 Zhenzhen Liu
Zhenzhen Liu Paul A. Roche
Paul A. Roche
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Physiol., 30 January 2015
Sec. Membrane Physiology and Membrane Biophysics
Volume 6 - 2015 | https://doi.org/10.3389/fphys.2015.00001
This article is part of the Research TopicPhysiological roles and regulatory mechanisms of macropinocytosis-mediated membrane retrieval in health and diseaseView all 8 articles
Dendritic cells (DCs) are outstanding antigen presenting cells (APCs) due to their robust ability to internalize extracellular antigens using endocytic processes such as receptor-mediated endocytosis, phagocytosis, and macropinocytosis. Macropinocytosis mediates the non-specific uptake of soluble antigens and occurs in DCs constitutively. Macropinocytosis plays a key role in DC-mediated antigen presentation to T cells against pathogens and the efficiency of macropinocytosis in antigen capture is regulated during the process of DC maturation. Here, we review the methods to study macropinocytosis, describe our current knowledge of the regulatory mechanisms of antigen uptake via macropinocytosis and the intracellular trafficking route followed by macropinocytosed antigens, and discuss the significance of macropinocytosis for DC function.
Dendritic cells (DCs) are the most accomplished of professional antigen presenting cells (APCs) and are responsible for initiating T cell responses against pathogens (Villadangos and Schnorrer, 2007). DCs are present throughout the body and function by sampling their microenvironment to “sense” pathogens. DCs express peptide fragments of degraded foreign antigens on their surface bound to MHC glycoproteins, molecules that function as “display platforms” presenting peptide-fragments for recognition by antigen-specific T cells (Blum et al., 2013). Peptide-MHC class I complexes (pMHC-I) are recognized by cytotoxic CD8+ T cells whereas peptide-MHC class II complexes (pMHC-II) are recognized by helper CD4+ T cells. Besides stimulating distinct classes of T cells, it is generally true that MHC-I and MHC-II differ in the sites of intracellular antigenic peptide generation. MHC-I-destined peptides are generated by cytosolic proteolysis of endogenous proteins and foreign (often viral) antigens, and these peptides are translocated into the endoplasmic reticulum for binding to nascent MHC-I. In specialized DC subsets, however, these peptides can be translocated directly into phagosomes for binding to MHC-I in a process termed cross-presentation (Joffre et al., 2012). Cross-presentation can also result from direct endo/lysosomal proteolysis of internalized antigens (Shen et al., 2004; Kreer et al., 2011), however this review will not address the endocytic mechanisms responsible for cross-presentation as this has been reviewed recently (Schuette and Burgdorf, 2014). By marked contrast with MHC-I, MHC-II-destined peptides are often generated by endo/lysosomal proteolysis of internalized pathogens. DCs generally recognize and capture foreign invaders in non-lymphoid organs. Upon pathogen capture, DCs concurrently mature and migrate to secondary lymphoid organs (such as lymph nodes) where their surface pMHC-I and pMHC-II present antigenic peptides to T cells to initiate antigen-specific adaptive immune responses.
One feature that makes DCs particularly effective stimulators of CD4+ T cell responses is their considerable capacity to acquire exogenous antigens/pathogens by a variety of different endocytic processes (Figure 1). Receptor-mediated endocytosis requires antigen (usually small soluble molecules) binding to a variety of receptors on DCs (most notably lectins, Fc receptors, and complement receptors) that are internalized in clathrin-coated and non-clathrin-coated vesicles. Phagocytosis is a form of endocytosis that mediates the internalization of a wide-variety of relatively-large insoluble particulate antigens including necrotic/apoptotic cells and opsonized pathogenic organisms/viruses (Stuart and Ezekowitz, 2005). Like receptor-mediated endocytosis, phagocytosis is usually initiated by antigen binding to DC surface “receptors,” however, DCs can also internalize large particles (such as antigen-coated latex beads) by non-specific phagocytosis. Lastly, macropinocytosis (Lim and Gleeson, 2011), or “cell drinking,” is the non-specific uptake of soluble molecules, nutrients and antigens. By contrast, phagocytosis and receptor-mediated endocytosis mediate specific uptake of extracellular materials via a number of distinct surface receptors (Burgdorf and Kurts, 2008). Macropinocytosis is an actin-dependent process initiated from plasma membrane ruffles that give rise to large endocytic vesicles called macropinosomes, which are distinct from other forms of endocytic vesicles. Unlike small (0.1 μm) clathrin-coated vesicles, macropinosomes have no apparent coat structure and are heterogeneous in size ranging from 0.2 to 5 μm in diameter (Hewlett et al., 1994; Swanson and Watts, 1995). Macropinocytosis occurs constitutively in DCs and DCs generated in vitro from mouse bone marrow (BMDC) can internalize approximately 2 fL/cell/min (Norbury et al., 1997), thereby providing these cells with a robust mechanism of non-specific foreign antigen uptake.
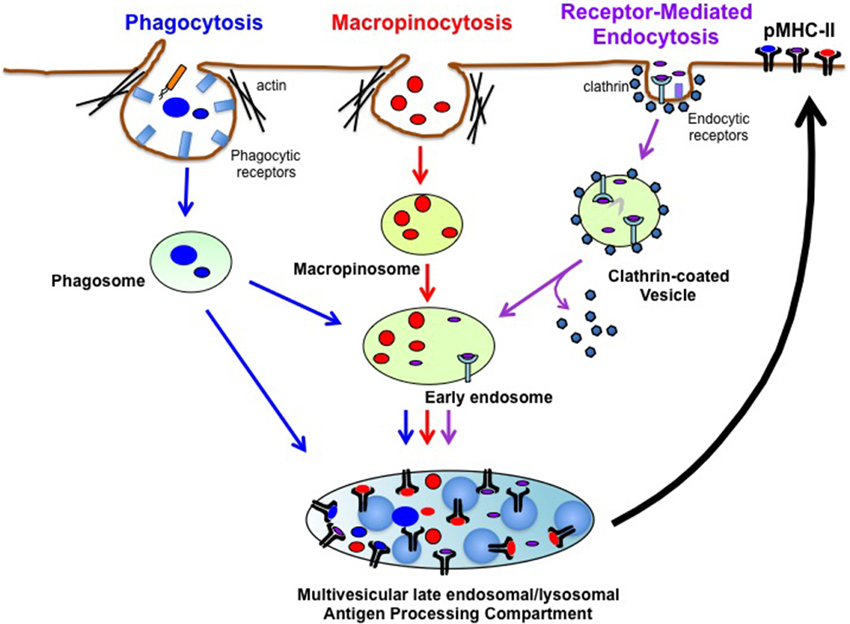
Figure 1. Pathways of Exogenous Antigen Uptake in DCs. DCs internalize extracellular antigens using three main endocytic pathways. Phagocytosis is an endocytic process in which opsonized particles bind to specific receptors on the DC surface and enter cells in membrane-derived phagosomes. Phagocytosed antigens (Blue) can fuse with MHC-II+ lysosomes to generate phagolysosomes (not shown) or can be directly targeted into late endosomal/lysosomal antigen processing compartments to generate pMHC-II complexes. Macropinocytosis mediates non-specific uptake of soluble antigens into the cell via macropinosomes. Antigens in macropinosomes (Red) are transferred into early endosomes that eventually fuse with multivesicular late endosomal/lysosomal antigen processing compartments. Macropinocytosed antigens are degraded and loaded on MHC-II in these compartments. Receptor-mediated endocytosis is a process in which small soluble antigens bind to specific receptors on the DC surface that internalize in clathrin-coated vesicles or in clathrin-uncoated vesicles (not shown). Following uncoating of clathrin, antigens in clathrin-coated vesicles (Purple) are delivered to early endosomes and eventually to antigen processing compartments for proteolytic degradation and pMHC-II formation. After formation in antigen processing compartments, pMHC-II complexes traffic to plasma membrane to initiate antigen-specific adaptive immune responses.
Since foreign-antigen uptake and proteolysis are such important functions of DCs, a variety of tools have been developed to study the mechanisms of antigen uptake in these cells. Quantitating the uptake of fluorescently-labeled molecules [often BSA, dextran, or ovalbumin (OVA)] or the fluorescent dye Lucifer Yellow (LY) is a standard method for measuring antigen uptake via macropinocytosis. However, in many of the studies performed to date it has been difficult to unambiguously distinguish between macropinocytosis and receptor-mediated endocytosis as the mechanims of “antigen” uptake in DCs. Whereas both fluorescently-labeled dextran and OVA have been used to measure uptake via “macropincytosis,” partial suppression of uptake by pretreatment with the mannose receptor-agonist mannan demonstrates that dextran and OVA are also internalized by mannose receptor-mediated endocytosis (Sallusto et al., 1995; Burgdorf et al., 2006; Platt et al., 2010). Thus, the choice of solute marker examined, and exact experimental protocol followed, are the key to knowing precisely which endocytic pathway is actually being assayed.
In addition, a number of inhibitors have been used to “specifically” study macropinocytosis. Inhibitiors of the amiloride family (amiloride, EIPA, HOE-694, and DMA) inhibit Na+/H+ exchanger activity that promotes actin polymerization during macropinocytosis. Sanglifehrin A and rapamycin are immunophilin-binding agents that exert immunosuppressive effects on DC functions including macropinocytosis (Sallusto et al., 1995; Hackstein et al., 2002, 2007). However, other studies revealed that these compounds also inhibit receptor-mediated endocytosis (Hackstein et al., 2002, 2007; Sarkar et al., 2005; Koivusalo et al., 2010). The same issue occurs when using PI3-kinase inhibitors (wortmannin and LY294002) and cytochalasins as inhibitors of macropinocytosis. These inhibitors function by blocking actin polymerization and therefore they not only reduce macropinocytosis but also inhibit phagocytosis in DCs (Araki et al., 1996, 2003). Low doses (<3 μM) of the PKCδ inhibitor Rottlerin significantly reduce uptake of Lucifer Yellow, but not transferrin, in DCs, suggesting that this compound preferentially blocks macropinocytosis in this cell type (Sarkar et al., 2005). However, Rottlerin has been shown to inhibit dectin-1-mediated phagocytosis of zymosan in monocytes when used at a concentration of 5 μM (Elsori et al., 2011), calling into question the specificity of this compound as an inhibitor of macropinocytosis. All the above data suggest that it is very difficult to specifically study the importance of macropinocytosis per se in DC antigen presenting function. More selective and efficient inhibitors need to be identified for investigation of the functional significance of macropinocytosis in the future.
Most DCs exist in vivo in a quiescent “immature” state that can be converted to an activated “mature” state by exposure to a wide variety of stimulatory ligands. Whereas immature DCs have a very robust process of constitutive macropinocytosis, DCs lose this ability upon activation/maturation. The original observation of this phenomenon was made in DCs generated from human blood monocytes in vitro (Sallusto et al., 1995). These in vitro-cultured immature DCs can take up about 1–1.5 pL of extracellular fluid per hour, representing nearly the entire volume of cell itself. Addition of stimuli that induce the maturation of DCs (including TNF-α, IL-1β, CD40L, and a variety of TLR-ligands) profoundly suppresses macropinocytosis. Efficient macropinocytosis cannot be recovered when these stimuli are removed, demonstrating that an irreversible termination of macropinocytosis occurs upon DC maturation. Suppression of macropinocytosis upon DC maturation has been shown in mouse BMDC (Garrett et al., 2000) and in in vitro activated spleen DCs (West et al., 2000).
As in other non-immune cells, macropinocytosis in DCs is controlled by the Rho GTPase Cdc42 and Rac-dependent remodeling of the actin cytoskeleton. Immature DCs possess large amounts of GTP-bound Cdc42 whose expression is greatly diminished upon DC activation (Garrett et al., 2000). Injection of dominant-negative forms of Cdc42 or Rac blocked macropinocytosis in immature DC, whereas overexpression of constitutively-active forms of these same GTPases restored macropinocytosis in mature DCs (Garrett et al., 2000), revealing a direct role of these proteins in the modulation of macropinocytosis in DCs. Interestingly, kinetic studies of macropinocytosis in DCs showed that TLR stimulation of DCs transiently enhanced antigen macropinocytosis in the first few hours, a process that was then followed by dramatic suppression of macropinocytosis hours later (West et al., 2004). LPS and other TLR ligands transiently activated MAP kinase activity and stimulated actin polymerization and disassembly of actin-rich podosomes, thereby resulting in increased accumulation of fluorescent antigen-filled macropinosomes in DCs. However, this acute actin remodeling was not dependent on Rac or Cdc42 (West et al., 2004). It is thought that this transient burst in macropinocytosis in immature DCs serves to maximize antigen capture and presentation of relevant antigens associated with the inflammatory source of DC activation. It is interesting to note that TLR signaling promotes the formation of phagolysosomes following phagocytosis (Blander and Medzhitov, 2006), and it is an intriguing possibility that TLR signaling enhances not only the kinetics of antigen uptake following macropinocytosis but also enhances the “maturation” of macropinosomes into proteolytic antigen processing compartments.
The finding that macropinocytosis is dramatically suppressed upon DCs maturation fits well with the idea that DCs specifically acquire pathogens at the time of their activation in vivo and that after activation additional pathogen uptake is unnecessary (or even undesirable). It has therefore been widely accepted that immature DCs are the primary antigen samplers of peripheral tissues and that maturation convert DCs from efficient antigen sampling cells to efficient antigen presenting cells. There are, however, compelling data challenging this hypothesis. Drutman et al. showed that injection of mice with the inflammatory mediators LPS or CpG resulted in efficient in vivo DC maturation. Surprisingly, subsequent immunization of these “in vivo activated” mice with the model antigen OVA resulted in efficient stimulation of OVA-specific CD8+ and CD4+ T cells both ex vivo and in vivo, suggesting that in vivo activated DCs are still able to take up antigens and present them to T cells (Drutman and Trombetta, 2010). In contradiction to results obtained in vitro, direct measurements of antigen uptake confirmed that antigen delivered in vivo was internalized by macropinocytosis equally well by immature DCs and in vivo matured DCs. Importantly, in this study the authors confirmed the many previous findings that in vitro activation almost completely blocks macropinocytosis in DCs, highlighting the differences in the regulation of macropinocytosis in DCs activated in vitro versus DCs activated in vivo. Platt et al. also found that mature spleen DCs isolated from mice injected with LPS possessed robust macropinocytosis activity and were able to stimulate OVA-specific CD4+ T cells when assayed ex vivo (Platt et al., 2010), supporting the idea that in vivo activation of DCs does not profoundly reduce macropinocytosis and that in vivo activated DCs can generate pMHC-II capable of stimulating naïve CD4+ T cells.
The robust ability of in vivo matured DCs to internalize soluble antigen and subsequently generate pMHC-II complexes capable of stimulating antigen-specific T cell has not been seen in other studies. While Young et al. also found that spleen DCs from control mice and CpG-injected (i.e., in vivo matured)-mice internalized comparable amounts of soluble OVA antigen ex vivo (Young et al., 2007), in their study in vivo-activated spleen DCs were unable to stimulate OVA-specific CD4+ T cells (Young et al., 2007). The authors attributed this finding to impaired formation of MHC-II-peptide complexes in the mature DCs (due to the down-regulation of MHC-II biosynthesis) but not to impaired antigen uptake. Despite conflicting results regarding the extent of T cell activation by in vivo-activated DCs, each of these studies demonstrates that unlike in vitro matured DCs, in vivo matured spleen DCs can still internalize foreign soluble antigens, most likely via macropinocytosis. It is important to remember, however, that simply internalizing antigens is not sufficient for APC function; DCs must generate antigen-specific pMHC-II complexes from the internalized antigens in order to activate antigen-specific T cells ex vivo or in vivo. Factors that affect pMHC-II formation in immature vs. mature DCs [such as diminished biosynthesis of MHC-II (Cella et al., 1997), altered intracellular localization of MHC-II (Turley et al., 2000), or differences in endo/lysosomal proteinase activity (Trombetta et al., 2003)] will contribute to the efficiency of pMHC-II formation in mature DCs regardless of whether macropinocytosis is intact or not, as highlighted by the findings of the abovementioned studies.
There is also evidence showing that the extent of macropinocytosis varies among different types of DCs. Conventional DCs (cDCs) in the spleen capture significantly more soluble fluorescent OVA than plasmacytoid DCs (pDCs) do in vitro, however this difference was not seen in vivo (Young et al., 2008). While CD8+ and CD8− cDCs internalize similar amounts of soluble OVA and BSA in vitro, these DC subsets target the internalized antigens into distinct intracellular compartments and present them to different classes of T cells (Schnorrer et al., 2006). In lung parenchyma, CD11bhiCD103− DCs preferentially internalize soluble OVA protein for presentation on MHC-II to CD4+ T cells, whereas CD11blowCD103+ DCs preferentially phagocytose OVA-coated latex beads for presentation on MHC-I to induce CD8+ T cell responses (Jakubzick et al., 2008). Finally, in the skin Langerin− dermal DCs internalize more soluble OVA than do Langerin+ epidermal DCs (Sparber et al., 2010). However, it is unclear whether OVA uptake in these distinct DC populations is mediated solely via macropinocytosis, as it has been shown that Langerin− dermal DCs express higher amounts of OVA-binding mannose receptors than do Langerin+ epidermal DCs (Turville et al., 2002).
Since one of the major functions of DCs is to process extracellular antigens into small peptides capable of generating pMHC-II complexes, the delivery of foreign antigens to the appropriate antigen-processing compartment is a crucial step in the induction of T cell responses. Sallusto et al. showed that immature DCs internalized and delivered the fluid phase marker FITC-dextran to a large compartment that contains MHC-II, cathepsin D, and LAMP-1, suggesting that macropinocytosed antigen was transported to late endosomes/lysosomes for degradation and pMHC-II formation (Sallusto et al., 1995). Subsequent studies confirmed this intracellular routing pathway of macropinocytosis. Macropinocytosed LY accumulates in late endosomes/lysosomes identified by Lysotracker but not in organelles expressing the early endosomal markers Rab5 and EEA-1 (Burgdorf et al., 2007). All these studies demonstrate that antigen endocytosed within macropinosomes is targeted to late endosomal/lysosomal MHC-II antigen processing compartments that are specialized for induction of CD4+ T cell responses.
Unlike macropinocytosis, receptor-mediated endocytosis can deliver antigens to distinct classes of endosomes in DCs. Mannose receptor-endocytosed OVA is preferentially delivered to an early endosomal compartment, and antigen targeting to this compartment generally leads to poor OVA processing and MHC-II presentation to CD4+ T cells (Burgdorf et al., 2007). By contrast, Fcγ receptor or DEC205-endocytosed OVA is transported rapidly to late endosomes/lysosomes for very efficient MHC-II antigen processing and presentation to OVA-specific T cells (Platt et al., 2010). These studies demonstrate that lysosomal targeting (and presumably proteolysis) of internalized antigens enhances MHC-II-restricted antigen presentation to CD4+ T cells.
All APC subtypes are able to take up extracellular molecules, antigens, and nutrients. Macrophages primarily use surface receptor-dependent phagocytosis to engulf pathogens and apoptotic cells (Aderem and Underhill, 1999), while B cells primarily use their antigen-specific B cell receptor (BCR) to target specific antigens to antigen processing compartments (Lanzavecchia, 1990). By contrast, DCs use macropinocytosis as a major mechanism to sample their microenvironment. Thus efficient, constitutive macropinocytosis distinguishes DCs from macrophages and B cells as APCs capable of sampling tissues without regard to endocytic “receptor” specificity.
In the absence of infection or inflammation, immature DCs use macropinocytosis to constitutively capture self-antigen and present self-pMHC-II complexes to induce regulatory T cells in lymphoid organs (Steinman et al., 2003; Wilson et al., 2004), strongly suggesting that macropinocytosis contributes to DC-induced peripheral tolerance. Recent studies showed that immature peripheral plasmacytoid DCs (pDCs) can transport OVA peptide or OVA expressed in a tissue-specific manner to the thymus and induce clonal depletion of OVA-specific thymocytes (Bonasio et al., 2006; Hadeiba et al., 2012), indicating that constitutive macropinocytosis of antigens by DCs in the periphery may also contribute to the establishment of central tolerance. However, given the demonstration that some fraction of OVA enters cell by receptor-mediated endocytosis (Burgdorf et al., 2007), it remains an open question as to whether macropinocytosis alone is responsible for central tolerance induction in the thymus. Further studies are needed to determine whether, and to what extent, DC macropinocytic uptake of peripheral antigens contributes to central tolerance.
In the presence of infection or under inflammatory conditions, DCs employ macropinocytosis to internalize the soluble pathogenic antigens, degrade them into antigenic peptides, and present these antigenic pMHC-II complexes to CD4+ T cells. A number of studies have shown that macropinocytosis is important for MHC-II antigen presentation by DCs both in vitro and in vivo. Administration of macropinocytosis inhibitors not only reduces the amount of antigen taken up by DCs but also limits their capacity to stimulate antigen-specific CD4+ T cells. This finding has been validated using various model systems, including adoptive transfer of alloantigen-pulsed DCs into syngeneic mice as well as in ex vivo studies using antigen-pulsed APCs to stimulate a wide-variety of mouse and human antigen-specific CD4+ T cells (Sarkar et al., 2005; Hackstein et al., 2007; von Delwig et al., 2006; Drutman and Trombetta, 2010). Together, these studies demonstrate that interfering with antigen uptake via macropinocytosis results in impaired MHC-II antigen presentation by DCs.
Although macropinocytosis plays an important role in DC-mediated immune protection, pathogens can exploit the macropinocytosis machinery to gain access to host cells. Recent studies have shown that many different viruses use the macropinocytosis pathway to enter host cells and cause infection. Viruses possessing this ability include vaccinia virus, adenovirus 3, echovirus 1, coxsackievirus B, Herpes simplex virus 1, and human immunodeficiency virus (Mercer and Helenius, 2009). Each of these viruses activates small GTPases or kinases that stimulate macropinocytosis in host cells, thereby enhancing viral entry. These viruses colocalize with fluid-phase makers within infected cells, providing another line of evidence that viruses enter host cells through macropinocytosis. Although these studies were not directly performed using DCs as host cells, there is no reason to believe that these viruses do not also affect macropinocytiosis in DCs. One consequence of macropinocytosed vaccinia virus into DCs is that DC maturation itself is suppressed (Engelmayer et al., 1999), thereby reducing the possibility of an anti-vaccinia virus immune response. HIV-1 enters DCs by both receptor mediated endocytosis and macropinocytosis, however it is the macropinocytosis pathway that contributes to DC-mediated HIV-1 trans-infection of CD4+ cells (Wang et al., 2008), revealing a role of macropinocytosis in propagating HIV infection. These data therefore provide new insights into the role of macropinocytosis in DC-mediated immune responses.
It is important to note that most of our current knowledge of the role of macropinocytosis in DC function has been acquired using in vitro culture systems. The contribution of macropinocytosis for DC function has not been fully tested in vivo, however this would require development of effective strategies to specifically disrupt macropinocytosis in vivo. Discovering specific inhibitors or generating mice containing mutations of known macropinocytosis regulators, such as Rac1 and Cdc42, is required to fully understand the contribution of this endocytic pathway to DC function in a physiological setting.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We acknowledge the many investigators whose important work could not be cited due to space limitations. The work was supported by the Intramural Research Program of the National Cancer Institute.
Aderem, A., and Underhill, D. M. (1999). Mechanisms of phagocytosis in macrophages. Annu. Rev. Immunol. 17, 593–623. doi: 10.1146/annurev.immunol.17.1.593
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Araki, N., Hatae, T., Furukawa, A., and Swanson, J. A. (2003). Phosphoinositide-3-kinase-independent contractile activities associated with Fcgamma-receptor-mediated phagocytosis and macropinocytosis in macrophages. J. Cell Sci. 116, 247–257. doi: 10.1242/jcs.00235
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Araki, N., Johnson, M. T., and Swanson, J. A. (1996). A role for phosphoinositide 3-kinase in the completion of macropinocytosis and phagocytosis by macrophages. J. Cell Biol. 135, 1249–1260. doi: 10.1083/jcb.135.5.1249
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Blander, J. M., and Medzhitov, R. (2006). Toll-dependent selection of microbial antigens for presentation by dendritic cells. Nature 440, 808–812. doi: 10.1038/nature04596
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Blum, J. S., Wearsch, P. A., and Cresswell, P. (2013). Pathways of antigen processing. Annu. Rev. Immunol. 31, 443–473. doi: 10.1146/annurev-immunol-032712-095910
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bonasio, R., Scimone, M. L., Schaerli, P., Grabie, N., Lichtman, A. H., and von Andrian, U. H. (2006). Clonal deletion of thymocytes by circulating dendritic cells homing to the thymus. Nat. Immunol. 7, 1092–1100. doi: 10.1038/ni1385
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Burgdorf, S., Kautz, A., Bohnert, V., Knolle, P. A., and Kurts, C. (2007). Distinct pathways of antigen uptake and intracellular routing in CD4 and CD8 T cell activation. Science 316, 612–616. doi: 10.1126/science.1137971
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Burgdorf, S., and Kurts, C. (2008). Endocytosis mechanisms and the cell biology of antigen presentation. Curr. Opin. Immunol. 20, 89–95. doi: 10.1016/j.coi.2007.12.002
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Burgdorf, S., Lukacs-Kornek, V., and Kurts, C. (2006). The mannose receptor mediates uptake of soluble but not of cell-associated antigen for cross-presentation. J. Immunol. 176, 6770–6776. doi: 10.4049/jimmunol.176.11.6770
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Cella, M., Engering, A., Pinet, V., Pieters, J., and Lanzavecchia, A. (1997). Inflammatory stimuli induce accumulation of MHC class II complexes on dendritic cells. Nature 388, 782–787. doi: 10.1038/42030
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Drutman, S. B., and Trombetta, E. S. (2010). Dendritic cells continue to capture and present antigens after maturation in vivo. J. Immunol. 185, 2140–2146. doi: 10.4049/jimmunol.1000642
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Elsori, D. H., Yakubenko, V. P., Roome, T., Thiagarajan, P. S., Bhattacharjee, A., Yadav, S. P., et al. (2011). Protein kinase Cdelta is a critical component of Dectin-1 signaling in primary human monocytes. J. Leukoc. Biol. 90, 599–611. doi: 10.1189/jlb.0610376
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Engelmayer, J., Larsson, M., Subklewe, M., Chahroudi, A., Cox, W. I., Steinman, R. M., et al. (1999). Vaccinia virus inhibits the maturation of human dendritic cells: a novel mechanism of immune evasion. J. Immunol. 163, 6762–6768.
Garrett, W. S., Chen, L. M., Kroschewski, R., Ebersold, M., Turley, S., Trombetta, S., et al. (2000). Developmental control of endocytosis in dendritic cells by Cdc42. Cell 102, 325–334. doi: 10.1016/S0092-8674(00)00038-6
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hackstein, H., Steinschulte, C., Fiedel, S., Eisele, A., Rathke, V., Stadlbauer, T., et al. (2007). Sanglifehrin a blocks key dendritic cell functions in vivo and promotes long-term allograft survival together with low-dose CsA. Am. J. Transplant. 7, 789–798. doi: 10.1111/j.1600-6143.2006.01729.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hackstein, H., Taner, T., Logar, A. J., and Thomson, A. W. (2002). Rapamycin inhibits macropinocytosis and mannose receptor-mediated endocytosis by bone marrow-derived dendritic cells. Blood 100, 1084–1087. doi: 10.1182/blood.V100.3.1084
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hadeiba, H., Lahl, K., Edalati, A., Oderup, C., Habtezion, A., Pachynski, R., et al. (2012). Plasmacytoid dendritic cells transport peripheral antigens to the thymus to promote central tolerance. Immunity 36, 438–450. doi: 10.1016/j.immuni.2012.01.017
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hewlett, L. J., Prescott, A. R., and Watts, C. (1994). The coated pit and macropinocytic pathways serve distinct endosome populations. J. Cell Biol. 124, 689–703. doi: 10.1083/jcb.124.5.689
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Jakubzick, C., Helft, J., Kaplan, T. J., and Randolph, G. J. (2008). Optimization of methods to study pulmonary dendritic cell migration reveals distinct capacities of DC subsets to acquire soluble versus particulate antigen. J. Immunol. Methods 337, 121–131. doi: 10.1016/j.jim.2008.07.005
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Joffre, O. P., Segura, E., Savina, A., and Amigorena, S. (2012). Cross-presentation by dendritic cells. Nat. Rev. Immunol. 12, 557–569. doi: 10.1038/nri3254
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Koivusalo, M., Welch, C., Hayashi, H., Scott, C. C., Kim, M., Alexander, T., et al. (2010). Amiloride inhibits macropinocytosis by lowering submembranous pH and preventing Rac1 and Cdc42 signaling. J. Cell Biol. 188, 547–563. doi: 10.1083/jcb.200908086
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kreer, C., Rauen, J., Zehner, M., and Burgdorf, S. (2011). Cross-presentation: how to get there - or how to get the ER. Front. Immunol. 2:87. doi: 10.3389/fimmu.2011.00087
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lanzavecchia, A. (1990). Receptor-mediated antigen uptake and its effect on antigen presentation to class II-restricted T lymphocytes. Annu. Rev. Immunol. 8, 773–793. doi: 10.1146/annurev.iy.08.040190.004013
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lim, J. P., and Gleeson, P. A. (2011). Macropinocytosis: an endocytic pathway for internalising large gulps. Immunol. Cell Biol. 89, 836–843. doi: 10.1038/icb.2011.20
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Mercer, J., and Helenius, A. (2009). Virus entry by macropinocytosis. Nat. Cell Biol. 11, 510–520. doi: 10.1038/ncb0509-510
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Norbury, C. C., Chambers, B. J., Prescott, A. R., Ljunggren, H. G., and Watts, C. (1997). Constitutive macropinocytosis allows TAP-dependent major histocompatibility complex class I presentation of exogenous soluble antigen by bone marrow-derived dendritic cells. Eur. J. Immunol. 27, 280–288. doi: 10.1002/eji.1830270141
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Platt, C. D., Ma, J. K., Chalouni, C., Ebersold, M., Bou-Reslan, H., Carano, R. A., et al. (2010). Mature dendritic cells use endocytic receptors to capture and present antigens. Proc. Natl. Acad. Sci. U.S.A. 107, 4287–4292. doi: 10.1073/pnas.0910609107
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Sallusto, F., Cella, M., Danieli, C., and Lanzavecchia, A. (1995). Dendritic cells use macropinocytosis and the mannose receptor to concentrate macromolecules in the major histocompatibility complex class II compartment: downregulation by cytokines and bacterial products. J. Exp. Med. 182, 389–400. doi: 10.1084/jem.182.2.389
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Sarkar, K., Kruhlak, M. J., Erlandsen, S. L., and Shaw, S. (2005). Selective inhibition by rottlerin of macropinocytosis in monocyte-derived dendritic cells. Immunology 116, 513–524. doi: 10.1111/j.1365-2567.2005.02253.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Schnorrer, P., Behrens, G. M., Wilson, N. S., Pooley, J. L., Smith, C. M., El-Sukkari, D., et al. (2006). The dominant role of CD8+ dendritic cells in cross-presentation is not dictated by antigen capture. Proc. Natl. Acad. Sci. U.S.A. 103, 10729–10734. doi: 10.1073/pnas.0601956103
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Schuette, V., and Burgdorf, S. (2014). The ins-and-outs of endosomal antigens for cross-presentation. Curr. Opin. Immunol. 26, 63–68. doi: 10.1016/j.coi.2013.11.001
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Shen, L., Sigal, L. J., Boes, M., and Rock, K. L. (2004). Important role of cathepsin S in generating peptides for TAP-independent MHC class I crosspresentation in vivo. Immunity 21, 155–165. doi: 10.1016/j.immuni.2004.07.004
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Sparber, F., Tripp, C. H., Hermann, M., Romani, N., and Stoitzner, P. (2010). Langerhans cells and dermal dendritic cells capture protein antigens in the skin: possible targets for vaccination through the skin. Immunobiology 215, 770–779. doi: 10.1016/j.imbio.2010.05.014
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Steinman, R. M., Hawiger, D., and Nussenzweig, M. C. (2003). Tolerogenic dendritic cells. Annu. Rev. Immunol. 21, 685–711. doi: 10.1146/annurev.immunol.21.120601.141040
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Stuart, L. M., and Ezekowitz, R. A. (2005). Phagocytosis: elegant complexity. Immunity 22, 539–550. doi: 10.1016/j.immuni.2005.05.002
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Swanson, J. A., and Watts, C. (1995). Macropinocytosis. Trends Cell Biol. 5, 424–428. doi: 10.1016/S0962-8924(00)89101-1
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Trombetta, E. S., Ebersold, M., Garrett, W., Pypaert, M., and Mellman, I. (2003). Activation of lysosomal function during dendritic cell maturation. Science 299, 1400–1403. doi: 10.1126/science.1080106
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Turley, S. J., Inaba, K., Garrett, W. S., Ebersold, M., Unternaehrer, J., Steinman, R. M., et al. (2000). Transport of peptide-MHC class II complexes in developing dendritic cells. Science 288, 522–527. doi: 10.1126/science.288.5465.522
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Turville, S. G., Cameron, P. U., Handley, A., Lin, G., Pohlmann, S., Doms, R. W., et al. (2002). Diversity of receptors binding HIV on dendritic cell subsets. Nat. Immunol. 3, 975–983. doi: 10.1038/ni841
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Villadangos, J. A., and Schnorrer, P. (2007). Intrinsic and cooperative antigen-presenting functions of dendritic-cell subsets in vivo. Nat. Rev. Immunol. 7, 543–555. doi: 10.1038/nri2103
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
von Delwig, A., Hilkens, C. M., Altmann, D. M., Holmdahl, R., Isaacs, J. D., Harding, C. V., et al. (2006). Inhibition of macropinocytosis blocks antigen presentation of type II collagen in vitro and in vivo in HLA-DR1 transgenic mice. Arthritis Res. Ther. 8, R93. doi: 10.1186/ar1964
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Wang, J. H., Wells, C., and Wu, L. (2008). Macropinocytosis and cytoskeleton contribute to dendritic cell-mediated HIV-1 transmission to CD4+ T cells. Virology 381, 143–154. doi: 10.1016/j.virol.2008.08.028
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
West, M. A., Prescott, A. R., Eskelinen, E. L., Ridley, A. J., and Watts, C. (2000). Rac is required for constitutive macropinocytosis by dendritic cells but does not control its downregulation. Curr. Biol. 10, 839–848. doi: 10.1016/S0960-9822(00)00595-9
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
West, M. A., Wallin, R. P., Matthews, S. P., Svensson, H. G., Zaru, R., Ljunggren, H. G., et al. (2004). Enhanced dendritic cell antigen capture via toll-like receptor-induced actin remodeling. Science 305, 1153–1157. doi: 10.1126/science.1099153
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Wilson, N. S., El-Sukkari, D., and Villadangos, J. A. (2004). Dendritic cells constitutively present self antigens in their immature state in vivo and regulate antigen presentation by controlling the rates of MHC class II synthesis and endocytosis. Blood 103, 2187–2195. doi: 10.1182/blood-2003-08-2729
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Young, L. J., Wilson, N. S., Schnorrer, P., Mount, A., Lundie, R. J., La Gruta, N. L., et al. (2007). Dendritic cell preactivation impairs MHC class II presentation of vaccines and endogenous viral antigens. Proc. Natl. Acad. Sci. U.S.A. 104, 17753–17758. doi: 10.1073/pnas.0708622104
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Young, L. J., Wilson, N. S., Schnorrer, P., Proietto, A., ten Broeke, T., Matsuki, Y., et al. (2008). Differential MHC class II synthesis and ubiquitination confers distinct antigen-presenting properties on conventional and plasmacytoid dendritic cells. Nat. Immunol. 9, 1244–1252. doi: 10.1038/ni.1665
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Keywords: dendritic cell, macropinocytosis, endocytosis, MHC class II, antigen presentation
Citation: Liu Z and Roche PA (2015) Macropinocytosis in phagocytes: regulation of MHC class-II-restricted antigen presentation in dendritic cells. Front. Physiol. 6:1. doi: 10.3389/fphys.2015.00001
Received: 11 November 2014; Accepted: 05 January 2015;
Published online: 30 January 2015.
Edited by:
Hiroyuki Kabayama, RIKEN Brain Science Institute, JapanReviewed by:
Bénédicte Manoury, Institut National de la Santé et de la Recherche Médicale, FranceCopyright © 2015 Liu and Roche. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Paul A. Roche, National Institutes of Health, Building 10, Room 4B36, Bethesda, MD 20892, USA e-mail:cGF1bC5yb2NoZUBuaWguZ292
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.