 Paulo Ávila-Gómez1
Paulo Ávila-Gómez1 Alba Vieites-Prado2,3
Alba Vieites-Prado2,3 Clara Correa-Paz4
Clara Correa-Paz4 Lucía Del Pozo-Filíu5
Lucía Del Pozo-Filíu5 Nuria Palomar-Alonso5
Nuria Palomar-Alonso5 Francisco Campos5*
Francisco Campos5* Esteban López-Arias5*
Esteban López-Arias5*- 1Department of Pathology and Laboratory Medicine, Weill Cornell Medicine, New York, NY, United States
- 2Brain Plasticity Laboratory, Health Research Institute of Santiago de Compostela (IDIS), Santiago de Compostela, Spain
- 3Brain Plasticity Laboratory, Centre for research in molecular medicine and chronic diseases (CIMUS), Universidade de Santiago de Compostela, Santiago de Compostela, Spain
- 4Neurological Sciences and Cerebrovascular Research Laboratory, Neurology and Cerebrovascular Disease Group, Department of Neurology and Stroke Centre, Neuroscience Area La Paz Institute for Health Research - idiPAZ (La Paz University Hospital- Universidad Autónoma de Madrid), Madrid, Spain
- 5Translational Stroke Laboratory Group (TREAT), Clinical Neurosciences Research Laboratory (LINC), Health Research Institute of Santiago de Compostela (IDIS), Santiago de Compostela, Spain
Several preclinical assays and clinical trials have found hypothermia as an efficient protective treatment for stroke. However, systemic hypothermia impairs several physiological functions being difficult to implement in acute critical patients. A deeper understanding of the mechanisms underlying the therapeutic effects of hypothermia could inspire new treatments based on the protective effects of cold. Furthermore, this could contribute to the reduction of the side effects associated with it. One of the metabolic landmarks of hypothermia is the overexpression of a small subset of shock proteins while global protein synthesis is reduced. Among these cold-shock proteins, RBM3 (RNA-binding motif protein 3) seems to play a central protective role. In physiological conditions, which is involved in the regulation of protein synthesis. In several models of cerebral diseases, in vitro and in vivo, RBM3 exhibited the ability to mitigate apoptosis or increase neural proliferation. In stroke models, RBM3 has shown specially promising effects attenuating neural damage and enhancing cell survival. Future prospects should be directed towards the design of efficient strategies to modulate RBM3 levels. This mini-review aims to summarize the progress made in understanding the role of RBM3 in cerebral tissue protection, while encouraging efforts to address research gaps, particularly in its modulation and clinical application.
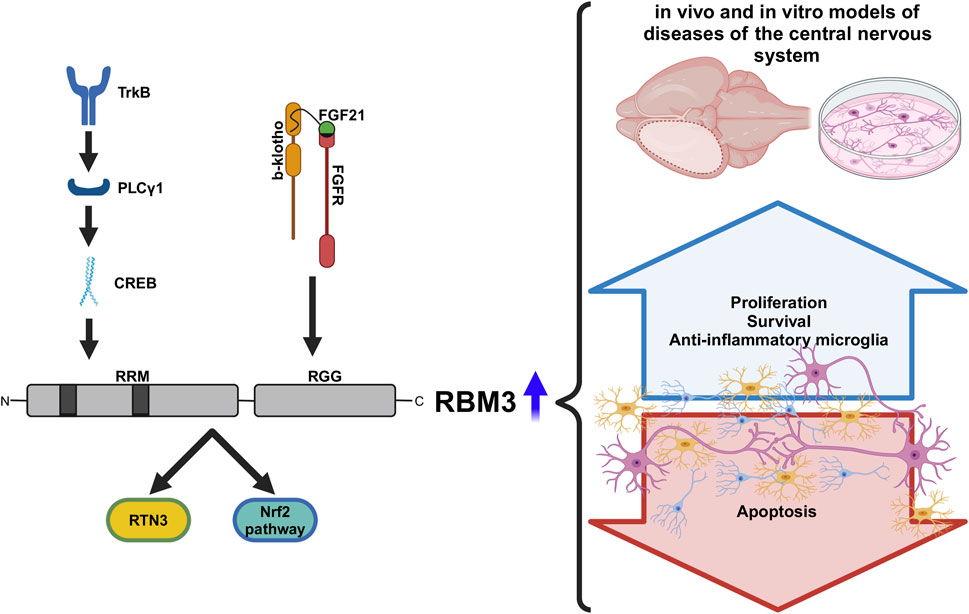
GRAPHICAL ABSTRACT | Summary of downstream and upstream pahtway of RBM3, with a detail of its structure. On the right we can see the effects of overexpression of RBM3 in several in vivo and in vitro models of deseases of central nervous system. Graphical abstract was created with BioRender (biorender.com/)
1 Introduction
Ischemic stroke occurs due to insufficient blood flow to cerebral tissue, leading to irreversible damage and, consequently, loss of function and structural support of the affected brain regions. Nowadays, it is still one of the leading causes of death and disability worldwide (World Health Organization, 2024). Approximately 85% of strokes are ischemic (Johnson et al., 2019), and their incidence is expected to increase in the coming years due to the progressive aging of the global population (Béjot et al., 2019).
Despite multiple neuroprotective strategies have been tested (Pérez-Mato et al., 2024), the only available treatment for ischemic stroke in the acute phase is the re-establishment of blood supply, either pharmacologically by intravenous fibrinolytic treatment, or by mechanical thrombectomy. Fibrinolytic treatment consists in rtPA (tissue-type plasminogen activator) injection by infusion that turns plasminogen into plasmin dissolving the clot, and mechanical thrombectomy is a surgical intervention under image guidance with endovascular devices in order to remove the clot (Powers et al., 2019).
Among other clinical parameters, temperature has a deep influence in the outcome of stroke patients (Campos et al., 2012). While hyperthermia worsens the functional outcome (Blanco et al., 2012; Campos et al., 2013; Saini et al., 2009), hypothermia correlates with positive prognosis (Dumitrascu et al., 2016; Miyazawa et al., 2003; Van Der Worp et al., 2007). However, the clinical use of hypothermia carries severe negative side effects such as the reduction of serum K+, impairing cardiac function (Mirzoyev et al., 2010); shivering, that requires the use of sedatives and hinders the management of the patient (Nakamura and Morrison, 2011); altered diuresis; disruption of the hemostatic function; immunosuppression; alteration of liver function, or decrease in drug clearance (Polderman, 2009; Wood and Thoresen, 2015). While the clinical use of hypothermia is limited, dissecting its protective mechanisms could inspire the development of new therapeutic strategies.
A hallmark of hypothermic effects over cellular metabolism is a depression of global protein synthesis, while a small subset of cold-shock proteins (CSPs) is overexpressed. Therefore, these CSP could play an important role in its neuroprotective effect. The most studied human CSPs are the cold-inducible RNA-binding protein (CIRP) and RNA-binding motif protein 3 (RBM3). CIRP has been related to increased neuroinflammation in an ischemic stroke model by increasing TNF-α expression and microglial activation resulting in bigger lesion volume (Zhou et al., 2014). Meanwhile, RBM3 has been widely studied and, as discussed in this mini-review, may play a beneficial role in ischemic stroke.
2 RBM3 protein
First described by Derry et al. (1995), RBM3 is a member of the glycine-rich RNA-binding protein family with RNA chaperone functions.
2.1 Structure, expression and functions
RBM3 contains 157 amino acids (17 kDa) (Derry et al., 1995), and is characterized by a highly conserved RNA recognition motif (RRM) at the N-terminus and a less conserved arginine-glycine-rich domain (RGG) at the C-terminus (Zhu et al., 2016a). The RRM is involved in mRNA processing, while the RGG domain regulates post-transcriptional modifications of RNA and interactions between RNA, DNA, and proteins (Hu et al., 2022).
RBM3 has been identified in embryonic development with a peak in postnatal phase in rats, decreasing after the second week. However, RBM3 can still be detected in the adulthood in the subventricular zone and in the rostral migratory stream, where cell proliferation and migration remain active, as well as in the cerebellum and olfactory bulb where the translation rate is elevated (Pilotte et al., 2009). Despite these facts, RBM3 knockout (KO) mice showed no differences in cerebral or neuronal characteristics compared to wild type that could suggest a redundant mechanism or, as we will discuss below, a prevention to pathophysiological conditions such as hypoxia or hypothermia (Matsuda et al., 2011; Zhu et al., 2019).
Similarly in humans, high levels of RBM3 have been observed in the infant brain (<1 year), especially in the hippocampus and prefrontal cortex; in adult brains, by contrast, RBM3 is nearly absent (Jackson et al., 2018). However, RBM3 has also been detected in higher concentrations in neurons than in glial cells (Jackson et al., 2019), perhaps because the high sensitivity of neurons requires greater prevention of unfavorable conditions.
It has been shown that RBM3 plays a role in several physiological processes at molecular and cellular levels, including: post-transcriptional and post-translational regulation (Sureban et al., 2008), modulation of alternative polyadenylation (Hu et al., 2014), mRNA alternative splicing (Smart et al., 2007), regulation of microRNA (Dresios et al., 2005; Wang et al., 2021), cell stemness and cycle progression (Matsuda et al., 2011), apoptosis (Chip et al., 2011; Zhuang et al., 2017), ER stress (Zhu et al., 2016b), dynamics of synaptic vesicles (Sertel et al., 2021), and circadian cycle (Liu et al., 2013; Sertel et al., 2021).
2.2 Relation between RBM3 and temperature
Danno et al. (1997) demonstrated for the first time that mild hypothermia (32°C) for 24 h induced transcription of RBM3 in several immortal human cell lines such as erythroleukemia type K562, hepatocellular carcinoma Hep62, epithelioid cervix carcinoma HeLa, and bladder carcinoma T24. This cold-induced increase has also been found in cortical human neurons differentiated from pluripotent stem cells (Rzechorzek et al., 2015), HEK293 (Zhu et al., 2016b), and human neural lineage cell lines SK-N-SH (Rosenthal et al., 2017), SH-SY5Y (Jackson et al., 2018) and hNSCs (Ma et al., 2024).
In healthy animals, RBM3 protein and mRNA levels were increased in cerebral tissue in rats following four hours of systemic hypothermia at 32°C induced using a rectal-thermostat-controlled electric pad, and remained elevated for a minimum of 6 h post-hypothermia. Focal hypothermia restricted to a single cerebral hemisphere was achieved with a metal coil connected to a cooled water circuit, this device was inserted between skull and temporalis muscle for 24 h, resulting in an increase of RBM3 protein and mRNA levels in the cooled hemisphere compared with the contralateral (normothermic) hemisphere. Therefore, reduction of temperature, whether systemic or localized, can increase RBM3 levels in the brain. Furthermore, similar results were reported in a rodent model of cerebral ischemia (Ávila-Gómez et al., 2022). In addition, pharmacological hypothermia using phenothiazine drugs (chlorpromazine and promethazine, an antipsychotic and a first-generation antihistamine, respectively) also resulted in increased RBM3 expression up to 24 h in a stroke mouse model (Jiang et al., 2022).
In stroke patients, a clinical retrospective study found a negative correlation between basal RBM3 levels in serum and body temperature at admission and at 24 h. Moreover, normothermic (<37.5°C) patients had higher RBM3 values at 24 h than hyperthermic (>37.5°C) patients (Ávila-Gomez et al., 2020). In the same line, analysis of blood samples from patients of the EuroHYP-1 study (a European multicenter, randomized, phase III clinical trial designed to evaluate therapeutic effect of hypothermia in acute ischemic stroke) (van der Worp et al., 2014) did not find significant differences on circulating RBM3 levels in patients who received hypothermic treatment compared with untreated subjects, although rewarming levels tended to be higher in hypothermic patients.
These clinical findings are in agreement with another study in which RBM3 levels increased in the blood of patients with congenital heart disease just after cardiopulmonary bypass (CPB) and therapeutic hypothermia compared with preoperative levels (Rosenthal et al., 2020). In these patients, RBM3 levels were not significantly elevated 24 h after surgery; neither in patients who received CPB but not therapeutic hypothermia. Finally, Wan et al. (2023) detected an elevation in RBM3 blood levels in stroke patients immediately after intra-arterial hypothermia and also 1 day after surgery, an increase that was not identified in the control group without hypothermia treatment.
3 RBM3-mediated effect on neuronal development and survival
Despite normal embryonic development in RBM3 KO, the absence of RBM3 causes lack of beneficial effects of cooling: structural and synaptic plasticity in neurons were abolished (Peretti et al., 2021) and neuronal proliferation and differentiation were altered in RBM3 KO mice (Xia et al., 2018; Yan et al., 2019), differentiation of hNSCs into neurons and neuroblasts was diminished after blocking RBM3 with siRNA (Ma et al., 2024), and knockdown of RBM3 by RNAi reduced the number of synapses and impaired novel memory in adult mice (Peretti et al., 2015). As we can see, alterations in RBM3 levels imply changes in the capability of cells and tissues of both neuroprotection and neurorepair processes.
Regarding neuroprotection, several studies have observed a correlation between increased RBM3 levels and an augmented resistance against damage. After staurosporine (STS, a caspase-dependent apoptosis inducer) treatment to induce apoptosis, hypothermia increased RBM3 levels and attenuated caspase-dependent apoptosis in primary murine neurons; blockage of RBM3 by siRNA almost abolished this damage attenuation, while over-expression of RBM3 without hypothermia mimicked the protective effect (Chip et al., 2011; Yang et al., 2017). In a cardiac arrest rat model, hypothermia increased RBM3 levels in certain brain regions resulting in less deficit and less apoptosis in the subgranular zone (SGZ) compared to normothermic controls (Zhang et al., 2021). Similarly, mild hypothermia increased RBM3 levels and decreased apoptosis in SH-SY5Y cells treated with sodium nitroprusside (SNP, a NO donor) with respect to SNP-treated cells under normothermic conditions (Yang et al., 2017).
Hypothermia after hypoxia in SK-N-SH neurons diminished neuronal damage as well as raised RBM3 levels (Rosenthal et al., 2017). According to that, treatment with RBM3 in hypoxic rats improved several neurological functions (Lin et al., 2024).
In different Parkinson disease (PD) models (Yang et al., 2018; Yang et al., 2019), a reduction of apoptosis has also been observed after overexpression of RBM3 in SH-SY5Y cells. Moreover, after adding 1-methyl-4-phenylpyridinium (a dopaminergic neurotoxin) to the medium as PD in vitro model, RBM3 overexpression was able to diminish several apoptosis markers (Yang et al., 2018). Yielding consistent results, in PC12 cell lines transfected with fragments of huntingtin with 23 and 74 glutamine repeats as a Huntington’s disease in vitro model, RBM3 transfection reduced cell death (Kita et al., 2002).
A reduction in synapse density was observed in wild-type mice immediately after treatment of 45 min at 16°C–18°C (a severe hypothermia), but normal synapse density was restored upon rewarming. This ability to recover was maintained in 5xFAD mice (Alzheimer model) at 2 months of age but lost at 3 months, which mirrors the ability of 5xFAD mice to acutely increase RBM3 in response to hypothermia, but lost at 3 months (Peretti et al., 2015). Similarly, in a prion disease model, synaptic recovery capacity was conserved until 5 weeks post-injection (w.p.i.) of prion and lost at 6 w.p.i., similar to the increase of RBM3 by cooling which was annulated at 6 w.p.i. (Peretti et al., 2015). These data reinforce the idea that RBM3 is essential in neuronal recovery after adverse conditions. Overexpression of RBM3 by lentivirus-mediated RNAi in this prion model resulted in rescue of synaptic deficit and global translation, improvement of synaptic transmission, reduction of memory deficit, and rise of survival; meanwhile, its blockage reverted the beneficial effects of cooling (Peretti et al., 2015).
Antisense oligonucleotide (ASO) targeting E3a poison exon (an exon within RBM3 pre-mRNA that mediates the degradation of the mRNA) was tested in the intrahippocampal kainate (KA) mouse model of epilepsy resulting in a significant reduction of neuronal loss after status epilepticus induction (Stauffenberg et al., 2024).
The influence of RBM3 in neurorepair was reported too. The same ASO against E3a was also administered in the abovementioned prion model resulting in overexpression of RBM3 and, consequently, in proliferation of pyramidal neurons in CA1 and attenuation of spongiosis (Preussner et al., 2023). Likewise, the overexpression of RBM3 in a hypoxia model in Neural stem cells (NSCs) succeeded in increasing live cells and promoting G1 to S phase transition (Yang et al., 2019).
4 RBM3 in ischemic stroke
In ischemic stroke models, in vitro and in vivo, many examples of activation of neuroprotection and neurorepair can also be found. Experiments with oxygen and glucose deprivation (OGD), as in vitro stroke model, revealed activation of neuroprotection when RBM3 was overexpressed, attenuating apoptosis in PC12 cells (Si et al., 2020), in N2a cells (Zhang et al., 2023), and even in organotypic brain slice culture (OBSC); in the last case by stimulating anti-inflammatory microglia proliferation (Zhao et al., 2024). Interestingly, PC12 cells from RBM3 KO presented an increase of apoptosis after OGD respect to cells from WT, moreover, inhibition of RBM3 by siRNA attenuated protective effect of mild hypothermia in N2a cells undergone OGD (Si et al., 2020; Zhang et al., 2023). Analogously, apoptosis induced by OGD in HT22 cells were reverted by adding RBM3 in culture (Lin et al., 2024).
Neurorepair activation has also been reported. For instance, the overexpression of RBM3 without hypothermia in NSPCs stimulated the profileration of these cells after OGD conditions (Zhu et al., 2019).
However, there are some differences in the response to RBM3 depending on the cell source. NSPCs from murine subventricular zone (SVZ) showed increased DCX positive cells when RBM3 was overexpressed with a recombinant form, but no differences in DCX positive cells were observed after OGD between vehicle and RBM3 overexpressed groups. Conversely, NSPCs from subgranular zone (SGZ) presented a rise of neural proliferation and neuronal differentiation when RBM3 was overexpressed in both conditions, but it is noteworthy that this cell type also increased the proliferation and differentiation after OGD in the vehicle group (Zhu et al., 2019).
Both neuroprotection and neurorepair activations were reflected in in vivo experiments. Administration of zr17-2 (a small-molecule that targets and modulate RBM3 and CIRP) in ischemic rats enhanced RBM3 protein level, and reduced infarction volume and glial scar formation, enhanced neurogenesis, promoted anti-inflammatory responses, decreased pro-inflammatory markers and improved motor function (Zhao et al., 2024). In line with this, an increase in Rbm3 levels following pharmacological hypothermia was related with better outcomes in stroke mice (Jiang et al., 2022). Reinforcing the importance of RBM3 in cerebral protection under ischemic circumstances, hypoxic-ischemic brain injury in KO mice resulted in an increase of neuronal loss, enhancement of apoptosis in ipsilateral hemisphere and decrease of neural proliferation in the SGZ (Zhu et al., 2019).
Finally, an independent association between RBM3 values at 24 h and good prognosis was found in stroke patients. And more importantly, a variation of RBM3 between admission and 24 h higher than 10% was an independent marker for good prognosis (Ávila-Gomez et al., 2020). Likewise, Wan et al. (2023) also found an independent positive correlation between RBM3 and good outcome at 90 days in stroke patients.
5 Non-hypothermic modulation of RBM3
Most of the above-discussed studies accomplished RBM3 overexpression directly by hypothermia or vector transfection, procedures hardly applicable in clinical practice. In search of alternative strategies to induce RBM3 increase, several approaches have been tested, and summarized in Table 1.
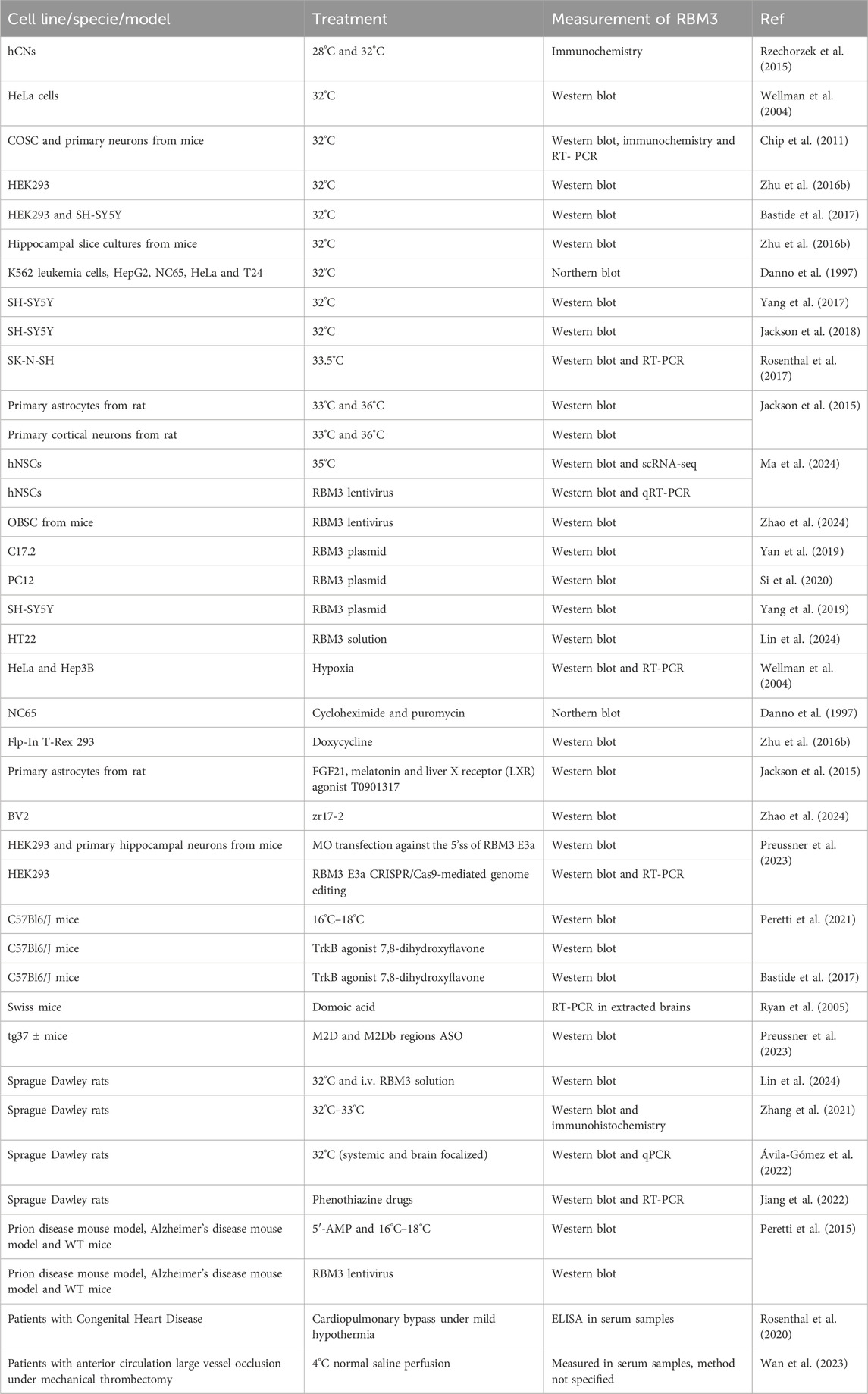
Table 1. Methods to increase RBM3 in vivo and in vitro (abbreviations: ASO, antisense oligonucleotide; COSC, cortical organotypic slice cultures; FGF21, fibroblast growth factor 21; hNSCs, human neural stem cells; i.v., intravenous; MO, morpholino; OBSC, organotypic brain slice culture; hCNs, human cortical neuronal cells).
Certain drugs have been shown to upregulate RBM3 expression in rodents and in vitro, such as neurotoxin domoic acid, fibroblast growth factor 21 (FGF21), melatonin and zr17-2 (Jackson et al., 2015; Ryan et al., 2005; Zhao et al., 2024). In contrast, metformin and the adenosine monophosphate (AMP) analog AICAR (5-aminoimidazole-4-carboxamide 1-β-D-ribofuranoside, Acadesine, N1-(β-D-ribofuranosyl)-5-aminoimidazole-4-carboxamide) have been shown to inhibit its expression in vivo and in vitro, both of them activators of 5′adenosine monophosphate-activated protein kinase pathway that finally modify alternative splicing (Laustriat et al., 2015).
FGF21, already approved for clinical trials of metabolic disorders (Shao and Jin, 2022), augmented RBM3 levels in young neuron culture (Jackson et al., 2015). In stroke patients, a strong relationship was found between serum FGF21 levels on admission and RBM3 levels 72 h after admission. Moreover, a negative correlation was found between both molecules and maximum temperature, and a positive correlation with good outcome (Ávila-Gómez et al., 2022).
The activation of the TrkB receptor induces upstream RBM3 through PLCγ1 and, in consequence, CREB phosphorylation. In fact, the activation of TrkB with the agonist 7,8-dihydroxyflavone increased RBM3 levels in vivo without cooling and induced neuroprotection in prion-infected mice (Peretti et al., 2021).
Downstream, RBM3 binds to reticulon-3 (RTN3) mRNA enhancing its translation, at least in HEK293 cells and in murine hippocampus. Moreover, overexpression of RTN3 promoted neuroprotection in an in vivo model of neurodegenerative disease (Bastide et al., 2017) and in vitro cell death models (Teng and Tang, 2013); its silencing weakened protective effect of hypothermia or RBM3 overexpression in N2a cells after OGD (Zhang et al., 2023). Conversely, RTN3 has also been identified as neurotoxic since its accumulation provokes the formation of immunoreactive dystrophic neurites, which impairs synaptic plasticity (Hu et al., 2007).
Similarly, Nuclear factor erythroid-derived 2-like 2 (Nrf2) signaling has been found downstream in the RBM3 through growth arrest specific 6. Nrf2 is involved in homeostasis process, mitochondrial function, or anti-inflammation by regulation of antioxidant genes, moreover, its silencing suppressed the anti-inflammatory effects of RBM3 in cell culture (Lin et al., 2024). Likewise, SOX11 pathway has been identified as necessary in neuronal differentiation stimulated by RBM3 (Ma et al., 2024).
A more sophisticated approach involves the use of ASO to promote RBM3 expression by inhibiting the poison exon E3a, as discussed in Section 3 (Preussner et al., 2023; Stauffenberg et al., 2024). This ASO technology is already approved by the Food and Drug Administration (FDA) for Spinal Muscular Atrophy (SMA) (Aartsma-Rus, 2017), making this new approach translational. However, several issues should be considered: both in animal models and in the clinic, the oligo is administered into the cerebrospinal fluid in order to facilitate access to the cerebral tissue. SMA was an orphan and mortal disease, but using this route of administration in stroke or other pathologies could raise ethical concerns. Consequently, alternative approaches have been proposed to vectorize the ASO, avoid its degradation, improve the stability and, ultimately, enable non-invasive administration. This includes approaches such as chemical modifications of the molecule or conjugation with peptides that bind the low-density lipoprotein receptor, aptamers capable to target receptors, or antibodies, among others (Benizri et al., 2019; Ferhat et al., 2024).
6 Conclusion
Targeting the RBM3 pathway is a promising strategy to benefit from cooling as treatment for stroke without its negative side effects. However, although multiple neuroprotective strategies have been veotested (summarized in Table 2), several knowledge gaps preclude its use, such as the optimal RBM3 levels required to obtain clinical benefit, the best pharmacological strategy to induce its effect in neuronal tissue, or the optimal therapeutic window.
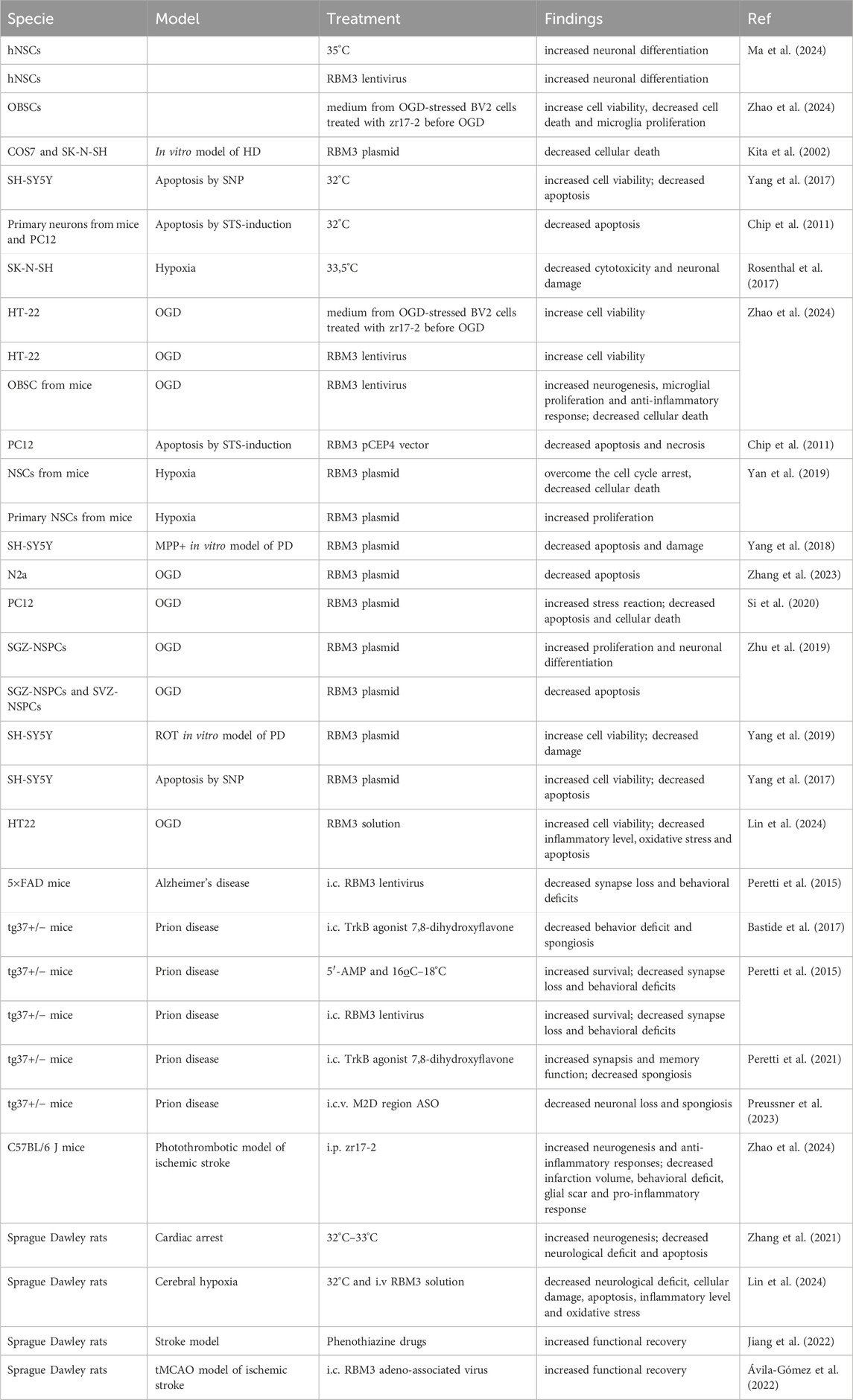
Table 2. Effects of RBM3 increasing (abbreviations: ASO, antisense oligonucleotide; HD, Huntington disease; hNSCs, human neural stem cells; i.c.v., intracerebroventricular; i.p., intraperitoneal; MPP+, 1-methyl-4-phenylpyridinium; NSCs, neural stem cells; OBSC, organotypic brain slice culture; OGD, oxygen and glucose deprivation; PC12, rat pheochromocytoma cells; PD, Parkinson disease; ROT, rotenone; SGZ, subgranular zone; SNP, Sodium nitroprusside; STS, staurosporine; SVZ, subventricular zone; tMCAO, transient middle cerebral artery occlusion; i.c., intracerebral).
Author contributions
PÁ-G: Writing–review and editing. AV-P: Writing–review and editing. CC-P: Writing–review and editing. LD-F: Writing–review and editing. NP-A: Writing–review and editing. FC: Funding acquisition, Supervision, Writing–review and editing. EL-A: Writing–original draft, Writing–review and editing.
Funding
The authors declare that financial support was received for the research, authorship, and publication of this article. The authors disclosed receipt of the following financial support for the research, authorship, and publication of this article: Instituto de Salud Carlos III (ISCIII) and co-funded by the European Union, grant numbers: ICI19/00032, PI20/01014, RD21/0006/0003 (RICORS-ICTUS network). Xunta de Galicia, grant number: IN607A2022/02.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Aartsma-Rus, A. (2017). FDA approval of nusinersen for spinal muscular atrophy makes 2016 the year of splice modulating oligonucleotides. Nucleic Acid. Ther. 27, 67–69. doi:10.1089/nat.2017.0665
Ávila-Gómez, P., Pérez-Mato, M., Hervella, P., Dopico-López, A., Silva-Candal, A. D., Bugallo-Casal, A., et al. (2022). Associations between RNA-binding motif protein 3, fibroblast growth factor 21, and clinical outcome in patients with stroke. J. Clin. Med. 11, 949. doi:10.3390/jcm11040949
Ávila-Gómez, P., Vieites-Prado, A., Dopico-López, A., Bashir, S., Fernandez-Susavila, H., Gubern, C., et al. (2020). Cold stress protein RBM3 responds to hypothermia and is associated with good stroke outcome. Brain. Commun. 2, fcaa078. doi:10.1093/braincomms/fcaa078
Bastide, A., Peretti, D., Knight, J. R., Grosso, S., Spriggs, R. V., Pichon, X., et al. (2017). RTN3 is a novel cold-induced protein and mediates neuroprotective effects of RBM3. Curr. Biol. 27, 638–650. doi:10.1016/j.cub.2017.01.047
Béjot, Y., Bailly, H., Graber, M., Garnier, L., Laville, A., Dubourget, L., et al. (2019). Impact of the ageing population on the burden of stroke: the dijon stroke registry. Neuroepidemiology 52, 78–85. doi:10.1159/000492820
Benizri, S., Gissot, A., Martin, A., Vialet, B., Grinstaff, M. W., and Barthélémy, P. (2019). Bioconjugated oligonucleotides: recent developments and therapeutic applications. Bioconjug. Chem. 30, 366–383. doi:10.1021/acs.bioconjchem.8b00761
Blanco, M., Campos, F., Rodriguez-Yanez, M., Arias, S., Fernandez-Ferro, J., Gómez-Sánchez, J. C., et al. (2012). Neuroprotection or increased brain damage mediated by temperature in stroke is time dependent. PLoS One 7, e30700. doi:10.1371/journal.pone.0030700
Campos, F., Blanco, M., Barral, D., Agulla, J., Ramos-Cabrer, P., and Castillo, J. (2012). Influence of temperature on ischemic brain: basic and clinical principles. Neurochem. Int. 60, 495–505. doi:10.1016/j.neuint.2012.02.003
Campos, F., Sobrino, T., Vieites-Prado, A., Perez-Mato, M., Rodriguez-Yanez, M., Blanco, M., et al. (2013). Hyperthermia in human ischemic and hemorrhagic stroke: similar outcome, different mechanisms. PLoS One 8, e78429. doi:10.1371/journal.pone.0078429
Chip, S., Zelmer, A., Ogunshola, O. O., Felderhoff-Mueser, U., Nitsch, C., Buhrer, C., et al. (2011). The RNA-binding protein RBM3 is involved in hypothermia induced neuroprotection. Neurobiol. Dis. 43, 388–396. doi:10.1016/j.nbd.2011.04.010
Danno, S., Nishiyama, H., Higashitsuji, H., Yokoi, H., Xue, J.-H., Itoh, K., et al. (1997). Increased transcript level of RBM3, a member of the glycine-rich RNA-binding protein family, in human cells in response to cold stress. Biochem. Biophys. Res. Commun. 236, 804–807. doi:10.1006/bbrc.1997.7059
Derry, J. M., Kerns, J. A., and Francke, U. (1995). RBM3, a novel human gene in Xp11. 23 with a putative RNA-binding domain. Hum. Mol. Genet. 4, 2307–2311. doi:10.1093/hmg/4.12.2307
Dresios, J., Aschrafi, A., Owens, G. C., Vanderklish, P. W., Edelman, G. M., and Mauro, V. P. (2005). Cold stress-induced protein Rbm3 binds 60S ribosomal subunits, alters microRNA levels, and enhances global protein synthesis. Proc. Natl. Acad. Sci. U. S. A. 102, 1865–1870. doi:10.1073/pnas.0409764102
Dumitrascu, O. M., Lamb, J., and Lyden, P. D. (2016). Still cooling after all these years: meta-analysis of pre-clinical trials of therapeutic hypothermia for acute ischemic stroke. J. Cereb. Blood Flow. Metab. 36, 1157–1164. doi:10.1177/0271678X16645112
Ferhat, L., Soussi, R., Masse, M., Kyriatzis, G., Girard, S. D., Gassiot, F., et al. (2024). A peptide-neurotensin conjugate that crosses the blood-brain barrier induces pharmacological hypothermia associated with anticonvulsant, neuroprotective and anti-inflammatory properties following status epilepticus in mice. Biorxiv. 2024.07 (15), 603208. doi:10.1101/2024.07.15.603208
Hu, W., Liu, Y., and Yan, J. (2014). Microarray meta-analysis of RNA-binding protein functions in alternative polyadenylation. PLoS One 9, e90774. doi:10.1371/journal.pone.0090774
Hu, X., Shi, Q., Zhou, X., He, W., Yi, H., Yin, X., et al. (2007). Transgenic mice overexpressing reticulon 3 develop neuritic abnormalities. EMBO J. 26, 2755–2767. doi:10.1038/sj.emboj.7601707
Hu, Y., Liu, Y., Quan, X., Fan, W., Xu, B., and Li, S. (2022). RBM3 is an outstanding cold shock protein with multiple physiological functions beyond hypothermia. J. Cell. Physiol. 237, 3788–3802. doi:10.1002/jcp.30852
Jackson, T. C., Janesko-Feldman, K., Carlson, S. W., Kotermanski, S. E., and Kochanek, P. M. (2019). Robust RBM3 and β-klotho expression in developing neurons in the human brain. J. Cereb. Blood Flow. Metab. 39, 2355–2367. doi:10.1177/0271678x19878889
Jackson, T. C., Kotermanski, S. E., and Kochanek, P. M. (2018). Infants uniquely express high levels of RBM3 and other cold-adaptive neuroprotectant proteins in the human brain. Dev. Neurosci. 40, 325–336. doi:10.1159/000493637
Jackson, T. C., Manole, M. D., Kotermanski, S. E., Jackson, E. K., Clark, R. S., and Kochanek, P. M. (2015). Cold stress protein RBM3 responds to temperature change in an ultra-sensitive manner in young neurons. Neurosci 305, 268–278. doi:10.1016/j.neuroscience.2015.08.012
Jiang, Q., Geng, X., and Ding, Y. (2022). Abstract WP24: pharmacological hypothermia (PH) promotes plasticity through RNA binding motif 3 (RBM3)-Mediated cold shock pathway following ischemic stroke. Stroke 53, AWP24. doi:10.1161/str.53.suppl_1.WP24
Johnson, C. O., Nguyen, M., Roth, G. A., Nichols, E., Alam, T., Abate, D., et al. (2019). Global, regional, and national burden of stroke, 1990–2016: a systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 18, 439–458. doi:10.1016/S1474-4422(19)30034-1
Kita, H., Carmichael, J., Swartz, J., Muro, S., Wyttenbach, A., Matsubara, K., et al. (2002). Modulation of polyglutamine-induced cell death by genes identified by expression profiling. Hum. Mol. Genet. 11, 2279–2287. doi:10.1093/hmg/11.19.2279
Laustriat, D., Gide, J., Barrault, L., Chautard, E., Benoit, C., Auboeuf, D., et al. (2015). In vitro and in vivo modulation of alternative splicing by the biguanide metformin. Mol. Ther. Nucleic Acids 4, E262. doi:10.1038/mtna.2015.35
Lin, P., Lin, C., and Diao, L. (2024). RBM3 ameliorates acute brain injury-induced inflammation and oxidative stress by stabilizing GAS6 mRNA through Nrf2 signaling pathway. Neurosci. Biobehav. Rev. 547, 74–87. doi:10.1016/j.neuroscience.2024.03.028
Liu, Y., Hu, W., Murakawa, Y., Yin, J., Wang, G., Landthaler, M., et al. (2013). Cold-induced RNA-binding proteins regulate circadian gene expression by controlling alternative polyadenylation. Sci. Rep. 3, 2054. doi:10.1038/srep02054
Ma, Y., He, Z., Wang, J., Zheng, P., Ma, Z., Liang, Q., et al. (2024). Mild hypothermia promotes neuronal differentiation of human neural stem cells via RBM3-SOX11 signaling pathway. iScience 27, 109435. doi:10.1016/j.isci.2024.109435
Matsuda, A., Ogawa, M., Yanai, H., Naka, D., Goto, A., Ao, T., et al. (2011). Generation of mice deficient in RNA-binding motif protein 3 (RBM3) and characterization of its role in innate immune responses and cell growth. Biochem. Biophys. Res. Commun. 411, 7–13. doi:10.1016/j.bbrc.2011.06.038
Mirzoyev, S. A., Mcleod, C. J., Bunch, T. J., Bell, M. R., and White, R. D. (2010). Hypokalemia during the cooling phase of therapeutic hypothermia and its impact on arrhythmogenesis. Resuscitation 81, 1632–1636. doi:10.1016/j.resuscitation.2010.08.007
Miyazawa, T., Tamura, A., Fukui, S., and Hossmann, K.-A. (2003). Effect of mild hypothermia on focal cerebral ischemia. Review of experimental studies. Neurol. Res. 25, 457–464. doi:10.1179/016164103101201850
Nakamura, K., and Morrison, S. F. (2011). Central efferent pathways for cold-defensive and febrile shivering. J. Physiol. 589, 3641–3658. doi:10.1113/jphysiol.2011.210047
Peretti, D., Bastide, A., Radford, H., Verity, N., Molloy, C., Martin, M. G., et al. (2015). RBM3 mediates structural plasticity and protective effects of cooling in neurodegeneration. Nature 518, 236–239. doi:10.1038/nature14142
Peretti, D., Smith, H. L., Verity, N., Humoud, I., De Weerd, L., Swinden, D. P., et al. (2021). TrkB signaling regulates the cold-shock protein RBM3-mediated neuroprotection. Life Sci. Alliance. 4, e202000884. doi:10.26508/lsa.202000884
Pérez-Mato, M., López-Arias, E., Bugallo-Casal, A., Correa-Paz, C., Arias, S., Rodríguez-Yáñez, M., et al. (2024). New perspectives in Neuroprotection for ischemic stroke. Neurosci 550, 30–42. doi:10.1016/j.neuroscience.2024.02.017
Pilotte, J., Cunningham, B. A., Edelman, G. M., and Vanderklish, P. W. (2009). Developmentally regulated expression of the cold-inducible RNA-binding motif protein 3 in euthermic rat brain. Brain Res. 1258, 12–24. doi:10.1016/j.brainres.2008.12.050
Polderman, K. H. (2009). Mechanisms of action, physiological effects, and complications of hypothermia. Crit. Care Med. 37, S186–S202. doi:10.1097/CCM.0b013e3181aa5241
Powers, W. J., Rabinstein, A. A., Ackerson, T., Adeoye, O. M., Bambakidis, N. C., Becker, K., et al. (2019). Guidelines for the early management of patients with acute ischemic stroke: 2019 update to the 2018 guidelines for the early management of acute ischemic stroke: a guideline for healthcare professionals from the American Heart Association/American Stroke Association. Stroke 50, e344–e418. doi:10.1161/STR.0000000000000211
Preussner, M., Smith, H. L., Hughes, D., Zhang, M., Emmerichs, A. K., Scalzitti, S., et al. (2023). ASO targeting RBM3 temperature-controlled poison exon splicing prevents neurodegeneration in vivo. EMBO Mol. Med. 15, e17157. doi:10.15252/emmm.202217157
Rosenthal, L. M., Tong, G., Walker, C., Wowro, S. J., Krech, J., Pfitzer, C., et al. (2017). Neuroprotection via RNA-binding protein RBM3 expression is regulated by hypothermia but not by hypoxia in human SK-N-SH neurons. Hypoxia 5, 33–43. doi:10.2147/HP.S132462
Rosenthal, L. M., Tong, G., Wowro, S., Walker, C., Pfitzer, C., Bottcher, W., et al. (2020). A prospective clinical trial measuring the effects of cardiopulmonary bypass under mild hypothermia on the inflammatory response and regulation of cold-shock protein RNA-binding motif 3. Ther. Hypothermia Temp. Manag. 10, 60–70. doi:10.1089/ther.2018.0038
Ryan, J., Morey, J., Ramsdell, J., and Van Dolah, F. (2005). Acute phase gene expression in mice exposed to the marine neurotoxin domoic acid. Neurosci 136, 1121–1132. doi:10.1016/j.neuroscience.2005.08.047
Rzechorzek, N. M., Connick, P., Patani, R., Selvaraj, B. T., and Chandran, S. (2015). Hypothermic preconditioning of human cortical neurons requires proteostatic priming. eBioMedicine 2, 528–535. doi:10.1016/j.ebiom.2015.04.004
Saini, M., Saqqur, M., Kamruzzaman, A., Lees, K. R., Shuaib, A., and Investigators, V. (2009). Effect of hyperthermia on prognosis after acute ischemic stroke. Stroke 40, 3051–3059. doi:10.1161/STROKEAHA.109.556134
Sertel, S. M., Von Elling-Tammen, M. S., and Rizzoli, S. O. (2021). The mRNA-binding protein RBM3 regulates activity patterns and local synaptic translation in cultured hippocampal neurons. J. Neurosci. 41, 1157–1173. doi:10.1523/jneurosci.0921-20.2020
Shao, W., and Jin, T. (2022). Hepatic hormone FGF21 and its analogues in clinical trials. Chronic Dis. Transl. Med. 8, 19–25. doi:10.1016/j.cdtm.2021.08.005
Si, W., Li, Z., Huang, Z., Ye, S., Li, X., Li, Y., et al. (2020). RNA binding protein motif 3 inhibits oxygen-glucose deprivation/reoxygenation-induced apoptosis through promoting stress granules formation in PC12 cells and rat primary cortical neurons. Front. Cell. Neurosci. 14, 559384. doi:10.3389/fncel.2020.559384
Smart, F., Aschrafi, A., Atkins, A., Owens, G. C., Pilotte, J., Cunningham, B. A., et al. (2007). Two isoforms of the cold-inducible mRNA-binding protein RBM3 localize to dendrites and promote translation. J. Neurochem. 101, 1367–1379. doi:10.1111/j.1471-4159.2007.04521.x
Stauffenberg, E., Pauletti, A., Haltenhof, T., Shao-Fang, Y., Heyd, F., and Bröer, S. (2024). Investigating the antiepileptogenic and neuroprotective potential of “hypothermia in a syringe”. AES Annu. Meet. 12/7/2024. Available at: https://aesnet.org/abstractslisting/investigating-the-antiepileptogenic-and-neuroprotective-potential-of-hypothermia-in-a-syringe (Accessed January 30, 2025).
Sureban, S., Ramalingam, S., Natarajan, G., May, R., Subramaniam, D., Bishnupuri, K., et al. (2008). Translation regulatory factor RBM3 is a proto-oncogene that prevents mitotic catastrophe. Oncogene 27, 4544–4556. doi:10.1038/onc.2008.97
Teng, F. Y. H., and Tang, B. L. (2013). Nogo/RTN4 isoforms and RTN3 expression protect SH-SY5Y cells against multiple death insults. Mol. Cell. Biochem. 384, 7–19. doi:10.1007/s11010-013-1776-6
Van Der Worp, H. B., Macleod, M. R., Bath, P. M., Demotes, J., Durand-Zaleski, I., Gebhardt, B., et al. (2014). EuroHYP-1: European multicenter, randomized, phase III clinical trial of therapeutic hypothermia plus best medical treatment vs. best medical treatment alone for acute ischemic stroke. Int. J. Stroke. 9, 642–645. doi:10.1111/ijs.12294
Van Der Worp, H. B., Sena, E. S., Donnan, G. A., Howells, D. W., and Macleod, M. R. (2007). Hypothermia in animal models of acute ischaemic stroke: a systematic review and meta-analysis. Brain 130, 3063–3074. doi:10.1093/brain/awm083
Wan, Y., Tian, H., Wang, H., Wang, D., Jiang, H., and Fang, Q. (2023). Selective intraarterial hypothermia combined with mechanical thrombectomy for acute cerebral infarction based on microcatheter technology: a single-center, randomized, single-blind controlled study. Front. Neurol. 14, 1039816. doi:10.3389/fneur.2023.1039816
Wang, S., Chen, X., and Qiao, T. (2021). Long non-coding RNA MIR4435-2HG promotes the progression of head and neck squamous cell carcinoma by regulating the miR-383-5p/RBM3 axis. Oncol. Rep. 45, 99–10. doi:10.3892/or.2021.8050
Wellmann, S., BüHrer, C., Moderegger, E., Zelmer, A., Kirschner, R., Koehne, P., et al. (2004). Oxygen-regulated expression of the RNA-binding proteins RBM3 and CIRP by a HIF-1-independent mechanism. J. Cell Sci. 117, 1785–1794. doi:10.1242/jcs.01026
Wood, T., and Thoresen, M. (2015). Physiological responses to hypothermia. Semin. Fetal Neonatal Med. 20, 87–96. doi:10.1016/j.siny.2014.10.005
World Health Organization (2024). The top 10 causes of death. Available at: https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (Accessed November 28, 2024).
Xia, W., Su, L., and Jiao, J. (2018). Cold-induced protein RBM3 orchestrates neurogenesis via modulating Yap mRNA stability in cold stress. J. Cell Biol. 217, 3464–3479. doi:10.1083/jcb.201801143
Yan, J., Goerne, T., Zelmer, A., Guzman, R., Kapfhammer, J. P., Wellmann, S., et al. (2019). The RNA-binding protein RBM3 promotes neural stem cell (NSC) proliferation under hypoxia. Front. Cell Dev. Biol. 7, 288. doi:10.3389/fcell.2019.00288
Yang, H. J., Ju, F., Guo, X. X., Ma, S. P., Wang, L., Cheng, B. F., et al. (2017). RNA-binding protein RBM3 prevents NO-induced apoptosis in human neuroblastoma cells by modulating p38 signaling and miR-143. Sci. Rep. 7, 41738. doi:10.1038/srep41738
Yang, H. J., Shi, X., Ju, F., Hao, B. N., Ma, S. P., Wang, L., et al. (2018). Cold shock induced protein RBM3 but not mild hypothermia protects human SH-SY5Y neuroblastoma cells from MPP(+)-Induced neurotoxicity. Front. Neurosci. 12, 298. doi:10.3389/fnins.2018.00298
Yang, H. J., Zhuang, R. J., Li, Y. B., Li, T., Yuan, X., Lei, B. B., et al. (2019). Cold-inducible protein RBM3 mediates hypothermic neuroprotection against neurotoxin rotenone via inhibition on MAPK signalling. J. Cell. Mol. Med. 23, 7010–7020. doi:10.1111/jcmm.14588
Zhang, L., Liang, W., Li, Y., Yan, J., Xue, J., Guo, Q., et al. (2021). Mild therapeutic hypothermia improves neurological outcomes in a rat model of cardiac arrest. Brain Res. Bull. 173, 97–107. doi:10.1016/j.brainresbull.2021.05.014
Zhang, Z., Liu, X., Yang, Z., and Mo, X. (2023). Study on the protective effect of RNA-binding motif protein 3 in mild hypothermia oxygen-glucose deprivation/reoxygenation cell model. Cryobiology 112, 104544. doi:10.1016/j.cryobiol.2023.05.001
Zhao, J., Liu, S., Li, K., Yang, Y., Zhao, Y., and Zhu, X. (2024). RBM3 promotes anti-inflammatory responses in microglia and serves as a neuroprotective target of ischemic stroke. Mol. Neurobiol. 61, 7384–7402. doi:10.1007/s12035-024-04052-4
Zhou, M., Yang, W.-L., Ji, Y., Qiang, X., and Wang, P. (2014). Cold-inducible RNA-binding protein mediates neuroinflammation in cerebral ischemia. B. B. A. - General Subjects 1840 (7), 2253–2261. doi:10.1016/j.bbagen.2014.02.027
Zhu, X., Bührer, C., and Wellmann, S. (2016a). Cold-inducible proteins CIRP and RBM3, a unique couple with activities far beyond the cold. Cell. Mol. Life Sci. 73, 3839–3859. doi:10.1007/s00018-016-2253-7
Zhu, X., Yan, J., Bregere, C., Zelmer, A., Goerne, T., Kapfhammer, J. P., et al. (2019). RBM3 promotes neurogenesis in a niche-dependent manner via IMP2-IGF2 signaling pathway after hypoxic-ischemic brain injury. Nat. Commun. 10, 3983. doi:10.1038/s41467-019-11870-x
Zhu, X., Zelmer, A., Kapfhammer, J. P., and Wellmann, S. (2016b). Cold-inducible RBM3 inhibits PERK phosphorylation through cooperation with NF90 to protect cells from endoplasmic reticulum stress. FASEB J. 30, 624–634. doi:10.1096/fj.15-274639
Keywords: brain, hypothermia, ischemia, modulation, neuroprotection, RBM3, stroke, therapy
Citation: Ávila-Gómez P, Vieites-Prado A, Correa-Paz C, Del Pozo-Filíu L, Palomar-Alonso N, Campos F and López-Arias E (2025) Therapeutic modulation of protein RBM3 for ischemic stroke treatment. Front. Pharmacol. 16:1555115. doi: 10.3389/fphar.2025.1555115
Received: 03 January 2025; Accepted: 12 February 2025;
Published: 07 March 2025.
Edited by:
Herve Boutin, INSERM U1253 Imagerie et Cerveau (iBrain), FranceReviewed by:
Lotfi Ferhat, L’Institut de NeuroPhysiopathologie, FranceXinzhou Zhu, Chinese Academy of Sciences (CAS), China
Copyright © 2025 Ávila-Gómez, Vieites-Prado, Correa-Paz, Del Pozo-Filíu, Palomar-Alonso, Campos and López-Arias. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Francisco Campos, ZnJhbmNpc2NvLmNhbXBvcy5wZXJlekBzZXJnYXMuZXM=; Esteban López-Arias, ZXN0ZWJhbi5sb3Blei5hcmlhc0BzZXJnYXMuZXM=