: 10.1016/j.jinsphys.2012.05.016
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Macedo, M., Avila, S., Zucchi, R. A., and Farias, A. F. (2017). “Mid-level image representation for fruit fly identification (Diptera: Tephritidae),” in Proceedings of the IEEE International Conference on eScience, (Auckland), 1–9.
Macrae, I. J., Zhou, K., Li, F., Repic, A., Brooks, A. N., Cande, W. Z., et al. (2006). Structural basis for double-stranded RNA processing by Dicer. Science 311, 195–198. doi: 10.1126/science.1121638
Nelson, N., Perzov, N., Cohen, A., Hagai, K., Padler, V., and Nelson, H. (2000). The cellular biology of proton-motive force generation by V-ATPases. J. Exp. Biol. 203, 89–95.
Niu, J., Taning, C. N. T., Christiaens, O., Smagghe, G., and Wang, J. J. (2018). Rethink RNAi in insect pest control: challenges and perspectives. Adv. Insect Physiol. 55, 1–17. doi: 10.1016/bs.aiip.2018.07.003
Nunes, A. M., Costa, K. Z., Faggioni, K. M., de Lourdes Zamboni Costa, M., da Silva, Gonçalves, R., et al. (2013). Dietas artificiais para a criação de larvas e adultos da mosca-das-frutas sul-americana. Pesqui. Agropecu. Bras. 48, 1309–1314. doi: 10.1590/S0100-204X2013001000001
Pane, A., Wehr, K., and Schüpbach, T. (2007). Zucchini and squash encode two putative nucleases required for rasiRNA production in the Drosophila germline. Dev. Cell 12, 851–862. doi: 10.1016/j.devcel.2007.03.022
Peng, W., Zheng, W., Handler, A. M., and Zhang, H. (2015). The role of the transformer gene in sex determination and reproduction in the tephritid fruit fly, Bactrocera dorsalis (Hendel). Genetica 143, 717–727. doi: 10.1007/s10709-015-9869-7
Prentice, K., Pertry, I., Christiaens, O., Bauters, L., Bailey, A., Niblett, C., et al. (2015). Transcriptome analysis and systemic RNAi response in the African sweetpotato weevil (Cylas puncticollis, Coleoptera, Brentidae). PLoS One 10:e0115336. doi: 10.1371/journal.pone.0115336
Prentice, K., Smagghe, G., Godelieve, G., and Christiaens, O. (2019). Nuclease activity decreases the RNAi response in the sweetpotato weevil Cylas puncticollis. Insect Biochem. Mol. Biol. 110, 80–89. doi: 10.1016/j.ibmb.2019.04.001
Sagri, E., Reczko, M., Gregoriou, M. E., Tsoumani, K. T., Zygouridis, N. E., Salpea, K. D., et al. (2014). Olive fly transcriptomics analysis implicates energy metabolism genes in spinosad resistance. BMC Genomics 15:714. doi: 10.1186/1471-2164-15-714
Sarles, L., Verhaeghe, A., Francis, F., and Verheggen, F. J. (2015). Semiochemicals of Rhagoletis fruit flies: potential for integrated pest management. Crop Prot. 78, 114–118. doi: 10.1016/j.cropro.2015.09.001
Schetelig, M. F., Milano, A., Saccone, G., and Handler, A. M. (2012). Male only progeny in Anastrepha suspensa by RNAi-induced sex reversion of chromosomal females. Insect Biochem. Mol. Biol. 42, 51–57. doi: 10.1016/j.ibmb.2011.10.007
Shen, G., Dou, W., Huang, Y., Jiang, X., Smagghe, G., and Wang, J. J. (2013). In silico cloning and annotation of genes involved in the digestion, detoxification and RNA interference mechanism in the midgut of Bactrocera dorsalis [Hendel (Diptera: Tephritidae)]. Insect Mol. Biol. 22, 354–365. doi: 10.1111/imb.12026
Shi, Y., Jiang, H., Gui, S., Liu, X., Pei, Y., and Xu, L. (2017). Ecdysis triggering hormone signaling (ETH/ETHR-A) is required for the larva-larva ecdysis in Bactrocera dorsalis (Diptera: Tephritidae). Front. Physiol. 8:587. doi: 10.3389/fphys.2017.00587
Singh, I. K., Singh, S., Mogilicherla, K., Shukla, J. N., and Palli, S. R. (2017). Comparative analysis of double-stranded RNA degradation and processing in insects. Sci. Rep. 7:17059. doi: 10.1038/s41598-017-17134-2
Suganya, R., Chen, S., and Lu, K. (2010). Target of rapamycin in the Oriental fruit fly Bactrocera dorsalis (Hendel): its cloning and effect on yolk protein expression. Arch. Insect. Biochem. Physiol. 75, 45–56. doi: 10.1002/arch.20383
Suganya, R., Chen, S., and Lu, K. (2011). cDNA cloning and characterization of S6 Kinase and its effect on yolk protein gene expression in the oriental fruit fly Bactrocera dorsalis (Hendel). Arch. Insect Biochem. Physiol. 78, 177–189. doi: 10.1002/arch.20446
Swevers, L., Huvenne, H., Menschaert, G., Kontogiannatos, D., Kourti, A., Pauchet, Y., et al. (2013). Colorado potato beetle (Coleoptera) gut transcriptome analysis: expression of RNA interference-related genes. Insect Mol. Biol. 22, 668–684. doi: 10.1111/imb.12054
Taning, C. N. T., Christiaens, O., Berkvens, N., Casteels, H., Maes, M., and Smagghe, G. (2016). Oral RNAi to control Drosophila suzukii: laboratory testing against larval and adult stages. J. Pest Sci. 89, 803–814. doi: 10.1007/s10340-016-0736-9
Ulvila, J., Parikka, M., Kleino, A., Sormunen, R., Ezekowitz, R. A., Kocks, C., et al. (2006). Double-stranded RNA is internalized by scavenger receptor-mediated endocytosis in Drosophila S2 cells. J. Biol. Chem. 281, 14370–14375. doi: 10.1074/jbc.M513868200
Vagin, V. V., Sigova, A., Li, C., Seitz, H., Gvozdev, V., and Zamore, P. D. (2006). A distinct small RNA pathway silences selfish genetic elements in the germline. Science 313, 320–324. doi: 10.1126/science.1129333
Vélez, A. M., Khajuria, C., Wang, H., Narva, K. E., and Siegfried, B. D. (2016). Knockdown of RNA interference pathway genes in western corn rootworms (Diabrotica virgifera virgifera Le Conte) demonstrates a possible mechanism of resistance to lethal dsRNA. PLoS One 11:e0157520. doi: 10.1371/journal.pone.0157520
Vitavska, O., Wieczorek, H., and Merzendorfer, H. (2003). A novel role for subunit C in mediating binding of the H+-V-ATPase to the actin cytoskeleton. J. Biol. Chem. 278, 18499–18505. doi: 10.1074/jbc.M212844200
Wang, K., Peng, Y., Pu, J., Fu, W., Wang, J., and Han, Z. (2016). Variation in RNAi efficacy among insect species is attributable to dsRNA degradation in vivo. Insect Biochem. Mol. Biol. 77, 1–9. doi: 10.1016/j.ibmb.2016.07.007
Whyard, S., Singh, A. D., and Wong, S. (2009). Ingested double-stranded RNAs can act as species-specific insecticides. Insect Biochem. Mol. Biol. 39, 824–832. doi: 10.1016/j.ibmb.2009.09.007
Winston, W. M., Molodowitch, C., and Hunter, C. P. (2002). Systemic RNAi in C. elegans requires the putative transmembrane protein SID-1. Science 295, 2456–2459. doi: 10.1126/science.1068836
Xie, Y. F., Niu, J. Z., Jiang, X. Z., Yang, W. J., Shen, G. M., Wei, D., et al. (2017). Influence of various stressors on the expression of core genes of the small interfering RNA pathway in the oriental fruit fly, Bactrocera dorsalis. Insect Sci. 24, 418–430. doi: 10.1111/1744-7917.12311
Xiong, K., Wang, J., Li, J., Deng, Y., Pu, P., Fan, H., et al. (2016). RNA interference of a trehalose-6-phosphate synthase gene reveals its roles during larval-pupal metamorphosis in Bactrocera minax (Diptera: Tephritidae). J. Insect Physiol. 92, 84–92. doi: 10.1016/j.jinsphys.2016.07.003
Yapici, N., Kim, Y. J., Ribeiro, C., and Dickson, B. J. (2008). A receptor that mediates the post-mating switch in Drosophila reproductive behaviour. Nature 451, 33–37. doi: 10.1038/nature06483
Yoon, J.-S., Shukla, J. N., Gong, Z. J., Mogilicherla, K., and Palli, S. R. (2016). RNA interference in the Colorado potato beetle, Leptinotarsa decemlineata: identification of key contributors. Insect Biochem. Mol. Biol. 78, 78–88. doi: 10.1016/j.ibmb.2016.09.002
Zheng, W., Liu, Y., Zheng, W., Xiao, Y., and Zhang, H. (2015). Influence of the silencing sex-peptide receptor on Bactrocera dorsalis adults and offspring by feeding with ds- spr. J. Asia Pac. Entomol. 18, 477–481. doi: 10.1016/j.aspen.2015.05.004
Zheng, W., Zhu, C., Peng, T., and Zhang, H. (2012). Odorant receptor co-receptor Orco is upregulated by methyl eugenol in male Bactrocera dorsalis (Diptera: Tephritidae). J. Insect Physiol. 58, 1122–1127. doi: 10.1016/j.jinsphys.2012.05.011
Zotti, M., dos Santos, E. A., Cagliari, D., Christiaens, O., Taning, C. N. T., and Smagghe, G. (2018). RNAi technology in crop protection against arthropod pests, pathogens and nematodes. Pest Manag. Sci. 74, 1239–1250. doi: 10.1002/ps.4813
Keywords: RNA interference, transcriptome, gene silencing, Diptera, Anastrepha fraterculus
Citation: Dias N, Cagliari D, Kremer FS, Rickes LN, Nava DE, Smagghe G and Zotti M (2019) The South American Fruit Fly: An Important Pest Insect With RNAi-Sensitive Larval Stages. Front. Physiol. 10:794. doi: 10.3389/fphys.2019.00794
Received: 27 February 2019; Accepted: 06 June 2019;
Published: 27 June 2019.
Edited by:
Arash Zibaee, University of Guilan, IranReviewed by:
Gabriella Mazzotta, University of Padova, ItalyChristian Wegener, University of Würzburg, Germany
Copyright © 2019 Dias, Cagliari, Kremer, Rickes, Nava, Smagghe and Zotti. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Naymã Dias, bmF5bWEuZGlhc0BnbWFpbC5jb20=; Guy Smagghe, Z3V5LnNtYWdnaGVAdWdlbnQuYmU=; Moisés Zotti, bW9pc2VzLnpvdHRpQHVmcGVsLmVkdS5icg==