 Yuquan Chen
Yuquan Chen Derek Hollenbeck
Derek Hollenbeck Yong Wang
Yong Wang YangQuan Chen
YangQuan Chen
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Phys., 04 October 2018
Sec. Statistical and Computational Physics
Volume 6 - 2018 | https://doi.org/10.3389/fphy.2018.00111
This article is part of the Research TopicFractional Calculus and its Applications in PhysicsView all 10 articles
Optimal random foraging strategy has gained increasing concentrations. It is shown that Lévy flight is more efficient compared with the Brownian motion when the targets are sparse. However, standard Lévy flight generally cannot be followed in practice. In this paper, we assume that each flight of the forager is possibly interrupted by some uncertain factors, such as obstacles on the flight direction, natural enemies within the vision distance, and restrictions in the energy storage for each flight, and introduce the tempered Lévy distribution p(l) ~ e−ρll−μ. It is validated by both theoretical analyses and simulation results that a higher searching efficiency can be derived when a smaller ρ or μ is chosen. Moreover, by taking the flight time as the waiting time, the master equation of the random searching procedure can be obtained. Interestingly, we build two different types of master equations: one is the standard diffusion equation and the other one is the tempered fractional diffusion equation.
One common approach to the animal movement patterns is to use the scheme of optimizing random search [1–3]. In a random search model, single or multiple individuals search a landscape to find targets whose locations are not known a priori, which is usually adopted to describe the scenario of animals foraging for food, prey or resources. The locomotion of the individual has a certain degree of freedom which is characterized by a specific search strategy such as a type of random walk and is also subject to other external or internal constraints, such as the environmental context of the landscape or the physical and psychological conditions of the individual. It is assumed that a strategy that optimizes the search efficiency can evolve in response to such constraints on a random search, and the movement is a consequence of the optimization on random search.
Many researchers have concentrated on the study of different animals' foraging movements. It is shown that when the environment contains a high density of food items, foragers tend to adopt Brownian walks, characterized by a great number of short step lengths in random directions that maintain foragers in a small portion of the available space [4, 5]. In contrast, when the density of food items is low, individuals tend to exhibit Lévy flights, where larger step lengths occasionally occur and relocate the foragers in the environment. Due to the fact that the density of food items is often low, many animals behave a Lévy flight when foraging and their movements have been found to fit closely to a Lévy distribution (power law distribution) with an exponent close to 2 [6, 7]. For instance, the foraging behavior of the wandering albatross on the ocean surface was found to obey a power law distribution [8]; the foraging patterns of a free-ranging spider monkey in the forests was also found to be a power law tailed distribution of steps consistent with Lévy walks [9, 10].
On this basis, researchers mainly consider two issues: one is to model the foraging behavior as a Lévy flight and the other one is to study the searching efficiency theoretically or experimentally. It is assumed that the forager takes a random walk according to the distribution p(l) ~ l−μ, 1 < μ < 3. Then it is proven that the highest searching efficiency can be obtained when μ is close to 2 for the non-destructive case (the same target site can be visited many times). While the searching efficiency is higher when μ tends to 1 for the destructive case (the target site found by the forager becomes undetectable in subsequent flights). Later, many more complex situations are considered. Due to the fact that foragers are always searching in a bounded area, Dybiec et al. [11] and Zhao et al. [12] studied the searching efficiency of Lévy flight in a bounded area. Kerster et al. [5] took the spatial memory of foragers into consideration and concluded that the spatial information influenced the foraging behavior significantly according to the experimental results. Interestingly, it was claimed that the Lévy flight foraging behavior can also be interpreted by a composite search model [13, 14]. The model consists of an intensive search phase, followed by an extensive phase, if no food is found in the intensive phase. Particularly, Zeng and Chen [15] considered the waiting time between two successive flights and formulated the master equation for such foraging behavior.
Though many studies have proven that it is usually more efficient to utilize Lévy flight foraging strategy, standard Lévy flight cannot be followed in practice because of many uncertain factors. For instance, the forager may encounter obstacles or natural enemies and extremely large flight distance cannot be reasonable due to the restriction of the forager's flight ability. In this paper, we take these conditions into consideration and temper the Lévy distribution with an exponential decaying function, which results in a tempered Lévy distribution p(l) ~ e−ρll−μ. It is then shown that a higher searching efficiency will be derived when a smaller ρ or μ is chosen, both by simulation and theoretical analyses. Further, two different types of master equations are derived: one is the standard diffusion equation and the other one is the tempered fractional diffusion equation. Since the first and second order moments exist, the foraging movement will finally result in a Gaussian motion, which indicates that the tempered fractional diffusion equation is in fact another expression for the standard diffusion.
The remainder of the paper is organized as follows. Section 2 provides the basic foraging model and some basic results are also given. In section 3, we study the searching efficiency when a tempered Lévy distribution is considered. Two different types of master equations are derived in section 4 after treating the flight time as the waiting time. The paper is concluded in section 5.
In this section, we mainly recall the original model and basic results of Lévy flight optimal random search. Assume that target sites are uniformly distributed and the forager behaves as follows
(1) If a target site lies within a “direct vision” distance rv, then the forager moves on a straight line to the nearest site. A finite value of rv, no matter how large, models the constraint that no forager can detect a target site located an arbitrarily large distance away.
(2) If there is no target site within a distance rv, then the forager chooses a direction uniformly and a distance lj from a probability distribution. It then incrementally moves to the new point, constantly looking for a target within a radius rv along its way. If it does not detect a target, it stops after traversing the distance lj and chooses a new direction and a new distance lj+1; otherwise, it proceeds to the target as rule (1).
In the case of non-destructive foraging, the forager can visit the same target site many times. In the case of destructive foraging, the target site found by the forager becomes undetectable in subsequent flights. Let λ be the mean free path of the forager between two successive target sites [for two dimensions where ϕ is the target-site area density].
On the basis of above behaviors, assume that the flight distance is distributed as the Lévy distribution
As shown in Figure 1 where η is the searching efficiency defined as (7), researchers find that μ ≈ 2 and μ → 1 will result in an optimal searching efficiency for the non-destructive case and destructive case, respectively. For more details about the model and existing results, one may refer to the works of Viswanathan et al. [6, 7] and references therein.
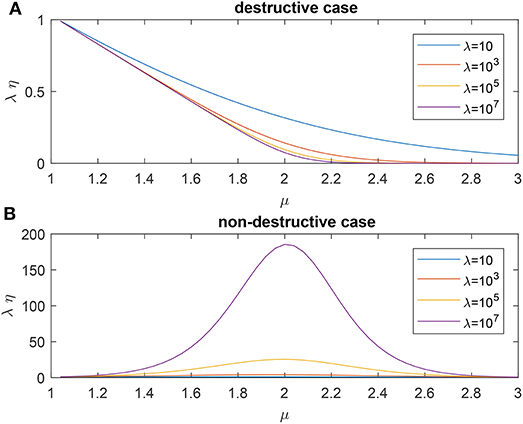
Figure 1. Searching efficiency of standard Lévy flight for different mean free path λ: (A) the destructive case, (B) the nondestructive case.
In almost all the existing literatures about Lévy flight foraging, it is assumed that the flight distance at each step is independently distributed as (1). Distribution (1) is power-law decaying, which indicates that a large jump length will appear more frequently compared with the traditional Gaussian distribution. In practical foraging, after the forager determines the flight distance at some step, the flight will be interrupted by some unknown reasons, such as obstacles on the flight direction, natural enemies in the vision distance, and restrictions in the energy storage for each flight. Because of these reasons, we can assume that the flight distance is distributed as
which indicates that the forager can keep the flight direction with the probability of an exponential distribution. Figures 2, 3 show the probability density function (pdf) of a tempered Lévy distribution for different μ and ρ respectively. One can find that the density decreases slower with a smaller μ or a smaller ρ, which means that a larger jump length is more likely to happen. Particularly, the μ = 0 case in Figure 2 is included to show that the tempered Lévy distribution always has a shorter tail than the pure μ = 0 Lévy distribution. The ρ = 0 case in Figure 3 is the Lévy distribution which has a heavier tail compared with other cases.
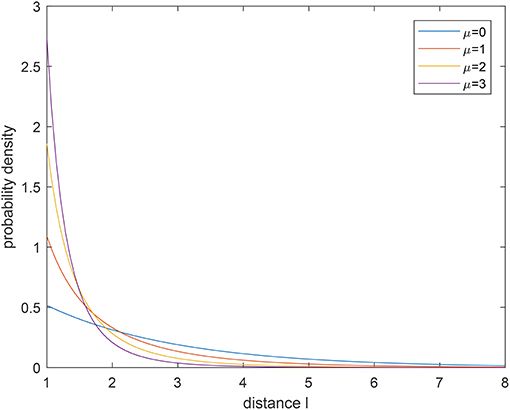
Figure 2. Probability density for tempered Lévy distribution with ρ = 0.5 for different μ.
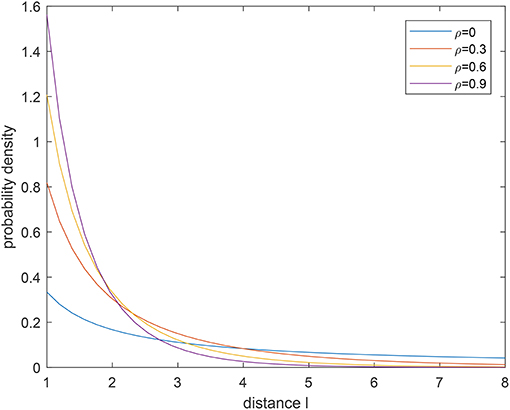
Figure 3. Probability density for tempered Lévy distribution with μ = 1 for different ρ.
Remark 1: The difference between (1) and (2) is that the power law distribution is tempered by an exponential decaying e−ρl. The exponential part e−ρl can be viewed as the probability density that the forager can keep its flight direction before he completes one flight in the existence of some unknown factors and ρ is determined by the environment. Because Lévy distribution is now tempered by e−ρl, the first and second order moments of distribution (2) exist for arbitrary μ ∈ ℝ. In the paper, we will discuss the problem in a wider range μ ∈ [1, ∞) rather than (1, 3) for the Lévy distribution.
In this part, we will borrow the idea from Viswanathan et al. [6] to optimize the searching efficiency. Given the pdf of the flight distance as (2), the mean flight distance can be calculated as
where, the incomplete gamma function Γup is defined as
Remark 2: In Viswanathan et al. [6], the Lévy distribution is truncated by the mean free path λ because it is assumed that the forager must find a target after flight for distance λ. Different from the idea in Viswanathan et al. [6], we assume that the flight distance may be truncated according to an exponential distribution which is used to describe the probability of encountering some uncertain factors. In this paper, the truncation is also considered when calculating the mean flight distance since the jump length may be larger than the mean free path. We have also to declare that the mean free path is generally very large since the targets are sparse and the decaying speed of tempered Lévy distribution is much faster. Thus, the integral of is very small such that it almost has no influence on the searching efficiency.
Let N be the mean number of flights taken by a Lévy forager while traveling between two successive target sites. Since the first and second order moments of tempered Lévy distribution exist, the trajectory of the forager will result in a Brownian motion. According to the existing results by Viswanathan et al. [6], for the non-destructive case, it follows that the mean flight number between two successive targets can be estimated as
where, D is the diffusion constant. According to the standard diffusion equation in section 4, it is found that the diffusion constant , where a is the second order moment of flight distance and b is the mean of the waiting time. Since we do not take the time into consideration, one can conclude that Nn is proportional to . Here, a can be calculated as
Based on the above analyses, we can then calculate the searching efficiency which is defined as
Take rv as 1 when simulating and the results for different mean free path λ are shown in Figure 4. Following observations can be drawn
(1) For fixed mean free path λ and ρ, a smaller μ will result in a higher searching efficiency.
(2) For fixed mean free path λ and μ, a smaller ρ will result in a higher searching efficiency.
(3) The mean free path λ almost has no influence on the choice of μ and ρ to derive the highest searching efficiency.
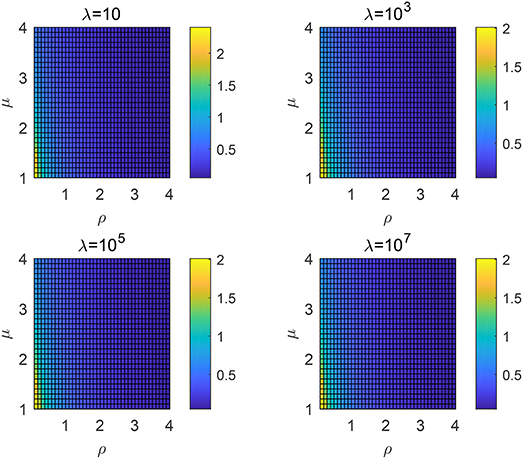
Figure 4. Searching efficiency ηλ for different order μ and ρ: the non-destructive case with different λ.
As interpreted in the existing papers, the Lévy distribution can lead to a higher efficiency in a sparse area due to the higher probability of large jump lengths. For this issue, a smaller μ or ρ will both decrease the decaying speed of the probability density, which means that the large jump lengths are more likely to appear. Hence, observations (1) and (2) can be explained since frequently large jump lengths can help covering a wider range where it is more likely to find a target in a sparse area. Generally, the density of target site is sparse in practice which means that λ is usually large. Due to the exponential decaying of tempered Lévy distribution, the value of is quite small and almost has no influence on the searching efficiency. It can then explain why the results of Figure 4 with different λ are similar.
One can also interpret the observations from the practical perspective. As discussed before, the tempered item e−ρl can be viewed as the probability density that the forager can keep its flight direction before he completes one flight in the existence of some unknown factors. Thus, a smaller ρ means that the probability of a forager to encounter some uncertain factors is lower and the foraging efficiency should be higher.
For the destructive case, the mean number N can be expressed as
Similar to the non-destructive case, one can then calculate the searching efficiency using (7). The results are shown in Figure 5, which is very similar to the non-destructive case. It is found that a smaller μ or ρ will both result in a higher search efficiency. The mean free path λ almost has no influence on the optimal choice of parameters μ and ρ. We have shown that for the Lévy distribution, μ → 1, where a large jump length appears more likely, will lead to a higher searching efficiency. Thus, a smaller ρ and μ will also result in a larger searching efficiency because large jump lengths are more likely to happen.
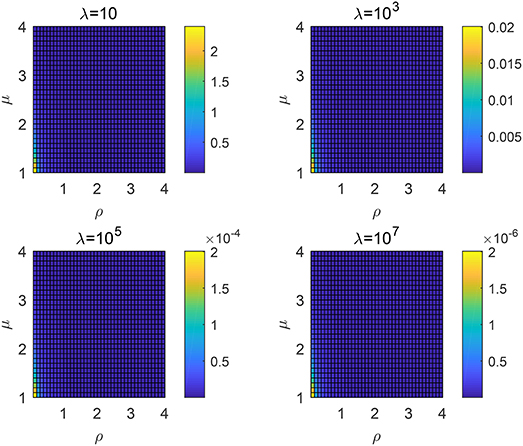
Figure 5. Searching efficiency ηλ for different order μ and ρ: the destructive case with different λ.
Remark 3: According to the results in Figure 6 where we obtain the mean flight number after averaging 100 independent runs, one can find that a larger variance will lead to a smaller mean flight number. Moreover, it is shown that there is a linear property between them with a correlation coefficient −0.9781, which indicates that the estimation of mean flight number is fine.
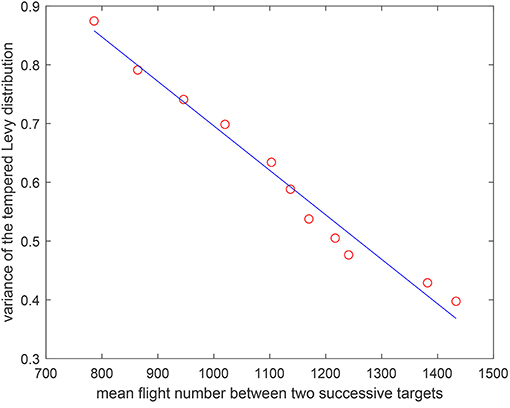
Figure 6. The relation between the mean flight number and the variance of the tempered Lévy distribution.
We also implement an experiment for validate the theoretical analyses. Consider a 200 × 200 area and 50 targets are uniformly distributed in this area. The vision distance is rv = 1 and the total flight distance is no longer than 10, 000 which can be viewed as the flight capability of the forager. The searching efficiency is estimated as where Nnum is the number of found targets and Ltotal is the total flight distance. From Figure 7 where the searching efficiency is derived by averaging 100 independent runs, one can find that a smaller ρ and μ will both lead to a higher searching efficiency, which is consistent with the theoretical analyses. Because a larger μ will make the density function decrease quickly, the range of jump lengths is then very tight. Thus, the searching efficiency is very close for a large μ where the jump lengths are all around the vision distance rv. Figures 8–10 give some typical foraging procedures for different parameters and one can find that all of them perform a Brownian motion which can be verified by the statistic results of averaging 100 independent generated jump lengths in Figure 11. Additionally, larger jump lengths frequently appear in Figure 8 compared with the other two figures, for which the searching efficiency is the highest. It is also shown in Figure 11, where larger jump lengths are most likely to appear for the λ = 0.5 and μ = 1 case.
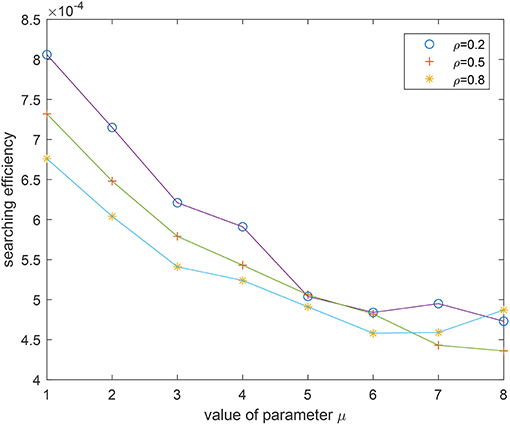
Figure 7. Experimental results of searching efficiency for different λ and μ.
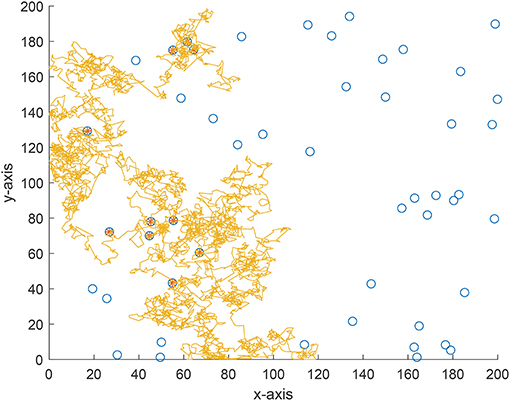
Figure 8. A typical example of foraging procedure with λ = 0.5 and μ = 1.
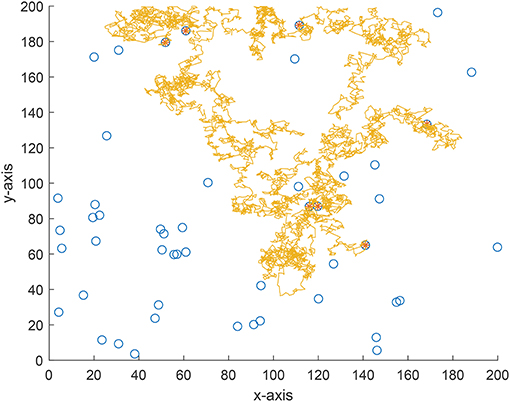
Figure 9. A typical example of foraging procedure with λ = 1 and μ = 1.
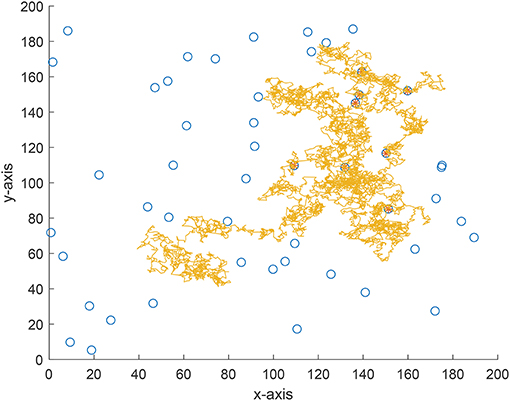
Figure 10. A typical example of foraging procedure with λ = 0.5 and μ = 3.
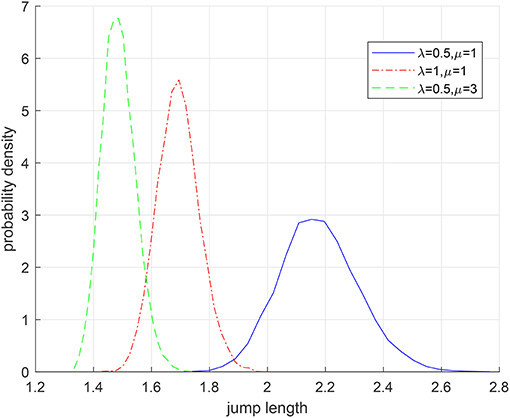
Figure 11. The statistic property of jump length with different λ and μ.
Remark 4: In this paper, we numerically generate the jump lengths distributed as a tempered Lévy distribution and Figure 12 shows the actual density function and the statistic result of generated jump lengths. It is found that the statistic result is very close to the actual density function. We have to declare here that all the foraging procedures in Figures 8–10 will result in a Brownian motion because of the Central Limit Theorem.
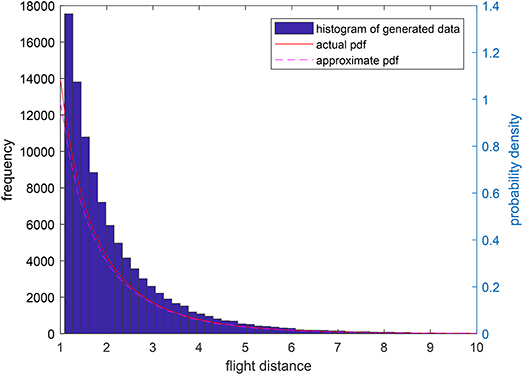
Figure 12. The actual tempered Lévy density function with ρ = 0.5 and μ = 1 and the statistic results of generated jump lengths.
Remark 5: Some conclusive remarks can be drawn as follows
(1) Tempered Lévy model is to assume the uncertain factors during the foraging procedure may happen according to an exponential distribution. Whenever such uncertain factor happens, the forager has to stop its flight, which seems like that the flight distance is truncated.
(2) If we take ρ = 0, the tempered Lévy fight will reduce to the standard Lévy flight.
(3) According to the results in Figure 5, it is found that a smaller ρ leads to a higher searching efficiency. In fact, the searching efficiency with standard Lévy distribution is higher than the truncated Lévy distribution. It is easy to understand since the forager will have a higher searching efficiency if there is no interruption during the foraging procedure.
In the previous, we have not taken the flight time into consideration. Assume that the flight speed v is constant during the foraging process and treat the flight time between two flights as the waiting time. Then, the pdf of waiting time is the same as the flight distance with a scaling parameter v, which can be expressed as
Let us introduce the Laplace transform for the waiting time as
The famous Montroll-Weiss Equation [16] in Fourier-Laplace space is in the following form
where W(k) is the Fourier transform for the flight distance in two dimensions which will be discussed later. Now consider the extreme distribution of Ψ(s) with s → 0. It is followed that
where, b is the mean of flight time t and o(·) means the higher order infinitesimal. In the following, we will present two different types of master equations for this foraging procedure via different treatments to the extreme distribution of W(k) with k → 0.
Assume that the searching direction θ is uniformly distributed in the interval [0, 2π). If the waiting time and the flight distance are independent, then the flight distance of the forager in two dimensions can be formulated as (x, y) = (lcos θ, lsin θ) and the following Fourier transform holds
where, k1 and k2 are the Fourier variables, k = (k1, k2), Θ = k1 cos θ + k2 sin θ and a is the second order moment of the flight distance.
Substitute (12) and (13) into the Montroll-Weiss equation and ignore the higher order infinitesimal, yielding,
Perform inverse Fourier-Laplace transform and one can derive the master equation
Remark 6: Unlike the master equation derived by Zeng and Chen [15], the master equation in this study is a normal diffusion equation since the first and second order moments exist. We have to mention that the master equation proposed by Zeng and Chen [15] should also be standard diffusion equation rather than fractional diffusion differential equation since the Lévy distribution is truncated by the mean free path λ. Moreover, the master equation should be two-dimensional rather than one-dimensional.
In this subsection, our purpose is to express the master equation as a tempered fractional diffusion equation and we have restrict μ varies from 1 to 2 to derive the tempered fractional derivative expression. The vector jump length can be described as lΘ, where Θ = (cos θ, sin θ). From Equation (7.9) in the book of Meerschaert and Sikorskii [17], it shows that
where k · Θ = k1 cos θ + k2 sin θ, M(dΘ) is a uniform distribution on a unit circle, and C is a constant relevant to coefficients ρ and μ.
Substitute (12) and (16) into the Montroll-Weiss Equation (11) and ignore the higher order infinitesimal, yielding,
Define
where,
with g(t) = f(x + tΘ) is the generator form for vector tempered fractional derivative.
Inverse (17) to derive the master equation
where, L is a vector (x, y).
Remark 7: Since the first order and second order moments of tempered Lévy distribution exist, the resulting standard diffusion equation (15) makes sense. Interestingly, we borrow the idea from Meerschaert and Sikorskii [17] and give another expression of the master equation, where vector tempered fractional derivative is used. In this paper, we do not give detailed proof for the derivation of vector tempered fractional derivative and one can refer to Chapter 6 and 7 in the book of Meerschaert and Sikorskii [17]. All these indicate that tempered fractional diffusion equation is in fact a different expression of the standard diffusion.
In this paper, we consider the optimal random foraging whose flight distance is distributed according to a tempered Lévy distribution p(l) ~ e−ρll−μ. It is found that a higher searching efficiency can be derived when we choose a smaller ρ or μ, which results in a slower decaying speed. Furthermore, we obtain the master equation of the random foraging. A standard diffusion equation is derived since the first and second order moments of the distribution for flight distance exist. Using the definition of tempered fractional derivative, a vector tempered fractional diffusion equation is then derived, which can be viewed as a special expression for the standard diffusion. A promising research topic can be directed to finding the optimal searching strategy for other types of flight distance distributions.
YC mainly contributed to the theoretical analysis and accomplishing the paper. DH mainly contributed to the numerical simulation. YW and YC contributed for providing the idea of using tempered Lévy distribution in foraging and helped revising the paper.
This work was fully supported by China Scholarship Council (No. 201706340089) and NSF NRT Fellowship (No. DGE-1633722).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
1. Bartumeus F, Da Luz ME, Viswanathan G, Catalan J. Animal search strategies: a quantitative random-walk analysis. Ecology (2005) 86:3078–87. doi: 10.1890/04-1806
2. Viswanathan G, Raposo E, Da Luz M. Lévy flights and superdiffusion in the context of biological encounters and random searches. Phys Life Rev. (2008) 5:133–50. doi: 10.1016/j.plrev.2008.03.002
3. Reynolds A. Optimal random Lévy-loop searching: new insights into the searching behaviours of central-place foragers. EPL (2008) 82:20001. doi: 10.1209/0295-5075/82/20001
4. Bartumeus F, Catalan J, Fulco U, Lyra M, Viswanathan G. Optimizing the encounter rate in biological interactions: Lévy versus Brownian strategies. Phys Rev Lett. (2002) 88:097901. doi: 10.1103/PhysRevLett.88.097901
5. Kerster BE, Rhodes T, Kello CT. Spatial memory in foraging games. Cognition (2016) 148:85–96. doi: 10.1016/j.cognition.2015.12.015
6. Viswanathan GM, Buldyrev SV, Havlin S, Da Luz M, Raposo E, Stanley HE. Optimizing the success of random searches. Nature (1999) 401:911–4.
7. Viswanathan G, Afanasyev V, Buldyrev SV, Havlin S, Da Luz M, Raposo E, et al. Lévy flights in random searches. Phys A Stat Mech Appl. (2000) 282:1–12. doi: 10.1016/S0378-4371(00)00071-6
8. Viswanathan GM, Afanasyev V, Buldyrev S, Murphy E, Prince P, Stanley HE. Lévy flight search patterns of wandering albatrosses. Nature (1996) 381:413–5.
9. Boyer D, Miramontes O, Ramos-Fernandez G, Mateos J, Cocho G. Modeling the searching behavior of social monkeys. Phys A Stat Mech Appl. (2004) 342:329–35. doi: 10.1016/j.physa.2004.04.091
10. Ramos-Fernández G, Mateos JL, Miramontes O, Cocho G, Larralde H, Ayala-Orozco B. Lévy walk patterns in the foraging movements of spider monkeys (Ateles geoffroyi). Behav Ecol Sociobiol. (2004) 55:223–30. doi: 10.1007/s00265-003-0700-6
11. Dybiec B, Gudowska-Nowak E, Barkai E, Dubkov AA. Lévy flights versus Lévy walks in bounded domains. Phys Rev E (2017) 95:052102. doi: 10.1103/PhysRevE.95.052102
12. Zhao K, Jurdak R, Liu J, Westcott D, Kusy B, Parry H, et al. Optimal Lévy-flight foraging in a finite landscape. J R Soc Interface (2015) 12:20141158. doi: 10.1098/rsif.2014.1158
13. Plank M, James A. Optimal foraging: Levy pattern or process? J R Soc Interface (2008) 5:1077–86. doi: 10.1098/rsif.2008.0006
14. Bartumeus F, Raposo EP, Viswanathan GM, da Luz MG. Stochastic optimal foraging: tuning intensive and extensive dynamics in random searches. PLoS ONE (2014) 9:e106373. doi: 10.1371/journal.pone.0106373
15. Zeng CB, Chen YQ. Optimal random search, fractional dynamics and fractional calculus. Fract Calculus Appl Anal. (2014) 17:321–32. doi: 10.2478/s13540-014-0171-7
16. Montroll EW, Weiss GH. Random walks on lattices. II. J Math Phys. (1965) 6:167–81. doi: 10.1063/1.1704269
Keywords: optimal random search, foraging, tempered Lévy distribution, master equation, tempered fractional derivative
Citation: Chen Y, Hollenbeck D, Wang Y and Chen Y (2018) On Optimal Tempered Lévy Flight Foraging. Front. Phys. 6:111. doi: 10.3389/fphy.2018.00111
Received: 27 May 2018; Accepted: 11 September 2018;
Published: 04 October 2018.
Edited by:
Dumitru Baleanu, University of Craiova, RomaniaReviewed by:
Jordan Yankov Hristov, University of Chemical Technology and Metallurgy, BulgariaCopyright © 2018 Chen, Hollenbeck, Wang and Chen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: YangQuan Chen, eWNoZW41M0B1Y21lcmNlZC5lZHU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.