 Dmitry Tsvetkov
Dmitry Tsvetkov Jean-Yves Tano
Jean-Yves Tano Mario Kassmann
Mario Kassmann Ning Wang
Ning Wang Rudolf Schubert
Rudolf Schubert Maik Gollasch
Maik Gollasch
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Physiol. , 04 August 2016
Sec. Vascular Physiology
Volume 7 - 2016 | https://doi.org/10.3389/fphys.2016.00335
The anti-contractile effect of perivascular adipose tissue (PVAT) is an important mechanism in the modulation of vascular tone in peripheral arteries. Recent evidence has implicated the XE991-sensitive voltage-gated KV (KCNQ) channels in the regulation of arterial tone by PVAT. However, until now the in vivo pharmacology of the involved vascular KV channels with regard to XE991 remains undetermined, since XE991 effects may involve Ca2+ activated BKCa channels and/or voltage-dependent KV1.5 channels sensitive to diphenyl phosphine oxide-1 (DPO-1). In this study, we tested whether KV1.5 channels are involved in the control of mesenteric arterial tone and its regulation by PVAT. Our study was also aimed at extending our current knowledge on the in situ vascular pharmacology of DPO-1 and XE991 regarding KV1.5 and BKCa channels, in helping to identify the nature of K+ channels that could contribute to PVAT-mediated relaxation. XE991 at 30 μM reduced the anti-contractile response of PVAT, but had no effects on vasocontraction induced by phenylephrine (PE) in the absence of PVAT. Similar effects were observed for XE991 at 0.3 μM, which is known to almost completely inhibit mesenteric artery VSMC KV currents. 30 μM XE991 did not affect BKCa currents in VSMCs. Kcna5−/− arteries and wild-type arteries incubated with 1 μM DPO-1 showed normal vasocontractions in response to PE in the presence and absence of PVAT. KV current density and inhibition by 30 μM XE991 were normal in mesenteric artery VSMCs isolated from Kcna5−/− mice. We conclude that KV channels are involved in the control of arterial vascular tone by PVAT. These channels are present in VSMCs and very potently inhibited by the KCNQ channel blocker XE991. BKCa channels and/or DPO-1 sensitive KV1.5 channels in VSMCs are not the downstream mediators of the XE991 effects on PVAT-dependent arterial vasorelaxation. Further studies will need to be undertaken to examine the role of other KV channels in the phenomenon.
Over the past decade, various potassium (K+) channels have been implicated as important players in the regulation of arterial vascular tone and its control by perivascular adipose tissue (PVAT). Opening of vascular smooth muscle cell (VSMC) K+ channels causes K+ efflux and membrane hyperpolarization, which leads to reduced Ca2+ influx though L-type CaV1.2 channels and consequently arterial relaxation (Nelson and Quayle, 1995). A variety of endogenous vasodilators, such as hypoxia, acidosis, as well as metabolites and autacoids (e.g., adenosine, prostacyclin) act as potent K+ channel openers to produce relaxation (Sobey, 2001; Tano and Gollasch, 2014). Noteworthy, many of these substances produce relaxation by opening maxi Ca2+ activated (BKCa) K+ channels in VSMCs (Bentzen et al., 2014). Only very few substances have been reported to relax vessels by opening arterial smooth muscle voltage-gated KV channels (Tanaka et al., 2006; Park et al., 2015). Among them adenosine and atrial natriuretic peptide (ANP) act via activation of the KCNQ (KV7) subfamily of KV channels (Khanamiri et al., 2013; Stott et al., 2015).
Recent studies have demonstrated a paracrine role for PVAT to produce relaxation of arterial smooth muscle cells in a number of vascular beds (Lohn et al., 2002; Verlohren et al., 2004; Gao et al., 2005; Zavaritskaya et al., 2013). Certain adipokines, such as adiponectin (Weston et al., 2013), angiotensin-1 to 7 (Lee R. M. K. W. et al., 2011), methyl palmitate (Lee Y.-C. et al., 2011), and notably H2S (Schleifenbaum et al., 2010) were recently proposed as potential perivascular-derived relaxing factors (PVRFs), which could mediate the anti-contractile properties of PVAT. The paracrine effects of PVAT involve the opening of K+ channels, however, the identity of the K+ channel subtype(s) involved is still a matter of debate (Tano et al., 2014).
Voltage-gated KV channels of the KCNQ (KV7) family have been proposed to play an important role in PVAT control of arterial tone. This conclusion is based on observations demonstrating that the anti-contractile effects of PVAT are inhibited by the pan KCNQ channel blocker XE991 at 30 μM or the pan KV channel blocker 4-aminopyridine (2 mmol/L; Fésüs et al., 2007; Schleifenbaum et al., 2010; Lee Y.-C. et al., 2011; Zavaritskaya et al., 2013). XE991 is a widely used pan KV7 channel blocker, which inhibits KV7.1 homomeric or KV7.1/KCNE channels (IC50 of ~0.8 μM and 11.1 μM, respectively; Wang et al., 2000), KCNQ2/3 channels (EC50 ~1 μM; Wang et al., 1998), KCNQ4 (EC50 ~5.5 μM; Søgaard et al., 2001), and KCNQ5 (EC50 ~65 μM; Schroeder et al., 2000). Noteworthy, XE991 can also inhibit other KV channels, such as ERG (KV11; EC50 ~110 μM) (Elmedyb et al., 2007) and KV1.2/1.5, KV2.1/KV9.3 channels (~30% inhibition at 10 μM) in heterologous expression systems (Zhong et al., 2010).
However, it is unknown whether XE991 is indeed specific for vascular KV channels in situ, and does not inhibit native BKCa channels. This is particularly relevant since BKCa channels have been proposed to play a role in PVAT control of arterial tone in other studies (Lynch et al., 2013; Weston et al., 2013), although studies using BKCa deficient mice gave opposing results (Fésüs et al., 2007). A recent study showed that KV channels in VSMCs of mouse mesenteric arteries are very sensitive to XE991 (EC50~60 nM), suggesting that these channels may contribute to PVAT control of arterial tone (Schleifenbaum et al., 2010, 2014). A very recent study suggested that diphenyl phosphine oxide-1 (DPO-1) sensitive KV1.5 channels could contribute to the KV current in VSMC (Fancher et al., 2015).
Therefore, we tested whether KV1.5 channels are involved in the control of arterial tone and its regulation by PVAT or not. Our study is also aimed at extending our current knowledge on the in situ vascular pharmacology of DPO-1 and XE991 regarding KV1.5 and BKCa channels, in helping to identify the nature of K+ channels that could contribute to PVAT-mediated relaxation.
We used Kcna5−/− mice as previously described (Pannasch et al., 2006). The mouse model was evaluated by RT-qPCR (Figure S1). Either litter- or age-matched (10–14 weeks old) male wild-type (129S6 background, previously known as 129SvEv-Ta) mice were used as controls. 250–300 g male Sprague Dawley rats were obtained from Charles River, Germany, Berlin. All experimental procedures were performed in accordance with the German legislation on protection of animals. Animal care followed American Physiological Society guidelines, and local authorities (Landesamt für Gesundheit und Soziales Berlin, LAGeSo) approved all protocols. Mice were housed in individually ventilated cages under standardized conditions with an artificial 12-h dark–light cycle with free access to water and food.
First order mesenteric arteries were removed immediately after killing the mice or rats under inhalation anesthesia with isoflurane by cervical dislocation, quickly transferred to cold (4°C), oxygenated (95% O2/5% CO2) physiological salt solution (PSS) containing (in mmol/L) 119 NaCl, 4.7 KCl, 1.2 KH2PO4, 25 NaHCO3, 1.2 MgSO4, 11.1 glucose, 1.6 CaCl2), and dissected into 2 mm rings whereby perivascular fat and connective tissue were either intact [(+) PVAT or removed (−) PVAT] without damaging the adventitia. Each ring was positioned on two stainless steel wires (diameter 0.0394 mm) in a 5-ml organ bath of a Mulvany Small Vessel Myograph (DMT 610 M; Danish Myo Technology, Denmark). The organ bath was filled with PSS. The bath solution was continuously oxygenated with a gas mixture of 95% O2 and 5% CO2, and kept at 37°C (pH 7.4) (Verlohren et al., 2004; Fésüs et al., 2007). The mesenteric rings were placed under a tension equivalent to that generated at 0.9 times the diameter of the vessel at 100 mm Hg by stepwise distending the vessel using LabChart DMT Normalization module. This normalization procedure was performed to obtain the passive diameter of the vessel at 100 mm Hg (Fésüs et al., 2007). The software Chart5 (AD Instruments Ltd. Spechbach, Germany) was used for data acquisition and display. After 60 min equilibration arteries were pre-contracted either with isotonic external 60 mmol/L KCl until a stable resting tension was acquired. The composition of 60 mM KCl (in mmol/L) was 63.7 NaCl, 60 KCl, 1.2 KH2PO4, 25 NaHCO3, 1.2 Mg2SO4, 11.1 glucose, and 1.6 CaCl2. Drugs were added to the bath solution if not indicated otherwise. Tension is expressed as a percentage of the steady-state tension (100%) obtained with isotonic external 60 mM KCl.
VSMCs from mesenteric arteries were isolated as described (Gollasch et al., 1998; Plüger et al., 2000). Briefly, the arteries were isolated and quickly transferred to cold (4°C) oxygenated (95% O2–5% CO2) PSS. The arteries were cleaned, cut into pieces, and placed into a Ca2+-free Hank's solution (in mmol/L): 55 NaCl, 80 sodium glutamate, 5.6 KCl, 2 MgCl2, 1 mg/ml bovine serum albumin (BSA, Sigma, Taufkirchen), 10 glucose, and 10 HEPES (pH 7.4 with NaOH) containing 0.5 mg/ml papain (Sigma) and 1.0 mg/ml DTT for 50 min at 37°C. The segments then were placed in Hank's solution containing 1 mg/ml collagenase (Sigma, type F and H, ratio 30 and 70%, respectively) and 0.1 mmol/L CaCl2 for 10 min at 37°C. Following several washes in Ca2+-free Hank's solution (containing 1 mg/ml BSA), single cells were dispersed from artery segments by gentle triturating. Cells were then stored in the same solution at 4°C.
Voltage dependent potassium (KV) currents and BKCa currents were measured in the conventional whole-cell configuration of the patch-clamp technique at room temperature as previously described (Gollasch et al., 1996; Essin et al., 2007; Schleifenbaum et al., 2014). Patch pipettes (resistance 3–5 MΩ) for recording KV currents were filled with a solution containing (in mmol/L): 130 KCl, 1 MgCl2, 3 Na2ATP, 0.1 Na3GTP, 10 HEPES, and 5 EGTA (pH 7.2; Yeung and Greenwood, 2005). Patch pipettes for recording BKCa currents contained (in mmol/L): 130 KCl, 1 MgCl2, 3 Na2ATP, 0.1 Na3GTP, 10 HEPES, 5 EGTA, and 4.3 CaCl2 (estimated [Ca2+] free, 10−6 mol/L; pH 7.2). The external bath solution contained (in mmol/L): 126 NaCl, 5 KCl, 1 MgCl2, 0.1 CaCl2, 11 glucose and 10 HEPES (pH 7.2; Yeung and Greenwood, 2005). Holding potential was −60 mV. Whole cell currents were recorded using an Axopatch 200B amplifier (Axon Instruments/Molecular Devices, Sunnyvale, CA, USA) or an EPC 7 amplifier (List, Darmstadt, Germany) and digitized at 5 kHz, using a Digidata 1440A digitizer (Axon CNS, Molecular Devices), and pClamp software versions 10.1 and 10.2 (Schleifenbaum et al., 2014).
Total RNA was isolated from snap-frozen heart and aortae tissues with or without KV1.5 by using the RNeasy RNA isolation kit (Qiagen, Hamburg, Germany) according to the manufacturer's instruction. Isolated RNA concentration was measured and RNA quality was tested by NanoDrop-1000 spectrophotometer (PeqLab, Erlangen, Germany). For the synthesis of cDNA, equivalent amounts of RNA (2 μg) were used and processed by a high capacity cDNA reverse transcription kit (Life Technologies GmbH, Darmstadt, Germany). Quantitative analysis of target mRNA expression was performed with real-time PCR using the relative standard curve method (Markó et al., 2016). TaqMan or SYBR green analysis was conducted according to the manufacturer's instructions, using an Applied Biosystems 7500 Sequence Detector (Life Technologies Corporation, Carlsbad, CA, USA). The expression level of the target genes was normalized by the expression of 18S. Primers for were synthesized by Biotez (Berlin, Germany) and the sequences are as follows: KV1.5 Forward sequence: 5′-GCTACTTCGATCCCTTGAGAAAT-3′; Reverse sequence: AGTAGTACAAAATGCCATCGAAGCT, 18S Forward sequence: 5′-ACATCCAAGGAAGGCAGCAG-3′; Reverse sequence 5′-TTTTCGTCACTACCTCCCCG-3′.
All salts and other chemicals were obtained from Sigma-Aldrich (Germany) or Merck (Germany). All drugs were freshly dissolved on the day of each experiment according to the material sheet. The following concentrations of drugs were used: phenylephrine (Sigma-Aldrich) ranged from 0.01 to 100 μmol/L, 5-HT from 0.01 to 10 μM, DPO-1 (Tocris) 1 and 10 μmol/L, 100 nmol/L iberiotoxin (Sigma Aldrich). XE991 (Tocris) was applied at concentrations between 0.3 and 30 μM.
Data represent mean ± SEM. EC50 values were calculated using a Hill equation: T = (B0 − Be)/(1 + ([D]/EC50)n) + Be, where T is the tension in response to the drug (D); Be is the maximum response induced by the drug; B0 is a constant; EC50 is the concentration of the drug that elicits a half-maximal response (Bychkov et al., 1998). Curve fittings were done by Prism 6 software using non-linear regression. Statistical significance was determined by two-way ANOVA or repeated-measures two-way ANOVA, followed by Bonferroni post hoc test, and using Prism 6 software. In case of unbalanced data, this software uses analysis of “unweighted means” to compare groups. Extra sum-of-squares F-test was performed for comparison of concentration-response curves and their 95% confidence intervals (CI). P-values < 0.05 were considered statistically significant. n represents the number of independent arteries tested or the number of cells measured. All rings were obtained from at least 3 different animals.
First, we examined the role of KV1.5 channels in the regulation of arterial tone by alpha1 adrenoceptor (alpha1-AR) stimulation. In this set of experiments, we used the KV1.5 channel blocker DPO-1 at concentrations assumed to be specific and potent for KV1.5 channel inhibition (Stump et al., 2005; Lagrutta et al., 2006; Regan et al., 2006). In the presence of 1 μM DPO-1, mesenteric artery rings without PVAT [(−) PVAT] displayed similar contractions in response to phenylephrine (PE) compared to non-treated (−) PVAT control rings (Figures 1A,B). The 95% CI for EC50 of control and DPO-1 treated rings were 1.21–1.79 μM and 0.78–1.40 μM, respectively. The anti-contractile effects of PVAT were also unchanged by 1 μM DPO-1: the 95% CI for EC50 of control (+) PVAT and 1 μM DPO-1 (+) PVAT treated rings were 3.99–8.14 μM and 4.99–10.05 μM, respectively. To further confirm the results obtained with the KV1.5 channel inhibitor, we performed similar experiments using mesenteric artery rings from Kcna5−/− mice. PE induced vasocontractions in (−) PVAT Kcna5−/− rings were not different from those observed in (−) PVAT Kcna5+/+ rings (Figures 1C,D). Similarly, we observed PE induced vasocontractions in (+) PVAT Kcna5−/− rings, which were not different from those observed in (+) PVAT Kcna5+/+ rings (Figures 1C,D). The 95% CI for EC50 of (−) PVAT and (+) PVAT arteries isolated from Kcna5−/− mice were 0.81–1.30 μM and 4.94–7.79 μM, respectively. Data are summarized in Table 1. Experiments on rat mesenteric arteries showed similar results: Cumulative dose-response curves in response to serotonin (5-HT) were similar in vessel rings in the absence or presence of DPO-1 (Figure 1E). Together, the results suggest that KV1.5 channels do not play a functionally relevant role in the control of arterial tone by PVAT, α1-AR and 5-HT agonists, in both mouse and rat mesenteric arteries.
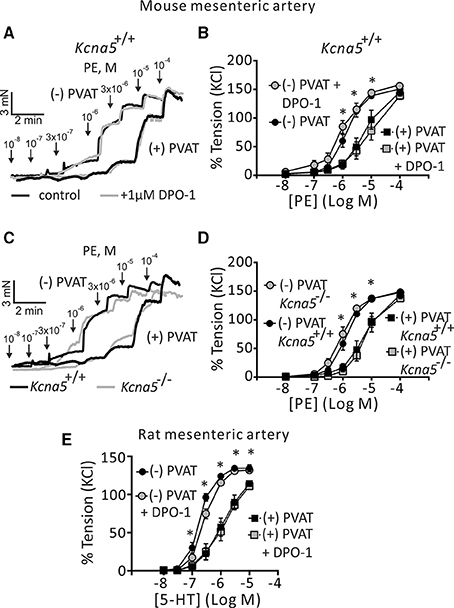
Figure 1. Effects of pharmacological blockade and genetic deletion of KV1.5 channels on regulation of arterial tone by perivascular adipose tissue (PVAT), phenylephrine (PE), and serotonin (5-HT). (A) Original traces showing the effects of 1 μM DPO-1 on PE-induced contractions in (−) PVAT and (+) PVAT mesenteric artery rings compared with control rings without DPO-1. (B) Concentration-response relationships for PE-induced contractions in Kcna5+/+ (+) PVAT (n = 8) or (−) PVAT (n = 9) mesenteric arteries in the absence of DPO-1 or in Kcna5+/+ (+) PVAT (n = 8) and (−) PVAT (n = 9) arteries after 30 min of pre-incubation with 1 μM DPO-1. *p < 0.05, for (−) PVAT vs. (+) PVAT or (−) PVAT + DPO-1 vs. (+) PVAT + DPO-1; repeated-measures two-way ANOVA, followed by Bonferroni post hoc test. (C) Original traces showing the effects of genetic deletion of Kcna5 on PE-induced contractions in (−) PVAT and (+) PVAT mesenteric artery rings. (D) PE-induced contractions of (+) PVAT and (−) PVAT artery rings isolated from Kcna5+/+ [(+) PVAT, n = 8; (−) PVAT, n = 9] and Kcna5−/− [(+) PVAT, n = 12; (−) PVAT, n = 12] mice. *p < 0.05, for (−) PVAT Kcna5+/+ vs. (+) PVAT Kcna5+/+ or (−) PVAT Kcna5−/− vs. (+) PVAT Kcna5−/−. (E) Cumulative concentration-response relationships to 5-HT in (+) PVAT (n = 7) and (−) PVAT rat mesenteric arteries (n = 6) and the effects of 1 μM DPO-1 on (+) PVAT (n = 6) and (−) PVAT rat mesenteric arteries (n = 8). *p < 0.05, for (−) PVAT vs. (+) PVAT or (−) PVAT + DPO-1 vs. (+) PVAT + DPO-1; repeated-measures two-way ANOVA, followed by Bonferroni post hoc test.
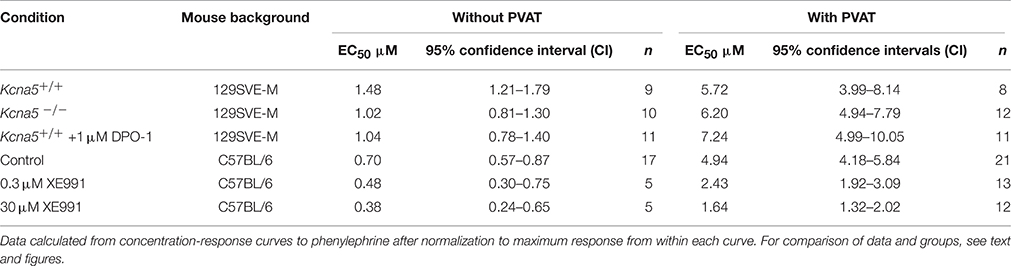
Table 1. EC50 and its confidence intervals.
Next, we studied putative non-KV1.5 channel dependent effects by using higher concentrations of DPO-1. Figure 2A shows that 1 μM DPO-1 had no effects on basal tone of Kcna5+/+ mesenteric artery rings with and without PVAT. Surprisingly, application of 10 μM DPO-1 resulted in a stable contraction of Kcna5+/+ mesenteric arteries without but not with PVAT (Figure 2B). This effect remained stable over 30 min and was observed also on rings isolated from Kcna5−/− mice (Figure 2C). Thus, unexpectedly, inhibition of DPO-1 sensitive KV channels distinct from KV1.5 channels or other pathways could contribute to vascular tone in this preparation.
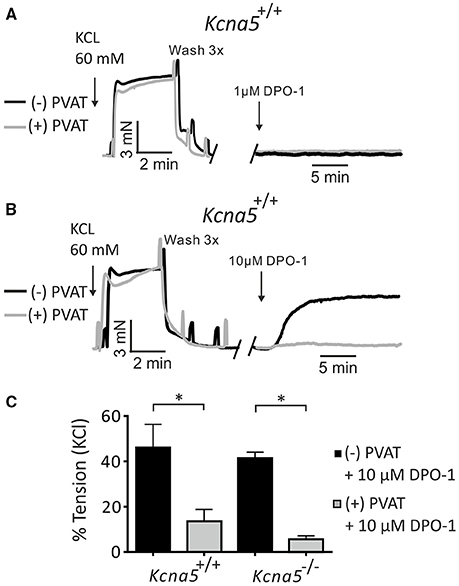
Figure 2. Effects of 1 μM and 10 μM DPO-1 on basal arterial tone of mesenteric artery rings isolated from Kcna5+/+ and Kcna5 −/− mice. Original traces showing contraction of mesenteric (−) and (+) PVAT rings induced by 60 mmol/L KCl and by 1 μM (A) and 10 μM DPO-1 (B). (C) Vessel tension induced by 10 μM DPO-1. Tension is expressed as a percentage of KCl contractions. Kcna5+/+ (−) PVAT, n = 9; (+) PVAT, n = 13. Kcna5−/− (−) PVAT, n = 7; (+) PVAT, n = 9. *p < 0.05, two-way ANOVA followed by Bonferroni post hoc test.
KV currents were recorded in mesenteric artery VSMCs freshly isolated from Kcna5+/+ and Kcna5−/− mice. We did not observe any difference between KV current densities in Kcna5+/+ and Kcna5−/− VSMCs. Moreover, KV current inhibition by 30 μM XE991 was not different between Kcna5+/+ and Kcna5−/− VSMCs (Figures 3A,B). 30 μM XE991 did not affect basal tone of mesenteric arteries prepared with or without PVAT (Figure S2).
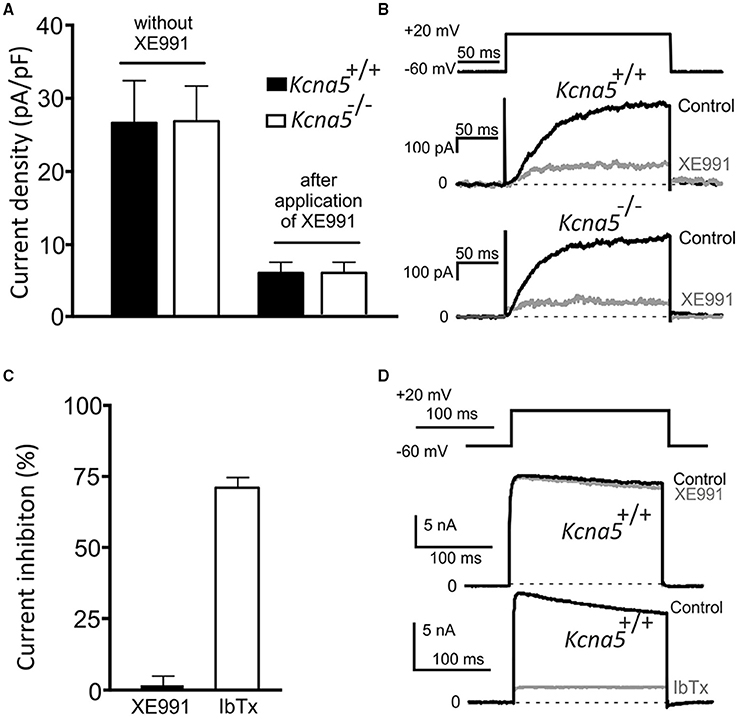
Figure 3. Voltage dependent (KV) K+ currents and BKCa currents in freshly-isolated mesenteric artery vascular smooth muscle cells (VSMCs) from Kcna5−/− and Kcna5+/+ mice. KV currents before (−XE991) and after (+XE991) application of 30 μM XE991. Current densities of peak KV currents (A) (n = 6 Kcna5−/− cells, n = 10 Kcna5+/+ cells, before and after application of XE991, respectively); original recordings (B). (C) Relative inhibition of peak BKCa current by iberiotoxion (IbTx) at 100 nM (n = 5) and XE991 at 30 μM (n = 9) in Kcna5+/+ VSMCs, original recordings are represented in (D).
In order to better understand the effects of XE991, we tested its actions on BKCa currents, potential mediators of the PVAT effect. VSMC Kcna5+/+ BKCa currents were recorded in the absence and presence of 30 μM XE991. 100 nM iberiotoxin (a potent and highly selective BKCa channel inhibitor) was used as positive control. While 30 μM XE991 did not affect the BKCa current, iberiotoxin almost completely inhibited the BKCa current. These results are consistent with plasma membrane VSMC BKCa channel activity resistant to XE991 in situ, at concentrations up to 30 μM of XE991 (Figures 3C,D).
Additionally, we tested the effects of 30 μM and 0.3 μM XE991 on the paracrine effects of PVAT on arterial tone. We found that XE991 even at the low concentration abolished the anti-contractile effects of PVAT. Interestingly, application of 0.3 and 30 μM XE991 resulted in a similar reduction of the anti-contractile effects of PVAT (Figures 4A,B). The EC50 95% CI values were 1.92–3.09 and 1.32–2.02 μM for (+) PVAT rings preincubated with 0.3 μM XE991 and 30 μM XE991, respectively; and 4.18–5.84 μM for (+) PVAT rings in the absence of XE991. (−) PVAT rings showed no difference, regardless of the absence or presence of 0.3 μM or 30 μM XE991. Data are presented in Table 1. Together, our data demonstrate that XE991 is a potent inhibitor of PVAT control of arterial tone at low concentrations similar to its potency of inhibiting KV currents in VSMCs (Schleifenbaum et al., 2014). BKCa channels are however exempt from this inhibitory effect of XE991 (up to 30 μM).
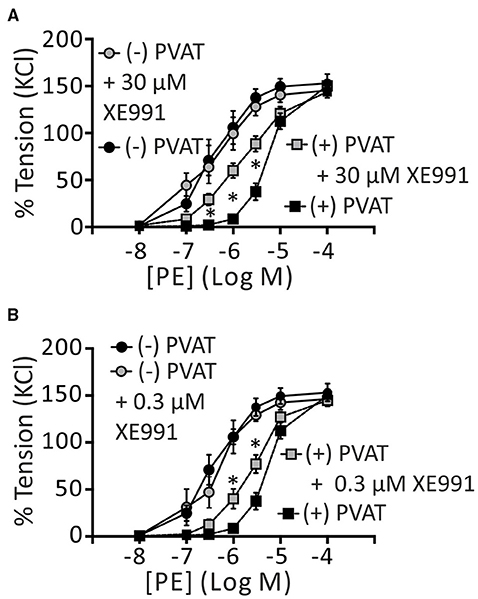
Figure 4. Effects of XE991 on regulation of arterial tone by perivascular adipose tissue. Cumulative concentration-response curves to PE in the presence of 30 μM XE991 (A) and 0.3 μM XE991 (B), expressed as a percentage of KCl contractions. (−) PVAT, n = 17; (+) PVAT, n = 21. (+) PVAT (+) 30 μM XE991, n = 12; (+) PVAT + 0.3 μM XE991, n = 13; (−) PVAT (+) 30 μM XE991, n = 5; (−) PVAT + 0.3 μM XE991, n = 5. *p < 0.05, repeated-measures two-way ANOVA, followed by Bonferroni post hoc test * in (A), (+) PVAT vs. (+) PVAT + 30 μM XE991. *In (B), (+) PVAT vs. (+) PVAT + 0.3 μM XE991. Experiments were performed on mouse mesenteric arteries.
Perivascular adipose tissue plays a potent anti-contractile role in the control of arterial tone along arterial segments of different vascular beds and species. The main findings of this study are threefold. First, XE991 inhibits the PVAT effect at nanomolar concentrations in mesenteric arteries of mice. Interestingly, similar concentrations were found in earlier studies to inhibit VSMC KV currents (50% inhibition at 60 nM XE991) (Schleifenbaum et al., 2014). Second, Kcna5−/− mice exhibited normal VSMC KV currents and arterial contractions in the absence and presence of PVAT, whose effects were insensitive to DPO-1. Third, the KCNQ channel blocker XE991 does not affect plasma membrane VSMC BKCa channels at concentrations, which inhibit the anti-contractile effects of PVAT. Together, the results of our current study implicate KCNQ-type KV channels in the XE991-mediated inhibition of the PVAT effects. Simultaneously, we exclude BKCa as well as KV1.5 channels as potential downstream candidates in this process.
In recent studies, KV1.5 channels have been shown to determine microvascular tone and the arteriolar response to vasoconstrictors in rat cerebral arteries (Chen et al., 2006; Fancher et al., 2015). Furthermore, KV1.5 channels in the heart are essential in coupling myocardial blood flow to cardiac metabolism (Ohanyan et al., 2015). Moreover, hypertension is associated with altered expression of vascular KV1.5 channels (Wang et al., 1997; Platoshyn et al., 2001; Cox and Rusch, 2002; Cox et al., 2008; Cidad et al., 2014). Therefore, KV1.5 channels may represent interesting putative targets of PVAT and that raised the question of their potential involvement in the regulation of arterial tone by phenylephrine in mouse mesenteric arteries. Our results suggest that KV1.5 channels are however not involved. In effect, the anti-contractile effects of PVAT were not different between Kcna5−/− and Kcna5+/+ arteries. Additionally, the KV1.5 channel inhibitor DPO-1 at 1 μM displayed no effect on vasocontractions in the absence and presence of PVAT. Notably, the mechanism of DPO-1 action (“open channel” blocker) might be a potential confounding factor, since KV1.5 channels are activated at V0.5 of −14 mV (Grissmer et al., 1994). However, the genetic approach had the advantage to study vascular effects in the absence of KV1.5 channels avoiding possible confounders related to membrane potential-dependent drug mechanisms. Furthermore, we did not observe any significant differences in the KV current density and inhibition by XE991 in Kcna5−/− and Kcna5+/+ VSMCs. Therefore, we conclude that KV1.5 channels have no apparent role in PVAT-dependent relaxation and are not the XE991 sensitive channels that contribute to this process. This conclusion is in line with our previous results obtained on cloned and heterologously expressed KV1.5 alpha subunits in HEK293 cells (Schleifenbaum et al., 2014). In these experiments, 100 nM XE991 failed to block KV1.5 currents. We also observed similar responses of arterial rings without PVAT to PE and 5-HT, regardless of genetic deletion of KV1.5 alpha subunits or pharmacological blockade of KV1.5 channels by DPO-1. To our knowledge, this is the first study to firmly establish that KV1.5 channels are not involved in the regulation of arterial tone of systemic visceral arteries of mice and rats, at least in mesenteric arteries. Our conclusions substantiate the work of other groups, namely that patients with genetic mutations of KCNA5 exhibit pulmonary arterial hypertension and arterial fibrillation but not systemic hypertension (Yang et al., 2009; Wipff et al., 2010; Machado et al., 2015). Together, our data questions the contribution of KV1.5 channels in a number of small resistance arteries to peripheral arterial resistance. The findings are however not generalizable to all vascular beds as KV1.5 channels were demonstrated to play important vasoregulatory functions in cerebral arteries and in Gracilis skeletal muscle arteries (Chen et al., 2006; Fancher et al., 2015).
DPO-1 was described as a specific KV1.5 inhibitor at micromolar concentrations (Stump et al., 2005; Lagrutta et al., 2006; Regan et al., 2006). It exerts its inhibitory effects through binding with several key residues in the S5- pore loop-S6 domains, thus resulting in blockade of the open state of the KV1.5 channel (Karczewski et al., 2009; Du et al., 2010). Other studies have suggested a DPO-1 preference for vascular KV1.5 channels, though with limited selectivity (Fancher et al., 2015). In this study, DPO-1 (1–10 μM) inhibited the outward K+ current in arterial smooth muscle cells from wild-type (Kcna5+/+) mice and mice lacking the Kcna5 gene; however, the inhibitory effect was much greater in cells from Kcna5+/+ mice (Fancher et al., 2015). Subsequently, our data in Figure 2 suggests that 10 μM DPO-1 induces contractions by inhibiting channels distinct from KV1.5 channels. Those channels appear to be important for the regulation of resting arterial tone. Interestingly, DPO-1 is able to block KV1.3 channel currents at EC50 of 3.1 μM in human T cells (Zhao et al., 2013). Kcna3 mRNA expression is also observed in mouse mesenteric arteries (Fountain et al., 2004; Cidad et al., 2014). Although the ability of KV1.5 and KV1.3 to form heteromers (Kv1.5/Kv1.3) (Villalonga et al., 2007) impede the study of their specific roles in native tissues in vivo, it is intriguing to speculate that KV1.3 channels could represent a putative target of PVAT regulation of arterial tone. Future studies are necessary to clarify their role.
In the mouse mesenteric arteries, Kcnq1, Kcnq4, and Kcnq5 expression was demonstrated at the mRNA level (Yeung et al., 2007), whereas mRNA expression of Kcnq2, Kcnq3 was not detectable or only at borderline low levels (Yeung et al., 2007; Schleifenbaum et al., 2014). We previously suggested that KCNQ type KV channels are key players in the paracrine role for periadventitial adipose tissue in the regulation of arterial tone (Schleifenbaum et al., 2010; Zavaritskaya et al., 2013); based on mRNA expression levels (see above), KCNQ1, KCNQ4, and/or KCNQ5 channels are likely candidates. This suggestion is also based on the ability of 30 μM XE991 (pan KCNQ blocker) and 2 mmol/L 4-aminopyridine (pan KV blocker) to block the anti-contractile effects of PVAT. Interestingly, KCNQ channel openers normalized reduced anti-contractile effects of PVAT in a rat model of hypertension (Zavaritskaya et al., 2013), which suggests therapeutic perspectives of KCNQ targeting in cardiovascular disease. It is thus imperative to better understand the actions of these compounds on the vasculature. In order to obtain more information about the potency of XE991 inhibition on PVAT-mediated anti-contractility, we performed experiments with a 100x lower concentration of XE991. The data show that this considerably reduced concentration still exerted an inhibitory impact on PVAT regulation of arterial tone (Figures 4A,B), while basal tone was unaffected in rings (+) or (−) PVAT (Figure S2). The incomplete inhibition observed, however suggests the possible involvement of additional K+ channels in this process. Furthermore, our data demonstrates that VSMC plasma membrane BKCa channels are not involved in the effects of XE991 on PVAT regulation, since XE991 does not inhibit BKCa currents (this study). This is in line with our previous findings indicating that the paracrine effects of PVAT on arterial tone are normal in the presence of BKCa channel blockers or in arteries that lack BKCa beta1 channel subunits (Fésüs et al., 2007; Zavaritskaya et al., 2013).
Previous studies examined the ability of XE991 to inhibit heterologously expressed KV1.5 alpha subunits. We found that 100 nM and 30 μM XE991 was unable to block monotetrameric KV1.5 channels heterologously expressed in HEK293 or CHO cells (Zavaritskaya et al., 2013; Schleifenbaum et al., 2014). Interestingly, Zhong et al. found a small (~20% at (+) 5 mV) inhibiting effect for 10 μM XE991 on heterologously expressed heterotetrameric KV1.2/KV1.5 and KV2.1/KV9.3 channel subunits (Zhong et al., 2010). The block of KV1.2/KV1.5 channels was voltage dependent, and evident only at voltages positive to -15 mV. Our present study contributes to the debate about the importance of accessory subunits for determining the pharmacological properties of vascular K+ channels in vivo. Since regulatory KVbeta1.3 subunits can decrease the sensitivity of KV1.5 channels to pharmacological inhibitors such as DPO-1 (Gonzalez et al., 2002; Arias et al., 2007; Du et al., 2010), one could argue that DPO-1 is not a reliable tool to study KV1.5 channels in native tissues. However, we believe that our pharmacological approach in combination with the Kcna5−/− mouse model firmly demonstrates that the XE991 sensitive regulation of arterial tone by PVAT regulation does not involve native vascular KV1.5 channels.
In conclusion, our results demonstrate that KV1.5 channels are not involved in the control of mesenteric arterial tone and its regulation by PVAT in mouse and rat mesenteric arteries. The nature of the 10 μM DPO-1 sensitive component is unclear, but is most likely related to non-specificity of this drug, for example in targeting vascular KV1.3 and/or KCNQ channels in situ. Importantly, the inhibitory effects of XE991 on PVAT vasorelaxation are rather related to inhibition of KCNQ-type KV channels than BKCa channels. These data unequivocally substantiate the hypothesis of different targets of perivascular relaxing factor(s), which employ distinct mechanisms to mediate an anti-contractile effect. Further studies should focus on the enhancement of these relaxing factors, as these will be beneficial for patients with cardiovascular diseases.
All authors planned and designed the experimental studies. DT and NW performed the wire myography experiments. MK and JT performed the electrophysiological experiments. DT and MG drafted the article, and all authors contributed to its completion.
This study was supported by grants from the Deutsche Forschungsgemeinschaft (DFG) to MG and the Deutsche Akademische Austauschdienst (DAAD) to MG and DT. DT is recipient of ERA/EDTA and DAAD fellowships and JT is an Alexander von Humboldt fellow.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank Dr. Helmut Kettenmann (MDC Berlin) for providing Kcna5−/− mice and Dr. Lajos Markó for helping with RT-qPCR.
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fphys.2016.00335
Arias, C., Guizy, M., David, M., Marzian, S., González, T., Decher, N., et al. (2007). Kvbeta1.3 reduces the degree of stereoselective bupivacaine block of Kv1.5 channels. Anesthesiology 107, 641–651. doi: 10.1097/01.anes.0000282100.32923.5c
Bentzen, B. H., Olesen, S. P., Ronn, L. C. B., and Grunnet, M. (2014). BK channel activators and their therapeutic perspectives. Front. Physiol. 5:389. doi: 10.3389/fphys.2014.00389
Bychkov, R., Gollasch, M., Steinke, T., Ried, C., Luft, F. C., and Haller, H. (1998). Calcium-activated potassium channels and nitrate-induced vasodilation in human coronary arteries. J. Pharmacol. Exp. Ther. 285, 293–298.
Chen, T. T., Luykenaar, K. D., Walsh, E. J., Walsh, M. P., and Cole, W. C. (2006). Key role of Kv1 channels in vasoregulation. Circ. Res. 99, 53–60. doi: 10.1161/01.RES.0000229654.45090.57
Cidad, P., Novensà, L., Garabito, M., Batlle, M., Dantas, A. P., Heras, M., et al. (2014). K(+) channels expression in hypertension after arterial injury, and effect of selective Kv1.3 blockade with PAP-1 on intimal hyperplasia formation. Cardiovasc. Drugs Ther. 28, 501–511. doi: 10.1007/s10557-014-6554-5
Cox, R. H., Fromme, S. J., Folander, K. L., and Swanson, R. J. (2008). Voltage gated K(+) channel expression in arteries of Wistar-Kyoto and spontaneously hypertensive rats. Am. J. Hypertens. 21, 213–218. doi: 10.1038/ajh.2007.44
Cox, R. H., and Rusch, N. J. (2002). New expression profiles of voltage-gated ion channels in arteries exposed to high blood pressure. Microcirculation 9, 243–257. doi: 10.1038/sj.mn.7800140
Du, Y., Zhang, X., Tu, D., Zhao, N., Liu, Y., Xiao, H., et al. (2010). Molecular determinants of Kv1.5 channel block by diphenyl phosphine oxide-1. J. Mol. Cell. Cardiol. 48, 1111–1120. doi: 10.1016/j.yjmcc.2010.02.010
Elmedyb, P., Calloe, K., Schmitt, N., Hansen, R. S., Grunnet, M., and Olesen, S.-P. (2007). Modulation of ERG channels by XE991. Basic Clin. Pharmacol. Toxicol. 100, 316–322. doi: 10.1111/j.1742-7843.2007.00048.x
Essin, K., Welling, A., Hofmann, F., Luft, F. C., Gollasch, M., and Moosmang, S. (2007). Indirect coupling between Cav1.2 channels and ryanodine receptors to generate Ca2+ sparks in murine arterial smooth muscle cells. J. Physiol. 584, 205–219. doi: 10.1113/jphysiol.2007.138982
Fancher, I. S., Butcher, J. T., Brooks, S. D., Rottgen, T. S., Skaff, P. R., Frisbee, J. C., et al. (2015). Diphenyl phosphine oxide-1-sensitive K+ channels contribute to the vascular tone and reactivity of resistance arteries from brain and skeletal muscle. Microcirculation 22, 315–325. doi: 10.1111/micc.12201
Fésüs, G., Dubrovska, G., Gorzelniak, K., Kluge, R., Huang, Y., Luft, F. C., et al. (2007). Adiponectin is a novel humoral vasodilator. Cardiovasc. Res. 75, 719–727. doi: 10.1016/j.cardiores.2007.05.025
Fountain, S. J., Cheong, A., Flemming, R., Mair, L., Sivaprasadarao, A., and Beech, D. J. (2004). Functional up-regulation of KCNA gene family expression in murine mesenteric resistance artery smooth muscle. J. Physiol. 556, 29–42. doi: 10.1113/jphysiol.2003.058594
Gao, Y.-J., Zeng, Z., Teoh, K., Sharma, A. M., Abouzahr, L., Cybulsky, I., et al. (2005). Perivascular adipose tissue modulates vascular function in the human internal thoracic artery. J. Thorac. Cardiovasc. Surg. 130, 1130–1136. doi: 10.1016/j.jtcvs.2005.05.028
Gollasch, M., Ried, C., Bychkov, R., Luft, F. C., and Haller, H. (1996). K+ currents in human coronary artery vascular smooth muscle cells. Circ. Res. 78, 676–688. doi: 10.1161/01.RES.78.4.676
Gollasch, M., Wellman, G. C., Knot, H. J., Jaggar, J. H., Damon, D. H., Bonev, A. D., et al. (1998). Ontogeny of local sarcoplasmic reticulum Ca2+ signals in cerebral arteries: Ca2+ sparks as elementary physiological events. Circ. Res. 83, 1104–1114.
Gonzalez, T., Navarro-Polanco, R., Arias, C., Caballero, R., Moreno, I., Delpon, E., et al. (2002). Assembly with the Kvbeta1.3 subunit modulates drug block of hKv1.5 channels. Mol. Pharmacol. 62, 1456–1463. doi: 10.1124/mol.62.6.1456
Grissmer, S., Nguyen, A. N., Aiyar, J., Hanson, D. C., Mather, R. J., Gutman, G. A., et al. (1994). Pharmacological characterization of five cloned voltage-gated K+ channels, types Kv1.1, 1.2, 1.3, 1.5, and 3.1, stably expressed in mammalian cell lines. Mol. Pharmacol. 45, 1227–1234.
Karczewski, J., Kiss, L., Kane, S. A., Koblan, K. S., Lynch, R. J., and Spencer, R. H. (2009). High-throughput analysis of drug binding interactions for the human cardiac channel, Kv1.5. Biochem. Pharmacol. 77, 177–185. doi: 10.1016/j.bcp.2008.09.035
Khanamiri, S., Soltysinska, E., Jepps, T. A., Bentzen, B. H., Chadha, P. S., Schmitt, N., et al. (2013). Contribution of Kv7 channels to basal coronary flow and active response to ischemia. Hypertension 62, 1090–1097. doi: 10.1161/HYPERTENSIONAHA.113.01244
Lagrutta, A., Wang, J., Fermini, B., and Salata, J. J. (2006). Novel, potent inhibitors of human Kv1.5 K+ channels and ultrarapidly activating delayed rectifier potassium current. J. Pharmacol. Exp. Ther. 317, 1054–1063. doi: 10.1124/jpet.106.101162
Lee, R. M. K. W., Bader, M., Alenina, N., Santos, R. A. S., Gao, Y.-J., and Lu, C. (2011). Mas receptors in modulating relaxation induced by perivascular adipose tissue. Life Sci. 89, 467–472. doi: 10.1016/j.lfs.2011.07.016
Lee, Y.-C., Chang, H.-H., Chiang, C.-L., Liu, C.-H., Yeh, J.-I., Chen, M.-F., et al. (2011). Role of perivascular adipose tissue-derived methyl palmitate in vascular tone regulation and pathogenesis of hypertension. Circulation 124, 1160–1171. doi: 10.1161/CIRCULATIONAHA.111.027375
Lohn, M., Dubrovska, G., Lauterbach, B., Luft, F. C., Gollasch, M., and Sharma, A. M. (2002). Periadventitial fat releases a vascular relaxing factor. FASEB J. 16, 1057–1063. doi: 10.1096/fj.02-0024com
Lynch, F. M., Withers, S. B., Yao, Z., Werner, M. E., Edwards, G., Weston, A. H., et al. (2013). Perivascular adipose tissue-derived adiponectin activates BK(Ca) channels to induce anticontractile responses. Am. J. Physiol. Hear. Circ. Physiol. 304, H786–H795. doi: 10.1152/ajpheart.00697.2012
Machado, R. D., Southgate, L., Eichstaedt, C. A., Aldred, M. A., Austin, E. D., Best, D. H., et al. (2015). Pulmonary arterial hypertension: a current perspective on established and emerging molecular genetic defects. Hum. Mutat. 36, 1113–1127. doi: 10.1002/humu.22904
Markó, L., Vigolo, E., Hinze, C., Park, J.-K., Roël, G., Balogh, A., et al. (2016). Tubular epithelial NF-κB activity regulates ischemic AKI. J. Am. Soc. Nephrol. doi: 10.1681/ASN.2015070748. [Epub ahead of print].
Nelson, M. T., and Quayle, J. M. (1995). Physiological roles and properties of potassium channels in arterial smooth muscle. Am. J. Physiol. 268, C799–C822.
Ohanyan, V., Yin, L., Bardakjian, R., Kolz, C., Enrick, M., Hakobyan, T., et al. (2015). Requisite role of Kv1.5 channels in coronary metabolic dilation. Circ. Res. 117, 612–621. doi: 10.1161/CIRCRESAHA.115.306642
Pannasch, U., Färber, K., Nolte, C., Blonski, M., Yan Chiu, S., Messing, A., et al. (2006). The potassium channels Kv1.5 and Kv1.3 modulate distinct functions of microglia. Mol. Cell. Neurosci. 33, 401–411. doi: 10.1016/j.mcn.2006.08.009
Park, S. W., Noh, H. J., Sung, D. J., Kim, J. G., Kim, J. M., Ryu, S.-Y., et al. (2015). Hydrogen peroxide induces vasorelaxation by enhancing 4-aminopyridine-sensitive Kv currents through S-glutathionylation. Pflügers Arch. Eur. J. Physiol. 467, 285–297. doi: 10.1007/s00424-014-1513-3
Platoshyn, O., Yu, Y., Golovina, V. A., McDaniel, S. S., Krick, S., Li, L., et al. (2001). Chronic hypoxia decreases K(V) channel expression and function in pulmonary artery myocytes. Am. J. Physiol. Lung. Cell Mol. Physiol. 280, L801–L812.
Plüger, S., Faulhaber, J., Fürstenau, M., Löhn, M., Waldschütz, R., Gollasch, M., et al. (2000). Mice with disrupted BK channel beta1 subunit gene feature abnormal Ca(2+) spark/STOC coupling and elevated blood pressure. Circ. Res. 87, E53–60. doi: 10.1161/01.RES.87.11.e53
Regan, C. P., Wallace, A. A., Cresswell, H. K., Atkins, C. L., and Lynch, Jr., J. J. (2006). In vivo cardiac electrophysiologic effects of a novel diphenylphosphine oxide IKur blocker, (2-Isopropyl-5-methylcyclohexyl) diphenylphosphine oxide, in rat and nonhuman primate. J. Pharmacol. Exp. Ther. 316, 727–732. doi: 10.1124/jpet.105.092197
Schleifenbaum, J., Kassmann, M., Szijártó, I. A., Hercule, H. C., Tano, J.-Y., Weinert, S., et al. (2014). Stretch-activation of angiotensin II type 1a receptors contributes to the myogenic response of mouse mesenteric and renal arteries. Circ. Res. 115, 263–272. doi: 10.1161/CIRCRESAHA.115.302882
Schleifenbaum, J., Köhn, C., Voblova, N., Dubrovska, G., Zavarirskaya, O., Gloe, T., et al. (2010). Systemic peripheral artery relaxation by KCNQ channel openers and hydrogen sulfide. J. Hypertens. 28, 1875–1882. doi: 10.1097/HJH.0b013e32833c20d5
Schroeder, B. C., Hechenberger, M., Weinreich, F., Kubisch, C., and Jentsch, T. J. (2000). KCNQ5, a novel potassium channel broadly expressed in brain, mediates m-type currents. J. Biol. Chem. 275, 24089–24095. doi: 10.1074/jbc.M003245200
Sobey, C. G. (2001). Potassium channel function in vascular disease. Arterioscler. Thromb. Vasc. Biol. 21, 28–38. doi: 10.1161/01.ATV.21.1.28
Søgaard, R., Ljungstrøm, T., Pedersen, K. A., Olesen, S. P., and Jensen, B. S. (2001). KCNQ4 channels expressed in mammalian cells: functional characteristics and pharmacology. Am. J. Physiol. Cell Physiol. 280, C859–C866.
Stott, J. B., Barrese, V., Jepps, T. A., Leighton, E. V., and Greenwood, I. A. (2015). Contribution of Kv7 channels to natriuretic peptide mediated vasodilation in normal and hypertensive rats. Hypertension 65, 676–682. doi: 10.1161/HYPERTENSIONAHA.114.04373
Stump, G. L., Wallace, A. A., Regan, C. P., and Lynch, J. J. (2005). In vivo antiarrhythmic and cardiac electrophysiologic effects of a novel diphenylphosphine oxide IKur blocker (2-isopropyl-5-methylcyclohexyl) diphenylphosphine oxide. J. Pharmacol. Exp. Ther. 315, 1362–1367. doi: 10.1124/jpet.105.092197
Tanaka, Y., Tang, G., Takizawa, K., Otsuka, K., Eghbali, M., Song, M., et al. (2006). Kv channels contribute to nitric oxide- and atrial natriuretic peptide-induced relaxation of a rat conduit artery. J. Pharmacol. Exp. Ther. 317, 341–354. doi: 10.1124/jpet.105.096115
Tano, J.-Y., and Gollasch, M. (2014). Calcium-activated potassium channels in ischemia reperfusion: a brief update. Front. Physiol. 5:381. doi: 10.3389/fphys.2014.00381
Tano, J. Y., Schleifenbaum, J., and Gollasch, M. (2014). Perivascular adipose tissue, potassium channels, and vascular dysfunction. Arterioscler. Thromb. Vasc. Biol. 34, 1827–1830. doi: 10.1161/ATVBAHA.114.303032
Verlohren, S., Dubrovska, G., Tsang, S. Y., Essin, K., Luft, F. C., Huang, Y., et al. (2004). Visceral periadventitial adipose tissue regulates arterial tone of mesenteric arteries. Hypertension 44, 271–276. doi: 10.1161/01.HYP.0000140058.28994.ec
Villalonga, N., Escalada, A., Vicente, R., Sánchez-Tilló, E., Celada, A., Solsona, C., et al. (2007). Kv1.3/Kv1.5 heteromeric channels compromise pharmacological responses in macrophages. Biochem. Biophys. Res. Commun. 352, 913–918. doi: 10.1016/j.bbrc.2006.11.120
Wang, H. S., Brown, B. S., McKinnon, D., and Cohen, I. S. (2000). Molecular basis for differential sensitivity of KCNQ and I(Ks) channels to the cognitive enhancer XE991. Mol. Pharmacol. 57, 1218–1223.
Wang, H. S., Pan, Z., Shi, W., Brown, B. S., Wymore, R. S., Cohen, I. S., et al. (1998). KCNQ2 and KCNQ3 potassium channel subunits: molecular correlates of the M-channel. Science 282, 1890–1893. doi: 10.1126/science.282.5395.1890
Wang, J., Juhaszova, M., Rubin, L. J., and Yuan, X. J. (1997). Hypoxia inhibits gene expression of voltage-gated K+ channel alpha subunits in pulmonary artery smooth muscle cells. J. Clin. Invest. 100, 2347–2353. doi: 10.1172/JCI119774
Weston, A. H., Egner, I., Dong, Y., Porter, E. L., Heagerty, A. M., and Edwards, G. (2013). Stimulated release of a hyperpolarizing factor (ADHF) from mesenteric artery perivascular adipose tissue: involvement of myocyte BKCa channels and adiponectin. Br. J. Pharmacol. 169, 1500–1509. doi: 10.1111/bph.12157
Wipff, J., Dieudé, P., Guedj, M., Ruiz, B., Riemekasten, G., Cracowski, J. L., et al. (2010). Association of a KCNA5 gene polymorphism with systemic sclerosis-associated pulmonary arterial hypertension in the European Caucasian population. Arthritis Rheum. 62, 3093–3100. doi: 10.1002/art.27607
Yang, Y., Li, J., Lin, X., Yang, Y., Hong, K., Wang, L., et al. (2009). Novel KCNA5 loss-of-function mutations responsible for atrial fibrillation. J. Hum. Genet. 54, 277–283. doi: 10.1038/jhg.2009.26
Yeung, S. Y. M., and Greenwood, I. A. (2005). Electrophysiological and functional effects of the KCNQ channel blocker XE991 on murine portal vein smooth muscle cells. Br. J. Pharmacol. 146, 585–595. doi: 10.1038/sj.bjp.0706342
Yeung, S. Y. M., Pucovskı, V., Moffatt, J. D., Saldanha, L., Schwake, M., Ohya, S., et al. (2007). Molecular expression and pharmacological identification of a role for K(v)7 channels in murine vascular reactivity. Br. J. Pharmacol. 151, 758–770. doi: 10.1038/sj.bjp.0707284
Zavaritskaya, O., Zhuravleva, N., Schleifenbaum, J., Gloe, T., Devermann, L., Kluge, R., et al. (2013). Role of KCNQ channels in skeletal muscle arteries and periadventitial vascular dysfunction. Hypertension 61, 151–159. doi: 10.1161/HYPERTENSIONAHA.112.197566
Zhao, N., Dong, Q., Du, L.-L., Fu, X.-X., Du, Y.-M., and Liao, Y.-H. (2013). Potent suppression of Kv1.3 potassium channel and IL-2 secretion by diphenyl phosphine oxide-1 in human T cells. PLoS ONE 8:e64629. doi: 10.1371/journal.pone.0064629
Keywords: XE991, KCNQ channels, KV1.5 channels, adipocyte-derived relaxing factor (ADRF), perivascular adipose tissue (PVAT), BK channels
Citation: Tsvetkov D, Tano J-Y, Kassmann M, Wang N, Schubert R and Gollasch M (2016) The Role of DPO-1 and XE991-Sensitive Potassium Channels in Perivascular Adipose Tissue-Mediated Regulation of Vascular Tone. Front. Physiol. 7:335. doi: 10.3389/fphys.2016.00335
Received: 21 April 2016; Accepted: 20 July 2016;
Published: 04 August 2016.
Edited by:
Iain A. Greenwood, St George's, University of London, UKReviewed by:
Sergey V. Smirnov, University of Bath, UKCopyright © 2016 Tsvetkov, Tano, Kassmann, Wang, Schubert and Gollasch. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Maik Gollasch, bWFpay5nb2xsYXNjaEBjaGFyaXRlLmRl
†These authors have contributed equally to this work.
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.