 Alexandre R. Vieira
Alexandre R. Vieira Merve Bayram3
Merve Bayram3 Frank Lippert
Frank Lippert
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Physiol. , 22 February 2017
Sec. Craniofacial Biology and Dental Research
Volume 8 - 2017 | https://doi.org/10.3389/fphys.2017.00104
This article is part of the Research Topic Tooth Enamel : Frontiers in Mineral Chemistry and Biochemistry, Integrative Cell Biology and Genetics View all 35 articles
We have previously shown that AQP5 and BTF3 genetic variation and expression in whole saliva are associated with caries experience suggesting that these genes may have a functional role in protecting against caries. To further explore these results, we tested ex vivo if variants in these genes are associated with subclinical dental enamel mineral loss. DNA and enamel samples were obtained from 53 individuals. Enamel samples were analyzed for Knoop hardness of sound enamel, integrated mineral loss after subclinical carious lesion creation, and change in integrated mineral loss after remineralization. DNA samples were genotyped for single nucleotide polymorphisms using TaqMan chemistry. Chi-square and Fisher's exact tests were used to compare individuals above and below the mean sound enamel microhardness of the cohort with alpha of 0.05. The A allele of BTF3 rs6862039 appears to be associated with harder enamel at baseline (p = 0.09), enamel more resistant to demineralization (p = 0.01), and enamel that more efficiently regain mineral and remineralize (p = 0.04). Similarly, the G allele of AQP5 marker rs3759129 and A allele of AQP5 marker rs296763 are associated with enamel more resistant to demineralization (p = 0.03 and 0.05, respectively). AQP5 and BTF3 genetic variations influence the initial subclinical stages of caries lesion formation in the subsurface of enamel.
Dental caries is a complex multifactorial disease and historically the biological factors operating within the host have been less explored. Our group has, for the last decade, investigated the possible role of genetic variation in the individual susceptibility to caries. Our results suggest, as expected, that several genes underlying multiple mechanisms (i.e., enamel formation, immune response, saliva composition, and quantity) are associated with dental caries (reviewed in Nibali et al., 2016).
One methodological challenge has been the use of past and current caries experience as a measure of disease. We believe that caries experience indicators do not capture fully the underlying mechanisms modulating the pathogenesis of dental caries in an individual, which may hinder discovery if one is trying to identify factors increasing or decreasing individual susceptibility to disease. With that idea in mind, we started to define the disease based on subclinical enamel loss (Shimizu et al., 2012; Weber et al., 2014; Bayram et al., 2015; Vieira et al., 2015) or presence of periapical pathology related to deep caries lesions in dentin (Menezes-Silva et al., 2012; Dill et al., 2015; Maheshwari et al., 2016). Typically, sound enamel microhardness is a poor indicator for susceptibility to demineralization (Lippert and Lynch, 2014). A myriad of factors are involved in the caries process and focusing only on enamel, structural (e.g., pore size and volume, ratio between interprismatic and prismatic enamel fractions) and compositional differences (e.g., Mg, Na, CO3, F contents) are likely the predetermining factors for caries susceptibility. We studied sound enamel microhardness presently to gain further insight into potential genetic factors predetermining susceptibility to demineralization and to explain variations in sound enamel microhardness. Here we are expanding this work to two loci that we previously showed are associated with caries experience, 5q13.2 and 12q13.12, to verify if we can still detect associations when a different phenotypical definition for dental caries is used.
This study was approved by the Ethics Committee of the Istanbul University, Medical Faculty, Istanbul, Turkey and the University of Pittsburgh Institutional Review Board (IRB# 11070236). Written informed consent was obtained from all participating individuals and parents/legal guardians. Fifty-three orthodontic patients from Istanbul University, Faculty of Dentistry, Department of Orthodontics, participated in this study during the period 5 September 2011 to 30 November 2012. Participants had an indication for extraction of pre-molars for orthodontic reasons and were consecutively invited to participate in the study during the period described above. They agreed to donate their extracted tooth (teeth). One first premolar, extracted for orthodontic reasons, was used from each participant as a source of enamel.
Unstimulated saliva samples were obtained from all participants and stored in Oragene DNA Self-Collection kits (DNA Genotek, ON, Canada) at room temperature until processed. DNA was extracted according to the manufacturer's instructions. Ten single nucleotide polymorphisms (SNPs) were selected, including six in the aquaporin locus 12q13.2 (rs3759129, rs10875989, rs1996315, rs2878771, rs296763, and rs467323) and four located in 5q13.2 (rs27565, rs4700418, rs875459, and rs6862039). These SNPs were chosen based on the results of our previous fine mapping studies on the two loci (Shimizu et al., 2013; Anjomshoaa et al., 2015). Polymerase chain reactions with TaqMan (hydrolysis probes that are designed to increase the specificity of quantitative PCR) SNP Genotyping Assays from Applied Biosystems (Valencia, CA, USA), with a total volume of 3 μl per reaction and 3.0 ng of DNA per reaction, were used for genotyping all selected markers in a Tetrad PTC225 thermocycler from MJ Research (Waltham, MA, USA). Genotype detection and analysis were performed using the ABI 7900HT with ABI SDS software (Applied Biosystems, Valencia, CA, USA).
Fifty-three caries-free premolar teeth (one from each participant), extracted for orthodontic reasons, were studied. Figure 1 summarizes the study design. Caries experience of this cohort was high, with a mean DMFT score of 5.19.
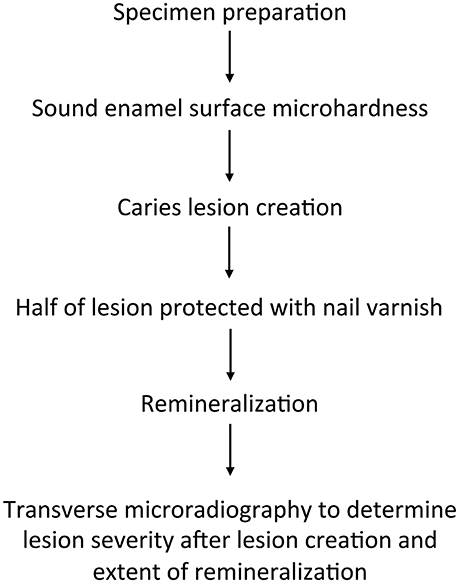
Figure 1. Summary of the study design.
The tissue remnants were cleaned from the teeth and then teeth were stored in 10% buffered formalin (pH 7.0) solution at 4°C until required for initial laboratory manipulation. The crowns were separated from the roots, and then each tooth was cut into 3 × 3-mm specimens using a low-speed saw (Isomet, Buehler, Lake Bluff, IL, USA). The teeth were stored in thymol during the sample preparation process. The specimens were embedded individually in acrylic resin (Varidur, Buehler) and polished to create flat surfaces to facilitate surface microhardness testing using Struers Rotopol 31/Rotoforce 4 polishing unit (Struers Inc., Cleveland, PA, USA). Specimens were ground flat and polished with water-cooled abrasive disks (500-, 1,200-, 2,400-, and 4,000-grit SiC papers; MDFuga, Struers Inc., Cleveland, Ohio, USA) and polishing cloth with diamond suspension (1 μm; Struers Inc.). After the polishing procedures, specimens were sonicated in neutral detergent solution and rinsed with deionized water. As a final cleaning step, the specimens were sonicated in a detergent solution (Micro-90 concentrated cleaning solution with 2 % dilution) for 3 min. The specimens were finally assessed under Nikon SMZ 1500 stereomicroscope at × 10 magnification. Accepted specimens had no obvious cracks, areas of hypomineralization, or other flaws in the enamel surface.
Initial hardness of each specimen was determined using a Knoop microhardness indenter (2100 HT; Wilson Instruments, Norwood, MA, USA) at a load of 50 g for 15 s. The average specimen surface microhardness was determined from five indentations placed in the center of the surface of each specimen, ~100 μm apart from one another.
Early carious lesions were created in the specimens utilizing a demineralization protocol based on that by White (1987), which has been extensively studied using a variety of techniques over the years (White, 1987; Churchley et al., 2011). Artificial lesions were formed in the enamel specimens of each disk by a 5-day immersion into a solution containing 0.1 M lactic acid, 4.1 mM CaCl2 × 2 H2O, 8.0 mM KH2PO4, and 0.2% w/v Carbopol 907 (BF Goodrich Co., USA), pH adjusted to 5.0 using KOH, for 5 days at 37°C. Demineralization was performed at a ratio of 10 ml of solution per specimen. The resulting lesions were early, shallow, subsurface1 lesions with a typical, average depth of approximately 50 μm. After lesion creation, approx. half of the lesion surface area was covered with colored, acid-resistant nail varnish to preserve a baseline lesion for future analysis.
All lesions were then remineralized for 4 days at 37°C using “resting plaque fluid” (Lynch et al., 2007) with the following composition: 10 mM acetic acid, 1.0 mM CaCl2 × 2 H2O, 12.7 mM KH2PO4, 130 mM KCl, 20 mM HEPES, 0.1 ppm F (NaF), pH adjusted to 6.5 using KOH.
Sections, approximately 100 μm in thickness and two per specimen, were cut from the center of the specimens across the varnish-covered lesion area and remineralized lesion window using a Silverstone-Taylor Hard Tissue Microtome (Scientific Fabrications Laboratories, USA). The sections were mounted, with an aluminum step wedge, on high resolution glass plate Type I A (Microchrome Technology Inc., San Jose, CA) and X-rayed at 20 kV and 30 mA at a distance of 42 cm for 65 min. The film was developed in Kodak D-19 developer for 3 min, placed in a stop bath (Kodak 146-4247) for 45 s, and then fixed (Kodak 146-4106) for 3 min. All plates were then rinsed in deionized water for 15 min and air-dried. Microradiographs were examined with a Zeiss EOM microscope in conjunction with the TMR software v.3.0.0.11. Sound enamel was assumed to be 87% v/v mineral. Integrated mineral loss (ΔZ) was recorded for both the varnish-covered lesion baseline area and the remineralized lesion. The difference was calculated (pre–post data) to assess the extent of remineralization.
Based on sound enamel hardness values, subjects were classified into dichotomous groups (baseline values or rate changes above or below the average of the group). Subjects were classified as having “softer enamel” (below the average of the group) and “harder enamel” (above the average of the group) for determination of hardness phenotypes. Chi-square and Fisher's exact tests were used to assess association between the SNPs and hardness values by the use of the PLINK software package (Purcell et al., 2007) with an established alpha of 0.05.
Table 1 shows the mean values of hardness and Table 2 summarizes all genotyping frequencies and comparisons were made between individuals that showed levels of enamel loss above or below the mean loss of the studied sample. The A allele of BTF3 rs6862039 appears to be associated with harder enamel at baseline (p = 0.09), enamel more resistant to demineralization (p = 0.01), and enamel that more efficiently regain mineral and remineralize (p = 0.04). Similarly, the G allele of AQP5 marker rs3759129 and A allele of AQP5 marker rs296763 are associated with enamel more resistant to demineralization (p = 0.03 and 0.05, respectively). No other markers studied showed statistical evidence of association.

Table 1. Mean hardness of studied specimens.
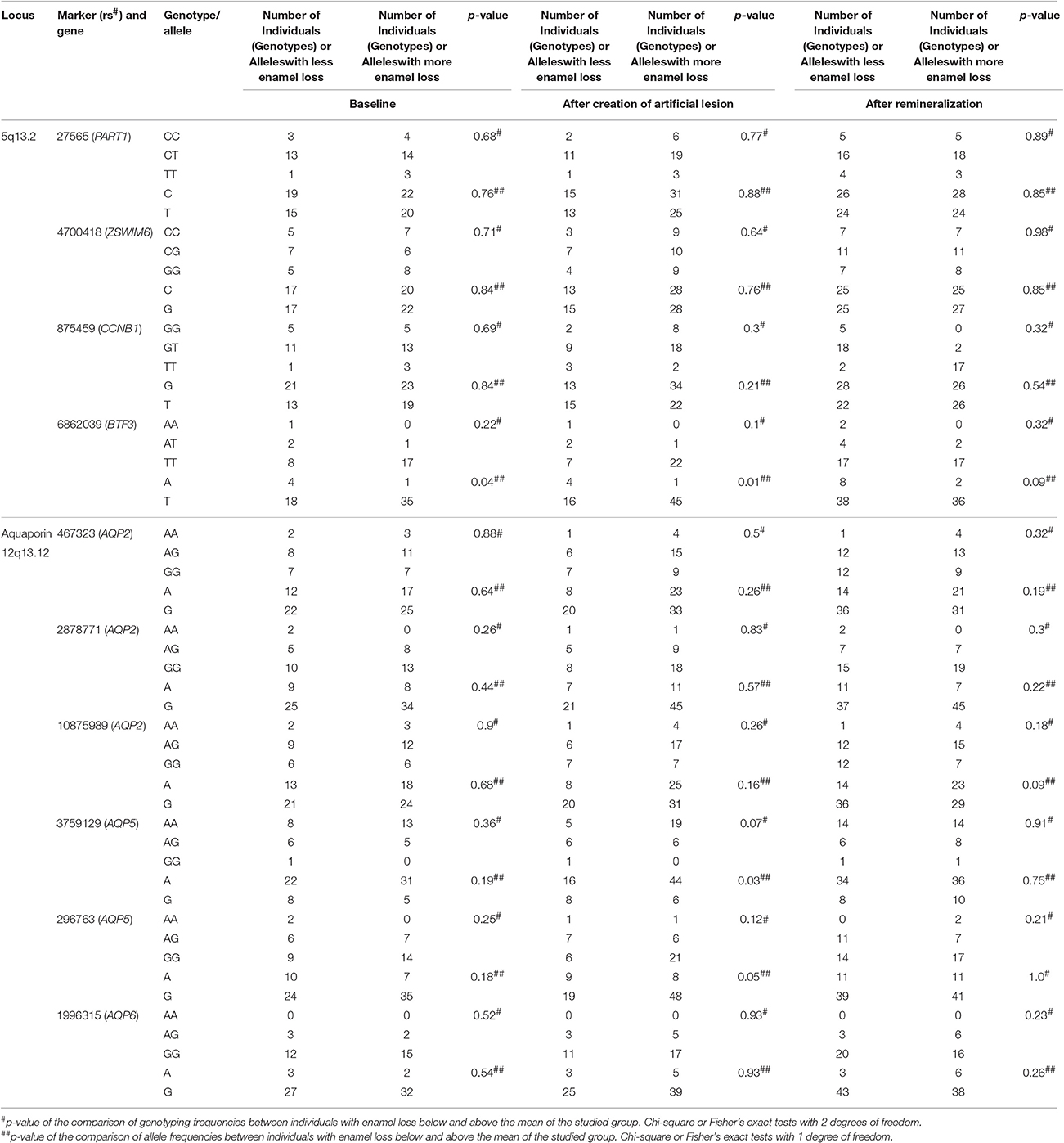
Table 2. Genotyping frequencies and summary results of the association studies.
Our data confirm the association between dental caries and genetic variation in BTF3 and AQP5 we have previously reported (Shimizu et al., 2013; Anjomshoaa et al., 2015). When we reported the association between caries experience and BTF3 (Shimizu et al., 2013), we tested if BTF3 was expressed in whole saliva and if expression that correlated with caries experience. We suggested based on our initial findings that BTF3 may be involved in caries susceptibility acting through saliva. This gene encodes the basic transcription factor 3, a protein that forms a stable complex with RNA polymerase IIB and is required for transcriptional initiation. Alternative splicing results in multiple transcript variants encoding different isoforms. BTF3 has multiple pseudogenes (https://www.ncbi.nlm.nih.gov/gene, Gene ID = 689). In gastric cancer, BTF3 expression is associated with enhanced cell proliferation, reduced cell cycle regulation, and apoptosis and its silencing inhibits proliferation of gastric cancer cells (Liu et al., 2013). Our data may suggest that BTF3 may be involved in the formation of the enamel, possibly acting as a transcription factor controlling cell proliferation.
Aquaporin 5 (AQP5) is a water channel protein expressed in salivary and lacrimal glands, various types of epithelial cells, and during tooth development (Ishida et al., 1997; Nielsen et al., 1997; Funaki et al., 1998; Hamann et al., 1998; Felszeghy et al., 2004). AQP5 interactions during dental development may impact the formation of dental enamel and susceptibility to dental caries (Anjomshoaa et al., 2015). Since the AQP locus was not associated with enamel hardness, we thought the role of AQP5 in caries was through salivation rather than influencing enamel development. We decided to reassess our microhardness experiments and this time we tested the specimens, instead directly at the treated surface, in the subsurface. These analyses suggested that genetic variants in AQP5 are associated with initial enamel loss, which is a surrogate for the development of early caries lesions. These data support the idea that AQP5 impacts enamel development possibly making it more susceptible to caries.
While concerned about multiple testing, we avoided to apply the strict Bonferroni correction and increase type II error. If we had used Bonferroni correction, we would have lowered the alpha to 0.005 (0.05/10). We have demonstrated previously (Vieira et al., 2008) that known true associations are missed when correction for multiple testing is implemented. The results of our work should be considered with caution and serve to generate a hypothesis to be directly tested in larger and more homogeneous samples. On the other hand, simply disregarding the nominal associations presented here may delay discovery by misleading the field to believe that no true biological relationships exist. Another limitation of our study is that our phenotype reflects subclinical caries lesions that cannot be detected by the naked eye (Shimizu et al., 2012). The SNPs we report here as associated with caries do not have known functional roles. Our work continues to support that individuals susceptibility is a factors in dental caries susceptibility, with some individuals more susceptible to mineral losses when pH lowers.
ARV designed the study, obtained support, analyzed and interpreted data, and wrote the first draft of the manuscript. MB and FS (Istanbul University) helped design the study facilitated DNA and sample collections, Kathleen Deeley (University of Pittsburgh) managed samples and generated genotypes, FL (Indiana University) helped design the study and generated enamel microhardness measurements, and RS (University of Pittsburgh) statistically analyzed the data. AM helped design the study and obtained support. All authors critical revised the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer CC and handling Editor declared their shared affiliation, and the handling Editor states that the process nevertheless met the standards of a fair and objective review.
Part of the data of this manuscript was presented during the Enamel 9 Conference, between October 30th and November 3rd 2016. The Enamel 9 Conference was supported in part by the NIH/NIDCR grant R13-DE026647, awarded to ARV. ARV's attendance to the conference was supported by Colgate Palmolive.
1. ^The chosen demineralization procedure will result in the formation of early caries lesions. The most prominent feature of these lesions is that demineralization is primarily confined to enamel mineral below the surface. Hence, the term “subsurface” demineralization has been introduced into the literature to differentiate to surface loss such as that occurring during dental erosion.
Anjomshoaa, I., Briseño-Ruiz, J., Deeley, K., Poletta, F. A., Mereb, J. C., Leite, A. L., et al. (2015). Aquaporin 5 interacts with fluoride and possibly protects against caries. PLoS ONE 10:e0143068. doi: 10.1371/journal.pone.0143068
Bayram, M., Deeley, K., Reis, M. F., Trombetta, V. M., Ruff, T. D., Sencak, R. C., et al. (2015). Genetic influences on dental enamel that impact caries differ between the primary and permanent dentitions. Eur. J. Oral Sci. 123, 327–334. doi: 10.1111/eos.12204
Churchley, D., Lynch, R. J., Lippert, F., Eder, J. S., Alton, J., and Gonzalez-Cabezas, C. (2011). Terahertz pulsed imaging study to assess remineralization of artificial caries lesions. J. Biomed. Opt. 16, 026001. doi: 10.1117/1.3540277
Dill, A., Letra, A., Chaves de Souza, L., Yadlapati, M., Bigueti, C. C., Garlet, G. P., et al. (2015). Analysis of multiple cytokine polymorphisms in individuals with untreated deep carious lesions reveals IL1B (rs1143643) as a susceptibility factor for periapical lesion development. J. Endod. 41, 197–200. doi: 10.1016/j.joen.2014.10.016
Felszeghy, S., Módis, L., Németh, P., Hagy, G., Zelles, T., Agre, P., et al. (2004). Expression of aquaporin isoforms during human and mouse tooth development. Arch. Oral Biol. 49, 247–257. doi: 10.1016/j.archoralbio.2003.09.011
Funaki, H., Yamamoto, T., Koyama, Y., Kondo, D., Yaoita, E., Kawasaki, K., et al. (1998). Localization and expression of AQP5 in cornea, serous salivary glands, and pulmonary epithelial cells. Am. J. Physiol. 275, C1151–C1157.
Hamann, S., Zeuthen, T., LaCour, M., Nagelhus, E. A., Ottersen, O. P., Agre, P., et al. (1998). Aquaporins in complex issues: distribution of aquaporins 1–5 in human and rat eye. Am. J. Physiol. 274, C1332–C1345.
Ishida, N., Hirai, S. I., and Mita, S. (1997). Immunolocalization of aquaporin homologs in mouse lacrimal glands. Biochem. Biophys. Res. Commun. 238, 891–895. doi: 10.1006/bbrc.1997.7396
Lippert, F., and Lynch, R. J. (2014). Comparison of knop and Vickers surface microhardness and transverse microradiography for the study of early caries lesion formation in human and bovine enamel. Arch. Oral Biol. 59, 704–710. doi: 10.1016/j.archoralbio.2014.04.005
Liu, Q., Zhou, J. P., Li, B., Huang, Z. C., Dong, H. Y., Li, G. Y., et al. (2013). Basic transcription factor 3 is involved in gastric cancer development and progression. World J. Gastroenterol. 19, 4495–4503. doi: 10.3748/wjg.v19.i28.4495
Lynch, R. J., Mony, U., and ten Cate, J. M. (2007). Effect of lesion characteristics and mineralizing solution type on enamel remineralization in vitro. Caries Res. 41, 257–262. doi: 10.1159/000101914
Maheshwari, K., Silva, R. M., Guajardo-Morales, L., Garlet, G. P., Vieira, A. R., and Letra, A. (2016). Heat shock 70 protein genes and genetic susceptibility to apical periodontitis. J. Endod. 42, 1467–1471. doi: 10.1016/j.joen.2016.07.010
Menezes-Silva, R., Khaliq, S., Deeley, K., Letra, A., and Vieira, A. R. (2012). Genetic susceptibility to periapical disease: conditional contribution of MMP2 and MMP3 genes to the development of periapical lesions and healing response. J. Endod. 38, 604–607. doi: 10.1016/j.joen.2012.02.009
Nibali, L., Di Iorio, A., Tu, Y. K., and Vieira, A. R. (2016). Host genetics role in the pathogenesis of periodontal disease and caries. J. Clin. Periodontol. doi: 10.1111/jcpe.12639. [Epub ahead of print].
Nielsen, S., King, L. S., Christensen, B. M., and Agre, P. (1997). Aquaporins in complex tissues. II. Subcellular distribution in respiratory and glandular tissues of rat. Am. J. Physiol. 273, C1549–C1561.
Purcell, S., Neale, B., Todd-Brown, K., Thomas, L., Ferreira, M. A., Bender, D., et al. (2007). PLINK: a toolset for whole-genome association and population-based linkage analysis. Am. J. Hum. Genet. 81, 559–575. doi: 10.1086/519795
Shimizu, T., Deeley, K., Briseño-Ruiz, J., Faraco, I. M. Jr., Poletta, F. A., Brancher, J. A., et al. (2013). Fine-mapping of 5q12.1-13.3 unveils new genetic contributors to caries. Caries Res. 47, 273–283. doi: 10.1159/000346278
Shimizu, T., Ho, B., Deeley, K., Briseño-Ruiz, J., Faraco, I. M. Jr., Schupack, B. I., et al. (2012). Enamel formation genes influence enamel microhardness before and after cariogenic challenge. PLoS ONE 7:e45022. doi: 10.1371/journal.pone.0045022
Vieira, A. R., Gibson, C. W., Deeley, K., Xue, H., and Li, Y. (2015). Weaker enamel explains dental decay. PLoS ONE 10:e0124236. doi: 10.1371/journal.pone.0124236
Vieira, A. R., McHenry, T. B., Daack- Hirsch, S., Murray, J. C., and Marazita, M. L. (2008). Candidate gene/loci studies in cleft lip/palate and dental anomalies finds novel susceptibility genes for clefts. Genet. Med. 10, 668–674. doi: 10.1097/GIM.0b013e3181833793
Weber, M. L., Hsin, H. Y., Kalay, E., Brožková, D. S., Shimizu, T., Bayram, M., et al. (2014). Role of estrogen related receptor beta (ESRRB) in DFN35B hearing impairment and dental decay. BMC Med. Genet. 15:81. doi: 10.1186/1471-2350-15-81
Keywords: dental caries, dental enamel, dental erosion, transcription factors, aquaporins
Citation: Vieira AR, Bayram M, Seymen F, Sencak RC, Lippert F and Modesto A (2017) In Vitro Acid-Mediated Initial Dental Enamel Loss Is Associated with Genetic Variants Previously Linked to Caries Experience. Front. Physiol. 8:104. doi: 10.3389/fphys.2017.00104
Received: 20 December 2016; Accepted: 08 February 2017;
Published: 22 February 2017.
Edited by:
Thimios Mitsiadis, University of Zurich, SwitzerlandReviewed by:
Claudio Cantù, University of Zurich, SwitzerlandCopyright © 2017 Vieira, Bayram, Seymen, Sencak, Lippert and Modesto. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alexandre R. Vieira, YXJ2MTFAcGl0dC5lZHU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.