
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Pharmacol. , 26 February 2025
Sec. Translational Pharmacology
Volume 16 - 2025 | https://doi.org/10.3389/fphar.2025.1519278
This article is part of the Research Topic Nutrition as a Pharmacological Approach to Metabolic Disorders and Ageing View all 7 articles
 Jaline Faiad†
Jaline Faiad† Márcia Fábia Andrade†
Márcia Fábia Andrade† Gabriela de Castro
Gabriela de Castro Joyce de Resende
Joyce de Resende Marina Coêlho
Marina Coêlho Giovana Aquino
Giovana Aquino Marilia Seelaender*
Marilia Seelaender*Cancer cachexia (CC) is characterized by significant skeletal muscle wasting, and contributes to diminished quality of life, while being associated with poorer response to treatment and with reduced survival. Chronic inflammation plays a central role in driving CC progression, within a complex interplay favoring catabolism. Although cachexia cannot be fully reversed by conventional nutritional support, nutritional intervention shows promise for the prevention and treatment of the syndrome. Of special interest are nutrients with antioxidant and anti-inflammatory potential and those that activate pathways involved in muscle mass synthesis and/or in the inhibition of muscle wasting. Extensive research has been carried out on novel nutritional supplements’ power to mitigate CC impact, while the mechanisms through which some nutrients or bioactive compounds exert beneficial effects on muscle mass are still not totally clear. Here, we discuss the most studied supplements and nutritional strategies for dealing with muscle loss in CC.
Cancer cachexia (CC) is characterized by progressive functional debilitation and significant skeletal muscle mass loss, often accompanied by fat mass wasting. The decrease in muscle mass contributes to diminished quality of life, increased fatigue and morbidity, and is associated with poorer responses to oncological therapy (von Haehling and Anker, 2014). Cachexia related muscle loss frequently provokes, the early discontinuation of treatment, the necessity of chemotherapeutic drug dose adjustment, and is robustly linked with worsened prognosis and decreased survival (Mattox, 2017).
Cancer cachexia definition and staging are still controversial, which makes it difficult to compare the existing data and to determine its prevalence (Wiegert et al., 2020). Within the most widely accepted consensus (Fearon et al., 2011), the syndrome is divided into three progressive stages based on clinical and biochemical parameters: pre-cachexia, cachexia, and refractory cachexia (RCa), the latter being characterized by non-responsiveness to anticancer therapies and an expected survival of up to 3 months, solely (Fearon et al., 2011). Wiegert et al. (2020) compared three diagnostic criteria for cachexia in 1,384 patients with incurable cancer under palliative care. According to their findings, 17.3% of the patients were cachectic, 20.8% were in pre-cachexia, 53.3% were in the RCa stage, and 8.2% were non-cachectic by Viagno et al.‘s classification. Applying Blum et al.'s criteria, 53.9% of the patients were classified as cachectic, 12.3% as pre-cachectic, 26.1 as RCa, and 9.7% as non-cachectic. In contrast, Wallengren et al.'s criteria identified 13.8% of the patients as cachectic and 86.2% as non-cachectic. Cachexia was found to be most prevalent among patients with gastrointestinal tract tumors (Wiegert et al., 2020). On the other hand, a study by Orellana López et al. (2023), which determined cachexia prevalence using the miniCASCO tool in a cohort of cancer patients in Chile, found that 27.5% presented cachexia. Within this group, 45.45% could be classified to be in the stage of pre-cachexia, and 36.36% in RCa, and 18.18%, cachexia. Both studies underscore the importance of CC classification to clinical practice and its potential to guide treatment decisions effectively, particularly at the early stages of the syndrome (Wiegert et al., 2020; Orellana López et al., 2023).
Despite the traditional definition that cachexia cannot be fully reversed by conventional nutritional support (Fearon et al., 2011), multimodal interventions at the onset of the syndrome appear to allow some gains. An anabolic window is observed in patients with cancer cachexia (CC), particularly at the earlier stages of the disease or during periods of clinical stability. These phases are characterized by improved symptom control, optimized pain management, enhanced nutritional intake, and better physical performance (Prado et al., 2013). Later along the progression of the disease, many factors lead to possible anabolic resistance, such as tumor site and stage, anticancer treatment side-effects, life expectancy, nutritional status and dietary intake, and inflammation (Engelen et al., 2016).
The difficulties in cachexia management arise from the fact that the disease is per se, multifactorial in nature, and is driven by a combination of inflammation, disrupted metabolic processes, and negative energy and protein balance (Fearon et al., 2011). Chronic inflammation plays a central role in CC onset and progression, promoting tumor-host interaction mediated by pro-inflammatory cytokines (Donohoe et al., 2011). Tumor Necrosis Factor-alpha (TNF-α), also known as cachectin, is one of the key cytokines involved in CC (Fearon et al., 2012). It promotes protein degradation by activating the intracellular NF-κB pathway, which induces the expression of genes associated with the process of proteolysis, including Muscle RING-Finger protein-1 (MuRF1) and atrogin-1 (Armstrong et al., 2020). Interleukin (IL)-6 is another crucial cytokine in cachexia (Fearon et al., 2012). It activates STAT3, which then translocates to the nucleus, promoting the expression of genes involved in protein degradation and inflammation, hence exacerbating tissue loss (Martin et al., 2023; Fonseca et al., 2020). Furthermore, myostatin and activin A, members of the TGF-β superfamily, act as negative regulators of muscle mass by activating the SMAD2/3 pathway, which represses muscle differentiation and regeneration (Fearon et al., 2012). FoxO transcription factors also contribute to muscle atrophy, by up regulating ubiquitin ligases and promoting autophagy (Martin et al., 2023). TWEAK, another cytokine reported to be augmented in CC, exacerbates muscle wasting by inducing proteolytic enzymes and promoting inflammation (Armstrong et al., 2020; Martin et al., 2023). Another highly relevant catabolic factor associated with CC is Growth Differentiation Factor 15 (GDF15), a cytokine released in response to several stress signals. Augmented circulating levels of GDF15 may lead to weight loss and anorexia in patients with CC and are associated with reduced survival (Lerner et al., 2015; Suzuki et al., 2021). To our knowledge, no published studies have established a correlation among nutritional supplementation, GDF15 levels, and muscle mass regulation.
The pathophysiology of CC involves a complex interplay between anabolic and catabolic pathways, with a pronounced shift towards protein degradation in detriment of synthesis. In anabolic states, protein synthesis exceeds degradation resulting in muscle protein gain, while the opposite leads to catabolism, resulting in muscle mass loss (Stipanuk, 2008). The Mammalian Target of the Rapamycin (mTOR) pathway, regulated through the IGF-1/PI3K/AKT cascade, has a central role in promoting muscle protein synthesis (Yoshida and Delafontaine, 2020). However, in CC, mTOR signaling is often inhibited due to increased levels of proinflammatory cytokines, such as TNF-α (Fonseca et al., 2020).
Muscle loss in CC is predominantly mediated by two main pathways: the Ubiquitin-proteasome system (UPS) and the Autophagy-lysosome pathway (ALP) (Martin et al., 2023; Aversa et al., 2016). The UPS is responsible for degrading damaged or malfunctioning intracellular proteins. This process begins with tagging the target by adding ubiquitin chains, a process mediated by MuRF1 and atrogin-1, also known as Muscle Atrophy F-box (MAFbx) (Foletta et al., 2011). These ubiquitinated proteins are then recognized and directed to the proteasome, a proteolytic complex that degrades the tagged proteins into smaller peptides (Foletta et al., 2011). In CC, there is increased expression of E3 ligases, what facilitates the ubiquitination of muscle proteins, leading to enhanced protein degradation and consequent muscle mass loss (Martin et al., 2023). Simultaneously, ALP, which generally maintains cellular homeostasis by degrading damaged organelles, is pathologically increased in CC, contributing to muscle atrophy through the degradation of both fiber proteins and organelles (Sandri, 2013). Catabolic factors derived from the tumor, such as proteolysis-inducing factor (PIF), directly activate the UPS, further exacerbating muscle proteolysis (Fearon et al., 2012).
Fearon et al. (2012) state that the impossibility of reversal of the cachectic state by conventional nutritional support is a marking feature of CC. Nevertheless, extensive research has been carried out on the potential of novel nutritional supplements to mitigate the syndrome’s deleterious impact, and understanding the balance between protein synthesis and degradation pathways is mandatory in this scenario. There is no standardized treatment for CC. Various nutritional intervention strategies have shown promise in preventing and treating this syndrome, with some nutrients or bioactive compounds demonstrating beneficial effects, whether alone or in combination. The mechanisms through which these interventions exert their effects on muscle mass are still uncovered, yet they offer a potential therapeutic avenue. One postulated mechanism by which nutrients can mitigate cachexia malnutrition is interfering with systemic inflammation. Overall, this review aims to explore the most studied nutritional supplements and strategies for treating and/or preventing muscle loss in CC.
Although recognized as the primary anabolic stimulus in skeletal muscle metabolism, studies investigating the effects of increased total protein intake in cancer patients are relatively recent. As revised by Prado and colleagues (Prado et al., 2020), current guidelines for adequate protein intake in oncologic patients fail to address factors such as body composition and muscle depletion. The recommended protein intake for cancer patients ranges from 1 to 1.5 g/kg (Muscaritoli et al., 2021), which appears to be insufficient. Colorectal cancer patients submitted to a high-protein diet at the pre-cachexia stage presented a reduction in subclinical inflammation, one of the main drivers of muscle wasting in cachexia, as well as an improvement in the nutritional status and in appetite (Ziętarska et al., 2017). Also, a systematic review (Capitão et al., 2022) comprising different types of cancer associated with sarcopenia (head and neck, lung, esophageal cancer) reported a minimum amount of protein of 1.4 g/kg to ensure muscle mass maintenance in this population, with lower amounts being associated with muscle loss along the treatment.
It is also important to consider the debate on higher protein intake and its association with tumor growth, since protein synthesis in the tumor, muscle, and immune cells involves the same signaling pathways (Butler et al., 2021; Li et al., 2007). A study with a model of cachectic colon tumor-bearing rats undergoing chemotherapy (Boutière et al., 2023) demonstrated that trends in tumor growth and response to chemotherapy remained unaltered despite enhanced protein dietary intake. Moreover, a modest improvement in nutritional status was observed in animals submitted to a high-protein diet, with an increase in relative fat-free mass. However, in neither of these studies, differences were reported regarding protein metabolism in skeletal muscle.
Most recently, a randomized clinical trial reported the effects of the total daily protein intake on patients with stage II-IV colorectal cancer submitted to chemotherapy (Ford et al., 2024). At baseline, most patients had lower protein ingestion than recommended by the guidelines, and individuals who managed to increase their protein ingestion favored the maintenance of muscle mass, physical function, and anabolism. One of the goals of the study was to assess the response to a diet containing 2.0 g/kg/day of protein versus another, in which 1.0 g/kg/day of protein was consumed, but only 35,3% of the patients in the 2.0 g/kg/day achieved the recommended protein intake. Despite not having succeeded in increasing protein ingestion to the proposed values, the study recognizes that individualized nutritional counselling had a promising effect on protein intake and muscle mass maintenance. Even though CC was an exclusion criterion in this pilot study, it represents the first attempt to demonstrate the effect of nutritional support alone on muscle loss prevention in patients with cancer, as well as providing an evidence-based optimal protein dose for this population, considering features such as acceptance, feasibility and efficacy of the diet regimen.
Along with increasing diet protein intake, the use of specific proteins and amino acids has also been the target of many studies. Among the strategies examined, we chose to address those with clearer evidence of beneficial impact.
The high concentration of easily digestible essential amino acids in whey protein (WP) renders it ideal as an effective way to add up proteins in the patient’s diet. WP has thus become a suitable choice for providing protein support for cancer patients, as it offers greater nutritional value and faster absorption compared to alternative dietary protein sources (Ramani et al., 2024).
WP is particularly rich in the branched-chain amino acids valine, leucine, and isoleucine, which play a crucial role in tissue growth and repair (Brestenský et al., 2015). Additionally, whey protein is a valuable source of cysteine and methionine, which are glutathione precursors and a key component for enhancing immune function (Mir Khan and Selamoglu, 2020). The whey protein subfractions most studied for their ability to disrupt tumor pathways include α-lactalbumin, bovine serum albumin, and lactoferrin (Teixeira et al., 2019). In vitro and in vivo research has demonstrated their anticancer properties, including decreased tumor occurrence, growth suppression, and improved antioxidant activity. Furthermore, it can potentially improve conventional cancer treatment efficacy and reduce side effects (Teixeira et al., 2019). In addition, WP has been shown to enhance immune function, improve nutritional status, and promote overall health, making it a beneficial strategy for cachectic patients (Zhao et al., 2022). Nonetheless, changes in muscle metabolism, such as a lower anabolic potential, are common in advanced CC (Prado et al., 2013), where protein synthesis remains impaired even when sufficient protein is consumed. Therefore, nutritional strategies should focus on both the increase of muscle protein synthesis and decrease of anabolic resistance (Van de Worp et al., 2020; Antoun and Raynard, 2018).
In a clinical trial involving cachectic patients undergoing chemotherapy and receiving nutritional supplementation, adding WP to the 3-month treatment regimen significantly improved body composition, muscle strength, and body weight, while reducing chemotherapy toxicity (Cereda et al., 2019).
Several studies indicate that WP supplementation, which is rich in leucine and other essential amino acids, can stimulate protein synthesis more effectively than other protein sources. This effect is attributed to the rapid digestion of WP, which leads to a swift increase in plasma amino acid levels, particularly essential amino acids, potentially mitigating muscle loss in cachectic patients (Cereda et al., 2019; Dangin et al., 2002; Deutz et al., 2011a; Dillon et al., 2007).
Therefore, WP may benefit patients with cancer-associated cachexia by improving muscle synthesis and immune function and reducing inflammation. While WP supplementation shows promise in improving clinical outcomes and quality of life, further studies are needed to define the optimal dosage and evaluate long-term effects.
The amino acids known as BCAA, leucine, isoleucine, and valine, are essential amino acids that play an important role in muscle metabolism. BCAAs have been studied in CC in regard to eventual potential to mitigate muscle mass loss (Ananieva et al., 2016). They interfere with the activation of catabolic pathways, such as the ubiquitin-proteasome system, which are exacerbated in cachexia (Setiawan et al., 2023). BCAA supplementation has been associated with improvement in muscle strength and function, modulation of inflammation, and improvement in energy metabolism, contributing to better quality of life (Storck et al., 2020; Gala et al., 2020).
The benefits of BCAA supplementation, including improved post-chemotherapy recovery - characterized by weight gain and increased energy levels - have been broadly examined (Deutz et al., 2011b; Nojiri et al., 2017; Katagiri et al., 2020; Hachiya et al., 2020). It has also been suggested that BCAAs may increase mitochondrial biogenesis (Valerio et al., 2011), potentially benefiting skeletal muscle energy metabolism (Borack and Volpi, 2016; van Dijk et al., 2015). On the other hand, recent studies have reported that BCAA supplementation may be detrimental to cancer patients, as potentially, tumor cells can consume these amino acids and thus remain alive even within anaerobic environments (Ananieva and Wilkinson, 2018; Ericksen et al., 2019; Lei et al., 2020; Lieu et al., 2020; Peng et al., 2020; Taherizadeh et al., 2021; Tang et al., 2010; Wang et al., 2018). Although limited by the absence of a control group and different types of tumors, two studies showed improvement in patient strength and quality of life (Van der Meij et al., 2019; Zanetti et al., 2020).
BCAAs act as metabolic regulators, influencing not only protein synthesis but also lipid and glucose metabolism (Zhang et al., 2017). Most circulating BCAAs are reincorporated into proteins, functioning as building blocks for muscle synthesis and providing nitrogen for the biosynthesis of nucleotides and nonessential amino acids (Jung et al., 2021). Supplementation in cachectic patients should be carefully monitored, as excessive intake of BCAAs may lead to an imbalance in the availability and metabolism of other essential amino acids (Storck et al., 2020), while the potential effect on tumor progression should be monitored.
Leucine is an essential amino acid from the BCAA group. It is known to be an agonist of the mTOR pathway, playing an important role as an anabolic mediator in protein metabolism (Ananieva et al., 2016; Tian et al., 2019). Studies have shown that leucine may play a role in immune function by activating T cells (Ananieva et al., 2016) and regulating the immune response through mTOR signaling, which is crucial in regulating pro- and anti-inflammatory cytokines (Thomson et al., 2009; Soares et al., 2020; Van der Ende et al., 2018).
A leucine-rich diet has shown promising effects in preserving serum insulin concentration along cancer progression, thus promoting muscle protein synthesis and attenuating anabolic resistance without impacting tumor growth in animal models (Cruz et al., 2019). Studies performed by Salomão et al. (2010) and Viana et al. (2016) reinforce these observations, having demonstrated that leucine supplementation can be an effective strategy to improve protein metabolism in cancer patients (Salomão et al., 2010; Viana et al., 2016).
Research in tumor-bearing animals (Cruz et al., 2019; Viana et al., 2016; Gomes-Marcondes et al., 2003; Peters et al., 2011; Xia et al., 2017) and studies with older sarcopenic individuals (Rasmussen et al., 2016; Dickinson et al., 2013) suggest that leucine is a safe and efficient supplement. It positively affects protein metabolism, muscle mass gain, protein and caloric intake, and effectively modulates the inflammatory process.
Although research has shown that leucine can reduce muscle degradation and improve protein metabolism and the inflammatory profile during cancer progression, few studies have evaluated its direct effects upon tumor growth, vascularization, and proliferation. More research is thus needed to understand its impact in the context of human cancer.
HMB is a metabolite derived from leucine metabolism. It has therapeutic potential in CC due to its effects in stimulating protein synthesis and in inhibiting muscle degradation. HMB was initially employed to improve wound healing (Williams et al., 2002), as this metabolite shows significant capacity to enhance muscle protein accretion and to induce collagen renewal (Zanchi et al., 2011).
The anabolic potential of HMB supplementation has been explored for the treatment of muscle loss in cancer, with results pointing out to its capacity to improve long-term outcomes (Prado et al., 2022). Tumor-bearing models show that HMB supplementation attenuates body weight and muscle mass loss at a dose similar to that adopted for human beings. Furthermore, HMB presents antitumor, anti-inflammatory, and anti-cachectic effects (Nunes et al., 2008; Zaira et al., 2011) Other studies showed increased survival time and promotion of favorable metabolic changes (Caperuto et al., 2007) and a significantly larger fiber cross-sectional area after HMB supplementation (Hao et al., 2011). Therefore, HMB supplementation may be a good option for complementary cancer therapy.
A more recent systematic review provided evidence that HMB supplementation benefits muscle mass and function in cancer patients. Additionally, the supplement has been demonstrated to be safe and tolerable, with no adverse effects (Prado et al., 2022). Studies suggest that HMB may effectively mitigate proteolysis associated with cachexia because of its capacity to reduce the activity of the ubiquitin-proteasome system, the main pathway for protein degradation and caspase activity (Prado et al., 2022). Furthermore, HMB stimulates proteogenesis through direct action on the mTOR pathway, the canonical protein synthesis pathway. HMB can also promote the activation of satellite cells in skeletal muscle and potentially enhance the tissue’s regenerative capacity, directly affecting cell proliferation and differentiation (Chodkowska et al., 2018; Yamada et al., 2022).
Nutrients and exercise activate the mTOR pathway in muscle, supporting muscle anabolism. In contrast, in cancer cells, genetic mutations can lead to chronic hyperactivation of the mTOR pathway, fueling tumor growth and bypassing the standard regulatory inhibitory mechanisms. However, studies have not shown a direct link between HMB supplementation and tumor growth (Tian et al., 2019; Prado et al., 2022).
HMB has also been proposed to operate through its capacity to stabilize the sarcolemma via cholesterol synthesis. It has been shown that the majority of HMB is converted into 3-hydroxy-3-methyl-glutaryl-coenzyme A reductase, which is the limiting step of cholesterol synthesis. As such, increased intramuscular levels of HMB may serve as the available substrate for cholesterol synthesis, and therefore for formation and stabilization of sarcolemma (Rossi et al., 2017). HMB is recognized as an effective anti-catabolic agent, reducing protein degradation while enhancing protein synthesis. The currently recommended daily dose is 3 g/day (Muscaritoli et al., 2021).
Glutamine is the most abundant amino acid in the plasma and in the skeletal muscle (Cruzat et al., 2018). In healthy individuals this amino acid plays an essential role in the regulation of cellular functions, among which immune cell activity, energy metabolism, maintenance of intestinal mucosal integrity, and protein synthesis (Newsholme et al., 2003). Glutamine metabolism is, however, disturbed in cancer patients, resulting in dysregulated protein synthesis, which can contribute to muscle wasting (Cruzat et al., 2018). Thus, the therapeutic potential of glutamine supplementation in cancer patients has been widely discussed, under the light of contributing to the preservation of muscle mass and warranting quality of life in patients with cancer-related wasting (Pradhan et al., 2024).
A cohort of 44 surgery patients with head and neck cancer received enteral glutamine supplementation for 4 weeks, having achieved significantly higher nutritional status and improved clinical outcomes than the control group. Notably, the intervention group maintained lean body mass, which correlated with a higher quality of life score during the postoperative period (Azman et al., 2015). Furthermore, other trials have investigated the effects of glutamine supplementation in combination with HMB and arginine, attenuating cancer-related wasting in association with increased free fat mass in patients with advanced disease, as observed by May et al. (2002).
In animal models, several studies demonstrated improvement in energy balance and inhibition of tumor growth following glutamine supplementation, as reviewed by Van de Worp et al. (2020). Walker-256 tumor-bearing rats supplemented with 2% l-glutamine exhibited reduced body weight loss and a lower percentage (%) loss of body mass index (BMI) (cachexia index), calculated according to tumor mass (Fracaro et al., 2016). These studies suggest that glutamine supplementation may improve muscle preservation and energy metabolism in CC patients.
Besides the promising findings, glutamine supplementation raises concerns due to its role in tumor growth. Since there is metabolic competition for glutamine between host cells and tumor cells, it is crucial to find a balance that supports normal cell function without promoting tumorigenesis during supplementation (Wang et al., 2024; Muranaka et al., 2024). Therefore, glutamine supplementation needs to be carefully monitored, and further research evaluating the potential benefits related to muscle loss in cancer-associated cachexia is necessary.
Arginine is a conditionally essential amino acid that depends on metabolic status. Arginine stimulates cell growth and protein synthesis by inducing the activation of the mTOR pathway in the muscle (Panwar et al., 2023). In addition, arginine serves as a substrate for nitric oxide synthesis, an important signaling molecule that affects immune function, vasodilation, and cicatrization (Wu et al., 2021). Considering its importance in several biological functions, this amino acid has been suggested to improve patient outcome in CC by regulating tumor growth and promoting anabolic effects in the muscle (Pradhan et al., 2024). In addition, in immune system cells, arginine supplementation has been shown to promote beneficial immunomodulatory gain. Finally, it has been shown to improve nutritional status (Soares et al., 2020).
Under catabolic conditions in cancer, arginine levels in plasma are decreased, as observed in patients with breast cancer, colon cancer, and pancreatic cancer, independent of weight loss or tumor stage (Vissers et al., 2005). This result indicates disturbed arginine metabolism in the disease.
A study with arginine supplementation during the perioperative period showed no beneficial effect upon outcome in patients with head and neck cancer; nonetheless, the arginine-supplemented group demonstrated a trend toward improved survival span (Van Bokhorst-de van der Schueren et al., 2001), consistent with findings reported by the same group in a previous study (Buijs et al., 2010). Antoun and Raynard (2018) discussed that arginine supplementation still lacks robust clinical evidence regarding its benefits on muscle wasting. Randomized trials with larger sample sizes, focusing on arginine supplementation in patients with cancer-associated cachexia, may in the future elucidate arginine’s direct and indirect effects on wasting conditions.
Creatine is synthesized endogenously from three amino acids. The initial stages occur in the kidney, where arginine and glycine are involved, and the subsequent steps in the liver, with the participation of methionine. It can also be acquired through an animal protein-rich diet (Tanaka et al., 2022). Most of creatine is absorbed, stored, and used by the skeletal muscle (Jung et al., 2013; Gualano et al., 2012), suggesting creatine supplementation to be able to increase muscle strength and lean body mass in myopathies (Harris et al., 1992).
The effects of creatine supplementation in cachectic patients must still be well established. A study with patients with colorectal cancer under chemotherapy and with CC showed no changes in neither muscle mass, nor body composition (Jatoi et al., 2017). Another randomized and double-blind clinical trial was carried out with 30 individuals with stage III-IV colorectal cancer who received creatine or placebo over an 8-week period. The creatine supplementation protocol included an initial loading phase of 4 × 5 g per day during the first week, followed by a maintenance phase of 2 × 2.5 g per day. Creatine was not effective in improving muscle mass gain or its function. In addition, there were no changes in the quality of life of these patients. However, it enhanced the bioimpedance phase angle, which is related to improved prognosis (Norman et al., 2006).
Studies in rats with cachexia showed that creatine supplementation attenuated weight loss and decreased tumor growth (Cella et al., 2020; Campos-Ferraz et al., 2016; Deminice et al., 2016; Wei et al., 2022). In addition, supplementation promoted lower plasma concentration of TNF-α and IL-6 (pro-inflammatory cytokines), while increasing the concentration of IL-10 (anti-inflammatory cytokine), and preventing atrogin-1 and MuRF-1, key regulators of muscle atrophy in the skeletal muscle (Cella et al., 2020).
Although the recommended dose for humans is 3–5 g/day (Tanaka et al., 2022), this dose did not show the same results in patients with cachexia (Jatoi et al., 2017; Norman et al., 2006), suggesting that changes in the metabolic pathways associated with the disease may limit the effects of creatine (Tanaka et al., 2022).
L-carnitine (LC) is among the most studied nutritional supplements in advanced cancer patients with malnutrition (Johal et al., 2022) due to its antioxidant activity and possible anti-wasting effect (Alhasaniah, 2023). It is synthesized in the liver and kidney by converting the amino acids lysine and methionine. Its primary function is to facilitate the transport of long-chain fatty acids to the mitochondrial matrix for β-oxidation and subsequent energy production (Alhasaniah, 2023; Longo et al., 2016). Failure in this process can lead to increased oxidative stress, metabolic dysfunctions, and increased pro-inflammatory cytokines. In this context, LC supplementation can reduce oxidative stress and the inflammatory response, leading to potential clinical benefits (Longo et al., 2016).
Gramignano et al. (2006) evaluated the effect of LC supplement intake in 12 patients with advanced cancer, having observed a significant increase in lean mass and an improvement in appetite after 4 weeks, suggesting that LC can mitigate muscle loss and improve nutritional status. In a phase III study by Mantovani et al. (2010), although the use of a LC supplement (4 g/day) for 4 months did not significantly improve primary outcomes, such as lean mass and fatigue, a positive impact on secondary indicators such as the Glasgow Prognostic Score and ECOG (Eastern Cooperative Oncology Group) performance status was reported. Similarly, the study by Kraft et al. (2012) in patients with advanced pancreatic cancer showed that LC supplementation (4 g/day) for 12 weeks increased BMI and improvement in quality of life, further supporting the therapeutic potential of LC in patients with cancer.
In experimental models, LC treatment improved food intake, reduced muscle mass loss, and led to the downregulation of atrogin-1 and MuRF1, biomarkers of skeletal muscle atrophy. In addition, LC intake decreased proteasome activity (degradation pathway) in the gastrocnemius muscle of cachectic animals (Busquets et al., 2012; Busquets et al., 2020) and increased physical activity levels by improving energy production (Busquets et al., 2012). Another intervention with the LC supplement led to an increase in both food intake and muscle mass. The study also examined the activity of carnitine palmitoyltransferase-1 (CPT-1), a marker of the effects of carnitine. The results demonstrated an upregulation of CPT-1, associated with reduced plasma levels of IL-6 and TNF-α (Liu et al., 2011). Other experimental studies support the finding that CPT-1 activity is higher in the muscles of cachectic animals supplemented with LC (Busquets et al., 2020; Silvério et al., 2012) in relation to controls. In cachectic animals, CPT-1 expression is typically reduced, contributing to an accumulation of triacylglycerols in the liver. A 28-day supplementation of LC administered intragastrically at a dose of 1 g/kg body weight/day, effectively restored CPT-1 activity, thereby preserving hepatic lipid metabolism. Additionally, LC modulated CPT I enzymatic activity and MTP gene expression, further supporting healthy hepatic lipid regulation. The supplementation also enabled normal weight gain in tumor-bearing animals and significantly inhibited tumor growth, demonstrating LC’s potential as a therapeutic strategy for managing cachexia (Silvério et al., 2012).
The available evidence suggests that LC has the potential to mitigate muscle mass loss, modulate lipid metabolism, and reduce inflammation associated with CC. Nonetheless, further studies are required to investigate the underlying molecular mechanisms to understand its therapeutic effects better.
Marine long-chain omega-3 (n-3) fatty acids, eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) are well-known for their anti-inflammatory properties. N-3 has been shown to decrease tumor angiogenesis and invasiveness (McCarty, 1996). N-3 supplementation can alter inflammatory markers in patients with colorectal cancer, decreasing IL-6 and increasing albuminemia (Camargo et al., 2016). Additionally, n-3 polyunsaturated fatty acids can induce ferroptosis through lipid peroxidation in acidic tumor environments, selectively promoting cancer cell death (Dierge et al., 2021), while also inhibiting tumor proliferation and modulating pro-resolving lipid mediators, which help suppress chronic inflammation (Liput et al., 2021).
We have previously discussed in a meta-analysis that n-3 fatty supplementation was not able to lower circulating inflammatory markers in cachexia (de Castro et al., 2022). Nevertheless, due to the low number of studies that matched our inclusion and exclusion criteria, only six articles were included in the systematic review, disallowing a strong conclusion (de Castro et al., 2022). Oral nutritional supplements with high protein and n-3 content have been shown to preserve lean body mass during chemo (radio) therapy (de van der Schueren et al., 2018). However, another meta-analysis could not support this result, indicating that oral n-3 supplements could not maintain muscle and body weight mass nor improve the quality of life in patients with cancer (Lam et al., 2021).
Despite these contradictions, the European Society for Clinical Nutrition and Metabolism (ESPEN) advocates, although with a weak recommendation, the supplementation of n-3 for some patients with cancer (Muscaritoli et al., 2021). Notably, patients with advanced cancer undergoing chemotherapy and endangered by weight loss or malnutrition could benefit from n-3 supplementation due to its ability to improve appetite, food consumption, lean body mass, and body weight (Muscaritoli et al., 2021). The safety and tolerability of n-3 supplements were also highlighted (Muscaritoli et al., 2021).
An increase in oxidative stress occurs when the normal redox equilibrium is altered, with increased presence of oxidative species and lower antioxidant defense status (Li et al., 2022). Patients with lung cancer and cachexia have higher skeletal muscle protein carbonyls and superoxide anions content compared with healthy controls (Puig-Vilanova et al., 2015). It has been proposed that dietary antioxidants may modulate the oxidative stress in CC (Li et al., 2022).
A comprehensive meta-analysis including approximately 68 clinical trials did not identify protective effects of antioxidants against cancer. On the contrary, an increase in mortality was observed in individuals who consumed β-carotene, vitamin A or vitamin E, as evidenced by Bjelakovic et al. (2007). These findings can be partially explained by the critical role of free radicals as signaling molecules in anabolic processes. Excess antioxidants can negatively interfere with these signaling pathways, compromising essential cellular mechanisms (Higgins et al., 2020).
Reactive oxygen species (ROS) play a complex role in cancer biology. Elevated ROS levels are essential for tumor cell proliferation, survival, and progression by activating signaling pathways such as NF-κB, STAT3, and MAPK, which promote cell growth, angiogenesis, and metastasis. Paradoxically, ROS are critical for the efficacy of chemotherapy, as many anticancer treatments rely on elevating ROS to toxic levels to induce oxidative stress and trigger tumor cell death. Excessive antioxidant supplementation can disrupt this delicate balance by lowering ROS levels below the threshold necessary for effective chemotherapeutic action (Moloney and Cotter, 2018). This highlights the complex role of ROS and the potential risks of antioxidant overuse in compromising tumor control and treatment outcomes.
Resveratrol is a polyphenol in the skin of red grapes and other fruits. It has been shown to inhibit NF-κB (p65) activity and gene expression of MURF1 in the skeletal muscle of mice bearing C-26 adenocarcinoma (Shadfar et al., 2011). In mice with lung cancer and cachexia, resveratrol (20 mg/kg body weight/day, for 15 days) was able to reduce tumor mass and preserve body weight, soleus and gastrocnemius weight, myofiber cross-sectional area, and decrease Atrogin and MURF1- protein (Penedo-Vázquez et al., 2021). However, in a previous study, resveratrol administered to rats bearing the Yoshida AH-130 ascites hepatoma (1 mg/kg body weight/day for 7 days) and mice bearing the Lewis lung carcinoma (at doses of 5 and 25 mg/kg body weight/day for 15 days) was not able to preserve skeletal muscle and body weight mass (Busquets et al., 2007). Of concern, rats bearing the Yoshida AH-130 ascites hepatoma that received resveratrol showed lower food intake, lower gastrocnemius mass, heart mass, and white adipose tissue and liver weight, compared with rats bearing the tumor that did not receive resveratrol (Busquets et al., 2007).
Curcumin is a polyphenolic compound found in turmeric. It is recognized as a potent antioxidant and has been studied in the context of CC (Li et al., 2022). Patients with pancreatic cancer who received 8 g of curcumin per day for up to 18 months showed lower expression of NF-κB proteins, cyclooxygenase-2, and phosphorylated signal transducer and activator of transcription 3 in peripheral blood mononuclear cells (Dhillon et al., 2008). In a lung cancer-induced cachexia model, treatment with curcumin (1 mg/kg body weight/day for 15 days) effectively mitigated muscle wasting. Curcumin effectively preserved body weight and increased the weight of both the gastrocnemius and soleus muscles. Additionally, it enhanced the cross-sectional area of type I and type II muscle fibers, indicating reduced muscle atrophy. Furthermore, it attenuated proteolysis by reducing the levels of Atrogin-1 and MuRF-1, while also improving overall muscle structure and function (Penedo-Vázquez et al., 2021). Patients with advanced colorectal cancer received doses between 0.45 and 3.6 g per day for up to 4 months (Sharma et al., 2004). A dose of 3.6 g per day decreased the production of a lipid mediator derived from arachidonic acid, prostaglandin E2 (PGE2), in blood samples taken 1 h after the dose on days 1 and 29, compared to the PGE2 concentration found before treatment (Sharma et al., 2004).
Epigallocatechin-3-gallate (EGCG), a polyphenol derived from green tea, shows promising properties in the context of cancer and cachexia. Studies have shown that EGCG negatively regulates the expression of genes associated with Atrogin-1, a key protein in the ubiquitin-mediated protein degradation process, while also acting on other proteins of the F-BOX family, contributing to the prevention or mitigation of the progression of cachexia both in vitro and in vivo (Wang et al., 2011). Additionally, EGCG exhibits anti-inflammatory properties, which may alleviate the systemic inflammation characteristic of cancer-associated cachexia, thus reducing the overall metabolic burden of the organism (Loyala et al., 2024).
Furthermore, EGCG inhibits tumor growth by inducing apoptosis through the mitochondrial pathway, arresting the cell cycle, and modulating key signaling pathways such as EGFR/RAS/RAF/MEK/ERK. These combined effects highlight its potential as a therapeutic agent against both cancer progression and muscle wasting (Sharifi-Rad et al., 2020).
A randomized, placebo-controlled trial assessed the safety of green tea catechins (Polyphenon E®) in 97 men with high-grade prostatic intraepithelial neoplasia (HGPIN) or atypical small acinar proliferation (ASAP) over 1 year. Participants received 200 mg of EGCG twice daily with food. The supplement was well-tolerated, with no significant treatment-related adverse events, including liver toxicity, compared to placebo. Plasma EGCG levels were significantly higher in the treatment group, confirming compliance (Kumar et al., 2016). The same group also showed that the supplemented group had lower rates of prostate cancer and Gleason scores after 1 year. Notably, in men with HGPIN but not ASAP at baseline, no progression to ASAP occurred in the treatment group compared to 20% in the placebo group (Kumar et al., 2016) These findings highlight the potential of green tea catechins to slow the progression of precursor lesions without increasing the risk of high-grade disease.
Despite these potential benefits, the mechanisms of action of EGCG are not yet fully elucidated. Green tea catechins exhibit very low stability after digestion, with EGCG being particularly sensitive. Its poor bioavailability restricts its therapeutic potential; however, it can be amplified when combined with other compounds, such as vitamin C, which has been shown to boost its biological activity in experimental models (Furniturewalla and Barve, 2022).
Patients with metastatic prostate cancer who received 2000 units of vitamin D per day for 12 weeks showed an improvement in muscle strength. Among the 16 patients enrolled, half of them showed an improvement in the timed chair rises, and the other half improved timed 10-meter walk (Van Veldhuizen et al., 2000). A meta-analysis evaluated the vitamin D levels and the outcomes in patients with lung cancer (Feng et al., 2017). This study showed that circulating 25-hydroxyvitamin D concentration was inversely associated with lung cancer risk and mortality but not with overall survival (Feng et al., 2017).
Higher (4000 IU) compared to standard dose (400 IU) of vitamin D supplementation together with chemotherapy to treat head and neck cancer was not able to improve patients’ body weight, BMI, muscle area, muscle attenuation, visceral adipose tissue area, or subcutaneous adipose tissue area after the first eight cycles of chemotherapy (Brown et al., 2020).
Cancer patients received arginine associated with omega-3 fatty acids and dietary nucleotides 5 days before and after radical cystectomy. They were compared with the control group that received BOOST Plus®, a commercially available nutritional drink (Hamilton-Reeves et al., 2018). The supplementation increased arginine levels from baseline to day 30 postoperative and immune support by maintaining the Th1-Th2 balance during surgery, leading to lower plasma IL-6 levels. Despite the advantageous modulation of the inflammatory response, there were no significant differences in appendicular muscle loss between the two groups (Hamilton-Reeves et al., 2018).
Chitapanarux et al. (2020) investigated the effects of arginine, glutamine, and fish oil supplementation in 88 cancer patients with head and neck cancer (45%), esophageal cancer (32%), and cervical cancer (23%) undergoing concurrent chemoradiotherapy (CCRT). The intervention group received 250 mL of supplementation twice daily, providing 500 kcal/day with additional protein (106.25 g/day total) for 40 days on average, corresponding to the full course of CCRT. The intervention group showed significantly lower severe hematologic toxicities (5% vs 23%, p = 0.03) and improved treatment completion rates compared to the placebo group. The study highlights the potential of immune-enhancing supplements to reduce CCRT-related toxicities and improve treatment adherence. Nonetheless, it may not be the exclusive beneficial factor, as it was not an isocaloric study, and the supplemented group received additional protein and calories (500 kcal/d) from the arginine, glutamine, and fish oil supplementation.
May et al. (2002) conducted a randomized, double-blind, controlled study to assess the effects of HMB, arginine, and glutamine supplementation in patients with advanced cancer cachexia. Thirty-two patients with solid tumors received either the HMB/Arg/Gln mixture (3 g HMB, 14 g arginine, 14 g glutamine daily) or an isonitrogenous control for 24 weeks. The supplemented group experienced an increase in body weight, primarily due to gains in lean body mass, while the control group showed continued muscle loss. These improvements were maintained throughout the study, and body composition changes were validated through multiple assessment methods. The supplementation was well-tolerated, with no adverse effects reported. Although no significant changes in fat mass or quality of life were observed, the results suggest that HMB, arginine, and glutamine may help counteract muscle wasting in CC, possibly by reducing protein breakdown and enhancing protein synthesis (May et al., 2002).
Changes in muscle mass and function were evaluated in 55 pre-cachectic and cachectic patients with lung cancer, who received an oral nutritional supplement containing ≈200 kcal, 10 g whey protein, 2.0 g eicosapentaenoic acid/docosahexaenoic acid in fish oil, and 10 µg 25-hydroxy-vitamin D3 during a 12-week randomized, double-blind, controlled pilot trial. The group of patients that received the supplement did not show differences in waist and calf circumference, appendicular lean body mass, grip strength, or daily walking distance compared to the control group (Laviano et al., 2020).
Another study evaluated the effects of an oral nutritional supplement containing several compounds, including vitamin C, vitamin B5, vitamin B9, and vitamin D alongside nutritional counseling, offered to 30 patients with lung cancer under chemotherapy (Torricelli et al., 2020). Compared to the control group, the patients who received the nutritional supplement maintained or increased their body weight after 90 days and reported improvements in chemotherapy-related symptoms, including anorexia, weakness, dyspnea, pain, and hemoptysis, leading to enhanced quality of life (Torricelli et al., 2020).
Nutritional intervention strategies have been explored owing to the potential impact on mitigation of muscle loss in patients with CC (Figure 1), a multifactorial condition marked by accelerated muscle protein degradation. Most cancer patients face treatment and disease side effects, compromising food intake of calories, protein, and bioactive compounds, worsening the nutritional status and aggravating the inflammatory and catabolic state.
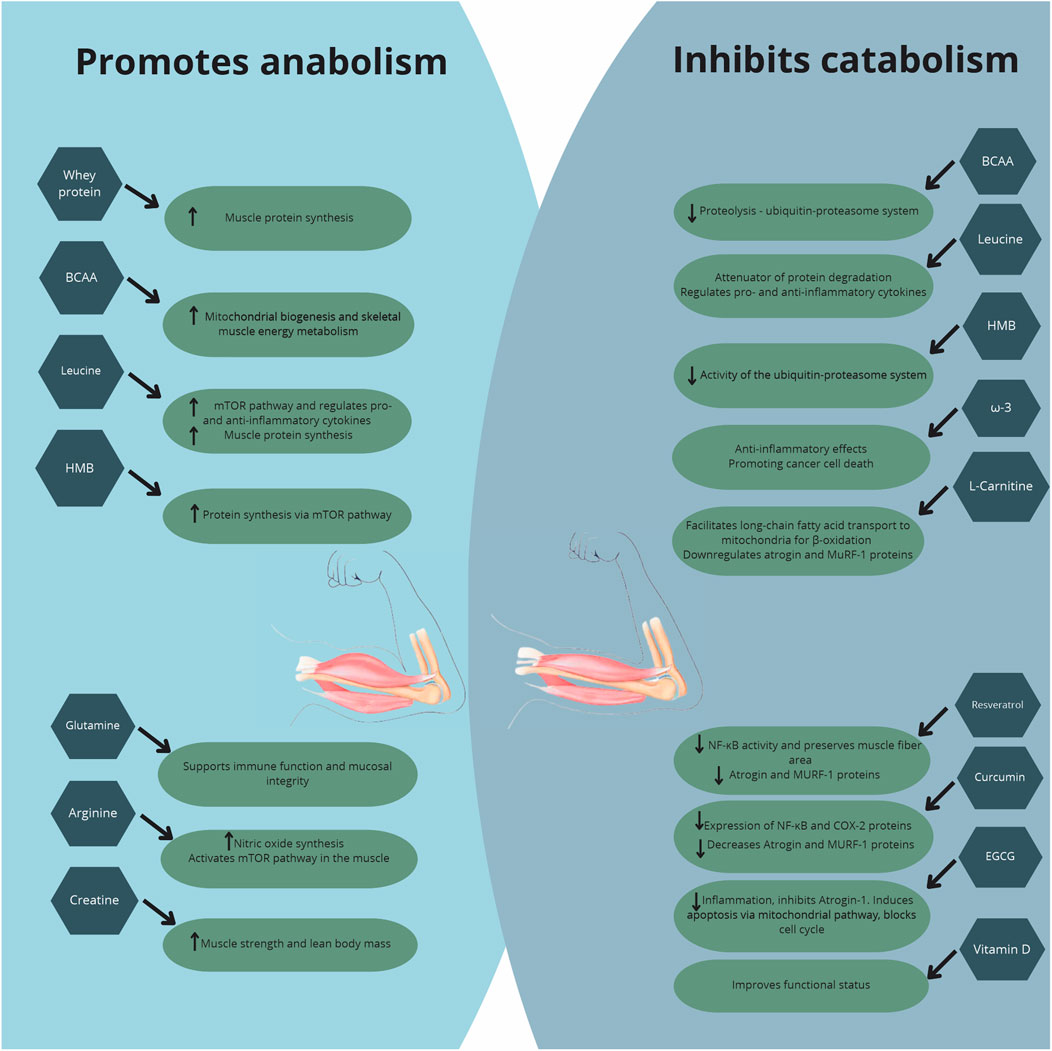
Figure 1. Major effects of nutritional supplementation in cancer cachexia.
To maximize the anabolic potential, it is crucial to detect risks of significant weight loss at the time of diagnosis and, through continuous nutritional screening, provide early nutritional intervention to prevent compromising of clinical outcome.
High-protein diets were reported to be associated with maintenance of muscle mass, while WP supplementation, a good strategy to increase protein intake, stands out for allowing rapid absorption and high plasma concentration of essential amino acids. WP improved body composition, muscle strength, and overall treatment tolerance in cancer patients undergoing chemotherapy. Additionally, HMB emerges as a promising option due to its dual action of promoting protein synthesis and inhibiting muscle degradation.
Creatine supplementation has yielded inconsistent results in human studies, with no significant improvements in muscle mass or quality of life observed in colorectal cancer patients. Despite promising results in improving lean mass and reducing fatigue in advanced cancer patients, LC requires further investigation to confirm efficacy.
The use of omega-3 fatty acids has shown contradictory results. When effectiveness was evaluated by meta-analysis, supplementation had no significant impact on inflammatory markers or muscle mass maintenance. Similarly, while antioxidants like resveratrol, curcumin, and EGCG have been explored for potential benefits, the clinical evidence in humans remained limited and inconclusive.
Combined nutrient supplementation, including arginine, omega-3 fatty acids, glutamine, and HMB, has demonstrated varying degrees of benefit in cancer patients by modulating the inflammatory response, reducing treatment-related toxicities, and preserving lean body mass. While improvements in immune function, symptom control, and quality of life were observed, the impact on muscle preservation and physical function remains inconsistent across studies.
In conclusion, the management of cancer-associated cachexia requires an individualized approach associated with nutritional counseling to manage treatment and disease side effects and to ensure feasible ways to achieve dietary requirements. An early intervention is mandatory or, at least, as soon as possible. The total protein intake matters and a dose of 1.4–2 g/kg of body weight is more effective in preventing muscle loss. When combined with other nutrients, it may improve the inflammatory profile. Further research, particularly well-designed clinical trials, is needed to determine optimal dosages and evaluate long-term outcomes, especially across different stages of cachexia and in conjunction with other therapeutic approaches to establish their full potential and clinical utility.
JF: Conceptualization, Writing–original draft, Writing–review and editing. MFA: Conceptualization, Writing–original draft, Writing–review and editing. GC: Conceptualization, Writing–original draft, Writing–review and editing. JR: Writing–original draft, Writing–review and editing. MC: Writing–original draft, Writing–review and editing. GA: Writing–original draft, Writing–review and editing. MS: Conceptualization, Supervision, Writing–original draft, Writing–review and editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. We acknowledge the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior Foundation (CAPES) and the Fundação Faculdade de Medicina for financial support. Financial support from Fundação de Amparo à Pesquisa do Estado de São Paulo, FAPESP (12/500792) was also received. MS is recipient of researcher award by CNPq 308715/2023-3. JF, MFA, JR, MC and GA are granted a research scholarship from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES)/Brazil (Process Numbers: 88887.653901/2021-00; 88887.895658/2023-00; 88887.928797/2023-00; 88887.843432/2023-00; 88887.984377/2024-00, respectively).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Alhasaniah, A. H. (2023). l-carnitine: nutrition, pathology, and health benefits. Saudi J. Biol. Sci. 30 (2), 103555. doi:10.1016/j.sjbs.2022.103555
Ananieva, E. A., Powell, J. D., and Hutson, S. M. (2016). Leucine metabolism in T cell activation: mTOR signaling and beyond. Adv. Nutr. 7 (4), 798S-805S–805S. doi:10.3945/an.115.011221
Ananieva, E. A., and Wilkinson, A. C. (2018). Branched-chain amino acid metabolism in cancer. Curr. Opin. Clin. Nutr. Metab. Care 21 (1), 64–70. doi:10.1097/MCO.0000000000000430
Antoun, S., and Raynard, B. (2018). Muscle protein anabolism in advanced cancer patients: response to protein and amino acids support, and to physical activity. Ann. Oncol. 29, ii10–7. doi:10.1093/annonc/mdx809
Armstrong, V. S., Fitzgerald, L. W., and Bathe, O. F. (2020). Cancer-associated muscle wasting—candidate mechanisms and molecular pathways. Int. J. Mol. Sci. 21 (23), 9268. doi:10.3390/ijms21239268
Aversa, Z., Pin, F., Lucia, S., Penna, F., Verzaro, R., Fazi, M., et al. (2016). Autophagy is induced in the skeletal muscle of cachectic cancer patients. Sci. Rep. 6 (1), 30340. doi:10.1038/srep30340
Azman, M., Mohd Yunus, M. R., Sulaiman, S., and Syed Omar, S. N. (2015). Enteral glutamine supplementation in surgical patients with head and neck malignancy: a randomized controlled trial. Head. Neck 37 (12), 1799–1807. doi:10.1002/hed.23839
Bjelakovic, G., Nikolova, D., Gluud, L. L., Simonetti, R. G., and Gluud, C. (2007). Mortality in randomized trials of antioxidant supplements for primary and secondary prevention: systematic review and meta-analysis. JAMA 297 (8), 842–857. doi:10.1001/jama.297.8.842
Borack, M. S., and Volpi, E. (2016). Efficacy and safety of leucine supplementation in the elderly. J. Nutr. 146 (12), 2625S-2629S–2629S. doi:10.3945/jn.116.230771
Boutière, M., Cottet-Rousselle, C., Coppard, C., Couturier, K., Féart, C., Couchet, M., et al. (2023). Protein intake in cancer: does it improve nutritional status and/or modify tumour response to chemotherapy? J. Cachexia Sarcopenia Muscle 14 (5), 2003–2015. doi:10.1002/jcsm.13276
Brestenský, M., Nitrayová, S., Patráš, P., Heger, J., and Nitray, J. (2015). Branched chain amino acids and their importance in nutrition. J. Microbiol. Biotechnol. food sci 5 (2), 197–202. doi:10.15414/jmbfs.2015.5.2.197-202
Brown, J. C., Rosenthal, M. H., Ma, C., Zhang, S., Nimeiri, H. S., McCleary, N. J., et al. (2020). Effect of high-dose vs standard-dose vitamin D3 supplementation on body composition among patients with advanced or metastatic colorectal cancer: a randomized trial. Cancers (Basel) 12 (11), 3451. doi:10.3390/cancers12113451
Buijs, N., van Bokhorst-de van der Schueren, M. A., Langius, J. A., Leemans, C. R., Kuik, D. J., Vermeulen, M. A., et al. (2010). Perioperative arginine-supplemented nutrition in malnourished patients with head and neck cancer improves long-term survival. Am. J. Clin. Nutr. 92 (5), 1151–1156. doi:10.3945/ajcn.2010.29532
Busquets, S., Fuster, G., Ametller, E., Olivan, M., Figueras, M., Costelli, P., et al. (2007). Resveratrol does not ameliorate muscle wasting in different types of cancer cachexia models. Clin. Nutr. 26 (2), 239–244. doi:10.1016/j.clnu.2006.12.001
Busquets, S., Pérez-Peiró, M., Salazar-Degracia, A., Argilés, J. M., Serpe, R., Rojano-Toimil, A., et al. (2020). Differential structural features in soleus and gastrocnemius of carnitine-treated cancer cachectic rats. J. Cell Physiol. 235 (1), 526–537. doi:10.1002/jcp.28992
Busquets, S., Serpe, R., Toledo, M., Betancourt, A., Marmonti, E., Orpí, M., et al. (2012). l-Carnitine: an adequate supplement for a multi-targeted anti-wasting therapy in cancer. Clin. Nutr. 31 (6), 889–895. doi:10.1016/j.clnu.2012.03.005
Butler, M., van der Meer, L. T., and van Leeuwen, F. N. (2021). Amino acid depletion therapies: starving cancer cells to death. Trends Endocrinol. and Metabolism 32 (6), 367–381. doi:10.1016/j.tem.2021.03.003
Camargo, C. de Q., Mocellin, M. C., Pastore Silva, J. de A., Fabre, M. E. de S., Nunes, E. A., and Trindade, E. B. S. de M. (2016). Fish oil supplementation during chemotherapy increases posterior time to tumor progression in colorectal cancer. Nutr. Cancer 68 (1), 70–76. doi:10.1080/01635581.2016.1115097
Campos-Ferraz, P. L., Gualano, B., das Neves, W., Andrade, I. T., Hangai, I., Pereira, R. T. S., et al. (2016). Exploratory studies of the potential anti-cancer effects of creatine. Amino Acids 48 (8), 1993–2001. doi:10.1007/s00726-016-2180-9
Caperuto, É. C., Tomatieli, R. V., Colquhoun, A., Seelaender, M. C. L., and Costa Rosa, LFBP (2007). Beta-hydoxy-beta-methylbutyrate supplementation affects Walker 256 tumor-bearing rats in a time-dependent manner. Clin. Nutr. 26 (1), 117–122. doi:10.1016/j.clnu.2006.05.007
Capitão, C., Coutinho, D., Neves, P. M., Capelas, M. L., Pimenta, N. M., Santos, T., et al. (2022). Protein intake and muscle mass maintenance in patients with cancer types with high prevalence of sarcopenia: a systematic review. Support. Care Cancer 30 (4), 3007–3015. doi:10.1007/s00520-021-06633-8
Cella, P. S., Marinello, P. C., Borges, F. H., Ribeiro, D. F., Chimin, P., Testa, M. T. J., et al. (2020). Creatine supplementation in Walker-256 tumor-bearing rats prevents skeletal muscle atrophy by attenuating systemic inflammation and protein degradation signaling. Eur. J. Nutr. 59 (2), 661–669. doi:10.1007/s00394-019-01933-6
Cereda, E., Turri, A., Klersy, C., Cappello, S., Ferrari, A., Filippi, A. R., et al. (2019). Whey protein isolate supplementation improves body composition, muscle strength, and treatment tolerance in malnourished advanced cancer patients undergoing chemotherapy. Cancer Med. 8 (16), 6923–6932. doi:10.1002/cam4.2517
Chitapanarux, I., Traisathit, P., Chitapanarux, T., Jiratrachu, R., Chottaweesak, P., Chakrabandhu, S., et al. (2020). Arginine, glutamine, and fish oil supplementation in cancer patients treated with concurrent chemoradiotherapy: a randomized control study. Curr. Probl. Cancer 44 (1), 100482. doi:10.1016/j.currproblcancer.2019.05.005
Chodkowska, K. A., Ciecierska, A., Majchrzak, K., Ostaszewski, P., and Sadkowski, T. (2018). Effect of β-hydroxy-β-methylbutyrate on miRNA expression in differentiating equine satellite cells exposed to hydrogen peroxide. Genes Nutr. 13 (1), 10. doi:10.1186/s12263-018-0598-2
Cruz, B., Oliveira, A., Ventrucci, G., and Gomes-Marcondes, M. C. C. (2019). A leucine-rich diet modulates the mTOR cell signalling pathway in the gastrocnemius muscle under different Walker-256 tumour growth conditions. BMC Cancer 19 (1), 349. doi:10.1186/s12885-019-5448-0
Cruzat, V., Macedo Rogero, M., Noel Keane, K., Curi, R., and Newsholme, P. (2018). Glutamine: metabolism and immune function, supplementation and clinical translation. Nutrients 10 (11), 1564. doi:10.3390/nu10111564
Dangin, M., Boirie, Y., Guillet, C., and Beaufrère, B. (2002). Influence of the protein digestion rate on protein turnover in young and elderly subjects. J. Nutr. 132 (10), 3228S-33S–3233S. doi:10.1093/jn/131.10.3228S
de Castro, G. S., Andrade, M. F., Pinto, F. C. S., Faiad, J. Z., and Seelaender, M. (2022). Omega-3 fatty acid supplementation and its impact on systemic inflammation and body weight in patients with cancer cachexia—a systematic review and meta-analysis. Front. Nutr. 8, 797513. doi:10.3389/fnut.2021.797513
Deminice, R., Cella, P. S., Padilha, C. S., Borges, F. H., da Silva, LECM, Campos-Ferraz, P. L., et al. (2016). Creatine supplementation prevents hyperhomocysteinemia, oxidative stress and cancer-induced cachexia progression in Walker-256 tumor-bearing rats. Amino Acids 48 (8), 2015–2024. doi:10.1007/s00726-016-2172-9
Deutz, N. E. P., Safar, A., Schutzler, S., Memelink, R., Ferrando, A., Spencer, H., et al. (2011a). Muscle protein synthesis in cancer patients can be stimulated with a specially formulated medical food. Clin. Nutr. 30 (6), 759–768. doi:10.1016/j.clnu.2011.05.008
Deutz, N. E. P., Safar, A., Schutzler, S., Memelink, R., Ferrando, A., Spencer, H., et al. (2011b). Muscle protein synthesis in cancer patients can be stimulated with a specially formulated medical food. Clin. Nutr. 30 (6), 759–768. doi:10.1016/j.clnu.2011.05.008
de van der Schueren, M. A. E., Laviano, A., Blanchard, H., Jourdan, M., Arends, J., and Baracos, V. E. (2018). Systematic review and meta-analysis of the evidence for oral nutritional intervention on nutritional and clinical outcomes during chemo(radio)therapy: current evidence and guidance for design of future trials. Ann. Oncol. 29 (5), 1141–1153. doi:10.1093/annonc/mdy114
Dhillon, N., Aggarwal, B. B., Newman, R. A., Wolff, R. A., Kunnumakkara, A. B., Abbruzzese, J. L., et al. (2008). Phase II trial of curcumin in patients with advanced pancreatic cancer. Clin. Cancer Res. 14 (14), 4491–4499. doi:10.1158/1078-0432.CCR-08-0024
Dickinson, J. M., Volpi, E., and Rasmussen, B. B. (2013). Exercise and nutrition to target protein synthesis impairments in aging skeletal muscle. Exerc Sport Sci. Rev. 41 (4), 216–223. doi:10.1097/JES.0b013e3182a4e699
Dierge, E., Debock, E., Guilbaud, C., Corbet, C., Mignolet, E., Mignard, L., et al. (2021). Peroxidation of n-3 and n-6 polyunsaturated fatty acids in the acidic tumor environment leads to ferroptosis-mediated anticancer effects. Cell Metab. 33 (8), 1701–1715.e5. doi:10.1016/j.cmet.2021.05.016
Dillon, E. L., Volpi, E., Wolfe, R. R., Sinha, S., Sanford, A. P., Arrastia, C. D., et al. (2007). Amino acid metabolism and inflammatory burden in ovarian cancer patients undergoing intense oncological therapy. Clin. Nutr. 26 (6), 736–743. doi:10.1016/j.clnu.2007.07.004
Donohoe, C. L., Ryan, A. M., and Reynolds, J. V. (2011). Cancer cachexia: mechanisms and clinical implications. Gastroenterol. Res. Pract. 2011, 601434–601513. doi:10.1155/2011/601434
Engelen, MPKJ, van der Meij, B. S., and Deutz, N. E. P. (2016). Protein anabolic resistance in cancer. Curr. Opin. Clin. Nutr. Metab. Care 19 (1), 39–47. doi:10.1097/MCO.0000000000000236
Ericksen, R. E., Lim, S. L., McDonnell, E., Shuen, W. H., Vadiveloo, M., White, P. J., et al. (2019). Loss of BCAA catabolism during carcinogenesis enhances mTORC1 activity and promotes tumor development and progression. Cell Metab. 29 (5), 1151–1165. doi:10.1016/j.cmet.2018.12.020
Fearon, K., Strasser, F., Anker, S. D., Bosaeus, I., Bruera, E., Fainsinger, R. L., et al. (2011). Definition and classification of cancer cachexia: an international consensus. Lancet Oncol. 12 (5), 489–495. doi:10.1016/S1470-2045(10)70218-7
Fearon, K. C. H., Glass, D. J., and Guttridge, D. C. (2012). Cancer cachexia: mediators, signaling, and metabolic pathways. Cell Metab. 16 (2), 153–166. doi:10.1016/j.cmet.2012.06.011
Feng, Q., Zhang, H., Dong, Z., Zhou, Y., and Ma, J. (2017). Circulating 25-hydroxyvitamin D and lung cancer risk and survival: a dose-response meta-analysis of prospective cohort studies. Medicine 96 (45), e8613. doi:10.1097/MD.0000000000008613
Foletta, V. C., White, L. J., Larsen, A. E., Léger, B., and Russell, A. P. (2011). The role and regulation of MAFbx/atrogin-1 and MuRF1 in skeletal muscle atrophy. Pflugers Arch. 461 (3), 325–335. doi:10.1007/s00424-010-0919-9
Fonseca, G. W. P. da, Farkas, J., Dora, E., von Haehling, S., and Lainscak, M. (2020). Cancer cachexia and related metabolic dysfunction. Int. J. Mol. Sci. 21 (7), 2321. doi:10.3390/ijms21072321
Ford, K. L., Sawyer, M. B., Ghosh, S., Trottier, C. F., Disi, I. R., Easaw, J., et al. (2024). Feasibility of two levels of protein intake in patients with colorectal cancer: findings from the Protein Recommendation to Increase Muscle (PRIMe) randomized controlled pilot trial. ESMO Open 9 (7), 103604. doi:10.1016/j.esmoop.2024.103604
Fracaro, L., Frez, F. C. V., Silva, B. C., Vicentini, G. E., de Souza, S. R. G., Martins, H. A., et al. (2016). Walker 256 tumor-bearing rats demonstrate altered interstitial cells of Cajal. Effects on ICC in the Walker 256 tumor model. Neurogastroenterol. and Motil. 28 (1), 101–115. doi:10.1111/nmo.12702
Furniturewalla, A., and Barve, K. (2022). Approaches to overcome bioavailability inconsistencies of epigallocatechin gallate, a powerful anti-oxidant in green tea. Food Chem. Adv. 1, 100037. doi:10.1016/j.focha.2022.100037
Gala, K., Desai, V., Liu, N., Omer, E. M., and McClave, S. A. (2020). How to increase muscle mass in critically ill patients: lessons learned from athletes and bodybuilders. Curr. Nutr. Rep. 9 (4), 369–380. doi:10.1007/s13668-020-00334-0
Gomes-Marcondes, M. C. C., Ventrucci, G., Toledo, M. T., Cury, L., and Cooper, J. C. (2003). A leucine-supplemented diet improved protein content of skeletal muscle in young tumor-bearing rats. Braz. J. Med. Biol. Res. 36 (11), 1589–1594. doi:10.1590/S0100-879X2003001100017
Gramignano, G., Lusso, M. R., Madeddu, C., Massa, E., Serpe, R., Deiana, L., et al. (2006). Efficacy of l-carnitine administration on fatigue, nutritional status, oxidative stress, and related quality of life in 12 advanced cancer patients undergoing anticancer therapy. Nutrition 22 (2), 136–145. doi:10.1016/j.nut.2005.06.003
Gualano, B., Roschel, H., Lancha, A. H., Brightbill, C. E., and Rawson, E. S. (2012). In sickness and in health: the widespread application of creatine supplementation. Amino Acids 43 (2), 519–529. doi:10.1007/s00726-011-1132-7
Hachiya, H., Aoki, T., Iso, Y., Shimizu, T., Tago, K., Park, K. H., et al. (2020). Effects of branched-chain amino acids on postoperative tumor recurrence in patients undergoing curative resection for hepatocellular carcinoma: a randomized clinical trial. J. Hepatobiliary Pancreat. Sci. 27 (11), 819–829. doi:10.1002/jhbp.830
Hamilton-Reeves, J. M., Stanley, A., Bechtel, M. D., Yankee, T. M., Chalise, P., Hand, L. K., et al. (2018). Perioperative immunonutrition modulates inflammatory response after radical cystectomy: results of a pilot randomized controlled clinical trial. J. Urology 200 (2), 292–301. doi:10.1016/j.juro.2018.03.001
Hao, Y., Jackson, J. R., Wang, Y., Edens, N., Pereira, S. L., and Alway, S. E. (2011). β-Hydroxy-β-methylbutyrate reduces myonuclear apoptosis during recovery from hind limb suspension-induced muscle fiber atrophy in aged rats. Am. J. Physiology-Regulatory, Integr. Comp. Physiology 301 (3), R701–R715. doi:10.1152/ajpregu.00840.2010
Harris, R. C., Söderlund, K., and Hultman, E. (1992). Elevation of creatine in resting and exercised muscle of normal subjects by creatine supplementation. Clin. Sci. 83 (3), 367–374. doi:10.1042/cs0830367
Higgins, M., Izadi, A., and Kaviani, M. (2020). Antioxidants and exercise performance: with a focus on vitamin E and C supplementation. Int. J. Environ. Res. Public Health 17 (22), 8452. doi:10.3390/ijerph17228452
Jatoi, A., Steen, P. D., Atherton, P. J., Moore, D. F., Rowland, K. M., Le-Lindqwister, N. A., et al. (2017). A double-blind, placebo-controlled randomized trial of creatine for the cancer anorexia/weight loss syndrome (N02C4): an Alliance trial. Ann. Oncol. 28 (8), 1957–1963. doi:10.1093/annonc/mdx232
Johal, J., Han, C. Y., Joseph, R., Munn, Z., Agbejule, O. A., Crawford-Williams, F., et al. (2022). Dietary supplements in people with metastatic cancer who are experiencing malnutrition, cachexia, sarcopenia, and frailty: a scoping review. Nutrients 14 (13), 2642. doi:10.3390/nu14132642
Jung, M. K., Okekunle, A. P., Lee, J. E., Sung, M. K., and Lim, Y. J. (2021). Role of branched-chain amino acid metabolism in tumor development and progression. J. Cancer Prev. 26 (4), 237–243. doi:10.15430/JCP.2021.26.4.237
Jung, S., Bae, Y. S., Kim, H. J., Jayasena, D. D., Lee, J. H., Park, H. B., et al. (2013). Carnosine, anserine, creatine, and inosine 5′-monophosphate contents in breast and thigh meats from 5 lines of Korean native chicken. Poult. Sci. 92 (12), 3275–3282. doi:10.3382/ps.2013-03441
Katagiri, R., Song, M., Zhang, X., Lee, D. H., Tabung, F. K., Fuchs, C. S., et al. (2020). Dietary intake of branched-chain amino acids and risk of colorectal cancer. Cancer. Cancer Prev. Res. 13 (1), 65–72. doi:10.1158/1940-6207.CAPR-19-0297
Kraft, M., Kraft, K., Gärtner, S., Mayerle, J., Simon, P., Weber, E., et al. (2012). L-Carnitine-supplementation in advanced pancreatic cancer (CARPAN) - a randomized multicentre trial. Nutr. J. 11 (1), 52. doi:10.1186/1475-2891-11-52
Kumar, N. B., Pow-Sang, J., Spiess, P. E., Park, J., Salup, R., Williams, C. R., et al. (2016). Randomized, placebo-controlled trial evaluating the safety of one-year administration of green tea catechins. Oncotarget 7 (43), 70794–70802. doi:10.18632/oncotarget.12222
Lam, C. N., Watt, A. E., Isenring, E. A., de van der Schueren, M. A. E., and van der Meij, B. S. (2021). The effect of oral omega-3 polyunsaturated fatty acid supplementation on muscle maintenance and quality of life in patients with cancer: a systematic review and meta-analysis. Clin. Nutr. 40 (6), 3815–3826. doi:10.1016/j.clnu.2021.04.031
Laviano, A., Calder, P. C., Schols, AMWJ, Lonnqvist, F., Bech, M., and Muscaritoli, M. (2020). Safety and tolerability of targeted medical nutrition for cachexia in non-small-cell lung cancer: a randomized, double-blind, controlled pilot trial. Nutr. Cancer 72 (3), 439–450. doi:10.1080/01635581.2019.1634746
Lei, M. Z., Li, X. X., Zhang, Y., Li, J. T., Zhang, F., Wang, Y. P., et al. (2020). Acetylation promotes BCAT2 degradation to suppress BCAA catabolism and pancreatic cancer growth. Signal Transduct. Target Ther. 5 (1), 70. doi:10.1038/s41392-020-0168-0
Lerner, L., Hayes, T. G., Tao, N., Krieger, B., Feng, B., Wu, Z., et al. (2015). Plasma growth differentiation factor 15 is associated with weight loss and mortality in cancer patients. J. Cachexia Sarcopenia Muscle 6 (4), 317–324. doi:10.1002/jcsm.12033
Li, P., Yin, Y. L., Li, D., Woo Kim, S., and Wu, G. (2007). Amino acids and immune function. Br. J. Nutr. 98 (2), 237–252. doi:10.1017/S000711450769936X
Li, Y., Li, S., and Wu, H. (2022). Ubiquitination-proteasome system (UPS) and autophagy two main protein degradation machineries in response to cell stress. Cells 11 (5), 851. doi:10.3390/cells11050851
Lieu, E. L., Nguyen, T., Rhyne, S., and Kim, J. (2020). Amino acids in cancer. Exp. Mol. Med. 52 (1), 15–30. doi:10.1038/s12276-020-0375-3
Liput, K. P., Lepczyński, A., Ogłuszka, M., Nawrocka, A., Poławska, E., Grzesiak, A., et al. (2021). Effects of dietary n–3 and n–6 polyunsaturated fatty acids in inflammation and cancerogenesis. Int. J. Mol. Sci. 22 (13), 6965. doi:10.3390/ijms22136965
Liu, S., Wu, H. J., Zhang, Z. Q., Chen, Q., Liu, B., Wu, J. P., et al. (2011). L-carnitine ameliorates cancer cachexia in mice by regulating the expression and activity of carnitine palmityl transferase. Cancer Biol. Ther. 12 (2), 125–130. doi:10.4161/cbt.12.2.15717
Longo, N., Frigeni, M., and Pasquali, M. (2016). Carnitine transport and fatty acid oxidation. Biochimica Biophysica Acta (BBA) - Mol. Cell Res. 1863 (10), 2422–2435. doi:10.1016/j.bbamcr.2016.01.023
Loyala, J. V., Down, B., Wong, E., and Tan, B. (2024). Treatment of cachexia in gastric cancer: exploring the use of anti-inflammatory natural products and their derivatives. Nutrients 16 (8), 1246. doi:10.3390/nu16081246
Mantovani, G., Macciò, A., Madeddu, C., Serpe, R., Massa, E., Dessì, M., et al. (2010). Randomized phase III clinical trial of five different arms of treatment in 332 patients with cancer cachexia. Oncologist 15 (2), 200–211. doi:10.1634/theoncologist.2009-0153
Martin, A., Gallot, Y. S., and Freyssenet, D. (2023). Molecular mechanisms of cancer cachexia-related loss of skeletal muscle mass: data analysis from preclinical and clinical studies. J. Cachexia Sarcopenia Muscle 14 (3), 1150–1167. doi:10.1002/jcsm.13073
Mattox, T. W. (2017). Cancer cachexia: cause, diagnosis, and treatment. Nutr. Clin. Pract. 32 (5), 599–606. doi:10.1177/0884533617722986
May, P. E., Barber, A., D’Olimpio, J. T., Hourihane, A., and Abumrad, N. N. (2002). Reversal of cancer-related wasting using oral supplementation with a combination of beta-hydroxy-beta-methylbutyrate, arginine, and glutamine. Am. J. Surg. 183 (4), 471–479. doi:10.1016/s0002-9610(02)00823-1
McCarty, M. F. (1996). Fish oil may impede tumour angiogenesis and invasiveness by down-regulating protein kinase C and modulating eicosanoid production. Med. Hypotheses 46 (2), 107–115. doi:10.1016/S0306-9877(96)90009-2
Mir Khan, U., and Selamoglu, Z. (2020). Nutritional and medical perspectives of whey protein: a historical overview. J. Pharmac. Care. 7 (4), 112–117. doi:10.18502/jpc.v7i4.2380
Moloney, J. N., and Cotter, T. G. (2018). ROS signalling in the biology of cancer. Semin. Cell Dev. Biol. 80, 50–64. doi:10.1016/j.semcdb.2017.05.023
Muranaka, H., Akinsola, R., Billet, S., Pandol, S. J., Hendifar, A. E., Bhowmick, N. A., et al. (2024). Glutamine supplementation as an anticancer strategy: a potential therapeutic alternative to the convention. Cancers (Basel) 16 (5), 1057. doi:10.3390/cancers16051057
Muscaritoli, M., Arends, J., Bachmann, P., Baracos, V., Barthelemy, N., Bertz, H., et al. (2021). ESPEN practical guideline: clinical Nutrition in cancer. Clin. Nutr. 40 (5), 2898–2913. doi:10.1016/j.clnu.2021.02.005
Newsholme, P., Lima, M. M. R., Procopio, J., Pithon-Curi, T. C., Doi, S. Q., Bazotte, R. B., et al. (2003). Glutamine and glutamate as vital metabolites. Braz. J. Med. Biol. Res. 36 (2), 153–163. doi:10.1590/S0100-879X2003000200002
Nojiri, S., Fujiwara, K., Shinkai, N., Iio, E., and Joh, T. (2017). Effects of branched-chain amino acid supplementation after radiofrequency ablation for hepatocellular carcinoma: a randomized trial. Nutrition 33, 20–27. doi:10.1016/j.nut.2016.07.013
Norman, K., Stübler, D., Baier, P., Schütz, T., Ocran, K., Holm, E., et al. (2006). Effects of creatine supplementation on nutritional status, muscle function and quality of life in patients with colorectal cancer—a double blind randomised controlled trial. Clin. Nutr. 25 (4), 596–605. doi:10.1016/j.clnu.2006.01.014
Nunes, E. A., Kuczera, D., Brito, G. A. P., Bonatto, S. J. R., Yamazaki, R. K., Tanhoffer, R. A., et al. (2008). Beta-hydroxy-beta-methylbutyrate supplementation reduces tumor growth and tumor cell proliferation ex vivo and prevents cachexia in Walker 256 tumor-bearing rats by modifying nuclear factor-kappaB expression. Nutr. Res. 28 (7), 487–493. doi:10.1016/j.nutres.2008.04.006
Orellana López, C., Leyton Estéfane, J., Ramos Rosales, M., Vásquez Ramirez, C., Manriquez Arriagada, C., Argilés, J. M., et al. (2023). Prevalence of cachexia in cancer patients. Eur. J. Cancer Care (Engl) 2023, 1–9. doi:10.1155/2023/5743872
Panwar, V., Singh, A., Bhatt, M., Tonk, R. K., Azizov, S., Raza, A. S., et al. (2023). Multifaceted role of mTOR (mammalian target of rapamycin) signaling pathway in human health and disease. Signal Transduct. Target Ther. 8 (1), 375. doi:10.1038/s41392-023-01608-z
Penedo-Vázquez, A., Duran, X., Mateu, J., López-Postigo, A., and Barreiro, E. (2021). Curcumin and resveratrol improve muscle function and structure through attenuation of proteolytic markers in experimental cancer-induced cachexia. Molecules 26 (16), 4904. doi:10.3390/molecules26164904
Peng, H., Wang, Y., and Luo, W. (2020). Multifaceted role of branched-chain amino acid metabolism in cancer. Oncogene 39 (44), 6747–6756. doi:10.1038/s41388-020-01480-z
Peters, S. J., van Helvoort, A., Kegler, D., Argilès, J. M., Luiking, Y. C., Laviano, A., et al. (2011). Dose-dependent effects of leucine supplementation on preservation of muscle mass in cancer cachectic mice. Oncol. Rep. 26 (1), 247–254. doi:10.3892/or.2011.1269
Pradhan, R., Dieterich, W., Natarajan, A., Schwappacher, R., Reljic, D., Herrmann, H. J., et al. (2024). Influence of amino acids and exercise on muscle protein turnover, particularly in cancer cachexia. Cancers (Basel) 16 (10), 1921. doi:10.3390/cancers16101921
Prado, C. M., Orsso, C. E., Pereira, S. L., Atherton, P. J., and Deutz, N. E. P. (2022). Effects of β-hydroxy β-methylbutyrate (HMB) supplementation on muscle mass, function, and other outcomes in patients with cancer: a systematic review. J. Cachexia, Sarcopenia Muscle 13, 1623–1641. doi:10.1002/jcsm.12952
Prado, C. M., Purcell, S. A., and Laviano, A. (2020). Nutrition interventions to treat low muscle mass in cancer. J. Cachexia Sarcopenia Muscle 11 (2), 366–380. doi:10.1002/jcsm.12525
Prado, C. M., Sawyer, M. B., Ghosh, S., Lieffers, J. R., Esfandiari, N., Antoun, S., et al. (2013). Central tenet of cancer cachexia therapy: do patients with advanced cancer have exploitable anabolic potential? Am. J. Clin. Nutr. 98 (4), 1012–1019. doi:10.3945/ajcn.113.060228
Puig-Vilanova, E., Rodriguez, D. A., Lloreta, J., Ausin, P., Pascual-Guardia, S., Broquetas, J., et al. (2015). Oxidative stress, redox signaling pathways, and autophagy in cachectic muscles of male patients with advanced COPD and lung cancer. Free Radic. Biol. Med. 79, 91–108. doi:10.1016/j.freeradbiomed.2014.11.006
Ramani, A., Hazra, T., Mudgil, S., and Mudgil, D. (2024). Emerging potential of whey proteins in prevention of cancer. Food Humanity 2, 100199. doi:10.1016/j.foohum.2023.12.007
Rasmussen, B., Gilbert, E., Turki, A., Madden, K., and Elango, R. (2016). Determination of the safety of leucine supplementation in healthy elderly men. Amino Acids 48 (7), 1707–1716. doi:10.1007/s00726-016-2241-0
Rossi, A. P., D’Introno, A., Rubele, S., Caliari, C., Gattazzo, S., Zoico, E., et al. (2017). The potential of β-Hydroxy-β-Methylbutyrate as a new strategy for the management of sarcopenia and sarcopenic obesity. Drugs Aging 34 (11), 833–840. doi:10.1007/s40266-017-0496-0
Salomão, E. M., Toneto, A. T., Silva, G. O., and Gomes-Marcondes, M. C. C. (2010). Physical exercise and a leucine-rich diet modulate the muscle protein metabolism in walker tumor-bearing rats. Nutr. Cancer 62 (8), 1095–1104. doi:10.1080/01635581.2010.492082
Sandri, M. (2013). Protein breakdown in muscle wasting: role of autophagy-lysosome and ubiquitin-proteasome. Int. J. Biochem. Cell Biol. 45 (10), 2121–2129. doi:10.1016/j.biocel.2013.04.023
Setiawan, T., Sari, I. N., Wijaya, Y. T., Julianto, N. M., Muhammad, J. A., Lee, H., et al. (2023). Cancer cachexia: molecular mechanisms and treatment strategies. J. Hematol. Oncol. 16 (1), 54. doi:10.1186/s13045-023-01454-0
Shadfar, S., Couch, M. E., McKinney, K. A., Weinstein, L. J., Yin, X., Rodríguez, J. E., et al. (2011). Oral resveratrol therapy inhibits cancer-induced skeletal muscle and cardiac atrophy in vivo. Nutr. Cancer 63 (5), 749–762. doi:10.1080/01635581.2011.563032
Sharifi-Rad, M., Pezzani, R., Redaelli, M., Zorzan, M., Imran, M., Ahmed, K. A., et al. (2020). Preclinical pharmacological activities of epigallocatechin-3-gallate in signaling pathways: an update on cancer. Molecules 25 (3), 467. doi:10.3390/molecules25030467
Sharma, R. A., Euden, S. A., Platton, S. L., Cooke, D. N., Shafayat, A., Hewitt, H. R., et al. (2004). Phase I clinical trial of oral curcumin: biomarkers of systemic activity and compliance. Clin. Cancer Res. 10 (20), 6847–6854. doi:10.1158/1078-0432.CCR-04-0744
Silvério, R., Laviano, A., Fanelli, F. R., and Seelaender, M. (2012). l-Carnitine induces recovery of liver lipid metabolism in cancer cachexia. Amino Acids 42 (5), 1783–1792. doi:10.1007/s00726-011-0898-y
Soares, J. D. P., Howell, S. L., Teixeira, F. J., and Pimentel, G. D. (2020). Dietary amino acids and immunonutrition supplementation in cancer-induced skeletal muscle mass depletion: a mini-review. Curr. Pharm. Des. 26 (9), 970–978. doi:10.2174/1381612826666200218100420
Stipanuk, M. H. (2008). Leucine and protein synthesis: mTOR and beyond. Nutr. Rev. 65 (3), 122–129. doi:10.1111/j.1753-4887.2007.tb00289.x
Storck, L. J., Ruehlin, M., Gaeumann, S., Gisi, D., Schmocker, M., Meffert, P. J., et al. (2020). Effect of a leucine-rich supplement in combination with nutrition and physical exercise in advanced cancer patients: a randomized controlled intervention trial. Clin. Nutr. 39 (12), 3637–3644. doi:10.1016/j.clnu.2020.04.008
Suzuki, H., Mitsunaga, S., Ikeda, M., Aoyama, T., Yoshizawa, K., Yoshimatsu, H., et al. (2021). Clinical and tumor characteristics of patients with high serum levels of growth differentiation factor 15 in advanced pancreatic cancer. Cancers (Basel) 13 (19), 4842. doi:10.3390/cancers13194842
Taherizadeh, M., Khoshnia, M., Shams, S., Hesari, Z., and Joshaghani, H. (2021). Plasma changes of branched-chain amino acid in patients with esophageal cancer. Middle East J. Dig. Dis. 13 (1), 49–53. doi:10.34172/mejdd.2021.203
Tanaka, K., Nakamura, S., and Narimatsu, H. (2022). Nutritional approach to cancer cachexia: a proposal for dietitians. Nutrients 14 (2), 345. doi:10.3390/nu14020345
Tang, H., Pang, S., Wang, M., Xiao, X., Rong, Y., Wang, H., et al. (2010). TLR4 activation is required for IL-17-induced multiple tissue inflammation and wasting in mice. J. Immunol. 185 (4), 2563–2569. doi:10.4049/jimmunol.0903664
Teixeira, F. J., Santos, H. O., Howell, S. L., and Pimentel, G. D. (2019). Whey protein in cancer therapy: a narrative review. Pharmacol. Res. 144, 245–256. doi:10.1016/j.phrs.2019.04.019
Thomson, A. W., Turnquist, H. R., and Raimondi, G. (2009). Immunoregulatory functions of mTOR inhibition. Nat. Rev. Immunol. 9 (5), 324–337. doi:10.1038/nri2546
Tian, T., Li, X., and Zhang, J. (2019). mTOR signaling in cancer and mTOR inhibitors in solid tumor targeting therapy. Int. J. Mol. Sci. 20 (3), 755. doi:10.3390/ijms20030755
Torricelli, P., Antonelli, F., Ferorelli, P., Borromeo, I., Shevchenko, A., Lenzi, S., et al. (2020). Oral nutritional supplement prevents weight loss and reduces side effects in patients in advanced lung cancer chemotherapy. Amino Acids 52 (3), 445–451. doi:10.1007/s00726-020-02822-7
Valerio, A., D’Antona, G., and Nisoli, E. (2011). Branched-chain amino acids, mitochondrial biogenesis, and healthspan: an evolutionary perspective. Aging 3 (5), 464–478. doi:10.18632/aging.100322
Van Bokhorst-de van der Schueren, M. A., Quak, J. J., von Blomberg-van der Flier, B. M. E., Kuik, D. J., Langendoen, S. I., Snow, G. B., et al. (2001). Effect of perioperative nutrition, with and without arginine supplementation, on nutritional status, immune function, postoperative morbidity, and survival in severely malnourished head and neck cancer patients. Am. J. Clin. Nutr. 73 (2), 323–332. doi:10.1093/ajcn/73.2.323
Van der Ende, M., Grefte, S., Plas, R., Meijerink, J., Witkamp, R. F., Keijer, J., et al. (2018). Mitochondrial dynamics in cancer-induced cachexia. Biochimica Biophysica Acta (BBA) - Rev. Cancer 1870 (2), 137–150. doi:10.1016/j.bbcan.2018.07.008
Van der Meij, B. S., Teleni, L., Engelen, MPKJ, and Deutz, N. E. P. (2019). Amino acid kinetics and the response to nutrition in patients with cancer. Int. J. Radiat. Biol. 95 (4), 480–492. doi:10.1080/09553002.2018.1466209
Van de Worp, WRPH, Schols, AMWJ, Theys, J., van Helvoort, A., and Langen, R. C. J. (2020). Nutritional interventions in cancer cachexia: evidence and perspectives from experimental models. Front. Nutr. 7, 601329. doi:10.3389/fnut.2020.601329
van Dijk, D. P., van de Poll, M. C., Moses, A. G., Preston, T., Olde Damink, S. W., Rensen, S. S., et al. (2015). Effects of oral meal feeding on whole body protein breakdown and protein synthesis in cachectic pancreatic cancer patients. J. Cachexia Sarcopenia Muscle 6 (3), 212–221. doi:10.1002/jcsm.12029
Van Veldhuizen, P. J., Taylor, S. A., Williamson, S., and Drees, B. M. (2000). Treatment of vitamin d deficiency in patients with metastatic prostate cancer may improve bone pain and muscle strength. J. Urology 163 (1), 187–190. doi:10.1097/00005392-200001000-00044
Viana, L. R., Canevarolo, R., Luiz, A. C. P., Soares, R. F., Lubaczeuski, C., Zeri, A. C. de M., et al. (2016). Leucine-rich diet alters the 1H-NMR based metabolomic profile without changing the Walker-256 tumour mass in rats. BMC Cancer 16 (1), 764. doi:10.1186/s12885-016-2811-2
Vissers, Y. L., Dejong, C. H., Luiking, Y. C., Fearon, K. C., von Meyenfeldt, M. F., and Deutz, N. E. (2005). Plasma arginine concentrations are reduced in cancer patients: evidence for arginine deficiency? Am. J. Clin. Nutr. 81 (5), 1142–1146. doi:10.1093/ajcn/81.5.1142
von Haehling, S., and Anker, S. D. (2014). Prevalence, incidence and clinical impact of cachexia: facts and numbers—update 2014. J. Cachexia Sarcopenia Muscle 5 (4), 261–263. doi:10.1007/s13539-014-0164-8
Wang, B., Pei, J., Xu, S., Liu, J., and Yu, J. (2024). A glutamine tug-of-war between cancer and immune cells: recent advances in unraveling the ongoing battle. J. Exp. and Clin. Cancer Res. 43 (1), 74. doi:10.1186/s13046-024-02994-0
Wang, H., Lai, Y. J., Chan, Y. L., Li, T. L., and Wu, C. J. (2011). Epigallocatechin-3-gallate effectively attenuates skeletal muscle atrophy caused by cancer cachexia. Cancer Lett. 305 (1), 40–49. doi:10.1016/j.canlet.2011.02.023
Wang, P., Wu, S., Zeng, X., Zhang, Y., Zhou, Y., Su, L., et al. (2018). BCAT1 promotes proliferation of endometrial cancer cells through reprogrammed BCAA metabolism. Int. J. Clin. Exp. Pathol. 11 (12), 5536–5546.
Wei, L., Wang, R., Lin, K., Jin, X., Li, L., Wazir, J., et al. (2022). Creatine modulates cellular energy metabolism and protects against cancer cachexia-associated muscle wasting. Front. Pharmacol. 13, 1086662. doi:10.3389/fphar.2022.1086662
Wiegert, E. V. M., de Oliveira, L. C., Calixto-Lima, L., Mota e Silva Lopes, M. S. da, and Peres, W. A. F. (2020). Cancer cachexia: comparing diagnostic criteria in patients with incurable cancer. Nutrition 79-80, 110945–110980. doi:10.1016/j.nut.2020.110945
Williams, J. Z., Abumrad, N., and Barbul, A. (2002). Effect of a specialized amino acid mixture on human collagen deposition. Ann. Surg. 236 (3), 369–374. doi:10.1097/00000658-200209000-00013
Wu, G., Meininger, C. J., McNeal, C. J., Bazer, F. W., and Rhoads, J. M. (2021). Role of L-arginine in nitric oxide synthesis and health in humans. Adv. Exp. Med. Biol. 1332, 167–187. doi:10.1007/978-3-030-74180-8_10
Xia, Z., Cholewa, J., Zhao, Y., Shang, H. Y., Yang, Y. Q., Araújo Pessôa, K., et al. (2017). Targeting inflammation and downstream protein metabolism in sarcopenia: a brief up-dated description of concurrent exercise and leucine-based multimodal intervention. Front. Physiol. 8, 434. doi:10.3389/fphys.2017.00434
Yamada, A. K., Ferretti, R., Matsumura, C. Y., Antunes, L., Silva, C. A. da, and Pertille, A. (2022). Beta-hydroxy-beta-methylbutyrate associated with low-intensity exercise training improves skeletal muscle regeneration through the IGF-Akt pathway. Braz. J. Med. Biol. Res. 55, e11597. doi:10.1590/1414-431x2021e11597
Yoshida, T., and Delafontaine, P. (2020). Mechanisms of IGF-1-mediated regulation of skeletal muscle hypertrophy and atrophy. Cells 9 (9), 1970. doi:10.3390/cells9091970
Zaira, A., Andrea, B., Paola, C., Valerio, G. M., Fabio, P., Francesco, M. B., et al. (2011). Muscaritoli. β-hydroxy-β-methylbutyrate (HMB) attenuates muscle and body weight loss in experimental cancer cachexia. Int. J. Oncol. 38(3), 713–20. doi:10.3892/ijo.2010.885
Zanchi, N. E., Gerlinger-Romero, F., Guimarães-Ferreira, L., de Siqueira Filho, M. A., Felitti, V., Lira, F. S., et al. (2011). HMB supplementation: clinical and athletic performance-related effects and mechanisms of action. Amino Acids 40 (4), 1015–1025. doi:10.1007/s00726-010-0678-0
Zanetti, M., Gortan Cappellari, G., Barazzoni, R., and Sanson, G. (2020). The impact of protein supplementation targeted at improving muscle mass on strength in cancer patients: a scoping review. Nutrients 12 (7), 2099. doi:10.3390/nu12072099
Zhang, S., Zeng, X., Ren, M., Mao, X., and Qiao, S. (2017). Novel metabolic and physiological functions of branched chain amino acids: a review. J. Anim. Sci. Biotechnol. 8 (1), 10. doi:10.1186/s40104-016-0139-z
Zhao, C., Chen, N., and Ashaolu, T. J. (2022). Whey proteins and peptides in health-promoting functions – a review. Int. Dairy J. 126, 105269. doi:10.1016/j.idairyj.2021.105269
Ziętarska, M., Krawczyk-Lipiec, J., Kraj, L., Zaucha, R., and Małgorzewicz, S. (2017). Chemotherapy-related toxicity, nutritional status and quality of life in precachectic oncologic patients with, or without, high protein nutritional support. A prospective, randomized study. Nutrients 9 (10), 1108. doi:10.3390/nu9101108
Keywords: nutritional supplementation, cancer cachexia, muscle wasting, chronic inflammation, protein synthesis
Citation: Faiad J, Andrade MF, de Castro G, de Resende J, Coêlho M, Aquino G and Seelaender M (2025) Muscle loss in cancer cachexia: what is the basis for nutritional support?. Front. Pharmacol. 16:1519278. doi: 10.3389/fphar.2025.1519278
Received: 29 October 2024; Accepted: 27 January 2025;
Published: 26 February 2025.
Edited by:
Maurizio Ragni, University of Milan, ItalyCopyright © 2025 Faiad, Andrade, de Castro, de Resende, Coêlho, Aquino and Seelaender. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marilia Seelaender, c2VlbGFlbmRlckB1c3AuYnI=
†These authors share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.