 Arturo Mendoza-Galván
Arturo Mendoza-Galván Roger Magnusson2
Roger Magnusson2 Hans Arwin
Hans Arwin Kenneth Järrendahl
Kenneth Järrendahl- 1Cinvestav – Querétaro, Juriquilla, Mexico
- 2Materials Optics, Department of Physics, Chemistry and Biology, Linköping University, Linköping, Sweden
- 3Conservation Ecology Group, Department of Physics, Chemistry and Biology, Linköping University, Linköping, Sweden
Many species of beetles from the family Scarabaeidae reflect light with near-circular polarization. In some cases, spectral narrow-band polarization phenomena result in a distinct color with a metallic shine. In other cases, broad-band features are seen, and these beetles have a silvery or goldish appearance. These features in the cuticles originate from helicoidal structures, so-called circular Bragg structures and also referred to as Bouligand structures. In this communication, Protaetia mirifica, exhibiting near-circular polarization properties in dual spectral regions, centered approximately at the wavelengths of 474 and 770 nm, is investigated in considerable detail using Mueller matrix spectroscopic ellipsometry (MMSE). From interference oscillations in the MMSE spectra, the pitch profile of the helicoidal structures in the beetle cuticle is extracted and further used in electromagnetic modeling of the cuticle structure, including the determination of epicuticle and exocuticle thicknesses (280 nm and 8.1
1 Introduction
Beetles reflecting light with a near-circular polarization (high degree of circular polarization) are in particular found in tribe Cetoniini in the subfamily Cetoniinae (flower chafers) and in tribe Rutelini in the subfamily Rutelinae (leaf chafers). An early observation was made more than 100 years ago by Michelson in his studies of Chrysina resplendens (Boucard, 1875) [1]. Neville and Caveney discussed these cuticle structures in terms of cholesteric liquid crystal analogs [2], and Bouligand suggested that a twisted structure of lamellae is the origin of these effects [3]. Today, these structures are often called Bouligand structures [4], and the associated polarization phenomenon is referred to as a circular Bragg resonance. The structural origin has been discussed in several reviews [5–8], and in a survey, Pye used circular polarizers to study more than 19,000 species of scarab beetles [9]. The twisted cuticle structure is generally considered to be composed of chitin molecules organized in crystals in the shape of fibers lying side by side, forming lamellae, which are stacked with an angle between each lamella, thus forming a twisted plywood structure [8].
Some of these beetles display narrow-band reflections like Cetonia aurata (Linné, 1761), which, in most specimens, has a green appearance but can also be red or blue [10]. Protaetia cuprea (Fabricius, 1775), which is another species in the same subfamily, exhibits a double resonance feature in its optical spectra [11]. Other beetles may show broadband reflections and appear as if they are made of silver, like Chrysina argenteola (Bates, 1888) [12], or gold, like in the widely studied C. resplendens [1, 7, 13, 14], to give a few examples. The elliptical polarization effects in the reflected light are manifested as non-zero values of the Mueller-matrix element
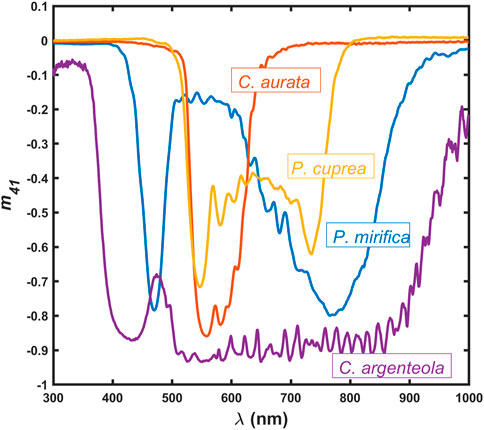
Figure 1. Mueller matrix element m41 for P. mirifica, C. aurata, P. cuprea, and C. argenteola, showing the capability of their cuticles to reflect near-circularly polarized light.
The objective of this report is to present a detailed study of dual chiral structures in beetle cuticles using Mueller-matrix spectroscopic ellipsometry (MMSE). The beetle chosen is P. mirifica. Multiple chiral structures in beetle cuticles have been studied earlier, and here, we give just a few examples. Carter et al. [15] studied variations in the circularly polarized reflection of several beetles, showing double peaks, including Lomaptera pygmaea (Kraatz, 1880) with a moderate separation of two peaks and Lomaptera geelvinkiana (Guérin-Méneville, 1830), showing a 300-nm reflectance peak separation. Several groups have studied C. resplendens. Vargas et al. [16] studied its dual chiral structure and observed that the total reflectance of this beetle exhibits two maxima, one at a wavelength of 575 nm and another at a wavelength of 755 nm. Its cuticle has a unidirectional layer between the two helicoidal structures, with the effect that both left- and right-handed polarized light is reflected, which increases the overall reflectance. In addition, the two structures have pitch variations with cuticle depth so that the cuticle becomes a broadband bio-reflector. Bagge et al. investigated C. resplendens using MMSE but in a smaller spectral range [12].
The preferred method to explore polarization and depolarization features of bio-reflectors is MMSE as it provides a complete description of specular reflection including depolarization [17]. Some early work using Mueller matrices was performed by Goldstein [13], and the methodology was further developed by Hodgkinson et al. [14] and Arwin et al. [18]. In addition to capabilities to completely map polarization features [10, 14] in specular reflection, Mueller matrix spectroscopy also offers possibilities to perform electromagnetic modeling of cuticle structures [19], detailed analysis of cuticle pitch grading [20, 21], imaging of polarization patterns [22, 23], bio-reflector characterization using sum decomposition [22], and chirality quantification using differential decomposition in Mueller matrix transmission studies [24].
Mueller matrix spectra were measured on P. mirifica, followed by a detailed pitch analysis presented in Section 3.3 and electromagnetic modeling presented in Section 3.4. Sum decomposition of the data was performed, as shown in Section 3.5, and related to the structure as observed by electron microscopy.
2 Materials and methods
2.1 Beetle specimens
Specimens of the flower chafer P. mirifica (Figure 2) were studied, and the data presented here were recorded on one specimen from the Mersin region and one from the Balikesir region in southern Turkey. These two specimens are referred to as PM1 and PM2, respectively. Most specimens of P. mirifica are dark purple with a metallic look and are 20–30 mm in size. Its natural habitat is wooded pastures and light forests with old hollow oaks (Quercus spp). The larvae live for 2–3 years in the compost at the bottom of tree trunk cavities, consuming dead fungi-infested wood. The adults are day-active and very good flyers, feeding on sap and fruits. The species is very rare with less than 20 known sites around the Mediterranean Sea in Europe. It is on the European Red List as vulnerable (VU) [25].

Figure 2. Photographs of Protaetia mirifica.
2.2 Methods and theory
A scanning electron microscope (SEM) (Zeiss Gemini 560) operated at 3 keV was used to obtain cross-sectional images. For preparation, the samples were cut with a razor blade and coated with a few nm-thin layer of Pt deposited during 10 s at 60 mA at a pressure of
where
The interaction with a sample is described with a normalized Mueller matrix
For incident unpolarized light
i.e., the polarization of the reflected light is completely described by the first column of
3 Results
3.1 SEM
An elytron of P. mirifica was investigated using SEM, and a cross-section image is shown in Figure 3. In the uppermost 15-μm region, two distinct regions can be identified, with the region closest to the surface having a layered structure with a smaller period compared to the lower layered region. These two regions are considered to constitute the so-called exocuticle, which is mainly responsible for the reflecting properties of the cuticle. The supporting endocuticle at the bottom is not involved in the reflection since the light will not penetrate to this depth. A thin epicuticle with a thickness of a few hundred nm is expected at the cuticle surface but cannot be distinguished.
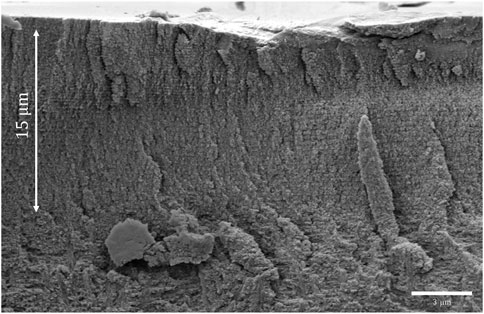
Figure 3. Cross-sectional SEM image of an elytron of Protaetia mirifica, PM1.
3.2 General features of primary Mueller matrix data
Figure 4 shows the Mueller matrix measured on the scutellum of the P. mirifica beetle PM1 at an angle of incidence
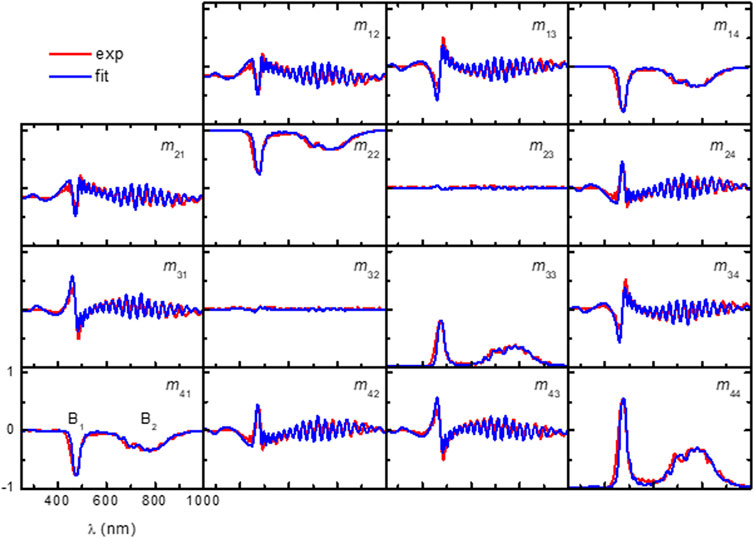
Figure 4. Experimental and model-calculated Mueller matrices from the scutellum of P. mirifica (beetle PM1) at
3.3 Pitch analysis
Pitch variation through the exocuticle is determined from the oscillations seen in several of the Mueller matrix elements. For a constant pitch, these oscillations would be equidistant if data are plotted versus
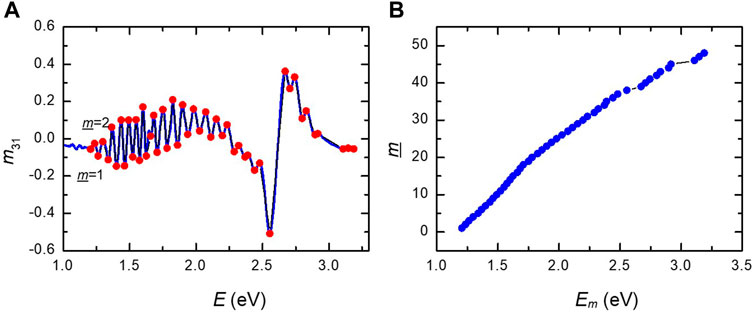
Figure 5. (A) Labeling from
In the C. chrysargyrea study [20], the effective penetration depth
where
where
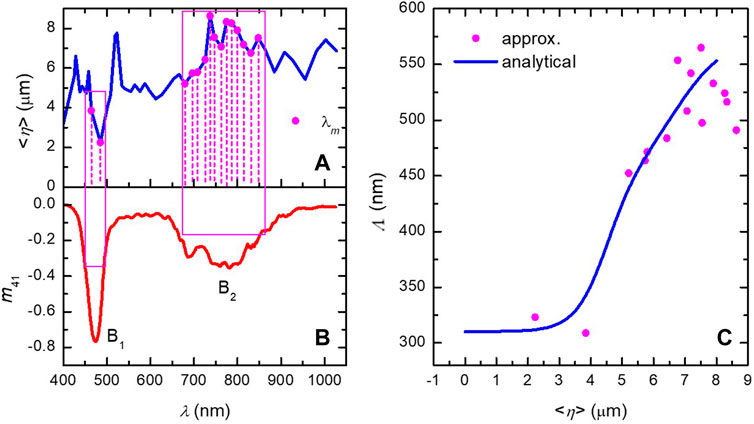
Figure 6. (A) Effective penetration depth
An analytical expression describing the pitch variation is given by [20]
where
3.4 Electromagnetic modeling
The next step is to model the exocuticle of the beetle as an optical active helicoidal (Bouligand) structure composed of biaxial slices stacked with a twist to each other relative to the cuticle normal, thus mimicking cholesteric liquid crystals [8]. Each slice is assigned to have refractive indices
where
In the optical model used, the helicoidal structure lies in between a substrate (the endocuticle) and a surface layer (the epicuticle). These parts are modeled with Cauchy dispersions (see Eq. S3 in the Supplementary Material for details). Non-linear regression is performed to fit the parameters in the model to minimize the differences between the experimental and model-generated data. The fit is shown in Figure 4 above, and parameter values can be found in Supplementary Figure S2 in Supplementary Material. The best-fit thicknesses and their 90% confidence intervals were found to be 8.1 ± 0.1 μm for the exocuticle and 0.28 ± 0.01 μm for the epicuticle. The best-fit refractive indices are shown in Supplementary Figure S3 in Supplementary Material.
The pitch profile
where
Figure 7A shows the azimuth from Eq. 8 versus the thickness-normalized position measured from the bottom of the exocuticle. In Figure 7B, the inverted relationship between
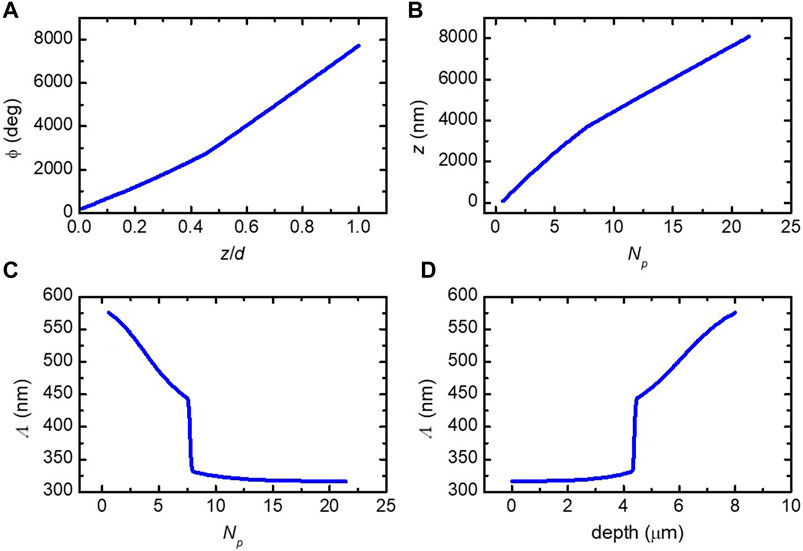
Figure 7. (A) Azimuth of the biaxial slices direction versus normalized depth from Eq. 8. (B) Inverted relationship between
3.5 Cuticle reflector characteristics from sum decomposition of its Mueller matrix
For the human eye, the beetle P. mirifica has a dark metallic shine, often purple or dark blue-green. Its polarizing properties can be revealed using polarizing filters. If observed through a circular polarizer, the human eye gives us the qualitative perception that the beetle appears almost black in a right-handed circular polarizer but similar as with the naked eye (except a little darker) in a left-handed circular polarizer. In this section, we analyze the reflecting characteristics of the cuticle in different spectral regions and describe the reflection with Mueller matrices in terms of basic reflector devices.
A Mueller matrix
where the coefficients
Figure 8A shows that the coefficients shown in Eq. 11 are obtained from a Cloude decomposition of
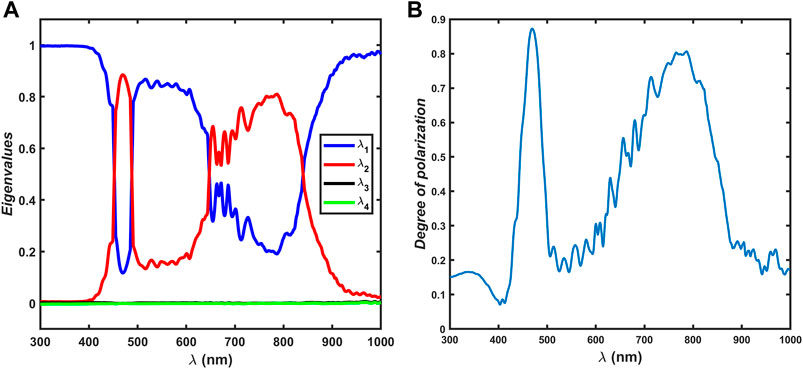
Figure 8. (A) Coefficients
From the sum decomposition of
Figure 8A shows that a left-handed circular polarizer is the dominating character in the B1 band with a maximum of
The strong interference oscillations in several off-diagonal elements in
4 Discussion
Details in the chiral structures vary from position to position on cuticles and among specimens, as well as on the size of the illuminated area, as shown by Vargas et al. [16]. These statements are general in studies of beetle reflectance and also hold for MMSE studies. In this report, data from two beetle specimens are included. The Mueller matrix of specimen PM1 has less pronounced near-infrared chirality and is used in pitch analysis and electromagnetic modeling. Specimen PM2 has a pronounced purple color and has a similar magnitude in
The reflection Mueller matrices were recorded at
Some of the elements in the Mueller matrices have pronounced oscillations, and their magnitude varies with the position on the cuticle and among specimens. Such oscillations are commonly observed in the optical studies of beetle cuticles and originate from interference in the cuticle. The understanding of the phenomenon is complicated by the fact that it is unclear if it is the actual cuticle thickness or the penetration depth of light that rules the interference. The exocuticle thickness in P. mirifica is of the order of 8 μm, which is rather small, and interference oscillations are expected to be larger compared to C. aurata with a cuticle thickness of 20 μm [22], thus having more periods in the helicoidal structure. More pronounced oscillations also indicate that the pitch is rather constant, which is the case in the near-surface region of P. mirifica, as shown in Figure 7D. A gradient in or a random variation in pitch will also decrease oscillations, and in the electromagnetic modeling of the cuticle of C. aurata, a pitch distribution was included to match the broadening of the circular Bragg resonance and reduce oscillations [18]. Another factor which may influence the magnitude of the oscillations is the spot size in the measurements. A larger spot will probe a larger area on the cuticle, and lateral inhomogeneities will then reduce the oscillations. This is shown in the work by Bagge et al., who used a beam size larger than the studied beetles [12].
At optical frequencies, light interacts with materials mainly through the excitation of dipoles by the electric field
Since differential decomposition provides
Perception of color depends on the source, the reflecting sample, and the detector. The cuticle of a P. mirifica specimen often appears purple, with a metallic shine for the naked eye in daylight, but hue and brilliance vary among specimens. The specimen PM1 studied here is less purple and has a green–blue color. The color depends on the spectral variation in the reflected irradiance, but the spectral distribution of reflectance is not contained in a normalized Mueller matrix as it only carries information about polarization properties of a reflector. However, electromagnetic modeling of a structure provides full detail about the structure and optical parameters of the sample, and the spectral reflectance can be derived. If, in addition, a source and detector are defined, the color coordinates L*a*b* can be calculated. This is implemented in the used software program (CompleteEASE), as detailed by Johs et al. [34]. From the electromagnetic model used for the structure of P. mirifica (specimen PM1), represented by the Mueller matrix in Figure 4, the coordinates were found to be L* = 32.1, a* = −14.6, and b* = 8.1. These coordinates correspond to a color with less lightness than what is observed by the eye. However, L*a*b* coordinates represent color perceived in the specular mode, whereas when a beetle is viewed in daylight, light is reflected in a range of incident angles, thus increasing the lightness.
It is not yet fully understood if circular polarization has some biological significance. Rather, few reports are found on the subject. Chrysina gloriosa (Leconte, 1854) has been found to have phototactic response and can discriminate between linearly and circularly polarized light, whereas its relative Chrysina woodi (Horn, 1883) cannot [35]. Polarized vision showing positive polarotaxis has been investigated in horseflies and deerflies by Horváth et al. [36]. The four scarab beetles Anomala dubia (Scopoli, 1763), Anomala vitis (Fabricius, 1775), C. aurata, and P. cuprea all possess left-circularly polarizing exocuticles. However, Blahó et al. [37] investigated their behavior and concluded that these four species are not attracted to circularly polarized light when feeding or for intraspecies communication. Recently Li et al. [38] reported that left-handed circularly polarized light probably mediates mating behavior in Anomala corpulenta (Motschulsky, 1854), but the mechanism for how this is done could not be found. Furthermore, they found by RNA sequencing that left-handed circularly polarized light also affects gene expression. Due to the limited information available, the hypothesis that circularly polarized light has a biological significance for insects needs more studies for verification. The studies in this report do not address this aspect but may, in the future, be found, a reason why evolution in some beetles has developed dual circular Bragg resonances in their cuticles.
5 Conclusion
Optical and structural properties of the scarab beetle P. mirifica have been studied with the following results:
• The cuticle’s purple color with metallic shine originates from dual circular Bragg resonances with one resonance near the ultraviolet spectral region and the other near the infrared region of the visible spectrum.
• The widths, strengths, and spectral positions of the resonances depend on measurement positions on the cuticle and vary among specimens.
• The pitch profile of the dual chiral structure in the beetle cuticle was determined and found qualitatively consistent with SEM analysis.
• The spectral reflection Mueller matrix has been used to model the cuticle structure, and cuticle layer thicknesses and refractive indices have been determined.
• Sum decomposition of the cuticle Mueller matrix reveals that reflection can be described either as a circular polarizer or as a dielectric mirror depending on the spectral region.
Data availability statement
The original contributions presented in the study are included in the article Supplementary Material; further inquiries can be directed to the corresponding author.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
AM-G: conceptualization, investigation, writing–review and editing, formal analysis, and methodology. RM: formal analysis, methodology, software, visualization, and writing–review and editing. NJ: data curation, writing–review and editing, and resources. HA: conceptualization, data curation, formal analysis, investigation, methodology, writing–original draft, and writing–review and editing. KJ: conceptualization, funding acquisition, investigation, project administration, resources, supervision, validation, and writing–review and editing.
Funding
The authors declare that financial support was received for the research, authorship, and/or publication of this article. This research was funded by Linköping University and the Swedish Government Strategic Research Area in Materials Science on Advanced Functional Materials at Linköping University (Faculty Grant SFO-Mat-Liu No. 2009-000971).
Acknowledgments
The authors would like to thank Enrique Garcia-Caurel (École Polytechnique, France) for sharing his MATLAB code for decomposition of Mueller matrices. They would also like to thank Anna Elsukova (Linköping University) for the microscope and the Swedish Research Council for access to ARTEMI, the Swedish National Infrastructure in Advanced Electron Microscopy, grant no. 2021-00171. Photographs of P. mirifica were kindly provided by Ogün Türkay (Figure 2, left) and Stanislav Snäll (Figure 2, right).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphy.2024.1444297/full#supplementary-material
References
2. Neville AC, Caveney S. Scarabaeid beetle exocuticle as an optical analogue of cholesteric liquid crystals. Biol Rev (1969) 44:531–562. doi:10.1111/j.1469-185x.1969.tb00611.x
3. Bouligand Y. Sur l’existence de “pseudomorphoses cholestertériques” chez divers organismes vivants. J Phys Colloques (1969) 30:C490–C103.
4. Weiglhofer S, Lakhtakia A. Introduction to complex mediums for optics and electromagnetics. Bellingham: SPIE Press (2003). doi:10.1117/3.504610.fm
6. Seago AE, Brady P, Vigneron JP, Schultz TD. Gold bugs and beyond: a review of iridescence and structural colour mechanisms in beetles (Coleoptera). J R Soc Interf (2009) 6:S165–184. doi:10.1098/rsif.2008.0354.focus
7. Vukusic P, Sambles JR. Photonic structures in biology. Nature (2003) 424:852–856. doi:10.1038/nature01941
8. Lenau T, Barfoed M. Colours and metallic sheen in beetle shells — a biomimetic search for material structuring principles causing light interference. Adv Eng Mat (2008) 10:299–314. doi:10.1002/adem.200700346
9. Pye JD. The distribution of circularly polarized light reflection in the Scarabaeoidea (Coleoptera). Biol J Linn Soc. (2010) 100:585–596. doi:10.1111/j.1095-8312.2010.01449.x
10. Arwin H, Fernández del Río L, Järrendahl K. Comparison and analysis of mueller-matrix spectra from exoskeletons of blue, green and red Cetonia aurata. Thin Solid Films (2014) 571:739–743. doi:10.1016/j.tsf.2014.02.012
11. Arwin H, Fernández del Río L, Åkerlind C, Valyukh S, Mendoza-Galván A, Magnusson R, et al. On the polarization of light reflected from beetle cuticles. Mater Today Proc (2017) 4:4933–4941.
12. Bagge LE, Kenton AC, Lyons BA, Wehling MF, Goldstein DH. Mueller matrix characterizations of circularly polarized reflections from golden scarab beetles. Appl Opt (2020) 59:F85–93. doi:10.1364/AO.398832
13. Goldstein DH. Polarization properties of Scarabaeidae. Appl Opt (2006) 45:7944–7950. doi:10.1364/ao.45.007944
14. Hodgkinson I, Lowrey S, Bourke L, Parker A, McCall MW. Mueller-matrix characterization of beetle cuticle: polarized and unpolarized reflections from representative architectures. Appl Opt (2010) 49:4558–4567. doi:10.1364/ao.49.004558
15. Carter IE, Weir K, McCall MW, Parker AR. Variation in the circularly polarized light reflection of Lomaptera (Scarabaeidae) beetles. J R Soc Interf (2016) 13:20160015. doi:10.1098/rsif.2016.0015
16. Vargas WE, Libby E, Alfaro-Córdoba M, Hernández-Jiménez M, Avendano E, Solís Á, et al. Optical and morphological properties of the cuticle of Chrysina resplendens scarabs: role of effective and structural pitches. Rec Adv Phot Opt (2021) 4:56–68. doi:10.36959/665/323
17. Fujiwara H. Spectroscopic ellipsometry principles and applications. Chichester: John Wiley and Sons Ltd. (2007).
18. Arwin H, Magnusson R, Landin J, Järrendahl K. Chirality-induced polarization effects in the cuticle of scarab beetles: 100 years after Michelson. Phil Mag (2012) 92:1583–1599. doi:10.1080/14786435.2011.648228
19. Arwin H, Berlind T, Johs B, Järrendahl K. Cuticle structure of the scarab beetle Cetonia aurata analyzed by regression analysis of Mueller-matrix ellipsometric data. Opt Express (2013) 21:22645–22656. doi:10.1364/oe.21.022645
20. Mendoza-Galván A, Fernández del Río L, Järrendahl K, Arwin H. Graded pitch profile for the helicoidal broadband reflector and left-handed circularly polarizing cuticle of the scarab beetle Chrysina chrysargyrea. Sci Rep (2018) 8:6456. doi:10.1038/s41598-018-24761-w
21. Mendoza-Galván A, Järrendahl K, Arwin H. Graded circular Bragg reflectors: a semi-analytical retrieval of approximate pitch profiles from Mueller-matrix data. J Opt (2019) 21:125401. doi:10.1088/2040-8986/ab4dc7
22. Arwin H, Magnusson R, Garcia-Caurel E, Fallet C, Järrendahl K, Foldyna M, et al. Sum decomposition of Mueller-matrix images and spectra of beetle cuticles. Opt Express (2015) 23:1951–1966. doi:10.1364/oe.23.001951
23. Ossikovski R, Foldyna M, Fallet C, De Martino A. Experimental evidence for naturally occurring nondiagonal depolarizers. Opt Lett (2009) 34:2426–2428. doi:10.1364/ol.34.002426
24. Arwin H, Mendoza-Galvan A, Magnusson R, Andersson A, Landin J, Järrendahl K, et al. Structural circular birefringence and dichroism quantified by differential decomposition of spectroscopic transmission Mueller matrices from Cetonia aurata. Opt Lett (2016) 41:3293–3296. doi:10.1364/OL.41.003293
25. Nieto A, Alexander KNA. European red list of saproxylic beetles. Luxembourg: Publications Office of the European Union (2010).
27. Muñoz-Pineda E, Järrendahl K, Arwin H, Mendoza-Galván A. Symmetries and relationships between elements of the Mueller matrix spectra of the cuticle of the beetle Cotinis mutabilis. Thin Solid Films (2014) 571:660–665. doi:10.1016/j.tsf.2013.11.144
28. Arteaga O. Natural optical activity vs circular Bragg reflection studied by Mueller matrix ellipsometry. Thin Solid Films (2016) 617:14–19. doi:10.1016/j.tsf.2016.01.012
29. Mendoza Galván A, Muñoz-Pineda E, Järrendahl K, Arwin H. Pitch profile across the cuticle of the scarab beetle Cotinis mutabilis determined by analysis of Mueller matrix measurements. R Soc Open Sci (2018) 5:181096. doi:10.1098/rsos.181096
31. Arwin H, Schoeche S, Hilfiker J, Hartveit M, Järrendahl K, Juárez-Rivera OR, et al. Optical chirality determined from mueller matrices. Appl Sci (2021) 11:6742. doi:10.3390/app11156742
32. Ossikovski R. Differential matrix formalism for depolarizing anisotropic media. Opt Lett (2011) 36:2330–2428. doi:10.1364/ol.36.002330
33. Schonhöfer A, Kuball H-G. Symmetry properties of the Mueller matrix. Chem Phys (1987) 115:159–67. doi:10.1016/0301-0104(87)80030-7
34. Johs B, Arwin H, Wagner T, Appel D, Peros D. Accuracy of color determination from spectroscopic ellipsometry measurements. Thin Solid Films (2011) 519:2711–2714. doi:10.1016/j.tsf.2010.12.058
35. Brady P, Cummings ME. Differential response to circularly polarized light by the jewel scarab beetle Chrysina gloriosa. Am Nat (2010) 175:614–620. doi:10.1086/651593
36. Horváth G, Majer J, Horváth L, Szivák I, Kriska G. Ventral polarization vision in tabanids: horseflies and deerflies (Diptera: Tabanidae) are attracted to horizontally polarized light. Naturwissenschaften (2008) 95:1093–1100. doi:10.1007/s00114-008-0425-5
37. Blahó M, Egri A, Hegedüs R, Jósvai J, Tóth M, Kertész K, et al. No evidence for behavioral responses to circularly polarized light in four scarab beetle species with circularly polarizing exocuticle. Physiol Behav (2012) 105:1067–1075. doi:10.1016/j.physbeh.2011.11.020
Keywords: Mueller matrix spectroscopy, ellipsometry, chirality, dual chiral structures, Protaetia mirifica, Cetoniinae
Citation: Mendoza-Galván A, Magnusson R, Jansson N, Arwin H and Järrendahl K (2024) Dual chiral structures in the cuticle of Protaetia mirifica analyzed with Mueller matrix spectroscopic ellipsometry. Front. Phys. 12:1444297. doi: 10.3389/fphy.2024.1444297
Received: 05 June 2024; Accepted: 03 July 2024;
Published: 25 July 2024.
Edited by:
Sébastien Robert Mouchet, University of Exeter, United KingdomReviewed by:
Nirmal Mazumder, Manipal Academy of Higher Education, IndiaRadu Hristu, National University of Science and Technology POLITEHNICA Bucharest, Romania
Copyright © 2024 Mendoza-Galván, Magnusson, Jansson, Arwin and Järrendahl. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kenneth Järrendahl, a2VubmV0aC5qYXJyZW5kYWhsQGxpdS5zZQ==