
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Pharmacol., 19 March 2025
Sec. Pharmacology of Anti-Cancer Drugs
Volume 16 - 2025 | https://doi.org/10.3389/fphar.2025.1540217
 Iera Hernandez-Unzueta1*Uxue Telleria-Gonzalez1
Iera Hernandez-Unzueta1*Uxue Telleria-Gonzalez1 Ana María Aransay2,3José Ezequiel Martin Rodriguez2Eduardo Sanz4
Ana María Aransay2,3José Ezequiel Martin Rodriguez2Eduardo Sanz4 Joana Márquez1*
Joana Márquez1*Cancer is one of the leading causes of death worldwide. Many therapies are being used to treat this disease, however, new treatments are now being implemented, since they are not always effective and their secondary effects represent one of the main reasons for cancer patients’ loss of life quality during the progression of the disease. In this scenario, Ocoxin is a mixture of plant extracts, amino acids, vitamins and minerals, known for its antioxidant, anti-inflammatory and immunoregulatory properties, which has shown to exert antitumor effects in many cancers. The aim of this study is to elucidate the mechanism of action of the compound in colorectal cancer, triple negative breast cancer, pancreatic cancer and prostate cancer. Analyses performed through RNA sequencing revealed that the main effect of Ocoxin appears to be the alteration of cell metabolism, especially inducing the process of ferroptosis. Nevertheless, the modulation of the cell cycle was also remarkable. Ocoxin altered 13 genes in common in all the four cancers that were not only associated to metabolism and cell cycle but were also involved in the integrated stress response and unfolded protein response, suggesting that the compound causes the induction of cell death through several pathways. Although the mechanisms vary according to the type of cancer, this study highlights the potential of Ocoxin as an adjunctive treatment to improve outcomes in cancer therapy.
Cancer poses a major challenge in our times, as it affects individuals all over the world, regardless of age, sex, race or socioeconomic status. Advances in research have led to a reduction of cancer-related deaths thanks to early detection methods and the improvement of treatments. However, cancer still poses a challenge to current medicine due to the limitations of therapies like toxicity and the fact that they occasionally become ineffective since tumors can develop resistance to them over time. In this context, radiotherapy and chemotherapy are the most extended remedies, which help to control tumor growth by destroying cancer cells, although they cause toxicity and weakness, resulting in a deterioration of patient’s health status. Additionally, less invasive methods are available, such as immunotherapy and targeted therapy, which reinforce and restore the immune system and target specific molecules, respectively. Still, these treatments can also cause side effects and tumors can develop resistance to them (Sarmento-Ribeiro et al., 2019; Labrie et al., 2022). Thus, further research is needed to develop more efficacious and less harmful treatments. As an alternative to prevent resistance and toxicity, combinations of several compounds are being administered to patients, which work together against cancer, each bringing their own unique strengths to bear on the disease (Woodring et al., 2023; Vladu et al., 2022). In this context, plant extracts obtained from seeds, leaves, roots or fruits are currently under investigation due to their demonstrated antitumor properties that target both cancer cells and the tumor microenvironment (Choudhari et al., 2020; Hosseini and Ghorbani, 2015; Pan et al., 2019; Rana et al., 2021; Xu et al., 2016; Khan et al., 2019; Masuelli et al., 2021). In fact, countless works report the capacity of flavonoids, polyphenols, vitamins, minerals and many other bioactive compounds to reduce tumor growth through numerous mechanisms, such as promoting apoptosis, blocking cell cycle checkpoints, inhibiting cell proliferation, reducing the migratory capacity, modulating the immune response and reducing the secretion of pro-tumoral factors, among others (Esmeeta et al., 2022; Lopes et al., 2017; Reed et al., 2018; Kim et al., 2021; Masuelli et al., 2021; Almatroodi et al., 2020; Khan et al., 2019; Zhang, W. et al., 2022; Zhang, Y. et al., 2023; Wang W. et al., 2023). In this regard, the possibility of combining first-line therapies used in clinics with these products is being studied. Indeed, certain natural compounds and their mixtures are already being administered to patients’ as adjuvants together with antitumor treatments, as they can enhance their effect (Harvie, 2014; Lee et al., 2021; Wang, S. et al., 2024; Farghadani and Naidu, 2022; Shi et al., 2023; Ng et al., 2022; Abadi et al., 2022; Talib et al., 2022; Chen, Z. et al., 2023). Besides, these supplements, as well as supporting nutrition, can help to mitigate adverse effects of treatments, such as loss of appetite, nausea and fatigue, thus improving tolerance to therapy and the patient’s quality of life (Harvie, 2014; Liu et al., 2021; Wang, K. et al., 2022). Green tea, for instance, is one of the most widely consumed beverages in the world that possesses many health benefits and may serve as a preventive element for cancer development. Its polyphenols, especially the epigallocatechin-3-gallate (EGCG), are known to protect against inflammation, inhibit tumor cell growth, modulate the secretion of metalloproteases and reduce the expression of immune checkpoint inhibitors, among others (Masuelli et al., 2021; Almatroodi et al., 2020). Similarly, many spices have shown antitumor effects against cancer: cinnamon induces apoptosis, impedes cell growth and hinders angiogenesis (Peng et al., 2024; Sadeghi et al., 2019); curcumin suppresses proliferation, enhances apoptosis and triggers mitochondrial stress in tumor cells (Abadi et al., 2022); ginger reduces cell viability, boosts cell death and modulates cytokine secretion (Mahomoodally et al., 2021); licorice also induces apoptosis, inhibits angiogenesis and impedes cell migration and invasion (Zhang, Y. et al., 2023); turmeric inhibits cell proliferation, promotes apoptosis and cell cycle arrest, and blocks Epithelial-Mesenchymal Transition (EMT) and invasion (Zhang, Y. et al., 2023; Wang X. et al., 2023). Likewise, some fruits like berries, grapes and many others, have shown beneficial effects against cancer by increasing cell death, causing cell cycle arrest and by reducing inflammation and the presence of Reactive Oxygen Species (ROS) (Masuelli et al., 2021; Esmeeta et al., 2022).
Related to this, Ocoxin is a mixture containing a combination of plant extracts (green tea, licorice, cinnamon), amino acids, minerals and vitamins. Although the compound was originally designed as a supplement to strengthen the immune system and to improve the general wellbeing of patients suffering from cancer and promote their overall health, several studies have reported its antitumor mechanisms. Briefly, Ocoxin showed to reduce tumor growth in vitro by decreasing cell proliferation, blocking cell cycle, augmenting apoptosis, modulating gene expression and altering the secretome of tumor cells while reducing chemoresistance and improving the effect of different chemotherapeutic drugs. Moreover, in vivo studies performed in mice revealed that, apart from reducing tumor volume, the natural supplement modulated the expression of secreted cytokines, diminished the infiltration of macrophages and Cancer Associated Fibroblasts (CAFs) and reduced angiogenesis (Hernandez-Unzueta et al., 2023; Hernandez-Unzueta et al., 2017; Benedicto et al., 2021; Márquez et al., 2016; Hernandez-Unzueta et al., 2019). Furthermore, several clinical trials have indicated the advantages of using Ocoxin together with cancer standard care therapies to alleviate side-effects and to improve the quality of life of patients (Shumsky et al., 2019; Fundora-Ramos et al., 2021; Kaidarova et al., 2019).
As we mentioned, extensive research has been performed on the antitumor effect of Ocoxin against both tumor cells and tumor microenvironment, which has permitted the comprehension of the biological processes in which the compound participates. However, much remains to be discovered about the underlying mechanisms and altered pathways that may be involved in the response to the treatment with Ocoxin. In order to gain a more complete understanding of how the natural mixture acts in the context of cancer, we have chosen to carry out RNA sequencing of the four cancer types mentioned above. This approach will allow us to identify which specific genes are altered in response to the compound and, in turn, help us to infer which biological pathways might be modified. This more detailed analysis will permit us to gain crucial insights into the possible mechanisms of action of Ocoxin in cancer and may provide a solid basis for future clinical studies and therapeutic developments.
Four different human cancer cell lines were cultured to carry out this study: the colorectal adenocarcinoma (COAD) SW620 cell line, the pancreatic adenocarcinoma (PAAD) BxPC-3 cell line, the prostatic adenocarcinoma (PRAD) LNCaP cell line and the triple-negative breast cancer (TNBC) HCC-70 cell line (ATCC, Manasas, VA, United States). All the cell lines were cultured with RPMI-1640 medium supplemented with 10% Fetal Bovine Serum (FBS) and Penicillin-Streptomycin-Amphotericin B solution (Gibco, Waltham, MA United States) at 37°C and 5% CO2 in a humidified atmosphere.
In order to assess the effect of Ocoxin against the mentioned cancer cell lines, several viability assays were carried out. To start with, 7 × 104 cells/mL were cultured onto 96-well plates under standard conditions. After 18 h, culture medium was removed and cells were treated with Ocoxin diluted (1:1000; 1:700; 1:500, 1:200, 1:100, 1:50, 1:25) in RPMI-1640 supplemented with 1% FBS for 24 h. Then, fresh Ocoxin was added again to the cells for another 24 h and cell viability was measured in the Fluoroscan Ascent (Thermo Labsystems, Waltham, MA, United States) with the Cell Counting Kit-8 (Sigma-Aldrich, St. Louis and Burlington, MO, United States) according to the manufacturer’s instructions. Finally, the half-maximal inhibitory concentration (IC50) of Ocoxin was established in all the cell lines to perform the following experiments.
Genetic alterations induced by Ocoxin were analyzed through mRNA sequencing in the previously mentioned human cancer cells. First, cells were cultured in 6-well plates under standard conditions at a concentration of 1.5 × 105 cells/mL. After 18 h, each cell line was treated with the IC50 dose of Ocoxin in 1% FBS supplemented medium for 48 h as previously described. The selected doses were as follows: PAAD cells with 1:500 of Ocoxin, TNBC with 1:700 of Ocoxin, PRAD with 1:50 of Ocoxin and COAD with 1:700 of Ocoxin. Then, cells were washed with PBS, the total RNA of adhered cells was isolated with the Total RNA purification kit (Norgen, Thorold, ON, Canada) and the quantity and integrity of the RNA was evaluated using the Qubit™ RNA HS Assay Kit (Invitrogen, Waltham, MA, United States) and Agilent RNA 600 NanoChips (Agilent Technologies, Santa Clara, CA, United States).
Afterwards, sequencing libraries were prepared using the TruSeq® Stranded mRNA Library Prep kit, TruSeq® RNA Single Indexes and TruSeq® RNA CD Index Plate (Illumina, San Diego, CA, United States). Then, in order to perform the mRNA sequencing, mRNA was purified, fragmented and primed for cDNA synthesis starting from 1 µg of total RNA with the SuperScriptTM II Reverse Transcriptase (Invitrogen, Waltham, MA, United States) for 10 min at 25°C, 15 min at 42°C, 15 min at 70°C and finished at 4°C. The second cDNA strand was synthesized with Illumina reagents at 16°C for 1 h, then A-tailing and adaptor ligation were performed and enrichment of libraries was achieved by PCR (30 s at 98°C; 15 cycles of 10 s at 98°C; 30 s at 55°C, 20 s at 72°C; 5 min at 72°C and pause at 4°C). Finally, libraries were visualized on an Agilent 2,100 Bioanalyzer using the Agilent High Sensitivity DNA kit (Agilent, Santa Clara, CA, United States) and quantified using QubitTM dsDNA HS DNA kit (Invitrogen, Waltham, MA, United States), quantified using Qubit™ dsDNA HS DNA kit (Invitrogen, Waltham, MA, United States) and sequenced in a NovaSeq 6,000 (Illumina, San Diego, CA, United States).
After, read adapter trimming was performed using fastp (OpenGene, 2022; Chen, 2023; Chen et al., 2018), alignment to reference genome hg38. bwa with STAR (Dobin et al., 2012; Dobin, 2022), PCR duplicates were removed from aligned BAM files with picard (Broad Institute MIT and Harvard, 2022; Dobin et al., 2012), and annotation followed by quantification of reads obtained through mRNAseq was got with SubRead’s FeatureCounts (Liao et al., 2013; Liao et al., 2022). Differential expression analysis was performed with the DESeq2 (Love et al., 2014) version 1.40.2 and the False Discovery Rate correction (FDR) was applied. Heatmaps were done with the library ComplexHeatmap version 2.16.0 (Gu, 2022; Gu et al., 2016).
In order to elucidate whether all the genes altered by Ocoxin were involved in any particular cellular pathway, several analyses were performed by using various softwares based on different databases. First, a systematic analysis of gene functions altered by Ocoxin was performed in each cancer model according to the Kyoto Encyclopedia of Genes and Genomes (KEGG) (Kanehisa and Sato, 2020; Kanehisa and Yoko, 2024). KEGG comprises three databases (PATHWAY, GENES and LIGAND) and enables the analysis of gene functions, linking genomic and functional information. Second, a functional protein association network analysis was performed based on the genes altered by Ocoxin with the STRING software (version 12.0) (Szklarczyk et al., 2023; Snel et al., 2000), which obtains data from Biocarta, BioCyc, GeneOntology, KEGGa and Reactome dabatases in order to predict physical and functional interactions between proteins. Finally, a Venn diagram (Oliveros, 2007) was utilized so as to find whether there were any common altered genes between the four cancer models.
Afterwards, the expression level of the genes which were in common in all the four tumors was measured by quantitative reverse transcription polymerase chain reaction (RT-qPCR). Specifically the analyzed genes were: Adrenomedullin 2 (ADM2), ChaC glutathione specific gamma-glutamylcyclotransferase 1 (CHAC1), Extra spindle pole bodies like 1, separase (ESPL1), Heme oxygenase 1 (HMOX1), Kinesin family member 20A (KIF20A), Metallothionein 1F (MT1F), NAD(P)H quinone dehydrogenase 1 (NQO1), RAD54 like (RAD54L), Sestrin 2 (SESN2), Solute carrier family 30 member 1 (SLC30A1), Solute carrier family 39 member 10 (SLC39A10), Tribbles pseudokinase 3 (TRIB3) and Thioredoxin reductase 1 (TXNRD1). To do so, cancer cells were treated with Ocoxin for 48 h as previously described and the RNA was again extracted and its quality evaluated as previously described in Section 2.3. Subsequently, 2 μg of mRNA were retrotranscribed into cDNA using the iScript cDNA Synthesis Kit (Bio-Rad, Hercules, CA, United States) and the RT-qPCR was performed using the Itaq Universal SYBR Green Supermix (Bio-Rad, Hercules, CA, United States) on the CFX96 Real-Time System (Bio-Rad, Hercules, CA, United States) and the primers shown on Table 1 (Invitrogen, Waltham, MA, United States). Finally, the relative expression of each gene was standarized to the internal control gene actin beta (ACTB).
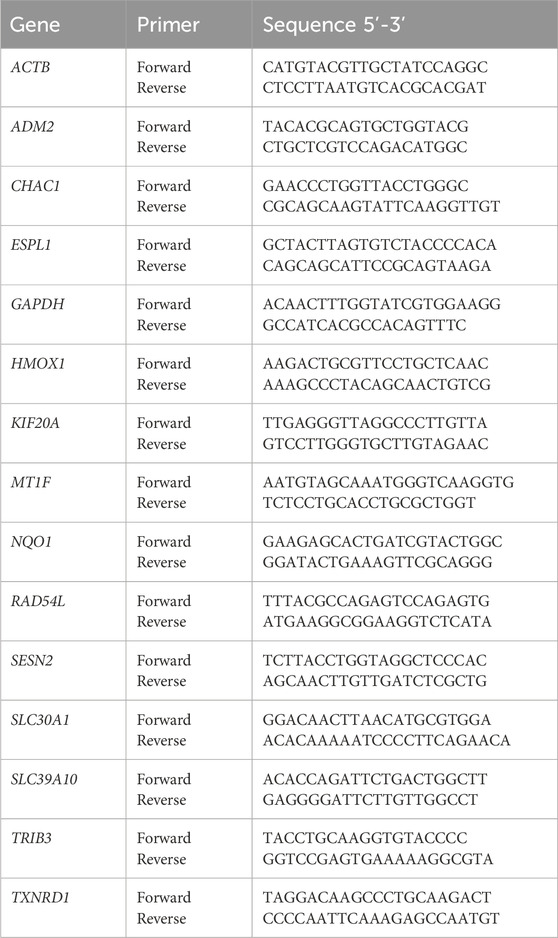
Table 1. Primers used for the RT-qPCR.
Ocoxin exhibited a dose-dependent cell viability reduction in all analyzed cell lines after 48 h of treatment. While PRAD cells showed to be quite resistant to Ocoxin up to the 1:50 dilution COAD, PAAD and TNBC were already affected by the natural supplement with the 1:1000 dilution (Figure 1). Approximately, IC50 was reached with the 1:700 dilution in TNBC and COAD cells, with the 1:500 dilution in PAAD cells and with the 1:50 dilution in PRAD cells (Figure 1).
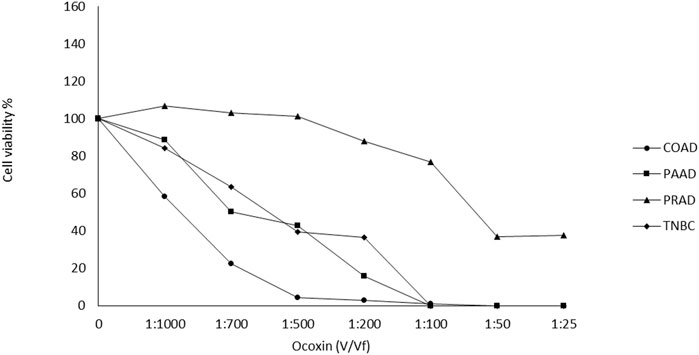
Figure 1. Effect of Ocoxin on the viability of tumor cells. Cancer cells were treated with different dilutions of Ocoxin for 48 h and cell viability was measured with the Cell Counting kit-8.
In order to study whether the effect of Ocoxin on the viability of tumor cells is mediated by genomic alterations, an RNAseq analysis was performed. As shown on Table 2, Ocoxin deregulated the expression of many genes in all the tested cancer cell lines. The number of altered genes was remarkably higher in PAAD cells and in TNBC cells compared to the others. Notably, TNBC exhibited the highest count of deregulated genes after the treatment with Ocoxin, with a total of 3,438 genes altered, followed by PAAD, which showed 1589 altered genes. On the contrary, COAD and PRAD only showed 511 and 594 deregulated genes respectively. Besides that, in COAD, PAAD and TNBC, approximately 50% of the altered genes were upregulated and the other 50% were downregulated, while in PRAD 73% of the deregulated genes were overexpressed.
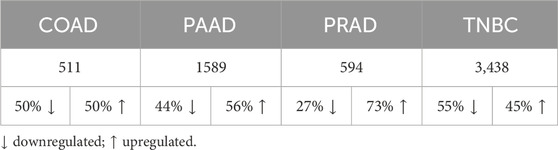
Table 2. Number of genes altered by Ocoxin in each cancer type.
Afterwards, the differentially expressed gene list was introduced in the KEGG mapper software to determine the KEGG pathways in which all the altered genes are implicated. Later, those pathways were classified according to the percentage of altered genes that belonged to that KEGG pathway versus the total number of deregulated genes in that cancer. As shown on Table 3 the majority of the differentially expressed genes were found to be involved in pathways related to metabolism, cancer and neurodegeneration in all the analyzed cancer types. However, in COAD and PRAD the cell cycle was one of the principal altered pathways, which was not present admist the most deregulated KEGG pathways in PAAD and TNBC.
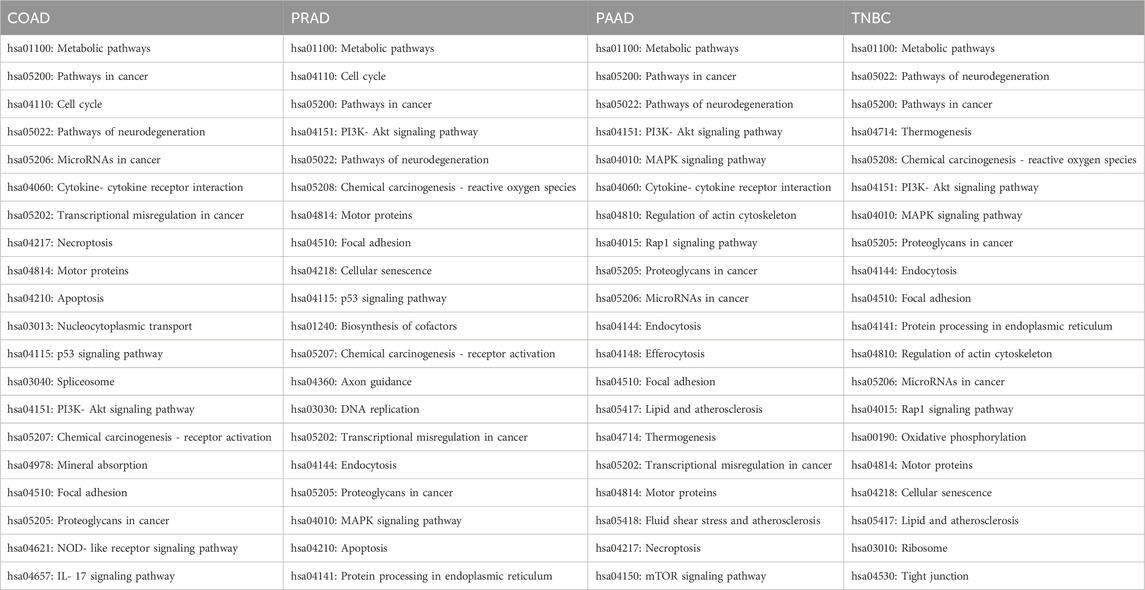
Table 3. Main KEGG pathways of each cancer in which all the genes altered by Ocoxin are implicated.
In addition to the aforementioned pathways, PAAD, PRAD and TNBC exhibited differentially expressed genes involved in the MAPK signaling pathway and PI3K-Akt signaling pathways; however, in COAD those pathways were relegated. Moreover, all the 4 cell lines showed alterations in motor proteins, in cancer-related proteoglycans and in genes involved in the focal adhesion and also pathways of miRNAs were found to be deregulated in COAD, PAAD, and TNBC. In turn, many other pathways were, as well, altered in two of the 4 cell lines like for instance the protein processing in endoplasmic reticulum (ER), which was altered in breast and PRAD, and also the p53 signaling pathway and the transcriptional misregulation in cancer in prostate and COAD.
Since the principal pathways altered by Ocoxin were involved in metabolic processes, we compared whether COAD, PAAD, PRAD and TNBC shared the same altered genes related to cell metabolism through a Venn diagram. Figure 2 shows that Ocoxin deregulated three genes in common in all the four cancers: CHAC1, HMOX1 and NQO1. Moreover, it was remarkable that PAAD and TNBC shared 67 genes (14%) involved in metabolic pathways, while the rest of paired comparisons shared less than 20 except for PRAD and TNBC, that shared 27 genes (6%). Thus, a more in-depth analysis was performed about those 67 metabolic genes by using the STRING software considering all the genes gathered in the “hsa01100: metabolic pathway” KEGG identifier. The network present in Figure 3 showed that all the genes were linked between them. However, five specific metabolic processes could be identified in the protein-protein interaction network: protein lysine methyltransferases methylate histone lysines, the citric acid cycle and respiratory electron transport and metabolism of nucleotides, lipids and carbohydrates.
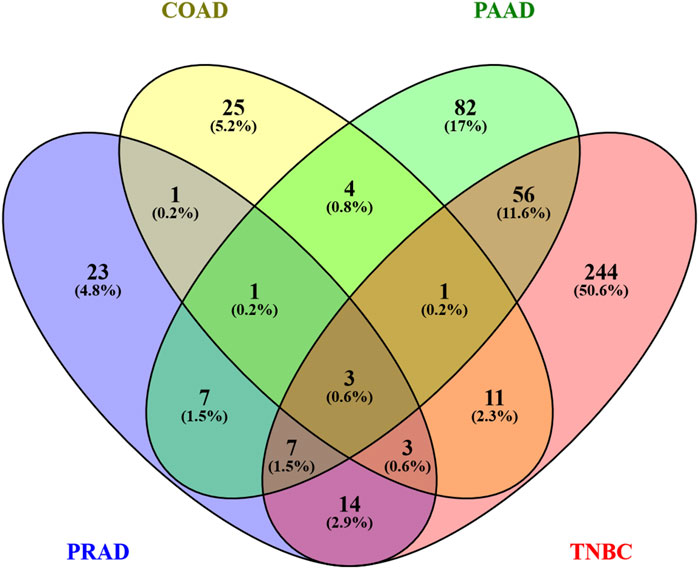
Figure 2. Venn diagram of genes altered by Ocoxin in COAD, PAAD, PRAD, and TNBC that were involved in metabolic pathways. All the genes related to metabolic processes according to the KEGG mapper were included in a Venn diagram in order to compare similarities between the four cancers.
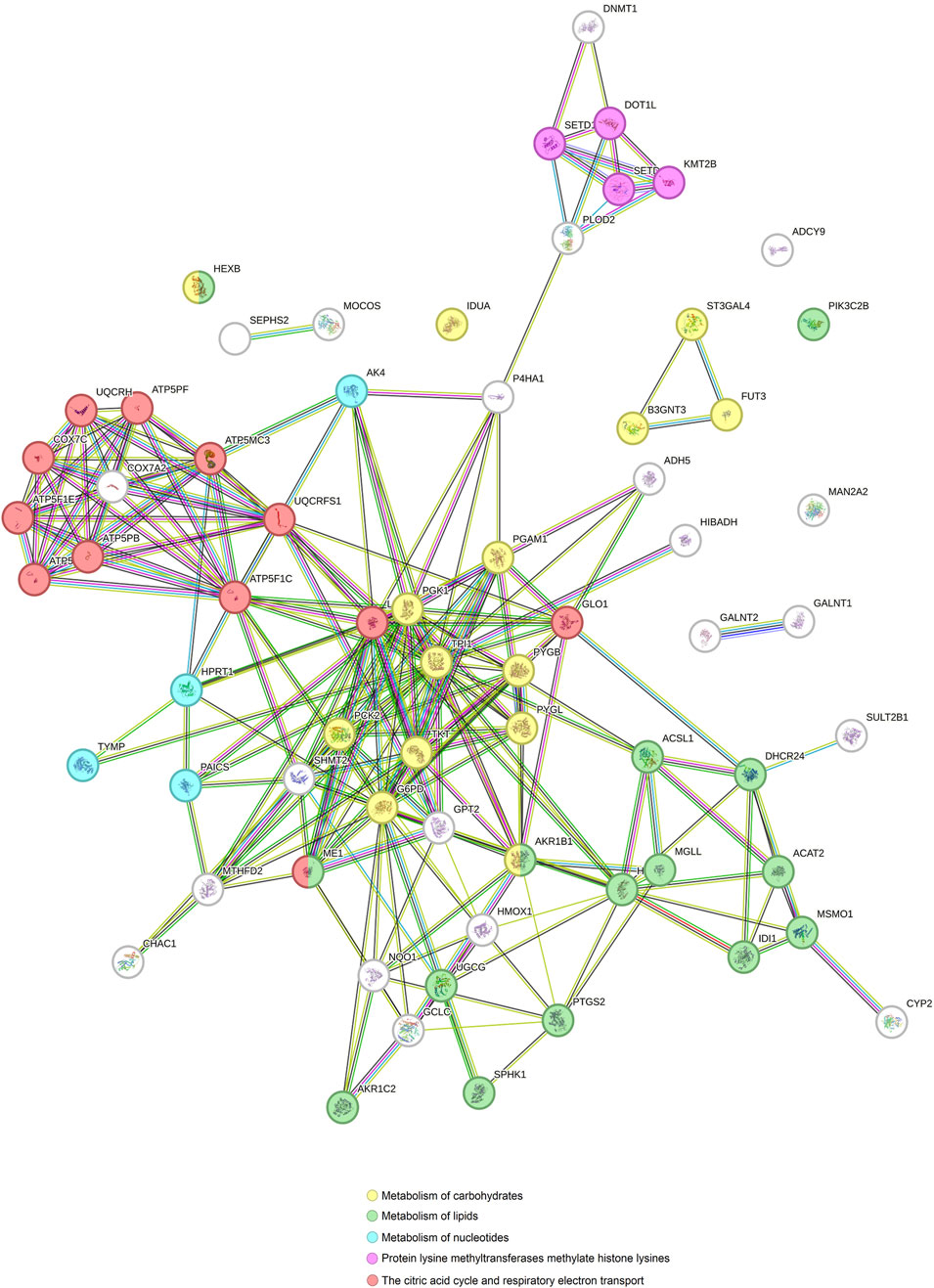
Figure 3. Protein-protein interaction network of genes involved in metabolic processes altered by Ocoxin in common in PAAD and TNBC. All the genes related to metabolic processes altered by Ocoxin in TNBC and PAAD were analyzed through the STRING software in order to analyze the relation between them.
Bearing in mind that the four analyzed cancers showed similar altered pathways with the KEGG software, a Venn diagram was utilized in order to determine whether any of the altered genes are shared among the different cancers. As Figure 4 shows, paired comparisons of the genes deregulated by Ocoxin revealed that most of the cancers shared around 5% of the altered genes with each other, while PAAD and TNBC shared 14% of them. Furthermore, 13 genes were found to be altered in common in the four cancers (Figure 4): ADM2, CHAC1, ESPL1, HMOX1, KIF20A, MT1F, NQO1, RAD54L, SESN2, SLC30A1, SLC39A10, TRIB3 and TXNRD1 (Table 4). Moreover, as shown in the heatmap of Figures 5, 11 of those genes were equally deregulated in all the samples, that is, nine genes (ADM2, CHAC1, HMOX1, MTF1, NQO1, SESN2, SLC30A1, TRIB3 and TXNRD1) were upregulated and 2 (KIF20A and SLC39A10) downregulated. On the contrary, two more genes (ESPL1 and RAD54L) showed to be upregulated PRAD and TNBC, and downregulated in COAD and PRAD (Figure 5).
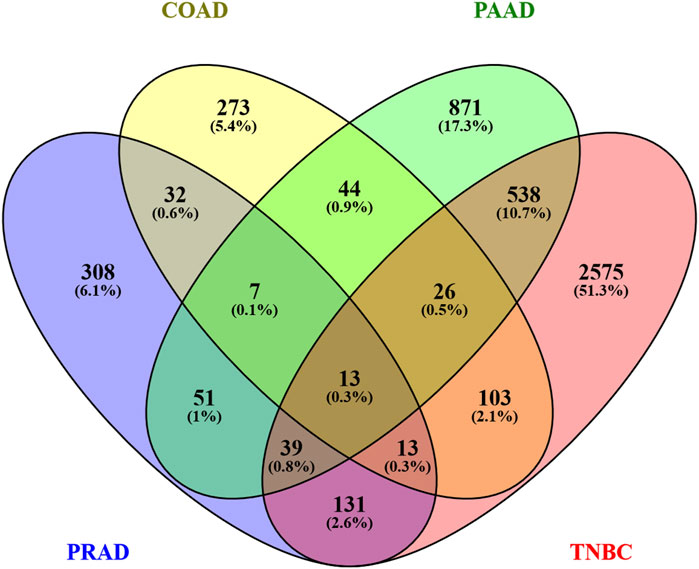
Figure 4. Venn diagram of all the genes altered by Ocoxin in COAD, PAAD, PRAD and TNBC. All the list genes deregulated by Ocoxin were gathered in a Venn diagram.
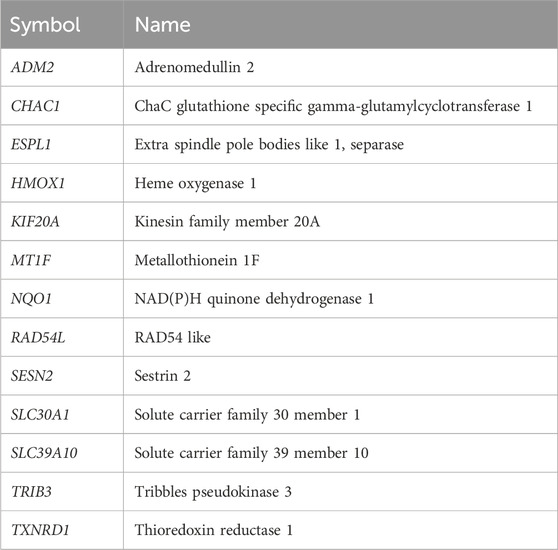
Table 4. Genes altered in common by Ocoxin in COAD, PAAD, PRAD and TNBC.
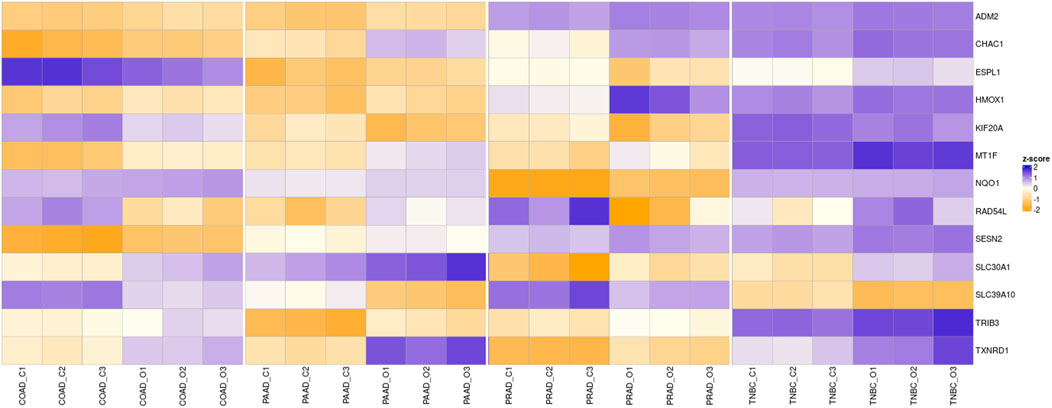
Figure 5. Overview of the RNAseq. The heatmap shows the Z-score of the genes commonly altered by Ocoxin in all the four analyzed cancers. The compound upregulated ADM2, CHAC1, HMOX1, MTF1, NQO1, SESN2, SLC30A1, TRIB3 and TXNRD1 and downregulated KIF20A and SLC39A10 in all the cancers compared to the untreated samples. ESPL1 and RAD54L were upregulated in PAAD and TNBC and downregulated in PRAD and COAD. Three samples were analyzed for each treatment condition and cancer (C: Control, O: Ocoxin).
Later, the expression level of those 13 genes deregulated by Ocoxin in the four cancers was validated through RT-qPCR. As shown in Figure 6, ADM2, CHAC1, HMOX1, MT1F, NQO1, SESN2, SLC30A1, TRIB3, and TXNRD1 were all upregulated in all the cancers while KIF20A and SLC39A10 were downregulated. Conversely, ESPL1 and RAD54L showed a different alteration pattern among the tumors being both upregulated in PAAD and TNBC and downregulated in COAD and PRAD.
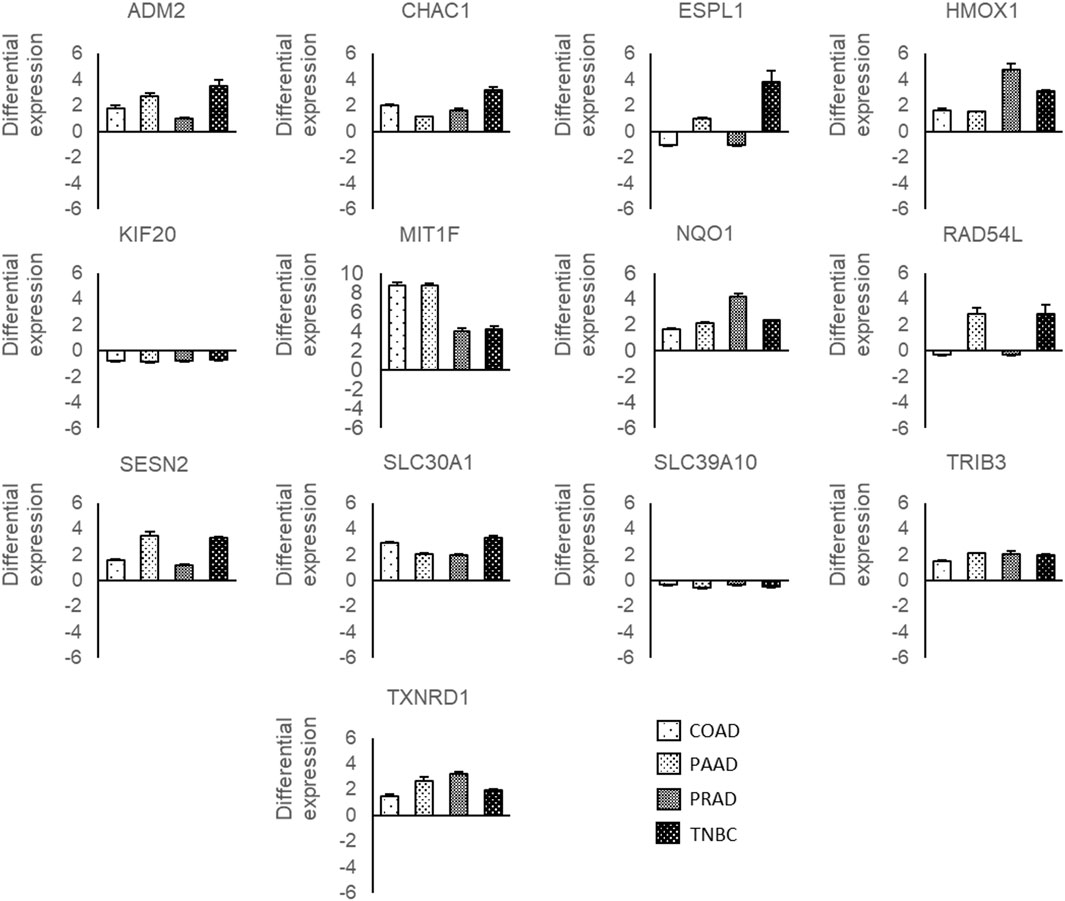
Figure 6. RT-qPCR of genes deregulated by Ocoxin in common in COAD, PAAD, PRAD and TNBC. Cancer cells were treated with the chosen dose of Ocoxin for 48 h (1:700 for COAD, 1:500 for PAAD, 1:50 for PRAD and 1:700 for TNBC), total mRNA was extracted and a RT-qPCR was carried out. Data were normalized to the actin beta expression of each tumor.
Afterwards, to further understand what the common genes altered by Ocoxin in TNBC, PAAD, PRAD and COAD are involved in, a protein-protein interaction analysis was performed through STRING. Figure 7 shows that the 13 proteins can be classified in three independent groups which are related to four protein biological functions. The first group is comprised of the proteins, TXNRD1, HMOX1 and NQO1, which are involved in the detoxification of Reactive Oxygen Species (ROS) and are related to MT1F, SLC30A1 and SCL39A10, proteins involved in Zinc homeostasis. The second group, constituted by ADM2, TRIB3, CHAC1 and SESN2, is involved in the unfolded protein response (UPR) and finally the third group contains KIF20A, RAD54L and ESPL1, which are involved in the cell cycle.
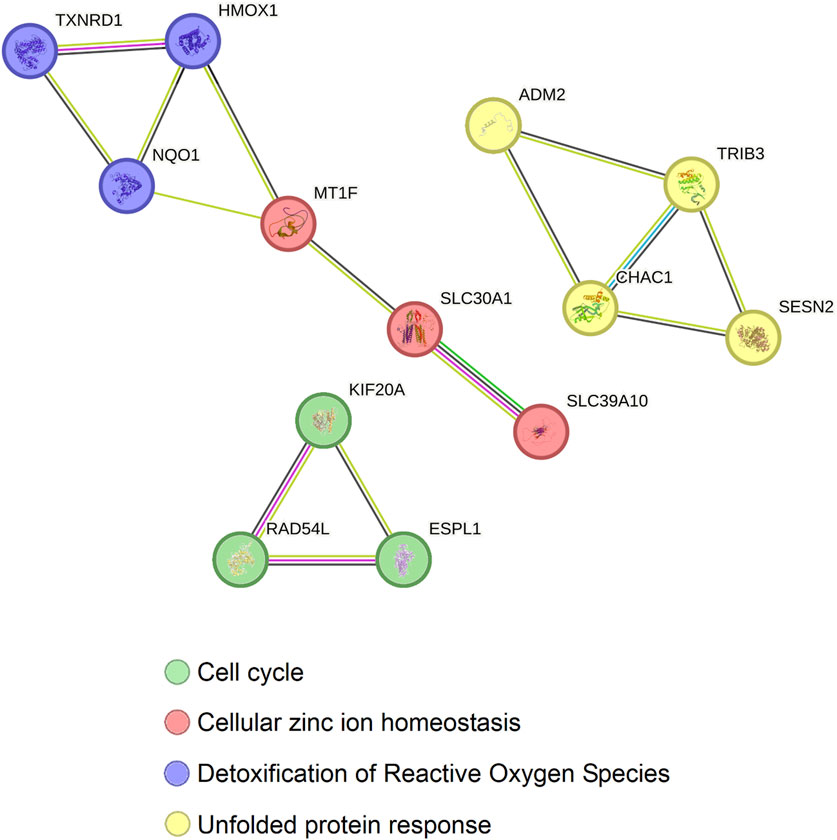
Figure 7. Protein-protein interaction between the 13 genes commonly altered by Ocoxin in COAD, PAAD, PRAD, and TNBC. All the genes altered by Ocoxin in common in the four cancers were analyzed through the STRING software in order to analyze the relation between them. Gene interactions were classified in three independent groups with four biological functions.
Afterwards, in order to examine the potential correlation between those proteins, we increased the number of predicted nodes and observed that all the proteins show to be correlated except for ESPL1, KIF20A and RAD54L through nine new nodes (Figure 8). As in the previous results, the proteins related to zinc homeostasis (MT1F, SLC30A1 and SLC39A10) together with more metallothioneins (MT1B, MT1E, MT1H, MT1X and MT2A) showed to be interconnected with the proteins involved in detoxification of ROS and, likewise, those molecules were linked to the proteins of the genes that take part in the UPR (ADM2, CHAC1, TRIB3 and SESN2) via the activating transcription factor 4 (ATF4), the glutation-S-transferases A1 and A2 (GSTA1 and GSTA2) and the sulfiredoxin 1 (SRXN1) (Figure 8).
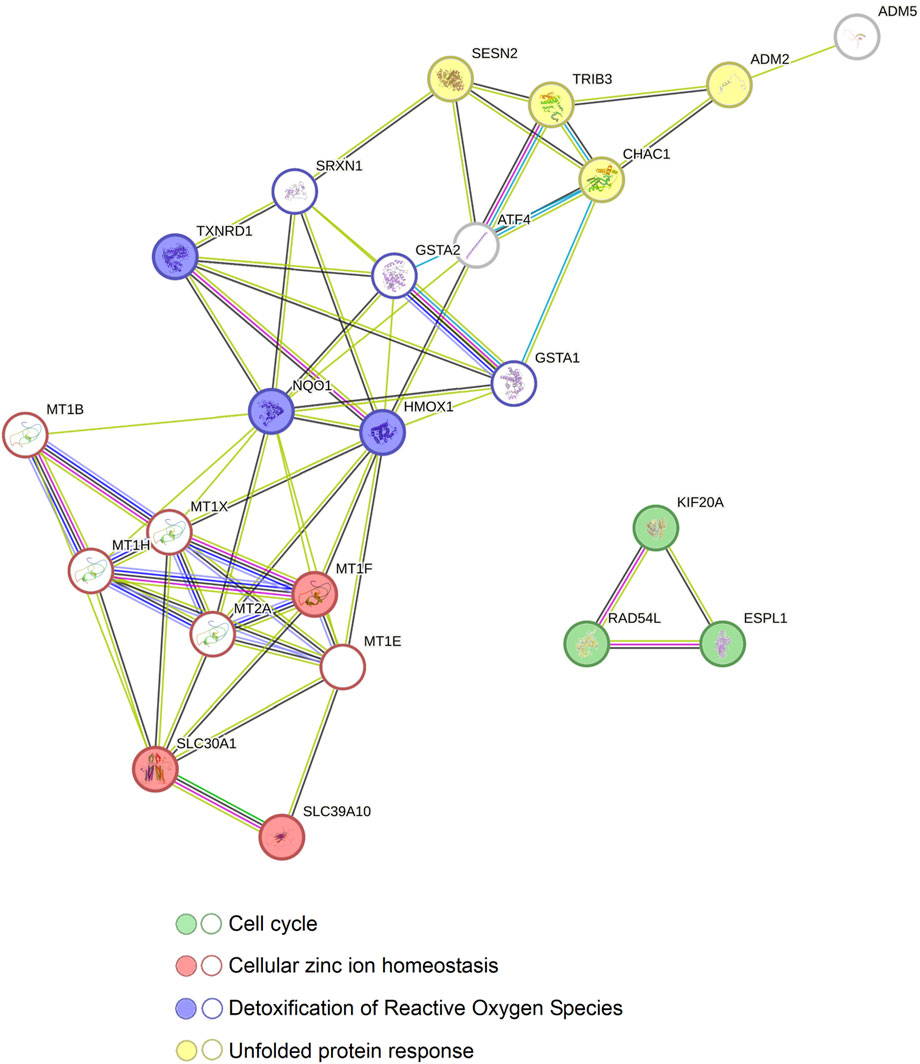
Figure 8. Relation between the protein-protein interaction networks of the genes altered by Ocoxin in common in COAD, PAAD, PRAD, and TNBC. The STRING software was used to reveal the connection between all the genes altered by Ocoxin in common in all the cancers. The 13 genes altered by Ocoxin in common in all the tumors (circles with colored background) were interconnected through nine new nodes (circles without a colored background) and classified in four biological functions.
Cancer is one of the most worrisome pathologies in the world. Nowadays, due to the prevention schedules, advances in oncological surgery and the identification of new therapeutic alternatives have permitted to treat, prolong and improve the life quality of patients suffering from many cancers. However, despite the progress, many of them experience recurrence and metastasis and although they can be treated with radiotherapy or chemotherapy, the adverse effects arising from those treatments considerably damage their quality of life. Thus, the latest research is focused on the identification of new therapeutic alternatives that not only treat cancer but also extend life expectancy and improve the wellbeing of cancer patients by reducing the undesired effects caused by the treatments. In this scenario, there is growing interest in the application of bioactive compounds of natural origin, such as, EGCG, curcumin, resveratrol, quercetin and many others, to treat different kinds of cancers and to reduce the non-desirable side effects produced by chemotherapy (Naujokat and McKee, 2021; Ding et al., 2020; Rana et al., 2021; Vladu et al., 2022; Maleki Dana et al., 2022).
Ocoxin is a natural mixture composed of plant extracts, amino acids, vitamins and minerals known for its established antioxidant, anti-inflammatory and immunoregulatory properties, which has further demonstrated to exert antitumor effects in several cancer models in vitro and in vivo by impairing tumor development at different levels. On the one hand, Ocoxin has reduced cell viability in vitro by decreasing cell proliferation and inducing apoptosis and cell cycle arrest in many cancers. On the other hand, the compound has increased the cytotoxic effect of first-line chemotherapeutic drugs while reverting the resistance to chemotherapy caused by CAFs (Hernandez-Unzueta et al., 2017; Benedicto et al., 2021; Hernandez-Unzueta et al., 2019). Besides, Ocoxin has been used in clinics and showed an improvement in the quality of life of patients with hepatocellular carcinoma, lung cancer, PRAD, PAAD, cervical cancer and endometrial adenocarcinoma, among others, while increasing survival in some patients (Al-Mahtab et al., 2015; Kaidarova et al., 2019; Ruiz Lorente et al., 2020; Lima-Pérez et al., 2024; García-Perdomo et al., 2021). Therefore, in the present work, we aimed to deepen into the fundamental mechanisms through which Ocoxin exerts its antitumoral activity against cancer, as well as to identify whether there are any commonalities among the various types of cancer that may offer insights into the functionality of the nutritional supplement.
In relation to that, previous works have described the role of natural products acting as multi-targeted elements to alter different signaling pathways by regulating gene expression or targeting proteins and their secretion (Luesch and Paavilainen, 2020; Irshad and Husain, 2021; Nasir et al., 2020). Considering that cancer is a dynamic disease and that tumor cells possess distinct molecular signatures and sensitivity to therapies, the capacity of bioactive compounds to target cancer through several mechanisms at once needs to be explored since those effects might be an interesting approach to personalized therapies. Therefore, once tumor and patient’s characteristics are defined, instead of using established first-line treatments, those could be combined with natural products, thereby increasing the potential for a successful treatment.
According to our results, the mixture altered several genes in each type of cancer. Notably, TNBC and PAAC showed a substantially higher number of significantly altered genes, 3,438 and 1589 respectively, from which 12% were in common. Contrarily, in PRAD and COAD the number of deregulated genes was around 500 in each cancer. Besides, taking into consideration all the results, Ocoxin deregulated the expression of 13 genes in common in the analyzed tumors, which can be classified in three principal independent groups based on four functional protein association networks. The first group is composed of proteins related to the UPR (ADM2, CHAC1, SESN2, TRIB3). The second group gathers proteins involved in two biological processes such as, zinc homeostasis (MT1F, SLC30A1, SLC39A10) and detoxification of ROS (HMOX1, NQO1, TXNRD1). These molecules can also be englobed as part of the nuclear factor erythroid 2-related factor 2 (NRF2) pathway, which is responsible for the defense against oxidative stress and the maintenance of stable conditions. Finally, the third group contains proteins related to the cell cycle (ESPL1, KIF20, RAD54L). In this context, the upregulation of the genes encoding for the proteins of the first and second group that deal with cell homeostasis could have led to the production of misfolded proteins that are not correctly eliminated by the ER-associated degradation (ERAD) (Smith et al., 2011; Travers et al., 2000), causing ER stress. In fact, some natural bioactive compounds are known to induce or reduce ER stress. Actually, Xue et al. reported that morusin, a flavonoid obtained from white mulberry, induces ER stress on human epithelial ovarian cancer cells by upregulating the heat shock protein family A member 5 (HSPA5), the DNA damage inducible transcript 3 (DDIT3 or CHOP), the endoplasmic reticulum to nucleus signaling 1 (ERN1) and the α subunit of the translation initiation factor 2 (eIF2α) driving cells to a paraptosis-like cell death (Xue et al., 2018) and Zhang et al. observed that curcumin boosts ER stress-associated apoptosis on human papillary thyroid carcinoma cells (Zhang, L. et al., 2018).
It is known that cells, in order to restore the proteostasis, activate a mechanism named the UPR, whereby modulation of the gene expression occurs and protein synthesis becomes diminished (Harding et al., 2000; Ron and Walter, 2007). In this scenario, cells need to make a decision: to continue living or to die. This choice is controlled by eIF2α, whose activation reduces the global protein synthesis and, at the same time, induces the activating transcription factor 4 (ATF4) (Harding et al., 2003). These genes are central players of the integrated stress response (ISR), an adaptive response by which cells have the ability to reprogram gene expression in order to provide an optimized response to deal with stressful conditions. Despite the fact that the ISR is mainly a homeostatic pro-survival mechanism, cell response will depend on the nature and intensity of the driving stimuli, which, in case of prolonged and severe signals, will drive cells to death (Pakos-Zebrucka et al., 2016). In fact, this signaling network could be useful to face cancer as it is known to contribute to drug sensitivity and to promote the expression of immunotherapeutic targets (Obiedat et al., 2020; Lines et al., 2023; Tian et al., 2021). Actually, over the last years ISR modulating therapies have attracted significant attention to such an extent that diverse ongoing clinical trials target different levels of this pathway (Lines et al., 2023). During the ISR, various kinases sense different stimuli, such as nutrient deprivation, heme deficiency, DNA damage, oxidative stress or ER stress and converge on the phosphorylation of eIF2α leading to the induction of the ATF4 (Lines et al., 2023; Tian et al., 2021). In this work, the treatment with Ocoxin only upregulated ATF4 in PAAD. However, ATF3, ATF5, PMAIP1, DDIT4 and CHOP, which are downstream genes of ATF4, were upregulated by Ocoxin in at least one of the studied cancers, while TRIB3 was overexpressed in all of them. Furthermore, despite the fact that ATF4 itself was not deregulated by Ocoxin in all the analyzed cancer models, it arises as a central protein in the correlation among the genes altered by the compound (except for KIF20A, RAD54L and ESPL1) in common in COAD, PAAD, PRAD and TNBC. In fact, most of the genes upregulated by Ocoxin, such as ADM2, CHAC1, HMOX1, MTF1, NQO1, SESN2, TRIB3 or TXNRD1, are related to the ER stress either directly or indirectly. On the one hand, ADM2, CHAC1, SESN2 and TRIB3 are stress sensors of UPR controlled by the ATF4 gene under ISR (Crawford et al., 2015; Kovaleva et al., 2016; Garaeva et al., 2016; Kalinin et al., 2023) and on the other hand, HMOX, MTF1, NQO1 and TXNRD1 are involved in ROS detoxification, whose imbalance causes ER stress (Shi et al., 2023; Dunn et al., 2014; Li, H. et al., 2021; Rodríguez-Menéndez et al., 2018; Scuto et al., 2022). Furthermore, MTF1 encodes for a zinc dependent transcription factor that responds to changes produced in the intracellular heavy-metal homeostasis. Briefly, MTF1 regulates cellular processes by binding zinc to metallothioneins (MTs) or through the transcription of metal transporters of the ZRT/IRT-like protein (ZIP) family and the zinc transporter (ZnT) family (Eide, 2006; Lichten et al., 2011; Lichten and Cousins, 2009). While ZIP family carriers import zinc from the extracellular space or subcellular organelles to the cytoplasm, ZnT family components export the metal from the cytoplasm to the extracellular milieu or organelles (Sun, C. et al., 2024; Cotrim et al., 2019). At this point, it needs to be mentioned that Ocoxin contains high levels of zinc, which could have altered the expression of genes that participate in its homeostasis. In particular, those genes deregulated by Ocoxin were SLC30A1 and SLC39A10 (also known as ZNT1 and ZIP10), which enable zinc discharge and acquisition respectively. In this regard, Ocoxin upregulated the expression of SLC30A1, whereas downregulated that of SLC39A10, resulting in an increase of export to the extracellular compartment and a reduction of intracellular zinc accumulation (Fukada and Kambe, 2011; Palmiter and Findley, 1995). Therefore, cancer cells may activate MTF1 in the presence of high concentrations of this metal, causing an underexpression of SLC39A10 and an overexpression of SLC30A1 in order to recover zinc homeostasis. In fact, this metal is an essential component for the correct functioning of the ER since any homeostatic disruption can originate ER stress and subsequent UPR activation, which, as previously described, are related to the ISR pathway (Homma et al., 2013; Nguyen et al., 2013; Kambe et al., 2017). Therefore, we deem that Ocoxin drives tumor cells to a stressful environment through the ISR. Bearing in mind the intertumoral and intratumoral heterogeneity, and given the implication of the ISR in cellular adaptation and tumor development, modulating its activity could be a formidable treatment for many cancers, especially in combination with other therapies. Due to the multiple cellular mechanisms controlled by the ISR, the manipulation of this pathway could reach several populations of cells with distinct genetic signatures within the same tumor, enabling the overcoming of barriers such as treatment resistance or relapse, which represents a major impediment in medical oncology.
Anyway, it is crucial to consider that our results correspond to cells that had been treated with Ocoxin for 48 h, a period that might not be sufficient for the cells to determine their fate. Probably, as described before, if the stress caused by Ocoxin persisted, cells would engage cell death signaling pathways. This is supported by unpublished data where we observed that the mixture led to an increase in the percentage of dead cells after 72 h, thereby confirming that cells ultimately succumb to death after sustained stress conditions. Nevertheless, all the analyzed tumors exhibited either unchanged or downregulated caspases, indicating that cells may be undergoing a caspase-independent form of programmed cell death or that a duration exceeding 48 h is required for the activation of apoptotic caspases. Here, we show that, among other processes, Ocoxin deregulated the expression of multiple genes associated with autophagy, necroptosis and especially ferroptosis, a type of programmed cell death characterized by the accumulation of free iron within cells and by lipid peroxidation. Due to its clinical importance, ferroptosis has gained attention over the last years as an apoptosis-independent programmed cell death. This mechanism is strongly associated with metabolic processes, such as, carbohydrate metabolism, lipid metabolism and amino-acid metabolism (Ye et al., 2024) as well as iron and selenium metabolism (Conrad and Proneth, 2020) among others. In this regard, keeping in mind that cell metabolism is a crucial process to sustain cell growth and self-renewal, it leads to the fact that metabolic activities and the metabolites produced in the tumor could have an influence on ferroptosis (Jiang et al., 2024). Interestingly, Ocoxin caused an overexpression of HMOX1, a critical mediator in ferroptosis that promotes cell death in the presence of oxidative stress and high iron concentrations inside the cells (Chiang et al., 2018). Moreover, the nutritional supplement also deregulated the expression of NQO1, a gene responsible for redox control that is also involved in ferroptosis (Zhang, L. et al., 2018; Wang W. et al., 2023). Actually, both HMOX1 and NQO1 are downstream genes of NRF2, which play a central role in the NRF2 pathway defending cells against ROS. In consonance, Liu et al. (2022) described that Vitamin C, which is one of the components of Ocoxin, sensitized pancreatic cancer cells to erastin-induced ferroptosis by activating the AMPK/NRF2/HMOX1 pathway (Liu, Y. et al., 2022). In line with this, Wang et al. reported that eriodictyol, a flavonoid obtained from certain plants, regulated ferroptosis through the NRF2/HMOX1/NQO1 signaling pathway in ovarian cancer (Wang X. et al., 2023). Thus, the overexpression of HMOX1 and NQO1 observed in cancer cells treated with Ocoxin could have accounted for the subsequent mortality of those cells when exposed to the compound. Besides, it is important to bear in mind that HMOX1 and NQO1 are also two of the genes related to the main pathway deregulated by Ocoxin in all the four analyzed cancers: cell metabolism. Interestingly, both HMOX1 and NQO1 are downstream genes of ATF4 and, therefore, they are implicated in the ISR pathway mentioned before. Wang W. et al. (2023) reported that a flavonoid derived from licorice increased HMOX1 and decreased metabolic glutathione peroxidase 4 (GPX4), which reduces lipid peroxidation, leading to ferroptosis (Wang X. et al., 2023; Koppula et al., 2021; Zhao et al., 2019; Li, S. et al., 2010). Additionally, Yang and colleagues (2021) described that the suppression of the KIF20A/NUAK1/NRF2/GPX4 signaling pathway induces ferroptosis and increases the sensitivity of COAD to chemotherapy. Interestingly, although the expression of NUAK1, NRF2 and GPX4 was not altered by Ocoxin, the supplement downregulated the expression of KIF20A in the four tumors (Yang et al., 2021). Nevertheless, a deregulation of SLC7A11 was detected in PRAD, PAAD and COAD, which is a downstream gene of NRF2 and the ISR (Koppula et al., 2021). This gene encodes for a cysteine/glutamate antitransporter system composed of two subunits (SLC7A11 and SLC3A2) that exchange extracellular cysteine for intracellular glutamate and regulate ferroptosis through the glutathione/glutathione peroxidase 4 (GSH/GPX4) pathway. Here we observed that Ocoxin upregulated SLC7A11 in three cancers along with SLC3A2 in PAAD, prompting an accumulation of cysteine, which is a component of Ocoxin, and a loss of glutamate inside the cell, impeding the function of GPX4 and, therefore, contributing to ferroptosis (Koppula et al., 2021). Correspondingly, Tang et al. described how certain metabolites obtained from plants used in Chinese medicine altered SLC7A11 and GPX4, among others, which reduced cancer development (Tang et al., 2024). Moreover, Ocoxin overexpressed CHAC1, a gene that also participates in the glutathione degrading metabolism, which curiously was connected to HMOX1 and NQO1 through ATF4 in the protein-protein interaction network analysis. According to several authors, CHAC1 is essential for the maintenance of NRF2 activation and is also involved in ferroptosis (Kreß et al., 2023; Kreß et al., 2023). Actually, He et al. reported that CHAC1 plays a role in hindering tumor development and enhancing the efficacy of docetaxel in prostate cancer by triggering ferroptosis and ER stress (He et al., 2021) and additionally, Chen et al. reported that this gene facilitates ferroptosis as well as necroptosis in breast cancer (Chen, M. et al., 2017). Furthermore, this is corroborated by Liu et al., who described the role of CHAC1 inducing ferroptosis through the glutathione metabolism in retinal pigment epithelial cells (Liu et al., 2023). Taking that into consideration, Ocoxin could have decreased intracellular glutathione concentration through the overexpression of CHAC1 and the subsequent glutathione degradation, driving cells to ferroptosis. Thus, we conclude that Ocoxin induces ferroptosis in tumor cells through several pathways, which could be beneficial for patients that do not respond to chemotherapy. According to several authors, apoptosis-resistant cancer cells have showed to be susceptible to ferroptosis inducers (Hangauer et al., 2017; Tsoi et al., 2018; Viswanathan et al., 2017). In this manner, a greater efficacy of current therapies could be achieved leading to the induction of apoptosis while simultaneously targeting resistant cells via ferroptosis. Moreover, this type of cell death has also been associated with the tumor microenvironment by several authors. On the one hand, Costa da Silva et al. reported that iron uptake by M2-like tumor-associated macrophages drives a differentiation to M1-proinflammatory phenotype which helps to reduce tumor growth (Costa da Silva et al., 2017). On the other hand, Wang et al. described that immunotherapy-activated CD8+ T cells enhance tumor ferroptosis by inhibiting the expression of molecules implicated in the glutamate–cystine antiporter system through interferon gamma (IFNγ) release, which causes lipid peroxidation and leads tumor to ferroptosis (Wang et al., 2019). In this regard, we previously mentioned that Ocoxin has an antitumor effect in different cells of the tumor stroma. However, additional analyses need to be carried out so as to identify the possible implication of the compound in the induction of ferroptosis through the tumor microenvironment. Nonetheless, the use of Ocoxin as an adjuvant agent could enhance first-line therapies by targeting the different populations of cancer cells and the tumor microenvironment, thereby increasing the susceptibility of therapy-persistent cancer cells to common therapies and preventing tumor recurrence and metastasis.
Nevertheless, those are not the unique metabolic genes targeted by Ocoxin. As previously mentioned, TNBC and PAAC shared a considerable number of altered genes. In particular, considering only those genes involved in metabolic pathways, those two cancers showed 14% of altered genes in common, while at the same time those were not altered in PRAD and COAD. In this regard, it is important to bear in mind that the analyzed PAAD and TNBC cells come from primary tumors, while those of PRAD and COAD were isolated from metastatic sites. A deeper analysis about the specific metabolic pathways where the genes altered by Ocoxin are involved, showed that in the primary tumors (PAAD and TNBC) the compound targeted genes from the carbohydrate, nucleotide and lipid metabolism, protein lysine methyltransferases (PKMTs) methylate histone lysine and the citric acid (TCA) cycle and respiratory electron transport. Specifically, regarding the respiratory electron transport, Ocoxin downregulated the expression of the mitochondrial oxidative phosphorylation (OXPHOS) genes ATP5F1C, ATP5MC3, ATP5F1E, ATP5PB, ATP5PF and ATP5MG, which could have been also driven by the ISR. Briefly, the phosphorylation of eIF2α mediated by the ISR stimulates the transcription of CHOP, ATF4 and ATF5 genes that participate in the activation of the mitochondrial UPR (UPRmt) (Zhang, G. et al., 2021; Ogilvie et al., 2005; Boyce et al., 2005) Since both, the ISR and the UPRmt, are two strongly related processes that function together to maintain the mitochondrial metabolism and function, we can assume that Ocoxin altered mitochondrial homeostasis. Interestingly, central players of this pathway, SIRT7 and HSP60 (HSPD1) (Lu et al., 2022), are overexpressed in the primary PAAD and TNBC but not in the metastatic COAD and PRAD. A similar phenomenon was also observed in this study, while the KEGG enrichment analysis indicated that cell cycle is one of the principal pathways altered in the metastatic PRAD and COAD tumors, in PAAD and TNBC the number of genes involved in the cell cycle was not that remarkable. In this context, it is widely described that one of the antitumor mechanisms of Ocoxin is the alteration of the cell cycle (Díaz-Rodríguez et al., 2018; Diaz-Rodriguez et al., 2017; Diaz-Rodriguez et al., 2016; Benedicto et al., 2021; Hernandez-Unzueta et al., 2023; Hernandez-Unzueta et al., 2019; Hernández-García et al., 2015; Pérez-Peña et al., 2019; Márquez et al., 2016). Among the 13 genes commonly altered by the mixture in the four cancer types, three of them, KIF20A, ESPL1 and RAD54L are implicated in the cell cycle progression. Curiously, although KIF20A is downregulated in all tumors, ESPL1 and RAD54L are differentially expressed between primary and metastatic tumors, being upregulated in PAAD and TNBC while downregulated in COAD and PRAD. We previously discussed the relation of KIF20A with ferroptosis; however, this gene mediates the spindle formation during mitosis and its overexpression is related to both, cancer development and a poor overall survival of patients. In fact, KIF20A is overexpressed in all the cancers present in the Gene Expression Profile Interactive Analysis (GEPIA) server (Tang et al., 2017), including COAD, PAAD, PRAD, and TNBC. Interestingly, in this work we demonstrated that Ocoxin downregulated KIF20A in all the analyzed tumors, that is, the compound reverted the expression of this gene. Besides, previous works report that the nutritional mixture also slowed down the cell cycle of tumor cells by causing an arrest on the G2/M phase in breast cancer while in colorectal carcinoma also affected the cell transition from G1 to S phase (Hernández-García et al., 2015; Márquez et al., 2016). Moreover, it is known that the inhibition of KIF20A hinders the growth of several cancer cells (Jin et al., 2023) and Nakamura and colleagues reported that the suppression of KIF20A arrested cells in the G2/M phase in breast cancer, while Zhang et al. reduced the colorectal cancer cell proliferation and migration by drecreasing the cell transition from G1 to S phase through the E2F1 transcription factor in hepatocellular carcinoma (Nakamura et al., 2020; Wang, H. et al., 2020). Considering all this, the reduction of KIF20A expression could be a specific therapeutic strategy to treat cancer, which proves that the understanding of factors such as the genetic profile of the patient and the mechanism of action of the compound might be a step towards more personalized therapies. In this regard, several authors have reported so far that Ocoxin slows down the cell cycle in different phases depending on the tumor type. Taking this into consideration, we can suggest that ESPL1 and RAD54L, which are differentially expressed between the primary TNBC and PAAC and the metastatic PRAD and COAD, are modulating the cell cycle in various ways. While RAD54L belongs to the DEAD-like helicase superfamily and is involved in the homologous recombination and DNA repair, ESPL1, also known as separase, initiates the final separation of sister chromatids during anaphase. ESPL1, which is typically upregulated in tumor tissue due to the rapid proliferation of cancer cells, is controlled by the PTTG1 regulator of sister chromatid separation, also known as securin, which has the ability to inhibit ESPL1 (Sun, Y. et al., 2009; Zhong et al., 2023). Here we observed that, on the one hand, the treatment with Ocoxin upregulated ESPL1 and downregulated PTTG1 in TNBC and PAAD, and on the other hand, ESPL1 was downregulated in PRAD and COAD, but PTTG1 was not altered. Moreover, while most of the cell cycle genes in common in metastatic tumors, such as, MCM2, MCM3, MCM4, MCM5, MCM7, BUB1, BUB1B and AURKB are downregulated, including CDKN2B, which is downregulated in TNBC and CDKN2C in PAAC, both encoded proteins that function as a cell growth regulator that controls cell cycle G1 progression. Nevertheless, most of the cell cycle genes of the metastatic COAD and PRAD tumors were upregulated, including the abovementioned CDKN2B, showing that Ocoxin regulates the G1 phase of the cell cycle through different routes.
To conclude, we can suggest that Ocoxin shows different mechanisms of action depending on the type and stage of the tumor. In fact, the compound demonstrated a greater sensitivity to primary tumors, where it altered more genes than in metastatic tumors. Nevertheless, the main altered pathway was cell metabolism, with a significant emphasis on the process of ferroptosis among the genes modulated by Ocoxin in common in all the studied cancers. Moreover, it must be mentioned that Ocoxin exerted a varied effect in each cancer by acting on distinct pathways, which, considering tumor heterogeneity, might reflect the multi-target action of the compound. Although the primary process modulated by the supplement was cell metabolism, the effect of Ocoxin in metastatic tumors exerted a substantial effect on the cell cycle. Therefore, we can presume that the antitumor action of Ocoxin is, in part, attributable to mechanisms englobed in the UPR, ISR and metabolic alterations leading to cell death, which may also explain its supportive role in chemotherapy. Modulating those pathways could be a crucial strategy to provide more specific treatments in clinical settings and improve oncological therapies, especially in aggressive or treatment-resistant tumors. However, further studies are needed to delve into the antitumoral mechanism of Ocoxin.
The data presented in the study are deposited in the GEO repository, accession number GSE291188.
Ethical approval was not required for the studies on humans in accordance with the local legislation and institutional requirements because only commercially available established cell lines were used.
IH-U: Data curation, Formal Analysis, Investigation, Methodology, Software, Validation, Visualization, Writing–original draft, Writing–review and editing. UT-G: Investigation, Methodology, Writing–review and editing. AA: Data curation, Formal Analysis, Methodology, Software, Visualization, Writing–review and editing. JM: Data curation, Formal Analysis, Methodology, Software, Visualization, Writing–review and editing. ES: Funding acquisition, Project administration, Writing–review and editing. JM: Conceptualization, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Writing–original draft, Writing–review and editing.
The author(s) declare that financial support was received for the research and/or publication of this article. This research was funded by Catalysis S.L. The funder had no role in the design of the study; in the collection, analysis and interpretation of data; in the writing of the manuscript or in the decision to publish the results.
ES was employed by Catalysis S.L.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abadi, A. J., Mirzaei, S., Mahabady, M. K., Hashemi, F., Zabolian, A., Hashemi, F., et al. (2022). Curcumin and its derivatives in cancer therapy: potentiating antitumor activity of cisplatin and reducing side effects. Phytother. Res. 36, 189–213. doi:10.1002/ptr.7305
Al-Mahtab, M., Akbar, S. M. F., Khan, M. S. I., and Rahman, S. (2015). Increased survival of patients with end-stage hepatocellular carcinoma due to intake of ONCOXIN®, a dietary supplement. Indian J. Cancer 52, 443–446. doi:10.4103/0019-509X.176699
Almatroodi, S. A., Almatroudi, A., Khan, A. A., Alhumaydhi, F. A., Alsahli, M. A., and Rahmani, A. H. (2020). Potential therapeutic targets of epigallocatechin gallate (EGCG), the most abundant catechin in green tea, and its role in the therapy of various types of cancer. Molecules 25, 3146. doi:10.3390/molecules25143146
Benedicto, A., Hernandez-Unzueta, I., Sanz, E., and Márquez, J. (2021). Ocoxin increases the antitumor effect of BRAF inhibition and reduces cancer associated fibroblast-mediated chemoresistance and protumoral activity in metastatic melanoma. Nutrients, 13, 686. doi:10.3390/nu13020686
Boyce, M., Bryant, K. F., Jousse, C., Long, K., Harding, H. P., Scheuner, D., et al. (2005). A selective inhibitor of eIF2alpha dephosphorylation protects cells from ER stress. Science 307, 935–939. doi:10.1126/science.1101902
Broad Institute MIT, and Harvard (2022). Picard. A set of command line tools (in Java) for manipulating high-throughput sequencing (HTS) data and formats such as SAM/BAM/CRAM and VCF. Available at: https://broadinstitute.github.io/picard/.
Chen, M., Wang, S., Hsu, C., Yin, P., Yeh, T., Lee, H., et al. (2017). CHAC1 degradation of glutathione enhances cystine-starvation-induced necroptosis and ferroptosis in human triple negative breast cancer cells via the GCN2-eIF2α-ATF4 pathway. Oncotarget 8, 114588–114602. doi:10.18632/oncotarget.23055
Chen, S. (2023). Ultrafast one-pass FASTQ data preprocessing, quality control, and deduplication using fastp. Imeta 2, e107. doi:10.1002/imt2.107
Chen, S., Zhou, Y., Chen, Y., and Gu, J. (2018). fastp: an ultra-fast all-in-one FASTQ preprocessor, 34, i884–i890. doi:10.1093/bioinformatics/bty560
Chen, Z., Li, Z., Yang, S., Wei, Y., and An, J. (2023). The prospect of Xihuang pill in the treatment of cancers, prospect Xihuang pill Treat. cancers 9, e15490. doi:10.1016/j.heliyon.2023.e15490
Chiang, S., Chen, S., and Chang, L. (2018). A dual role of heme oxygenase-1 in cancer cells. Int. J. Mol. Sci. 20, 39. doi:10.3390/ijms20010039
Choudhari, A. S., Mandave, P. C., Deshpande, M., Ranjekar, P., and Prakash, O. (2020). Phytochemicals in cancer treatment: from preclinical studies to clinical practice. Front. Pharmacol. 10, 1614. doi:10.3389/fphar.2019.01614
Conrad, M., and Proneth, B. (2020). Selenium: tracing another essential element of ferroptotic cell death. Cell. Chem. Biol. 27, 409–419. doi:10.1016/j.chembiol.2020.03.012
Costa da Silva, M., Breckwoldt, M. O., Vinchi, F., Correia, M. P., Stojanovic, A., Thielmann, C. M., et al. (2017). Iron induces anti-tumor activity in tumor-associated macrophages. Front. Immunol. 8, 1479. doi:10.3389/fimmu.2017.01479
Cotrim, C. A., Jarrott, R. J., Martin, J. L., and Drew, D. (2019). A structural overview of the zinc transporters in the cation diffusion facilitator family. Acta Crystallogr. D. Struct. Biol. 75, 357–367. doi:10.1107/S2059798319003814
Crawford, R. R., Prescott, E. T., Sylvester, C. F., Higdon, A. N., Shan, J., Kilberg, M. S., et al. (2015). Human CHAC1 protein degrades glutathione, and mRNA induction is regulated by the transcription factors ATF4 and ATF3 and a bipartite ATF/CRE regulatory element. J. Biol. Chem. 290, 15878–15891. doi:10.1074/jbc.M114.635144
Diaz-Rodriguez, E., El-Mallah, A. M., Sanz, E., and Pandiella, A. (2017). Antitumoral effect of Ocoxin in hepatocellular carcinoma, Antitumoral Eff. Ocoxin Hepatocell. carcinoma 14, 1950–1958. doi:10.3892/ol.2017.6440
Diaz-Rodriguez, E., Hernandez-Garcia, S., Sanz, E., and Pandiella, A. (2016). Antitumoral effect of Ocoxin on acute myeloid leukemia, Antitumoral Eff. Ocoxin acute myeloid leukemia 7, 6231–6242. doi:10.18632/oncotarget.6862
Díaz-Rodríguez, E., Sanz, E., and Pandiella, A. (2018). Antitumoral effect of Ocoxin, a natural compound-containing nutritional supplement, in small cell lung cancer. Int. J. Oncol. 53, 113–123. doi:10.3892/ijo.2018.4373
Ding, S., Xu, S., Fang, J., and Jiang, H. (2020). The protective effect of polyphenols for colorectal cancer. Front. Immunol. 11, 1407. doi:10.3389/fimmu.2020.01407
Dobin, A., Davis, C. A., Schlesinger, F., Drenkow, J., Zaleski, C., Jha, S., et al. (2012). STAR: ultrafast universal RNA-seq aligner. Star. ultrafast Univers. RNA-seq aligner 29, 15–21. doi:10.1093/bioinformatics/bts635
Dunn, L. L., Midwinter, R. G., Ni, J., Hamid, H. A., Parish, C. R., and Stocker, R. (2014). New insights into intracellular locations and functions of heme oxygenase-1. Antioxid. Redox Signal. 20, 1723–1742. doi:10.1089/ars.2013.5675
Eide, D. J. (2006). Zinc transporters and the cellular trafficking of zinc. Biochim. Biophys. Acta 1763, 711–722. doi:10.1016/j.bbamcr.2006.03.005
Esmeeta, A., Adhikary, S., Dharshnaa, V., Swarnamughi, P., Ummul Maqsummiya, Z., Banerjee, A., et al. (2022). Plant-derived bioactive compounds in colon cancer treatment: an updated review. Biomed. Pharmacother. 153, 113384. doi:10.1016/j.biopha.2022.113384
Farghadani, R., and Naidu, R. (2022). Curcumin as an enhancer of therapeutic efficiency of chemotherapy drugs in breast cancer. Int. J. Mol. Sci. 23, 2144. doi:10.3390/ijms23042144
Fukada, T., and Kambe, T. (2011). Molecular and genetic features of zinc transporters in physiology and pathogenesis. Metallomics 3, 662–674. doi:10.1039/c1mt00011j
Fundora-Ramos, M. I., Maden, L. B., Casanova, F. O., Hernandez Cruz, F., Reyes, C. S., Hernandez Gato, A., et al. (2021). Oncoxin-Viusid® may improve quality of life and survival in patients with hormone-refractory prostate cancer undergoing onco-specific treatments. Mol. Clin. Oncol. 14, 5. doi:10.3892/mco.2020.2167
Garaeva, A. A., Kovaleva, I. E., Chumakov, P. M., and Evstafieva, A. G. (2016). Mitochondrial dysfunction induces SESN2 gene expression through Activating Transcription Factor 4. Cell. Cycle 15, 64–71. doi:10.1080/15384101.2015.1120929
García-Perdomo, H. A., Gómez-Ospina, J. C., and Reis, L. O. (2021). Immunonutrition hope? Oral nutritional supplement on cancer treatment. Int. J. Clin. Pract. 75, e14625. doi:10.1111/ijcp.14625
Gu, Z., Eils, R., and Schlesner, M. (2016). Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 32, 2847–2849. doi:10.1093/bioinformatics/btw313
Hangauer, M. J., Viswanathan, V. S., Ryan, M. J., Bole, D., Eaton, J. K., Matov, A., et al. (2017). Drug-tolerant persister cancer cells are vulnerable to GPX4 inhibition, 551, 247–250. doi:10.1038/nature24297
Harding, H. P., Novoa, I., Zhang, Y., Zeng, H., Wek, R., Schapira, M., et al. (2000). Regulated translation initiation controls stress-induced gene expression in mammalian cells. Mol. Cell 6, 1099–1108. doi:10.1016/s1097-2765(00)00108-8
Harding, H. P., Zhang, Y., Zeng, H., Novoa, I., Lu, P. D., Calfon, M., et al. (2003). An integrated stress response regulates amino acid metabolism and resistance to oxidative stress. Mol. Cell 11, 619–633. doi:10.1016/s1097-2765(03)00105-9
Harvie, M. (2014). Nutritional supplements and cancer: potential benefits and proven harms. Am. Soc. Clin. Oncol. Educ. Book 478, e478–e486. doi:10.14694/EdBook_AM.2014.34.e478
He, S., Zhang, M., Ye, Y., Zhuang, J., Ma, X., Song, Y., et al. (2021). ChaC glutathione specific γ-glutamylcyclotransferase 1 inhibits cell viability and increases the sensitivity of prostate cancer cells to docetaxel by inducing endoplasmic reticulum stress and ferroptosis. Exp. Ther. Med. 22, 997. doi:10.3892/etm.2021.10429
Hernández-García, S., González, V., Sanz, E., and Pandiella, A. (2015). Effect of oncoxin oral solution in HER2-overexpressing breast cancer. Nutr. Cancer 67, 1159–1169. doi:10.1080/01635581.2015.1068819
Hernandez-Unzueta, I., Benedicto, A., Olaso, E., Sanz, E., Viera, C., Arteta, B., et al. (2017). Ocoxin oral solution® as a complement to irinotecan chemotherapy in the metastatic progression of colorectal cancer to the liver. Oncol. Lett. 13, 4002–4012. doi:10.3892/ol.2017.6016
Hernandez-Unzueta, I., Benedicto, A., Romayor, I., Herrero, A., Sanz, E., Arteta, B., et al. (2019). Ocoxin oral solution exerts an antitumoral effect in pancreatic cancer and reduces the stromal-mediated chemoresistance. Stromal-Mediated Chemoresistance 48, 555–567. doi:10.1097/MPA.0000000000001277
Hernandez-Unzueta, I., Benedicto, A., Telleria, U., Sanz, E., and Márquez, J. (2023). Improving the antitumor effect of chemotherapy with Ocoxin as a novel adjuvant agent to treat prostate cancer Prostate Cancer 15, 2536. doi:10.3390/nu15112536
Homma, K., Fujisawa, T., Tsuburaya, N., Yamaguchi, N., Kadowaki, H., Takeda, K., et al. (2013). SOD1 as a molecular switch for initiating the homeostatic ER stress response under zinc deficiency. Mol. Cell 52, 75–86. doi:10.1016/j.molcel.2013.08.038
Hosseini, A., and Ghorbani, A. (2015). Cancer therapy with phytochemicals: evidence from clinical studies. Avicenna J. Phytomed 5, 84–97.
Irshad, R., and Husain, M. (2021). Natural products in the reprogramming of cancer epigenetics. Toxicol. Appl. Pharmacol. 417, 115467. doi:10.1016/j.taap.2021.115467
Jiang, X., Peng, Q., Peng, M., Oyang, L., Wang, H., Liu, Q., et al. (2024). Cellular metabolism: a key player in cancer ferroptosis. Cancer. Commun. (Lond) 44, 185–204. doi:10.1002/cac2.12519
Jin, Z., Peng, F., Zhang, C., Tao, S., Xu, D., and Zhu, Z. (2023). Expression, regulating mechanism and therapeutic target of KIF20A in multiple cancer, Expr. Regul. Mech. Ther. target KIF20A multiple cancer 9, e13195. doi:10.1016/j.heliyon.2023.e13195
Kaidarova, D. R., Kopp, M. V., Pokrovsky, V. S., Dzhugashvili, M., Akimzhanova, Z. M., Abdrakhmanov, R. Z., et al. (2019). Multicomponent nutritional supplement Oncoxin and its influence on quality of life and therapy toxicity in patients receiving adjuvant chemotherapy. Oncol. Lett. 18, 5644–5652. doi:10.3892/ol.2019.10868
Kalinin, A., Zubkova, E., and Menshikov, M. (2023). Integrated stress response (ISR) pathway: unraveling its role in cellular senescence. Int. J. Mol. Sci. 24, 17423. doi:10.3390/ijms242417423
Kambe, T., Matsunaga, M., and Takeda, T. (2017). Understanding the contribution of zinc transporters in the function of the early secretory pathway. Int. J. Mol. Sci. 18, 2179. doi:10.3390/ijms18102179
Kanehisa, M., and Sato, Y. (2020). KEGG Mapper for inferring cellular functions from protein sequences. Protein Sci. 29, 28–35. doi:10.1002/pro.3711
Khan, T., Ali, M., Khan, A., Nisar, P., Jan, S. A., Afridi, S., et al. (2019). Anticancer plants: a review of the active phytochemicals, applications in animal models, and regulatory aspects. Appl. Animal Models, Regul. Aspects 10, 47. doi:10.3390/biom10010047
Kim, A., Ha, J., Kim, J., Cho, Y., Ahn, J., Cheon, C., et al. (2021). Natural products for pancreatic cancer treatment: from traditional medicine to modern drug Discovery. Nutrients 13, 3801. doi:10.3390/nu13113801
Koppula, P., Zhuang, L., and Gan, B. (2021). Cystine transporter SLC7A11/xCT in cancer: ferroptosis, nutrient dependency, and cancer therapy. Protein Cell 12, 599–620. doi:10.1007/s13238-020-00789-5
Kovaleva, I. E., Garaeva, A. A., Chumakov, P. M., and Evstafieva, A. G. (2016). Intermedin/adrenomedullin 2 is a stress-inducible gene controlled by activating transcription factor 4. Gene X. 590, 177–185. doi:10.1016/j.gene.2016.06.037
Kreß, J. K. C., Jessen, C., Hufnagel, A., Schmitz, W., Xavier da Silva, T. N., Ferreira Dos Santos, A., et al. (2023). The integrated stress response effector ATF4 is an obligatory metabolic activator of NRF2. Cell. Rep. 42, 112724. doi:10.1016/j.celrep.2023.112724
Labrie, M., Brugge, J. S., Mills, G. B., and Zervantonakis, I. K. (2022). Therapy resistance: opportunities created by adaptive responses to targeted therapies in cancer. Nat. Rev. Cancer. 22, 323–339. doi:10.1038/s41568-022-00454-5
Lee, G., Lee, J., Son, C., and Lee, N. (2021). Combating drug resistance in colorectal cancer using herbal medicines. Chin. J. Integr. Med. 27, 551–560. doi:10.1007/s11655-020-3425-8
Li, H., Liu, B., Lian, L., Zhou, J., Xiang, S., Zhai, Y., et al. (2021). High dose expression of heme oxigenase-1 induces retinal degeneration through ER stress-related DDIT3. Mol. Neurodegener. 16 (1), 16. doi:10.1186/s13024-021-00437-4
Li, S., Dong, P., Wang, J., Zhang, J., Gu, J., Wu, X., et al. (2010). Icariin, a natural flavonol glycoside, induces apoptosis in human hepatoma SMMC-7721 cells via a ROS/JNK-dependent mitochondrial pathway. Cancer Lett. 298, 222–230. doi:10.1016/j.canlet.2010.07.009
Liao, Y., Smyth, G. K., and Shi, W. (2013). featureCounts: an efficient general purpose program for assigning sequence reads to genomic features , 30, 923–930. doi:10.1093/bioinformatics/btt656
Liao, Y., Smyth, G. K., and Shi, W. (2022). featureCounts. Available at: https://subread.sourceforge.net/.
Lichten, L. A., and Cousins, R. J. (2009). Mammalian zinc transporters: nutritional and physiologic regulation. Annu. Rev. Nutr. 29, 153–176. doi:10.1146/annurev-nutr-033009-083312
Lichten, L. A., Ryu, M., Guo, L., Embury, J., and Cousins, R. J. (2011). MTF-1-mediated repression of the zinc transporter Zip10 is alleviated by zinc restriction. PLoS One 6, e21526. doi:10.1371/journal.pone.0021526
Lima-Pérez, M., Soriano-García, J. L., González-Meisozo, M., Soriano-Lorenzo, J. L., Fleites-Calvo, V., Morales-Morgado, D., et al. (2024). Nutritional supplement Ocoxin® combined with gemcitabine-based chemotherapy in patients with advanced pancreatic adenocarcinoma. Open J. Gastroenterol. 14, 267–287. doi:10.4236/ojgas.2024.148030
Lines, C. L., McGrath, M. J., Dorwart, T., and Conn, C. S. (2023). The integrated stress response in cancer progression: a force for plasticity and resistance. Front. Oncol. 13, 1206561. doi:10.3389/fonc.2023.1206561
Liu, Y., Huang, P., Li, Z., Xu, C., Wang, H., Jia, B., et al. (2022). Vitamin C sensitizes pancreatic cancer cells to erastin-induced ferroptosis by activating the AMPK/Nrf2/HMOX1 pathway. Oxid. Med. Cell. Longev. 2022, 5361241. doi:10.1155/2022/5361241
Liu, Y., Wang, X., He, D., and Cheng, Y. (2021). Protection against chemotherapy- and radiotherapy-induced side effects: a review based on the mechanisms and therapeutic opportunities of phytochemicals. Phytomedicine 80, 153402. doi:10.1016/j.phymed.2020.153402
Liu, Y., Wu, D., Fu, Q., Hao, S., Gu, Y., Zhao, W., et al. (2023). CHAC1 as a novel contributor of ferroptosis in retinal pigment epithelial cells with oxidative damage. Int. J. Mol. Sci. 24, 1582. doi:10.3390/ijms24021582
Lopes, C. M., Dourado, A., and Oliveira, R. (2017). Phytotherapy and nutritional supplements on breast cancer. Biomed. Res. Int. 2017, 7207983. doi:10.1155/2017/7207983
Love, M. I., Huber, W., and Anders, S. (2014). Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550–558. doi:10.1186/s13059-014-0550-8
Lu, H., Wang, X., Li, M., Ji, D., Liang, D., Liang, C., et al. (2022). Mitochondrial unfolded protein response and integrated stress response as promising therapeutic targets for mitochondrial diseases Targets Mitochondrial Dis. 12, 20. doi:10.3390/cells12010020
Luesch, H., and Paavilainen, V. O. (2020). Natural products as modulators of eukaryotic protein secretion. Nat. Prod. Rep. 37, 717–736. doi:10.1039/c9np00066f
Mahomoodally, M. F., Aumeeruddy, M. Z., Rengasamy, K. R. R., Roshan, S., Hammad, S., Pandohee, J., et al. (2021). Ginger and its active compounds in cancer therapy: from folk uses to nano-therapeutic applications. Semin. Cancer Biol. 69, 140–149. doi:10.1016/j.semcancer.2019.08.009
Maleki Dana, P., Sadoughi, F., Asemi, Z., and Yousefi, B. (2022). The role of polyphenols in overcoming cancer drug resistance: a comprehensive review. Cell. Mol. Biol. Lett. 27, 1–9. doi:10.1186/s11658-021-00301-9
Márquez, J., Mena, J., Hernandez-Unzueta, I., Benedicto, A., Sanz, E., Arteta, B., et al. (2016). Ocoxin® oral solution slows down tumor growth in an experimental model of colorectal cancer metastasis to the liver in Balb/c mice. Oncol. Rep. 35, 1265–1272. doi:10.3892/or.2015.4486
Masuelli, L., Benvenuto, M., Focaccetti, C., Ciuffa, S., Fazi, S., Bei, A., et al. (2021). Targeting the tumor immune microenvironment with “nutraceuticals”: from bench to clinical trials. Pharmacol. Ther. 219, 107700. doi:10.1016/j.pharmthera.2020.107700
Nakamura, M., Takano, A., Thang, P. M., Tsevegjav, B., Zhu, M., Yokose, T., et al. (2020). Characterization of KIF20A as a prognostic biomarker and therapeutic target for different subtypes of breast cancer. Int. J. Oncol. 57, 277–288. doi:10.3892/ijo.2020.5060
Nasir, A., Bullo, M. M. H., Ahmed, Z., Imtiaz, A., Yaqoob, E., Jadoon, M., et al. (2020). Nutrigenomics: epigenetics and cancer prevention: a comprehensive review. Crit. Rev. Food Sci. Nutr. 60, 1375–1387. doi:10.1080/10408398.2019.1571480
Naujokat, C., and McKee, D. L. (2021). The “big five” phytochemicals targeting cancer stem cells: curcumin, EGCG, sulforaphane, resveratrol and genistein. Curr. Med. Chem. 28, 4321–4342. doi:10.2174/0929867327666200228110738
Ng, C. X., Affendi, M. M., Chong, P. P., and Lee, S. H. (2022). The potential of plant-derived extracts and compounds to augment anticancer effects of chemotherapeutic drugs. Nutr. Cancer 74, 3058–3076. doi:10.1080/01635581.2022.2069274
Nguyen, T. S. L., Kohno, K., and Kimata, Y. (2013). Zinc depletion activates the endoplasmic reticulum-stress sensor Ire1 via pleiotropic mechanisms. Biosci. Biotechnol. Biochem. 77, 1337–1339. doi:10.1271/bbb.130130
Obiedat, A., Charpak-Amikam, Y., Tai-Schmiedel, J., Seidel, E., Mahameed, M., Avril, T., et al. (2020). The integrated stress response promotes B7H6 expression. J. Mol. Med. Berl. 98, 135–148. doi:10.1007/s00109-019-01859-w
Ogilvie, I., Kennaway, N. G., and Shoubridge, E. A. (2005). A molecular chaperone for mitochondrial complex I assembly is mutated in a progressive encephalopathy. J. Clin. Invest. 115, 2784–2792. doi:10.1172/JCI26020
Pakos-Zebrucka, K., Koryga, I., Mnich, K., Ljujic, M., Samali, A., and Gorman, A. M. (2016). The integrated stress response. EMBO Rep. 17, 1374–1395. doi:10.15252/embr.201642195
Palmiter, R. D., and Findley, S. D. (1995). Cloning and functional characterization of a mammalian zinc transporter that confers resistance to zinc. EMBO J. 14, 639–649. doi:10.1002/j.1460-2075.1995.tb07042.x
Pan, P., Huang, Y., Oshima, K., Yearsley, M., Zhang, J., Arnold, M., et al. (2019). The immunomodulatory potential of natural compounds in tumor-bearing mice and humans. Crit. Rev. Food Sci. Nutr. 59, 992–1007. doi:10.1080/10408398.2018.1537237
Peng, J., Song, X., Yu, W., Pan, Y., Zhang, Y., Jian, H., et al. (2024). The role and mechanism of cinnamaldehyde in cancer. J. Food Drug Anal. 32, 140–154. doi:10.38212/2224-6614.3502
Pérez-Peña, J., Díaz-Rodríguez, E., Sanz, E., and Pandiella, A. (2019). Central role of cell cycle regulation in the antitumoral action of Ocoxin. Nutrients 11, 1068. doi:10.3390/nu11051068
Rana, P., Shrama, A., and Mandal, C. C. (2021). Molecular insights into phytochemicals-driven break function in tumor microenvironment. J. Food Biochem. 45, e13824. doi:10.1111/jfbc.13824
Reed, D., Raina, K., and Agarwal, R. (2018). Nutraceuticals in prostate cancer therapeutic strategies and their neo-adjuvant use in diverse populations. NPJ Precis. Oncol. 2, 15–x. doi:10.1038/s41698-018-0058-x
Rodríguez-Menéndez, S., García, M., Fernández, B., Álvarez, L., Fernández-Vega-Cueto, A., Coca-Prados, M., et al. (2018). The zinc-metallothionein redox system reduces oxidative stress in retinal pigment epithelial cells, Nutrients, 10, 1874. doi:10.3390/nu10121874
Ron, D., and Walter, P. (2007). Signal integration in the endoplasmic reticulum unfolded protein response. Nat. Rev. Mol. Cell Biol. 8, 519–529. doi:10.1038/nrm2199
Ruiz Lorente, R., Hernández Durán, D., García Viamontes, J., Lence Anta, J., Ortiz Reyes, R., and Sanz Navares, E. (2020). Efficacy of oncoxin-viusid on the reduction of adverse reactions to chemotherapy and radiotherapy in patients diagnosed with cervical cancer and endometrial adenocarcinoma. Adenocarcinoma 11, 276–295. doi:10.4236/jct.2020.115023
Sadeghi, S., Davoodvandi, A., Pourhanifeh, M. H., Sharifi, N., ArefNezhad, R., Sahebnasagh, R., et al. (2019). Anti-cancer effects of cinnamon: insights into its apoptosis effects. Eur. J. Med. Chem. 178, 131–140. doi:10.1016/j.ejmech.2019.05.067
Sarmento-Ribeiro, A. B., Scorilas, A., Gonçalves, A. C., Efferth, T., and Trougakos, I. P. (2019). The emergence of drug resistance to targeted cancer therapies: clinical evidence. Drug Resist Updat 47, 100646. doi:10.1016/j.drup.2019.100646
Scuto, M., Ontario, M. L., Salinaro, A. T., Caligiuri, I., Rampulla, F., Zimbone, V., et al. (2022). Redox modulation by plant polyphenols targeting vitagenes for chemoprevention and therapy: relevance to novel anti-cancer interventions and mini-brain organoid technology. Free Radic. Biol. Med. 179, 59–75. doi:10.1016/j.freeradbiomed.2021.12.267
Shi, Q., Qian, Y., Wang, B., Liu, L., Chen, Y., Chen, C., et al. (2023). Glycyrrhizin protects against particulate matter-induced lung injury via regulation of endoplasmic reticulum stress and NLRP3 inflammasome-mediated pyroptosis through Nrf2/HO-1/NQO1 signaling pathway. Int. Immunopharmacol. 120, 110371. doi:10.1016/j.intimp.2023.110371
Shumsky, A., Bilan, E., Sanz, E., and Petrovskiy, F. (2019). Oncoxin nutritional supplement in the management of chemotherapy- and/or radiotherapy-associated oral mucositis. Mol. Clin. Oncol. 10, 463–468. doi:10.3892/mco.2019.1809
Smith, M. H., Ploegh, H. L., and Weissman, J. S. (2011). Road to ruin: targeting proteins for degradation in the endoplasmic reticulum. Science 334, 1086–1090. doi:10.1126/science.1209235
Snel, B., Lehmann, G., Bork, P., and Huynen, M. A. (2000). STRING: a web-server to retrieve and display the repeatedly occurring neighbourhood of a gene. Nucleic Acids Res. 28, 3442–3444. doi:10.1093/nar/28.18.3442
Sun, C., He, B., Gao, Y., Wang, X., Liu, X., and Sun, L. (2024). Structural insights into the calcium-coupled zinc export of human ZnT1. Sci. Adv. 10, eadk5128. doi:10.1126/sciadv.adk5128
Sun, Y., Kucej, M., Fan, H., Yu, H., Sun, Q., and Zou, H. (2009). Separase is recruited to mitotic chromosomes to dissolve sister chromatid cohesion in a DNA-dependent manner, 137, 123–132. doi:10.1016/j.cell.2009.01.040
Szklarczyk, D., Kirsch, R., Koutrouli, M., Nastou, K., Mehryary, F., Hachilif, R., et al. (2023). The STRING database in 2023: protein-protein association networks and functional enrichment analyses for any sequenced genome of interest. Nucleic Acids Res. 51, D638–D646. doi:10.1093/nar/gkac1000
Talib, W. H., Awajan, D., Hamed, R. A., Azzam, A. O., Mahmod, A. I., and Al-Yasari, I. H. (2022). Combination anticancer therapies using selected phytochemicals. Molecules 27, 5452. doi:10.3390/molecules27175452
Tang, Y., Zhuang, Y., Zhao, C., Gu, S., Zhang, J., Bi, S., et al. (2024). The metabolites from traditional Chinese medicine targeting ferroptosis for cancer therapy. Front. Pharmacol. 15, 1280779. doi:10.3389/fphar.2024.1280779
Tang, Z., Li, C., Kang, B., Gao, G., Li, C., and Zhang, Z. (2017). GEPIA: a web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 45, W98-W102–W102. doi:10.1093/nar/gkx247
Tian, X., Zhang, S., Zhou, L., Seyhan, A. A., Hernandez Borrero, L., Zhang, Y., et al. (2021). Targeting the integrated stress response in cancer therapy. Front. Pharmacol. 12, 747837. doi:10.3389/fphar.2021.747837
Travers, K. J., Patil, C. K., Wodicka, L., Lockhart, D. J., Weissman, J. S., and Walter, P. (2000). Functional and genomic analyses reveal an essential coordination between the unfolded protein response and ER-associated degradation, 101, 249–258. ddoi:10.1016/s0092-8674(00)80835-1
Tsoi, J., Robert, L., Paraiso, K., Galvan, C., Sheu, K. M., Lay, J., et al. (2018). Multi-stage differentiation defines melanoma subtypes with differential vulnerability to drug-induced iron-dependent oxidative stress. Cancer. Cell. 33, 890–904. doi:10.1016/j.ccell.2018.03.017
Viswanathan, V. S., Ryan, M. J., Dhruv, H. D., Gill, S., Eichhoff, O. M., Seashore-Ludlow, B., et al. (2017). Dependency of a therapy-resistant state of cancer cells on a lipid peroxidase pathway, 547, 453–457. doi:10.1038/nature23007
Vladu, A. F., Ficai, D., Ene, A. G., and Ficai, A. (2022). Combination therapy using polyphenols: an efficient way to improve antitumoral activity and reduce resistance. Int. J. Mol. Sci. 23, 10244. doi:10.3390/ijms231810244
Wang, H., Wang, X., Xu, L., Zhang, J., and Cao, H. (2020). Integrated analysis of the E2F transcription factors across cancer types. Oncol. Rep. 43, 1133–1146. doi:10.3892/or.2020.7504
Wang, K., Yu, Y., Chen, H., Chiang, Y., Ali, M., Shieh, T., et al. (2022). Recent advances in Glycyrrhiza glabra (Licorice)-Containing herbs alleviating radiotherapy- and chemotherapy-induced adverse reactions in cancer treatment Metabolites, 12, 535. doi:10.3390/metabo12060535
Wang, S., Zhang, F., Liu, J., and Qin, P. (2024). Efficacy and safety of Chinese patent medicine in the adjuvant treatment of prostate cancer: a Bayesian network meta-analysis. Med. Baltim. 103, e37180. doi:10.1097/MD.0000000000037180
Wang, W., Green, M., Choi, J. E., Gijón, M., Kennedy, P. D., Johnson, J. K., et al. (2019). CD8+ T cells regulate tumour ferroptosis during cancer immunotherapy. cancer Immunother. 569, 270–274. doi:10.1038/s41586-019-1170-y
Wang, W., Li, M., Wang, L., Chen, L., and Goh, B. (2023a). Curcumin in cancer therapy: exploring molecular mechanisms and overcoming clinical challenges. Cancer Lett. 570, 216332. doi:10.1016/j.canlet.2023.216332
Wang, X., Chen, J., Tie, H., Tian, W., Zhao, Y., Qin, L., et al. (2023b). Eriodictyol regulated ferroptosis, mitochondrial dysfunction, and cell viability via Nrf2/HO-1/NQO1 signaling pathway in ovarian cancer cells. J. Biochem. Mol. Toxicol. 37, e23368. doi:10.1002/jbt.23368
Woodring, R. N., Gurysh, E. G., Bachelder, E. M., and Ainslie, K. M. (2023). Drug delivery systems for localized cancer combination therapy. ACS Appl. Bio Mater. 6, 934–950. doi:10.1021/acsabm.2c00973
Xu, J., Song, Z., Guo, Q., and Li, J. (2016). Synergistic effect and molecular mechanisms of traditional Chinese medicine on regulating tumor microenvironment and cancer cells. Biomed. Res. Int. 2016, 1490738. doi:10.1155/2016/1490738
Xue, J., Li, R., Zhao, X., Ma, C., Lv, X., Liu, L., et al. (2018). Morusin induces paraptosis-like cell death through mitochondrial calcium overload and dysfunction in epithelial ovarian cancer. Chem. Biol. Interact. 283, 59–74. doi:10.1016/j.cbi.2018.02.003
Yang, C., Zhang, Y., Lin, S., Liu, Y., and Li, W. (2021). Suppressing the KIF20A/NUAK1/Nrf2/GPX4 signaling pathway induces ferroptosis and enhances the sensitivity of colorectal cancer to oxaliplatin. Aging 13, 13515–13534. doi:10.18632/aging.202774
Ye, L., Wen, X., Qin, J., Zhang, X., Wang, Y., Wang, Z., et al. (2024). Metabolism-regulated ferroptosis in cancer progression and therapy. Cell. Death Dis. 15, 196–y. doi:10.1038/s41419-024-06584-y
Zhang, G., Wang, X., Li, C., Li, Q., An, Y. A., Luo, X., et al. (2021). Integrated stress response couples mitochondrial protein translation with oxidative stress control, 144, 1500–1515. doi:10.1161/CIRCULATIONAHA.120.053125
Zhang, L., Cheng, X., Xu, S., Bao, J., and Yu, H. (2018). Curcumin induces endoplasmic reticulum stress-associated apoptosis in human papillary thyroid carcinoma BCPAP cells via disruption of intracellular calcium homeostasis. Med. Baltim. 97, e11095. doi:10.1097/MD.0000000000011095
Zhang, W., Li, S., Li, C., Li, T., and Huang, Y. (2022). Remodeling tumor microenvironment with natural products to overcome drug resistance. Front. Immunol. 13, 1051998. doi:10.3389/fimmu.2022.1051998
Zhang, Y., Sheng, Z., Xiao, J., Li, Y., Huang, J., Jia, J., et al. (2023). Advances in the roles of glycyrrhizic acid in cancer therapy. Front. Pharmacol. 14, 1265172. doi:10.3389/fphar.2023.1265172
Zhao, T., Xu, Y., Hu, H., Gong, H., and Zhu, H. (2019). Isoliquiritigenin (ISL) and its formulations: potential antitumor agents. Curr. Med. Chem. 26, 6786–6796. doi:10.2174/0929867325666181112091700
Keywords: Ocoxin, cancer, plant extract, ferroptosis, adjuvant, chemotherapy
Citation: Hernandez-Unzueta I, Telleria-Gonzalez U, Aransay AM, Martin Rodriguez JE, Sanz E and Márquez J (2025) Unravelling the antitumor mechanism of Ocoxin through cancer cell genomics. Front. Pharmacol. 16:1540217. doi: 10.3389/fphar.2025.1540217
Received: 05 December 2024; Accepted: 24 February 2025;
Published: 19 March 2025.
Edited by:
Zhenhua Chen, Jinzhou Medical University, ChinaReviewed by:
Ruoxi Zhang, University of Texas Southwestern Medical Center, United StatesCopyright © 2025 Hernandez-Unzueta, Telleria-Gonzalez, Aransay, Martin Rodriguez, Sanz and Márquez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Joana Márquez, am9hbmEubWFycXVlekBlaHUuZXVz; Iera Hernandez-Unzueta, aWVyYS5oZXJuYW5kZXpAZWh1LmV1cw==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.