 Jian Li
Jian Li Xin Li
Xin Li Hongwei Liu*
Hongwei Liu*
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Pharmacol., 20 February 2025
Sec. Pharmacology of Anti-Cancer Drugs
Volume 16 - 2025 | https://doi.org/10.3389/fphar.2025.1551115
This article is part of the Research TopicDecoding Tumor Drug Resistance: Machine Learning’s Role from Molecules to TreatmentView all 10 articles
The isolation and application of biological macromolecules (BMMs) have become central in applied science today, with these compounds serving as anticancer, antimicrobial, and anti-inflammatory agents. Parthenolide (PTL), a naturally occurring sesquiterpene lactone derived from Tanacetum parthenium (feverfew), is among the most important of these BMMs. PTL has been extensively studied for its anticancer and anti-inflammatory properties, making it a promising candidate for further research and drug development. This review summarizes the anticancer and anti-inflammatory effects of PTL and its derivatives, with a focus on Micheliolide (MCL) and Dimethylaminomicheliolide (DMAMCL). These compounds, derived from PTL, have been developed to overcome PTL’s instability in acidic and basic conditions and its low solubility. We also explore their potential in targeted and combination therapies, providing a comprehensive overview of their therapeutic mechanisms and highlighting their significance in future cancer treatment strategies.
Medicinal plants have long been revered for their therapeutic properties. These plants produce a wide variety of compounds, both constitutive and secondary metabolites, many of which are biologically active. Aromatic compounds, often phenolic or oxygen-substituted derivatives, are commonly found in plant-based extracts used in traditional medicine (Newman and Cragg, 2020). Sesquiterpene lactones (SLs), a group of plant metabolites, have been utilized for centuries to treat various inflammatory conditions, including high fevers, headaches, stomachaches, toothaches, rheumatoid arthritis (RA), menstrual irregularities, and other inflammatory disorders (Mathema et al., 2012). Parthenolide (PTL), a notable sesquiterpene lactone, is primarily derived from Tanacetum parthenium and exhibits both anti-inflammatory and anticancer properties. The nucleophilic lactone ring, methylene, and epoxide groups of PTL enable rapid interactions with biological targets (Smolinski and Pestka, 2005).
SLs are predominantly sourced from the Asteraceae family and have been recognized since the 1970s for their anti-inflammatory and various other biological activities, including antitumor, cytotoxic, and antibacterial effects (Merfort, 2011). PTL, Micheliolide (MCL), and Dimethylaminomicheliolide (DMAMCL) are key members of the SL family, exhibiting both anti-inflammatory and antitumor activities. However, PTL’s instability in acidic and basic conditions limits its use (Jin et al., 2007). MCL, which is more stable than PTL, exhibits a half-life of 2.64 h, though it remains unstable at room temperature (Zhang et al., 2012). DMAMCL, a prodrug of MCL, continuously releasing MCL over an 8-h period in the plasma. It demonstrates improved stability, stronger activity, and lower toxicity, while also having the ability to cross the blood-brain barrier (Zhang et al., 2012; An et al., 2015; Xi X. N. et al., 2019). The growing body of research on these compounds’ anti-inflammatory and anticancer activities suggests that they hold significant therapeutic potential. This review aims to compile and synthesize available data, providing insights and opening new research avenues.
PTL, a potent anti-inflammatory agent, exerts its effects primarily through the inhibition of the Toll-like receptor 4 (TLR4)-mediated activation of Akt, mTOR, and NF-κB pathways. This reduces the production of key inflammatory mediators, positioning PTL as a potential therapeutic for various inflammation-related disorders, including psoriasis (Zhan et al., 2024), RA (Williams et al., 2020), and colon inflammation (Liu Y. J. et al., 2020). PTL has also demonstrated promising results in reducing mortality in patients with severe COVID-19, likely due to its ability to lower IL-6 cytokine levels (Liu Y. J. et al., 2020; Bahrami et al., 2020).
PTL has emerged as the first small molecule identified to target cancer stem cells (Ghantous et al., 2013). Recent studies have revealed PTL’s potential to induce apoptosis in various cancer cell lines, including acute lymphoblastic leukemia (Ortiz-Reyes et al., 2024), melanoma (Dorostgou et al., 2024), lung cancer (Cai et al., 2024), colorectal cancer (Liu Y. C. et al., 2017), and pancreatic cancer (Liu W. et al., 2017; Denda et al., 2024). PTL has shown significant antiproliferative effects in multiple cancer models, including lung cancer, where it suppresses IGF-1R-mediated PI3K/Akt/FoxO3α signaling (Lin et al., 2017) (Table 1). PTL has also been found to inhibit the growth of colorectal cancer cells through the inhibition of USP7/Wnt signaling (Li et al., 2020) and affect cell migration and invasion via the TGF-β1 and NF-κB pathways (Zhu et al., 2019; Kim et al., 2017a). However, the drug’s high lipophilicity and poor solubility in blood plasma limit its oral bioavailability. To overcome these limitations, a water-soluble analogue, Dimethylaminoparthenolide (DMAPT), has been developed, showing improved bioavailability and enhanced therapeutic efficacy (Deraska et al., 2018; Flores-Lopez et al., 2018). Furthermore, DMAPT was also regarded as a potential agent for breast cancer, bladder cancer, and lung cancer (Wang et al., 2017; Shanmugam et al., 2011; Song et al., 2014).
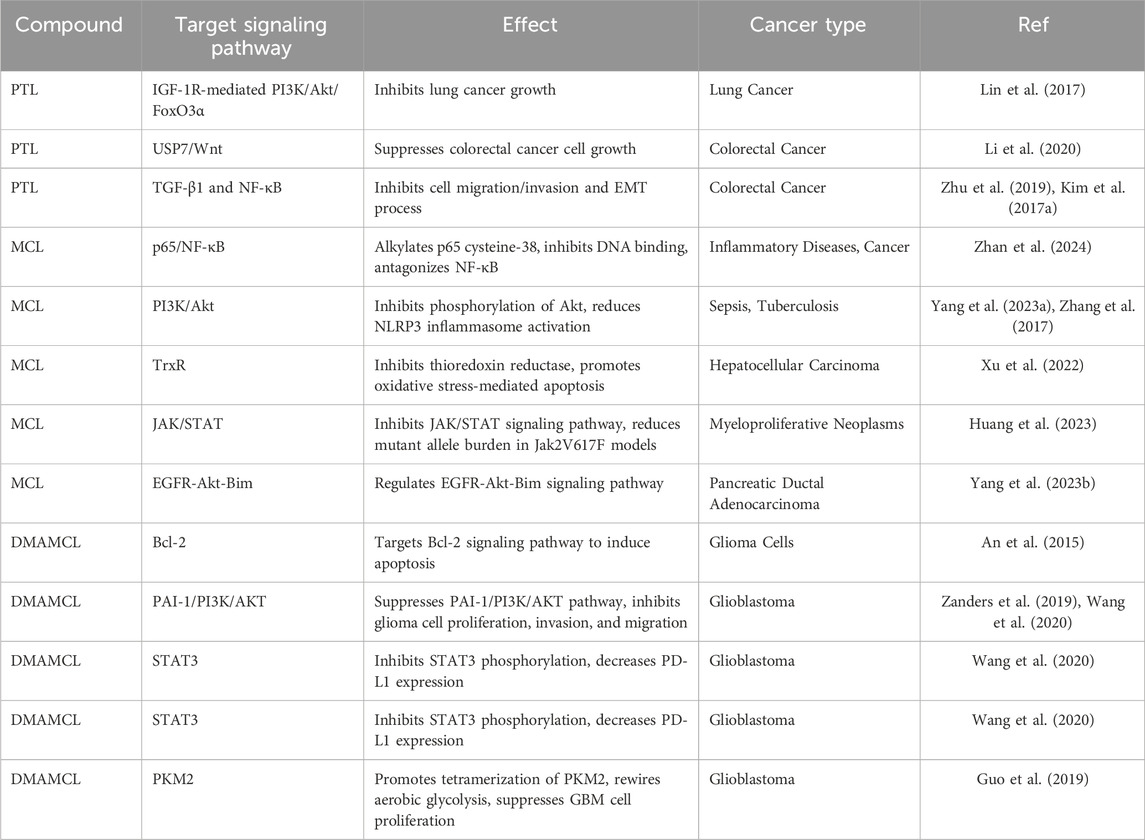
Table 1. Anti-tumor mechanisms involving inhibition of signaling pathways.
Despite the promising antitumor effects of PTL and DMAPT, their lack of selectivity between normal and cancerous cells remains a significant challenge. To address this issue, tumor-targeted formulations such as nanocarriers and liposomes have been developed, which improve drug efficacy while minimizing side effects. For example, the combination of PTL and ginsenoside CK in liposomes has shown improved efficacy in lung cancer treatment (Jin et al., 2018). Furthermore, PLGA-antiCD44-PTL nanoparticles have enhanced drug delivery and bioavailability by selectively targeting leukemic cells (Darwish et al., 2019). However, previous studies have highlighted several disadvantages. For instance, issues such as poor bioavailability and low water solubility have been noted in some studies (An et al., 2022). Another disadvantage is that these biological extracts are obtained from the plants. However, the seasonal availability of plants makes it challenging to consistently source the proper and unique plants needed to obtain these biologically active molecules, posing a significant hurdle in the development of PTL derivatives.
MCL has been synthesized to overcome the instability of PTL under acidic and basic conditions. MCL is a natural guaianolide sesquiterpene lactone found in Michelia compressa and Michelia champaca, and can also be synthesized from PTL in vitro. MCL exhibits anti-inflammatory and anticancer activities similar to PTL but is more stable under both in vitro and in vivo conditions (Peng et al., 2019). MCL inhibits NF-κB activation by alkylating p65 cysteine-38, preventing DNA binding and exerting an antagonist effect on NF-κB (Ghantous et al., 2013). Given NF-κB’s pivotal role in inflammation and immune responses, MCL’s ability to modulate this pathway holds significant therapeutic potential in treating inflammatory diseases such as RA and inflammatory bowel disease (Viennois et al., 2014).
Besides, MCL exerts anti-inflammatory effects by modulating the PI3K/Akt signaling pathway. In Alzheimer’s disease (AD) model mice, MCL improves cognitive function and reduces brain inflammation by inhibiting glial cell activation and cytokine secretion. These effects are linked to the reduction of Akt phosphorylation and the downregulation of key inflammatory mediators (Yang G. et al., 2023). Similarly, in a doxorubicin (DOX)-induced cardiotoxicity model, MCL mitigates cardiac injury, improves cardiac function, and alleviates histopathological damage through the regulation of the PI3K/Akt pathway, decreasing phosphorylated Akt, Bad, and caspase-3 levels, while increasing phosphorylated PTEN (Kalantary-Charvadeh et al., 2019). In addition to its effects on the NF-κB signaling pathway and the PI3K/Akt pathway, MCL also activates the MAPK signaling pathway in cancer and fibrotic models. In pancreatic and colon cancer cells, MCL enhances ROS production and regulates cell death mechanisms such as autophagy, paraptosis, and ferroptosis via MAPK activation (Yang M. H. et al., 2024). Furthermore, MCL suppress the Mtdh/BMP/MAPK pathway in renal fibrosis, reducing fibrotic markers and restoring epithelial function in both in vivo and in vitro models (Peng et al., 2019).
Inflammatory factors are pivotal in inflammatory diseases progression and significantly influence the efficacy of therapies (Zhai et al., 2024; Xiao et al., 2019; Xiao et al., 2020; Zhang et al., 2024; Ni et al., 2021; Zhang et al., 2023a; Ni et al., 2020). The NF-κB signaling pathway has been directly implicated in the development of various inflammation-associated disorders, as demonstrated in studies using transgenic mouse models. In a dextran sodium sulfate (DSS)-induced murine model of colitis, DSS treatment resulted in a significant increase in the expression levels of pro-inflammatory cytokines such as IL-1β, TNF-α, and IL-6, which play key roles in the pathogenesis of inflammatory bowel disease (IBD). Treatment with MCL was shown to significantly reduce the DSS-induced release of these cytokines by modulating the NF-κB signaling pathway (Viennois et al., 2014). Similarly, these cytokines are central to the pathophysiology of rheumatoid arthritis (RA) (Chen et al., 2019). The therapeutic effects of MCL in RA were further confirmed in a murine model, with potential mechanisms including alterations in the expression of cytokines such as C5/C5a, TIMP-1, M-CSF, and BLC (Xu et al., 2015).
Sepsis, another inflammation-associated condition, is triggered when lipopolysaccharide binds to TLR4, initiating the production of large quantities of inflammatory cytokines, which play a pivotal role in the onset of sepsis (Savva and Roger, 2013). In addition to its effect on NF-κB activation, MCL has been shown to reduce the levels of various LPS-induced inflammatory cytokines by inhibiting the phosphorylation of p70S6K (Thr389) and Akt (Ser473) (Qin et al., 2016). Moreover, a recent study highlighted MCL’s therapeutic potential in sepsis caused by Gram-positive bacteria and antibiotic-resistant bacteria through similar mechanisms (Jiang et al., 2017). Further investigation into MCL’s regulatory effects on the immunopathology of tuberculosis, induced by Mycobacterium tuberculosis (Mtb), reinforced its therapeutic potential in treating inflammation-related disorders. MCL dose-dependently reduced the secretion of inflammatory cytokines such as TNF-α and IL-1β in response to Mtb. Additionally, MCL inhibited Mtb-induced phosphorylation of Akt (Ser473) in Raw264.7 cells, thereby attenuating the activation of the NLRP3 inflammasome (Zhang et al., 2017).
Recent studies have also expanded on the therapeutic potential of MCL in other inflammation-related diseases. A study synthesizing novel ferulic acid-michelin lactone (FA-MCL) derivatives found that one such derivative exhibited significant anti-inflammatory activity in LPS-induced inflammatory responses, suggesting its potential as a candidate drug for acute lung injury (ALI) (Duan et al., 2024). Additionally, MCL has been shown to alleviate severe acute pancreatitis (SAP)-induced pancreatic injury by activating Nrf2-regulated antioxidant pathways and inhibiting NF-κB p65-mediated inflammation, further emphasizing its anti-inflammatory therapeutic potential in SAP (Wu et al., 2024). As a neuroprotective agent, MCL may also inhibit neuroinflammation in neurodegenerative disorders (Sun et al., 2017). In Alzheimer’s disease (AD), MCL mitigates neuroinflammation by inhibiting the NF-κB and PI3K/Akt pathways in glial cells, leading to a reduction in inflammatory mediators and improvements in cognitive function, positioning MCL as a promising therapeutic candidate for AD (Yang G. et al., 2023). Furthermore, by promoting autophagy and facilitating the degradation of the NLR pyrin domain three (NLRP3) inflammasome, MCL reduces inflammation and tissue damage, offering potential as a novel anti-inflammatory therapy for radiation-induced enteropathy (RIE) (Wu et al., 2021).
Immune microenvironment is pivotal in tumor progression (Yang H. et al., 2024; Sun Z. et al., 2024; Wang J. et al., 2023), and significantly influence the efficacy of cancer therapies (Deng et al., 2024; Xia et al., 2023; Sun L. et al., 2024; Zhang J. et al., 2023; Li X. et al., 2024). The NF-κB signaling pathway, a central component of the inflammation-fibrosis-cancer axis, is implicated not only in inflammatory diseases but also in pathological states where inflammation serves as a fundamental factor, such as in various forms of cancer (Czauderna et al., 2019; Zhao et al., 2023). The transition from inflammation to cancer may be mediated by several regulatory molecules (Wang Y. et al., 2023; Wang et al., 2025), including chemokines, proinflammatory cytokines, TNF, and IL-6, which are crucial in promoting the growth, proliferation, and invasion of cancer cells (Singh et al., 2019; Jiang et al., 2024; Zhang et al., 2023c; Zhao et al., 2024; Shao et al., 2024; Wang et al., 2024). Notably, colitis-associated cancer (CAC), which arises from chronic colonic inflammation, highlights the potential therapeutic role of MCL in this context (Viennois et al., 2014). Specifically, IKKβ-induced NF-κB activation in enterocytes plays a significant role in tumor initiation during the early stages, while IKK2-mediated NF-κB activity in myeloid cells promotes tumor progression by stimulating the expression of proinflammatory cytokines that act as tumor growth factors (Greten et al., 2004). Therefore, MCL could be considered a potential therapeutic agent for treating CAC through its inhibitory effects on NF-κB.
Hepatocellular carcinoma (HCC) is another inflammation-driven cancer (Zhai et al., 2023; Zhang S. et al., 2023; Zhang et al., 2023e), where inflammatory cytokines such as TNF-α and IL-6 activate downstream targets including NF-κB, JNK, and STAT3, thereby promoting tumorigenesis (Czauderna et al., 2019; Yang et al., 2019; Kumari et al., 2016; Kitamura et al., 2017). MCL, a natural thioredoxin reductase (TrxR) inhibitor, has emerged as a promising therapeutic agent for HCC. It induces immunogenic cell death (ICD) in HCC cells via the generation of reactive oxygen species (ROS) and the induction of endoplasmic reticulum (ER) stress (Xu et al., 2022) (Table 2). The anticancer efficacy of MCL is primarily attributed to its specific inhibition of TrxR, which results in the accumulation of ROS and the promotion of oxidative stress-induced apoptosis in malignant cells. MCL covalently binds to the Sec residue at position 498 of TrxR, thereby impairing its biological function, reducing TrxR activity, increasing oxidized Trx, and accumulating ROS. This disruption of redox homeostasis sensitizes cancer cells to ionizing radiation (IR) and enhances their cytotoxic response. Mechanistic studies have further suggested that MCL’s inhibition of TrxR contributes significantly to enhanced radiosensitivity, offering potential as both a chemotherapeutic agent and a radiosensitizer in cancer treatment (Zhang et al., 2022). In a xenograft liver cancer model, MCL was found to inhibit tumor growth by promoting apoptosis and disrupting the actin cytoskeleton (Yu et al., 2019). Thus, MCL presents substantial therapeutic potential for the treatment of liver cancer.
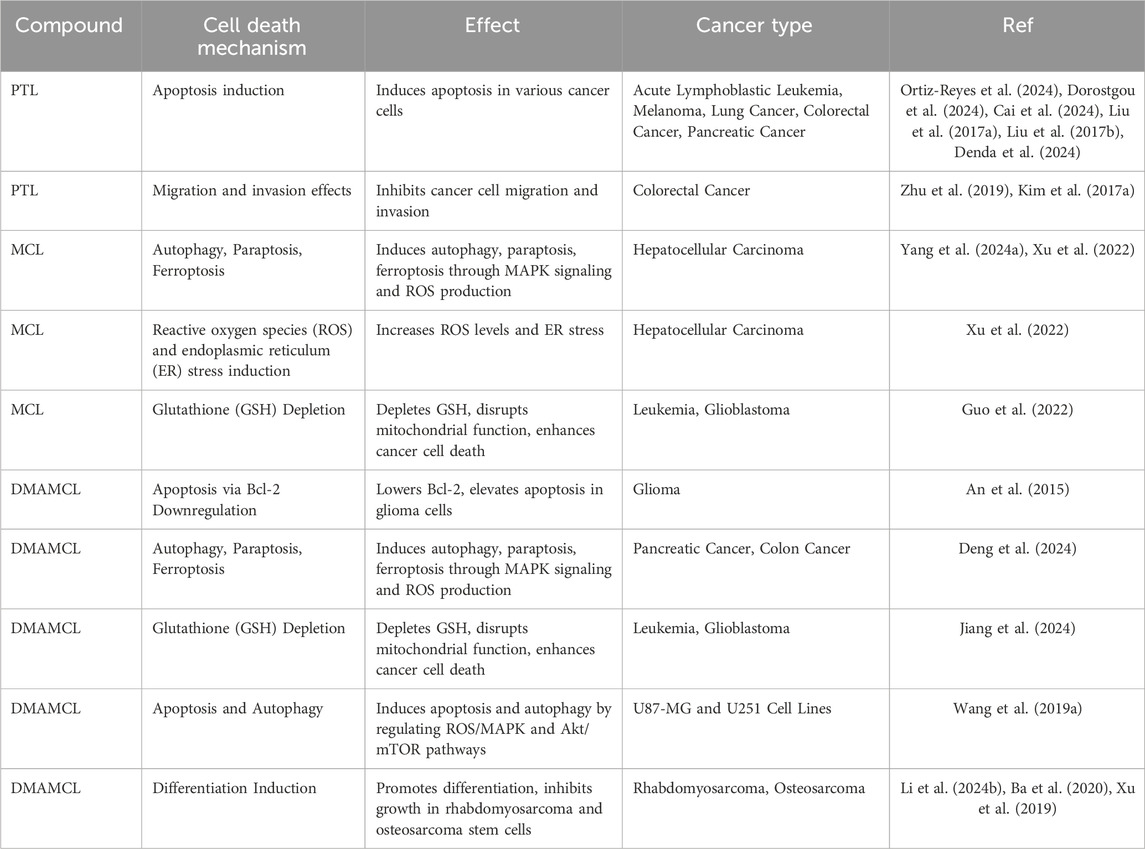
Table 2. Anti-tumor mechanisms involving induction of cell death and apoptosis.
Gastric carcinoma, originating from the epithelium of the gastric mucosa, is one of the most prevalent malignancies globally, with an increasing incidence, particularly among younger populations, largely due to dietary changes, rising work-related stress, and Helicobacter pylori infection. The primary treatment for advanced gastric cancer involves a combination of neoadjuvant chemoradiotherapy, molecular-targeted therapy, and immunotherapy (Song et al., 2017). However, prognosis remains poor for patients with advanced disease (Tirino et al., 2018), highlighting the critical need to improve therapeutic outcomes. Recent studies have indicated that IL-6 is upregulated in gastric cancer tissues, serving as a marker for poor prognosis. This upregulation is associated with IL-6’s role in promoting tumor cell invasion, metastasis, and angiogenesis (Unver and McAllister, 2018). Additionally, a mouse study on gastric tumorigenesis induced by N-methyl-N-nitrosourea revealed that IL-6 promotes gastric cancer cell proliferation through the activation of STAT3, as demonstrated by comparing IL-6 knockout mice with wild-type mice (Kinoshita et al., 2013). Targeting and blocking the IL-6/STAT3 pathway therefore represents an effective therapeutic strategy for gastric cancer. Studies on MCL have shown that it suppresses gastric cancer growth by blocking the IL-6/STAT3 pathway (Tang et al., 2019). Specifically, in both cellular and in vivo studies, treatment with MCL resulted in a reduction of IL-6 expression and inhibited the STAT3 signaling pathway. Furthermore, the restoration of the STAT3 pathway through the addition of exogenous IL-6 supports the notion that MCL exerts its anti-cancer effects by downregulating IL-6, thereby blocking STAT3 activation (Tang et al., 2019). Hence, MCL not only holds potential as a treatment for gastric cancer but could also improve the prognosis of patients with advanced gastric cancer.
Beyond its impact on previously discussed malignancies, MCL exhibits considerable therapeutic promise across a range of other cancers, including pancreatic, hematologic, and breast cancers. In pancreatic and colon cancer cell lines, MCL elicits anticancer responses by stimulating autophagy, paraptosis, and ferroptosis, processes that are mediated through the activation of MAPK signaling and the generation of reactive oxygen species (ROS) (Xu et al., 2022). Moreover, MCL augments the effectiveness of ruxolitinib in managing myeloproliferative neoplasms (MPNs) by targeting the JAK/STAT signaling cascade. This enhancement is accomplished through covalent attachment to cysteine residues on STAT3 and STAT5, leading to diminished phosphorylation and a reduction in mutant allele burden in Jak2V617F-mutated models (Huang et al., 2023). The active metabolite of MCL, known as ACT001, demonstrates antitumor activity in pancreatic ductal adenocarcinoma (PDAC) by restraining cell proliferation, promoting apoptosis, and increasing ROS levels. The fundamental mechanism encompasses modulation of the EGFR-Akt-Bim signaling pathway, as observed in both in vitro and in vivo studies (Yang J. et al., 2023). In leukemia and glioblastoma cell types, MCL elicits strong anticancer effects by initiating oxidative stress, mainly via the depletion of glutathione (GSH), thereby impairing mitochondrial function and promoting cancer cell apoptosis. The combination with GSH biosynthesis inhibitors, such as L-buthionine sulfoximine (BSO), intensifies this effect, presenting a viable approach to surmount resistance to conventional cancer treatments (Guo et al., 2022). Additionally, MCL has been demonstrated to counteract tamoxifen resistance in breast cancer (BC) by downregulating the expression of Amidohydrolase 1 (ASAH1), adjusting the ROS/AKT signaling pathway, and activating the NRF2/KEAP1 antioxidant mechanism. This results in the suppression of cell proliferation and improved therapeutic effectiveness against tamoxifen-resistant BC cells (Han et al., 2024). In summary, MCL exhibits extensive anticancer potential through the activation of multiple cell death pathways, the augmentation of other therapeutic modalities, and the ability to overcome drug resistance, thereby establishing it as a multifaceted candidate for cancer therapy.
DMAMCL, the dimethyl amino Michael adduct of MCL, demonstrates superior stability, enhanced activity, and reduced toxicity compared to MCL. When administered orally, DMAMCL exhibits a significantly longer half-life than its counterpart, DMAPT, and exerts potent anti-inflammatory effects (Viennois et al., 2014). As previously discussed, MCL has shown efficacy in treating inflammatory bowel diseases and other inflammatory conditions. Furthermore, DMAMCL has also been identified as a promising therapeutic agent for diabetic kidney disease (DKD) and neuroinflammation, exhibiting effects similar to those of MCL. Notably, DMAMCL has been observed to induce downregulation of metadherin (Mtdh), thereby inhibiting NF-κB signaling and suppressing downstream inflammatory cytokines. This mechanism suggests that DMAMCL could attenuate Mtdh-mediated renal inflammation in DKD (Liu et al., 2019). Additionally, DMAMCL has been reported to alleviate NLRP3-mediated neuroinflammation, potentially inhibiting the progression of Parkinson’s disease (PD) by modulating the NLRP3 inflammasome (Liu et al., 2020b).
DMAMCL metabolizes slowly in plasma, gradually releasing MCL as its active metabolite, which contributes to its improved pharmacokinetic properties and potential as a promising anticancer agent. In hepatocellular carcinoma (HCC), DMAMCL triggers apoptosis and induces cell cycle arrest at the G2/M phase by activating reactive oxygen species (ROS) production and inhibiting the PI3K/Akt pathway (Yao et al., 2020). Thus, DMAMCL presents as a viable therapeutic candidate for the treatment of HCC.
Glioblastoma (GBM), recognized as one of the most malignant and aggressive primary brain tumors, presents significant treatment challenges. Standard therapy, which includes surgical resection followed by radiotherapy and chemotherapy with temozolomide (TMZ), often results in poor prognosis due to tumor heterogeneity, limited central nervous system (CNS) penetration, and drug resistance (Zanders et al., 2019). Consequently, there is an urgent need for new therapeutic agents or combination therapies to address these obstacles. DMAMCL is currently undergoing phase I clinical trials for the treatment of GBM (trial ID: ACTRN12616000228482) and has been designated as an orphan drug for GBM by the FDA. Previous studies have elucidated the mechanisms by which MCL targets gliomas, particularly its interaction with PAI-1, a direct target of DMAMCL. ACT001, a metabolite of DMAMCL, binds to PAI-1, thereby inhibiting the PAI-1/PI3K/AKT pathway, which in turn suppresses glioma cell proliferation, invasion, and migration. Additionally, DMAMCL has been shown to inhibit STAT3 phosphorylation by directly binding to STAT3, reducing PD-L1 expression in glioma cells, as phosphorylated STAT3 binds to the PD-L1 promoter to modulate its transcription (Zanders et al., 2019; Wang et al., 2020). Furthermore, MCL, released from DMAMCL, selectively targets monomeric PKM2 and promotes its tetramerization, thereby altering the glycolytic metabolism of GBM cells and suppressing their proliferation (Guo et al., 2019). Additionally, DMAMCL may target the Bcl-2 signaling pathway, further enhancing its antitumor efficacy in glioma cells (An et al., 2015). Finally, DMAMCL induces apoptosis and autophagy through regulation of the ROS/MAPK and Akt/mTOR signaling pathways in U87-MG and U251 glioblastoma cell lines (Wang Y. et al., 2019).
DMAMCL promotes differentiation and inhibits the growth of rhabdomyosarcoma (RMS) by inducing morphological changes, enhancing muscle differentiation markers, and downregulating DLL1, a key ligand in the Notch signaling pathway. This positions DMAMCL as a potential agent for differentiation-based therapies in RMS (Li Q. et al., 2024). In addition, DMAMCL has demonstrated promising antitumor effects in RMS and osteosarcoma, the most common soft tissue sarcoma and primary bone malignancy, respectively, particularly in pediatric and adolescent populations. The underlying mechanism of DMAMCL’s efficacy in these cancers is likely associated with its induction of Bim, modulation of the NF-κB pathway, ROS generation in RMS cell lines, and reduction of osteosarcoma stem cell stemness (Ba et al., 2020; Xu et al., 2019).
In acute myeloid leukemia (AML), DMAMCL has been shown to decrease the percentage of stem cells in primary AML cells (Zhang et al., 2012). As mentioned earlier, MCL targets PKM2 and induces its irreversible tetramerization, which suppresses the proliferation of GBM cells. Furthermore, DMAMCL significantly inhibits leukemia cell growth as a PKM2-targeted therapeutic agent (Li J. et al., 2018).
In conclusion, DMAMCL represents an eco-friendly, less toxic alternative to many conventional chemical anticancer agents. Given its promising therapeutic profiles across various cancers, DMAMCL could emerge as a versatile, effective compound for cancer treatment.
DMAMCL has been extensively characterized and compared with other natural compounds, such as MCL and PLT. The published findings indicate that DMAMCL offers several advantages over both MCL and PLT. Notably, DMAMCL has been identified as a potent suppressor of GBM cell proliferation by targeting pyruvate kinase M2 (PKM2) and facilitating the rewiring of aerobic glycolysis (Guo et al., 2019). In the context of peritoneal fibrosis (PF), a primary cause of ultrafiltration failure in patients undergoing long-term peritoneal dialysis, DMAMCL emerges as a promising new compound due to its low toxicity, high stability, and sustained release of MCL, presenting a significant advantage in terms of safety and efficacy (Li et al., 2019). Furthermore, research has demonstrated that the LC50 values of DMAMCL against C6 and U-87MG glioma cell lines in vitro were 27.18 ± 1.89 μM and 20.58 ± 1.61 μM, respectively. Importantly, DMAMCL was found to significantly reduce the expression of the anti-apoptotic gene Bcl-2 and promote apoptosis in C6 and U-87MG cells in a dose-dependent manner. These findings suggest that DMAMCL holds considerable potential for the treatment of glioma (An et al., 2015).
Combination therapy has been shown to provide superior therapeutic effects compared to monotherapy by inhibiting tumor growth and improving survival rates (Zhou et al., 2021; Liu et al., 2022; Li et al., 2023; Zhang et al., 2023f). Moreover, long-term high-dose monotherapy often results in severe side effects and the development of drug resistance (Jin et al., 2024). Consequently, combining two or more drugs can enhance therapeutic efficacy, reduce adverse drug reactions, and target various symptoms or comorbidities. In recent years, PTL, MCL, and DMAMCL have demonstrated significant potential in combination therapy, with promising applications in the treatment of various diseases and in improving existing treatment regimens.
The mechanisms and therapeutic effects of PTL in treating various diseases have been extensively studied. Notably, the synergistic benefits of PTL in combination with other drugs have gradually been recognized as more effective than monotherapy. In human acute myeloid leukemia (AML) cells, combining PTL with pan-histone deacetylase inhibitors (HDACIs) potentiates HDACI lethality by inhibiting NF-κB and the subsequent activation of the SAPK/JNK pathway via MKK7 (Dai et al., 2010). Moreover, it was found that PTL treatment upregulates NADPH levels via the pentose phosphate pathway (PPP) and activates Nrf2-mediated responses. To overcome these challenges, a novel triple-drug regimen known as PDT—comprising 2-deoxyglucose, PTL, and temsirolimus—has been designed and shown to exhibit potent therapeutic effects on primary AML cells, including those resistant to PTL alone (Pei et al., 2016).
Balsalazide, serving as a prodrug for 5-aminosalicylate, has exhibited promise in chemoprevention for patients with ulcerative colitis within the scope of treating human colorectal cancer and colitis-associated cancer (CAC) (Do et al., 2016). The simultaneous use of PTL with balsalazide leads to a synergistic inhibition of NF-κB activation, thereby enhancing therapeutic efficacy (Kim et al., 2017b). In the MDA-MB-468 breast cancer cell line, epirubicin promotes apoptosis and demonstrates anticancer properties, yet it also induces cytotoxic effects in normal cells. Combining PTL with epirubicin achieves similar antiproliferative and proapoptotic outcomes while reducing cytotoxicity in healthy cells, thereby mitigating side effects (Ghorbani-Abdi-Saedabad et al., 2020). Triple-negative breast cancer (TNBC) presents the poorest prognosis and the highest risk of distant metastasis among all breast cancer subtypes, highlighting the need for innovative strategies to improve tumor sensitivity to treatments. Research by Carlisi et al. showed that the combination of SAHA and PTL increased the sensitivity of MDA-MB-231 breast cancer cells to PTL’s cytotoxic effects (Carlisi et al., 2015). Moreover, PTL was demonstrated to prevent TNBC cells from developing resistance to doxorubicin and mitoxantrone by suppressing the overexpression of Nrf2 (Carlisi et al., 2017).
Recent research also highlighted the potential of combination therapy in treating advanced hepatocellular carcinoma (HCC). The PTL nanocrystal delivery system, in combination with sorafenib (Sora), was used to treat tumors in mice with HCC, achieving a tumor inhibition rate of 81.86%, significantly higher than that of either PTL or Sora alone, indicating the combination’s ability to enhance the therapeutic effect while overcoming the poor water solubility of PTL (Liang et al., 2020). Moreover, PTL has been reported to enhance the sensitivity of gastric cancer cells to cisplatin (DDP) and to reverse drug resistance by inhibiting the STAT3 signaling pathway (Li H. et al., 2018). In the treatment of glioblastoma multiforme (GBM), temozolomide (TMZ) remains the standard chemotherapeutic drug, but its effectiveness is limited by chemoresistance. Inhibition of NF-κB has been shown to enhance anti-glioma activity and reduce TMZ-induced resistance by downregulating MGMT gene expression (Yu et al., 2018).
Given that DMAMCL, MCL, and PTL alone may not fully eradicate GBM, combination treatments involving ACT001 with other therapies have been explored. ACT001, in combination with cisplatin, has shown synergistic effects by suppressing the PAI-1/PI3K/AKT pathway, enhancing apoptosis in glioma cells and overcoming cisplatin resistance (Xi X. et al., 2019). DMAMCL is capable of crossing the blood-brain barrier (BBB) and accumulating in the brain (Xi X. N. et al., 2019), leading to decreased PD-L1 expression. Although side effects need to be addressed, DMAMCL, when combined with TMZ, may serve as an optimal treatment for GBM, reversing immunosuppression and inhibiting immune escape in GBM cells induced by TMZ (Wang S. et al., 2019). In Parkinson’s disease (PD), L-3,4-dihydroxyphenylalanine (L-DOPA) is the primary treatment, though long-term use may result in severe side effects. Studies have shown that DMAMCL, when administered in conjunction with a low dose of L-DOPA, yields therapeutic effects equivalent to those of high-dose L-DOPA in mouse models of PD, suggesting that DMAMCL enhances the curative effect of L-DOPA (Liu et al., 2020c). Furthermore, the combination of MCL and radiotherapy has been shown to improve the sensitivity of cancer tissues to radiation, enhancing the therapeutic effect. MCL promotes radiosensitivity in p53-deficient non-small cell lung cancer (NSCLC) by inhibiting the HIF-1α pathway, thereby promoting its degradation (Kong et al., 2020). Additionally, MCL-mediated depletion of glutathione (GSH) reduces KLF4-mediated cisplatin resistance in breast cancer cells (Jia et al., 2015).
Inflammation plays a significant role in tumor initiation and progression, with carcinogens in colon cancer and inflammatory bowel disease serving as prominent examples. Over the years, various biological macromolecules have been identified and developed as anticancer and anti-inflammatory agents to address these issues and associated infections. Notably, PLTs have achieved extensive application in the treatment of cancer and inflammatory disorders via modulating key signaling pathways, including NF-κB, STAT3, and MAPK, which are essential to both oncogenic progression and the regulation of immune responses. This review underscores the ability of PLTs to shield normal cells from apoptosis while selectively triggering apoptotic mechanisms in cancerous cells. Consequently, the findings presented herein offer significant insights that may drive future investigations, potentially leading to the development of targeted therapies that leverage the anticancer and anti-inflammatory properties of PLTs.
JL: Writing–original draft. XL: Writing–original draft. HL: Writing–original draft, Writing–review and editing.
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
An, T., Yin, H., Lu, Y., and Liu, F. (2022). The emerging potential of parthenolide nanoformulations in tumor therapy. Drug Des. Devel Ther. 16, 1255–1272. doi:10.2147/DDDT.S355059
An, Y., Guo, W., Li, L., Xu, C., Yang, D., Wang, S., et al. (2015). Micheliolide derivative DMAMCL inhibits glioma cell growth in vitro and in vivo. PLoS One 10, e0116202. doi:10.1371/journal.pone.0116202
Ba, G., Hua, Z., Xu, N., Zhang, S., Liu, Z., Thiele, C. J., et al. (2020). Novel agent DMAMCL suppresses osteosarcoma growth and decreases the stemness of osteosarcoma stem cell. Cell Cycle 19, 1530–1544. doi:10.1080/15384101.2020.1762041
Bahrami, M., Kamalinejad, M., Latifi, S. A., Seif, F., and Dadmehr, M. (2020). Cytokine storm in COVID-19 and parthenolide: preclinical evidence. Phytother. Res. 34, 2429–2430. doi:10.1002/ptr.6776
Cai, Z., Gao, L., Hu, K., and Wang, Q. M. (2024). Parthenolide enhances the metronomic chemotherapy effect of cyclophosphamide in lung cancer by inhibiting the NF-kB signaling pathway. World J. Clin. Oncol. 15, 895–907. doi:10.5306/wjco.v15.i7.895
Carlisi, D., De Blasio, A., Drago-Ferrante, R., Di Fiore, R., Buttitta, G., Morreale, M., et al. (2017). Parthenolide prevents resistance of MDA-MB231 cells to doxorubicin and mitoxantrone: the role of Nrf2. Cell Death Discov. 3, 17078. doi:10.1038/cddiscovery.2017.78
Carlisi, D., Lauricella, M., D'Anneo, A., Buttitta, G., Emanuele, S., di Fiore, R., et al. (2015). The synergistic effect of SAHA and parthenolide in MDA-MB231 breast cancer cells. J. Cell Physiol. 230, 1276–1289. doi:10.1002/jcp.24863
Chen, Z., Bozec, A., Ramming, A., and Schett, G. (2019). Anti-inflammatory and immune-regulatory cytokines in rheumatoid arthritis. Nat. Rev. Rheumatol. 15, 9–17. doi:10.1038/s41584-018-0109-2
Czauderna, C., Castven, D., Mahn, F. L., and Marquardt, J. U. (2019). Context-dependent role of NF-κB signaling in primary liver cancer-from tumor development to therapeutic implications. Cancers (Basel) 11, 1053. doi:10.3390/cancers11081053
Dai, Y., Guzman, M. L., Chen, S., Wang, L., Yeung, S. K., Pei, X. Y., et al. (2010). The NF (Nuclear factor)-κB inhibitor parthenolide interacts with histone deacetylase inhibitors to induce MKK7/JNK1-dependent apoptosis in human acute myeloid leukaemia cells. Br. J. Haematol. 151, 70–83. doi:10.1111/j.1365-2141.2010.08319.x
Darwish, N. H. E., Sudha, T., Godugu, K., Bharali, D. J., Elbaz, O., El-Ghaffar, H. A. A., et al. (2019). Novel targeted nano-parthenolide molecule against NF-kB in acute myeloid leukemia. Molecules 24, 2103. doi:10.3390/molecules24112103
Denda, Y., Matsuo, Y., Sugita, S., Eguchi, Y., Nonoyama, K., Murase, H., et al. (2024). The natural product parthenolide inhibits both angiogenesis and invasiveness and improves gemcitabine resistance by suppressing nuclear factor κB activation in pancreatic cancer cell lines. Nutrients 16, 705. doi:10.3390/nu16050705
Deng, Y., Shi, M., Yi, L., Naveed Khan, M., Xia, Z., and Li, X. (2024). Eliminating a barrier: aiming at VISTA, reversing MDSC-mediated T cell suppression in the tumor microenvironment. Heliyon 10, e37060. doi:10.1016/j.heliyon.2024.e37060
Deraska, P. V., O'Leary, C., Reavis, H. D., Labe, S., Dinh, T. K., Lazaro, J. B., et al. (2018). NF-kappaB inhibition by dimethylaminoparthenolide radiosensitizes non-small-cell lung carcinoma by blocking DNA double-strand break repair. Cell Death Discov. 4, 10. doi:10.1038/s41420-017-0008-3
Do, E. J., Hwang, S. W., Kim, S. Y., Ryu, Y. M., Cho, E. A., Chung, E. J., et al. (2016). Suppression of colitis-associated carcinogenesis through modulation of IL-6/STAT3 pathway by balsalazide and VSL#3. J. Gastroenterol. Hepatol. 31, 1453–1461. doi:10.1111/jgh.13280
Dorostgou, Z., Hosseiny, M., Bahrami, A., Zhiani, R., and Mohtashami, M. (2024). Parthenolide inhibits tumor cell growth and metastasis in melanoma A2058 cells. Curr. Med. Chem. 32. doi:10.2174/0109298673334309240924081449
Duan, X., Liu, N., Lv, K., Wang, J., Li, M., Zhang, Y., et al. (2024). Synthesis and anti-inflammatory activity of ferulic acid-sesquiterpene lactone hybrids. Molecules 29, 936. doi:10.3390/molecules29050936
Flores-Lopez, G., Moreno-Lorenzana, D., Ayala-Sanchez, M., Aviles-Vazquez, S., Torres-Martinez, H., Crooks, P. A., et al. (2018). Parthenolide and DMAPT induce cell death in primitive CML cells through reactive oxygen species. J. Cell Mol. Med. 22, 4899–4912. doi:10.1111/jcmm.13755
Ghantous, A., Sinjab, A., Herceg, Z., and Darwiche, N. (2013). Parthenolide: from plant shoots to cancer roots. Drug Discov. Today 18, 894–905. doi:10.1016/j.drudis.2013.05.005
Ghorbani-Abdi-Saedabad, A., Hanafi-Bojd, M. Y., Parsamanesh, N., Tayarani-Najaran, Z., Mollaei, H., and Hoshyar, R. (2020). Anticancer and apoptotic activities of parthenolide in combination with epirubicin in mda-mb-468 breast cancer cells. Mol. Biol. Rep. 47, 5807–5815. doi:10.1007/s11033-020-05649-3
Greten, F. R., Eckmann, L., Greten, T. F., Park, J. M., Li, Z. W., Egan, L. J., et al. (2004). IKKbeta links inflammation and tumorigenesis in a mouse model of colitis-associated cancer. Cell 118, 285–296. doi:10.1016/j.cell.2004.07.013
Guo, J., Liu, K., Wang, J., Jiang, H., Zhang, M., Liu, Y., et al. (2022). A rational foundation for Micheliolide-based combination strategy by targeting redox and metabolic circuit in cancer cells. Biochem. Pharmacol. 200, 115037. doi:10.1016/j.bcp.2022.115037
Guo, J., Xue, Q., Liu, K., Ge, W., Liu, W., Wang, J., et al. (2019). Dimethylaminomicheliolide (DMAMCL) suppresses the proliferation of glioblastoma cells via targeting pyruvate kinase 2 (PKM2) and rewiring aerobic glycolysis. Front. Oncol. 9, 993. doi:10.3389/fonc.2019.00993
Han, X., Zhang, Y., Li, Y., Lin, Z., Fu, Z., Wang, C., et al. (2024). MCL restrained ROS/AKT/ASAH1 pathway to therapy tamoxifen resistance breast cancer by stabilizing NRF2. Cell Prolif. 57, e13700. doi:10.1111/cpr.13700
Huang, H., Liu, J., Yang, L., Yan, Y., Chen, M., Li, B., et al. (2023). Micheliolide exerts effects in myeloproliferative neoplasms through inhibiting STAT3/5 phosphorylation via covalent binding to STAT3/5 proteins. Blood Sci. 5, 258–268. doi:10.1097/BS9.0000000000000168
Jia, Y., Zhang, C., Zhou, L., Xu, H., Shi, Y., and Tong, Z. (2015). Micheliolide overcomes KLF4-mediated cisplatin resistance in breast cancer cells by downregulating glutathione. Onco Targets Ther. 8, 2319–2327. doi:10.2147/OTT.S88661
Jiang, S., Yang, X., Lin, Y., Liu, Y., Tran, L. J., Zhang, J., et al. (2024). Unveiling Anoikis-related genes: a breakthrough in the prognosis of bladder cancer. J. Gene Med. 26, e3651. doi:10.1002/jgm.3651
Jiang, X., Wang, Y., Qin, Y., He, W., Benlahrech, A., Zhang, Q., et al. (2017). Micheliolide provides protection of mice against Staphylococcus aureus and MRSA infection by down-regulating inflammatory response. Sci. Rep. 7, 41964. doi:10.1038/srep41964
Jin, P., Madieh, S., and Augsburger, L. L. (2007). The solution and solid state stability and excipient compatibility of parthenolide in feverfew. AAPS PharmSciTech 8, E105. doi:10.1208/pt0804105
Jin, W., Zhang, Y., Zhao, Z., and Gao, M. (2024). Developing targeted therapies for neuroblastoma by dissecting the effects of metabolic reprogramming on tumor microenvironments and progression. Theranostics 14, 3439–3469. doi:10.7150/thno.93962
Jin, X., Zhou, J., Zhang, Z., and Lv, H. (2018). The combined administration of parthenolide and ginsenoside CK in long circulation liposomes with targeted tLyp-1 ligand induce mitochondria-mediated lung cancer apoptosis. Artif. Cells Nanomed Biotechnol. 46, S931–S942. doi:10.1080/21691401.2018.1518913
Kalantary-Charvadeh, A., Sanajou, D., Hemmati-Dinarvand, M., Marandi, Y., Khojastehfard, M., Hajipour, H., et al. (2019). Micheliolide protects against doxorubicin-induced cardiotoxicity in mice by regulating PI3K/Akt/NF-kB signaling pathway. Cardiovasc Toxicol. 19, 297–305. doi:10.1007/s12012-019-09511-2
Kim, S. L., Kim, S. H., Park, Y. R., Liu, Y. C., Kim, E. M., Jeong, H. J., et al. (2017b). Combined parthenolide and balsalazide have enhanced antitumor efficacy through blockade of NF-κB activation. Mol. Cancer Res. 15, 141–151. doi:10.1158/1541-7786.MCR-16-0101
Kim, S. L., Park, Y. R., Lee, S. T., and Kim, S. W. (2017a). Parthenolide suppresses hypoxia-inducible factor-1α signaling and hypoxia induced epithelial-mesenchymal transition in colorectal cancer. Int. J. Oncol. 51, 1809–1820. doi:10.3892/ijo.2017.4166
Kinoshita, H., Hirata, Y., Nakagawa, H., Sakamoto, K., Hayakawa, Y., Takahashi, R., et al. (2013). Interleukin-6 mediates epithelial-stromal interactions and promotes gastric tumorigenesis. PLoS One 8, e60914. doi:10.1371/journal.pone.0060914
Kitamura, H., Ohno, Y., Toyoshima, Y., Ohtake, J., Homma, S., Kawamura, H., et al. (2017). Interleukin-6/STAT3 signaling as a promising target to improve the efficacy of cancer immunotherapy. Cancer Sci. 108, 1947–1952. doi:10.1111/cas.13332
Kong, P., Yu, K. N., Yang, M., Almahi, W. A., Nie, L., Chen, G., et al. (2020). Micheliolide enhances radiosensitivities of p53-deficient non-small-cell lung cancer via promoting HIF-1α degradation. Int. J. Mol. Sci. 21, 3392. doi:10.3390/ijms21093392
Kumari, N., Dwarakanath, B. S., Das, A., and Bhatt, A. N. (2016). Role of interleukin-6 in cancer progression and therapeutic resistance. Tumour Biol. 37, 11553–11572. doi:10.1007/s13277-016-5098-7
Li, H., Lu, H., Lv, M., Wang, Q., and Sun, Y. (2018b). Parthenolide facilitates apoptosis and reverses drug-resistance of human gastric carcinoma cells by inhibiting the STAT3 signaling pathway. Oncol. Lett. 15, 3572–3579. doi:10.3892/ol.2018.7739
Li, J., Li, S., Guo, J., Li, Q., Long, J., Ma, C., et al. (2018a). Natural product Micheliolide (MCL) irreversibly activates pyruvate kinase M2 and suppresses leukemia. J. Med. Chem. 61, 4155–4164. doi:10.1021/acs.jmedchem.8b00241
Li, Q., Chen, Y., Chen, Y., Hua, Z., Gong, B., Liu, Z., et al. (2024b). Novel small molecule DMAMCL induces differentiation in rhabdomyosarcoma by downregulating of DLL1. Biomed. Pharmacother. 174, 116562. doi:10.1016/j.biopha.2024.116562
Li, S., Peng, F., Gong, W., Wu, J., Wang, Y., Xu, Z., et al. (2019). Dimethylaminomicheliolide ameliorates peritoneal fibrosis through the activation of autophagy. J. Mol. Med. Berl. 97, 659–674. doi:10.1007/s00109-019-01757-1
Li, X., Kong, L., Yang, Q., Duan, A., Ju, X., Cai, B., et al. (2020). Parthenolide inhibits ubiquitin-specific peptidase 7 (USP7), Wnt signaling, and colorectal cancer cell growth. J. Biol. Chem. 295, 3576–3589. doi:10.1074/jbc.RA119.011396
Li, X., Lin, Z., Zhao, F., Huang, T., Fan, W., Cen, L., et al. (2024a). Unveiling the cellular landscape: insights from single-cell RNA sequencing in multiple myeloma. Front. Immunol. 15, 1458638. doi:10.3389/fimmu.2024.1458638
Li, Z., Zhou, H., Xia, Z., Xia, T., Du, G., Franziska, S. D., et al. (2023). HMGA1 augments palbociclib efficacy via PI3K/mTOR signaling in intrahepatic cholangiocarcinoma. Biomark. Res. 11, 33. doi:10.1186/s40364-023-00473-w
Liang, P., Wu, H., Zhang, Z., Jiang, S., and Lv, H. (2020). Preparation and characterization of parthenolide nanocrystals for enhancing therapeutic effects of sorafenib against advanced hepatocellular carcinoma. Int. J. Pharm. 583, 119375. doi:10.1016/j.ijpharm.2020.119375
Lin, M., Bi, H., Yan, Y., Huang, W., Zhang, G., Zhang, G., et al. (2017). Parthenolide suppresses non-small cell lung cancer GLC-82 cells growth via B-Raf/MAPK/Erk pathway. Oncotarget 8, 23436–23447. doi:10.18632/oncotarget.15584
Liu, Q., Guo, X., Huang, Z., He, Q., Zhu, D., Zhang, S., et al. (2020b). Anti-neuroinflammatory effects of dimethylaminomylide (DMAMCL, i.e., ACT001) are associated with attenuating the NLRP3 in fl ammasome in MPTP-induced Parkinson disease in mice. Behav. Brain Res. 383, 112539. doi:10.1016/j.bbr.2020.112539
Liu, Q., Zhang, S., Zhu, D., Tang, X., Che, Y., and Feng, X. (2020c). The parthenolide derivative ACT001 synergizes with low doses of L-DOPA to improve MPTP-induced Parkinson's disease in mice. Behav. Brain Res. 379, 112337. doi:10.1016/j.bbr.2019.112337
Liu, W., Chen, X., Wang, Y., Chen, Y., Chen, S., Gong, W., et al. (2019). Micheliolide ameliorates diabetic kidney disease by inhibiting Mtdh-mediated renal inflammation in type 2 diabetic db/db mice. Pharmacol. Res. 150, 104506. doi:10.1016/j.phrs.2019.104506
Liu, W., Wang, X., Sun, J., Yang, Y., Li, W., and Song, J. (2017b). Parthenolide suppresses pancreatic cell growth by autophagy-mediated apoptosis. Onco Targets Ther. 10, 453–461. doi:10.2147/OTT.S117250
Liu, Y., Cheng, L., Song, X., Li, C., Zhang, J., and Wang, L. (2022). A TP53-associated immune prognostic signature for the prediction of the overall survival and therapeutic responses in pancreatic cancer. Math. Biosci. Eng. 19, 191–208. doi:10.3934/mbe.2022010
Liu, Y. C., Kim, S. L., Park, Y. R., Lee, S. T., and Kim, S. W. (2017a). Parthenolide promotes apoptotic cell death and inhibits the migration and invasion of SW620 cells. Intest. Res. 15, 174–181. doi:10.5217/ir.2017.15.2.174
Liu, Y. J., Tang, B., Wang, F. C., Tang, L., Lei, Y. Y., Luo, Y., et al. (2020a). Parthenolide ameliorates colon inflammation through regulating Treg/Th17 balance in a gut microbiota-dependent manner. Theranostics 10, 5225–5241. doi:10.7150/thno.43716
Mathema, V. B., Koh, Y. S., Thakuri, B. C., and Sillanpaa, M. (2012). Parthenolide, a sesquiterpene lactone, expresses multiple anti-cancer and anti-inflammatory activities. Inflammation 35, 560–565. doi:10.1007/s10753-011-9346-0
Merfort, I. (2011). Perspectives on sesquiterpene lactones in inflammation and cancer. Curr. Drug Targets 12, 1560–1573. doi:10.2174/138945011798109437
Newman, D. J., and Cragg, G. M. (2020). Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 83, 770–803. doi:10.1021/acs.jnatprod.9b01285
Ni, M., Zhang, J., Sosa, R., Zhang, H., Wang, H., Jin, D., et al. (2021). T-cell immunoglobulin and mucin domain-containing protein-4 is critical for kupffer cell homeostatic function in the activation and resolution of liver ischemia reperfusion injury. Hepatology 74, 2118–2132. doi:10.1002/hep.31906
Ni, M., Zhou, H., Zhang, J., Jin, D., Lu, T., Busuttil, R. W., et al. (2020). Isoform- and cell type-specific roles of glycogen synthase kinase 3 N-terminal serine phosphorylation in liver ischemia reperfusion injury. J. Immunol. 205, 1147–1156. doi:10.4049/jimmunol.2000397
Ortiz-Reyes, A. E., Garcia-Sanchez, S., Serrano, M., Nunez-Enriquez, J. C., Alvarado-Moreno, J. A., Montesinos, J. J., et al. (2024). In vitro purging of acute lymphoblastic leukemia (B-ALL) cells with the use of PTL, DMAPT, or PU-H71. Int. J. Mol. Sci. 25, 11707. doi:10.3390/ijms252111707
Pei, S., Minhajuddin, M., D'Alessandro, A., Nemkov, T., Stevens, B. M., Adane, B., et al. (2016). Rational design of a parthenolide-based drug regimen that selectively eradicates acute myelogenous leukemia stem cells. J. Biol. Chem. 291, 25280. doi:10.1074/jbc.A116.750653
Peng, F., Li, H., Li, S., Wang, Y., Liu, W., Gong, W., et al. (2019). Micheliolide ameliorates renal fibrosis by suppressing the Mtdh/BMP/MAPK pathway. Lab. Invest. 99, 1092–1106. doi:10.1038/s41374-019-0245-6
Qin, X., Jiang, X., Jiang, X., Wang, Y., Miao, Z., He, W., et al. (2016). Micheliolide inhibits LPS-induced inflammatory response and protects mice from LPS challenge. Sci. Rep. 6, 23240. doi:10.1038/srep23240
Savva, A., and Roger, T. (2013). Targeting toll-like receptors: promising therapeutic strategies for the management of sepsis-associated pathology and infectious diseases. Front. Immunol. 4, 387. doi:10.3389/fimmu.2013.00387
Shanmugam, R., Kusumanchi, P., Appaiah, H., Cheng, L., Crooks, P., Neelakantan, S., et al. (2011). A water soluble parthenolide analog suppresses in vivo tumor growth of two tobacco-associated cancers, lung and bladder cancer, by targeting NF-κB and generating reactive oxygen species. Int. J. Cancer 128, 2481–2494. doi:10.1002/ijc.25587
Shao, W., Lin, Z., Xiahou, Z., Zhao, F., Xu, J., Liu, X., et al. (2024). Single-cell RNA sequencing reveals that MYBL2 in malignant epithelial cells is involved in the development and progression of ovarian cancer. Front. Immunol. 15, 1438198. doi:10.3389/fimmu.2024.1438198
Singh, N., Baby, D., Rajguru, J. P., Patil, P. B., Thakkannavar, S. S., and Pujari, V. B. (2019). Inflammation and cancer. Ann. Afr. Med. 18, 121–126. doi:10.4103/aam.aam_56_18
Smolinski, A. T., and Pestka, J. J. (2005). Comparative effects of the herbal constituent parthenolide (Feverfew) on lipopolysaccharide-induced inflammatory gene expression in murine spleen and liver. J. Inflamm. (Lond) 2, 6. doi:10.1186/1476-9255-2-6
Song, J. M., Qian, X., Upadhyayya, P., Hong, K. H., and Kassie, F. (2014). Dimethylaminoparthenolide, a water soluble parthenolide, suppresses lung tumorigenesis through down-regulating the STAT3 signaling pathway. Curr. Cancer Drug Targets 14, 59–69. doi:10.2174/15680096113136660108
Song, Z., Wu, Y., Yang, J., Yang, D., and Fang, X. (2017). Progress in the treatment of advanced gastric cancer. Tumour Biol. 39, 1010428317714626. doi:10.1177/1010428317714626
Sun, L., Shao, W., Lin, Z., Lin, J., Zhao, F., and Yu, J. (2024b). Single-cell RNA sequencing explored potential therapeutic targets by revealing the tumor microenvironment of neuroblastoma and its expression in cell death. Discov. Oncol. 15, 409. doi:10.1007/s12672-024-01286-5
Sun, Z., Li, G., Tong, T., and Chen, J. (2017). Micheliolide suppresses LPS-induced neuroinflammatory responses. PLoS One 12, e0186592. doi:10.1371/journal.pone.0186592
Sun, Z., Wang, J., Fan, Z., Yang, Y., Meng, X., Ma, Z., et al. (2024a). Investigating the prognostic role of lncRNAs associated with disulfidptosis-related genes in clear cell renal cell carcinoma. J. Gene Med. 26, e3608. doi:10.1002/jgm.3608
Tang, X., Ding, Q., Chen, C., Chen, F., Zhou, X., Hong, C. J., et al. (2019). Micheliolide inhibits gastric cancer growth in vitro and in vivo via blockade of the IL-6/STAT3 pathway. Pharmazie 74, 175–178. doi:10.1691/ph.2019.8816
Tirino, G., Pompella, L., Petrillo, A., Laterza, M. M., Pappalardo, A., Caterino, M., et al. (2018). What's new in gastric cancer: the therapeutic implications of molecular classifications and future perspectives. Int. J. Mol. Sci. 19, 2659. doi:10.3390/ijms19092659
Unver, N., and McAllister, F. (2018). IL-6 family cytokines: key inflammatory mediators as biomarkers and potential therapeutic targets. Cytokine Growth Factor Rev. 41, 10–17. doi:10.1016/j.cytogfr.2018.04.004
Viennois, E., Xiao, B., Ayyadurai, S., Wang, L., Wang, P. G., Zhang, Q., et al. (2014). Micheliolide, a new sesquiterpene lactone that inhibits intestinal inflammation and colitis-associated cancer. Lab. Invest. 94, 950–965. doi:10.1038/labinvest.2014.89
Wang, B., Zhou, Y., Zhang, J., Jin, X., Wu, H., and Huang, H. (2020). Fructose-1,6-bisphosphatase loss modulates STAT3-dependent expression of PD-L1 and cancer immunity. Theranostics 10, 1033–1045. doi:10.7150/thno.38137
Wang, J., Zuo, Z., Yu, Z., Chen, Z., Tran, L. J., Zhang, J., et al. (2023a). Collaborating single-cell and bulk RNA sequencing for comprehensive characterization of the intratumor heterogeneity and prognostic model development for bladder cancer. Aging (Albany NY) 15, 12104–12119. doi:10.18632/aging.205166
Wang, J. F., Wang, J. S., Liu, Y., Ji, B., Ding, B. C., Wang, Y. X., et al. (2025). Knockdown of integrin β1 inhibits proliferation and promotes apoptosis in bladder cancer cells. Biofactors 51, e2150. doi:10.1002/biof.2150
Wang, R., Bhat-Nakshatri, P., Padua, M. B., Prasad, M. S., Anjanappa, M., Jacobson, M., et al. (2017). Pharmacological dual inhibition of tumor and tumor-induced functional limitations in a transgenic model of breast cancer. Mol. Cancer Ther. 16, 2747–2758. doi:10.1158/1535-7163.MCT-17-0717
Wang, S., Yao, F., Lu, X., Li, Q., Su, Z., Lee, J. H., et al. (2019b). Temozolomide promotes immune escape of GBM cells via upregulating PD-L1. Am. J. Cancer Res. 9, 1161–1171.
Wang, Y., Wang, J., He, J., Ji, B., Pang, Z., Wang, J., et al. (2023b). Comprehensive analysis of PRPF19 immune infiltrates, DNA methylation, senescence-associated secretory phenotype and ceRNA network in bladder cancer. Front. Immunol. 14, 1289198. doi:10.3389/fimmu.2023.1289198
Wang, Y., Wang, J., Liu, Y., Wang, X., and Ren, M. (2024). Multidimensional pan-cancer analysis of HSPA5 and its validation in the prognostic value of bladder cancer. Heliyon 10, e27184. doi:10.1016/j.heliyon.2024.e27184
Wang, Y., Zhang, J., Yang, Y., Liu, Q., Xu, G., Zhang, R., et al. (2019a). ROS generation and autophagosome accumulation contribute to the DMAMCL-induced inhibition of glioma cell proliferation by regulating the ROS/MAPK signaling pathway and suppressing the Akt/mTOR signaling pathway. Onco Targets Ther. 12, 1867–1880. doi:10.2147/OTT.S195329
Williams, B., Lees, F., Tsangari, H., Hutchinson, M. R., Perilli, E., and Crotti, T. N. (2020). Assessing the effects of parthenolide on inflammation, bone loss, and glial cells within a collagen antibody-induced arthritis mouse model. Mediat. Inflamm. 2020, 6245798. doi:10.1155/2020/6245798
Wu, C. Y., Wang, K. Q., Qin, Y. Y., Wang, H. W., Wu, M. M., Zhu, X. D., et al. (2024). Micheliolide ameliorates severe acute pancreatitis in mice through potentiating Nrf2-mediated anti-inflammation and anti-oxidation effects. Int. Immunopharmacol. 143, 113490. doi:10.1016/j.intimp.2024.113490
Wu, D. M., Li, J., Shen, R., Li, J., Yu, Y., Li, L., et al. (2021). Autophagy induced by Micheliolide alleviates acute irradiation-induced intestinal injury via inhibition of the NLRP3 inflammasome. Front. Pharmacol. 12, 773150. doi:10.3389/fphar.2021.773150
Xi, X., Liu, N., Wang, Q., Chu, Y., Yin, Z., Ding, Y., et al. (2019b). ACT001, a novel PAI-1 inhibitor, exerts synergistic effects in combination with cisplatin by inhibiting PI3K/AKT pathway in glioma. Cell Death Dis. 10, 757. doi:10.1038/s41419-019-1986-2
Xi, X. N., Liu, N., Wang, Q. Q., Wu, H. T., He, H. B., Wang, L. L., et al. (2019a). Pharmacokinetics, tissue distribution and excretion of ACT001 in Sprague-Dawley rats and metabolism of ACT001. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 1104, 29–39. doi:10.1016/j.jchromb.2018.11.004
Xia, Z., Chen, S., He, M., Li, B., Deng, Y., Yi, L., et al. (2023). Editorial: targeting metabolism to activate T cells and enhance the efficacy of checkpoint blockade immunotherapy in solid tumors. Front. Immunol. 14, 1247178. doi:10.3389/fimmu.2023.1247178
Xiao, J., Huang, K., Lin, H., Xia, Z., Zhang, J., Li, D., et al. (2020). Mogroside II(E) inhibits digestive enzymes via suppression of interleukin 9/interleukin 9 receptor signalling in acute pancreatitis. Front. Pharmacol. 11, 859. doi:10.3389/fphar.2020.00859
Xiao, J., Lin, H., Liu, B., Xia, Z., Zhang, J., and Jin, J. (2019). Decreased S1P and SPHK2 are involved in pancreatic acinar cell injury. Biomark. Med. 13, 627–637. doi:10.2217/bmm-2018-0404
Xu, H., Wang, J., Wang, C., Chang, G., Lin, Y., Zhang, H., et al. (2015). Therapeutic effects of Micheliolide on a murine model of rheumatoid arthritis. Mol. Med. Rep. 11, 489–493. doi:10.3892/mmr.2014.2767
Xu, N., Hua, Z., Ba, G., Zhang, S., Liu, Z., Thiele, C. J., et al. (2019). The anti-tumor growth effect of a novel agent DMAMCL in rhabdomyosarcoma in vitro and in vivo. J. Exp. Clin. Cancer Res. 38, 118. doi:10.1186/s13046-019-1107-1
Xu, Z., Xu, J., Sun, S., Lin, W., Li, Y., Lu, Q., et al. (2022). Mecheliolide elicits ROS-mediated ERS driven immunogenic cell death in hepatocellular carcinoma. Redox Biol. 54, 102351. doi:10.1016/j.redox.2022.102351
Yang, G., Hu, Y., Qin, X., Sun, J., Miao, Z., Wang, L., et al. (2023a). Micheliolide attenuates neuroinflammation to improve cognitive impairment of Alzheimer's disease by inhibiting NF-κB and PI3K/Akt signaling pathways. Heliyon 9, e17848. doi:10.1016/j.heliyon.2023.e17848
Yang, H., Li, Z., Zhu, S., Wang, W., Zhang, J., Zhao, D., et al. (2024b). Molecular mechanisms of pancreatic cancer liver metastasis: the role of PAK2. Front. Immunol. 15, 1347683. doi:10.3389/fimmu.2024.1347683
Yang, J., Li, Y., Han, X., Pei, X., Lin, Z., and Li, C. (2023b). The antitumor effect of the novel agent MCL/ACT001 in pancreatic ductal adenocarcinoma. J. Cancer Res. Clin. Oncol. 149, 5717–5728. doi:10.1007/s00432-022-04542-9
Yang, M. H., Baek, S. H., Jung, Y. Y., Um, J. Y., and Ahn, K. S. (2024a). Activation of autophagy, paraptosis, and ferroptosis by Micheliolide through modulation of the MAPK signaling pathway in pancreatic and colon tumor cells. Pathol. Res. Pract. 263, 155654. doi:10.1016/j.prp.2024.155654
Yang, Y. M., Kim, S. Y., and Seki, E. (2019). Inflammation and liver cancer: molecular mechanisms and therapeutic targets. Semin. Liver Dis. 39, 26–42. doi:10.1055/s-0038-1676806
Yao, S., Ye, J., Yin, M., and Yu, R. (2020). DMAMCL exerts antitumor effects on hepatocellular carcinoma both in vitro and in vivo. Cancer Lett. 483, 87–97. doi:10.1016/j.canlet.2020.04.003
Yu, L., Chen, W., Tang, Q., and Ji, K. Y. (2019). Micheliolide inhibits liver cancer cell growth via inducing apoptosis and perturbing actin cytoskeleton. Cancer Manag. Res. 11, 9203–9212. doi:10.2147/CMAR.S216870
Yu, Z., Chen, Y., Wang, S., Li, P., Zhou, G., and Yuan, Y. (2018). Inhibition of NF-κB results in anti-glioma activity and reduces temozolomide-induced chemoresistance by down-regulating MGMT gene expression. Cancer Lett. 428, 77–89. doi:10.1016/j.canlet.2018.04.033
Zanders, E. D., Svensson, F., and Bailey, D. S. (2019). Therapy for glioblastoma: is it working? Drug Discov. Today 24, 1193–1201. doi:10.1016/j.drudis.2019.03.008
Zhai, X., Xia, Z., Du, G., Zhang, X., Xia, T., Ma, D., et al. (2023). LRP1B suppresses HCC progression through the NCSTN/PI3K/AKT signaling axis and affects doxorubicin resistance. Genes Dis. 10, 2082–2096. doi:10.1016/j.gendis.2022.10.021
Zhai, X., Zhang, H., Xia, Z., Liu, M., Du, G., Jiang, Z., et al. (2024). Oxytocin alleviates liver fibrosis via hepatic macrophages. JHEP Rep. 6, 101032. doi:10.1016/j.jhepr.2024.101032
Zhan, Z. Y., Zhang, Z. H., Sun, R. H., Wu, Y. L., Nan, J. X., and Lian, L. H. (2024). A therapeutic strategy of parthenolide in improving imiquimod-induced psoriasis-like skin inflammation targeting IL-36/NETs through skin transdermal therapeutic system. Int. Immunopharmacol. 131, 111824. doi:10.1016/j.intimp.2024.111824
Zhang, H., Ni, M., Wang, H., Zhang, J., Jin, D., Busuttil, R. W., et al. (2023a). Gsk3β regulates the resolution of liver ischemia/reperfusion injury via MerTK. JCI Insight 8, e151819. doi:10.1172/jci.insight.151819
Zhang, H., Xia, T., Xia, Z., Zhou, H., Li, Z., Wang, W., et al. (2024). KIF18A inactivates hepatic stellate cells and alleviates liver fibrosis through the TTC3/Akt/mTOR pathway. Cell Mol. Life Sci. 81, 96. doi:10.1007/s00018-024-05114-5
Zhang, H., Zhai, X., Liu, Y., Xia, Z., Xia, T., Du, G., et al. (2023f). NOP2-mediated m5C modification of c-Myc in an EIF3A-dependent manner to reprogram glucose metabolism and promote hepatocellular carcinoma progression. Res. Wash D.C. 6, 0184. doi:10.34133/research.0184
Zhang, J., Chen, Y., and Fang, J. (2022). Targeting thioredoxin reductase by Micheliolide contributes to radiosensitizing and inducing apoptosis of HeLa cells. Free Radic. Biol. Med. 186, 99–109. doi:10.1016/j.freeradbiomed.2022.05.007
Zhang, J., Peng, G., Chi, H., Yang, J., Xie, X., Song, G., et al. (2023b). CD8 + T-cell marker genes reveal different immune subtypes of oral lichen planus by integrating single-cell RNA-seq and bulk RNA-sequencing. BMC Oral Health 23, 464. doi:10.1186/s12903-023-03138-0
Zhang, Q., Jiang, X., He, W., Wei, K., Sun, J., Qin, X., et al. (2017). MCL plays an anti-inflammatory role in Mycobacterium tuberculosis-induced immune response by inhibiting NF-κB and NLRP3 inflammasome activation. Mediat. Inflamm. 2017, 2432904. doi:10.1155/2017/2432904
Zhang, Q., Lu, Y., Ding, Y., Zhai, J., Ji, Q., Ma, W., et al. (2012). Guaianolide sesquiterpene lactones, a source to discover agents that selectively inhibit acute myelogenous leukemia stem and progenitor cells. J. Med. Chem. 55, 8757–8769. doi:10.1021/jm301064b
Zhang, S., Jiang, C., Jiang, L., Chen, H., Huang, J., Gao, X., et al. (2023d). Construction of a diagnostic model for hepatitis B-related hepatocellular carcinoma using machine learning and artificial neural networks and revealing the correlation by immunoassay. Tumour Virus Res. 16, 200271. doi:10.1016/j.tvr.2023.200271
Zhang, X., Zhang, P., Cong, A., Feng, Y., Chi, H., Xia, Z., et al. (2023c). Unraveling molecular networks in thymic epithelial tumors: deciphering the unique signatures. Front. Immunol. 14, 1264325. doi:10.3389/fimmu.2023.1264325
Zhang, X., Zhuge, J., Liu, J., Xia, Z., Wang, H., Gao, Q., et al. (2023e). Prognostic signatures of sphingolipids: understanding the immune landscape and predictive role in immunotherapy response and outcomes of hepatocellular carcinoma. Front. Immunol. 14, 1153423. doi:10.3389/fimmu.2023.1153423
Zhao, F., Hong, J., Zhou, G., Huang, T., Lin, Z., Zhang, Y., et al. (2024). Elucidating the role of tumor-associated ALOX5+ mast cells with transformative function in cervical cancer progression via single-cell RNA sequencing. Front. Immunol. 15, 1434450. doi:10.3389/fimmu.2024.1434450
Zhao, Z., Luo, Q., Liu, Y., Jiang, K., Zhou, L., Dai, R., et al. (2023). Multi-level integrative analysis of the roles of lncRNAs and differential mRNAs in the progression of chronic pancreatitis to pancreatic ductal adenocarcinoma. BMC Genomics 24, 101. doi:10.1186/s12864-023-09209-4
Zhou, W., Yun, Z., Wang, T., Li, C., and Zhang, J. (2021). BTF3-mediated regulation of BMI1 promotes colorectal cancer through influencing epithelial-mesenchymal transition and stem cell-like traits. Int. J. Biol. Macromol. 187, 800–810. doi:10.1016/j.ijbiomac.2021.07.106
Keywords: parthenolide, Micheliolide, Dimethylaminomicheliolide, antiinflammatory, anti-cancer
Citation: Li J, Li X and Liu H (2025) Sesquiterpene lactones and cancer: new insight into antitumor and anti-inflammatory effects of parthenolide-derived Dimethylaminomicheliolide and Micheliolide. Front. Pharmacol. 16:1551115. doi: 10.3389/fphar.2025.1551115
Received: 24 December 2024; Accepted: 28 January 2025;
Published: 20 February 2025.
Edited by:
Jing Zhang, University of South Dakota, United StatesReviewed by:
Zhijia Xia, Affiliated Hospital of Guilin Medical University, ChinaCopyright © 2025 Li, Li and Liu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hongwei Liu, c3l0amxod0AxNjMuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.