
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Physiol. , 14 June 2019
Sec. Exercise Physiology
Volume 10 - 2019 | https://doi.org/10.3389/fphys.2019.00730
This article is part of the Research Topic Cardiovascular Adjustments and Adaptations to Exercise: From the Athlete to the Patient View all 14 articles
 D. A. Cardinale1*
D. A. Cardinale1* F. J. Larsen1
F. J. Larsen1 J. Lännerström1T. Manselin1O. Södergård1
J. Lännerström1T. Manselin1O. Södergård1 S. Mijwel2
S. Mijwel2 P. Lindholm2
P. Lindholm2 B. Ekblom1
B. Ekblom1 R. Boushel3*
R. Boushel3*Background: Hyperoxia (HYPER) increases O2 carrying capacity resulting in a higher O2 delivery to the working muscles during exercise. Several lines of evidence indicate that lactate metabolism, power output, and endurance are improved by HYPER compared to normoxia (NORM). Since HYPER enables a higher exercise power output compared to NORM and considering the O2 delivery limitation at exercise intensities near to maximum, we hypothesized that hyperoxic-supplemented high-intensity interval training (HIIT) would upregulate muscle mitochondrial oxidative capacity and enhance endurance cycling performance compared to training in normoxia.
Methods: 23 trained cyclists, age 35.3 ± 6.4 years, body mass 75.2 ± 9.6 kg, height 179.8 ± 7.9 m, and VO2max 4.5 ± 0.7 L min−1 performed 6 weeks polarized and periodized endurance training on a cycle ergometer consisting of supervised HIIT sessions 3 days/week and additional low-intensity training 2 days/week. Participants were randomly assigned to either HYPER (FIO2 0.30; n = 12) or NORM (FIO2 0.21; n = 11) breathing condition during HIIT. Mitochondrial respiration in permeabilized fibers and isolated mitochondria together with maximal and submaximal VO2, hematological parameters, and self-paced endurance cycling performance were tested pre- and posttraining intervention.
Results: Hyperoxic training led to a small, non-significant change in performance compared to normoxic training (HYPER 6.0 ± 3.7%, NORM 2.4 ± 5.0%; p = 0.073, ES = 0.32). This small, beneficial effect on the self-paced endurance cycling performance was not explained by the change in VO2max (HYPER 1.1 ± 3.8%, NORM 0.0 ± 3.7%; p = 0.55, ES = 0.08), blood volume and hemoglobin mass, mitochondrial oxidative phosphorylation capacity (permeabilized fibers: HYPER 27.3 ± 46.0%, NORM 16.5 ± 49.1%; p = 0.37, ES = 3.24 and in isolated mitochondria: HYPER 26.1 ± 80.1%, NORM 15.9 ± 73.3%; p = 0.66, ES = 0.51), or markers of mitochondrial content which were similar between groups post intervention.
Conclusions: This study showed that 6 weeks hyperoxic-supplemented HIIT led to marginal gain in cycle performance in already trained cyclists without change in VO2max, blood volume, hemoglobin mass, mitochondrial oxidative phosphorylation capacity, or exercise efficiency. The underlying mechanisms for the potentially meaningful performance effects of hyperoxia training remain unexplained and may raise ethical questions for elite sport.
It is still under the debate whether strategies such as hyperoxia supplementation enabling acute improvement of exercise performance leads to superior training adaptations and therefore greater performance enhancement (Ploutz-Snyder et al., 1996; Hamalainen et al., 2000; Morris et al., 2000; Perry et al., 2005, 2007; Kilding et al., 2012; Przyklenk et al., 2017). However, despite the fact that previous studies have employed different study designs, training intervention lengths (3–6 weeks), exercise mode (continuous or high-intensity interval training), fractions of O2 inspired (FIO2 range: 0.26–0.70), training status of the participants (i.e., untrained or trained cyclists), and different physical performance tests (all-out effort or time to exhaustion), an overall “likely positive” effect on performance has been found following hyperoxic-supplemented training compared to normoxic training (Mallette et al., 2017; Cardinale and Ekblom, 2018).
As recently reviewed (Cardinale and Ekblom, 2018), hyperoxia acutely improves lactate metabolism (Ekblom et al., 1975), reduces muscle glycogen utilization (Stellingwerff et al., 2006), and enables a higher exercise work rate compared to exercise in normoxia (Powers et al., 1989; Nielsen et al., 1999) while reducing submaximal exercise efficiency (Manselin et al., 2017). Acutely, hyperoxia increases oxygen (O2) delivery to the working muscles (Ekblom et al., 1975) and completely prevents exercise-induced arterial hypoxemia (EIAH), i.e., “oxyhemoglobin SaO2 below 95%” (Powers et al., 1989), a condition often found in individuals exercising at intensities approaching maximal oxygen uptake (VO2max) (Dempsey and Wagner, 1999; Dempsey et al., 2008). Considering the O2 delivery limitation at near maximal exercise intensities (Saltin and Calbet, 2006) and that the mitochondria possess an excess capacity above the O2 delivery (Boushel et al., 2011), we postulated that hyperoxic-supplemented exercise training allowing a higher training load leads to a greater training stimulus at the muscle level and therefore greater performance enhancement compared to the same training regimen breathing normoxia. This hypothesis is supported by the finding that exercise training with recombinant human erythropoietin treatment enhanced skeletal muscle mitochondrial capacity compared to controls (Plenge et al., 2012).
Only a few studies have assessed markers of skeletal muscle metabolic adaptions following hyperoxic-supplemented endurance training (Ploutz-Snyder et al., 1996; Perry et al., 2007; Przyklenk et al., 2017). The greater enzyme activity involved in β-oxidation following hyperoxic training indicated by the higher β-hydroxyacyl-coenzyme A dehydrogenase shown by Ploutz-Snyder et al. (1996), was not later found by Perry et al. (2007). In both studies, glycolytic enzymes activities (phosphofructokinase, creatine kinase, and glyceraldehyde phosphate dehydrogenase) and components of oxidative phosphorylation (cytochrome c-oxidase and citrate synthase) were not enhanced with hyperoxic training. Furthermore, previous studies have been conducted on untrained or recreationally trained individuals, and the mechanism of action of hyperoxic-supplemented endurance training may differ in trained individuals. Only two studies have been conducted on trained individuals (Hamalainen et al., 2000; Kilding et al., 2012) with opposite results on performance, and without detail on potential physiological mechanisms. For these reasons, we designed a 6-week randomized controlled training study with parallel groups where both participants and training supervisors were blinded to the type of gas inhaled with the aim to assess the effect of hyperoxic-supplemented high-intensity interval training on physiological and performance outcomes in trained cyclists.
We hypothesized that hyperoxic-supplemented high-intensity interval training in trained cyclists would enhance the training stimulus on skeletal muscle and thereby improve cycle performance to a greater extent than a normoxic breathing condition due to an upregulated skeletal muscle mitochondrial oxidative phosphorylation capacity independent of cardiorespiratory or hematological adaptations.
This study used a 6-week double-blind randomized controlled training study design with parallel groups stratified for participants’ baseline VO2max. Participants were randomly assigned to either an experimental group that trained breathing hyperoxia (FIO2 0.30; HYPER) or a sham-hyperoxia control group that trained in normoxia (FIO2 0.21; NORM). Within a week prior to and posttraining intervention, participants reported to the laboratory three times for baseline and post-tests, respectively. The training intervention consisted of 15 supervised high-intensity interval training (HIIT) bouts breathing either HYPER or NORM and 10 non-supervised low-intensity exercise sessions distributed within a 6-week period (explained in detail in “the training intervention” section). Inclusion criteria in the final analysis were an attendance of at least 85% of the HIIT and low-intensity training sessions. Participants were informed of the possible risks and discomfort involved before giving their written consent to participate. The study was undertaken according to the Declaration of Helsinki and was approved by the Swedish Regional Ethics Committee (2014/1764-31/2 and 2017/630-32).
A group of 32 trained cyclists (24 men and 7 women), age 34.8 ± 7.3 years [mean ± standard deviation (SD)], body mass 72.9 ± 10.8 kg, height 177.7 ± 9.6 m, and VO2max 4.4 ± 0.8 L min−1 participated in this study. Prior to inclusion, a larger group of cyclists completed a health screening survey and exercise tests for assessment of their cardiorespiratory fitness. Only the healthy subjects who regularly conducted endurance and ultra-endurance races and had been competing at a national or amateur level in the last 5–10 years prior to this study were selected for this study. The aim of including experienced cyclists is that this group of athletes shows a small magnitude of change in performance, cardiorespiratory fitness as well as metabolic adaptation even when intensifying their training. On average, each subject was accustomed to about 10–15 h exercise training per week. High-intensity interval training and resistance training was consistently implemented in their normal training regimen on average once per week; however 80–90% of the whole training time was spent at low and average exercise intensity.
The training intervention consisted of a 6-week polarized and periodized endurance training on a cycle ergometer. The term polarized refers to the combination of both HIIT and low-intensity exercise sessions within the same mesocycle (i.e., 6 weeks) (Stöggl and Sperlich, 2014), whereas the term periodized refers to the variation of training load and intensity within the mesocycle. The 6-week intervention protocol used a periodization model with a relationship of 2:1 between hard weeks and easy weeks. This approach was taken to reduce the risk of overtraining syndrome and included a tapering period of a week before the post-tests assessments. Participants were scheduled to perform 15 supervised HIIT sessions breathing either HYPER or NORM and 10 low-intensity exercise sessions.
The HIIT sessions, scheduled on Monday, Wednesday and Friday each week, consisted of either three times 8-min intervals or four times 4-min intervals performed at maximal sustainable effort [equivalent to a rating perceived exertion, RPE, of 18–20 (Borg, 1970)], with 3-min active relief in between the exercise bouts on a cycle ergometer. The longer intervals were performed three times per week during weeks 1, 2, 4, and 5 of the training intervention, whereas the 4-by-4-min intervals were performed twice in week 3 and once in week 6. To progressively increase the participants’ training load during the training intervention, a fourth interval of 4 min in length in week 4 and a fourth interval of 8 min in length in week 5 was added, in addition to the three times 8-min intervals as described above.
Each HIIT session was preceded by a standard 20-min warm up protocol on a cycle ergometer which included two bouts of 1 min at moderate/high-intensity exercise. This type of warm up has been shown to be superior to continuous warm up either at an intensity below or above threshold (McGowan et al., 2015). Overall, the 20-min warm up was performed as follows: 10 min cycling at an exercise intensity equivalent to 10–12 RPE, 1 min at 14–16 RPE, 3 min at 11–14 RPE, 1 min at 15–17 RPE, and 5 min at 10–12 RPE. During the last 30 s of the warm-up protocol, the participants wore a full-face mask with headgear used for gas administration (refer to section “Gas Administration” for the complete setup description). The participants removed the mask during the relief periods. At the completion of the HIIT session, a 10-min cool-down was performed at an exercise intensity equivalent to 10–12 RPE.
During the HIIT sessions, the only data shown to the participants were the cycling cadence and the elapsed exercise time, whereas the participants were blinded to breathing condition, power output, and heart rate. Since the HIIT sessions were self-paced by the participants who were blinded to power output, one or two training supervisors guided and encouraged the participants to assure that each interval of each session was performed at the highest sustainable intensity possible by the participants. For motivational and time-efficiency purposes, participants completed HIIT sessions in the company of 2–4 other participants. The training supervisors were also blinded to the breathing condition that the participants were assigned to during the whole duration of the training intervention.
Participants were encouraged to perform two low-intensity exercise training sessions consisting of about 2 h (LOW1) and 4 h (LOW2) long continuous exercise at ~75% of the individual maximal heart rate (equivalent to an RPE of 14–15). The participants conducted the LOW1 cycling either outdoors or on a stationary bike indoors on Tuesday or Thursday of weeks 1, 2, 4, 5, and 6 of the training intervention. A LOW2 session was scheduled each weekend during the 6-week training intervention.
Participants were requested to maintain supplementary training involving flexibility exercise and exercise specifically recruiting upper body muscles. No other strenuous exercise training was allowed during the 6-week intervention except for the one above described.
The gas was delivered to the participants through a face mask which was connected, by tubing, to a dosage unit (Oxelerate, Tumba, Sweden) which in turn was connected to a gas tank filled with either pure medical oxygen or medical air gas (i.e., 21% O2). For subjects assigned to the hyperoxia group, the dosage unit intermittently delivered a gas bolus at the beginning of each participant’s inhalation that was mixed with room air inside the mask cavity resulting in a final FIO2 of ~0.30, as established by previous work (Lindholm et al., 2017). The participants and the training supervisors were blinded to the type of gas inhaled.
Participants reported to the laboratory on three separate occasions with 2–3 days in between within the week prior to the start of the exercise intervention and a week after the last training session. Participants abstained from strenuous physical activity 24–48 h prior to each occasion.
The first occasion consisted of skeletal muscle biopsy collection and total hemoglobin mass assessment which was scheduled in the morning between 07:00 and 10:00 h to limit the circadian influence. At the second occasion, participants performed a submaximal and maximal incremental test on cycle ergometer in the afternoon between 15:00 and 19:00 h. The third occasion consisted of a self-paced cycling performance test scheduled between 09:00 and 17:00 h.
A skeletal muscle sample was obtained from the middle portion of the vastus lateralis muscle at a depth of 2–3 cm, about one-third of the distance from the upper margin of the patella to the anterior superior iliac spine with the participants resting in a semi recumbent position lying on a bench. After local anesthesia (2–4 ml carbocaine 20 mg ml−1; AstraZeneca, Södertälje, Sweden), an incision (0.5–1 cm) was made through the skin and fascia and a muscle sample (50–100 mg) was obtained with the Weil-Blakesley chonchotome technique. A portion of the sample was snap frozen in liquid nitrogen, while second and third portions were rapidly placed in ice-cold mitochondrial isolation medium and relaxing medium (see section below), followed by mitochondria isolation and muscle fiber permeabilization, respectively, as later described.
A subgroup of 12 participants (n = 6 per group) was tested for total hemoglobin mass (Hbmass) assessment performed as described elsewhere (Burge and Skinner, 1995) with some minor modification of the rebreathing technique. Briefly, with the participants still lying on the bench in a semi recumbent position following the biopsy collection, 15 ml of blood was sampled from an antecubital vein via a 20-gauge venflon and analyzed immediately for Hb concentration (Hb) using HemoCue® Hb 201+ System (HemoCue AB, Ängelholm, Sweden); and hematocrit in quadruplets with micro-method (3 min at 13,500 rpm). About 1.5 ml of the blood sample was then quickly transferred to a 2-ml Eppendorf tube and stored at −80°C until percent carboxyhemoglobin (%HbCO) analysis using an hemoximeter (ABL800, Radiometer, Copenhagen, Denmark). After baseline collection, the participants breathed from a Douglas bag previously filled with pure oxygen for 4 min to flush nitrogen from the airways. During this time, the operator flushed the re-breathing circuit with the pure oxygen which was then closed. After 4 min, the operator switched the participant to the rebreathing circuit and a precisely measured bolus of 1.2 ml kg−1 body mass of 99.997% chemically pure CO (CO N47, Air Liquide, Paris, France) was injected into the circuit. The participants then breathed the gas mixture for 10 min. Thereafter, an additional venous blood sample was collected from an antecubital vein for assessment of the change in (%HbCO) accounting for the CO remaining in the re-breathing circuit which was determined (Monoxor III, Bacharach Inc., New Kensington, USA). Hbmass was calculated from the change in %HbCO and total red blood cell volume (RCV), blood volume (BV), and plasma volume (PV) were derived (Burge and Skinner, 1995).
To determine the power output at lactate inflection point using a modified Dmax method, a sub-group of 12 participants (n = 6 per group) cycled at 4–6 intervals (i.e., 30, 150, 185, 220, 255, 290, and 325 W) each of 4 min in length on a cycle ergometer (Monark LC6, Monark Exercise AB, Vansbro, Sweden). Participants cycled with a constant, freely chosen cadence until blood lactate concentration (Biosen C-Line Clinic; EKF-diagnostic GmbH, Barleben, Germany) measured at the end of each interval from fingertips, reached a concentration higher than 4 mMol L−1. Participants cycled wearing a Hans Rudolf mask (Hans Rudolph Inc., Kansas, USA) which covered mouth and nose for assessment of pulmonary oxygen consumption (Jaeger Oxycon Pro; CareFusion GmbH, Hoechberg, Germany). The metabolic cart was calibrated prior to each test according to the manufacturer’s instructions, with high-grade calibration gases (Air Liquide, Paris, France). Respiratory variables were measured and averaged every 10 s. The averaged VO2 recorded during the last minute of each interval was taken as the representative oxygen consumption for that specific power output. After the last interval, participants were allowed 10 min cycling at an intensity equal to 10–12 RPE to recover and get ready for subsequent graded incremental exercise test.
The blood lactate concentrations representative for each exercise intensity were used to calculate the power output at lactate inflection point using a modified Dmax method (Cheng et al., 1992), i.e., defined as the derivate to the exponential curve created from exponential lactate increase, including maximal lactate concentration obtained from the successive maximal increment test. The increase in lactate relative to power output was defined as the increase in blood lactate from the point where the exponential curve crossed the lactate baseline.
The mean oxygen consumption and the respiratory exchange ratio between minute 3 and 4 at each cycled stage was used to calculate energy expenditure with the equation developed by Brouwer (1957). Cycling efficiency when cycling at 150 W was expressed as gross efficiency (GE) and work efficiency (WE) as described elsewhere (Mogensen et al., 2006).
All participants performed a graded incremental exercise test until volitional exhaustion on a cycle ergometer to determine VO2max and time to exhaustion. Participants pedaled at a fixed cadence and the load was increased by 25 and 20 W min−1 for men and women, respectively. Participants cycled wearing a Hans Rudolf mask (Hans Rudolph Inc., Kansas, USA) which covered mouth and nose for assessment of pulmonary oxygen consumption (Jaeger Oxycon Pro; CareFusion GmbH, Hoechberg, Germany). VO2max leveling-off criteria were applied (i.e., a VO2 plateau, followed by exercise cessation or decrease of VO2 at higher work rates, with an RER >1.10). The power output that the participants pedaled at the time of the volitional exhaustion was taken as the maximal power output (Winc.). O2 consumption was measured with a metabolic cart (OxyconPro, Jaeger GmbH, Germany), calibrated prior to each test according to the manufacturer’s instructions, with high-grade calibration gases (Air Liquide, Paris, France). Respiratory variables were measured and averaged every 10 s. The highest 60 s averaged VO2 recorded was taken as the VO2max.
The self-paced endurance cycling performance test consisted of pedaling for 20 min at the maximal sustainable effort with the intent to obtain the highest mean power output on a cycle ergometer (Monark LC2, Monark Exercise AB, Vansbro, Sweden) equipped with a power meter (SRM power meter science road; SRM International, Jülich, Germany) for power output assessment. The test was preceded by a standard 20-min warmup as described in the “Training intervention” section. Briefly, the participants cycled for 10 min at an exercise intensity equivalent to 10–12 RPE, 1 min at 14–16 RPE, 3 min at 11–14 RPE, 1 min at 15–17 RPE, and 5 min at 10–12 RPE. During the last 5 min of the warm up, the participants were equipped with a Hans Rudolf mask (Hans Rudolph Inc., Kansas, USA) which covered mouth and nose for assessment of pulmonary oxygen consumption (Jaeger Oxycon Pro; CareFusion GmbH, Hoechberg, Germany) and a pulse oximetry sensor (Rad-97 Masimo Corporation; Neuchatel, Switzerland) was positioned on the participant’s forehead for assessment of the peripheral capillary oxygen saturation (SpO2).
During the test, the only data shown to participants were the cycling cadence and the elapsed exercise time, whereas the participants were blinded to power output and heart rate. The test leader verbally encouraged the participants during the whole test length. For motivational purposes, music chosen by the participant was played during test. The participants completed the test with a 10 min cool-down performed at an exercise intensity equivalent to 10–12 RPE. Participants were all familiar to this test and performed one or two familiarization tests prior the baseline test, which showed a typical error of measurement of about 2%.
Muscle fiber bundle permeabilization was performed as previously described (Pesta and Gnaiger, 2012; Cardinale et al., 2018a). Briefly, a portion of the muscle biopsy (~5 mg wet weight) was immediately transferred into ice-cold relaxing medium (BIOPS) containing 10 mM/L Ca2+/EGTA buffer, 20 mM/L imidazole, 50 mM/L K+-4-morpholinoethanesulfonic acid (Mes), 0.5 mMol/L dithiothreitol, 6.56 mM/L MgCl2, 5.77 mM/L ATP, and 15 mMol/L phosphocreatine at pH 7.1. A portion of the 5 mg wet weight sample (~1–3 mg) was transferred into BIOPS in a small petri dish on an ice-cold metal plate where the fiber bundles were mechanically separated using forceps and needles. Thereafter, approximately 10–15 fibers were slowly agitated for 30 min on a platform shaker in BIOPS containing saponin (5 mg/ml saponin) solution at 4°C. Fibers were then washed for 10 min at 4°C in ice-cold mitochondrial respiration medium (MiR06; 0.5 mM EGTA, 3 mM MgCl2, 60 mM K-lactobionate, 20 mM taurine, 10 mM KH2PO4, 20 mM HEPES, 110 mM sucrose, and 1 g/L BSA essentially fatty acid free, adjusted to pH 7.1, 2.8 U/mg solid catalase lypophilized powder). The fibers were weighed on a microbalance after having been blotted on filter paper and transferred into the respirometry chamber.
A portion of the muscle biopsy (40–80 mg wet weight) designated for mitochondrial isolation was first weighed and then cut in ice-cold isolation medium (sucrose 100 mM, KCl 100 mM, Tris-HCl 50 mM, KH2PO4 1 mM, EGTA 100 μM, BSA 0.1%; final pH was set to 7.4). The homogenate was washed in 1 ml isolation medium and the supernatant was removed. One ml of isolation medium containing 0.2 mg ml−1 bacterial protease was added to the homogenate. The homogenate was gently agitated every 30 s for ~2-min time and then transferred in a pre-cooled glass jacket connected to an ice-cold bath pump and further homogenized with a hand held electrically driven drill (80 rpm). The final homogenate was then transferred to a falcon tube containing 3 ml isolation medium and then subsequently centrifuged at 700 g at 4°C for 10 min. After removing the pellet, the suspension was again centrifuged at 10,000 g at 4°C. The resultant mitochondrial pellet was re-suspended in the same medium. The Eppendorf was then centrifuged at 7,000 g for 5 min and the pellet was dissolved in 0.6 μl preservation medium (EGTA 0.5 mM, MgCl2⋅6H2O 3 mM, K-lactobionate 60 mM, Taurine 20 mM, KH2PO4 10 mM, HEPES 20 mM, sucrose 110 mM, BSA 1 g L−1 histidine 20 mM, vitamin E succinate 20 μM, glutathione 3 mM, leupeptine 1 μM, glutamate 2 mM, malate 2 mM, and Mg-ATP 2 mM) per mg wet weight.
Mitochondrial respiration was performed in a two-channel high-resolution respirometer (Oroboros Oxygraph, Paar, Graz, Austria). The glass chamber volume (2 ml capacity) was sealed with rubber-ringed stoppers to minimize the O2 back diffusion into the chamber (Steinlechner-Maran et al., 1996). Polyvinylidene difluoride magnetic stirrers set to 750 rpm were used to stir the sample in to the medium. Data were collected at 1-s intervals and averaged over 40 s. All experiments were run in duplicate and the respiration data for each of the two chambers were then averaged. The medium in the respiration chamber was MiR05 containing EGTA 0.5 mM, MgCl2⋅6H2O 3 mM, K-lactobionate 60 mM, taurine 20 mM, KH2PO4 10 mM, HEPES 20 mM, sucrose 110 mM, and BSA 1 g L−1. All experiments were performed at 37°C. Time constants for complete mixing in the chamber were calculated by briefly stopping and starting the stirrers. O2 consumption and zero-drift of the O2 electrode were calculated using DatLab 5.2 software (Oroboros, Paar, Graz, Austria). At least five different O2 tensions (400–0 nMol/ml) were used during the background calibration to calculate and account for the diffusion of O2 into the chamber.
Mitochondrial respiration was measured by adding the following substrates into the chambers (final concentrations): octanoylcarnitine (0.2 mM), malate (2 mM) for assessment of leak respiration (EFTL), ADP (2.5 mM) to support electron entry from fatty acid β-oxidation through electron-transferring flavoprotein and complex I (EFTP), followed by pyruvate (5 mM) and glutamate (10 mM) to stimulate complex I (CIP), and succinate (10 mM) to stimulate complex I and II linked respiration (CI + IIP). Following inhibition of complex III with antimycin A (mM), N,N,N′,N′-tetramethyl-p-phenylenediamine dihydrochloride (0.5 mM) and ascorbate (2 mM) were added followed by sodium azide (100 mM) for cytochrome C oxidase activity assessment. O2 flux from the permeabilized fiber preparation was normalized per initial fiber wet weight.
The same titration protocol described above was used to measure O2 flux in isolated mitochondria with the exception that ADP was left to be completely phosphorylated to ATP following β-oxidation respiration allowing respiration proceed to state 4 (S4). Furthermore, carbonyl cyanide m-chloro phenyl hydrazine (0.05 μM steps) was used to measure maximal uncoupled oxidative phosphorylation (Unc). O2 flux from isolated mitochondria was normalized by protein concentrations determined in aliquots of supernatant diluted 1:10 in distilled water using the Pierce 660 nm protein assay (Thermo Scientific).
A portion of the freeze-dried muscle samples was first cleansed of visible blood, fat and connective tissue and subsequently homogenized in ice-cold buffer (100 μl/mg dry weight) consisting of 50 mM KH2PO4, 1 mM EDTA and 0.05% Triton X-100 using a Bullet Blender (NextAdvance, Averill Park, NY) with 0.5 mm ZrO beads. The Eppendorf tubes containing the homogenates were rotated for 30 min at 4°C before being centrifuged at 1,400 g for 1 min at 4°C. CS activity was measured on a 96-well plate in a reagent solution (in mM): 50 Tris-HCl, 0.2 DTNB, and 30 acetyl-CoA. The reaction was initiated by adding oxaloacetate (10 mM) and the change in absorbance at 412 nm was measured spectrophotometrically at 25°C.
A portion of the snap frozen biopsy sample was (1) freeze-dried, (2) cleansed of visible blood, fat, and connective tissue and subsequently, and (3) homogenized in ice-cold buffer (100 μl/mg dry weight) consisting of 2 mM HEPES (pH 7.4), 1 mM EDTA, 5 mM EGTA, 10 mM MgCl2, 1% Triton X-100, 2 mM dithiothreitol, and 1.5% phosphatase and protease inhibitor cocktail (Halt™, Thermo Scientific, Rockford, MD) using a Bullet Blender (NextAdvance, Averill Park, NY) with 0.5 mm ZrO beads. The Eppendorf tubes containing the homogenates were 360° rotated for 30 min at 4°C before being centrifuged at 10,000 g for 10 min at 4°C. The obtained supernatant was stored at −80°C. Protein concentrations of the homogenates were determined using the Pierce 660 nm protein assay (Thermo Scientific). Muscle homogenates were diluted with 4× Laemmli sample buffer (Bio-Rad, Richmond, CA) and homogenizing buffer to obtain a final protein concentration that was similar between all samples. Subsequently, all samples were heated at 95°C for 5 min to denature proteins, and then stored at −20°C until further analysis.
Samples were separated by SDS polyacrylamide gel electrophoresis (PAGE) on 26-well Criterion TGX gradient gels (4–20% acrylamide; Bio-Rad). Samples from all three groups were loaded on the same gel. The blots were quantified using Quantity One software version 4.6.3 (BIORAD). To control for appropriate loading and transfer, target proteins were expressed relative to total protein stained at ~95 kDa obtained by staining the membranes with MemCode Reversible Protein Stain Kit (Thermo Scientific) (Antharavally et al., 2004).
The monoclonal primary cytochrome c oxidase antibody (#4850; 1:1,000; Cell Signaling Technology, Danvers, USA) conjugated with a secondary anti-rabbit antibody (#7074; 1:10,000; Cell Signaling Technology, Danvers, USA) was used for the detection of target total protein.
Normal distribution of the data was checked by assessing skewness. Baseline characteristics of each group were summarized using descriptive statistics. Exact χ2 tests were used to evaluate if differences existed between groups for categorical variables at baseline. For between-groups analyses, one-way analysis of variance (one-way ANOVA) was conducted using the variable changes pre to post intervention. A two-tailed p < 0.05 was considered significant. Analysis of covariance (ANCOVA) with baseline-test results as a covariate and the post-test as the dependent variable was also performed on the same dataset to ensure that baseline measurements did not affect the statistical results. The use of one-way ANOVA or ANCOVA did not influence the interpretation of the study results. Effect sizes and associated confidence intervals were interpreted according to Cohen’s guidelines (Cohen, 1988), effect sizes with scores of 0.2–0.5, 0.5–0.8, and >0.8 were considered small, medium, and large effects, respectively. Paired sample t-test was used to assess within-group differences pre- to post measurement. Statistical analyses were carried out using SPSS statistical software version 21 (SPSS Inc., Chicago, Illinois, USA).
Of the initial 32 trained cyclists, 23 of those (19 men and 4 women), with age 35.3 ± 6.4 years [mean ± standard deviation (SD)], body mass 75.2 ± 9.6 kg, height 179.8 ± 7.9 m, and VO2max of 4.5 ± 0.7 L·min−1, successfully adhered to the training regimen (NORM attended 95.8 ± 5.4% of the HIIT sessions and 90.1 ± 8.6% of the LOW1/LOW2 sessions; HYPER attended 94.4 ± 5.6% of the HIIT sessions and 95.5 ± 4.7% of the LOW1/LOW2 sessions) and were included for further analysis. The two groups NORM (n = 11) and HYPER (n = 12) did not differ at baseline for any measured variables. Participants’ characteristics are shown in Table 1. During the HIIT sessions, HYPER consistently trained at 3.3 ± 1.2% higher relative intensity than NORM despite a similar rating of perceived exertion (i.e., NORM RPE 8.3 ± 1.0 and HYPER RPE 8.2 ± 0.7). However, the higher training intensity did not lead to a significantly higher training load over the intervention compared to NORM (p = 0.37) (Figure 1).
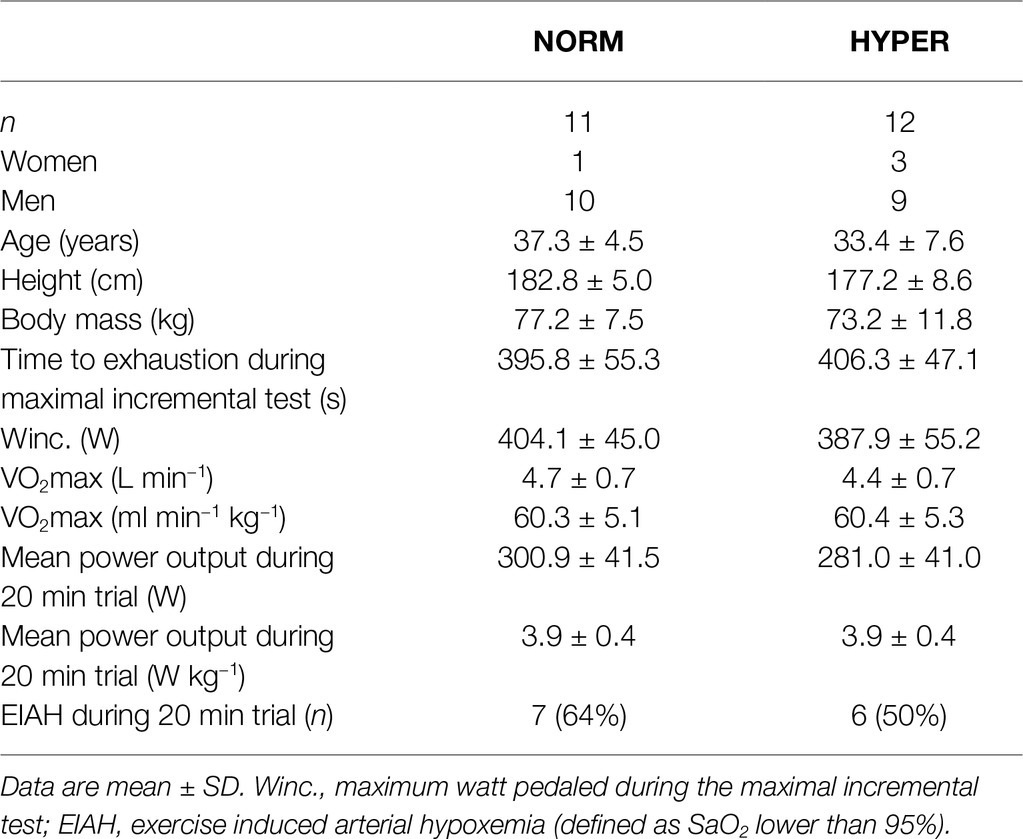
Table 1. Characteristics and baseline measures of the hyperoxia (HYPER) and normoxia (NORM) group.
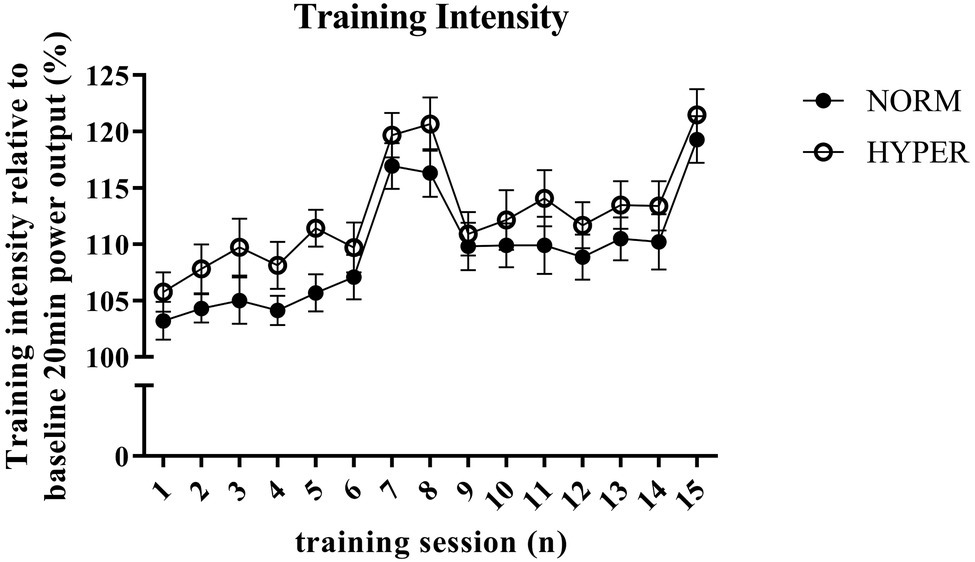
Figure 1. Overview of the training intensity and progression of the normoxic (NORM) and hyperoxic (HYPER) group during the 6-week training intervention. Training intensity of each high-intensity interval session is expressed in percentage of the mean power output obtained during the baseline self-paced 20-min cycling trial. Note that the higher training intensity during training session number 7, 8, and 15 is attributed to the higher exercise intensity obtained when performing 4 × 4 min interval training session instead of the otherwise performed 3 × 8 min intervals. This figure shows (1) the numerically but not significantly higher training intensity performed by HYPER compared to NORM throughout the training intervention which was independent of the interval duration, (2) the increasingly higher training intensity performed by both groups during the intervention period which indicated that cycle performance was enhanced in both groups independent of hyperoxia.
The permeabilized fibers assay revealed that the training intervention had no effect on mass-specific (normalized to tissue wet weight) leak respiration, fatty acid oxidation, maximal oxidative phosphorylation respiratory, and electron transfer system capacity of the skeletal muscle with no difference between groups (Figure 2A). Maximal oxidative phosphorylation numerically increased (22.6 ± 46.1%) but did not reach the level of significance over the intervention (p = 0.20), between (p = 0.37, ES = 3.24) or within groups (NORM 16.5 ± 49.1% p = 0.86; HYPER 27.3 ± 46.0%, p = 0.15) and showed a great individual variability in response to exercise training (Figure 3). Although no differences over the intervention were detected between groups, fatty acid oxidation decreased over the intervention only in NORM (p = 0.03).
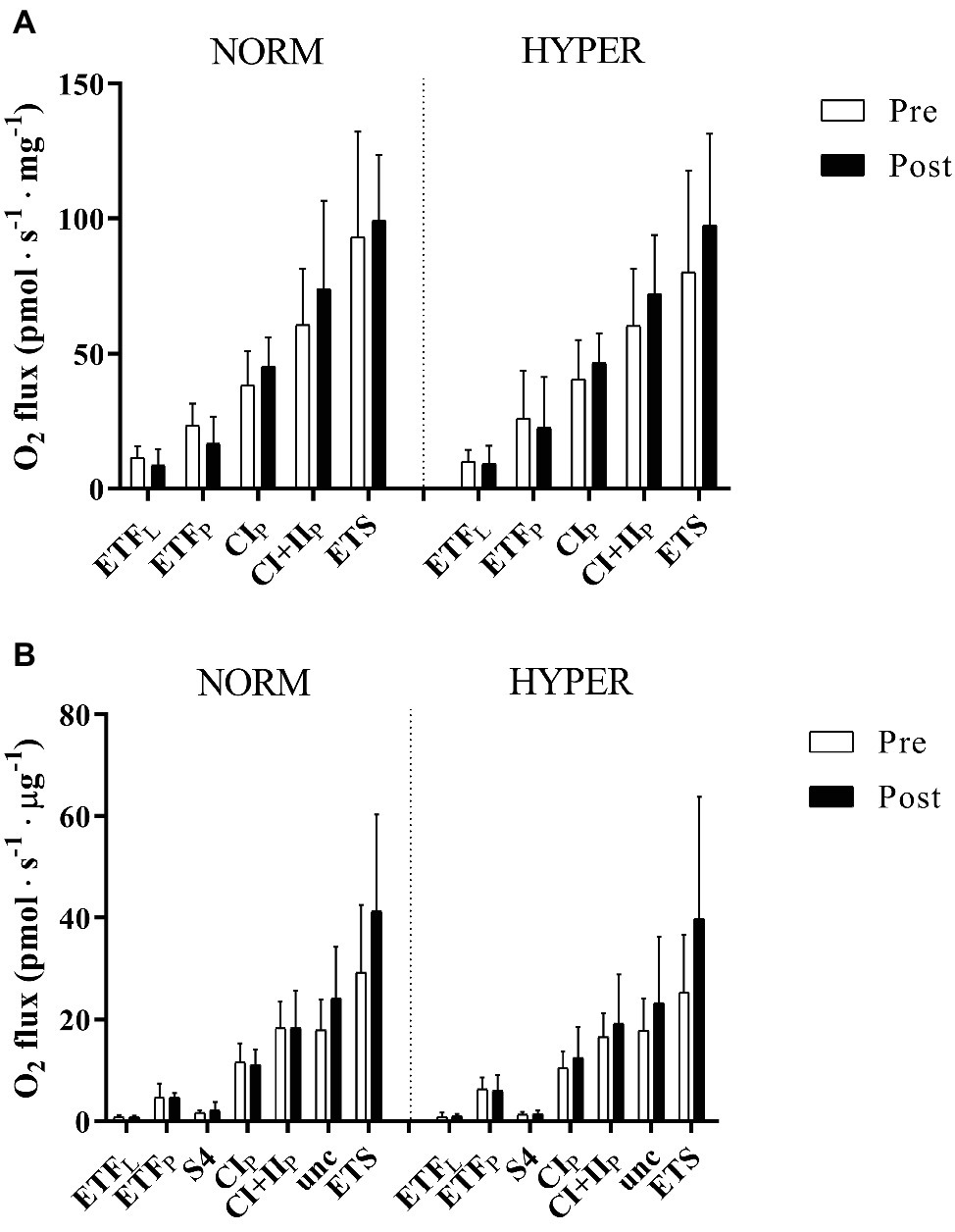
Figure 2. Mitochondrial respiration pre and post 6 weeks polarized and periodized high-intensity interval training either in normoxia (NORM) or hyperoxic-supplemented (HYPER). All values are expressed as means ± SD. In panel (A), O2 flux rates obtained from measurement of mitochondrial respiration using permeabilized fibers technique are expressed in pmol s−1 mg−1 fibers initial wet weight, whereas in panel (B), O2 flux obtained from measurement of isolated mitochondria the O2 flux rates are presented in pmol s−1 μg−1 protein. Electron-transferring flavoprotein complex (ETF), state 4 (S4), complex I (CI), complex I + II-linked substrate state (CI + II), and uncoupling state (Unc) electron transport system capacity (ETS). Leak respiration and oxidative phosphorylation are indicated by subscripts L and P, respectively.
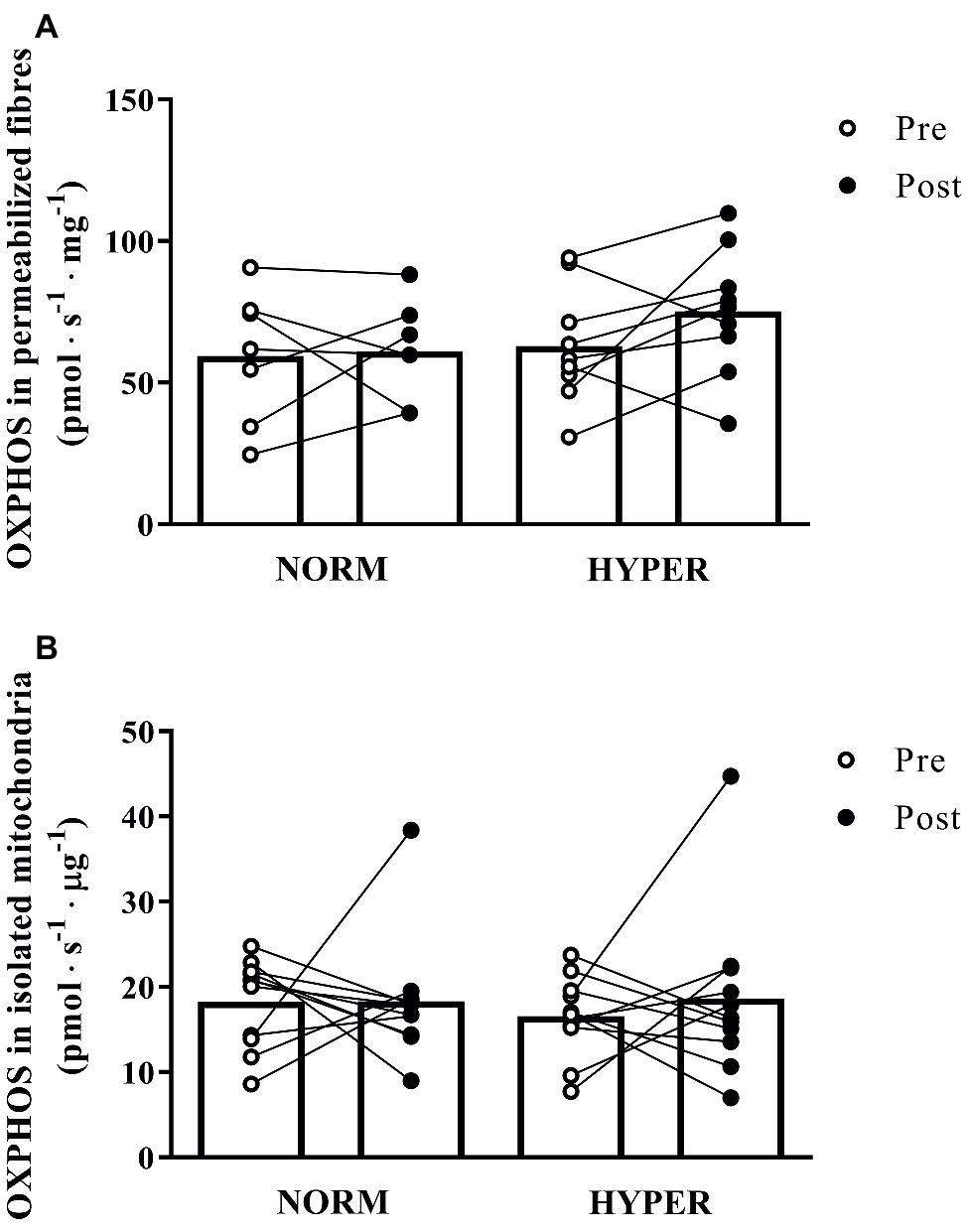
Figure 3. Maximal mitochondrial respiration (OXPHOS) pre and post 6 weeks polarized and periodized high-intensity interval training either in normoxia (NORM) or hyperoxic-supplemented (HYPER). All values are shown as mean and individual response following the intervention. In panel (A), O2 flux rates obtained from measurement of mitochondrial respiration using permeabilized fibers technique are expressed in pmol s−1 mg−1 fibers initial wet weight, whereas in panel (B), O2 flux obtained from measurement of isolated mitochondria are presented in pmol s−1 μg−1 protein.
Similarly, the isolated mitochondria assay showed only a small relative increase (21.0 ± 75.1% p = 0.90) in intrinsic maximal mitochondrial respiration over the intervention (Figure 2B) with no change between (p = 0.66, ES = 0.51) or within groups (NORM 15.9 ± 73.3% p = 0.99; HYPER 26.1 ± 80.1%, p = 0.54).
The unchanged ex vivo mitochondrial oxidative phosphorylation over the intervention paralleled the unchanged biomarkers of mitochondrial content, i.e., citrate synthase (CS) activity (p = 0.42 and between groups p = 0.41). CS activity in NORM went from 238.7 ± 53.2 to 239.3 ± 39.5 nM min−1 mg−1 (p = 0.97) and HYPER from 239.4 ± 37.3 to 221.9 ± 48.6 nM min−1 mg−1 (p = 0.28). Similarly, cytochrome C oxidase protein levels did not change over the intervention (p = 0.93) and between groups (p = 0.95). Cytochrome C oxidase protein levels in NORM went from 11.6 ± 6.4 to 11.6 ± 4.2 a.u. (p = 0.99) and HYPER from 7.7 ± 3.6 to 7.6 ± 3.6 a.u. (p = 0.90).
Hematology variables are presented in Table 2. All parameters were unaltered by the training intervention and no differences were detected between groups.
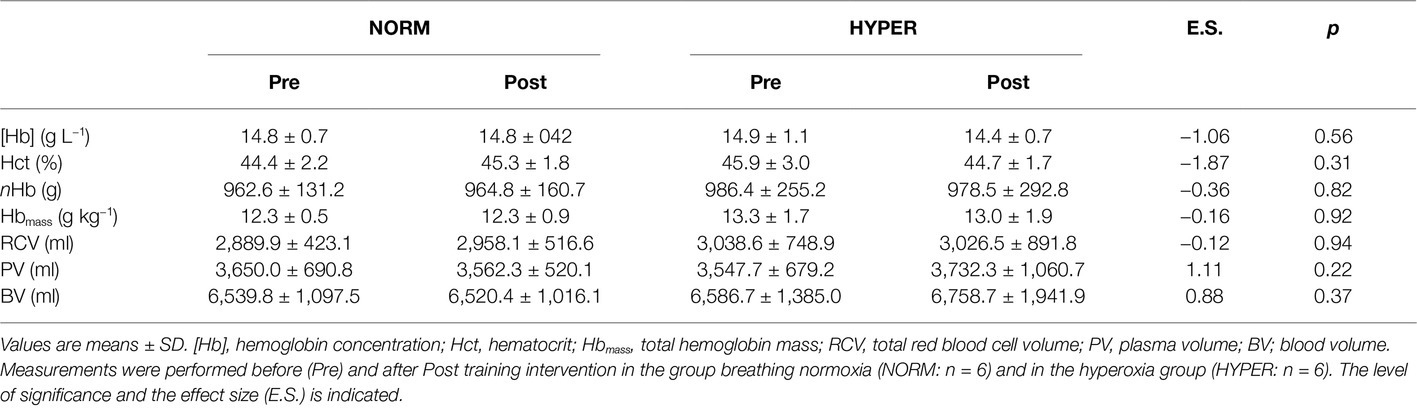
Table 2. Body mass and intravascular volumes before and after training intervention.
Power outputs at lactate inflection point did not change over the intervention (p = 0.73). NORM went from a power output at lactate inflection point of 281.8 ± 42.1 to 282.9 ± 44.9 W (p = 0.78) and HYPER from 288.2 ± 37.2 to 289.3 ± 29.2 W (p = 0.84). No change in gross efficiency (GE) and work efficiency (WE) when cycling at 150 W were observed pre to post intervention with no difference between groups (NORM GE from 19.0 ± 0.9 to 19.3 ± 1.1% and HYPER GE 19.7 ± 0.1 to 20.0 ± 1.3%; NORM WE from 24.2 ± 1.0 to 24.9 ± 0.5% and HYPER WE from 25.0 ± 1.3 to 25.7 ± 1.7%).
Overall, VO2max was maintained over the course of the intervention (p = 0.58) with no difference between groups (p = 0.55, ES = 0.08). NORM went from a VO2max of 4.7 ± 0.7 to 4.6 ± 0.7 L min−1 (0.0 ± 3.7%, p = 0.84) and HYPER from 4.4 ± 0.7 to 4.5 ± 0.8 L min−1 (1.1 ± 3.8%, p = 0.33). Time to exhaustion significantly improved pre to post intervention from 401.3 ± 42.1 to 426.8 ± 49.7 s (p = 0.001) with no difference between groups (p = 0.31, ES = −0.18).
Overall, the 6-week training intervention enhanced the absolute (3.7 ± 4.6%; p = 0.001) and relative to body mass (4.3 ± 4.7%; p = 0.0002) mean power output during the self-paced 20-min cycling trial (Figure 4). The magnitude of improvement in mean power output during the 20-min cycling trial was larger in HYPER compared to NORM (for mean power output, p = 0.07, ES = 0.22; for mean power output relative to body mass, p = 0.06, ES = 0.32). NORM numerically increased absolute mean power output during the 20-min cycling trial by 1.6 ± 4.3% (p = 0.23), whereas HYPER significantly enhanced power output by 5.6 ± 4.2% (p = 0.0006) (Figure 4). The power output during the 20-min test normalized per body mass numerically increased by 2.4 ± 6.0% in NORM (p = 0.15) and significantly in HYPER by 6.0 ± 3.7% (p = 0.0001).
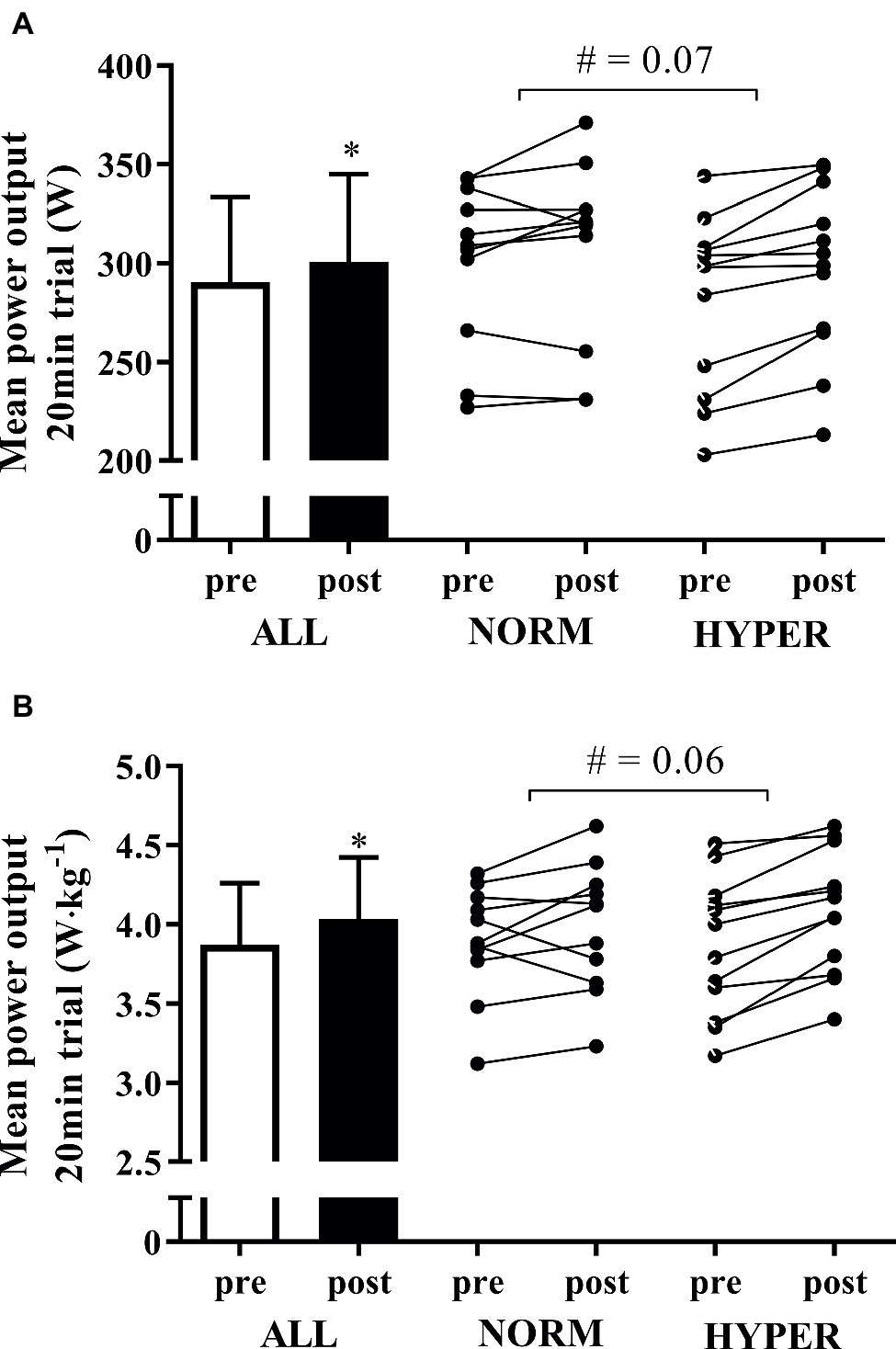
Figure 4. Mean ± SD of the mean power output during the self-paced 20-min cycling trial pre and post intervention expressed in absolute Watts (A) and Watts relative to body mass (B) for all the participants as well as the individual response of the participants in normoxia (NORM) and hyperoxia (HYPER) groups. Note: *p < 0.05, #p between groups.
This study presents novel findings on performance effects and physiological responses to high-intensity interval training with hyperoxia in already trained cyclists. We showed that 6-week high-intensity interval training induced non-significant, but potentially meaningful performance gains without affecting VO2max, hematological parameters, mitochondrial oxidative phosphorylation capacity, and biomarkers of mitochondrial content in already trained cyclists. Our results are in line with the study of Kilding et al. (2012) who, using a similar polarized training (Seiler, 2010) intervention with a FIO2 of 0.60, showed that training with hyperoxia had no significant physiological benefit in trained cyclists of a similar performance level to those tested in our study. This study shows that training with hyperoxia induces no change in hematological or muscle oxidative metabolic capacity as underlying mechanisms for endurance performance.
Hyperoxia acutely enables a higher exercise intensity and therefore larger mechanical work produced over the intervention compared to when breathing normoxia (Perry et al., 2005, 2007; Kilding et al., 2012). Despite a numerically greater exercise intensity in HYPER compared to NORM, contrary to our hypothesis, the higher relative intensity and greater mechanical work led to a small positive effect in performance compared to normoxia but did not induce superior physiological training adaptations. Of note, cyclists could only see cadence data during the HIIT sessions and were blind to breathing assignment, power output and heart rate while pedaling at the maximum effort during the 4- and 8-min intervals. Our findings indicate that a further increase in exercise intensity of an already high-intense exercise regimen does not necessarily lead to additional gains in skeletal muscle training adaptations. However, individual performance change over the intervention (Figure 4) revealed that breathing hyperoxia allowed a positive training response compared to normoxia in some individuals (2 of 11 cyclists in NORM slightly decreased power output during the 20-min cycling trial pre- to postintervention). We speculate that hyperoxic-supplemented training may be advantageous in individuals who show a lower magnitude of performance change compared to the group response. With this consideration, hyperoxic-supplemented endurance high-intensity training may have an effect in erroneously categorized “non-responders” prior to increasing training frequency as previously suggested (Montero and Lundby, 2017).
Improved arterial oxygen saturation with hyperoxia has been reported at exercise intensities near to VO2max thus preventing EIAH (Nielsen, 2003). EIAH is linked to the alveolar-capillary diffusion limitation due to decreased Hb mean-transit time in the lung (Dempsey and Wagner, 1999) caused by mechanical ventilatory constraint during exercise (Dominelli et al., 2013). EIAH has been reported to occur in ~50% of highly trained endurance athletes at sea level (Powers et al., 1988) and is more pronounced in both active and well-trained females (St Croix et al., 1998; Guenette and Sheel, 2007) than in male athletes (Anselme et al., 1994). In line with previous findings, 57% of our participants showed EIAH at intensities near to maximum; 7 of the 11 cyclists in the NORM, and 6 of the 12 cyclists in HYPER. The lower the SaO2 during exercise intensity near to maximum the larger is the effect of acutely breathing hyperoxia on exercise tolerance (Ohya et al., 2016). Therefore, it can be expected that maintaining Hb fully saturated at an exercise intensity near to VO2max in cyclists who otherwise exhibit EIAH in normoxia would lead to improved skeletal muscle training adaptations resulting from the increased O2 delivery. Contrary to what we hypothesized, increasing O2 delivery, partially preventing EIAH to occur, did not improve exercise adaptations when breathing hyperoxia. Furthermore, in HYPER, the change in performance did not correlate with the SaO2 levels during the cycle performance test (Figure 5). By contrast, there was a trend for an opposite relationship whereby cyclists manifesting high SaO2 during the cycle performance test in normoxia demonstrated a larger magnitude of change in performance over the hyperoxic intervention.
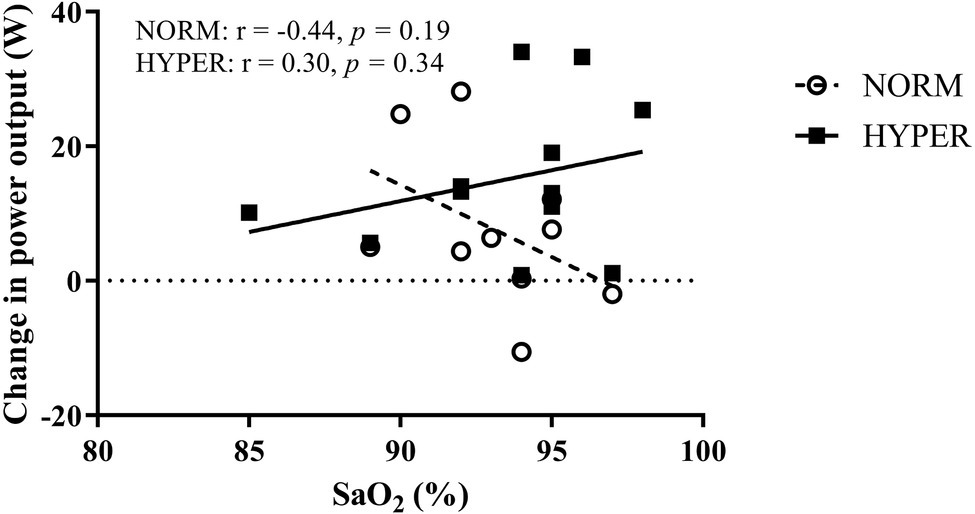
Figure 5. Relationship between the change in power output during the 20-min self-paced cycling test over the training intervention and the mean arterial oxygen saturation (SaO2) measured during the same test in participants in normoxia (NORM) and hyperoxia (HYPER) groups.
Fundamental papers have shown that the convective component of the oxygen cascade limits VO2max (Ekblom et al., 1968; Saltin and Calbet, 2006) and that the maximal mitochondrial oxidative phosphorylation is in excess over the O2 delivery (Boushel et al., 2011) in healthy individuals. We hypothesized that increasing microvasculature PO2 by hyperoxia would increase in vivo mitochondrial relative activation (Cardinale et al., 2018b), such that each mitochondrion would respire at a higher rate in vivo and in turn be exposed to a higher adaptive training stimulus. This hypothesis is supported by the greatly enhanced peripheral adaptations of skeletal muscle following one-legged cycling where higher O2 delivery per active muscle mass occurs compared to cycling (Abbiss et al., 2011). By contrast, we did not find support for this notion. It is likely that mitochondrial respiratory capacity may be still in excess of the O2 delivery despite the increased O2 carrying capacity due to hyperoxia while cycling. Our findings indicate that the greater mitochondrial activation induced when breathing hyperoxia compared to normoxia during cycling did not induce greater mitochondrial biogenesis and/or intrinsic function. It has been shown that hyperoxia does not increase in vivo mitochondrial respiration during exercise in obese untrained but only in patients with type II diabetes with impaired ex vivo mitochondrial respiration (Cree-Green et al., 2018). The unchanged in vivo mitochondrial respiration in obese untrained but overall healthy individuals when breathing hyperoxia can be explained by a lower mitochondrial O2 affinity (p50mito), a novel mechanism regulating oxygen diffusion from microvessels to muscle mitochondria with direct effects on oxygen consumption (Cardinale et al., 2018b). In the present study, the lack of change in the hematological variables that could have altered the O2 carry capacity and therefore O2 delivery over the intervention indicates that training with hyperoxia does not induce changes in the capacity for oxygen delivery or utilization.
The unchanged mitochondrial content pre to post intervention suggests either that our trained cyclists had already reached a mitochondrial content plateau prior to the intervention (Montero and Lundby, 2017), that the increase in O2 delivery with HYPER remained below mitochondrial capacity (maintained mitochondrial excess capacity) or that the training intervention did not increase the training volume to which our participants were accustomed to before the start of this study. The latter is supported by recent findings indicating that training volume significantly relates to CS activity (Granata et al., 2018) and that no plateau in CS activity should occur if training volume is constantly increased (Granata et al., 2018). However, the coefficient of variation of the change in OXPHOS measurements pre to post intervention (coefficient of variation of ~40%) was much larger than previously found in our laboratory (Cardinale et al., 2018a). The reason for this is unknown; we cannot discriminate if the variation came from the mitochondrial isolation procedure itself or if it was due to a large individual response to the hyperoxic and normoxic training stimulus. Nevertheless, the results on mitochondrial respiration should be interpreted with caution.
This study did not include direct neuromuscular measurements and it cannot be excluded that the numerically higher training intensity induced by hyperoxia increased muscle contractile properties which in turn led to small improvements in performance compared to normoxia. However, cycling efficiency was unchanged in both groups pre to post intervention. Furthermore, in Kilding et al., 2012’s study, hyperoxic training did not improve peak and mean power output during a 60-s cycle sprint compared to normoxia. Therefore, it is unlikely that our results are explained by improved neuromuscular properties post intervention in the hyperoxic group.
This study attempted to recreate the real training scenario of trained cyclists including (1) fluctuations of training intensities within the micro-cycle, (2) recovery days, and (3) a tapering period prior to post testing (Meeusen et al., 2013) in a periodized and polarized training intervention program while still maintaining the rigor of controlled trials with a parallel group study design. The significant improvement in performance in our participants indicates that the overall training intervention was successful.
In conclusion, this study showed that 6-week hyperoxic-supplemented high-intensity interval training produced a small, potentially meaningful effect on cycling performance. This response was not explained by cardiorespiratory, hematological, or mitochondrial factors measured in this study. The underlying mechanisms for the performance effects of hyperoxia training remain unexplained, and may raise ethical questions for elite sport.
The datasets generated for this study can be found in figshare, https://figshare.com/s/90ea6a92bec059112c6b.
This study was carried out in accordance with the recommendations of the Swedish Ethics Committee with written informed consent from all subjects. All subjects gave written informed consent in accordance with the Declaration of Helsinki. The protocol was approved by the Regional Ethical Review Board in Stockholm.
DC, RB, BE, FL, and PL contributed to the conception and design of the experiment. JL, TM, and OS supervised the training. DC, RB, FL, JL, TM, OS, and SM contributed to the data collection. DC analyzed and interpreted the data and wrote the first draft of the manuscript which was reviewed by the other co-authors. All authors read and approved the final manuscript.
This study was funded by The Swedish National Centre for Research in Sports (P2017-0054 and P2015-0133).
PL declares to have conflicts of interest and financial interest as co-founder of Oxelerate.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors would like to thank all the participants, Arndt T. for the administrative support, and Larsson Å. for lending the Oxelerate devices and help in making the gas administration setup.
Abbiss, C. R., Karagounis, L. G., Laursen, P. B., Peiffer, J. J., Martin, D. T., Hawley, J. A., et al. (2011). Single-leg cycle training is superior to double-leg cycling in improving the oxidative potential and metabolic profile of trained skeletal muscle. J. Appl. Physiol. 110, 1248–1255. doi: 10.1152/japplphysiol.01247.2010
Anselme, F., Caillaud, C., Couret, I., Rossi, M., and Prefaut, C. (1994). Histamine and exercise-induced hypoxemia in highly trained athletes. J. Appl. Physiol. 76, 127–132. doi: 10.1152/jappl.1994.76.1.127
Antharavally, B. S., Carter, B., Bell, P. A., and Krishna Mallia, A. (2004). A high-affinity reversible protein stain for Western blots. Anal. Biochem. 329, 276–280. doi: 10.1016/j.ab.2004.02.049
Borg, G. (1970). Perceived exertion as an indicator of somatic stress. Scand. J. Rehabil. Med. 2, 92–98.
Boushel, R., Gnaiger, E., Calbet, J. A., Gonzalez-Alonso, J., Wright-Paradis, C., Sondergaard, H., et al. (2011). Muscle mitochondrial capacity exceeds maximal oxygen delivery in humans. Mitochondrion 11, 303–307. doi: 10.1016/j.mito.2010.12.006
Brouwer, E. (1957). On simple formulae for calculating the heat expenditure and the quantities of carbohydrate and fat oxidized in metabolism of men and animals, from gaseous exchange (oxygen intake and carbonic acid output) and urine-N. Acta Physiol. Pharmacol. Neerl. 6, 795–802.
Burge, C. M., and Skinner, S. L. (1995). Determination of hemoglobin mass and blood volume with CO: evaluation and application of a method. J. Appl. Physiol. 79, 623–631.
Cardinale, D. A., and Ekblom, B. (2018). Hyperoxia for performance and training. J. Sports Sci. 36, 1515–1522. doi: 10.1080/02640414.2017.1398893
Cardinale, D. A., Gejl, K. D., Ørtenblad, N., Ekblom, B., Blomstrand, E., and Larsen, F. J. (2018a). Reliability of maximal mitochondrial oxidative phosphorylation in permeabilized fibers from the vastus lateralis employing high-resolution respirometry. Physiol. Rep. 6:e13611. doi: 10.14814/phy2.13611
Cardinale, D. A., Larsen, F. J., Jensen-Urstad, M., Rullman, E., Sondergaard, H., Morales-Alamo, D., et al. (2018b). Muscle mass and inspired oxygen influence oxygen extraction at maximal exercise: role of mitochondrial oxygen affinity. Acta Physiol. e13110. doi: 10.1111/apha.13110
Cheng, B., Kuipers, H., Snyder, A. C., Keizer, H. A., Jeukendrup, A., and Hesselink, M. (1992). A new approach for the determination of ventilatory and lactate thresholds. Int. J. Sports Med. 13, 518–522. doi: 10.1055/s-2007-1021309
Cohen, J. (1988). Statistical power analysis for the behavioral sciences (Hilldale, NJ: Lawrence Earlbaum).
Cree-Green, M., Scalzo, R. L., Harrall, K., Newcomer, B. R., Schauer, I. E., Huebschmann, A. G., et al. (2018). Supplemental oxygen improves in-vivo mitochondrial oxidative phosphorylation flux in sedentary obese adults with type 2 diabetes. Diabetes 67, 1369–1379. doi: 10.2337/db17-1124
Dempsey, J. A., McKenzie, D. C., Haverkamp, H. C., and Eldridge, M. W. (2008). Update in the understanding of respiratory limitations to exercise performance in fit, active adults. Chest 134, 613–622. doi: 10.1378/chest.07-2730
Dempsey, J. A., and Wagner, P. D. (1999). Exercise-induced arterial hypoxemia. J. Appl. Physiol. 87, 1997–2006. doi: 10.1152/jappl.1999.87.6.1997
Dominelli, P. B., Foster, G. E., Dominelli, G. S., Henderson, W. R., Koehle, M. S., McKenzie, D. C., et al. (2013). Exercise-induced arterial hypoxaemia and the mechanics of breathing in healthy young women. J. Physiol. 591, 3017–3034. doi: 10.1113/jphysiol.2013.252767
Ekblom, B., Astrand, P. O., Saltin, B., Stenberg, J., and Wallström, B. (1968). Effect of training on circulatory response to exercise. J. Appl. Physiol. 24, 518–528. doi: 10.1152/jappl.1968.24.4.518
Ekblom, B., Huot, R., Stein, E. M., and Thorstensson, A. T. (1975). Effect of changes in arterial oxygen content on circulation and physical performance. J. Appl. Physiol. 39, 71–75. doi: 10.1152/jappl.1975.39.1.71
Granata, C., Jamnick, N. A., and Bishop, D. J. (2018). Training-induced changes in mitochondrial content and respiratory function in human skeletal muscle. Sports Med. 48, 1809–1828. doi: 10.1007/s40279-018-0936-y
Guenette, J. A., and Sheel, A. W. (2007). Exercise-induced arterial hypoxaemia in active young women. Appl. Physiol. Nutr. Metab. 32, 1263–1273. doi: 10.1139/H07-122
Hamalainen, I. T., Nummela, A. T., and Rusko, H. K. (2000). Training in hyperoxia improves 3000-m running performance in national level athletes. Med. Sci. Sports Exerc. 32:S47.
Kilding, A. E., Wood, M., Sequira, G., and Bonetti, D. L. (2012). Effect of hyperoxic-supplemented interval training on endurance performance in trained cyclists. Int. J. Sports Med. 33, 359–363. doi: 10.1055/s-0031-1297999
Lindholm, P., Larsson, Å., Frånberg, O., and Gullstrand, L. (2017). A portable device for intermittent oxygen supplementation during high-intensity exercise. J. Biomed. Sci. Eng. 10:13. doi: 10.4236/jbise.2017.105023
Mallette, M. M., Stewart, D. G., and Cheung, S. S. (2017). The effects of hyperoxia on sea-level exercise performance, training, and recovery: a meta-analysis. Sports Med. 48, 153–175. doi: 10.1007/s40279-017-0791-2
Manselin, T. A., Södergård, O., Larsen, F. J., and Lindholm, P. (2017). Aerobic efficiency is associated with the improvement in maximal power output during acute hyperoxia. Physiol. Rep. 5. doi: 10.14814/phy2.13119
McGowan, C. J., Pyne, D. B., Thompson, K. G., and Rattray, B. (2015). Warm-up strategies for sport and exercise: mechanisms and applications. Sports Med. 45, 1523–1546. doi: 10.1007/s40279-015-0376-x
Meeusen, R., Duclos, M., Foster, C., Fry, A., Gleeson, M., Nieman, D., et al. (2013). Prevention, diagnosis, and treatment of the overtraining syndrome: joint consensus statement of the European College of Sport Science and the American College of Sports Medicine. Med. Sci. Sports Exerc. 45, 186–205. doi: 10.1249/MSS.0b013e318279a10a
Mogensen, M., Bagger, M., Pedersen, P. K., Fernström, M., and Sahlin, K. (2006). Cycling efficiency in humans is related to low UCP3 content and to type I fibres but not to mitochondrial efficiency. J. Physiol. 571, 669–681. doi: 10.1113/jphysiol.2005.101691
Montero, D., and Lundby, C. (2017). Refuting the myth of non-response to exercise training: ‘non-responders’ do respond to higher dose of training. J. Physiol. 595, 3377–3387. doi: 10.1113/JP273480
Morris, D. M., Kearney, J. T., and Burke, E. R. (2000). The effects of breathing supplemental oxygen during altitude training on cycling performance. J. Sci. Med. Sport 3, 165–175. doi: 10.1016/S1440-2440(00)80078-X
Nielsen, H. B. (2003). Arterial desaturation during exercise in man: implication for O2 uptake and work capacity. Scand. J. Med. Sci. Sports 13, 339–358. doi: 10.1046/j.1600-0838.2003.00325.x
Nielsen, H. B., Boushel, R., Madsen, P., and Secher, N. H. (1999). Cerebral desaturation during exercise reversed by O2 supplementation. Am. J. Phys. 277, H1045–H1052. doi: 10.1152/ajpheart.1999.277.3.H1045
Ohya, T., Yamanaka, R., Ohnuma, H., Hagiwara, M., and Suzuki, Y. (2016). Hyperoxia extends time to exhaustion during high-intensity intermittent exercise: a randomized, crossover study in male cyclists. Sports Med. Open 2:34. doi: 10.1186/s40798-016-0059-7
Perry, C. G., Reid, J., Perry, W., and Wilson, B. A. (2005). Effects of hyperoxic training on performance and cardiorespiratory response to exercise. Med. Sci. Sports Exerc. 37, 1175–1179. doi: 10.1249/01.mss.0000169610.33318.54
Perry, C. G., Talanian, J. L., Heigenhauser, G. J., and Spriet, L. L. (2007). The effects of training in hyperoxia vs. normoxia on skeletal muscle enzyme activities and exercise performance. J. Appl. Physiol. 102, 1022–1027. doi: 10.1152/japplphysiol.01215.2006
Pesta, D., and Gnaiger, E. (2012). High-resolution respirometry: OXPHOS protocols for human cells and permeabilized fibers from small biopsies of human muscle. Methods Mol. Biol. 810, 25–58. doi: 10.1007/978-1-61779-382-0_3
Plenge, U., Belhage, B., Guadalupe-Grau, A., Andersen, P. R., Lundby, C., Dela, F., et al. (2012). Erythropoietin treatment enhances muscle mitochondrial capacity in humans. Front. Physiol. 3:50. doi: 10.3389/fphys.2012.00050
Ploutz-Snyder, L. L., Simoneau, J. A., Gilders, R. M., Staron, R. S., and Hagerman, F. C. (1996). Cardiorespiratory and metabolic adaptations to hyperoxic training. Eur. J. Appl. Physiol. Occup. Physiol. 73, 38–48. doi: 10.1007/BF00262807
Powers, S. K., Dodd, S., Lawler, J., Landry, G., Kirtley, M., McKnight, T., et al. (1988). Incidence of exercise induced hypoxemia in elite endurance athletes at sea level. Eur. J. Appl. Physiol. Occup. Physiol. 58, 298–302. doi: 10.1007/BF00417266
Powers, S. K., Lawler, J., Dempsey, J. A., Dodd, S., and Landry, G. (1989). Effects of incomplete pulmonary gas exchange on VO2 max. J. Appl. Physiol. 66, 2491–2495. doi: 10.1152/jappl.1989.66.6.2491
Przyklenk, A., Gutmann, B., Schiffer, T., Hollmann, W., Strueder, H. K., Bloch, W., et al. (2017). Endurance exercise in hypoxia, hyperoxia and normoxia: mitochondrial and global adaptations. Int. J. Sports Med. 38, 588–596. doi: 10.1055/s-0043-106740
Saltin, B., and Calbet, J. A. L. (2006). Point: in health and in a normoxic environment, V̇O2 max is limited primarily by cardiac output and locomotor muscle blood flow. J. Appl. Physiol. 100, 744–745. doi: 10.1152/japplphysiol.01395.2005
Seiler, S. (2010). What is best practice for training intensity and duration distribution in endurance athletes? Int. J. Sports Physiol. Perform. 5, 276–291. doi: 10.1123/ijspp.5.3.276
St Croix, C. M., Harms, C. A., McClaran, S. R., Nickele, G. A., Pegelow, D. F., Nelson, W. B., et al. (1998). Effects of prior exercise on exercise-induced arterial hypoxemia in young women. J. Appl. Physiol. 85, 1556–1563. doi: 10.1152/jappl.1998.85.4.1556
Steinlechner-Maran, R., Eberl, T., Kunc, M., Margreiter, R., and Gnaiger, E. (1996). Oxygen dependence of respiration in coupled and uncoupled endothelial cells. Am. J. Phys. 271, C2053–C2061.
Stellingwerff, T., Leblanc, P. J., Hollidge, M. G., Heigenhauser, G. J., and Spriet, L. L. (2006). Hyperoxia decreases muscle glycogenolysis, lactate production, and lactate efflux during steady-state exercise. Am. J. Physiol. Endocrinol. Metab. 290, E1180–E1190. doi: 10.1152/ajpendo.00499.2005
Keywords: cycling performance, hyperoxia, high-intensity interval training, mitochondria, OXPHOS, VO2max
Citation: Cardinale DA, Larsen FJ, Lännerström J, Manselin T, Södergård O, Mijwel S, Lindholm P, Ekblom B and Boushel R (2019) Influence of Hyperoxic-Supplemented High-Intensity Interval Training on Hemotological and Muscle Mitochondrial Adaptations in Trained Cyclists. Front. Physiol. 10:730. doi: 10.3389/fphys.2019.00730
Edited by:
Antonio Crisafulli, University of Cagliari, ItalyReviewed by:
Peter Rasmussen, University of Zurich, SwitzerlandCopyright © 2019 Cardinale, Larsen, Lännerström, Manselin, Södergård, Mijwel, Lindholm, Ekblom and Boushel. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: D. A. Cardinale, ZGFuaWVsZS5jYXJkaW5hbGVAZ2loLnNl; R. Boushel, cm9iZXJ0LmJvdXNoZWxAdWJjLmNh
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.