
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Phys., 07 July 2022
Sec. Social Physics
Volume 10 - 2022 | https://doi.org/10.3389/fphy.2022.923099
 Saulo D. S. Reis1*
Saulo D. S. Reis1* Lucas Böttcher2,3,4
Lucas Böttcher2,3,4 João P. da C. Nogueira1Geziel S. Sousa5,6Antonio S. Lima Neto5,7Hans J. Herrmann1,8,9
João P. da C. Nogueira1Geziel S. Sousa5,6Antonio S. Lima Neto5,7Hans J. Herrmann1,8,9 José S. Andrade1
José S. Andrade1Based on a data set of dengue cases in the Brazilian city of Fortaleza, collected from 2011 to 2016, we study the spatio-temporal characteristics of dengue outbreaks to characterize epidemic and non-epidemic years. First, we identify regions that show a high incidence of dengue cases and mosquito larvae in different years and analyze their corresponding correlations. We find that the characteristic correlation length of the epidemic is at least of the order of the system size, suggesting that factors such as citizen mobility may play a significant role in driving the spatial spread of the disease. We also find that in epidemic years, spatial correlations are substantially higher with a distribution skewed towards large values than non-epidemic years. Inspired by this observation, we perform a mean-field estimation of the basic reproduction number and find that the estimated values agree well with the values reported for other regions in Brazil and other countries, pointing towards similar underlying spreading mechanisms. These findings provide insights into the spreading characteristics of dengue in densely populated areas and may be relevant for improving disease containment strategies.
According to a recent report of the world health organization (WHO), over 40% of the world’s population are at the risk of a dengue infection [1, 2]. Alarmingly, in the last half-century, a 20 to 30-fold increase of dengue cases has been monitored world-wide [2]. As a vector-borne disease, the transmission of dengue fever occurs from infected to susceptible humans through female Aedes aegypti mosquitoes. Up to 12 days after symptom onset, infected individuals can transmit the virus to mosquitoes, which can further transmit the virus after an incubation period of 4–10 days [1–3]. Mosquitoes can remain alive and infectious for at least 2 weeks [1]. Symptoms include high fever, headache, vomiting, skin rash, and muscle and joint pains [1]. Aedes aegypti is also responsible for the transmission of other severe vector-borne diseases such as yellow fever, chikungunya and zika whose recent outbreaks are challenging health officials in different countries [4–6]. Vector-borne diseases are responsible for the death of more than one million humans each year, disrupt health systems, and obstruct the development of many countries [2]. As a consequence of the unavailability of vaccines and/or drug resistance, as is the case for many vector-borne diseases, control measures such as personal protection, reduction of vector breading habitats, and usage of insecticides are essential to contain outbreaks [2, 7].
Recent studies have demonstrated the poor correspondence between levels of infestation measured by entomological surveys at national and local level [8–10]. The correlation between high infestation, measured by larval entomological indicators, and risk of dengue epidemic has been shown to be weak in the most diverse scenarios [11–15]. Despite the evidence of the low usefulness and incipient positive predictive value of these larval surveys being extensively demonstrated, some countries still consider the thresholds of infestation indices as a main indicator of epidemic risk and trigger of spatially oriented vector control actions [8, 9, 15].
In fact, there is some skepticism about the suitability of larval indicators for the prediction of epidemics. Instead, several authors have suggested the systematic use of pupal indices and the direct measurement of the density of the adult mosquito, even if these imply a radical reformulation in the routines of vector control programs. They argue that the larval indices are no longer able to fulfill their main objective, which is to anticipate the transmission of the disease [8, 10, 13, 15–17]. Moreover, pupal surveys and mosquito capture are of low cost-effectiveness and inaccurate in low infestation scenarios where the mosquito has high survival due to optimal climatic conditions and other factors, increasing vector competence and causing sustained transmission of the virus with low pupal and mosquito densities.
Urban mobility has become a central concept in studies on a wide range of interests [18, 19]. In what concerns the transmission of dengue fever, its importance is even comparable to the levels of infestation and abundance of the Aedes aegypti vector [20, 21]. Several studies point out the importance of the human movement in the spatio-temporal dynamics of urban arboviruses transmitted by Aedes aegypti, in particular, dengue fever [22–24]. In 2019, the Pan American Health Organization (PAHO) published a technical document, based on the most diverse information available, reporting that the risk stratification of transmission is the best path to achieve efficient vector control [25]. Theoretically, mapping cities in micro-areas with different levels of transmission risk could make vector control strategies more effective, especially in regions with limited human and financial resources [26–28]. However, human mobility patterns are still not subject to accurate monitoring and can be crucial for the definition of high-risk spatial units that should be prioritized by the national and local programs for the control of urban arboviruses transmitted by Aedes aegypti [8].
In this work, we study the spatio-temporal characteristics of dengue outbreaks in Fortaleza from 2011–2016 to characterize epidemic and non-epidemic years. Fortaleza, the capital of Ceará state, is one of the largest cities in Brazil and is located in the North-East of the country where dengue and other neglected tropical diseases show a high prevalence [16, 29]. We show in Figure 1 that up to 1,000 dengue cases have been reported per day in Fortaleza during 2011 and 2016. In our analysis, we identify regions that exhibit a large number of dengue infections and mosquito larvae in different years, and also analyze the corresponding correlations throughout all neighborhoods of Fortaleza. We show that the characteristic length scale of correlations between case numbers at different locations in the system (i.e., the correlation length) is at least of the order of the system size. This provides support to the hypothesis that factors such as citizen mobility drive the spatial spreading of the disease. Motivated by the observation that people interact across long distances, we also use a mean-field model to compare the outbreak dynamics of two characteristic epidemic years with corresponding analyses made for other regions [30]. In particular, we perform a Bayesian Markov chain Monte Carlo parameter estimation for a mean-field susceptible-infected-recovered (SIR) model which has been previously successfully applied to the modeling of dengue outbreaks [30]. Our results provide insights into the spatio-temporal characteristics of dengue outbreaks in densely populated areas and may be relevant for making dengue containment strategies more effective.
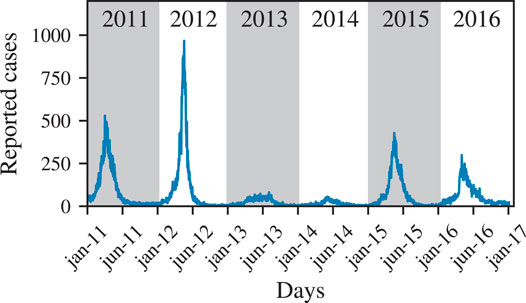
FIGURE 1. Temporal evolution of the number of dengue cases. Here, we show the number of reported dengue cases per day in Fortaleza from 2011 to 2016.
In total, we use four data sets. The first one consists of spatio-temporal information on dengue infections in the city of Fortaleza from 2011 to 2016. These data have been provided by the Epidemiological Surveillance Division of Fortaleza’s Health Secretariat, and contains both the date when an infected person reports a potential dengue infection to a physician and the corresponding geographic location with a granularity of approximately 200 m. Serology and clinical diagnosis were used to confirm dengue infections. This dataset was duly anonymized, epidemiological and clinical variables were removed as well as those cases of infected tourists and individuals who live outside the area of the city of Fortaleza. The total number of dengue cases between 2011 and 2016 in our data set after data processing is 133,540. Table 1 shows the number of cases reported for each year. The second data set contains information about Aedes aegypti larvae measurements in Fortaleza from 2011 to 2016. Measurement data is available in intervals of 2 weeks from 2,701 strategic points (SP) (e.g., junk yards, garages, buildings under construction, and warehouses) that are monitored by health authorities of Fortaleza. A SP is said to be positive if Aedes aegypti larvae have been found independent of the actual amount. Data on Aedes aegypti infestation according to each Strategic Point are available on the Fortaleza Daily Disease Monitoring System 1 and were tabulated and consolidated by the epidemiological surveillance team. Due to the influence of precipitation on the mosquito population size [31], we also consider data from three different rain gauges in Fortaleza during 2011–2016. In addition to the temporal information on dengue cases, we collected data about the geographical locations of all infection cases. Additionally, in order to investigate the influence on human mobility, we use a origin-destiny matrix generated from electronic bus pass data of Fortaleza’s transportation system for the year of 2015 [32–34].
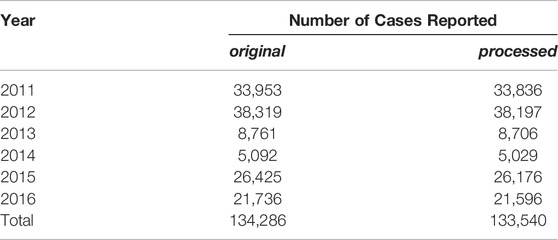
TABLE 1. Number of dengue cases reported in Fortaleza between 2011 and 2016. The processed subcolumn contains the original number of cases reported after removal of cases of infected tourists and people who live in Fortaleza metropolitan area.
The spreading dynamics of dengue in an urban evironment can be better understood in terms of underlying spatio-temporal correlations, namely, the cross-correlations between the time series of dengue incidence calculated for different pairs of city neighborhoods. We therefore now focus on spatio-temporal correlations between dengue incidence time series of different districts. In total, there are 118 neighborhoods in Fortaleza, and for each neighborhood i we use
We characterize the spatio-temporal correlations of dengue outbreaks between sites i and j by
where τ denotes a time lag, rij is the distance between neighborhood i and neighborhood j,
where
According to the Brazilian ministry of health and the health administration of the City Hall of Fortaleza, the population of Fortaleza had been at a high risk of a dengue infection during the years 2011, 2012, 2015, and 2016, being therefore classified as epidemic years [35]. The corresponding number of reported dengue cases are given in Table 1 and shown in Figure 1 (right). Due to the equatorial location of Fortaleza, the mean temperature over 1 year is 26.3 ± 0.6 °C and can be regarded as constant with only limited impact on the local dengue spreading dynamics 2. Considering the population size of 2.6 million 3, an alarmingly high number of several hundred up to almost 1,000 new infections per day have been reported from 2011 to 2016.
We illustrate the geographical distribution of dengue cases in Figure 2 from 2011 until 2016. If the total number of dengue infections in a certain area is large over the course of 1 year, this area appears in red. Areas with small numbers of dengue infections are colored blue. In 2011 and 2012 the main outbreaks occur in similar regions and cover almost the whole city except some parts in the Eastern outskirts. In the non-epidemic years 2013 and 2014, the overall dengue incidence is clearly reduced. However, some parts in the city center in the North-West that also exhibit large numbers of dengue cases in the epidemic years are still at a high risk of dengue outbreaks. Also in the epidemic years 2015 and 2016, some neighborhoods in the south show a high dengue incidence.
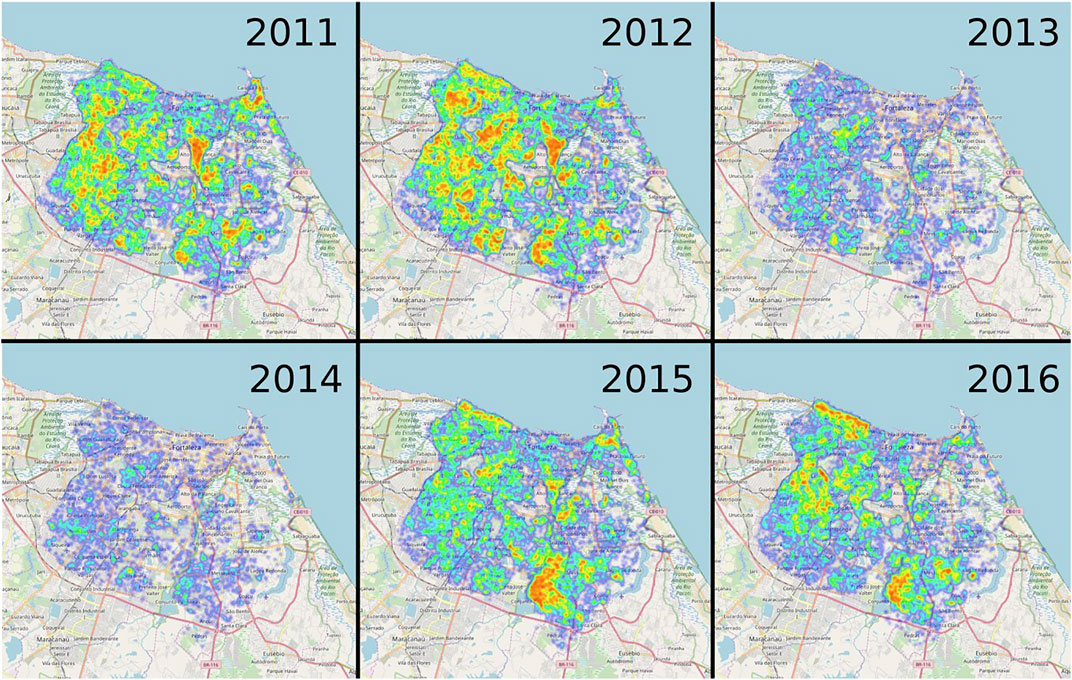
FIGURE 2. Dengue outbreaks in Fortaleza from 2011 until 2016. Heat maps of all dengue cases in Fortaleza from 2011 until 2016. Blue areas correspond to regions with a low incidence of dengue cases whereas red ones indicate large dengue outbreaks. As depicted, the heatmaps show a clear the difference between epidemic years (2011, 2012, 2015, and 2016) and non-epidemic years (2013 and 2014).
In addition to the geographical location of dengue cases, we also show a heat map of the spatial distribution of positive SPs in Figure 3. Interestingly, and in contrast to the spatial distribution of dengue cases, the majority of positive SPs are located at the same places in Fortaleza regardless of the year. In particular, the city center and some parts in the North and South-West of the town exhibit a large number of positive SPs. While according to Figure 3, the regions of major outbreaks change substantially between 2011, 2013, and 2015, the number of positive SPs does not at all. Concerning the outbreak year of 2015, the recurrent dengue outbreak and number of positive SPs seem to be even anti-correlated, since the number of dengue cases are concentrated at the southeast region of the city, a region with low incidence of positive SPs. These observations confirm the low usefulness of these larval surveys, as has been extensively demonstrated before and suggest that an effective disease intervention measure should target the neighborhoods which exhibit recurrent dengue outbreaks instead of numbers of positive SPs. In particular, the density of SPs is not homogeneous, and in fact Figure 3 is mostly reproducing the distribution of SPs, because it is common to find at least one larvae during the year and thus it is likely that most SPs will be identified as positive.
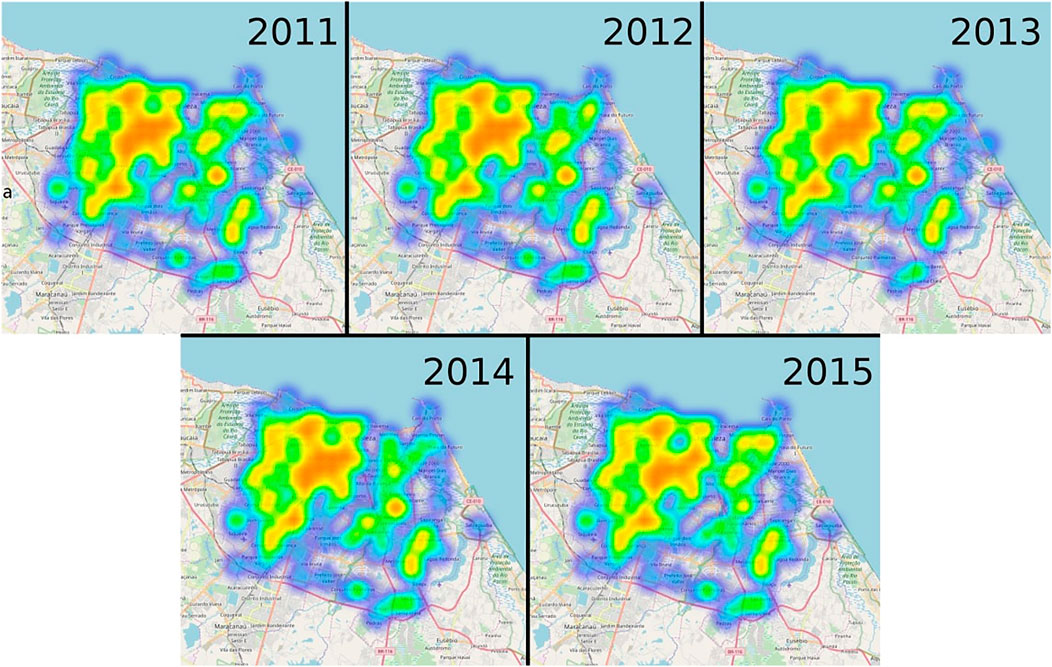
FIGURE 3. Positive SPs in Fortaleza from 2011 until 2015. Heat maps of the larvae measurement outcomes in Fortaleza from 2011 until 2015. If larvae are found, the SP measurement is said to be positive. Blue areas correspond to regions with a low number of positive SPs, whereas red ones indicate the opposite.
As shown in Figure 4, the number of reported dengue cases starts to increase in January and February just shortly after the beginning of the rain season and typically reaches its peak before July. This shows that the corresponding climate conditions facilitate the growth of mosquito populations. In addition, we also show the time evolution of the number of positive SPs. The data in Figure 4 indicates that the way the number of positive SPs changes in time is similar to the rainfall, although their relative amplitudes vary substantially from year to year.
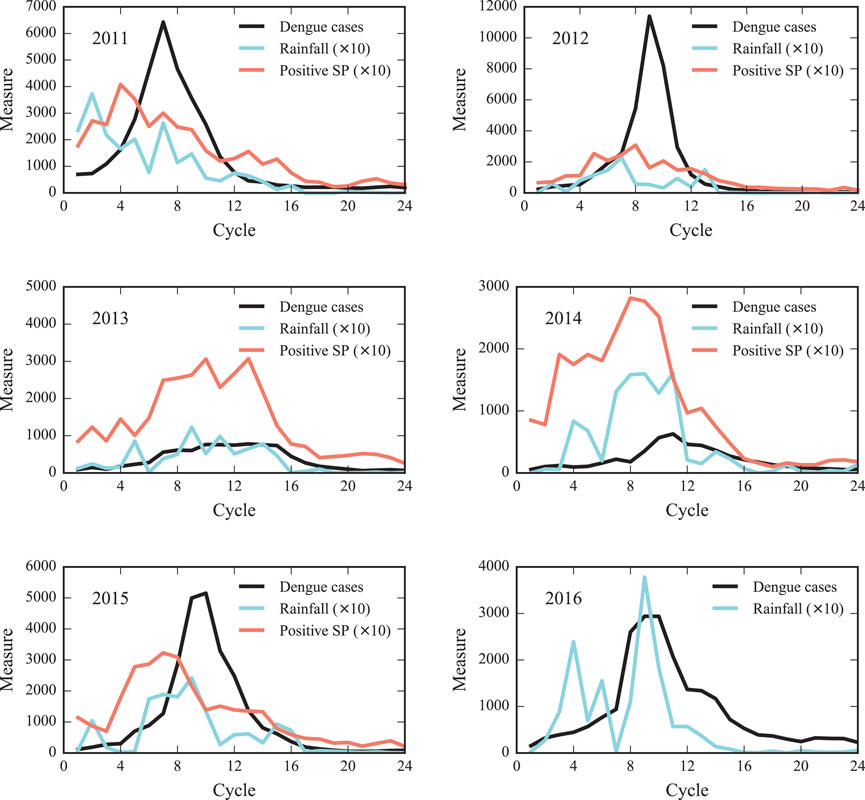
FIGURE 4. The number of reported dengue cases, positive SPs and rainfall as afunction of time from 2011 to 2016. For each year from 2011 until 2016, we show the number of reported dengue cases, positive SPs, and the precipitation level as a function of time. One time interval (cycle) is one fortnight, starting from the first fortnight of the year (cycle = 1). The number of positive SPs and the amount of rain in liters per square meter have been rescaled by a factor of 10. There is no larvae measurement data available for 2016.
In Table 2, we show the time lags τmax that correspond to the maximum correlation, as defined by Eq. 3. In the case of dengue occurrences and rainfall, we find a mean value of
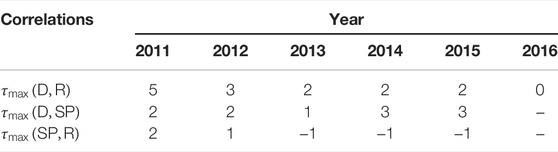
TABLE 2. The time lags from 2011 until 2016. For each year from 2011 until 2016, we compute the correlation between the number of reported dengue cases (D), positive SPs and the rainfall (R) for different time lags τ. The value of τ = 1 corresponds to one fortnight. We show the time lags that correspond to the largest correlation in the corresponding year. There is no SP data available for 2016.
Despite the very similar curve shapes, the results are less conclusive in the case of correlations between rainfall and positive SPs. In some years, the maximum number of positive SPs is found after the rainfall maximum, whereas the opposite situation occurs in other years. In any case, the time lag is zero when averaged over the 5-years reporting period, suggesting that both phenomena occur almost simultaneosly, but are subject to strong fluctuations. This can be also observed by comparing the time evolutions of rainfall and the number of positive SPs in Figure 4.
A possible explanation for this result may be that a SP measure, as provided by the health authorities of Fortaleza, do not provide a quantitative measure for the amount larvae, but only provides a binary indicator which is considered positive even if only a small amount of larvae was found at the location. Due to this qualitative nature of the SP, its positiveness indicates the spatial extent of mosquitoes and not its concentration. Therefore, correlations between rainfall and positive SPs have to be interpreted with caution. Still, the similar shapes of the positive SP and rainfall curves indicate that the spatial spread of mosquitoes is correlated with the precipitation level. The influence of climate conditions on dengue outbreaks in Brazil has also been analyzed in Ref. [31].
Dengue outbreaks that occur in a certain region of the city may lead to outbreaks in another more distant region due to human mobility [21, 36, 37]. To analyze this effect and identify characteristic correlation length-scales, we study the correlation between the dengue incidence in neighborhood i and in another neighborhood j located within a distance of rij, which is measured in kilometers. Here, rij is the distance between the barycenters of neighborhoods i and j extracted from their geographical contours. Specifically, we study the correlations between neighborhoods i and j, i.e. cij (τ, rij) as defined by Eq. 2, for different time lags τ and radii rij. According to the definition of cij (τ, rij) in Eq. 2, a correlation length ξ smaller than our considered system size would lead to an observable decay of cij (τ, rij) for rij > ξ. In all considered years, the correlations cij(τ) only vary slightly with the distance rij, as shown in Figure 5. Within the error bars of our data, we do not observe a substantial decay of cij(τ) for values of rij smaller than the system size of about 15 km. We thus conclude that the characteristic correlation length ξ is at least of the order of the system size.
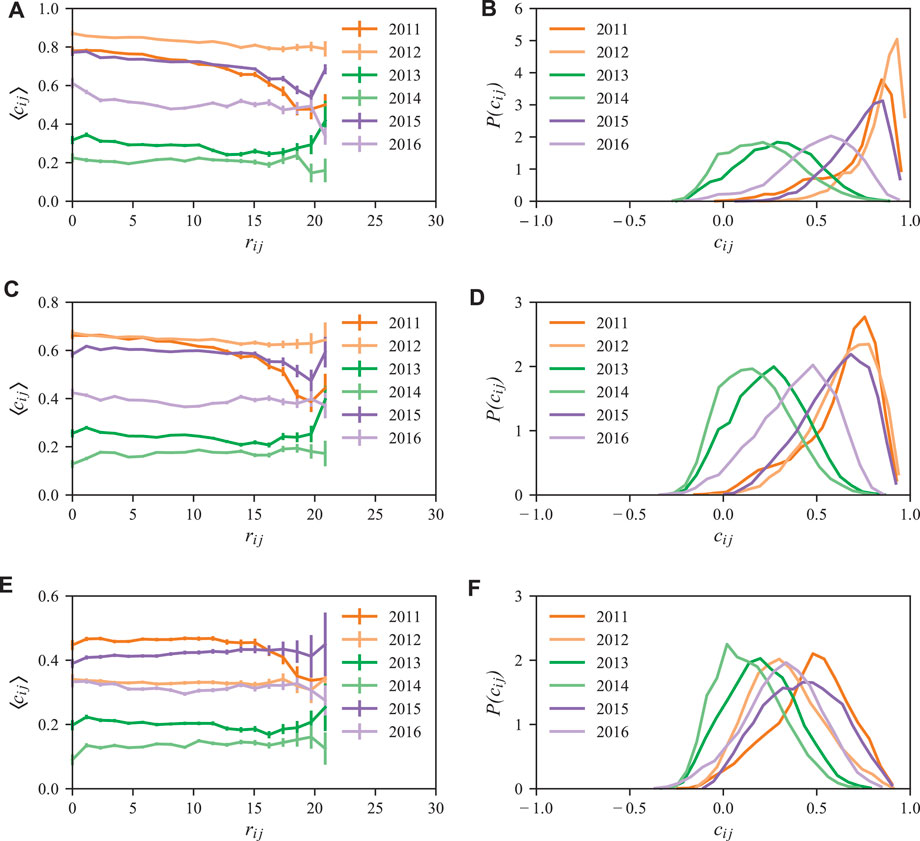
FIGURE 5. Correlations for different time lags, distances and years and their corresponding distributions. The left panels show the average of the spatio-temporal correlations ⟨cij(τ)⟩ as a function of the distance rij between neighborhoods i and j, and calculated for time lags of τ0 = 0 weeks (A,B), τ1 = 2 weeks (C,D), and τ2 = 4 weeks (E,F). As depicted, epidemic years (2011, 2012, 2015, and 2016) present a higher ⟨cij(τ)⟩ when compared to non-epidemic years (2013 and 2014). The right panels show the corresponding distribution
It is unlikely that disease vectors are responsible for such correlation effects over distances of multiple kilometers due to their limited movement capabilities. In particular, it is known that the maximum flight distance reached by the Aedes aegypti from its breeding location is of the order of approximately 100 m. Humans, however, travel through densely populated urban regions on a daily basis, and may transfer the virus to Aedes aegypti mosquitoes at different locations. This virus transfer mechanism is therefore compatible with the large correlation lengths revealed in this study and also agrees well with recent studies that suggest that human mobility is a key component of dengue spreading [21, 36, 37].
We choose three time lags, namely, τ0 = 0 weeks, τ1 = 2 weeks and τ2 = 4 weeks which are of the order of the transmission time scale of 12 days [2]. The dependence of the correlations on different time lags, distances, and years is shown in Figure 5 (left). For no time lag, τ0 = 0 weeks, or a time lag of τ1 = 2 weeks, we find that the correlations are substantially larger in the epidemic years 2011, 2012, 2015, and 2016 as compared to the non-epidemic years 2013 and 2014. However, in 2016 the correlations are less pronounced compared to other epidemic years since the outbreaks are widely distributed and their densities rather small, as shown in Figure 2. For the larger time lag, τ2 = 4 weeks, the average correlation ⟨cij(τ2)⟩ is smaller than that obtained for time lag τ1. This behavior is compatible with the fact that a time lag of 4 weeks exceeds the dengue transmission period, and we expect to find smaller correlations. In Figure 5 (right), we show the corresponding distributions of P(cij) and find that they allow to clearly distinguish between epidemic and non-epidemic years for no time lag and a time lag of τ1 = 2 weeks. In the particular case of no time lag, the distributions are strongly skewed towards larger correlations in epidemic years compared to the non-epidemic ones.
In order to better understand this result, we used an origin-destination matrix between pairs of neighborhoods of Fortaleza, made available in the mentioned Refs. [33, 34]. Precisely, this matrix was built from an extensive dataset of human mobility in the city collected during 2015, consisting of public bus validations records with smart cards of passengers. As reported in Ref. [33], the average flux of passengers fij between neighborhoods i and j is defined as,
where the brackets represent an average over 52 weeks. As shown in Figure 6A, apart from small random fluctuations, the fluxes fij show practically no dependency on the neighborhood distance rij. Similarly, the Figure 6B shows that the average spatio-temporal correlations cij remains practically invariant with fij. This result is consistent with our finding that the correlation length of the contagion process of dengue fever in Fortaleza between pairs of distinct neighborhoods should be at least larger than the system size. Furthermore, it also indicates that the human mobility at the city scale has a more profound impact on the transmission of the disease than expected since it connects the neighborhoods in such an efficient way that even the largest distances between them are too small to make a difference in the contagion process. It is interesting to compare our findings with the results obtained by Brockmann and Helbing presented in the Ref. [38], where the network-driven contagion is investigated at larger scales, such as the global viral epidemics of SARS in 2003, and MERS in 2009. In these cases, the variability in the fluxes between different regions play an important role on the disease spreading.
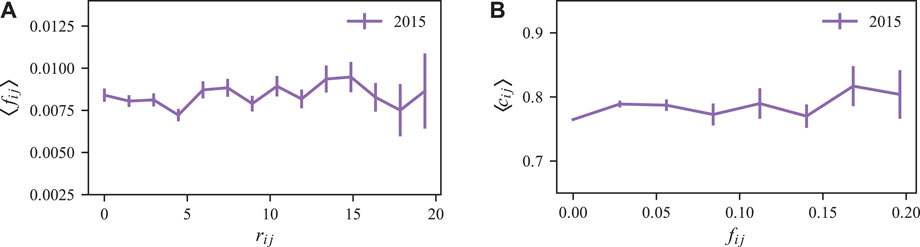
FIGURE 6. Spatio-temporal characteristics of human mobility between neighborhoods. (A) Average neighborhood-neighborhood flux ⟨fij⟩ as a function of the distance rij between neighborhoods i and j for the year of 2015. As shown, the rate flux of bus trips between neighborhoods of Fortaleza is independent of the distance between the neighborhoods. (B) The right panel shows the spatio-temporal correlations ⟨cij⟩ with τ = 0 as a function of the neighborhood-neighborhood flux fij for the year of 2015. As presented, the spatio-temporal correlations of dengue cases have no significant dependence on fij. In both panels, the bars are the standard deviations.
As described in the previous section, the studied dengue outbreaks in Fortaleza exhibit a correlation length ξ which is at least of the order of the underlying system size. Based on this observation, we can apply a mean-field epidemic model, as an approximation, to further characterize the observed spreading dynamics. In particular, we aim at comparing the disease transmission parameters of dengue outbreaks in Fortaleza with the results of previous studies [30]. To do so, we consider the two epidemic years 2012 and 2015 and perform a Bayesian Markov chain Monte Carlo parameter estimation for an SIR model that has been previously applied in the context of dengue outbreaks in Thailand [30]. More details about the methodology are presented in Supplementary Material.
We determine the basic reproduction number R0 of dengue outbreaks in Fortaleza and compare it with the values of other disease outbreaks. The basic reproduction number is an important epidemiological measure and is defined as the average number of secondary cases originating from one infectious individual during the initial outbreak period (i.e., in a fully susceptible population) [39]. Different models exist to characterize and predict epidemic outbreaks [39–43]. In the case of dengue, some models explicitly incorporate a mosquito population, whereas others take into account such effects by using an effective spreading rate [30, 44]. According to Ref. [30], an explicit treatment of vector populations just increases the number of modeling parameters and may not lead to a better agreement between model and data. We therefore do not explicitly model a vector population and consider an SIR model with an effective spreading rate which has been found to capture the essential features of dengue outbreaks in Thailand [30]. The governing equations are.
where N = S + I + R is the human population size and S = S(t), I = I(t), R = R(t) denote the numbers of susceptible, infected, and recovered individuals at time t, respectively. The human mortality and recovery rates are denoted by μH and γH, respectively. The term μHN in Eq. 5 corresponds to the birth rate and is chosen such that the population size is kept constant. Furthermore, the composite human-to-human transmission rate β accounts for transitions from susceptible to infected. The relation
connects β with the mosquito-to-human transmission rate per bite βH, human-to-mosquito transmission rate βV, number of mosquitoes per person m, mean rate of bites per mosquito c, and mosquito mortality rate μV [30]. The basic reproduction number of the described SIR model variant is [39].
For R0 > 1 there exists a stable endemic state, whereas the disease-free equilibrium is stable for R0 ≤ 1. In addition to Eqs. 5–7, 10, the cumulative number of reported dengue cases C = C(t) evoles according to
where p is the fraction of infected individuals that were diagnosed with dengue and reported to the health officials. As mentioned above, Fortaleza has a population of N = 2.6 million. We use a mortality rate of μH = 1/76 y−1 in accordance with the latest World Bank life expectancy estimates 4.
A parameter estimation based on Eqs. 5–7, 10 means that we have to determine the parameters β, γH, p and the initial fraction of recovered individuals r0 = R(0)/N that best describe the observed dengue outbreaks. To do so, we use Bayes’ theorem to determine the posterior parameter distribution
based on the likelihood function
To model one complete outbreak period, we use November as the starting month in our simulations since the precipitation level and case numbers then typically reach a minimum. In this way, we can compare our results with those of Ref. [30]. In Table 3, we summarize the inferred model parameters, median values, and 95% confidence intervals of the posterior distributions. In addition, we also present the maximum likelihood (ML) estimates which we use to compare the SIR model with the actual dengue case data in Figure 7. We find good agreement between the model predictions and the reported numbers of dengue cases. Only the dengue outbreak peak between April and June 2012 is difficult to capture due to an overwhelming number of up to 1,000 new infections per day.
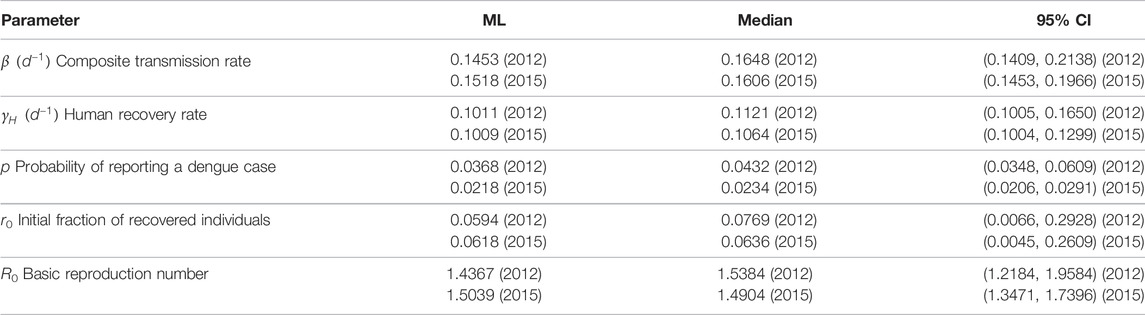
TABLE 3. Posterior parameter estimations. Based on uniform prior SIR parameter distributions, the corresponding posterior distributions for the dengue outbreaks in 2012 and 2015 have been obtained using Bayesian Markov chain Monte Carlo sampling. For both years an additional maximum likelihood (ML) parameter estimate is given. The posterior distributions are characterized by their median values and their 95% confidence intervals (CI).
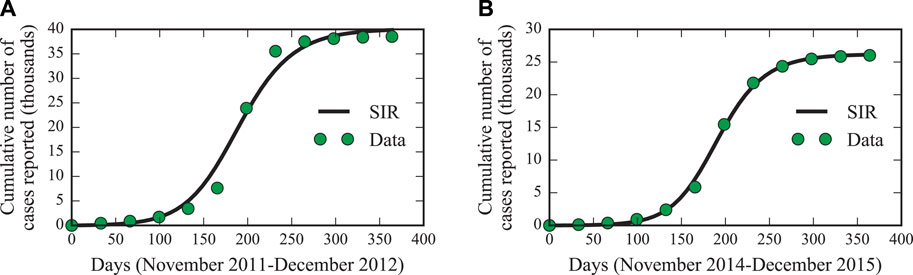
FIGURE 7. SIR model fit for the epidemic years 2012 and 2015. The maximum likelihood fits of the SIR model (black solid lines) as defined by Eqs. 5–7, 10 for the epidemic years 2012 (A) and 2015 (B). The cumulative number of reported dengue cases according to our data set is indicated by green dots.
We now compare the obtained estimates with those of Ref. [30] which focuses on dengue outbreaks in Thailand. In Ref. [30], the number of reported dengue hemorrhagic fever (DHF) cases, a more severe form of dengue fever, has been used, while we considered the number of all reported dengue cases. The total number of mentioned DHF cases in Thailand is roughly 75,000 with a total population size of 46.8 million in 1984 [30]. In contrast to these results, the number of reported dengue cases in Fortaleza in 2012 is almost 40,000 with a total population size of 2.5 million. The attack rate in Fortaleza is thus about one order of magnitude larger than that in Thailand. This alarming difference may be partially a result of the different definitions of reported cases (“DHF” versus “dengue fever”) as well as due to sub-notification, but it also points out to the need of better control measures to contain the outbreaks in Fortaleza. Overall, the parameter estimation for the epidemic years 2012 and 2015 leads to values in a similar range compared to the results of Ref. [30]. The ML estimates of the probability of reporting a dengue case, p, and the initial fraction of recovered individuals, r0, are by a factor 2–3 larger in our parameter estimation. On the other hand, the ML estimates of β and γ are slightly smaller in Fortaleza. According to our statistical analysis, the basic reproduction number as defined in Eq. 9 (basically the fraction of β and γH) is 1.44 (2012) and 1.50 (2015). These ML estimates are about 30–40% larger than the corresponding ML estimate reported for the aforementioned dengue outbreak in Thailand [30].
The re-emergence of dengue and other neglected tropical diseases is a major threat to public health in different countries. Various factors such as environmental conditions and human mobility affect the spread of dengue [21, 31, 36, 37], making it difficult to contain an outbreak [45, 46].
We analyzed recent dengue outbreaks in Fortaleza, one of the largest Brazilian cities, and identified regions which exhibit a large number of dengue infections and Aedes aegypti larvae over different years. Our results show that the characteristic length scale of correlations between the number of cases at different locations is at least of the order of the system size. Using data from bus transportation between pairs of neighborhoods in the city, we propose that a plausible explanation for this observation is that the human mobility can affect the dissemination of the virus. Indeed, human movement seems to be a factor in the transmission dynamics, particularly in large tropical cities where successive dengue epidemics have been recorded. Interestingly, there is an extensive literature on the investigation of spatial correlations of events in urban areas in terms of clustering and percolation transition (see, for example, Refs. [47, 48]). In these studies, the correlations between pairs of sites usually decay with their geographical distance. In the present case, due to urban mobility, the correlation length of the contagion process is at least of the order of the system size, leading to the observed flat behavior of the correlations with distance.
We also compared the disease transmission characteristics of dengue outbreaks in Fortaleza with those reported for Thailand [30]. The inferred basic reproduction number of dengue outbreaks in Fortaleza is about 30–40% larger than the ML estimate reported in Ref. [30]. Moreover, we classified epidemic and non-epidemic years based on an analysis of spatio-temporal correlations and their corresponding distributions. We found that for non-epidemic years the distribution of spatial correlations is strongly skewed towards strong correlations in epidemic years. This means that in an epidemic year, spatial correlations are typically of the same size, pointing towards recurrent connections between neighborhoods. By comparing spatial correlations between epidemic and non-epidemic years, we found that in epidemic years spatial correlations are higher by a factor of about three as compared to non-epidemic years. This shows that during an epidemic the outbreaks do not occur independently in different neighborhoods but affect each other. Using spatial-correlation analyses may also allow to predict at an early stage if a year tends to be epidemic or not.
Publicly available datasets were analyzed in this study. This data can be found here: https://github.com/rafaellpontes/dengue_mobility_paper.
All authors designed research. GS and AL collected and prepared the Dengue data sets. SS, LB, and JN processed the Dengue data. All authors analyzed and interpreted the Dengue data. SS, LB, AL, HH, and JA wrote the manuscript. All authors read and approved the final manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We acknowledge financial support from the Brazilian agencies CNPq, CAPES, the FUNCAP, and from the National Institute of Science and Technology for Complex Systems (INCT-SC) in Brazil. LB acknowledges financial support from the SNF Early Postdoc. Mobility fellowship on “Multispecies interacting stochastic systems in biology” and the Army Research Office (W911NF-18-1-0345).
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphy.2022.923099/full#supplementary-material
1Fortaleza Daily Disease Monitoring System (SIMDA). https://pt.climate-data.org/america-do-sul/brasil/ceara/fortaleza-2031/, Retrieved 3 June 2020.
2[Dataset] https://pt.climate-data.org/america-do-sul/brasil/ceara/fortaleza-2031/, Retrieved 5 December 2020.
3[Dataset] https://cidades.ibge.gov.br/brasil/ce/fortaleza/panorama, Retrieved 6 June 2021.
4[Dataset] https://data.worldbank.org/indicator/SP.DYN.LE00.IN, Retrieved 5 December 2020.
1.World Health Organization. Dengue: Guidelines for Diagnosis, Treatment, Prevention and Control. Geneva: World Health Organization (2009).
2.World Health Organization. Sustaining the Drive to Overcome the Global Impact of Neglected Tropical Diseases. Geneva: World Health Organization (2013).
3.World Health Organization. A Global Brief on Vector-Borne Diseases. Geneva: World Health Organization (2014).
4. Barrett ADT, Higgs S. Yellow Fever: a Disease that Has yet to Be Conquered. Annu Rev Entomol (2007) 52:209–29. doi:10.1146/annurev.ento.52.110405.091454
5. Charrel RN, de Lamballerie X, Raoult D. Chikungunya Outbreaks - the Globalization of Vectorborne Diseases. N Engl J Med (2007) 356:769–71. doi:10.1056/nejmp078013
6. Brasil P, Pereira JP, Moreira ME, Ribeiro Nogueira RM, Damasceno L, Wakimoto M. Zika virus infection in pregnant women in rio de janeiro. N Engl J Med (2016) 375:2321–34. doi:10.1056/nejmoa1602412
7. Hotez PJ, Bottazzi ME, Franco-Paredes C, Ault SK, Periago MR. The Neglected Tropical Diseases of Latin america and the Caribbean: a Review of Disease burden and Distribution and a Roadmap for Control and Elimination. Plos Negl Trop Dis (2008) 2:e300. doi:10.1371/journal.pntd.0000300
8. Enslen AW, Lima Neto AS, Castro MC. Infestation Measured by aedes Aegypti Larval Surveys as an Indication of Future Dengue Epidemics: an Evaluation for Brazil. Trans Roy Soc Trop Med Hyg (2020) 114:506–12. doi:10.1093/trstmh/traa021
9. MacCormack-Gelles B, Neto ASL, Sousa GS, do Nascimento OJ, Castro MC. Evaluation of the Usefulness of aedes Aegypti Rapid Larval Surveys to Anticipate Seasonal Dengue Transmission between 2012–2015 in Fortaleza, Brazil. Acta Trop (2020) 205:105391. doi:10.1016/j.actatropica.2020.105391
10. Sanchez L, Vanlerberghe V, Alfonso L, del Carmen Marquetti M, Guzman MG, Bisset J, et al. Aedes aegypti Larval Indices and Risk for Dengue Epidemics. Emerg Infect Dis (2006) 12:800. doi:10.3201/eid1205.050866
11. Cromwell EA, Stoddard ST, Barker CM, Van Rie A, Messer WB, Meshnick SR, et al. The Relationship between Entomological Indicators of aedes Aegypti Abundance and Dengue Virus Infection. PLoS Neglect Trop Dis (2017) 11:e0005429. doi:10.1371/journal.pntd.0005429
12. Focks DA, Chadee DD. Pupal Survey: an Epidemiologically Significant Surveillance Method for aedes Aegypti: an Example Using Data from trinidad. Am J Trop Med Hyg (1997) 56:159–67. doi:10.4269/ajtmh.1997.56.159
13. Focks DA. A Review of Entological Sampling gMethods and Indicatiors for Dengue Vectors. Techinical report. Geneva: Tech. rep., World Health Organization (2004).
14. Gubler DJ. Dengue and Dengue Hemorrhagic Fever. Clin Microbiol Rev (1998) 11:480–96. doi:10.1128/cmr.11.3.480
15. Bowman LR, Runge-Ranzinger S, McCall P. Assessing the Relationship between Vector Indices and Dengue Transmission: a Systematic Review of the Evidence. PLoS Neglect Trop Dis (2014) 8:e2848. doi:10.1371/journal.pntd.0002848
16. MacCormack-Gelles B, Neto ASL, Sousa GS, Nascimento OJ, Machado MM, Wilson ME, et al. Epidemiological Characteristics and Determinants of Dengue Transmission during Epidemic and Non-epidemic Years in Fortaleza, brazil: 2011-2015. PLoS Neglect Trop Dis (2018) 12:e0006990. doi:10.1371/journal.pntd.0006990
17. Tun-Lin W, Kay B, Barnes A, Forsyth S. Critical Examination of aedes Aegypti Indices: Correlations with Abundance. Am J Trop Med Hyg (1996) 54:543–7. doi:10.4269/ajtmh.1996.54.543
18. Jusup M, Holme P, Kanazawa K, Takayasu M, Romić I, Wang Z, et al. Social Physics. Phys Rep (2022) 948:1–148. doi:10.1016/j.physrep.2021.10.005
19. Helbing D, Brockmann D, Chadefaux T, Donnay K, Blanke U, Woolley-Meza O, et al. Saving Human Lives: What Complexity Science and Information Systems Can Contribute. J Stat Phys (2015) 158:735–81. doi:10.1007/s10955-014-1024-9
20. Bouzid M, Brainard J, Hooper L, Hunter PR. Public Health Interventions for aedes Control in the Time of Zikavirus–A Meta-Review on Effectiveness of Vector Control Strategies. PLoS Neglect Trop Dis (2016) 10:e0005176. doi:10.1371/journal.pntd.0005176
21. Stoddard ST, Forshey BM, Morrison AC, Paz-Soldan VA, Vazquez-Prokopec GM, Astete H, et al. House-to-house Human Movement Drives Dengue Virus Transmission. Proc Natl Acad Sci USA (2013) 110:994–9. doi:10.1073/pnas.1213349110
22. Vazquez-Prokopec GM, Bisanzio D, Stoddard ST, Paz-Soldan V, Morrison AC, Elder JP, et al. Using Gps Technology to Quantify Human Mobility, Dynamic Contacts and Infectious Disease Dynamics in a Resource-Poor Urban Environment. PLoS ONE (2013) 8:e58802. doi:10.1371/journal.pone.0058802
23. Vazquez-Prokopec GM, Kitron U, Montgomery B, Horne P, Ritchie SA. Quantifying the Spatial Dimension of Dengue Virus Epidemic Spread within a Tropical Urban Environment. PLoS Neglect Trop Dis (2010) 4:e920. doi:10.1371/journal.pntd.0000920
24. Vazquez-Prokopec GM, Stoddard ST, Paz-Soldan V, Morrison AC, Elder JP, Kochel TJ, et al. Usefulness of Commercially Available Gps Data-Loggers for Tracking Human Movement and Exposure to Dengue Virus. Int J Health Geogr (2009) 8:68. doi:10.1186/1476-072x-8-68
25.[Dataset] Pan American Health Organization. Technical Document for the Implementation of Interventions Based on Generic Operational Scenarios for aedes Aegypti Control. Washington, D.C.: PAHO (2019).
26.World Health Organization. Handbook for Integrated Vector Management. Geneva: World Health Organization (2012).
27. Quintero J, Brochero H, Manrique-Saide P, Barrera-Pérez M, Basso C, Romero S, et al. Ecological, Biological and Social Dimensions of Dengue Vector Breeding in Five Urban Settings of Latin america: a Multi-Country Study. BMC Infect Dis (2014) 14:38. doi:10.1186/1471-2334-14-38
28. Caprara A, Lima JWO, Marinho ACP, Calvasina PG, Landim LP, Sommerfeld J. Irregular Water Supply, Household Usage and Dengue: a Bio-Social Study in the Brazilian Northeast. Cad Saude Publica (2009) 25:S125–S136. doi:10.1590/s0102-311x2009001300012
29. Lindoso JAL, Lindoso AAB. Neglected Tropical Diseases in Brazil. Rev Inst Med Trop Sao Paulo (2009) 51:247–53. doi:10.1590/s0036-46652009000500003
30. Pandey A, Mubayi A, Medlock J. Comparing Vector–Host and Sir Models for Dengue Transmission. Math Biosci (2013) 246:252–9. doi:10.1016/j.mbs.2013.10.007
31. Stolerman LM, Maia PD, Kutz JN. Forecasting Dengue Fever in Brazil: An Assessment of Climate Conditions. PLoS One (2019) 14:e0220106. doi:10.1371/journal.pone.0220106
32. Caminha C, Furtado V, Pinheiro V, Silva C. Micro-interventions in Urban Transportation from Pattern Discovery on the Flow of Passengers and on the Bus Network. In: 2016 IEEE International Smart Cities Conference (ISC2). Trento, Italy: IEEE (2016). p. 1–6. doi:10.1109/isc2.2016.7580776
33. Caminha C, Furtado V, Pequeno TH, Ponte C, Melo HP, Oliveira EA, et al. Human Mobility in Large Cities as a Proxy for Crime. PLoS ONE (2017) 12:e0171609. doi:10.1371/journal.pone.0171609
34. Bomfim R, Pei S, Shaman J, Yamana T, Makse HA, Andrade JS, et al. Predicting Dengue Outbreaks at Neighbourhood Level Using Human Mobility in Urban Areas. J R Soc Interf (2020) 17:20200691. doi:10.1098/rsif.2020.0691
35.Ministério da Saúde do Brasil. Diretrizes nacionais para prevenção e controle de epidemias de dengue (2009). Available from: https://bvsms.saude.gov.br/bvs/publicacoes/diretrizes_nacionais_prevencao_controle_dengue.pdf (Accessed June 8, 2017).
36. Reiner RC, Stoddard ST, Scott TW. Socially Structured Human Movement Shapes Dengue Transmission Despite the Diffusive Effect of Mosquito Dispersal. Epidemics (2014) 6:30–6. doi:10.1016/j.epidem.2013.12.003
37. Stone CM, Schwab SR, Fonseca DM, Fefferman NH. Human Movement, Cooperation and the Effectiveness of Coordinated Vector Control Strategies. J R Soc Interf (2017) 14:20170336. doi:10.1098/rsif.2017.0336
38. Brockmann D, Helbing D. The Hidden Geometry of Complex, Network-Driven Contagion Phenomena. Science (2013) 342:1337–42. doi:10.1126/science.1245200
39. Keeling MJ, Rohani P. Modeling Infectious Diseases in Humans and Animals. USA: Princeton University Press (2008).
40. Böttcher L, Woolley-Meza O, Araújo NAM, Herrmann HJ, Helbing D. Disease-induced Resource Constraints Can Trigger Explosive Epidemics. Sci Rep (2015) 5:16571. doi:10.1038/srep16571
41. Böttcher L, Woolley-Meza O, Goles E, Helbing D, Herrmann HJ. Connectivity Disruption sparks Explosive Epidemic Spreading. Phys Rev E (2016) 93:042315. doi:10.1103/PhysRevE.93.042315
42. Böttcher L, Nagler J, Herrmann HJ. Critical Behaviors in Contagion Dynamics. Phys Rev Lett (2017) 118:088301. doi:10.1103/PhysRevLett.118.088301
43. Böttcher L, Andrade JS, Herrmann HJ. Targeted Recovery as an Effective Strategy against Epidemic Spreading. Sci Rep (2017) 7:14356. doi:10.1038/s41598-017-14763-5
44. Fitzgibbon W, Morgan J, Webb G. An Outbreak Vector-Host Epidemic Model with Spatial Structure: the 2015–2016 Zika Outbreak in Rio De Janeiro. Theor Biol Med Model (2017) 14:7. doi:10.1186/s12976-017-0051-z
45. Guzzetta G, Marques-Toledo CA, Rosà R, Teixeira M, Merler S. Quantifying the Spatial Spread of Dengue in a Non-endemic Brazilian metropolis via Transmission Chain Reconstruction. Nat Commun (2018) 9:2837. doi:10.1038/s41467-018-05230-4
46. Antonio FJ, Itami AS, de Picoli S, Teixeira JJV, dos Santos Mendes R. Spatial Patterns of Dengue Cases in Brazil. PLoS ONE (2017) 12:e0180715. doi:10.1371/journal.pone.0180715
47. Gallos LK, Barttfeld P, Havlin S, Sigman M, Makse HA. Collective Behavior in the Spatial Spreading of Obesity. Sci Rep (2012) 2:1–9. doi:10.1038/srep00454
Keywords: dengue fever, Bayesian inference, disease spreading, mathematical modelling, data analysis
Citation: S. Reis SD, Böttcher L, Nogueira JPdC, Sousa GS, Lima Neto AS, Herrmann HJ and Andrade JS (2022) Spatio-Temporal Characteristics of Dengue Outbreaks. Front. Phys. 10:923099. doi: 10.3389/fphy.2022.923099
Received: 18 April 2022; Accepted: 06 June 2022;
Published: 07 July 2022.
Edited by:
Haroldo V. Ribeiro, State University of Maringá, BrazilReviewed by:
Matjaž Perc, University of Maribor, SloveniaCopyright © 2022 S. Reis, Böttcher, Nogueira, Sousa, Lima Neto, Herrmann and Andrade. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Saulo D. S. Reis, c2F1bG9AZmlzaWNhLnVmYy5icg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.