 Ilka Starke1,2
Ilka Starke1,2 Michael Börsch
Michael Börsch- 1Single-Molecule Microscopy Group, Jena University Hospital, Friedrich Schiller University, Jena, Germany
- 2Institute for Physical Chemistry, Albert Ludwigs University of Freiburg, Freiburg, Germany
- 3Department of Chemistry, University of Michigan, Ann Arbor, MI, United States
- 4Abbe Center of Photonics, Friedrich Schiller University, Jena, Germany
Targeting the mitochondrial enzyme FoF1-ATP synthase and modulating its catalytic activities with small molecules is a promising new approach for treatment of autoimmune diseases. The immunomodulatory compound Bz-423 is such a drug that binds to subunit OSCP of the mitochondrial FoF1-ATP synthase and induces apoptosis via increased reactive oxygen production in coupled, actively respiring mitochondria. Here, we review the experimental progress to reveal the binding of Bz-423 to the mitochondrial target and discuss how subunit rotation of FoF1-ATP synthase is affected by Bz-423. Briefly, we report how Förster resonance energy transfer can be employed to colocalize the enzyme and the fluorescently tagged Bz-423 within the mitochondria of living cells with nanometer resolution.
Introduction
Cellular processes as metabolism and transport are powered by the universal chemical “energy currency” that is the molecule adenosine triphosphate (ATP). Therefore, millimolar ATP concentrations inside the cell have to be produced and maintained through sequential catalytic reactions by the glycolysis pathway or more efficiently by the FoF1-ATP synthase as part of the oxidative phosphorylation (OXPHOS) pathway. FoF1-ATP synthases are working in the plasma membrane of bacteria or in small organelles inside of eukaryotes, i.e., the thylakoid membrane in chloroplasts or the inner mitochondrial membrane. In case of mitochondrial FoF1-ATP synthase a proton motive force (PMF) comprising a concentration difference of protons across the membrane (ΔpH) plus an electric membrane potential (ΔΨ) is required for ATP synthesis. The PMF is generated by sequential redox processes and associated proton pumping of the enzyme complexes I to IV of the respiratory chain across the inner mitochondrial membrane.
If we consider autoimmune diseases, for example systemic lupus erythematodes, being caused by hyperactivity of pathogenic T cells of the immune system, then controlling their cellular ATP concentration with drugs and reducing their activity could become a promising approach for clinical treatment. Modulating T cell activity temperately could circumvent a complete shut-down of the normal immune function. Therefore, one option would be controlling the PMF by targeting any of the enzyme complexes I to IV of the respiratory chain with inhibitors. Vice versa, controlling the efficiency of converting PMF to ATP synthesis by FoF1-ATP synthase would be a possible approach. This latter process is called uncoupling.
More than a decade ago, a 1,4-benzodiazepine, Bz-423 (Figure 1A), has been found to target lymphoid cells in a murine model of lupus erythematodes (Blatt et al., 2002). Bz-423 specifically induced apoptosis of pathogenic lymphocytes and attenuated disease progression. As a result, the treated mice showed a prolonged survival at the therapeutic dosage without adverse toxicity and with maintained immune function (Bednarski et al., 2003). The mechanism of Bz-423 action was revealed and subsequently the molecular target was identified – the mitochondrial FoF1-ATP synthase (Johnson et al., 2005). Here we focus on the discovery of the drug binding site and discuss a recent microscopy approach using Förster resonance energy transfer (FRET) that has directly demonstrated the binding of a fluorescent Bz-423 derivative to the mitochondrial enzyme in living cells (Starke et al., 2016).
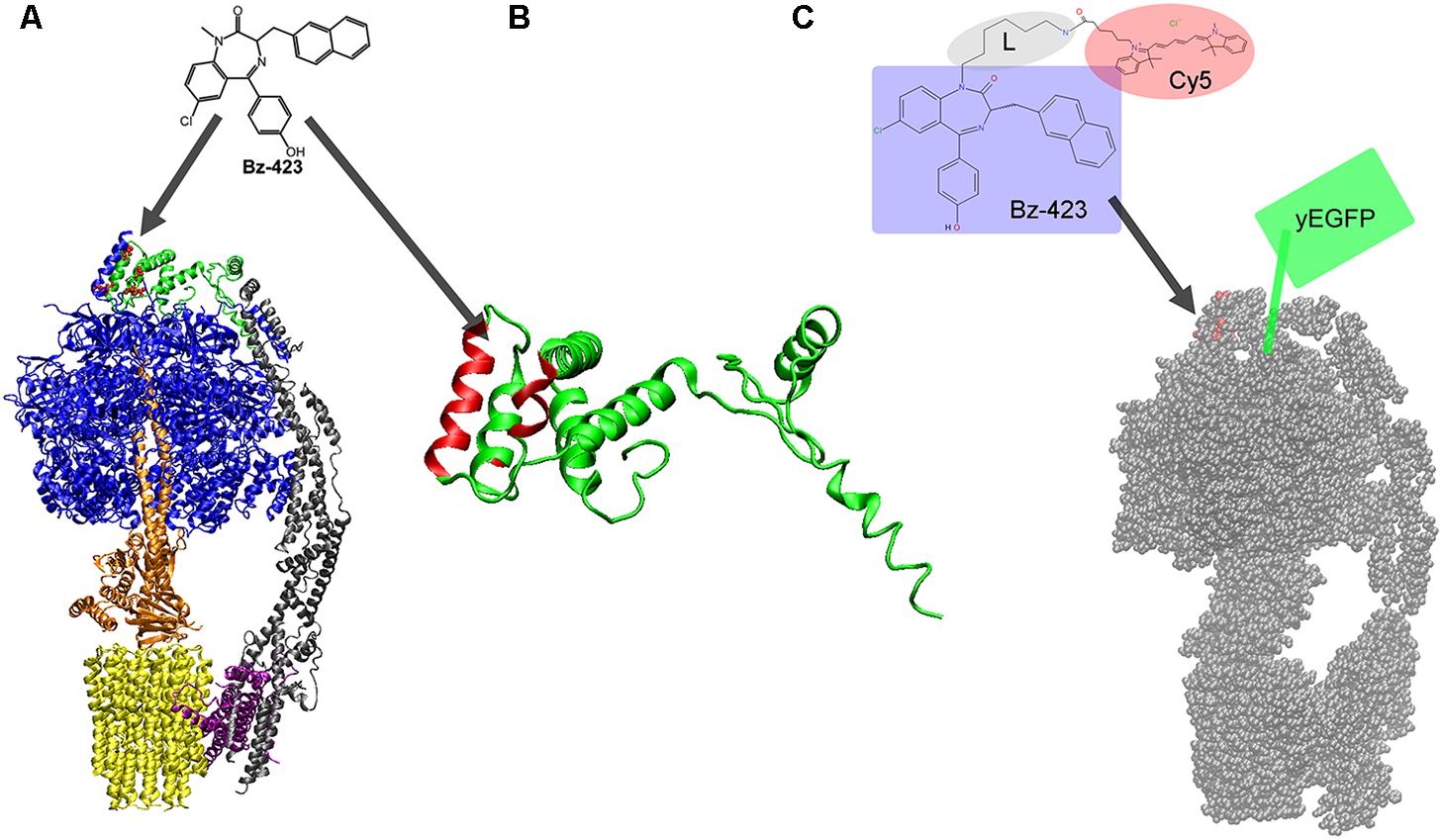
FIGURE 1. (A) Structure of the monomeric mitochondrial FoF1-ATP synthase from yeast with binding site for Bz-423 (red dots) on subunit OSCP (in green, on the top; from cryoEM data, PDB 6CP3 (Srivastava et al., 2018). Subunits α and β are shown in blue and the rotary subunits γ, δ, and 𝜀 in orange of the F1 part. The peripheral static domain consists of subunits b, d, F6, f, 8, i, j (in gray) and OSCP. The proton-translocating subunit a is depicted in purple, and the rotor ring of the Fo part comprising eight c subunits is shown in yellow. (B) Structure of OSCP from yeast (PDB 6CP3) with helices highlighted in red (including residues 51, 55, 65, 66, 75, 77, and 92) shown to be involved in Bz-423 binding. (C) Bz-423 derivative with the 1,4-benzodiazepine moiety highlighted by the bluish box, a flexible hexyl linker L (gray ellipse) and the FRET acceptor fluorophore Cy5 (red ellipse). Bz-423-Cy5 is expected to bind to OSCP of the yeast mitochondrial FoF1-ATP synthase (red dots on the top of the monomer of the yeast enzyme, cryoEM data, PDB 6B8H (Guo et al., 2017). The FRET donor yeast-enhanced green fluorescent protein (yEGFP) is fused to the extended C-terminus of the γ subunit (symbolized by the green box).
Bz-423 Binds to Oscp of Mitochondrial FoF1-ATP Synthase
Initially, Bz-423 was identified as a potential drug candidate from a library of 1,4-benzodiazepines generated by diversity-oriented chemical synthesis (Blatt et al., 2002). Phenotype screening of Ramos B cells revealed that Bz-423 caused cell shrinkage, nuclear condensation, cytoplasmic vacuolization, membrane blebbing, and DNA fragmentation. Other 1,4-benzodiazepine derivatives were found to selectively target T cells (Francis et al., 2006; Gatza et al., 2011; Wahl et al., 2012; Tkachev et al., 2015). Bz-423 did not strongly bind to the peripheral benzodiazepine receptor. Cytotoxicity of Bz-423 was related to rapidly generated superoxide (O2∙-) in mitochondria. Superoxide is one of the reactive oxygen species (ROS) that can chemically damage cellular macromolecules at higher concentrations. However, Bz-423 superoxide signaling for induced apoptosis was proven as the underlying mechanism (Blatt et al., 2008, 2009).
In the presence of 1 mM sodium azide, the proapoptotic O2∙- generation by Bz-423 was abolished (Blatt et al., 2002). Because sodium azide is an inhibitor of cytochrome c oxidase, i.e., complex IV of the mitochondrial respiratory chain, Bz-423 was proposed to bind to a mitochondrial OXPHOS protein. Binding of Bz-423 did not alter or collapse the electric potential ΔΨ across the inner mitochondrial membrane. The superoxide response by Bz-423 required active mitochondria in state 3 respiration, but not mitochondria in state 4 with minimal respiration and in the absence of ADP. Comparing the superoxide generation mechanism induced by oligomycin that induces a state-3-to-4 transition of mitochondrial respiration (Korshunov et al., 1997) lead to the hypothesis that Bz-423 might cause a state-3-to-4 transition as well and might bind to FoF1-ATP synthase.
The validation of FoF1-ATP synthase as the mitochondrial target of Bz-423 was achieved by phage display screening (Johnson et al., 2005). Briefly, the oligomycin sensitivity conferring protein (OSCP) being a subunit of FoF1-ATP synthase was determined. Subsequently the binding site of Bz-423 was located by NMR spectroscopy using the isolated subunit OSCP in solution (Stelzer et al., 2010). Figure 1A shows the structure of the yeast FoF1-ATP synthase with the highlighted subunit OSCP in green and the amino acid residues involved in binding of Bz-423 as red dots. Accordingly Bz-423 binds to the top of the membrane enzyme at the interface of OSCP with one pair of α, β subunits (blue in Figure 1A) of the F1 part. Binding of water-soluble Bz-423 analogs to OSCP in a chemical shift perturbation NMR measurement revealed specific residues 51, 55, 65, 66, 75, 77, and 92 that might form a hydrophobic pocket to accommodate the drug (Figure 1B). Furthermore, Bz-423 binding resulted in a conformational rearrangement of helices in OSCP and might alter the interaction of OSCP with F1 in an allosteric manner (Stelzer et al., 2010).
Bz-423 Requires Oscp to Modulate FoF1-ATP Synthase Activity In Vitro and in Cells
Binding of Bz-423 to OSCP in the intact FoF1-ATP synthase inhibited both synthesis and hydrolysis of ATP in isolated sub-mitochondrial particles (SMPs) in vitro (Johnson et al., 2005, 2006). Both maximal turnover Vmax and KM were changed, in contrast to the inhibitor oligomycin which reduced Vmax only. ATP hydrolysis by the soluble mitochondrial F1 part (comprising the blue and orange colored subunits in Figure 1A) was reduced but only when F1 was assembled with OSCP. The IC50 for Bz-423 was about 5 μM. In perfused HEK cells ATP synthesis rates of mitochondria were reduced, with IC50 of less than 5 μM for Bz-423. Engineered HEK cells with a specifically reduced content of OSCP by siRNA showed alleviated apoptosis by Bz-423, and the residual amount of cellular OSCP correlated well with the percentage of apoptotic cells (Johnson et al., 2005).
FoF1-ATP synthase accomplishes ATP synthesis by mechanochemical energy conversion with two rotary subunit motors. The PMF drives the Fo motor when protons enter the half-channel of the membrane-embedded a subunit (purple in Figure 1A) from the intermembrane space, i.e., from below and along the horizontally tilted helices (Allegretti et al., 2015). Binding to a specific residue on one c subunit (yellow in Figure 1A) compensates electrostatic forces and allows the ring of c subunits to rotate one step forward. Rotation of the c-ring moves the elastically-coupled attached F1 motor (orange in Figure 1A) comprising subunits γ, δ, and 𝜀. The F1 motor rotates in three 120° steps at high PMF and stops at each of the three catalytic sites where ATP is synthesized in F1. These distinct step sizes of the rotary Fo and F1 motors during ATP synthesis have been measured in vitro in single-molecule experiments using bacterial FoF1-ATP synthases (Diez et al., 2004; Zimmermann et al., 2005; Duser et al., 2009). FoF1-ATP synthase can also run in reverse by hydrolyzing ATP. ATP hydrolysis has been used to investigate the F1 motor in great detail since 20 years (Noji et al., 1997) and revealed torque, elastic domains (Wachter et al., 2011; Sielaff and Borsch, 2013; Martin et al., 2018), substeps (Yasuda et al., 2001; Suzuki et al., 2014), breaks and other motor properties (Yoshida et al., 2001; Junge et al., 2009; Borsch and Duncan, 2013).
Internal subunit rotation with high torque requires a mechanically stable stator counterpart of the enzyme. The static domain of the mitochondrial FoF1-ATP synthase comprises the six F1 subunits α3β3 (blue in Figure 1A), subunits b, d, F6, f, 8, i, j (gray in Figure 1A), the a subunit (purple in Figure 1A) and OSCP (green in Figure 1A). Binding of Bz-423 to the interface of OSCP with α3β3 might weaken the stator/F1 assembly. Alternatively, Bz-423 might influence the subtle conformational changes of OSCP bound to the top part of the catalytic α3β3 subunits and thereby provokes reduced rates of ATP synthesis and hydrolysis. Quantitative enzymatic analysis revealed that Bz-423 is an uncompetitive inhibitor of mitochondrial FoF1-ATP synthase decreasing Vmax for ATP synthesis to 50% in the presence of ∼10 μM Bz-423 (Johnson et al., 2006). Inhibition by μM amounts of Bz-423 corresponded to fast off-rates <0.3 s-1 of the drug from FoF1-ATP synthase and a 90% recovery of ATP synthesis rates after 10 s.
Imaging the Drug and Its Molecular Target in Mitochondria
Localizing a drug at a specific target in life cells can be achieved using fluorescence microscopy with high spatial and temporal resolution as well as single molecule sensitivity. A variety of functional Bz-423 derivatives was synthesized with a flexible linker to rhodamine- or cyanine fluorophores, for example Bz-423-Cy5 (Figure 1C). Because μM concentrations of Bz-423 are required to bind to OSCP and to induce apoptosis, direct imaging of the fluorescent drug bound to FoF1-ATP synthase in the inner mitochondrial membrane is not possible due to a high fluorescent background of unbound Bz-423 throughout the cell. Fast off-rates of Bz-423 and its fluorescent derivatives prevent extensive washing of the cells which is needed to obtain a good imaging contrast. Therefore, confocal microscopy with about 250 nm resolution or superresolution microscopies like structured illumination microscopy (SIM) (Shao et al., 2011) (resolution limit of about 100 nm) or stimulated emission depletion (STED) (Hell, 2007) (resolution limit of about 20 nm) cannot be used to identify bound Bz-423 on OSCP.
Instead, FRET (Förster, 1946, 2012) as a distance measurement approach in the 2 to 9 nm range is applicable. The dipole–dipole interaction of FRET between two nearby fluorophores causes a relative loss of fluorescence intensity of the FRET donor (excited by the laser) and an increase of fluorescence intensity of the FRET acceptor. Thus FRET can be used to relate the spatial position of the fluorescently labeled drug to its cellular target that is tagged with a different fluorophore. Mitochondrial FoF1-ATP synthase can be tagged with fluorescent proteins, for example at OSCP (Prescott et al., 1997) or at the rotary γ subunit without affecting function (Prescott et al., 2003; Muster et al., 2010; Foertsch et al., 2017). The benefit of using a genetic fusion to the enzyme is a negligible fluorescent background from other parts of the cell than the cristae of the inner mitochondrial membrane.
Revealing Bz-423-Cy5 Binding to FoF1-ATP Synthase by Fret Acceptor Photobleaching
To detect binding of fluorescent Bz-423 to mitochondrial FoF1-ATP synthase in living Saccharomyces cerevisiae cells, we used a FoF1-ATP synthase mutant designed by J. Petersen and P. Gräber (I. Starke, Ph.D. thesis, University of Freiburg, 2015). A fusion of the yeast-enhanced green fluorescent protein (yEGFP) linked to the C-terminus of the γ subunit (Figure 1C) provided the donor for FRET imaging (Figure 2). The fully functional mutant was checked for ATP synthesis, and catalytic rates were also determined in vitro after protein purification and reconstitution to liposomes. Staining mitochondria was achieved by incubating the S. cerevisiae cells with 4 μm Bz-423-Cy5 (see structure in Figure 1C) in the presence of 2% EtOH for 8 h at 28°C. After washing, the bluish cells (Figure 2) were imaged immediately on the microscope at 22°C (Starke et al., 2016).
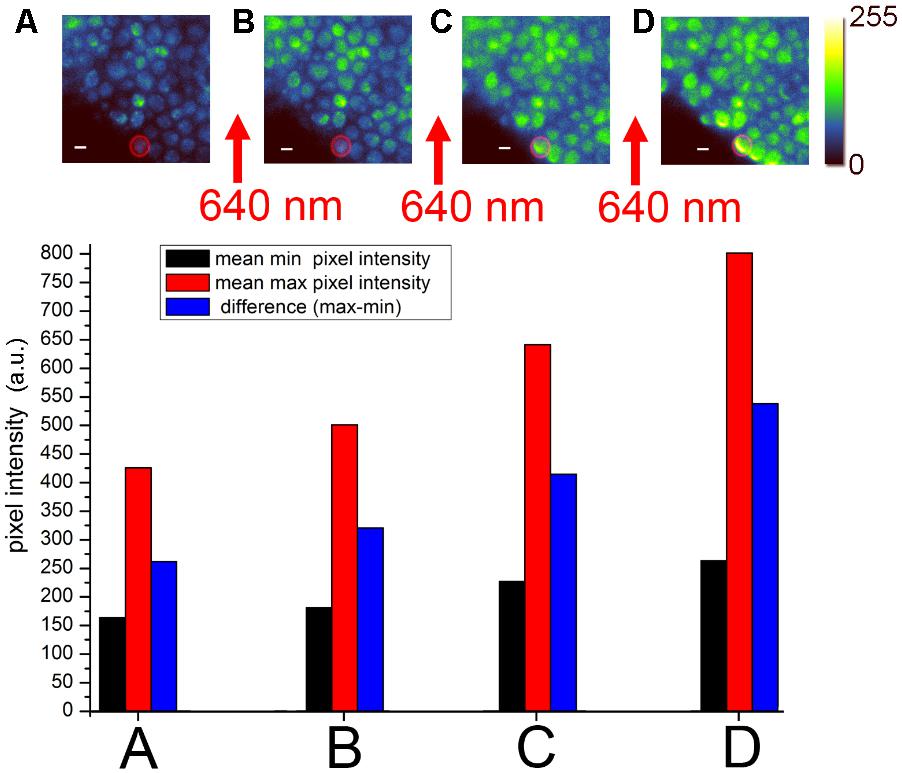
FIGURE 2. Förster resonance energy transfer (FRET) acceptor photobleaching of Bz-423-Cy5 bound to mitochondrial FoF1-ATP synthase with yEGFP fusion to the γ subunit in living Saccharomyces cerevisiae (Starke et al., 2016). EMCCD-based widefield fluorescence microscopy used laser excitation of yEGFP with 488 nm and fluorescence detection of yEGFP between 500 and 550 nm. (A–D) Sequential imaging of yEGFP-tagged FoF1-ATP synthases with recalculated, false-colored intensities in 8 bit (0–255; upper panel). Between each image, a 30-s high-power laser pulse with 640 nm was applied to partially photobleach the Cy5 chromophore. The individual mean pixel intensities of the highlighted cell [red circle as the region-of-interest (ROI)] are plotted in the histograms in the lower panel as recorded by the EMCCD camera [modified from Starke et al. (2016) with permission].
Widefield fluorescence microscopy of the stained yeast showed spherical cells with bright spots suggesting fluorescent FoF1-ATP synthase in mitochondria (Figure 2A). SIM imaging of unstained yeast confirmed that only mitochondria were fluorescent (Starke et al., 2016). Repeated imaging of the same field of view indicated comparable pixel intensities in these cells revealing only minor photobleaching of yEGFP. However, subsequent excitation of the cells with 640 nm at high power photobleached the FRET acceptor Cy5 dye on Bz-423, and the loss of FRET acceptor resulted in an increase of the FRET donor intensity when imaged again with 488 nm (Figure 2B). Stepwise photobleaching of Cy5 correlated with a stepwise increase of FRET donor fluorescence (Figures 2C,D). Analysis of the intensity increase in individual cells (see histograms of the single cell highlighted by the red circle in Figures 2A–D) due to Cy5 photobleaching unequivocally corroborated the binding of Bz-423-Cy5 to a position on FoF1-ATP synthase only few nm away from the yEGFP chromophore at the extended C-terminus of the γ subunit. A background fluorescence increase (black bars in the histograms) after each 640 nm exposure was subtracted from the increase of maximum fluorescence (red bars) to obtain the effective increase of FRET donor fluorescence (blue bars). Most likely, Bz-423-Cy5 was bound to OSCP.
Förster resonance energy transfer acceptor photobleaching to confirm the molecular target of a drug in living cells is a fluorescence microscopy approach that requires fast imaging capability but not necessarily high photon counts rates per pixel to achieve accurate colocalization. Despite weak (μM) binding affinities of Bz-423-Cy5 and fast exchange of the immunomodulator on OSCP, a significant fraction of bound Bz-423-Cy5 was revealed by FRET. Slow transport of Bz-423 across the membranes into the matrix of mitochondria as the final destination was indicated by long incubation times needed for staining the yeast cells. Accordingly, a significant fraction of Bz-423-Cy5 still remained in the cytosol and outside of the mitochondria during FRET acceptor photobleaching, and did not contribute to apoptotic action. However, slow exchange of photobleached Bz-423-Cy5 into and out of the matrix compartment facilitated the FRET detection.
Future Developments
Following the first demonstration that the 1,4-benzodiazepine Bz-423 induced apoptosis in living B cells by stimulating superoxid production of the OXPHOS complexes (Blatt et al., 2002) the identification of its molecular target in mitochondria was accomplished by biochemical methods. Subunit OSCP of FoF1-ATP synthase being the destination of the drug was unraveled by human cDNA T7 phage display screening. Genetic removal of OSCP in mutant FoF1-ATP synthases proved that apoptosis required binding of Bz-423 to this subunit. Using purified soluble OSCP, the amino acids involved in the binding site were discovered by NMR spectroscopy (Stelzer et al., 2010).
A detailed view on the Bz-423 binding site at the interface of OSCP with the N-termini of α and β subunits of F1 is permitted by the recent high-resolution CryoEM structures of the complete mitochondrial enzymes from bovine heart and from yeast S. cerevisiae (Zhou et al., 2015; Guo et al., 2017; Srivastava et al., 2018). Both OSCP and the N-termini of α and β subunits change their conformations during catalysis, and Bz-423 binding might interfere with these changes and might retard the turnover. Subsequently the mitochondrial PMF builds up, mitochondria switch from respirational state 3 to 4, superoxide is produced, and Ca2+ signaling and apoptosis are initiated. Bz-423 induced the opening of the mitochondrial permeability transition pore and thus decreased the Ca2+ retention capability (Giorgio et al., 2013, 2017; Antoniel et al., 2014, 2018; Bernardi et al., 2015).
The weak binding affinities of Bz-423 and its fluorescent derivatives like Bz-423-Cy5 prevented direct fluorescence microscopy approaches in living cells. In initial confocal microscopy experiments we noticed a broad spatial distribution of Bz-423-Cy5 in S. cerevisiae cells but not a specific staining of the mitochondria. Therefore we evaluated the use of SMPs with fluorescently tagged FoF1-ATP synthase for FRET in vitro but failed to detect sensitized FRET acceptor emission due to the high fluorescent background of unbound Bz-423-Cy5. The solution for a FRET-based direct detection of Bz-423 binding to OSCP was FRET acceptor photobleaching in mitochondria of living cells. Here the pool of unbound Bz-423-Cy5 is limited, and photobleaching Cy5 with 640 nm at high power is possible without destroying the FRET donor fluorophore yEGFP on FoF1-ATP synthase. Now, similar FRET experiments are under way with human HEK cells and by using the brightest and more photostable green fluorescent protein mNeonGreen (Shaner et al., 2013) fused to the C-terminus of the γ subunit of FoF1-ATP synthase (Foertsch et al., 2017).
To improve specificity of the cellular distribution of the hydrophobic Bz-423 derivatives and to accelerate an uptake into mitochondria, attaching cationic dyes could be used. As shown by Irion et al. (1993), Rottele and Zimmermann (1993), Schneider and Zimmermann (1994), and Schneider et al. (1994), almost all fluorophores being lipophilic cations transfer quickly to the inner mitochondrial membrane. There, they can bind to proteins. One identified target was cytochrome C oxidase. This finding could be used to specifically induce photodamage by singlet oxygen as a ROS (Cernay and Zimmermann, 1996; Dummin et al., 1997; Borsch, 2010) for Photodynamic Therapy. Beside photoaffinity labeling approaches, time-resolved FRET was applied to reveal that cytochrome C oxidase was a binding site of these lipophilic cationic photosensitizers acting as FRET donors (Huglin et al., 1995; Borsch, 2010). Similarly, confocal imaging FRET donor lifetimes of mitochondrial FoF1-ATP synthases in the presence and absence of FRET acceptor-tagged Bz-423 derivatives could be employed to provide direct optical evidence for Bz-423 binding to FoF1-ATP synthase. Alternatively, a photoaffinity tag on Bz-423 might be necessary to detect binding of the drug to the multimeric form of the enzyme.
Förster resonance energy transfer-based direct evidence for Bz-423 acting at the mitochondrial FoF1-ATP synthase needs to be complemented by detailed analysis of cristae morphology changes. In mitochondria of living cells, this can be accomplished by superresolution microscopy approaches, for example SIM (Shao et al., 2011) or STED (Schmidt et al., 2009) microscopy. Especially, a rearrangement of row assembly of dimeric FoF1-ATP synthases at the rim of the cristae could indicate the beginning and early events of apoptosis. Finally, an unequivocal mechanistic demonstration of how Bz-423 affects the catalytic activity and the rotary motors of mitochondrial FoF1-ATP synthase is awaited. Single-molecule FRET between different fluorophores attached to rotor and stator of the mitochondrial enzyme has already been developed. Accordingly, the inhibition mechanism of Bz-423 could be unraveled by single-molecule rotation experiments, as shown previously for the inhibitor Aurovertin (Johnson et al., 2009) or of phytopolyphenols (Sekiya et al., 2017).
Author Contributions
All authors designed the experiments and provided chemicals, biological materials and microscopy tools. All authors wrote the manuscript and approved it for publication.
Funding
This work was supported by NIH grant AI-47450 to GG and in part by DFG grant BO 1891/15-1 and by the DFG Collaborative Research Center Transregio 166 “ReceptorLight” project A1 to MB. The N-SIM/N-STORM superresolution microscope was funded by the State of Thuringa grant FKZ 12026-515 to MB.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We gratefully acknowledge the collaboration with Mark Prescott (Monash University), Jan Petersen and Peter Gräber (Albert Ludwigs University of Freiburg) who designed and provided γ subunit mutants of the mitochondrial enzyme of S. cerevisiae. Kathryn M. Johnson synthesized the Cy5-tagged Bz-423 (University of Michigan).
References
Allegretti, M., Klusch, N., Mills, D. J., Vonck, J., Kuhlbrandt, W., and Davies, K. M. (2015). Horizontal membrane-intrinsic α-helices in the stator a-subunit of an F-type ATP synthase. Nature 521, 237–240. doi: 10.1038/nature14185
Antoniel, M., Giorgio, V., Fogolari, F., Glick, G. D., Bernardi, P., and Lippe, G. (2014). The oligomycin-sensitivity conferring protein of mitochondrial ATP synthase: emerging new roles in mitochondrial pathophysiology. Int. J. Mol. Sci. 15, 7513–7536. doi: 10.3390/ijms15057513
Antoniel, M., Jones, K., Antonucci, S., Spolaore, B., Fogolari, F., Petronilli, V., et al. (2018). The unique histidine in OSCP subunit of F-ATP synthase mediates inhibition of the permeability transition pore by acidic pH. EMBO Rep. 19, 257–268. doi: 10.15252/embr.201744705
Bednarski, J. J., Warner, R. E., Rao, T., Leonetti, F., Yung, R., Richardson, B. C., et al. (2003). Attenuation of autoimmune disease in Fas-deficient mice by treatment with a cytotoxic benzodiazepine. Arthritis Rheum. 48, 757–766. doi: 10.1002/art.10968
Bernardi, P., Rasola, A., Forte, M., and Lippe, G. (2015). The mitochondrial permeability transition pore: channel formation by F-ATP synthase, integration in signal transduction, and role in pathophysiology. Physiol. Rev. 95, 1111–1155. doi: 10.1152/physrev.00001.2015
Blatt, N. B., Bednarski, J. J., Warner, R. E., Leonetti, F., Johnson, K. M., Boitano, A., et al. (2002). Benzodiazepine-induced superoxide signals B cell apoptosis: mechanistic insight and potential therapeutic utility. J. Clin. Invest. 110, 1123–1132. doi: 10.1172/JCI16029
Blatt, N. B., Boitano, A. E., Lyssiotis, C. A., Opipari, A. W. Jr., and Glick, G. D. (2008). Bz-423 superoxide signals apoptosis via selective activation of JNK, Bak, and Bax. Free Radic. Biol. Med. 45, 1232–1242. doi: 10.1016/j.freeradbiomed.2008.07.022
Blatt, N. B., Boitano, A. E., Lyssiotis, C. A., Opipari, A. W. Jr., and Glick, G. D. (2009). Bz-423 superoxide signals B cell apoptosis via Mcl-1, Bak, and Bax. Biochem. Pharmacol. 78, 966–973. doi: 10.1016/j.bcp.2009.05.025
Borsch, M. (2010). Targeting cytochrome C oxidase in mitochondria with Pt(II)-porphyrins for photodynamic therapy. Prog. Biomed. Opt. Imaging Proc. SPIE 7551:75510G. doi: 10.1117/12.841284
Borsch, M., and Duncan, T. M. (2013). Spotlighting motors and controls of single FoF1-ATP synthase. Biochem. Soc. Trans. 41, 1219–1226. doi: 10.1042/BST20130101
Cernay, T., and Zimmermann, H. W. (1996). Selective photosensitization of mitochondria by the lipophilic cationic porphyrin POR10. J. Photochem. Photobiol. B 34, 191–196. doi: 10.1016/1011-1344(95)07267-5
Diez, M., Zimmermann, B., Borsch, M., Konig, M., Schweinberger, E., Steigmiller, S., et al. (2004). Proton-powered subunit rotation in single membrane-bound FoF1-ATP synthase. Nat. Struct. Mol. Biol. 11, 135–141. doi: 10.1038/nsmb718
Dummin, H., Cernay, T., and Zimmermann, H. W. (1997). Selective photosensitization of mitochondria in HeLa cells by cationic Zn (II) phthalocyanines with lipophilic side-chains. J. Photochem. Photobiol. B 37, 219–229. doi: 10.1016/S1011-1344(96)07416-7
Duser, M. G., Zarrabi, N., Cipriano, D. J., Ernst, S., Glick, G. D., Dunn, S. D., et al. (2009). 36 degrees step size of proton-driven c-ring rotation in FoF1-ATP synthase. EMBO J. 28, 2689–2696. doi: 10.1038/emboj.2009.213
Foertsch, F., Ilchenko, M., Heitkamp, T., Nossmann, S., Hoffmann, B., Starke, I., et al. (2017). Imaging cytochrome C oxidase and F0F1-ATP synthase in mitochondrial cristae of living human cells by FLIM and superresolution microscopy. Proc. SPIE 10071:100710. doi: 10.1117/12.2251210
Förster, T. (1946). Energiewanderung und fluoreszenz. Naturwissenschaften 33, 166–175. doi: 10.1007/BF00585226
Förster, T. (2012). Energy migration and fluorescence. J. Biomed. Opt. 17:011002. doi: 10.1117/1.JBO.17.1.011002
Francis, T. M., Sundberg, T. B., Cleary, J., Groendyke, T., Opipari, A. W. Jr., and Glick, G. D. (2006). Identification of cytotoxic, T-cell-selective 1,4-benzodiazepine-2,5-diones. Bioorg. Med. Chem. Lett. 16, 2423–2427. doi: 10.1016/j.bmcl.2006.01.113
Gatza, E., Wahl, D. R., Opipari, A. W., Sundberg, T. B., Reddy, P., Liu, C., et al. (2011). Manipulating the bioenergetics of alloreactive T cells causes their selective apoptosis and arrests graft-versus-host disease. Sci. Transl. Med. 3:67ra68. doi: 10.1126/scitranslmed.3001975
Giorgio, V., Burchell, V., Schiavone, M., Bassot, C., Minervini, G., Petronilli, V., et al. (2017). Ca2+ binding to F-ATP synthase beta subunit triggers the mitochondrial permeability transition. EMBO Rep. 18, 1065–1076. doi: 10.15252/embr.201643354
Giorgio, V., von Stockum, S., Antoniel, M., Fabbro, A., Fogolari, F., Forte, M., et al. (2013). Dimers of mitochondrial ATP synthase form the permeability transition pore. Proc. Natl. Acad. Sci. U.S.A. 110, 5887–5892. doi: 10.1073/pnas.1217823110
Guo, H., Bueler, S. A., and Rubinstein, J. L. (2017). Atomic model for the dimeric FO region of mitochondrial ATP synthase. Science 358, 936–940. doi: 10.1126/science.aao4815
Hell, S. W. (2007). Far-field optical nanoscopy. Science 316, 1153–1158. doi: 10.1126/science.1137395
Huglin, D., Seiffert, W., and Zimmermann, H. W. (1995). Time-resolved microfluorometric study of the binding sites of lipophilic cationic pyrene probes in mitochondria of living HeLa cells. J. Photochem. Photobiol. B 31, 145–158. doi: 10.1016/1011-1344(95)07191-1
Irion, G., Ochsenfeld, L., Naujok, A., and Zimmermann, H. W. (1993). The concentration jump method. Kinetics of vital staining of mitochondria in HeLa cells with lipophilic cationic fluorescent dyes. Histochemistry 99, 75–83. doi: 10.1007/BF00268024
Johnson, K. M., Chen, X., Boitano, A., Swenson, L., Opipari, A. W. Jr., and Glick, G. D. (2005). Identification and validation of the mitochondrial F1F0-ATPase as the molecular target of the immunomodulatory benzodiazepine Bz-423. Chem. Biol. 12, 485–496. doi: 10.1016/j.chembiol.2005.02.012
Johnson, K. M., Cleary, J., Fierke, C. A., Opipari, A. W. Jr., and Glick, G. D. (2006). Mechanistic basis for therapeutic targeting of the mitochondrial F1F0-ATPase. ACS Chem. Biol. 1, 304–308. doi: 10.1021/cb600143j
Johnson, K. M., Swenson, L., Opipari, A. W. Jr., Reuter, R., Zarrabi, N., Fierke, C. A., et al. (2009). Mechanistic basis for differential inhibition of the F1Fo-ATPase by aurovertin. Biopolymers 91, 830–840. doi: 10.1002/bip.21262
Junge, W., Sielaff, H., and Engelbrecht, S. (2009). Torque generation and elastic power transmission in the rotary FOF1-ATPase. Nature 459, 364–370. doi: 10.1038/nature08145
Korshunov, S. S., Skulachev, V. P., and Starkov, A. A. (1997). High protonic potential actuates a mechanism of production of reactive oxygen species in mitochondria. FEBS Lett. 416, 15–18. doi: 10.1016/S0014-5793(97)01159-9
Martin, J. L., Ishmukhametov, R., Spetzler, D., Hornung, T., and Frasch, W. D. (2018). Elastic coupling power stroke mechanism of the F1-ATPase molecular motor. Proc. Natl. Acad. Sci. U.S.A. 115, 5750–5755. doi: 10.1073/pnas.1803147115
Muster, B., Kohl, W., Wittig, I., Strecker, V., Joos, F., Haase, W., et al. (2010). Respiratory chain complexes in dynamic mitochondria display a patchy distribution in life cells. PLoS One 5:e11910. doi: 10.1371/journal.pone.0011910
Noji, H., Yasuda, R., Yoshida, M., and Kinosita, K. Jr. (1997). Direct observation of the rotation of F1-ATPase. Nature 386, 299–302. doi: 10.1038/386299a0
Prescott, M., Lourbakos, A., Bateson, M., Boyle, G., Nagley, P., and Devenish, R. J. (1997). A novel fluorescent marker for assembled mitochondria ATP synthase of yeast. OSCP subunit fused to green fluorescent protein is assembled into the complex in vivo. FEBS Lett. 411, 97–101. doi: 10.1016/S0014-5793(97)00670-4
Prescott, M., Nowakowski, S., Gavin, P., Nagley, P., Whisstock, J. C., and Devenish, R. J. (2003). Subunit gamma-green fluorescent protein fusions are functionally incorporated into mitochondrial F1F0-ATP synthase, arguing against a rigid cap structure at the top of F1. J. Biol. Chem. 278, 251–256. doi: 10.1074/jbc.M204556200
Rottele, J., and Zimmermann, H. W. (1993). Transport and accumulation of lipophilic dye cations at the mitochondria of HeLa cells in situ. Cell. Mol. Biol. 39, 739–756.
Schmidt, R., Wurm, C. A., Punge, A., Egner, A., Jakobs, S., and Hell, S. W. (2009). Mitochondrial cristae revealed with focused light. Nano Lett. 9, 2508–2510. doi: 10.1021/nl901398t
Schneider, K., Naujok, A., and Zimmermann, H. W. (1994). Influence of trans-membrane potential and of hydrophobic interactions on dye accumulation in mitochondria of living cells. Photoaffinity labelling of mitochondrial proteins, action of potential dissipating drugs, and competitive staining. Histochemistry 101, 455–461. doi: 10.1007/BF00269496
Schneider, K., and Zimmermann, H. W. (1994). Photoaffinity labelling with fluorescence detection. Dye accumulation at four mitochondrial proteins in HeLa and LM cells. Histochemistry 101, 135–142. doi: 10.1007/BF00269360
Sekiya, M., Sakamoto, Y., Futai, M., and Nakanishi-Matsui, M. (2017). Role of alpha/beta interface in F1 ATPase rotational catalysis probed by inhibitors and mutations. Int. J. Biol. Macromol. 99, 615–621. doi: 10.1016/j.ijbiomac.2017.02.089
Shaner, N. C., Lambert, G. G., Chammas, A., Ni, Y., Cranfill, P. J., Baird, M. A., et al. (2013). A bright monomeric green fluorescent protein derived from Branchiostoma lanceolatum. Nat. Methods 10, 407–409. doi: 10.1038/nmeth.2413
Shao, L., Kner, P., Rego, E. H., and Gustafsson, M. G. (2011). Super-resolution 3D microscopy of live whole cells using structured illumination. Nat. Methods 8, 1044–1046. doi: 10.1038/nmeth.1734
Sielaff, H., and Borsch, M. (2013). Twisting and subunit rotation in single FOF1-ATP synthase. Philos. Trans. R. Soc. Lond. B Biol. Sci. 368:20120024. doi: 10.1098/rstb.2012.0024
Srivastava, A. P., Luo, M., Zhou, W., Symersky, J., Bai, D., Chambers, M. G., et al. (2018). High-resolution cryo-EM analysis of the yeast ATP synthase in a lipid membrane. Science 360:aas9699. doi: 10.1126/science.aas9699
Starke, I., Johnson, K., Petersen, J., Graber, P., Opipari, A. W., Glick, G. D., et al. (2016). Binding of the immunomodulatory drug Bz-423 to mitochondrial FoF1-ATP synthase in living cells by FRET acceptor photobleaching. Proc. SPIE 9712:97120. doi: 10.1117/12.2209645
Stelzer, A. C., Frazee, R. W., Van Huis, C., Cleary, J., Opipari, A. W. Jr., Glick, G. D., et al. (2010). NMR studies of an immunomodulatory benzodiazepine binding to its molecular target on the mitochondrial F(1)F(0)-ATPase. Biopolymers 93, 85–92. doi: 10.1002/bip.21306
Suzuki, T., Tanaka, K., Wakabayashi, C., Saita, E.-I., and Yoshida, M. (2014). Chemomechanical coupling of human mitochondrial F1-ATPase motor. Nat. Chem. Biol. 10, 930–936. doi: 10.1038/nchembio.1635
Tkachev, V., Goodell, S., Opipari, A. W., Hao, L. Y., Franchi, L., Glick, G. D., et al. (2015). Programmed death-1 controls T cell survival by regulating oxidative metabolism. J. Immunol. 194, 5789–5800. doi: 10.4049/jimmunol.1402180
Wachter, A., Bi, Y., Dunn, S. D., Cain, B. D., Sielaff, H., Wintermann, F., et al. (2011). Two rotary motors in F-ATP synthase are elastically coupled by a flexible rotor and a stiff stator stalk. Proc. Natl. Acad. Sci. U.S.A. 108, 3924–3929. doi: 10.1073/pnas.1011581108
Wahl, D. R., Byersdorfer, C. A., Ferrara, J. L., Opipari, A. W. Jr., and Glick, G. D. (2012). Distinct metabolic programs in activated T cells: opportunities for selective immunomodulation. Immunol. Rev. 249, 104–115. doi: 10.1111/j.1600-065X.2012.01148.x
Yasuda, R., Noji, H., Yoshida, M., Kinosita, K. Jr., and Itoh, H. (2001). Resolution of distinct rotational substeps by submillisecond kinetic analysis of F1-ATPase. Nature 410, 898–904. doi: 10.1038/35073513
Yoshida, M., Muneyuki, E., and Hisabori, T. (2001). ATP synthase - a marvellous rotary engine of the cell. Nat. Rev. Mol. Cell Biol. 2, 669–677. doi: 10.1038/35089509
Zhou, A., Rohou, A., Schep, D. G., Bason, J. V., Montgomery, M. G., and Walker, J. E. (2015). Structure and conformational states of the bovine mitochondrial ATP synthase by cryo-EM. eLife 4:e10180. doi: 10.7554/eLife.10180
Keywords: FoF1-ATP synthase, mitochondria, Bz-423, immunomodulator, drug target, Förster resonance energy transfer FRET, FRET acceptor photobleaching
Citation: Starke I, Glick GD and Börsch M (2018) Visualizing Mitochondrial FoF1-ATP Synthase as the Target of the Immunomodulatory Drug Bz-423. Front. Physiol. 9:803. doi: 10.3389/fphys.2018.00803
Received: 30 April 2018; Accepted: 07 June 2018;
Published: 04 July 2018.
Edited by:
Paolo Bernardi, Università degli Studi di Padova, ItalyReviewed by:
Valentina Giorgio, Istituto di Neuroscienze (IN), ItalyJosé J. Garcia-Trejo, National Autonomous University of Mexico, Mexico
Copyright © 2018 Starke, Glick and Börsch. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Michael Börsch, bWljaGFlbC5ib2Vyc2NoQG1lZC51bmktamVuYS5kZQ==