 Diana Carreño
Diana Carreño Antonella Facundo
Antonella Facundo Anjelica Cardenas
Anjelica Cardenas Shahrdad Lotfipour
Shahrdad Lotfipour- 1Department of Pharmaceutical Sciences, University of California, Irvine, Irvine, CA, United States
- 2Department of Emergency Medicine, University of California, Irvine, Irvine, CA, United States
- 3Department of Radiation Oncology, David Geffen School of Medicine, University of California, Los Angeles, Los Angeles, CA, United States
- 4Department of Pathology and Laboratory Medicine, University of California, Irvine, Irvine, CA, United States
Introduction: The rewarding effects of drugs of abuse are associated with the dopaminergic system in the limbic circuitry. Nicotine exposure during adolescence is linked to increased use of drugs of abuse with nicotine and methamphetamine (METH) commonly used together. Nicotine acts on neuronal nicotinic acetylcholine receptor (nAChR) systems, critical for reward processing and drug reinforcement, while METH leads to a higher dopamine (DA) efflux in brain reward regions. A human single nucleotide polymorphism (SNP) in the 3′-untranslated region (UTR) of the α6 nicotinic receptor subunit gene (CHRNA6, rs2304297), has been linked with tobacco/nicotine and general substance use during adolescence. Using CRISPR-Cas9 genomic engineering, our lab recapitulated the CHRNA6 3′UTRC123G SNP, generating α6CC and α6GG allele carriers in Sprague Dawley rats. We hypothesized the CHRNA6 3′UTRC123G SNP would sex- and genotype-dependently enhance nicotine-induced METH self-administration as well as nicotine-induced DA overflow in the nucleus accumbens shell of adolescent α6GG and α6CC carriers.
Methods: Adolescent male and female rats underwent a 4-day sub-chronic, low-dose (0.03 mg/kg/0.1 mL, x2) nicotine pretreatment paradigm to assess intravenous METH (0.02 mg/kg/0.1 mL) self-administration as well as nicotine- and METH (0.02 mg/kg/0.1 mL)-induced DA overflow in the nucleus accumbens shell (NAcS) using in vivo microdialysis coupled with high-performance liquid-chromatography-electrochemical detection (HPLC-ECD).
Results: Nicotine pretreatment sex- and genotype-dependently enhanced subsequent METH self-administration in adolescent CHRNA6 3′UTRC123G SNP rats. Further nicotine and METH-induced DA overflow is observed in α6CC females as compared to α6GG females, with METH-induced DA overflow enhanced in α6GG males when compared to α6CC males.
Conclusion: These findings demonstrate that the CHRNA6 3′-UTRC123G SNP can sex- and genotype-dependently impact adolescent nicotine-induced effects on METH self-administration and stimulant-induced DA overflow in reward regions of the brain.
1 Introduction
Adolescence is a critical period of development characterized by heightened susceptibility to environmental influences, including substance use. Nicotine and methamphetamine (METH) are two commonly abused substances that can have profound, long lasting consequences on the developing brain. Adolescents predominantly favor electronic cigarettes as their primary choice among tobacco products (Gorukanti et al., 2017). In 2023, the Annual National Youth Tobacco Survey reported more than one in four of current youth e-cigarette users use an e-cigarette product every day (Birdsey et al., 2023). Most e-cigarettes contain nicotine, a highly addictive substance that can harm the developing brain of adolescents (Goriounova and Mansvelder, 2012; Yuan et al., 2015). Adolescent exposure to nicotine may lead to the use of other tobacco products and drugs of abuse and amplify common underlying risk factors, i.e., genetic predisposition, peer influence, socioeconomic status and family history (McCabe et al., 2018; Berry et al., 2019; Ren and Lotfipour, 2019).
The neurobiological actions of nicotine are similar to other psychomotor stimulants including cocaine and METH (Laviolette and van der Kooy, 2004). Nicotine binds to nAChRs, leading to increased dopamine (DA) release and neurotransmitter modulation, resulting in enhanced alertness, reward and potential dependence (Di Chiara and Imperato, 1988; Albuquerque et al., 2009; Dani, 2015). METH is a potent analog of the stimulant, amphetamine. METH increases DA levels by inhibiting the reuptake and promoting release of DA, causing intense euphoria, heightened alertness, and potential neurotoxicity with chronic use (Rothman et al., 2001; Elkashef et al., 2008; Rusyniak, 2011; Ferrucci et al., 2019). It has been hypothesized that nicotine addiction is mediated by DA release in the mesolimbic system via local action at somatodendritic sites in the ventral tegmental area (VTA) (Di Chiara and Imperato, 1988; Nisell et al., 1994). Indeed, injections of 6-hydroxydopamine, a neurotoxin used to selectively destroy dopaminergic and noradrenergic neurons, into the nucleus accumbens (NAc) blocked nicotine self-administration in adult rats (Corrigall et al., 1994). In vitro studies have shown that METH evokes greater efflux of DA via the dopamine transporter (DAT) than its counterpart, amphetamine (Goodwin et al., 2009). In vivo studies have shown that nicotine pretreatment enhances the acquisition of METH self-administration and increases its intake in adolescent male, but not female rats (Dao et al., 2011; Cardenas and Lotfipour, 2022). Additionally, repeated administration of METH and nicotine in mice produced locomotor sensitization effects and a symmetrical cross-sensitization (Kuriban, 1999).
During adolescence, the α6 nAChR subunit reaches peak mRNA expression in dopaminergic cell bodies in the VTA and substantia nigra (SNg) (Azam et al., 2007). Moreover, the α6 nAChR subunit is localized in the mesolimbic and nigrostriatal DA pathway (Le Novère et al., 2002; Azam et al., 2007; Jackson et al., 2009) which may indicate a role of α6*-containing nAChRs in nicotine-induced behaviors and DA release (*denotes other associated nAChR subunits) (Le Novère et al., 2002; Azam et al., 2007; Pons et al., 2008; Jackson et al., 2009; Zoli et al., 2015). In vivo studies have demonstrated the α6* nAChRs have been shown to be involved in nicotine-induced locomotion (Drenan et al., 2008; Cardenas et al., 2022), nicotine self-administration and drug-seeking behavior (Pons et al., 2008; Brunzell et al., 2010; Carreño and Lotfipour, 2023; Carreño et al., 2024). Nevertheless, there is a scarcity of studies that examine the impact of α6* nAChRs in adolescent nicotine-induced behaviors. Despite this gap in research, it is important to note a robust correlation between tobacco/nicotine intake during adolescence and subsequent METH use (Brecht et al., 2007; Russell et al., 2008). Repeated self-administration of METH upregulated the CHRNA6 mRNA in the VTA of adult male rats (Bosch et al., 2015). Given these results, exploring how nicotine affects METH self-administration and the underlying mechanisms for how nicotine and METH exposure impact DA overflow in the NAc shell in adolescent rodent lines carrying a human CHRNA6 3′-UTR SNP becomes crucial.
To model adolescent initiation of nicotine smoking behavior in rats, a low-dose 4-day intravenous nicotine pretreatment paradigm was used. Previously, this paradigm has shown nicotine-induced acquisition of subsequent cocaine, fentanyl, METH, and alcohol in adolescent rats. (McQuown et al., 2007; Dao et al., 2011; Linker et al., 2020; Cardenas et al., 2021). Following nicotine treatment, animals underwent 2-h operant intravenous METH self-administration over 5-days (Dao et al., 2011; Cardenas and Lotfipour, 2022). Given our prior results that nicotine exposure can sex- and age-dependently enhance METH self-administration (Cardenas and Lotfipour, 2022), our current studies evaluated the combined interactions in our CHRNA6 3′UTR SNP rat lines in order to test the hypothesis that α6*-containing nAChRs mechanisms influence these relationships. In parallel, we used the same 4-day sub-chronic paradigm to evaluate nicotine- and METH-induced DA overflow in the NAc shell in adolescent male and female CHNRA6 3′-UTR SNP rats. Our recent studies illustrate that adolescent nicotine reinstatement behavior predicts DA tissue levels in CHRNA6 3′UTR SNP rat lines providing support for our hypothesis (Carreño et al., 2024). To better understand mechanisms underlying our observed effects, male and female adolescent rats (PN 31) underwent in vivo analytical quantification of neurotransmitters collected from the interstitial fluid from the NAc shell. DA, and its metabolites, 3,4-Dihydroxyphenylacetic acid (DOPAC), and homovanillic acid (HVA) from the extracellular space were measured by high performance liquid chromatography coupled with electrochemical detection (HPLC-ECD) methods. Our studies test the hypothesis that sex- and genotype-dependent effects will be observed for nicotine-induced METH self-administration and drug-induced DA overflow in adolescent CHRNA6 3′-UTR SNP rats. The objective of this study is to fill critical gaps in understanding how nicotine exposure during adolescence and the CHRNA6 3′-UTR SNP influences METH self-administration and associated neurobiological changes. By exploring sex- and genotype-dependent effects, we gain insights into the complex interplay between genetics, sex, and co-substance use in adolescents, thereby informing future strategies for the prevention and intervention of substance abuse disorders.
2 Methods
2.1 Animals
Male and female, CHRNA63′-UTR SNP knock-in rats were developed and bred in house as previously described (Cardenas et al., 2022; Carreño and Lotfipour, 2023). Juveniles were weaned at postnatal day (PN 21), separated by sex, and handled for 3 days prior to experimentation. Rats were group-housed in a controlled 12-h light–dark cycle in an AAALAC-accredited vivarium. Food and water were available ad libitum except when indicated. Animals were weighed daily to ensure the maintenance of normal growth. To avoid potential litter effects, one pup per litter per experimental group was used for data collection. Animals were allocated to experimental groups using a random sequence generator. The study was conducted according to the guidelines of the Institutional Animal Care and Use Committee (IACUC) at the University of California, Irvine (UCI) (protocol: AUP-21-022, 4 February 2021). UCI’s IACUC has Animal Welfare Assurance #D16-00259 (#A3416.01) approval on file with the Office of Laboratory Animal Welfare at NIH and is fully accredited by AAALAC, International. CHRNA6 3′UTR SNP rat strains (SD-CHRNA6em1Slot 984, SD-CHRNA6em2Slot 985) have been donated to the Rat Resource and Research Center (RRRC).
2.2 Drugs
Nicotine tartrate (Glentham Life Sciences, Corsham, Wiltshire, United Kingdom) was calculated as a base, dissolved in saline, with a final pH of 7.2–7.4. Methamphetamine (METH) (National Institute of Drug Abuse (NIDA), Bethesda, MD) was dissolved in saline and filtered via 0.22 μm sterile filters (VWR). Equithesin (Pentobarbital and Chloral Hydrate (Sigma Aldrich, St. Louis, MO, United States), propylene glycol, ethanol and magnesium sulfate) is made in the laboratory, similar to our prior studies (Carreño and Lotfipour, 2023; Carreño et al., 2024). Carprofen (Zoetis, Parsippany, NJ, United States) was diluted in saline. Nicotine, Equithesin, and carprofen were filtered via 0.22 μm sterile filters (VWR, Radnor, PA, United States). Propofol (Medvet, Mettawa, IL, United States) in a 5 mg/kg, intravenous (i.v.) was administered to test for catheter patency.
2.3 Surgical implantation of intravenous catheter
Catheter construction and implantation was done as described previously (Belluzzi et al., 2005). Animals were anesthetized with Equithesin (0.0035 mL/g body weight), and a chronic catheter was surgically implanted into the right external jugular vein. The catheter was passed subcutaneously from the animal’s back to the jugular vein where the tubing was inserted. The cannula assembly was affixed to the animal’s back and sealed to maintain an unobstructed system. Wound closures were made with wound clips, antiseptic ointment was applied to the incisions, and carprofen (5 mg/kg, subcutaneous) was injected to prevent infection. The animals were kept in a warm cage for post-surgical observation until they emerged from anesthesia. Catheter patency was tested for rapid (5–10 s) anesthesia by infusion of propofol (5 mg/kg) before and after completion of self-administration and in vivo microdialysis experiments (Figure 1).
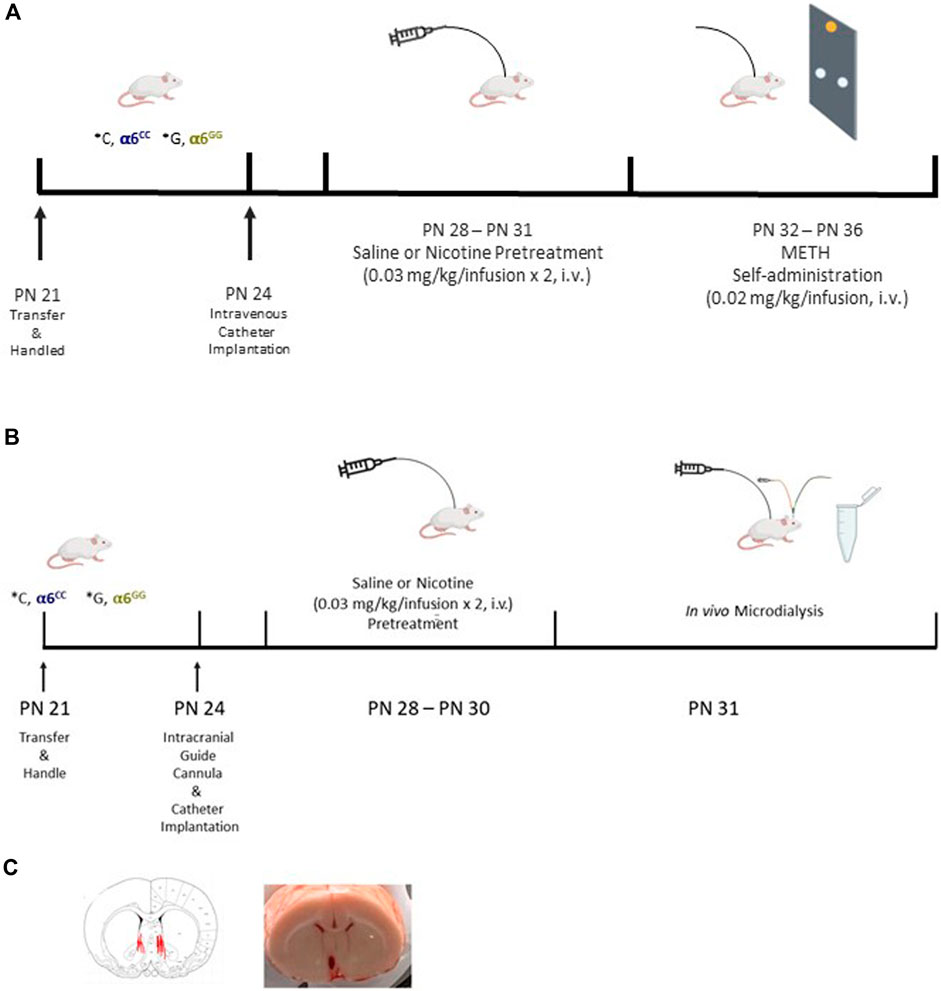
Figure 1. Experimental Design in the CHRNA6 3′UTR SNP Knock-In. Male and female CHRNA6 3′-UTR SNP knock-in rats were bred in-house and underwent a series of procedures including nicotine pretreatment and METH self-administration (A). In vivo microdialysis was performed to assess dopamine and its metabolites after drug administration (B). Probe placement confirmation in male and female CHRNA6 3′-UTR SNP knock-in rats (C).
2.4 Nicotine pretreatment
Starting on PN 28 rats were administered nicotine (2 × 0.03 mg/kg/0.1 mL, i.v.) or saline injections spaced 1 minute apart daily for 4 days (Figure 1A). The dual injection of 0.03 mg/kg nicotine daily over 4 days was chosen to model early initiation of nicotine use during adolescence (Cardenas et al., 2021), models prior studies using the same paradigm (Cardenas et al., 2021), and reduced receptor desensitization and toxic effects (McQuown et al., 2007). Furthermore, the 0.6 mg/kg dose is the optimal dose regularly chosen in nicotine self-administration studies, is equivalent to 1-2 cigarettes worth of nicotine/day and produces 30 ng/mL peak plasma levels in adolescent and adult rats (Cao et al., 2007; Matta et al., 2007).
2.5 METH self-administration
One day after nicotine pretreatment, animals were tested in 28 × 25 × 30 cm self-administration chambers with two nose pokes holes for 5 days (PN 32–36) (Figure 1A). Adolescent animals underwent a 2-h nose poke session on a fixed ratio 1 (FR1) schedule to administer METH (0.02 mg/kg/infusion (inf)) (Dao et al., 2011; Cardenas and Lotfipour, 2021) with a time-out period of 20 s signaled by a cue light over the reinforced hole. During this time-out period, animals were restricted from receiving another reinforced response (inf) and the house light was turned off. Nose poke of the nonreinforced hole was scored but did not result in a signal or inf. To determine catheter patency, after the last intravenous self-administration session, rats were administered propofol, a rapid anesthetic (0.05 mL for adolescents, 0.1 mL for adults, i.v.). Propofol negative animals were removed from the study analyses.
2.6 Surgery, in vivo microdialysis, and HPLC-ECD
2.6.1 Stereotaxic surgery
Following the placement of the i.v. Catheter, animals designated for microdialysis underwent the implantation of a cranial guide cannula (PN 24). Animals were placed in a stereotaxic frame (Stoelting Co., Chicago, Il, United States), their skulls revealed and drilled to expose the dura. A chronic guide cannula (Bioanalytical Systems, Inc., West Lafayette, IN) was stereotaxically implanted 2.0 mm above the target area, fixed to the skull with acrylic dental cement, and sealed with a dummy cannula. Anatomical coordinates for adolescent animals (PN 28-29) were determined from the adult atlas (Paxinos and Watson, 1989), then empirically identified in a preliminary experiment with histological confirmation. The following guide cannula coordinates were measured from the dura; nucleus accumbens shell AP, + 2.1 mm; ML, ± 0.6 mm; DV,-6 mm.
2.6.2 In Vivo microdialysis procedure
Animals were given 3 days to recover after surgeries with daily handling after surgeries and catheters flushed daily. Beginning on PN 28, the animals received either saline (acute condition) or nicotine (sub-chronic condition) pretreatment (PN 28-31, Figure 1A). Intravenous catheter patency was tested by propofol 1 day before the experiment. On the experimental day (PN 31), the dummy cannula was replaced with a 2 mm microdialysis probe with a 30 kDa cut-off membrane (MD-2200; Bioanalytical Systems, Inc., West Lafayette, IN). The probe membrane extended 2 mm beyond the tip of the guide cannula. The quality of probes was tested in vitro before the experiment with an average recovery of 10% (data not shown). Microdialysis was carried out under a free-moving condition in Culex NxT with Raturn–Multi Animal (Bioanalytical Systems, Inc., West Lafayette, IN), with the probe continuously perfused with artificial cerebrospinal fluid (Ringers Solution 147 mM NaCl, 2.2 mM CaCl2, 4 mM KCl) at a constant flow rate of 1.1 mL/min delivered by a Empis infusion pump (CX-300; Bioanalytical Systems, Inc., West Lafayette, IN) (Figure 1B). Dialysate was collected into a Honeycomb refrigerated fraction collector (MD-1201; Bioanalytical Systems, Inc., West Lafayette, IN) that maintained the samples at 4°C. After 4-h perfusion to establish an equilibration between the probes’ internal and external environment, baseline samples were collected every 20 min for 60 min. When DA levels reached a stable baseline, animals were given two 0.100 mL injections (i.v.) of saline, 1 min apart. After 100 min, nicotine (0.03 mg/kg/0.100 mL injection, i.v.) was injected twice at a 1 min interval and samples were collected for another 200 min. After 220 min, METH (0.02 mg/kg/0.100 mL injection, i.v.) was injected twice at 1 min interval and samples were collected for another 100 min (Figure 1). DA and its metabolite levels were quantified by HPLC-ECD. The position of microdialysis probes was verified histologically and mapped onto relevant atlas sections (Paxinos and Watson, 1986) (Figure 1C).
2.6.3 HPLC-ECD
Microdialysate samples (20 μL) were automatically injected by an ESA 542 refrigerated auto-sampler onto a 150 × 3 mm ODS C18 column (ESA Inc., Chelmsford MA) connected to an ESA 580 HPLC pump. The column was kept at 35°C and perfused by MD-TM mobile phase (ESA, Chelmsford, MA) at a rate of 0.6 mL/min. DA and metabolite levels were determined by an electrochemical ESA 5600 detector with an ESA 5011 microdialysis cell with the dominant potential of 160 mV. The sensitivity of the detector is 500 fg. Measurements were analyzed using CoulArray for Windows Software 2.0 (ESA Inc., Chelmsford, MA, United States). Standard curves were generated with catecholamine (ThermoScientific, Wlatham, MA), DOPAC, and HVA (Sigma-Aldrich, St Louis, MO) standards, and levels in experimental samples were determined from the curve and expressed as ng/20 μL injection (Equation 1), as there were no significant differences in probe recovery. Basal levels of DA and its metabolites were determined by averaging the samples before nicotine injection. Nicotine- and METH-induced changes in DA and DA metabolite levels were expressed as area under the curve (AUC) given by Equation 1 below.
2.7 Statistics
Data were analyzed with JMP statistical analysis software (SAS Institute, Cary, NC). A total of 186 rats were used in our studies. Animals that did not exhibit anesthesia from propofol were removed from the analysis (n = 41). Further exclusions were applied to animals that fell outside the boundaries defined by predetermined box and whisker plots for each group (n = 6 for METH self-administration and n = 8 for microdialysis). The 5-day METH acquisition mean response data over days were analyzed by a repeated measure five-way analysis of variance (ANOVA) for sex × genotype × pretreatment × response × day, with a repeated measure on day and response. Microdialysate samples AUC were analyzed by a four-way ANOVA for sex x genotype x pretreatment (acute vs. sub-chronic) x drug (nicotine, METH, saline) with repeated measures for drug. Significant main effects or interactions were further tested by t-test with the Bonferroni adjustment for multiple comparisons.
3 Results
3.1 Nicotine pretreatment enhances subsequent METH self-administration in the CHRNA6 3′-UTR SNP knock-in rats in a sex- and genotype-dependent manner
To determine whether the human SNP in the 3′-UTR of the CHRNA6 gene influences nicotine-induced METH self-administration, we evaluated mean response data over time. Overall ANOVA showed a main effect for reinforced/nonreinforced response [F1,72 = 41.86, p = 0.0001] with sex × reinforced/nonreinforced [F1,72 = 6.58, p = 0.012], pretreatment × reinforced/nonreinforced [F1,72 = 7.05, p = 0.0098], genotype × sex × pretreatment × reinforced/nonreinforced [F1,72 = 4.35, p = 0.04], response × day [F4,288 = 7.48, p = 0.0001], response × day × sex × pretreatment [F4,288 = 2.44, p = 0.047] and day × genotype [F4,288 = 2.47, p = 0.045] interactive effects. Since we identified interactions for every measure, we separated the data by genotype, sex, pretreatment, response, and day (Figures 2A–D). For females, nicotine-treated α6CC and saline-treated α6GG animals acquired and maintained METH self-administration on days 1–5 as measured by preference for reinforced over non-reinforced nose pokes (p < 0.05, Figures 2A, B). Conversely, saline-treated α6CC female rats demonstrated preference for reinforced over nonreinforced nose poke on day 1 of METH self-administration (p = 0.023) whereas nicotine-treated α6GG female rats acquired METH self-administration on days 4–5 (p < 0.05, Figures 2A, B). Nicotine-treated males responded for METH significantly more than saline-treated males on days 2–4, independent of genotype (p < 0.05, Figures 2C, D). Taken together, our findings demonstrate that adolescent sub-chronic nicotine exposure influences the acquisition and maintenance of METH in a sex-and genotype-dependent manner.
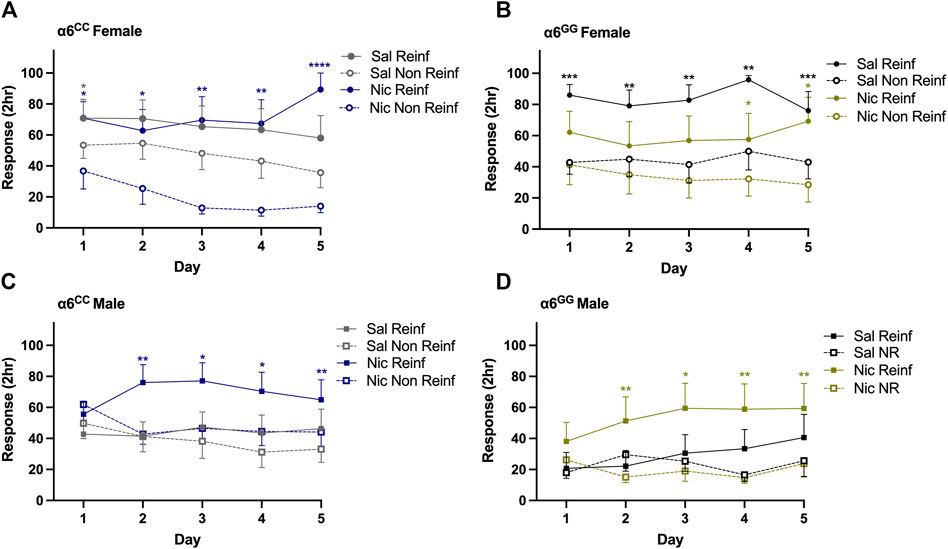
Figure 2. Sex- and genotype-dependent nicotine-induced METH self-administration in male and female CHRNA6 3′-UTR SNP rats. METH self-administration mean 2-h responses ±SEM across 5 days in (A) α6CC (B) α6GG females, (C) α6CC and (D) α6GG males. Nicotine-treated α6CC and saline-treated α6GG females maintain consistent METH self-administration, saline-pretreated α6CC females initially prefer reinforced responding. Nicotine-treated males show sustained METH self-administration from days 2–5, independent of genotype. Open circles represent female data; open and closed squares represent male data. ****p < 0.0001, ***p < 0.001, **p < 0.01, *p < 0.05 Reinf versus Non Reinf responses; n = 9/12/group. METH = methamphetamine, Reinf = Reinforced, Non Reinf = Non-Reinforced, Sal = Saline (grey/black), Nic = Nicotine (blue/gold).
3.2 Nicotine pretreatment impacts drug-induced DA overflow in a sex- and genotype-dependent manner in the CHRNA6 3′-UTR SNP rats
The effects of nicotine- and METH-induced DA overflow in the NAc shell, were assessed in male and female adolescent (PN 31) CHRNA6 3′-UTR SNP rats. An overall ANOVA for DA overflow showed a between main effect for pretreatment [F1,46 = 5.9267, p = 0.0189], as well as between interactive effects for genotype x sex [F1,46 = 7.3049, p = 0.0096], genotype x sex x pretreatment [F1,46 = 8.6829, p = 0.0050]. In addition, we observed a within main effect for drug [F2,92 = 12.1493], as well as within interactive effects for drug x genotype x sex [F2,92 = 7.3632, p = 0.0011], drug x pretreatment [F2,92 = 4.9620, p = 0.0090], and drug x genotype x sex x pretreatment [F1,92 = 6.2993, p = 0.0027]. Thus, we evaluated these parameters separately. Nicotine and METH induced DA overflow is enhanced in α6CC females as compared to α6GG females (p < 0.05), primarily in acute condition (Figures 3A–D). METH induced DA overflow is enhanced in α6GG males in the acute condition, but not sub-chronic condition. No variation in genotype were noted for nicotine effects in both acute and sub-chronic conditions in males. (Figures 3E–H).
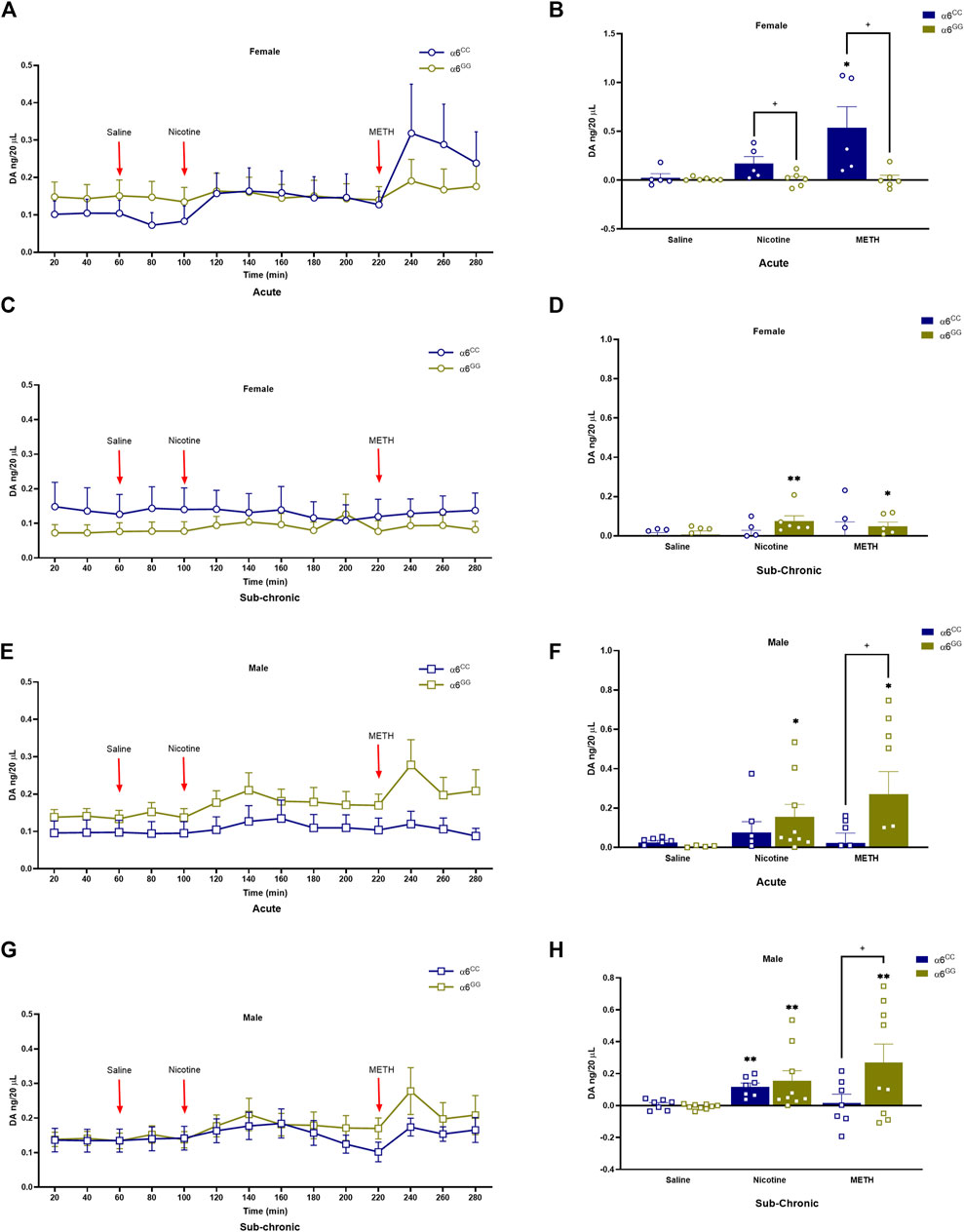
Figure 3. DA levels in male and female in CHRNA6 3′-UTR SNP Rats. The effects of nicotine- and METH-induced DA overflow in the NAc shell, were assessed in male and female adolescent (PN 31) CHRNA6 3′-UTR SNP rats. Female (A) acute timeline and (B) AUC (C) sub-chronic timeline and (D) AUC for saline, nicotine, and METH; Male (E) acute timeline and (F) AUC (G) sub-chronic timeline and (H) AUC for saline, nicotine, and METH. Nicotine and METH induce greater DA overflow in α6CC females than α6GG females in the acute condition, while α6GG males show enhanced METH-induced DA overflow only acute condition. No significant genotype differences are noted in nicotine-induced DA overflow in males under either acute or sub-chronic conditions.**p < 0.01 vs. saline; *p < 0.05 vs. saline; +p < 0.05 α6CC vs. α6GG. AUC = area under the curve METH = Methamphetamine N = 5-12/group.
We assessed DA metabolites, DOPAC and HVA, to determine their change in release in acute and sub-chronic condition. For DOPAC, a between main effect for pretreatment [F1,52 = 4.6560, p = 0.0356], in addition a between interactive effects for genotype x sex [F1,52 = 13.2859, p = 0.0006] and genotype x pretreatment [F1,52 = 12.0334, p = 0.0011] were observed. A 4-way multivariate ANOVA revealed drug x genotype x sex x genotype [F2,104 = 4.4833, p = 0.0136] interactive effects (Figures 4A–H). Data were separated by sex, genotype and pretreatment. In the acute condition, α6CC females displayed higher DOPAC overflow for saline and nicotine for the acute condition as compared to α6GG females (p < 0.001), this distinction was not evident in males (Figures 4A–F). In the sub-chronic condition, α6GG males exhibited a greater METH-induced DOPAC overflow compared to α6CC males (p < 0.001) (Figures 4G, H). HVA, an extracellular DA metabolite showed a between main effect for genotype [F1,47 = 12.4396, p = 0.0010], and between interactive effects for genotype x sex [F1,47 = 19.5262, p = 0.001], genotype x pretreatment [F1,47 = 7.2047, p = 0.01] and genotype x sex x pretreatment [F1,47 = 4.0431, p = 0.0501]. A 4-way multivariate ANOVA revealed drug x sex x genotype × pretreatment interaction [F2,98 = 3.7283, p = 0.0313]. Data were separated for sex, genotype and pretreatment. Nicotine and METH altered HVA overflow in saline- and nicotine-treated α6CC females as compared to α6GG females, these effects were not observed in males (Figures 5A–H).
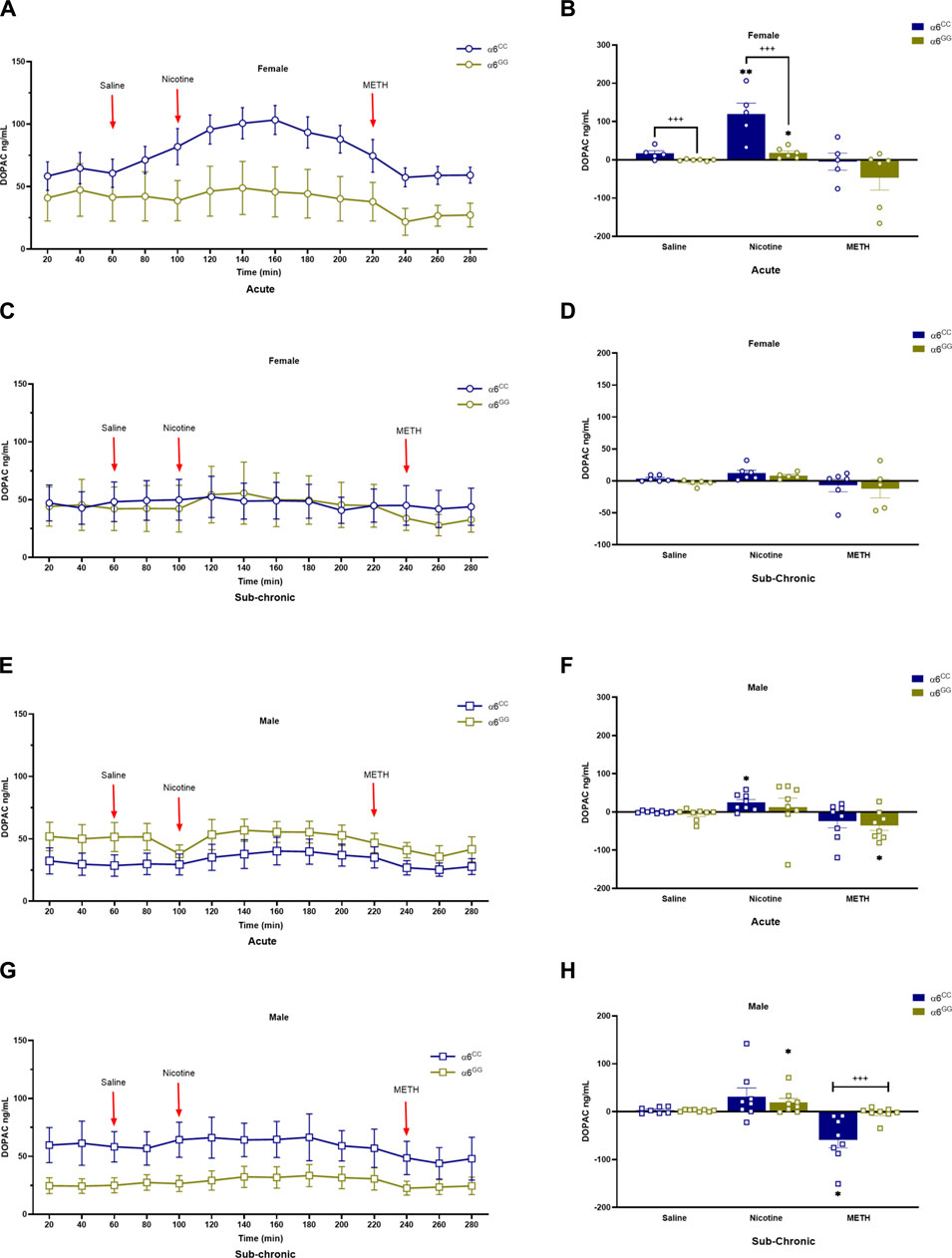
Figure 4. DOPAC levels in male and female in CHRNA6 3′-UTR SNP Rats. Female (A) acute timeline and (B) AUC (C) sub-chronic timeline and (D) AUC for saline, nicotine, and METH; Male (E) acute timeline and (F) AUC (G) sub-chronic timeline and (H) AUC for saline, nicotine, and METH. α6CC females exhibit higher DOPAC levels than α6GG females in response to saline and nicotine pretreatments, indicating genotype-specific DA metabolism in females. In males, this effect emerges under sub-chronic METH treatment, with α6GG males showing higher DOPAC levels than α6CC males. During acute conditions, α6CC females display significantly higher DOPAC overflow for both saline and nicotine compared to α6GG females (p < 0.001), a disparity not seen in males. Sub-chronic nicotine-treated α6GG males exhibit greater METH-induced DOPAC overflow than α6CC males. **p < 0.01, *p < 0.05 vs. saline; +++p < 0.001 α6CC vs. α6GG. AUC = area under the curve; METH = Methamphetamine N = 5-12/group.
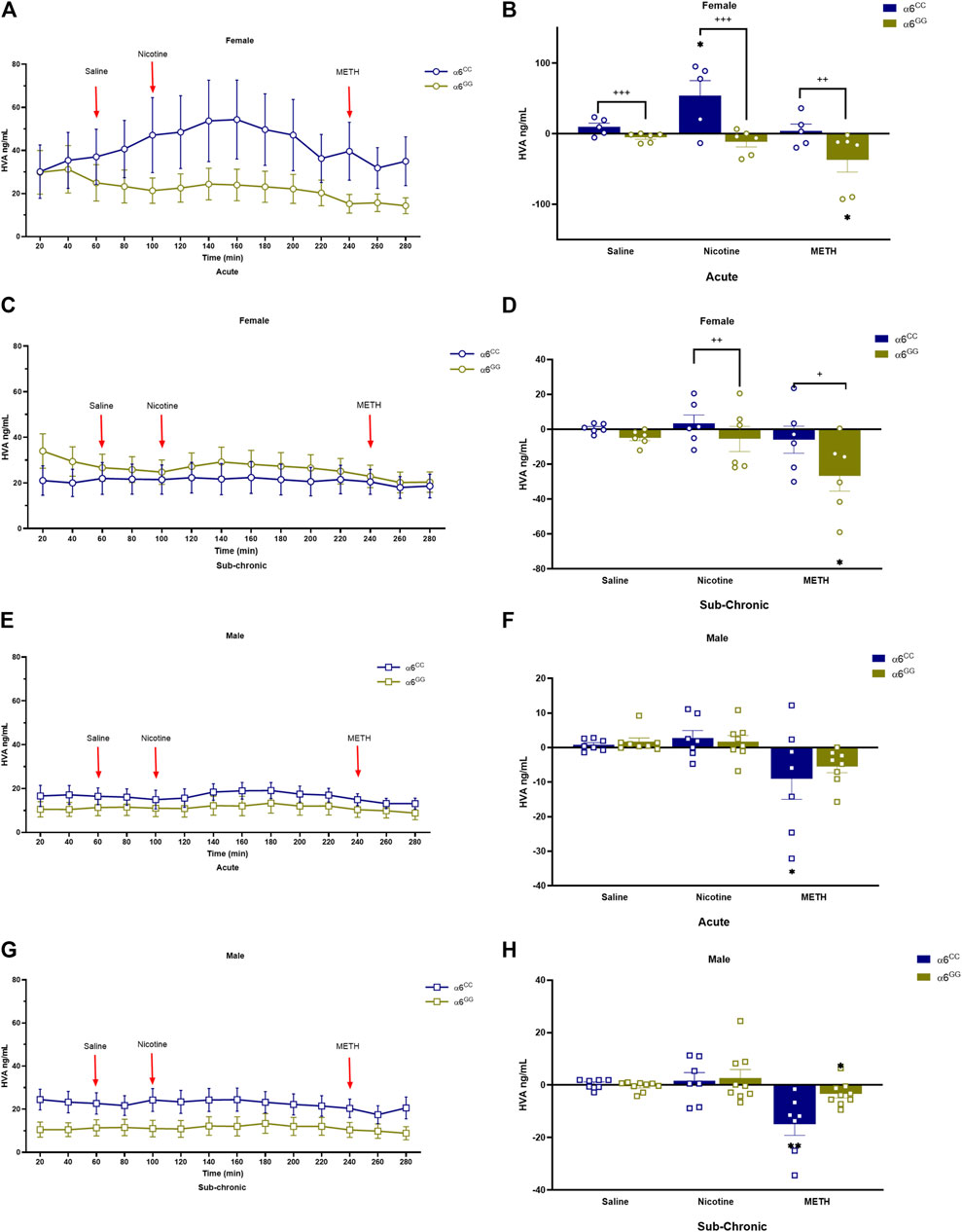
Figure 5. Figure 6 HVA levels in male and female in CHRNA6 3′-UTR SNP Rats. Female (A) acute timeline and (B) AUC (C) sub-chronic timeline and (D) AUC for Sal, Nic, and METH; Male (E) acute timeline and (F) AUC (G) sub-chronic timeline and (H) AUC for saline, nicotine, and METH. Saline- and nicotine-treated α6CC females show altered HVA overflow compared to α6GG females, a difference not observed in males **p < 0.01, *p < 0.05 vs. saline; +++p < 0.001 α6CC vs. α6GG. AUC = area under the curve, METH = Methamphetamine N = 5-12/group.
4 Discussion
In the present investigation, the impact of the CHRNA6 3′-UTR SNP on nicotine-induced METH self-administration and the overflow of DA in the NAc shell revealed multiple interactions, indicating an intricate interrelationship involving sex, pretreatment, and genotype. Notably, nicotine-treated α6CC and saline-treated α6GG females exhibited METH self-administration over several days. α6CC females when pretreated with saline (and not nicotine) initially showed a preference for reinforced versus non-reinforced responding for Day 1 METH self-administration. Nicotine-treated males, regardless of genotype, exhibited METH self-administration on days 2–5. These results highlight the sex- and genotype-dependent effects of adolescent sub-chronic nicotine exposure on the acquisition and maintenance of METH self-administration in rats with the human CHRNA6 3′-UTR SNP. Moreover, nicotine- and METH-induced DA overflow are enhanced in α6CC females as compared to α6GG females primarily in acute condition whereas α6GG males showed enhanced METH-induced DA overflow in acute, but not sub-chronic condition. No genotype differences were observed for males in either acute- or sub-chronic condition for nicotine-induced DA overflow. Taken together, these findings illustrate the complex interplay of sex, genotype, and pretreatment in influencing the effects of nicotine and METH on DA dynamics and self-administration behavior.
4.1 The influence of nicotine pre-treatment on METH self-administration in the CHRNA6 3′-UTR knock-in rats
Previous behavioral studies in rodents with the CHRNA6 3′-UTR SNP knock-in have shown that α6CC females and α6GG males displayed nicotine-induced enhanced locomotor and anxiolytic behavior when compared to their saline-treated counterparts (Cardenas et al., 2022). In addition, nicotine-induced locomotion and anxiolytic behavior in α6CC females was significantly higher than α6GG females. In contrast, nicotine-induced anxiolytic behavior in males was significantly higher in α6GG as compared to α6CC males (Cardenas et al., 2022). In the current study, we illustrate additional sex- and genotype-dependent adolescent nicotine exposure effects on subsequent METH self-administration. Nicotine pretreatment enhanced self-administration for METH in α6CC female rats. In contrast, saline-, but not nicotine-treated, α6GG females show self-administration for METH. In comparison, Sprague Dawley WT adolescent females did not exhibit increased METH responsiveness, whether or not they received nicotine pretreatment (Cardenas and Lotfipour, 2021). In male adolescents, prior nicotine exposure increased subsequent METH self-administration, its maintenance, and intake, independent of CHRNA6 3′-UTR SNP genotype. CHRNA6 3′-UTR SNP knock-in adolescent male rat data replicates adolescent WT male nicotine-induced enhancement of METH self-administration (Dao et al., 2011; Cardenas and Lotfipour, 2022). Sex and nicotine effects have been observed for METH and amphetamine self-administration as well as stimulant locomotor sensitization which may be further impacted by genetics (Collins et al., 2004; Collins and Izenwasser, 2004; Harmony et al., 2020). A potential mechanism for developmental nicotine exposure effects on subsequent METH self-administration is regulation of α6* nAChRs. A bidirectional effect of sex on transcriptional regulation of α6 nAChR mRNA expression and nicotine behaviors has been observed though protein kinase C epsilon in adult mice (Moen et al., 2021). However, we do not observe baseline or nicotine-induced α6 nAChR subunit mRNA differences in the CHRNA6 3′-UTR SNP knock-in rats (Cardenas et al., 2022). Whether the CHRNA6 3′-UTR SNP impact the translation of the α6 nAChR subunit protein needs to be evaluated in future studies. However, we are limited by lack of α6 specific antibodies available (Cardenas et al., 2020). Either way, our current findings suggest functional changes of α6 nAChRs in reward regions of the brain based on the CHRNA6 3′-UTR SNP genotype, sex, and drug treatment effects observed in our studies. These findings emphasize the critical role of using translational models to understand the complex interplay of sex, genotype, and pretreatment in influencing nicotine and METH effects. This approach can help in developing more targeted and effective interventions for substance use disorders. While our current studies have focused on drug reinforcement and neurotransmitter overflow, future studies could evaluate the effects of natural rewards as well in the CHRNA6 3′UTR SNP rodent line. Indeed, we have previously illustrated that our adolescent humanized CHRNA6 3′UTR SNP rats do not differ based on natural food rewards (Carreño and Lotfipour, 2023). Further, prior studies in wild type Sprague Dawley rats have illustrated that the 4-day nicotine pretreatment paradigm has minimal impacts on sucrose taking behavior, with no impacts on extinction and reinstatement during adolescents or adults, which is different than what is observed for stimulants (Mojica et al., 2014). For these reasons, we have not assessed sucrose reinforcement, extinction and reinstatement in our studies. Nevertheless, future studies could evaluate this to confirm whether similar results would be observed in the CHRNA6 3′UTR SNP rats. Furthermore, future studies should investigate sex-, genotype-, and nicotine-dependent α6* nicotinic receptor protein expression via binding assays in tissue of humanized CHRNA6 3-UTR SNP rats.
4.2 Nicotine pretreatments effects on drug-induced DA overflow are impacted in a sex- and genotype-dependent manner in CHRNA6 3′-UTR SNP rats
We have previously shown sex, age and genotype difference of DA tissue levels in adolescent, adult and nicotine-seeking CHRNA6 3′-UTR SNP knock-in rats (Carreño et al., 2024). Post-reinstatement, male α6GG rats show suppressed DA levels in the NAc shell compared to baseline (Carreño et al., 2024). The present study emphasizes the distinctions of DA transmission in the CHRNA6 3′-UTR SNP knock-in adolescent female and male rats of nicotine and METH. In the acute condition, α6CC females demonstrated heighten DA overflow by nicotine- and METH as compared to the α6GG females. In contrast, α6GG males exhibited greater METH-induced DA overflow when compared to α6CC males in the acute, but not sub-chronic condition, which may suggest tolerance effects. No genotype differences were observed for nicotine-induced DA overflow in the acute or sub-chronic condition in adolescent males. While there is nicotine-induced DA overflow among adolescent α6CC and α6GG males, there is no genotype difference in animals pretreated with saline or nicotine. However, we observe a genotype difference for METH-induced DA overflow in adolescent male saline-pretreated α6CC and α6GG rats. In contrast, we observe a more pronounced genotype difference in saline pretreated α6CC females for both nicotine- and METH-induced DA release when compared to α6GG females. Notably, α6CC females exhibit greater DA overflow in response to nicotine and METH compared to α6GG females particularly in the saline pretreatment condition. These results are in accordance with a previous study reporting acute nicotine increases DA transmission while long-term nicotine exposure reduces the release of DA in the NAc (Perez et al., 2012). In addition, the significant main and interactive effects of drugs indicate that nicotine and METH differentially influence DA overflow and its metabolites depending on genotype, sex, and pretreatment.
There is a significant difference in DOPAC levels based on genotype and sex, with α6CC females showing higher levels of DOPAC in response to saline and nicotine pretreatments compared to α6GG females, suggesting that genotype influences DA metabolism differently in females. In males, this genotype effect is not evident under acute conditions but becomes apparent under sub-chronic conditions with METH treatment, where α6GG males exhibit higher DOPAC levels than α6CC males. In the acute condition, α6CC females exhibit higher DOPAC overflow for saline and nicotine compared to α6GG females (p < 0.001), a difference not observed in males, while in the sub-chronic condition, α6GG males show greater METH-induced DOPAC overflow compared to α6CC males (p < 0.001). DOPAC levels suggest that the DAT, which is responsible for reuptake of DA from the synapse, may be influenced by genotype, sex, and drug exposure. Higher DOPAC levels in response to saline and nicotine pretreatments in α6CC females compared to α6GG females could indicate differences in DA reuptake or metabolism related to the DAT. Similarly, the differences observed in males under sub-chronic conditions with METH treatment suggest a potential role of the DA transporter in mediating these effects. Furthermore, the analysis of HVA levels revealed significant main and interactive effects influenced by genotype, sex, and pretreatment, as well as their interactions with drugs. Specifically, HVA overflow was altered in saline- and nicotine-treated α6CC females compared to α6GG females, an effect not observed in males. These results suggest that DA metabolism to DOPAC and HVA in the brain varies depending on the genetic background, sex, and prior exposure to substances like nicotine in female rats. The variations in HVA levels further suggest that the DA transporter may play a role in modulating DA metabolism in response to genetic and environmental factors. The altered HVA overflow in saline- and nicotine-pretreated α6CC females compared to α6GG females, specifically, could indicate differences in DA reuptake or metabolism that are influenced by the DAT. This variability in DA metabolism could potentially have implications for understanding how these factors contribute to differences in behavior or susceptibility to certain conditions. Future research that centers on the DA transporter has the potential to reveal additional insights into its function in these processes and its possible impact on behavior and vulnerability to specific conditions. Such studies could expand on these discoveries to develop a more thorough comprehension of how genetic and environmental factors influence DA metabolism and how these influences contribute to behavior and susceptibility to certain conditions.
Some potential limitations of our studies include the lack of assessment of drug-induced neurotransmitter release during METH self-administration. Given the technical difficulty of coupling in vivo microdialysis with METH self-administration, we have instead used a method that provides a potential readout of what our self-administration studies could provide if neurotransmitter release was simultaneously assessed during behavior. Given that contingent as compared with non-contingent delivery of drugs could differentially impact drug-induced neurotransmitter release, future studies could aim to couple these techniques with either in vivo microdialysis and/or fiber photometry to better understand the temporal dynamics of drug-induced neurotransmitter release and behavior. Further, our self-administration studies focus primarily on reinforcement behavior to METH at a fixed ratio one schedule of reinforcement. Thus, future studies could look at different schedules of reinforcement (e.g., fixed ratio 2-5), extended access and/or progressive ratio schedules of reinforcement, as well as extinction or reinstatement schedules of reinforcement, in order to determine how our findings impact other parameters of METH self-administration. Finally, while our studies focus on drug associated rewards, we could also evaluate whether our results impact natural rewards. While this is unlikely, given prior results related to this in wild type rats (Mojica et al., 2014) and the CHRNA6 SNP rats (Carreño and Lotfipour, 2023; Carreño et al., 2024), it would be an important question to assess in CHRNA6 3′UTR SNP rats in order to confirm that drug, not natural, rewards are what are most impacted by adolescent nicotine exposure.
5 Conclusion
The study investigated the effects of adolescent nicotine exposure on METH self-administration and DA overflow in the NAc shell of CHRNA6 3′-UTR SNP knock-in rats. The findings revealed significant interactions between sex, genotype, and nicotine pretreatment in modulating drug-induced DA overflow, metabolites, and METH self-administration in these rats. Nicotine pretreatment enhanced discrimination for reinforced non-reinforced responding for METH self-administration in α6CC females rats, whereas saline-treated, but not nicotine-treated α6GG females showed this type of discrimination for METH self-administration. Additionally, in male adolescents, prior nicotine exposure increased subsequent METH self-administration, maintenance, and intake, independent of the CHRNA6 3′-UTR SNP genotype. Female α6CC rats showed higher DA overflow in response to nicotine and METH compared to α6GG females. Male α6GG rats exhibited higher METH-induced DA overflow than α6CC males under sub-chronic pretreatment conditions, α6CC females had higher DOPAC levels after saline and nicotine pretreatments than α6GG females, indicating genotype influences DA metabolism differently in females. Significant differences in HVA levels were found based on genotype, sex, and pretreatment, with altered HVA overflow in saline- and nicotine pretreated α6CC females compared to α6GG females. In males, this effect appeared under sub-chronic METH conditions. The study suggests that the dopamine transporter plays a crucial role in modulating DA metabolism, with implications for behavior and susceptibility to conditions based on genetic and environmental factors. Understanding the complex interactions between sex, genotype, and nicotine exposure in modulating drug-induced behaviors and DA overflow can contribute to the development of more effective prevention and intervention strategies in the future. This can pave the way for more customized combined treatments such as pharmacotherapy, psychotherapy, and education to counter or lessen addiction behavior. Unlike conventional addiction methods that often follow a one-size-fits-all model with little consideration of a person’s genetics, personalized approaches can significantly improve treatment outcomes by tailoring interventions to an individual’s genetic profile and specific needs.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Ethics statement
The animal study was approved by Institutional Animal Care and Use Committee (IACUC) at the University of California, Irvine (protocol: AUP-21-022, 4 February 2021). The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
DC: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing–original draft, Writing–review and editing. AF: Data curation, Formal Analysis, Funding acquisition, Investigation, Writing–review and editing. AC: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Validation, Writing–review and editing. SL: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing–original draft, Writing–review and editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. The research was supported by the Tobacco and Related Disease Research Program (TRDRP) award [T31IP1427C, T33IP6417, T34IR8049] (SL), UCI School of Medicine Startup Funds (SL), Institute of Clinical Translational Science (ICTS) Pilot Award [TR001414] (SL). DC was supported by the Vertex Fellowship and UCI Graduate Dean Dissertation Fellowship. AF was supported by the National Science Foundation Fellowship. AC was supported by the Ford Foundation Predoctoral Fellowship and UC President’s Pre-Professoriate Fellowship.
Acknowledgments
The authors would like to thank Drs. Frances Leslie, James Belluzzi, and Emily Castro for their critical insights in this manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Albuquerque, E. X., Pereira, E. F. R., Alkondon, M., and Rogers, S. W. (2009). Mammalian nicotinic acetylcholine receptors: from structure to function. Physiol. Rev. 89, 73–120. doi:10.1152/physrev.00015.2008
Azam, L., Chen, Y., and Leslie, F. M. (2007). Developmental Regulation of Nicotinic Acetylcholine receptors within midbrain dopamine neurons. Neuroscience 23, 1347–1360. doi:10.1016/j.neuroscience.2006.11.011
Belluzzi, J. D., Wang, R., and Leslie, F. M. (2005). Acetaldehyde enhances acquisition of nicotine self-administration in adolescent rats. Neuropsychopharmacology 30, 705–712. doi:10.1038/sj.npp.1300586
Berry, K. M., Fetterman, J. L., Benjamin, E. J., Bhatnagar, A., Barrington-Trimis, J. L., Leventhal, A. M., et al. (2019). Association of electronic cigarette use with subsequent initiation of tobacco cigarettes in US youths. JAMA Netw. Open 2, e187794. doi:10.1001/jamanetworkopen.2018.7794
Birdsey, J., Cornelius, M., Jamal, A., Park-Lee, E., Cooper, M. R., Wang, J., et al. (2023). Tobacco product use among U.S. Middle and high School students — national youth tobacco Survey, 2023. MMWR 72, 1173–1182. doi:10.15585/mmwr.mm7244a1
Bosch, P. J., Benton, M. C., Macartney-Coxson, D., and Kivell, B. M. (2015). mRNA and microRNA analysis reveals modulation of biochemical pathways related to addiction in the ventral tegmental area of methamphetamine self-administering rats. BMC Neurosci. 16, 43. doi:10.1186/s12868-015-0186-y
Brecht, M.-L., Greenwell, L., and Anglin, M. D. (2007). Substance use pathways to methamphetamine use among treated users. Addict. Behav. 32, 24–38. doi:10.1016/j.addbeh.2006.03.017
Brunzell, D. H., Boschen, K. E., Hendrick, E. S., Beardsley, P. M., and Mcintosh, J. M. (2010). Alpha-conotoxin MII-sensitive nicotinic acetylcholine receptors in the nucleus accumbens shell regulate progressive ratio responding maintained by nicotine. Neuropsychopharmacology 35, 665–673. doi:10.1038/npp.2009.171
Cao, J., Belluzzi, J. D., Loughlin, S. E., Keyler, D. E., Pentel, P. R., and Leslie, F. M. (2007). Acetaldehyde, a major constituent of tobacco smoke, enhances behavioral, endocrine, and neuronal responses to nicotine in adolescent and adult rats. Neuropsychopharmacology 32, 2025–2035. doi:10.1038/sj.npp.1301327
Cardenas, A., Bai, Y., Hajy Heydary, Y., Li, J., Leslie, F. M., and Lotfipour, S. (2022). Sex-and genotype-dependent nicotine-induced behaviors in adolescent rats with a human polymorphism (rs2304297) in the 3-UTR of the CHRNA6 gene. Int. J. Mol. Sci. 23, 3145. doi:10.3390/ijms23063145
Cardenas, A., Elabd, M., and Lotfipour, S. (2020). Specificity of a rodent alpha(α)6 nicotinic acetylcholine receptor subunit antibody. Psychopharmacol. Berl. 237, 283–285. doi:10.1007/s00213-019-05413-x
Cardenas, A., and Lotfipour, S. (2021). Age- and sex-dependent nicotine pretreatment effects on the enhancement of methamphetamine self-administration in sprague-dawley rats. Nicotine Tob. Res. 24, 1186–1192. doi:10.1093/ntr/ntab218
Cardenas, A., and Lotfipour, S. (2022). Age- and sex-dependent nicotine pretreatment effects on the enhancement of methamphetamine self-administration in sprague-dawley rats. Nicotine Tob. Res. 24, 1186–1192. doi:10.1093/ntr/ntab218
Cardenas, A., Martinez, M., Saenz Mejia, A., and Lotfipour, S. (2021). Early adolescent subchronic low-dose nicotine exposure increases subsequent cocaine and fentanyl self-administration in Sprague-Dawley rats. Behav. Pharmacol. 32, 86–91. doi:10.1097/FBP.0000000000000593
Carreño, D., Facundo, A., Nguyen, M. T. T., and Lotfipour, S. (2024). Dopamine and norepinephrine tissue levels in the developing limbic brain are impacted by the human CHRNA6 3′-UTR single-nucleotide polymorphism (rs2304297) in rats. IJMS 25, 3676. doi:10.3390/ijms25073676
Carreño, D., and Lotfipour, S. (2023). Sex- and genotype-dependent nicotine plus cue-primed reinstatement is enhanced in adolescent Sprague Dawley rats containing the human CHRNA6 3′-UTR polymorphism (rs2304297). Front. Psychiatry 13, 1064211. doi:10.3389/fpsyt.2022.1064211
Collins, S. L., and Izenwasser, S. (2004). Chronic nicotine differentially alters cocaine-induced locomotor activity in adolescent vs. adult male and female rats. Neuropharmacology 46, 349–362. doi:10.1016/j.neuropharm.2003.09.024
Collins, S. L., Montano, R., and Izenwasser, S. (2004). Nicotine treatment produces persistent increases in amphetamine-stimulated locomotor activity in periadolescent male but not female or adult male rats. Brain Res. Dev. 153, 175–187. doi:10.1016/j.devbrainres.2004.08.004
Corrigall, W. A., Coen, K. M., and Adamson, K. L. (1994). Self-administered nicotine activates the mesolimbic dopamine system through the ventral tegmental area.
Dani, J. A. (2015). Neuronal nicotinic acetylcholine receptor structure and function and response to nicotine. Int. Rev. Neurobiol. 124, 3–19. doi:10.1016/BS.IRN.2015.07.001
Dao, J. M., McQuown, S. C., Loughlin, S. E., Belluzzi, J. D., and Leslie, F. M. (2011). Nicotine alters limbic function in adolescent rat by a 5-HT1A receptor mechanism. Neuropsychopharmacology 36, 1319–1331. doi:10.1038/npp.2011.8
Di Chiara, G., and Imperato, A. (1988). Drugs abused by humans preferentially increase synaptic dopamine concentrations in the mesolimbic system of freely moving rats. Proc. Natl. Acad. Sci. USA. 85, 5274–5278. (amphetamine/cocaine/ethanol/nicotine/opiates). doi:10.1073/pnas.85.14.5274
Drenan, R. M., Grady, S. R., Whiteaker, P., McClure-Begley, T., McKinney, S., Miwa, J. M., et al. (2008). In vivo activation of midbrain dopamine neurons via sensitized, high-affinity alpha 6 nicotinic acetylcholine receptors. Neuron 60, 123–136. doi:10.1016/j.neuron.2008.09.009
Elkashef, A., Vocci, F., Hanson, G., White, J., Wickes, W., and Tiihonen, J. (2008). Pharmacotherapy of methamphetamine addiction: an update. Subst. Abus 29, 31–49. doi:10.1080/08897070802218554
Ferrucci, M., Limanaqi, F., Ryskalin, L., Biagioni, F., Busceti, C. L., and Fornai, F. (2019). The effects of amphetamine and methamphetamine on the release of norepinephrine, dopamine and acetylcholine from the brainstem reticular formation. Front. Neuroanat. 13, 48. doi:10.3389/fnana.2019.00048
Goodwin, J. S., Larson, G. A., Swant, J., Sen, N., Javitch, J. A., Zahniser, N. R., et al. (2009). Amphetamine and methamphetamine differentially affect dopamine transporters in vitro and in vivo. J. Biol. Chem. 284, 2978–2989. doi:10.1074/jbc.M805298200
Goriounova, N. A., and Mansvelder, H. D. (2012). Short-and long-term consequences of nicotine exposure during adolescence for prefrontal cortex neuronal network function. Cold Spring Harb. Perspect. Med. 2, a012120. doi:10.1101/cshperspect.a012120
Gorukanti, A., Delucchi, K., Ling, P., Fisher-Travis, R., and Halpern-Felsher, B. (2017). Adolescents’ attitudes towards e-cigarette ingredients, safety, addictive properties, social norms, and regulation. Prev. Med. 94, 65–71. doi:10.1016/j.ypmed.2016.10.019
Harmony, Z. R., Alderson, E. M., Garcia-Carachure, I., Bituin, L. D., and Crawford, C. A. (2020). Effects of nicotine exposure on oral methamphetamine self-administration, extinction, and drug-primed reinstatement in adolescent male and female rats. Drug Alcohol Depend. 209, 107927. doi:10.1016/j.drugalcdep.2020.107927
Jackson, K. J., Mcintosh, J. M., Brunzell, D. H., Sanjakdar, S. S., and Damaj, M. I. (2009). The role of 6-containing nicotinic acetylcholine receptors in nicotine reward and withdrawal. doi:10.1124/jpet.109.155457
Kuriban, H. (1999). Does nicotine modify the psychotoxic effect of methamphetamine? Assessment in terms of locomotor sensitization in mice. J. Toxicol. Sci. 24, 55–62. doi:10.2131/jts.24.55
Laviolette, S., and van der Kooy, D. (2004). The neurobiology of nicotine addiction: bridging the gap from molecules to behaviour. Nat. Rev. Neurosci. 5, 55–65. doi:10.1038/nrn1298
Le Novère, N., Corringer, P.-J., and Changeux, J.-P. (2002). The diversity of subunit composition in nAChRs: evolutionary origins, physiologic and pharmacologic consequences. J. Neurobiol. 53, 447–456. doi:10.1002/neu.10153
Linker, K. E., Elabd, M. G., Tawadrous, P., Cano, M., Green, K. N., Wood, M. A., et al. (2020). Microglial activation increases cocaine self-administration following adolescent nicotine exposure. Nat. Commun. 11, 1–14. doi:10.1038/s41467-019-14173-3
Matta, S. G., Balfour, D. J., Benowitz, N. L., Boyd, R. T., Buccafusco, J. J., Caggiula, A. R., et al. (2007). Guidelines on nicotine dose selection for in vivo research. Psychopharmacol. Berl. 190, 269–319. doi:10.1007/s00213-006-0441-0
McCabe, S. E., West, B. T., and McCabe, V. V. (2018). Associations between early onset of E-cigarette use and cigarette smoking and other substance use among us adolescents: a national study. Nicotine Tob. Res. 20, 923–930. doi:10.1093/ntr/ntx231
McQuown, S. C., Belluzzi, J. D., and Leslie, F. M. (2007). Low dose nicotine treatment during early adolescence increases subsequent cocaine reward. Neurotoxicol Teratol. 29, 66–73. doi:10.1016/j.ntt.2006.10.012
Moen, J. K., DeBaker, M. C., Myjak, J. E., Wickman, K., and Lee, A. M. (2021). Bidirectional sex-dependent regulation of α6 and β3 nicotinic acetylcholine receptors by protein kinase Cε. Addict. Biol. 26, e12954. doi:10.1111/ADB.12954
Mojica, C. Y., Belluzzi, J. D., and Leslie, F. M. (2014). Age-dependent alterations in reward-seeking behavior after brief nicotine exposure. Psychopharmacol. Berl. 231, 1763–1773. doi:10.1007/s00213-013-3266-7
Nisell, M., Nomikos, G. G., and Svensson, T. H. (1994). Systemic nicotine-induced dopamine release in the rat nucleus accumbens is regulated by nicotinic receptors in the ventral tegmental area. Synapse 16, 36–44. doi:10.1002/syn.890160105
Paxinos, G., and Watson, C. (1989). The rat brain in stereotaxic coordinates. Fourth Edition. London: Academic Press.
Perez, X. A., Ly, J., McIntosh, J. M., and Quik, M. (2012). Long-term nicotine exposure depresses dopamine release in nonhuman primate nucleus accumbens. JPET 342 (2), 335–344. doi:10.1124/jpet.112.194084
Pons, S., Fattore, L., Cossu, G., Tolu, S., Porcu, E., McIntosh, J. M., et al. (2008). Crucial role of alpha4 and alpha6 nicotinic acetylcholine receptor subunits from ventral tegmental area in systemic nicotine self-administration. J. Neurosci. 28, 12318–12327. doi:10.1523/JNEUROSCI.3918-08.2008
Ren, M., and Lotfipour, S. (2019). Nicotine gateway effects on adolescent substance use. West. J. Emerg. Med. 696, 696–709. doi:10.5811/westjem.2019.7.41661
Rothman, R. B., Baumann, M. H., Dersch, C. M., Romero, D. V., Rice, K. C., Carroll, F. I., et al. (2001). Amphetamine-type central nervous system stimulants release norepinephrine more potently than they release dopamine and serotonin. Synapse 39, 32–41. doi:10.1002/1098-2396(20010101)39:1<32::AID-SYN5>3.0.CO;2-3
Russell, K., Dryden, D. M., Liang, Y., Friesen, C., O’Gorman, K., Durec, T., et al. (2008). Risk factors for methamphetamine use in youth: a systematic review. BMC Pediatr. 8, 48. doi:10.1186/1471-2431-8-48
Rusyniak, D. E. (2011). Neurologic manifestations of chronic methamphetamine abuse. Neurol. Clin. 29, 641–655. doi:10.1016/j.ncl.2011.05.004
Yuan, M., Cross, S. J., Loughlin, S. E., and Leslie, F. M. (2015). Nicotine and the adolescent brain. J. Physiol. 593, 3397–3412. doi:10.1113/JP270492
Keywords: electronic cigarettes, addiction, developing brain, microdialysis, tobacco, co-morbid drug use
Citation: Carreño D, Facundo A, Cardenas A and Lotfipour S (2024) Sub-chronic nicotine exposure influences methamphetamine self-administration and dopamine overflow in a sex-and genotype-dependent manner in humanized CHRNA6 3′-UTR SNP (rs2304297) adolescent rats. Front. Pharmacol. 15:1445303. doi: 10.3389/fphar.2024.1445303
Received: 07 June 2024; Accepted: 02 August 2024;
Published: 14 August 2024.
Edited by:
Vijayapandi Pandy, MIT World Peace University, IndiaReviewed by:
Marsida Kallupi, University of California, San Diego, United StatesMaw Shin Sim, University of Malaya, Malaysia
Copyright © 2024 Carreño, Facundo, Cardenas and Lotfipour. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Diana Carreño, bGRjYXJyZW5vQGdtYWlsLmNvbQ==