 Feng He
Feng He Juan Li2†
Juan Li2† Chia-Chen Chuang
Chia-Chen Chuang Li Zuo
Li Zuo
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Physiol. , 07 November 2016
Sec. Exercise Physiology
Volume 7 - 2016 | https://doi.org/10.3389/fphys.2016.00486
It is well known that regular exercise can benefit health by enhancing antioxidant defenses in the body. However, unaccustomed and/or exhaustive exercise can generate excessive reactive oxygen species (ROS), leading to oxidative stress-related tissue damages and impaired muscle contractility. ROS are produced in both aerobic and anaerobic exercise. Mitochondria, NADPH oxidases and xanthine oxidases have all been identified as potential contributors to ROS production, yet the exact redox mechanisms underlying exercise-induced oxidative stress remain elusive. Interestingly, moderate exposure to ROS is necessary to induce body's adaptive responses such as the activation of antioxidant defense mechanisms. Dietary antioxidant manipulation can also reduce ROS levels and muscle fatigue, as well as enhance exercise recovery. To elucidate the complex role of ROS in exercise, this review updates on new findings of ROS origins within skeletal muscles associated with various types of exercises such as endurance, sprint and mountain climbing. In addition, we will examine the corresponding antioxidant defense systems as well as dietary manipulation against damages caused by ROS.
Regular exercise is beneficial to our health. However, unaccustomed or exhaustive exercise can result in detrimental health effects such as muscle damage, inflammation and oxidative stress. Specifically, repetitive muscle contraction involves accumulation of reactive oxygen species (ROS) (Zuo et al., 2011a, 2014, 2015b). These oxygen-derived free radicals or reactive derivatives, including superoxide (), hydroxyl radical (•OH), and hydrogen peroxide (H2O2), have been implicated in various diseases and physiological conditions (Alfadda and Sallam, 2012). Acting as signaling molecules, a physiological level of ROS is essential for normal cellular functions. For instance, exogenous antioxidant supplements have been shown to suppress muscle contractility while the addition of H2O2 relieves such an effect, suggesting that oxidants (at low levels) may be imperative in facilitating muscle contraction (Reid et al., 1993; Powers and Jackson, 2008). However, the overproduction of ROS induced by exhaustive exercise training or other stresses, along with compromised antioxidant defenses, can lead to oxidative stress and related tissue damage (Powers et al., 2011b; Zuo et al., 2012). Interestingly, proper exercise (moderate to high intensity exercise) stimulates the adaptive responses and strengthens the endogenous antioxidant defense systems to combat excessive ROS thereby maintaining muscle redox balance (Parker et al., 2014; Zuo et al., 2015b).
Several techniques have been reported to examine oxidative stress in muscle tissues of both human and animal models (Powers and Jackson, 2008; Cheng et al., 2016). It is worth noting that the direct and quantitative measurement of ROS production continues to remain challenging in muscle redox biology due to the reactive nature of ROS as well as the methodological shortcomings. Commonly used indicators of ROS alteration in intact muscle fibers, such as fluorescent probes and spin traps, have limited specificity to the types of ROS (Powers and Jackson, 2008; Cheng et al., 2016). It is also difficult to assess the subtle changes in ROS levels during repeated muscle contractions directly using fluorescence (Cheng et al., 2016). Other indirect evaluation of oxidative stress includes the measurement of antioxidants, reduced/oxidized glutathione (GSH/GSSH) ratio, and oxidative modified molecules such as malondialdehyde for lipid peroxidation and 8-hydroxy-2′-deoxyguanosine for DNA oxidation (Powers and Jackson, 2008; Cakir-Atabek et al., 2010). These approaches are likely subject to experimental artifacts (Powers and Jackson, 2008). Along with limitations of these techniques on the accuracy of ROS measurement, the variation in specific ROS sources and oxidative modifications in different exercise protocols further contribute to the inconsistency and difficulty seen in this type of study.
Currently, the exact redox mechanisms underlying exercise-induced oxidative stress and exercise-induced adaptation remain unclear. Exploring ROS pathways may advance our understanding of muscle fatigue and recovery in exercise, as well as the development of potential tools for ROS assessment in exercising muscles. Although, mounting evidence has shown an elevation of oxidative stress associated with exercise, there is a lack of systemic review on how the activities of exercise (i.e., exercise type, intensity, and duration) affect ROS production. Therefore, this review aims to provide a timely update on the sources of ROS in different types of exercise, as well as the paradoxical role of ROS in acute and chronic exercise.
Muscle activity has been shown to associate with ROS production, yet the extents and sources of ROS differ based on types of exercise (Steinbacher and Eckl, 2015). There is a general consensus that ROS are generated predominantly by contracting skeletal muscles during physical activity. Indeed, moderate levels of ROS are necessary for the production of normal muscle force; however, excess ROS can lead to muscle fatigue and contractile dysfunction (Powers et al., 2011a). Major endogenous sources of ROS in skeletal muscle include mitochondria, NADPH oxidase (NOX), and xanthine oxidase (XO) (Steinbacher and Eckl, 2015). Under physiological conditions, ROS are released as byproducts of cellular respiration by mitochondria. Accordingly, mitochondria-derived can be observed in both resting and exercising muscle (Sakellariou et al., 2014; Zuo et al., 2015b). Mitochondrial respiration is in state 4 (basal) at rest, and enters active state 3 when muscle contraction begins, which is characterized by an increase in mitochondrial ADP levels due to rapid breakdown of ATP. Interestingly, the rate of production is normally higher at basal mitochondrial respiration (state 4) than state 3 in both skeletal muscle and the diaphragm, suggesting that mitochondria might not be the major source of ROS in exercising muscles (Powers and Jackson, 2008; Kavazis et al., 2009; Sakellariou et al., 2014). On the other hand, NOX is a key ROS generator during muscle contractions, contributing to a larger extent of cytosolic than mitochondria (Powers et al., 2011a; Steinbacher and Eckl, 2015). NOX is a multi-component enzyme located on the plasma membrane of phagocytic cells and several subcellular sites of skeletal muscle fibers (e.g., T-tubules and sarcolemma) (Michaelson et al., 2010; Zuo et al., 2011b; Sakellariou et al., 2013, 2014). NOX-induced ROS in the T-tubules can directly activate ryanodine receptor type 1 to enhance calcium (Ca2+) release and muscle contractions during exercise (Espinosa et al., 2006; Hidalgo et al., 2006). Other factors, such as phospholipase A2(PLA2), have been shown to stimulate NOX to produce ROS. PLA2 also facilitates phospholipid turnover and releases arachidonic acid (a substrate for lipoxygenases), leading to further ROS formation and lipid peroxidation related damage (Zuo et al., 2004; Steinbacher and Eckl, 2015). Found in the endothelium and cytosol of muscle, XO contributes to the production of extracellular during isometric contraction. This XO-derived plays a critical role in the muscle force generation (Powers and Jackson, 2008; Gomez-Cabrera et al., 2010). Moreover, the auto-oxidation of myoglobin or the oxidation of hemoglobin to methemoglobin further contributes to oxidative stress in the muscle by inducing peroxide formation (Marciniak et al., 2009).
In addition to endogenous sources of ROS described above, strenuous exercise-induced muscle injuries involve oxidative burst from immune cells, leading to a rapid ROS formation and subsequent oxidative damage (Steinbacher and Eckl, 2015). Particularly, untrained individuals are more prone to the detrimental effects exerted by the enhanced oxidative stress, while the trained subjects normally experience lessened effects due to increased oxidative tolerance (Steinbacher and Eckl, 2015). Aging or pathophysiological states of muscle are also associated with ROS elevation and contractile dysfunction (Steinbacher and Eckl, 2015). For example, greater endogenous oxidant generation has been observed in the isolated skeletal muscle fiber of old mice compared to young mice at rest (Palomero et al., 2013; Vasilaki and Jackson, 2013). It is suggested that such changes in ROS levels can be attributed to chronic inactivity of the muscle, which provides a possible explanation for the age-related ROS overproduction in muscle (Talbert et al., 2013; Vasilaki and Jackson, 2013). In addition, under disease states such as muscle dystrophy, simple stretch contractions can lead to significant muscle damage that is associated with ROS generation, through both increased NOX activation and cytosolic Ca2+ levels (Whitehead et al., 2010).
In skeletal muscle, both enzymatic (e.g., glutathione peroxidase (GPx) and catalase) and non-enzymatic (e.g., GSH, uric acid, bilirubin, vitamin E, vitamin C, etc.) antioxidants function as a unified complex to scavenge ROS (Powers and Jackson, 2008). These intracellular antioxidants are normally located within cells, cytoplasm, and organelles (e.g., mitochondria) to protect muscle fibers from ROS-induced damage (Powers and Jackson, 2008; Powers et al., 2011a). However, excessive ROS formation can offset these protective mechanisms during intense and exhaustive exercise. In general, the intensity of aerobic exercise is represented by maximal oxygen uptake (%VO2max) and the intensity of anaerobic exercise is described by repetition maximum (% RM). The extents and sources of ROS production can be influenced by the intensity, type, and duration of exercise, in which details will be discussed in latter paragraphs.
Strenuous aerobic or endurance exercise is commonly known to induce ROS and reactive nitrogen species overproduction due to enhanced metabolism, leading to oxidative stress and related injuries (Powers and Jackson, 2008; Neubauer et al., 2010; Gomes et al., 2012). It has been estimated that aerobic exercise results in a 1–3-folds increase of during muscle contraction (Sakellariou et al., 2014; Figure 1). However, mitochondria only account for a small portion of generation during aerobic exercise (Sakellariou et al., 2014; Zuo et al., 2015b). In fact, mitochondria-derived formation in skeletal muscle is decreased during exercise compared to that at rest. This is because contractile activities alter the redox status in muscles toward a more oxidative state, leading to a lowered mitochondrial NADH/NAD+ ratio. The decline in NADH/NAD+ ratio is linked with reduced complex I-dependent release (Sakellariou et al., 2014). During endurance exercise, ATP is broken down to release energy and support continuous muscle contraction. In some instances, AMP is formed which can be further degraded to hypoxanthine, xanthine, and uric acid through a biochemical process involving XO. As described previously, XO induces formation by utilizing molecular oxygen, thereby exacerbating oxidative stress (Mastaloudis et al., 2001). Elevated lipid peroxidation and DNA oxidative damage have been observed following a single bout of intensive exercise. Such acute inflammatory and oxidative responses can be induced by vigorous aerobic exercise, which resemble the stress responses following ischemic stroke and myocardial infarction (Mastaloudis et al., 2004). In addition, oxidative burst induced by leukocytes is an effective mechanism for fighting against microbes during infection (Saran et al., 1999; Agarwal et al., 2003). The long-lasting endurance exercise may compromise the ROS-generation capability of leukocytes, resulting in an increased susceptibility to infectious diseases in athletes (Nielsen et al., 2004). Moreover, for exercising people with diseases such as asthma, special cautions must be taken since asthma may cause substantial ROS formation and oxidative stress thus compromising exercise-induced benefits (Jiang et al., 2014).
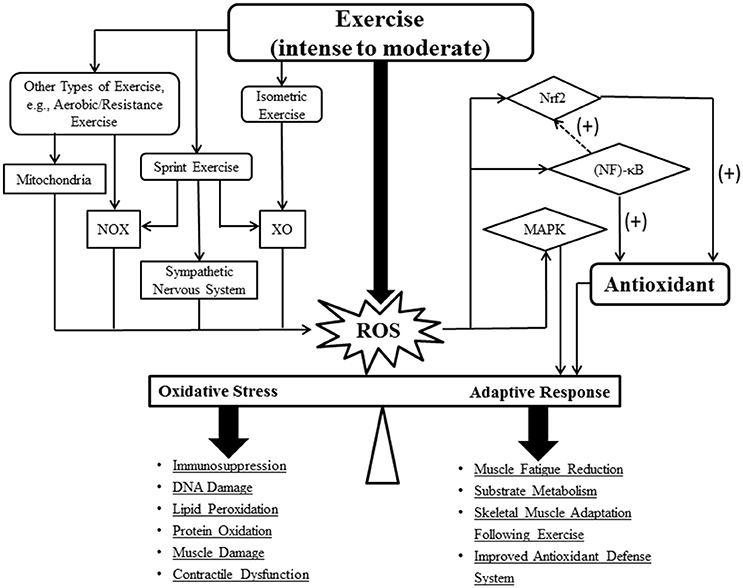
Figure 1. Schematic illustrating ROS generation during different types of exercise and their associated roles in adaptive response. The dash arrow represents an indirect effect. Abbreviations: reactive oxygen species (ROS); NADPH oxidase (NOX); xanthine oxidase (XO); mitogen-activated protein kinase (MAPK); nuclear erythroid 2 p45-related factor 2 (Nrf2); nuclear factor κB (NF-κB).
Although, single bouts of intensive aerobic exercise may cause potential oxidative damage to muscle fibers, regular aerobic exercise will help enhance the cellular ability to detoxify ROS over-accumulation (Radak et al., 2013). Regular/moderate exercise has been shown to enhance antioxidant defense by incrementing the activity of endogenous antioxidant enzymes such as superoxide dismutase (SOD), GPx, and catalase (Miyata et al., 2008). Exercise protects the body against constant mild or moderate ROS exposure through redox-associated preconditioning including oxidative damage repair systems (Radak et al., 2013). This moderate exercise-mediated adaptation also involves increased myocellular antioxidant capacity which helps to lower ROS levels (Mastaloudis et al., 2001; Knez et al., 2006). Moreover, increased ROS formation in active skeletal muscles plays a critical role in exercise adaptation by modulating muscle contraction (Mastaloudis et al., 2004; Radak et al., 2013). For example, endurance running is regarded as important for survival in human evolution since it can trigger exercise-associated adaptive responses through metabolic and redox challenges (Radak et al., 2013; Ferraro et al., 2014; Wiggs, 2015). However, contemporary lifestyles decrease physical activities and suppress human adaptive capacity of metabolism and redox homeostasis (Radak et al., 2013). Substantial evidence has suggested that at least 30 min of accumulated physical activity (moderate-intensity) each day is necessary to maintain good health and reduce potential disease risks (Knez et al., 2006). Accordingly, Berzosa et al. and Georgakouli et al. both observed a significant elevation of plasma total antioxidant capacity in healthy individuals after a 30 min of submaximal exercise (70% of maximum workload and 50–60% of the heart rate reserve, respectively) on cycle ergometer (Berzosa et al., 2011; Georgakouli et al., 2015).
Although the main source of ROS during aerobic exercise has been thoroughly reviewed in a previous study (Powers and Jackson, 2008), little is known regarding the potential source of ROS during short-term intensive (anaerobic) exercise such as sprints. The redox mechanisms of anaerobic work have been investigated in a variety of exercise models including sprinting trainings as well as isometric and eccentric exercises (Nikolaidis et al., 2007, 2008; Stagos et al., 2015).
Unlike other exercises, sprints predominantly rely on anaerobic energy pathways due to its high energy demand. While sprinting, a small portion (0.15%) of is produced in the mitochondria (St-Pierre et al., 2002). This lower than usual ROS production in skeletal muscle mitochondria can be attributed to relatively low amounts of oxygen consumption and increased ADP (state 3) during sprints (Herrero and Barja, 1997; Morales-Alamo and Calbet, 2014). NOX is one of the potential sites of production associated with intense muscle contractions (Sakellariou et al., 2013; Figure 1). Additionally, XO activation triggered by an elevation in hypoxanthine during and following sprints, is regarded as another important contributor for ROS production (Kang et al., 2009; Figure 1). Intensive exercise accelerates ATP degradation, leading to elevated formation of AMP, hypoxanthine, xanthine, and uric acid. Particularly, the increased levels of xanthine facilitate ROS generation by XO, thereby exacerbating oxidative stress in anaerobic exercise (Mastaloudis et al., 2001; Radak et al., 2013). In response to intense exercise, the active sympathetic nervous system can also play a role in ROS formation (Figure 1). Accordingly, Bors et al. demonstrated that adrenaline administration largely increased H2O2 levels in vitro (Bors et al., 1978).
In static positions, isometric exercise is common in daily activities such as holding weighted objects. A variety of oxidative stress biomarkers have been examined in response to isometric exercise; yet mixed results can be produced. For instance, isometric contractions result in increased levels of hydroperoxide and elevations in blood protein carbonyls. However, there is no change in plasma malondialdehyde (a useful indicator of lipid peroxidation) (Rodriguez et al., 2003; Urso and Clarkson, 2003). Moreover, repetitive static exercise (RSE) can be considered as a similar condition to partial ischemia/reperfusion, which may protect the tissues against oxidative stress (Zuo et al., 2013). However, Sahlini et al. observed no signs of ROS elevation during prolonged RSE despite a manifestation of decreased mechanical efficiency and force generation (Sahlin et al., 1992). Furthermore, isometric exercise was reported to induce an increase in the GSSH/GSH ratio, but intense isometric contraction can lead to lactic acidosis and stimulate the conversion of to highly reactive •OH (Waterfall et al., 1996; Groussard et al., 2000; Garatachea et al., 2012).
A handful studies have also assessed the oxidative stress resulting from eccentric exercise (Nikolaidis et al., 2007, 2008), a physical activity that can induce sarcolemma inflammation and subsequent ROS overproduction and muscular damage (Nikolaidis et al., 2007, 2008). One study reported that ROS formation peaked after the large muscle function decline in downhill running (Close et al., 2004). Other study showed that eccentric contraction likely causes secondary muscle damage due to ROS–induced inflammation (Nikolaidis et al., 2007; Silva et al., 2010).
A good example for exploring the influence of ROS on physical activity is mountain climbing. Mountain climbing involves the exposure to extreme environmental conditions caused by high altitudes, stimulating ROS generation in the body (Miller et al., 2013). Mountaineers generally experience various undesirable conditions at altitudes of 2 km or above (Basnyat, 2001; Hackett and Roach, 2001; Basnyat et al., 2003). For example, long-term exposures to an altitude above 4 km could induce a loss of appetite, leading to nutrition deficiency and weight loss (Siesjö et al., 1996; Wasse et al., 2012). Collectively, these symptoms associated with acute mountain sickness are related to harsh environmental factors such as low oxygen, cold, and ultraviolet rays (Askew, 2002; Smedley and Grocott, 2013). Particularly, hypobaric hypoxia generates a large amount of ROS, resulting in the subsequent tissue injuries in mountaineers (Askew, 2002; Julian et al., 2014).
As altitude increases, lower atmospheric pressures lead to reduced atmospheric oxygen partial pressures and arterial blood oxygen levels, causing hypoxic damage (Askew, 2002; Vallecilla et al., 2014). Under normal circumstances, people are able to resist mild oxidative stress and restore redox balance via the body's naturally equipped antioxidant system. However, overwhelmed antioxidant defenses due to severe oxidative stress (e.g., inappropriate exercise exertion) can promote cell damage or death (Bakonyi and Radak, 2004; Zuo et al., 2015b). Oxidative stress induced by hypoxia at high altitudes results in intracellular Ca2+ overflow, energy metabolism disruption and cellular organelles oxidation (Askew, 2002; Mungai et al., 2011). It is noted that such damage can occur in both aerobic and anaerobic exercises at any exercise intensity under hypoxic conditions (Bakonyi and Radak, 2004).
Moreover, physical exercise associated with mountain climbing also plays an important role in ROS production (Askew, 2002), as physical workouts at high altitudes can aggravate oxidative stress (Bakonyi and Radak, 2004; Miller et al., 2013). For example, enhanced DNA breakage and oxidation were frequently observed in exercising subjects at high altitudes compared to sea level (Møller et al., 2001; Ziogas et al., 2010). The antioxidant system in the body is particularly vulnerable under stressed conditions such as hypoxia, and is unable to prevent DNA damage caused by exercise at high altitudes (Møller et al., 2001). In addition to physical exercise and hypobaric hypoxia, other environmental factors including coldness, sunburn and diet also contribute to the augmentation of oxidative stress at high altitudes (Askew, 2002). Insufficient antioxidant intake may exacerbate high altitude-induced anorexia as well as tissue damage (Askew, 2002; Bailey et al., 2004). Thus, caution should be taken at high altitudes as mountaineers could experience intense oxidative stress from both high altitude environments and physical workouts.
In the past decades, majority of studies mainly emphasize on the detrimental effects of exercised-induced oxidative stress on muscles, whereas researchers recently reported the significance of ROS in triggering and mediating body's adaptive responses to exercise (Yavari et al., 2015). Acute exercise generates excessive ROS that cause damage in the body, while regular exercise results in bodily adaptations leading to resistance against oxidative damage via antioxidant pathways (Yavari et al., 2015). It has been observed that the antioxidant capacity of skeletal muscle can be altered by exercise training. For example, SOD levels are commonly higher in the resting blood and muscle of trained individuals compared to those of control groups (Tiidus et al., 1996). Endurance training may increase the activities of SOD and GPx in both plasma and exercised muscles (Lambertucci et al., 2007; Brooks et al., 2008; Vieira Junior et al., 2013; Azizbeigi et al., 2014). This magnitude of exercise-mediated changes in SOD or GPx activities is dependent on the intensity and duration of that specific exercise. For example, high-intensity exercises may lead to a higher muscular GPx activity than that in low-intensity ones (Powers et al., 1994; Fisher et al., 2011). Similarly, long-duration exercise trainings (e.g., 60 min/day) increase more muscular GPx function than short-duration (30 min/day) exercise bouts (Powers et al., 1994). The enhancement of exercise-induced SOD and GPx activity is fiber type-specific, and a greater increase is normally observed in skeletal muscles mainly composed of highly oxidative fibers (e.g., type I and type IIa) (Powers et al., 1994; Gonchar, 2005; Ferraro et al., 2014). However, whether catalase (another major antioxidant enzyme) expression or activity can be affected by chronic exercise remains controversial, as previous studies reported mixed results (Vincent et al., 2000; Brooks et al., 2008; Liberali et al., 2016).
Several important pathways have been proposed in mediating the adaptive responses to exercise training (Morris et al., 2008; Samjoo et al., 2013; Csala et al., 2015). It is suggested that mitochondrial ROS generated during regular exercise are necessary for the activation of primary signaling pathways associated with muscle adaptation (Yavari et al., 2015). Nuclear factor erythroid 2-related factor (Nrf2), a redox-sensing transcription factor, is the primary regulator of antioxidants as well as other cytoprotective cofactors that are responsible for the enhanced antioxidant defense system (Osburn and Kensler, 2008; Muthusamy et al., 2012). Upregulated Nrf2 expression occurs after high-intensity exercise (Gounder et al., 2012). In a mouse myocardium, acute exercise activates Nrf2 signaling via increased ROS production, which in turn, promotes the trans-activation of antioxidant genes, leading to improved cardioprotection (Muthusamy et al., 2012; Figure 1). However, there is a lack of human studies that address the Nrf2-mediated adaptive responses generated by exercise. Another adaptation to exercise involves the enhancement of mitochondrial biogenesis via upregulated peroxisome proliferator-activated receptor-γ coactivator-1α (PGC-1α) gene expression (Steinbacher and Eckl, 2015). PGC-1α has been demonstrated to upregulate Nrf2 in order to control mitochondrial biogenesis (Wu et al., 1999; Wright et al., 2007). The upstream signals that regulate PGC-1α expression such as mitogen-activated protein kinase (MAPK) and nuclear factor (NF)-κB are redox-sensitive (Dodd et al., 2010; Derbre et al., 2012). In addition, proteasome inhibition, which reduces NF-κB activation, has been shown to enhance cellular antioxidant defenses via an Nrf2-dependent transcriptional mechanism, suggesting the indirect effects of NF-κB on antioxidant regulation (Karin and Ben-Neriah, 2000; Elliott et al., 2003; Dreger et al., 2009; Figure 1). Exercise-induced ROS also plays a role in adaptation through the oxidation of cysteine residue in various proteins. For example, cysteine-rich peroxiredoxin, an antioxidant responsible for H2O2 catalysis, is oxidized and formed stable dimers in response to elevated H2O2 levels during exercise, managing H2O2 gradients and regulating extracellular redox-signaling (Wadley et al., 2016). Moreover, the disulfide bonds formed by oxidized cysteine residues likely enhance protein synthesis in active individuals (Buresh and Berg, 2015).
Exhaustive endurance and/or resistance exercise may induce temporary immunosuppression (i.e., a reduction in CD4/CD8) (Jin et al., 2015). Particularly, the elevated oxidative and physical stress reflected by the level of intracellular ROS and cortisol, respectively, may contribute to the immunosuppression (Jin et al., 2015; Figure 1). For example, NF-κB is activated in response to stimulants such as H2O2, TNF-α, and other proinflammatory cytokines (e.g., IL-6). The activated NF-κB then binds to a specific DNA binding domain and upregulates the corresponding antioxidant gene expression (e.g., SOD) (Morgan and Liu, 2011; Figure 1). Accordingly, the NF-κB signaling pathway can be activated following an acute bout of exercise in rats (Ji, 2007). In addition, low levels of inflammatory markers have been observed in the elderly who frequently exercise (Marzatico et al., 1997). As mentioned previously, MAPK also plays an important role in exercise-induced adaptation in skeletal muscle. MAPK is composed of four subfamilies (ERK1/2, JNK, p38 MAPK, and ERK5) (Kramer and Goodyear, 2007; Figure 1). The activities of ERK and MEK have a positive correlation with exercise intensity in human skeletal muscle (Widegren et al., 2000). ROS such as H2O2, can induce the activation of ERK, JNK, and p38 MAPK in skeletal myoblasts in a dose- and time-dependent manner (Kefaloyianni et al., 2006). Oxidative stress could also modulate the MAPK signaling pathway through insulin signaling and glucose transport (Kim et al., 2006; Sandström et al., 2006; Kramer and Goodyear, 2007; Figure 1).
Growing evidence on exercise-induced oxidative damage and impaired muscle performance has prompted intensive research into the efficacy of antioxidant supplementation in exercising individuals (Gomes et al., 2012). It has been suggested that oral antioxidant supplements, which are common intakes among athletes, support endogenous antioxidant defense system against oxidative stress (Peternelj and Coombes, 2011). However, studies on the effects of antioxidant supplements in muscle damage prevention and recovery remain inconsistent, mostly due to different exercise protocols, research designs, and analytical methods (Peternelj and Coombes, 2011).
Most commonly known antioxidants are vitamins, which can be obtained readily through natural foods such as vegetables and fruits (Trapp et al., 2010). Indeed, vegetarians have been shown to have higher levels of endogenous vitamin than omnivores due to antioxidant–rich diets, providing effective protections against exercise-induced oxidative stress (Rauma and Mykkanen, 2000; Trapp et al., 2010). Similar nutritional strategy is wisely utilized by the athletes to improve performance and promote hastened muscle recovery (Margaritis and Rousseau, 2008). Antioxidant vitamins have demonstrated potential prophylactic effects. In the study performed by He et al., short-term combined vitamin C and E supplementation not only attenuated levels of creatine kinase (a muscle damage marker) and muscle soreness, but also enhanced muscle protection following the second bout of aerobic exercise (He et al., 2015). Moreover, a supplemental or adequate intake of nutritional antioxidants is necessary for endurance athletes (Wagner et al., 2010). For example, long-distance runners who took vitamin C and E for 4 or 5 weeks prior to a marathon experienced less muscle damage (Urso and Clarkson, 2003). Likewise, Fogarty et al. reported that both short- and long-term supplementation of watercress, which is rich in lipid soluble antioxidants (i.e., α-tocopherol, β-carotene, and xanthophyll), can reduce exhaustive exercise-associated lipid peroxidation and DNA damage (Fogarty et al., 2011). Phenolic compounds found in grapes also exhibited great antioxidant and anti-inflammatory properties, and has been shown to improve exercise performance in recreational runners (15% increase in time-to-exhaustion running) (Ali et al., 2010; Toscano et al., 2015). Moreover, Pala et al. suggested that coenzyme Q10 supplementation protects tissue from oxidative injury during exercise training through a mechanism involving Nrf2 expressions (Pala et al., 2016).
Despite beneficial effects mentioned above, a thorough understanding on the application of vitamin and antioxidant supplements such as effective dosage and administration method is necessary to avoid undesirable effects. Some studies have indicated that antioxidant supplements fail to protect against the damaging effects of oxidative stress such as exercise-induced lipid peroxidation and inflammation, both of which hinder muscle recovery (Teixeira et al., 2009). Specifically, prolonged antioxidant supplementation is not recommended since it can disrupt endogenous antioxidant levels and interfere exercise-induced adaptation, thereby blunting body's defense against oxidative stress (Peternelj and Coombes, 2011; Rowlands et al., 2012). Excessive antioxidant intake, such as vitamin C and E supplementation, has been shown to delay healing process and muscle strength restoration in athletes following an exhaustive exercise training (Margaritis and Rousseau, 2008; Theodorou et al., 2011). Additionally, an increased exercise-induced oxidative stress is observed in individual taking high-doses of α-tocopherol (Margaritis and Rousseau, 2008). In short term, N-acetyl-cysteine (NAC; antioxidant) and allopurinol (an inhibitor of XO) do attenuate muscle damage and lipid oxidation caused by acute exhaustive exercise (Gómez-Cabrera et al., 2003; Braakhuis and Hopkins, 2015). Nevertheless, long-term intakes of these antioxidants may not be beneficial (Braakhuis and Hopkins, 2015). Gomez-Cabrera et al. further suggested that 8 weeks of vitamin C supplementation prevents training-induced mitochondrial biogenesis by suppressing the expression of SOD and GPx (Gomez-Cabrera et al., 2008). A double-blinded and placebo-controlled study also showed that the combination of vitamins C and E blunts mitochondrial adaptive responses (i.e., increase in COX4 protein) after 11 weeks of endurance training (Paulsen et al., 2014).
Collectively, mixed results from antioxidant intervention studies may be interpreted by the variances in participants' baseline redox status, the dose and length of the antioxidant supplementation, and the choice of oxidative stress markers. Instead of antioxidant supplements, a balanced diet consisting natural antioxidants from fruits and vegetables is sufficient to meet the dietary requirement for physically active individuals (Bloomer et al., 2007; Poljsak et al., 2013; Yavari et al., 2015).
In the past decades, exercise-induced oxidative stress and its effects have been largely studied. Despite the increasingly sophisticated approaches on the study of ROS in skeletal muscle, inconsistency in the results of several studies remains, which is likely associated with different methodology of ROS measurements and exercise protocols. It is therefore essential to determine an appropriate measuring module for various types of exercises and muscles in order to obtain reliable and valid data (Zuo et al., 2015a; Jackson, 2016). Excessive ROS production beyond the capability of antioxidant defense following exhaustive and/or unaccustomed exercise could adversely affect human adaptive responses. The current challenge is the lack of in-depth human studies that explore the molecular mechanisms of how ROS regulate the key redox-sensitive transcription factors including Nrf2, NF-κB, MAPK and PGC-1α. Further studies focusing on minimizing oxidative damage and maximizing adaptive response induced by exercise are indispensable. Developing promising strategies that combine an effective natural antioxidant diet with customized exercise within a variety of populations (e.g., disease population, obese individuals, the elderly, and trained/untrained individuals) could tremendously improve health and quality of life. Moreover, identifying the effective and reliable biomarkers of alterations in redox homeostasis is critical in monitoring the training tolerance of individuals and may shed a light on optimizing a customized training program.
LZ, FH designed the outline, FH, JL, ZL, CC, WY, LZ wrote the paper.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
This study was funded by 2016 American Physiology Society S&R Foundation Ryuji Ueno Award, OSU-HRS #013000, Initiative Grant for International Collaboration of Biomedical Research (#22010130), China Scholarship Council and China University of Geosciences Collaboration Research Fund. We acknowledge the generous assistance from Tingyang Zhou, Andrew Graef, Michael Chien, Zan Xu, Nicole Otenbaker, Joshua Stringer and Alexander Ziegler.
Agarwal, A., Saleh, R. A., and Bedaiwy, M. A. (2003). Role of reactive oxygen species in the pathophysiology of human reproduction. Fertil. Steril. 79, 829–843. doi: 10.1016/S0015-0282(02)04948-8
Alfadda, A. A., and Sallam, R. M. (2012). Reactive oxygen species in health and disease. J. Biomed. Biotechnol. 2012:936486. doi: 10.1155/2012/936486
Ali, K., Maltese, F., Choi, Y. H., and Verpoorte, R. (2010). Metabolic constituents of grapevine and grape-derived products. Phytochem. Rev. 9, 357–378. doi: 10.1007/s11101-009-9158-0
Askew, E. W. (2002). Work at high altitude and oxidative stress: antioxidant nutrients. Toxicology 180, 107–119. doi: 10.1016/S0300-483X(02)00385-2
Azizbeigi, K., Stannard, S. R., Atashak, S., and Haghighi, M. M. (2014). Antioxidant enzymes and oxidative stress adaptation to exercise training: comparison of endurance, resistance, and concurrent training in untrained males. J. Exerc. Sci. Fit. 12, 1–6. doi: 10.1016/j.jesf.2013.12.001
Bailey, D. M., Ainslie, P. N., Jackson, S. K., Richardson, R. S., and Ghatei, M. (2004). Evidence against redox regulation of energy homoeostasis in humans at high altitude. Clin. Sci. 107, 589–600. doi: 10.1042/CS20040085
Basnyat, B. (2001). High-altitude illness. N. Engl. J. Med. 345, 1279. author reply: 1280–1281. doi: 10.1056/NEJM200110253451713
Basnyat, B., Gertsch, J. H., Johnson, E. W., Castro-Marin, F., Inoue, Y., and Yeh, C. (2003). Efficacy of low-dose acetazolamide (125 mg BID) for the prophylaxis of acute mountain sickness: a prospective, double-blind, randomized, placebo-controlled trial. High Alt. Med. Biol. 4, 45–52. doi: 10.1089/152702903321488979
Berzosa, C., Cebrián, I., Fuentes-Broto, L., Gómez-Trullén, E., Piedrafita, E., Martínez-Ballarín, E., et al. (2011). Acute exercise increases plasma total antioxidant status and antioxidant enzyme activities in untrained men. J. Biomed. Biotechnol. 2011:540458. doi: 10.1155/2011/540458
Bloomer, R. J., Falvo, M. J., Schilling, B. K., and Smith, W. A. (2007). Prior exercise and antioxidant supplementation: effect on oxidative stress and muscle injury. J. Int. Soc. Sports Nutr. 4:9. doi: 10.1186/1550-2783-4-9
Bors, W., Michel, C., Saran, M., and Lengfelder, E. (1978). The involvement of oxygen radicals during the autoxidation of adrenalin. Biochim. Biophys. Acta 540, 162–172. doi: 10.1016/0304-4165(78)90445-2
Braakhuis, A. J., and Hopkins, W. G. (2015). Impact of dietary antioxidants on sport performance: a review. Sports Med. 45, 939–955. doi: 10.1007/s40279-015-0323-x
Brooks, S. V., Vasilaki, A., Larkin, L. M., McArdle, A., and Jackson, M. J. (2008). Repeated bouts of aerobic exercise lead to reductions in skeletal muscle free radical generation and nuclear factor κB activation. J. Physiol. 586, 3979–3990. doi: 10.1113/jphysiol.2008.155382
Buresh, R., and Berg, K. (2015). A tutorial on oxidative stress and redox signaling with application to exercise and sedentariness. Sports Med. 1:3. doi: 10.1186/s40798-40014-40003-40797
Cakir-Atabek, H., Demir, S., Pinarbasili, R. D., and Gündüz, N. (2010). Effects of different resistance training intensity on indices of oxidative stress. J. Strength Cond. Res. 24, 2491–2497. doi: 10.1519/JSC.0b013e3181ddb111
Cheng, A. J., Yamada, T., Rassier, D., Andersson, D. C., Westerblad, H., and Lanner, J. T. (2016). ROS/RNS and contractile function in skeletal muscle during fatigue and recovery. J. Physiol. 594, 5149–5160. doi: 10.1113/JP270650
Close, G. L., Ashton, T., Cable, T., Doran, D., and Maclaren, D. P. M. (2004). Eccentric exercise, isokinetic muscle torque and delayed onset muscle soreness: the role of reactive oxygen species. Eur. J. Appl. Physiol. 91, 615–621. doi: 10.1007/s00421-003-1012-2
Csala, M., Kardon, T., Legeza, B., Lizák, B., Mandl, J., Margittai, É., et al. (2015). On the role of 4-hydroxynonenal in health and disease. Biochim. Biophys Acta. 1852, 826–838. doi: 10.1016/j.bbadis.2015.01.015
Derbre, F., Ferrando, B., Gomez-Cabrera, M. C., Sanchis-Gomar, F., Martinez-Bello, V. E., Olaso-Gonzalez, G., et al. (2012). Inhibition of xanthine oxidase by allopurinol prevents skeletal muscle atrophy: role of p38 MAPKinase and E3 ubiquitin ligases. PLoS ONE 7:e46668. doi: 10.1371/journal.pone.0046668
Dodd, S. L., Gagnon, B. J., Senf, S. M., Hain, B. A., and Judge, A. R. (2010). Ros-mediated activation of NF-kappaB and Foxo during muscle disuse. Muscle Nerve 41, 110–113. doi: 10.1002/mus.21526
Dreger, H., Westphal, K., Weller, A., Baumann, G., Stangl, V., Meiners, S., et al. (2009). Nrf2-dependent upregulation of antioxidative enzymes: a novel pathway for proteasome inhibitor-mediated cardioprotection. Cardiovasc. Res. 83, 354–361. doi: 10.1093/cvr/cvp107
Elliott, P. J., Soucy, T. A., Pien, C. S., Adams, J., and Lightcap, E. S. (2003). Assays for proteasome inhibition. Methods Mol. Med. 85, 163–172. doi: 10.1385/1-59259-380-1:163
Espinosa, A., Leiva, A., Peña, M., Müller, M., Debandi, A., Hidalgo, C., et al. (2006). Myotube depolarization generates reactive oxygen species through NAD (P) H oxidase; ROS-elicited Ca2+ stimulates ERK, CREB, early genes. J. Cell. Physiol. 209, 379–388. doi: 10.1002/jcp.20745
Ferraro, E., Giammarioli, A. M., Chiandotto, S., Spoletini, I., and Rosano, G. (2014). Exercise-induced skeletal muscle remodeling and metabolic adaptation: redox signaling and role of autophagy. Antioxid. Redox Signal. 21, 154–176. doi: 10.1089/ars.2013.5773
Fisher, G., Schwartz, D. D., Quindry, J., Barberio, M. D., Foster, E. B., Jones, K. W., et al. (2011). Lymphocyte enzymatic antioxidant responses to oxidative stress following high-intensity interval exercise. J. Appl. Physiol. (1985) 110, 730–737. doi: 10.1152/japplphysiol.00575.2010
Fogarty, M. C., Hughes, C. M., Burke, G., Brown, J. C., Trinick, T. R., Duly, E., et al. (2011). Exercise-induced lipid peroxidation: implications for deoxyribonucleic acid damage and systemic free radical generation. Environ. Mol. Mutagen. 52, 35–42. doi: 10.1002/em.20572
Garatachea, N., Garcia-Lopez, D., Bernal, A., Almar, M., and Gonzalez-Gallego, J. (2012). Oxidative stress response to isometric exercise in women: effect of age and exercise intensity. Int. Sports Med. J. 13, 85–95.
Georgakouli, K., Manthou, E., Fatouros, I. G., Deli, C. K., Spandidos, D. A., Tsatsakis, A. M., et al. (2015). Effects of acute exercise on liver function and blood redox status in heavy drinkers. Exp. Ther. Med. 10, 2015–2022. doi: 10.3892/etm.2015.2792
Gomes, E. C., Silva, A. N., and de Oliveira, M. R. (2012). Oxidants, antioxidants, and the beneficial roles of exercise-induced production of reactive species. Oxid. Med. Cell. Longev. 2012:756132. doi: 10.1155/2012/756132
Gomez-Cabrera, M. C., Close, G. L., Kayani, A., McArdle, A., Vina, J., and Jackson, M. J. (2010). Effect of xanthine oxidase-generated extracellular superoxide on skeletal muscle force generation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 298, R2–R8. doi: 10.1152/ajpregu.00142.2009
Gomez-Cabrera, M. C., Domenech, E., Romagnoli, M., Arduini, A., Borras, C., Pallardo, F. V., et al. (2008). Oral administration of vitamin C decreases muscle mitochondrial biogenesis and hampers training-induced adaptations in endurance performance. Am. J. Clin. Nutr. 87, 142–149.
Gómez-Cabrera, M. C., Pallardó, F. V., Sastre, J., Viña, J., and García-del-Moral, L. (2003). Allopurinol and markers of muscle damage among participants in the Tour de France. JAMA 289, 2503–2504. doi: 10.1001/jama.289.19.2503-b
Gonchar, O. (2005). Muscle fiber specific antioxidative system adaptation to swim training in rats: influence of intermittent hypoxia. J. Sports Sci. Med. 4, 160–169.
Gounder, S. S., Kannan, S., Devadoss, D., Miller, C. J., Whitehead, K. J., Odelberg, S. J., et al. (2012). Impaired transcriptional activity of Nrf2 in age-related myocardial oxidative stress is reversible by moderate exercise training. PLoS ONE 7:e45697. doi: 10.1371/journal.pone.0045697
Groussard, C., Morel, I., Chevanne, M., Monnier, M., Cillard, J., and Delamarche, A. (2000). Free radical scavenging and antioxidant effects of lactate ion: an in vitro study. J. Appl. Physiol. (1985) 89, 169–175.
Hackett, P. H., and Roach, R. C. (2001). High-altitude illness. N. Engl. J. Med. 345, 107–114. doi: 10.1056/NEJM200107123450206
He, F., Hockemeyer, J. A., and Sedlock, D. (2015). Does combined antioxidant vitamin supplementation blunt repeated bout effect? Int. J. Sports Med. 36, 407–413. doi: 10.1055/s-0034-1395630
Herrero, A., and Barja, G. (1997). ADP-regulation of mitochondrial free radical production is different with complex I- or complex II-linked substrates: implications for the exercise paradox and brain hypermetabolism. J. Bioenerg. Biomembr. 29, 241–249. doi: 10.1023/A:1022458010266
Hidalgo, C., Sánchez, G., Barrientos, G., and Aracena-Parks, P. (2006). A transverse tubule NADPH oxidase activity stimulates calcium release from isolated triads via ryanodine receptor type 1 S-glutathionylation. J. Biol. Chem. 281, 26473–26482. doi: 10.1074/jbc.M600451200
Jackson, M. J. (2016). Recent advances and long-standing problems in detecting oxidative damage and reactive oxygen species in skeletal muscle. J. Physiol. 594, 5185–5193. doi: 10.1113/JP270657
Ji, L. L. (2007). Antioxidant signaling in skeletal muscle: a brief review. Exp. Gerontol. 42, 582–593. doi: 10.1016/j.exger.2007.03.002
Jiang, L., Diaz, P. T., Best, T. M., Stimpfl, J. N., He, F., and Zuo, L. (2014). Molecular characterization of redox mechanisms in allergic asthma. Ann. Allergy Asthma Immunol. 113, 137–142. doi: 10.1016/j.anai.2014.05.030
Jin, C. H., Paik, I. Y., Kwak, Y. S., Jee, Y. S., and Kim, J. Y. (2015). Exhaustive submaximal endurance and resistance exercises induce temporary immunosuppression via physical and oxidative stress. J. Exerc. Rehabil. 11, 198–203. doi: 10.12965/jer.150221
Julian, C. G., Subudhi, A. W., Hill, R. C., Wilson, M. J., Dimmen, A. C., Hansen, K. C., et al. (2014). Exploratory proteomic analysis of hypobaric hypoxia and acute mountain sickness in humans. J. Appl. Physiol. (1985) 116, 937–944. doi: 10.1152/japplphysiol.00362.2013
Kang, C., O'moore, K. M., Dickman, J. R., and Ji, L. L. (2009). Exercise activation of muscle peroxisome proliferator-activated receptor-gamma coactivator-1alpha signaling is redox sensitive. Free Radic. Biol. Med. 47, 1394–1400. doi: 10.1016/j.freeradbiomed.2009.08.007
Karin, M., and Ben-Neriah, Y. (2000). Phosphorylation meets ubiquitination: the control of NF-[kappa]B activity. Annu. Rev. Immunol. 18, 621–663. doi: 10.1146/annurev.immunol.18.1.621
Kavazis, A. N., Talbert, E. E., Smuder, A. J., Hudson, M. B., Nelson, W. B., and Powers, S. K. (2009). Mechanical ventilation induces diaphragmatic mitochondrial dysfunction and increased oxidant production. Free Radic. Biol. Med. 46, 842–850. doi: 10.1016/j.freeradbiomed.2009.01.002
Kefaloyianni, E., Gaitanaki, C., and Beis, I. (2006). ERK1/2 and p38-MAPK signalling pathways, through MSK1, are involved in NF-kappaB transactivation during oxidative stress in skeletal myoblasts. Cell. Signal. 18, 2238–2251. doi: 10.1016/j.cellsig.2006.05.004
Kim, J. S., Saengsirisuwan, V., Sloniger, J. A., Teachey, M. K., and Henriksen, E. J. (2006). Oxidant stress and skeletal muscle glucose transport: roles of insulin signaling and p38 MAPK. Free Radic. Biol. Med. 41, 818–824. doi: 10.1016/j.freeradbiomed.2006.05.031
Knez, W. L., Coombes, J. S., and Jenkins, D. G. (2006). Ultra-endurance exercise and oxidative damage: implications for cardiovascular health. Sports Med. 36, 429–441. doi: 10.2165/00007256-200636050-00005
Kramer, H. F., and Goodyear, L. J. (2007). Exercise, MAPK, and NF-kappaB signaling in skeletal muscle. J. Appl. Physiol. (1985) 103, 388–395. doi: 10.1152/japplphysiol.00085.2007
Lambertucci, R. H., Levada-Pires, A. C., Rossoni, L. V., Curi, R., and Pithon-Curi, T. C. (2007). Effects of aerobic exercise training on antioxidant enzyme activities and mRNA levels in soleus muscle from young and aged rats. Mech. Ageing Dev. 128, 267–275. doi: 10.1016/j.mad.2006.12.006
Liberali, R., Wilhelm Filho, D., and Petroski, E. L. (2016). Aerobic and anaerobic training sessions promote antioxidant changes in young male soccer players. Med. Exp. 3:1. doi: 10.5935/medicalexpress.2016.01.07
Marciniak, A., Brzeszczynska, J., Gwozdzinski, K., and Jegier, A. (2009). Antioxidant capacity and physical exercise. Biol. Sport 26:197. doi: 10.5604/20831862.894649
Margaritis, I., and Rousseau, A. S. (2008). Does physical exercise modify antioxidant requirements? Nutr. Res. Rev. 21, 3–12. doi: 10.1017/S0954422408018076
Marzatico, F., Pansarasa, O., Bertorelli, L., Somenzini, L., and Della Valle, G. (1997). Blood free radical antioxidant enzymes and lipid peroxides following long-distance and lactacidemic performances in highly trained aerobic and sprint athletes. J. Sports Med. Phys. Fitness 37, 235–239.
Mastaloudis, A., Leonard, S. W., and Traber, M. G. (2001). Oxidative stress in athletes during extreme endurance exercise. Free Radic. Biol. Med. 31, 911–922. doi: 10.1016/S0891-5849(01)00667-0
Mastaloudis, A., Morrow, J. D., Hopkins, D. W., Devaraj, S., and Traber, M. G. (2004). Antioxidant supplementation prevents exercise-induced lipid peroxidation, but not inflammation, in ultramarathon runners. Free Radic. Biol. Med. 36, 1329–1341. doi: 10.1016/j.freeradbiomed.2004.02.069
Michaelson, L. P., Shi, G., Ward, C. W., and Rodney, G. G. (2010). Mitochondrial redox potential during contraction in single intact muscle fibers. Muscle Nerve 42, 522–529. doi: 10.1002/mus.21724
Miller, L. E., McGinnis, G. R., Kliszczewicz, B., Slivka, D., Hailes, W., Cuddy, J., et al. (2013). Blood oxidative-stress markers during a high-altitude trek. Int. J. Sport Nutr. Exerc. Metab. 23, 65–72. doi: 10.1123/ijsnem.23.1.65
Miyata, M., Kasai, H., Kawai, K., Yamada, N., Tokudome, M., Ichikawa, H., et al. (2008). Changes of urinary 8-hydroxydeoxyguanosine levels during a two-day ultramarathon race period in Japanese non-professional runners. Int. J. Sports Med. 29, 27–33. doi: 10.1055/s-2007-965072
Møller, P., Loft, S., Lundby, C., and Olsen, N. V. (2001). Acute hypoxia and hypoxic exercise induce DNA strand breaks and oxidative DNA damage in humans. FASEB J. 15, 1181–1186. doi: 10.1096/fj.00-0703com
Morales-Alamo, D., and Calbet, J. A. (2014). Free radicals and sprint exercise in humans. Free Radic. Res. 48, 30–42. doi: 10.3109/10715762.2013.825043
Morgan, M. J., and Liu, Z. G. (2011). Crosstalk of reactive oxygen species and NF-kappaB signaling. Cell Res. 21, 103–115. doi: 10.1038/cr.2010.178
Morris, R. T., Laye, M. J., Lees, S. J., Rector, R. S., Thyfault, J. P., and Booth, F. W. (2008). Exercise-induced attenuation of obesity, hyperinsulinemia, and skeletal muscle lipid peroxidation in the OLETF rat. J. Appl. Physiol. (1985) 104, 708–715. doi: 10.1152/japplphysiol.01034.2007
Mungai, P. T., Waypa, G. B., Jairaman, A., Prakriya, M., Dokic, D., Ball, M. K., et al. (2011). Hypoxia triggers AMPK activation through reactive oxygen species-mediated activation of calcium release-activated calcium channels. Mol. Cell. Biol. 31, 3531–3545. doi: 10.1128/MCB.05124-11
Muthusamy, V. R., Kannan, S., Sadhaasivam, K., Gounder, S. S., Davidson, C. J., Boeheme, C., et al. (2012). Acute exercise stress activates Nrf2/ARE signaling and promotes antioxidant mechanisms in the myocardium. Free Radic. Biol. Med. 52, 366–376. doi: 10.1016/j.freeradbiomed.2011.10.440
Neubauer, O., Reichhold, S., Nics, L., Hoelzl, C., Valentini, J., Stadlmayr, B., et al. (2010). Antioxidant responses to an acute ultra-endurance exercise: impact on DNA stability and indications for an increased need for nutritive antioxidants in the early recovery phase. Br. J. Nutr. 104, 1129–1138. doi: 10.1017/S0007114510001856
Nielsen, H. G., Hagberg, I. A., and Lyberg, T. (2004). Marathon running leads to partial exhaustion of ROS-generating capacity in leukocytes. Med. Sci. Sports Exerc. 36, 68–73. doi: 10.1249/01.MSS.0000106168.12113.95
Nikolaidis, M. G., Jamurtas, A. Z., Paschalis, V., Fatouros, I. G., Koutedakis, Y., and Kouretas, D. (2008). The effect of muscle-damaging exercise on blood and skeletal muscle oxidative stress. Sports Med. 38, 579–606. doi: 10.2165/00007256-200838070-00005
Nikolaidis, M. G., Paschalis, V., Giakas, G., Fatouros, I. G., Koutedakis, Y., Kouretas, D., et al. (2007). Decreased blood oxidative stress after repeated muscle-damaging exercise. Med. Sci. Sports Exerc. 39, 1080–1089. doi: 10.1249/mss.0b013e31804ca10c
Osburn, W. O., and Kensler, T. W. (2008). Nrf2 signaling: an adaptive response pathway for protection against environmental toxic insults. Mutat. Res. 659, 31–39. doi: 10.1016/j.mrrev.2007.11.006
Pala, R., Orhan, C., Tuzcu, M., Sahin, N., Ali, S., Cinar, V., et al. (2016). Coenzyme Q10 Supplementation Modulates NFkappaB and Nrf2 Pathways in Exercise Training. J. Sports Sci. Med. 15, 196–203.
Palomero, J., Vasilaki, A., Pye, D., McArdle, A., and Jackson, M. J. (2013). Aging increases the oxidation of dichlorohydrofluorescein in single isolated skeletal muscle fibers at rest, but not during contractions. Am. J. Physiol. Regul. Integr. Comp. Physiol. 305, R351–R358. doi: 10.1152/ajpregu.00530.2012
Parker, L., McGuckin, T. A., and Leicht, A. S. (2014). Influence of exercise intensity on systemic oxidative stress and antioxidant capacity. Clin. Physiol. Funct. Imaging 34, 377–383. doi: 10.1111/cpf.12108
Paulsen, G., Cumming, K. T., Holden, G., Hallén, J., Rønnestad, B. R., Sveen, O., et al. (2014). Vitamin C and E supplementation hampers cellular adaptation to endurance training in humans: a double-blind, randomised, controlled trial. J. Physiol. 592, 1887–1901. doi: 10.1113/jphysiol.2013.267419
Peternelj, T. T., and Coombes, J. S. (2011). Antioxidant supplementation during exercise training: beneficial or detrimental? Sports Med. 41, 1043–1069. doi: 10.2165/11594400-000000000-00000
Poljsak, B., Šuput, D., and Milisav, I. (2013). Achieving the balance between ROS and antioxidants: when to use the synthetic antioxidants. Oxid. Med. Cell. Longev. 2013:956792. doi: 10.1155/2013/956792
Powers, S. K., Criswell, D., Lawler, J., Ji, L. L., Martin, D., Herb, R. A., et al. (1994). Influence of exercise and fiber type on antioxidant enzyme activity in rat skeletal muscle. Am. J. Physiol. Regul. Integr. Comp. Physiol. 266, R375–R380.
Powers, S. K., and Jackson, M. J. (2008). Exercise-induced oxidative stress: cellular mechanisms and impact on muscle force production. Physiol. Rev. 88, 1243–1276. doi: 10.1152/physrev.00031.2007
Powers, S. K., Ji, L. L., Kavazis, A. N., and Jackson, M. J. (2011a). Reactive oxygen species: impact on skeletal muscle. Compr. Physiol. 1, 941–969. doi: 10.1002/cphy.c100054
Powers, S. K., Nelson, W. B., and Hudson, M. B. (2011b). Exercise-induced oxidative stress in humans: cause and consequences. Free Radic. Biol. Med. 51, 942–950. doi: 10.1016/j.freeradbiomed.2010.12.009
Radak, Z., Zhao, Z., Koltai, E., Ohno, H., and Atalay, M. (2013). Oxygen consumption and usage during physical exercise: the balance between oxidative stress and ROS-dependent adaptive signaling. Antioxid. Redox Signal. 18, 1208–1246. doi: 10.1089/ars.2011.4498
Rauma, A. L., and Mykkanen, H. (2000). Antioxidant status in vegetarians versus omnivores. Nutrition 16, 111–119. doi: 10.1016/S0899-9007(99)00267-1
Reid, M. B., Khawli, F. A., and Moody, M. R. (1993). Reactive oxygen in skeletal muscle. III. Contractility of unfatigued muscle. J. Appl. Physiol. (1985) 75, 1081–1087.
Rodriguez, M. C., Rosenfeld, J., and Tarnopolsky, M. A. (2003). Plasma malondialdehyde increases transiently after ischemic forearm exercise. Med. Sci. Sports Exerc. 35, 1859–1865. doi: 10.1249/01.MSS.0000093609.75937.70
Rowlands, D. S., Pearce, E., Aboud, A., Gillen, J. B., Gibala, M. J., Donato, S., et al. (2012). Oxidative stress, inflammation, and muscle soreness in an 894-km relay trail run. Eur. J. Appl. Physiol. 112, 1839–1848. doi: 10.1007/s00421-011-2163-1
Sahlin, K., Cizinsky, S., Warholm, M., and Höberg, J. (1992). Repetitive static muscle contractions in humans—a trigger of metabolic and oxidative stress? Eur. J. Appl. Physiol. 64, 228–236. doi: 10.1007/BF00626285
Sakellariou, G. K., Jackson, M. J., and Vasilaki, A. (2014). Redefining the major contributors to superoxide production in contracting skeletal muscle. The role of NAD(P)H oxidases. Free Radic. Res. 48, 12–29. doi: 10.3109/10715762.2013.830718
Sakellariou, G. K., Vasilaki, A., Palomero, J., Kayani, A., Zibrik, L., McArdle, A., et al. (2013). Studies of mitochondrial and nonmitochondrial sources implicate nicotinamide adenine dinucleotide phosphate oxidase(s) in the increased skeletal muscle superoxide generation that occurs during contractile activity. Antioxid. Redox Signal. 18, 603–621. doi: 10.1089/ars.2012.4623
Samjoo, I. A., Safdar, A., Hamadeh, M. J., Raha, S., and Tarnopolsky, M. A. (2013). The effect of endurance exercise on both skeletal muscle and systemic oxidative stress in previously sedentary obese men. Nutr. Diabetes 3:e88. doi: 10.1038/nutd.2013.30
Sandström, M. E., Zhang, S. J., Bruton, J., Silva, J. P., Reid, M. B., Westerblad, H., et al. (2006). Role of reactive oxygen species in contraction-mediated glucose transport in mouse skeletal muscle. J. Physiol. 575, 251–262. doi: 10.1113/jphysiol.2006.110601
Saran, M., Beck-Speier, I., Fellerhoff, B., and Bauer, G. (1999). Phagocytic killing of microorganisms by radical processes: consequences of the reaction of hydroxyl radicals with chloride yielding chlorine atoms. Free Radic. Biol. Med. 26, 482–490. doi: 10.1016/S0891-5849(98)00187-7
Siesjö, B. K., Katsura, K. I., Kristián, T., Li, P. A., and Siesjö, P. (1996). Molecular mechanisms of acidosis-mediated damage. Acta Neurochir. Suppl. 66, 8–14. doi: 10.1007/978-3-7091-9465-2_2
Silva, L. A., Pinho, C. A., Silveira, P. C., Tuon, T., De Souza, C. T., Dal-Pizzol, F., et al. (2010). Vitamin E supplementation decreases muscular and oxidative damage but not inflammatory response induced by eccentric contraction. J. Physiol. Sci. 60, 51–57. doi: 10.1007/s12576-009-0065-3
Smedley, T., and Grocott, M. P. (2013). Acute high-altitude illness: a clinically orientated review. Br. J. Pain 7, 85–94. doi: 10.1177/2049463713489539
Stagos, D., Goutzourelas, N., Ntontou, A. M., Kafantaris, I., Deli, C. K., Poulios, A., et al. (2015). Assessment of eccentric exercise-induced oxidative stress using oxidation-reduction potential markers. Oxid. Med. Cell. Longev. 2015:204615. doi: 10.1155/2015/204615
Steinbacher, P., and Eckl, P. (2015). Impact of oxidative stress on exercising skeletal muscle. Biomolecules 5, 356–377. doi: 10.3390/biom5020356
St-Pierre, J., Buckingham, J. A., Roebuck, S. J., and Brand, M. D. (2002). Topology of superoxide production from different sites in the mitochondrial electron transport chain. J. Biol. Chem. 277, 44784–44790. doi: 10.1074/jbc.M207217200
Talbert, E. E., Smuder, A. J., Min, K., Kwon, O. S., Szeto, H. H., and Powers, S. K. (2013). Immobilization-induced activation of key proteolytic systems in skeletal muscles is prevented by a mitochondria-targeted antioxidant. J. Appl. Physiol. (1985) 115, 529–538. doi: 10.1152/japplphysiol.00471.2013
Teixeira, V. H., Valente, H. F., Casal, S. I., Marques, A. F., and Moreira, P. A. (2009). Antioxidants do not prevent postexercise peroxidation and may delay muscle recovery. Med. Sci. Sports Exerc. 41, 1752–1760. doi: 10.1249/MSS.0b013e31819fe8e3
Theodorou, A. A., Nikolaidis, M. G., Paschalis, V., Koutsias, S., Panayiotou, G., Fatouros, I. G., et al. (2011). No effect of antioxidant supplementation on muscle performance and blood redox status adaptations to eccentric training. Am. J. Clin. Nutr. 93, 1373–1383. doi: 10.3945/ajcn.110.009266
Tiidus, P. M., Pushkarenko, J., and Houston, M. E. (1996). Lack of antioxidant adaptation to short-term aerobic training in human muscle. Am. J. Physiol. 271, R832–R836.
Toscano, L. T., Tavares, R. L., Toscano, L. T., Silva, C. S., Almeida, A. E., Biasoto, A. C., et al. (2015). Potential ergogenic activity of grape juice in runners. Appl. Physiol. Nutr. Metab. 40, 899–906. doi: 10.1139/apnm-2015-0152
Trapp, D., Knez, W., and Sinclair, W. (2010). Could a vegetarian diet reduce exercise-induced oxidative stress? A review of the literature. J. Sports Sci. 28, 1261–1268. doi: 10.1080/02640414.2010.507676
Urso, M. L., and Clarkson, P. M. (2003). Oxidative stress, exercise, and antioxidant supplementation. Toxicology 189, 41–54. doi: 10.1016/S0300-483X(03)00151-3
Vallecilla, C., Khiabani, R. H., Sandoval, N., Fogel, M., Briceño, J. C., and Yoganathan, A. P. (2014). Effect of high altitude exposure on the hemodynamics of the bidirectional Glenn physiology: modeling incremented pulmonary vascular resistance and heart rate. J. Biomech. 47, 1846–1852. doi: 10.1016/j.jbiomech.2014.03.021
Vasilaki, A., and Jackson, M. J. (2013). Role of reactive oxygen species in the defective regeneration seen in aging muscle. Free Radic. Biol. Med. 65, 317–323. doi: 10.1016/j.freeradbiomed.2013.07.008
Vieira Junior, R. C., Silva, C. M. S., Araújo, M. B. D., Garcia, A., Voltarelli, V. A., Reis Filho, A. D. D., et al. (2013). Aerobic swimming training increases the actitivy of antioxidant enzymes and the glycogen content in the skeletal muscle of rats. Rev. Brasileira Med. Esporte 19, 204–208. doi: 10.1590/S1517-86922013000300012
Vincent, H. K., Powers, S. K., Stewart, D. J., Demirel, H. A., Shanely, R. A., and Naito, H. (2000). Short-term exercise training improves diaphragm antioxidant capacity and endurance. Eur. J. Appl. Physiol. 81, 67–74. doi: 10.1007/PL00013799
Wadley, A. J., Aldred, S., and Coles, S. J. (2016). An unexplored role for Peroxiredoxin in exercise-induced redox signalling? Redox Biol. 8, 51–58. doi: 10.1016/j.redox.2015.10.003
Wagner, K. H., Reichhold, S., Hölzl, C., Knasmüller, S., Nics, L., Meisel, M., et al. (2010). Well-trained, healthy triathletes experience no adverse health risks regarding oxidative stress and DNA damage by participating in an ultra-endurance event. Toxicology 278, 211–216. doi: 10.1016/j.tox.2009.09.006
Wasse, L. K., Sunderland, C., King, J. A., Batterham, R. L., and Stensel, D. J. (2012). Influence of rest and exercise at a simulated altitude of 4000 m on appetite, energy intake, and plasma concentrations of acylated ghrelin and peptide YY. J. Appl. Physiol. (1985) 112, 552–559. doi: 10.1152/japplphysiol.00090.2011
Waterfall, A. H., Singh, G., Fry, J. R., and Marsden, C. A. (1996). Acute acidosis elevates malonaldehyde in rat brain in vivo. Brain Res. 712, 102–106. doi: 10.1016/0006-8993(95)01540-X
Whitehead, N. P., Yeung, E. W., Froehner, S. C., and Allen, D. G. (2010). Skeletal muscle NADPH oxidase is increased and triggers stretch-induced damage in the mdx mouse. PLoS ONE 5:e15354. doi: 10.1371/journal.pone.0015354
Widegren, U., Wretman, C., Lionikas, A., Hedin, G., and Henriksson, J. (2000). Influence of exercise intensity on ERK/MAP kinase signalling in human skeletal muscle. Pflugers Arch. 441, 317–322. doi: 10.1007/s004240000417
Wiggs, M. P. (2015). Can endurance exercise preconditioning prevention disuse muscle atrophy? Front. Physiol. 6:63. doi: 10.3389/fphys.2015.00063
Wright, D. C., Han, D. H., Garcia-Roves, P. M., Geiger, P. C., Jones, T. E., and Holloszy, J. O. (2007). Exercise-induced mitochondrial biogenesis begins before the increase in muscle PGC-1alpha expression. J. Biol. Chem. 282, 194–199. doi: 10.1074/jbc.M606116200
Wu, Z., Puigserver, P., Andersson, U., Zhang, C., Adelmant, G., Mootha, V., et al. (1999). Mechanisms controlling mitochondrial biogenesis and respiration through the thermogenic coactivator PGC-1. Cell 98, 115–124. doi: 10.1016/S0092-8674(00)80611-X
Yavari, A., Javadi, M., Mirmiran, P., and Bahadoran, Z. (2015). Exercise-induced oxidative stress and dietary antioxidants. Asian J. Sports Med. 6:e24898. doi: 10.5812/asjsm.24898
Ziogas, V., Tanou, G., Molassiotis, A., Diamantidis, G., and Vasilakakis, M. (2010). Antioxidant and free radical-scavenging activities of phenolic extracts of olive fruits. Food Chem. 120, 1097–1103. doi: 10.1016/j.foodchem.2009.11.058
Zuo, L., Best, T. M., Roberts, W. J., Diaz, P. T., and Wagner, P. D. (2015a). Characterization of reactive oxygen species in diaphragm. Acta Physiol. (Oxf.) 213, 700–710. doi: 10.1111/apha.12410
Zuo, L., Christofi, F. L., Wright, V. P., Bao, S., and Clanton, T. L. (2004). Lipoxygenase-dependent superoxide release in skeletal muscle. J. Appl. Physiol. (1985) 97, 661–668. doi: 10.1152/japplphysiol.00096.2004
Zuo, L., Hallman, A. H., Roberts, W. J., Wagner, P. D., and Hogan, M. C. (2014). Superoxide release from contracting skeletal muscle in pulmonary TNF-alpha overexpression mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 306, R75–R81. doi: 10.1152/ajpregu.00425.2013
Zuo, L., Hallman, A. H., Yousif, M. K., and Chien, M. T. (2012). Oxidative stress, respiratory muscle dysfunction, and potential therapeutics in chronic obstructive pulmonary disease. Front. Biol. 7:506. doi: 10.1007/s11515-012-1251-x
Zuo, L., Nogueira, L., and Hogan, M. C. (2011a). Reactive oxygen species formation during tetanic contractions in single isolated Xenopus myofibers. J. Appl. Physiol. (1985) 111, 898–904. doi: 10.1152/japplphysiol.00398
Zuo, L., Roberts, W. J., Tolomello, R. C., and Goins, A. T. (2013). Ischemic and hypoxic preconditioning protect cardiac muscles via intracellular ROS signaling. Front. Biol. 8:305. doi: 10.1007/s11515-012-1225-z
Zuo, L., Youtz, D. J., and Wold, L. E. (2011b). Particulate matter exposure exacerbates high glucose-induced cardiomyocyte dysfunction through ROS generation. PLoS ONE 6:e23116. doi: 10.1371/journal.pone.0023116
Keywords: dietary antioxidant, exercise, exercise-induced adaptation, ROS, skeletal muscle
Citation: He F, Li J, Liu Z, Chuang C-C, Yang W and Zuo L (2016) Redox Mechanism of Reactive Oxygen Species in Exercise. Front. Physiol. 7:486. doi: 10.3389/fphys.2016.00486
Received: 19 April 2016; Accepted: 10 October 2016;
Published: 07 November 2016.
Edited by:
Vincent Pialoux, Claude Bernard University Lyon 1, FranceReviewed by:
Camille Faes, University of North Carolina, Chapel Hill, USACopyright © 2016 He, Li, Liu, Chuang, Yang and Zuo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Li Zuo, enVvLjRAb3N1LmVkdQ==
†Co-first authors.
‡These authors have contributed equally to this work.
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.