 Nan Wang
Nan Wang Qianqian Ma1,2†
Qianqian Ma1,2† Yan Liang
Yan Liang Xueqiong Wu
Xueqiong Wu- 1Tuberculosis Prevention and Control Key Laboratory, Beijing Key Laboratory of New Techniques of Tuberculosis Diagnosis and Treatment, Department of Tuberculosis, The Eighth Medical Center of PLA General Hospital, PLA General Hospital, Beijing, China
- 2Graduate School, Hebei North University, Zhangjiakou, Hebei, China
Background: Integrated traditional Chinese medicine and biomedicine is an effective method to treat tuberculosis (TB). In our previous research, traditional Chinese medicine preparation NiuBeiXiaoHe (NBXH) achieved obvious anti-TB effects in animal experiments and clinical practice. However, the action mechanism of NBXH has not been elucidated.
Method: Peripheral blood mononuclear cells (PBMCs) were collected to extract mRNA and differentially expressed (DE) genes were obtained using gene microarray technology. Finally, GEO databases and RT-qPCR were used to verify the results of expression profile.
Result: After MTB infection, most upregulated DE genes in mice were immune-related genes, including cxcl9, camp, cfb, c4b, serpina3g, and ngp. Downregulated DE genes included lrrc74b, sult1d1, cxxc4, and grip2. After treatment with NBXH, especially high-dose NBXH, the abnormal gene expression was significantly corrected. Some DE genes have been confirmed in multiple GEO datasets or in pulmonary TB patients through RT-qPCR.
Conclusion: MTB infection led to extensive changes in host gene expression and mainly caused the host’s anti-TB immune responses. The treatment using high-dose NBXH partially repaired the abnormal gene expression, further enhanced the anti-TB immunity included autophagy and NK cell-mediated cytotoxicity, and had a certain inhibitory effect on overactivated immune responses.
1 Introduction
Tuberculosis (TB) epidemic is still a serious public health problem. According to the WHO Global Tuberculosis Report 2023, there were about 10.6 million new TB patients, 1.3 million deaths from TB, 410,000 rifampicin-resistant TB (RR-TB), and the burden of RR-TB and multidrug-resistant TB (MDR-TB) continues to increase, with a low global success rate of RR/MDR-TB treatment (only 63%) worldwide in 2022 (Global Tuberculosis Report, 2023). At present, there are many problems in TB treatment: 1) The current anti-TB chemotherapy needs to take a long time, and these long and complex chemotherapy regimens are a severe challenge for TB patients and the public health system. 2) The bacteria have special regulations and drug resistance mechanisms on the intervention of antibiotics, which leads to the gradual increase of drug resistance of Mycobacterium tuberculosis (MTB) and needs more expensive and intolerable drugs used for longer treatment (Pontali et al., 2019; Arora et al., 2021). In addition, new drug research and development also need a long time. 3) Anti-TB drugs have great toxic and side effects. Long-term chemotherapy has different degrees of damage to liver function, renal function, gastrointestinal tract, etc., especially about 10% of TB patients have severe liver damage and have to stop chemotherapy (An et al., 2013); 4) The onset and development of TB are closely related to the deficiency of immune function, imbalance of Th1/Th2 immune response, and low immunity (Cai et al., 2019; An et al., 2022). Anti-TB treatment is facing great challenges. Therefore, it is extremely urgent to develop new therapeutic agents, explore new therapeutic approaches, and establish new joint anti-TB treatment programs.
Traditional Chinese medicine (TCM) has been used to treat TB for more than 2000 years, which was recorded in the Internal Classic, the Miraculous Book of Ten Prescriptions, the Medical Biography, etc., Before the invention of chemotherapeutic drugs, TCM played an extremely important role in the treatment and control of TB and accumulated rich clinical experience. It was found that the deficiency of Vital Energy was an important basis for the onset of pulmonary TB, and the treatment principle of “tonifying deficiency and killing insects” was established, which emphasized adjusting the internal factors of the human body and enhancing self-repair ability to achieve the goal of cure TB. TCM has the advantage of comprehensive treatment of TB with multi-components, multi-targets, and multi-systems. Therefore, the auxiliary treatment of pulmonary TB with TCM can build up healthful vital energy, improve the patient’s immunity, repair the damage, cooperate with biomedicine to play an anti-TB role, to improve the patient’s symptoms of TB poisoning, promote the absorption of lesions and the closure of the cavity, and accelerate the negative conversion of sputum bacteria, thus improving the clinical treatment effect (Li et al., 2020). In recent years, modern experimental methods have been used to clarify the material basis of the efficacy of many TCM prescriptions for the treatment of TB, which has promoted the modernization process of TCM prescriptions. A variety of new anti-TB Chinese polyherbal preparations (such as JieHeWan, FeiTai capsule, QinJiaLiFei capsule, etc.,) have obtained new drug certificates approved by the National Medical Products Administration in China.
The TCM compound NiuBeiXiaoHe (NBXH) is an effective anti-TB empirical formula that has been used for a long time in clinical practice, mainly composed of six traditional Chinese medicines, including Fritillaria cirrhosa, Bletilla striata, Houttuynia cordata, Platycodon grandiflorum, Fructus arctii, and glutinous rice. Among them, Fritillaria cirrhosa and Bletilla striata are used as monarch medicine. Fritillaria cirrhosa has the effects of clearing heat and resolving phlegm, moistening the lungs and relieving cough, dispersing nodules and eliminating carbuncle, and is used to relieve cough, expectorate, relieve asthma, inhibit bacteria and inflammation, and widely used in the clinical treatment of respiratory diseases (Wang et al., 2016; Chen et al., 2020; Zhang et al., 2020). Bletilla striata has the effects of astringent hemostasis, dispersing swelling and engendering flesh, clearing heat and removing dampness, and is used to stanch bleeding, resist pathogenic microorganisms, inhibit inflammation, regulate immunity, and used for anti-fibrosis, anti-oxidation, etc., (Jiang et al., 2019; Zhai et al., 2021; Zhang et al., 2021). Therefore, Bletilla striata is mainly used to treat hematemesis, hemoptysis, pulmonary TB, and various ulcer bleeding in clinics. Houttuynia cordata, Platycodon grandiflorum, and Fructus arctii are all minister medicines. Houttuynia cordata has the effects of clearing heat and detoxifying, eliminating carbuncle and discharging pus, promoting diuresis and relieving stranguria, clearing heat and stopping diarrhea, and is used for anti-bacteria, anti-viruses, anti-inflammation, enhancing the immune function, anti-allergy, relieving asthma and cough, etc., (Sekita et al., 2016; Shingnaisui et al., 2018; Utaiwat et al., 2021). Platycodon grandiflorum has the effect of dispersing lung and eliminating phlegm, facilitating pharynx and discharging pus, and is mainly used for relieving cough and eliminating phlegm, anti-inflammation, anti-oxidation, regulating immunity, protecting liver and kidney, etc., (Choi et al., 2017; Zhang et al., 2018; Li and Yang, 2021). Fructus arctii has the effects of dispersing lung and eliminating phlegm, promoting pharynx and rash, detoxifying and detumescence, moistening intestines and relieving constipation, and is used for anti-bacteria, anti-viruses, anti-inflammation, and liver protection (Gao et al., 2018; Li et al., 2022). Glutinous rice is an assistant medicine, which has the effects of tonifying the middle and replenishing qi, strengthening the spleen and stomach, and stopping sweating. To sum up, the main function of this prescription is to moisten the lung and relieve cough, clear heat and eliminate phlegm, and eliminate carbuncle and discharge pus, which can be used for pulmonary TB and lung carbuncle with yin deficiency and phlegm heat, with symptoms of cough, yellow purulent sputum, hemoptysis, fever or low fever in the afternoon, chest tightness or dry stool. The results of our previous studies showed that the extract of the effective TCM compound NBXH in the clinical treatment of TB could improve the immune function, inhibit the growth of MTB, and reduce the degree of the lesion in the treatment of the mouse TB model (Liang et al., 2017; Duan et al., 2021; Liang et al., 2021). At present, this Chinese medicine compound has been developed into a traditional Chinese polyherbal preparation. The preparation process is different from that of TCM decoction. It is necessary to understand its mechanism of action and clarify its target and regulatory pathway. Therefore, based on previous studies (Liang et al., 2017; Duan et al., 2021; Liang et al., 2021), this study constructed a mouse model of MTB acute infection. With traditional Chinese polyherbal preparations JieHeWan (JHW) powder as a control, we studied the changes in gene transcription expression and regulation pathway after MTB infection and after treatment with traditional Chinese polyherbal preparation NBXH intermediates through gene expression microarray technology, and verified the expression of some genes in TB patients through GEO database and RT-PCR, to clarify the mechanism of "NBXH - TB targets - biomolecular network” regulating the body to play an overall anti-TB role.
2 Materials and methods
2.1 The preparation of NBXH intermediate
NBXH intermediate (lot number H200828) was produced by Xi’an Xintong Pharmaceutical Research Co., Ltd. (Xi’an, China). The preparation process is shown as follows: Taked 167 g Houttuynia cordata, 208 g Platycodon grandiflorum, and 167 g Fructus arctii, with 10 times the amount of water to boil three times continuously. The first and second times were for 2 h and the third was for 1 h. Combined and filtered the decoctions, then concentrated the filtrate under reduced pressure (60°C–70°C) to a thick paste with a relative density of 1.20–1.25 (60°C), dried under reduced pressure, ground the dry extracts and passed through a 100-mesh sieve, and then mixed it with the powder of 208 g Fritillaria cirrhosa, 312 g Bletilla striata, and 188 g glutinous rice, which directly ground and passed through a 100-mesh sieve.
2.2 Experimental design and grouping
Based on the previous animal experimental study on TB treatment with NBXH intermediates (Liang et al., 2017; Duan et al., 2021; Liang et al., 2021), this study further studied the anti-TB action mechanism of NBXH in mice. The complete research design, including animal model construction, grouping, medicine administration plan, gene microarray, and validation of gene expression profile are shown in Figure 1. Briefly, 3 days after MTB infection, 50 mice were randomly divided into the following five groups and orally administered 5 times a week: TB model group (0.5 mL distilled water/day per mouse), JHW treatment group (12 mg/day per mouse), low-dose (2.4 mg/day per mouse) of NBXH intermediate group, middle-dose (4.8 mg/day per mouse) of NBXH intermediate group, and high-dose (9.6 mg/day per mouse) of NBXH intermediate group. On the 87th day after treatment, the mice were euthanized and the total RNAs of peripheral blood mononuclear cells (PBMCs) from three mice were extracted to perform gene microarray analysis to obtain differential gene (DE) profile. Finally, the GEO database and RT-qPCR assay were used to validate the reliability of the DE gene profile.
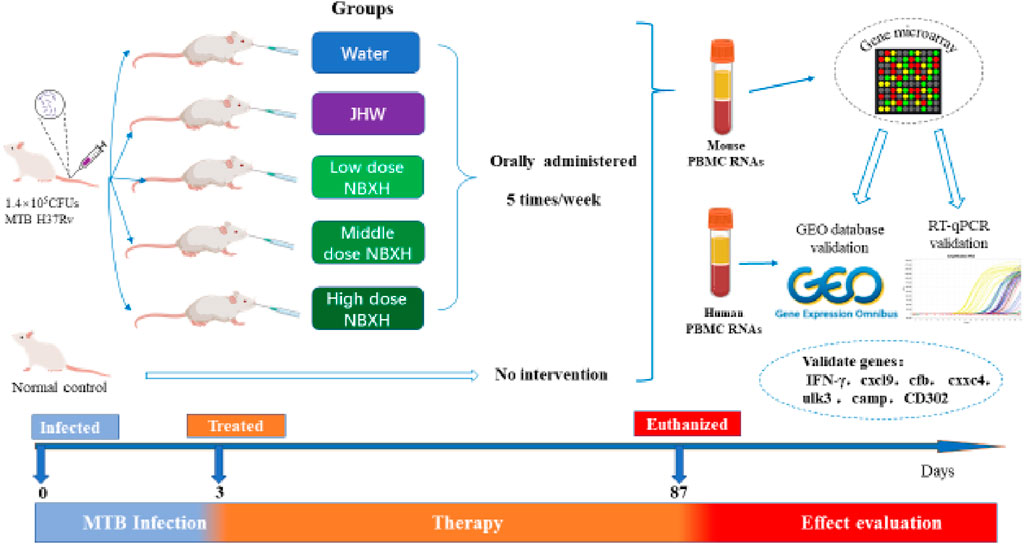
Figure 1. The study flowchart.
2.3 Experimental animals and ethics
Sixty BALB/c 42–62-day-old female mice were purchased from Beijing Vitalriver Experimental Animal Technology Co., LTD., and put into the animal laboratory of the Eighth Medical Center of the PLA General Hospital for adaptive feeding. The mouse model of acute MTB infection was prepared by injecting 1.4 × 105 CFUs MTB H37Rv standard strain (Laboratory of National Institute for Food and Drug Control) suspension through the tail vein. The animal experiments have been approved by the Animal Ethics Committee of the Eighth Medical Center of the PLA General Hospital (309,202,108,101,027). Animal feeding and management strictly follow the animal management and usage regulations of the animal laboratory of the Eighth Medical Center of the PLA General Hospital.
2.4 PBMC isolation and RNA extraction
Three weeks after the last immunization, three mice in each group mentioned above were euthanized, and then their blood was collected in heparin sodium anticoagulant tubes. According to the manufacturer’s instructions, PBMCs were isolated with the Mouse PBMC Isolation Kit (Haoyang Biological Products Technology, Tianjin, China), and total RNA was extracted from PBMCs using TRIzol Reagent (Invitrogen, United States).
2.5 Gene microarray expression analysis
The gene microarray experiments were conducted by Kangcheng Biotechnology Co., LTD. (Shanghai, China) using the Agilent Array platform. In brief, RNA samples were identified using a NanoDrop ND-1000 ultraviolet spectrophotometer (Implen, Germany) and agarose gel electrophoresis. Then, according to the experimental instructions of Agilent One-Color Microarray-Based Gene Expression Analysis (Agilent Technology, United States), RNA samples were labeled using Agilent Quick Amp Labeling kit, and hybridization experiments were performed using Agilent mouse 4 × 44 K gene expression profile chip v2 (4 × 44 K, Agilent, United States). The hybrid chip was washed, fixed, and scanned using the Agilent DNA Microarray Scanner (part number G2505°C). Agilent Feature Extraction software (v11.0.1.1) was used to collect the chip probe signal value to obtain the original data. Finally, the GeneSpring GX v12.1 software (Agilent Technology) was used for Quantile standardization and subsequent processing of the original data.
2.6 DE genes screening and identification
The DE genes in the TB model group vs. normal group, JHW group vs. TB model group, as well as low, middle, and high dose NBXH groups vs. TB model group were screened by analyzing hierarchical clustering heatmaps, scatter plots, and volcano plots. The Hierarchical cluster plot showed the differences in gene expression among different groups of samples. The scatter plot showed the DE gene correlation between two groups of samples, where the x-axis values and the y-axis values were the results of mean normalization processing (log2 scaling). The volcano plot showed the distribution of DE genes between two groups of samples, with the x-axis representing the log2 value (Fold Change) and the y-axis representing the -log10 value (p-value).
2.7 GO analysis and KEGG analysis
Gene Ontology (GO) is an internationally standardized gene functional classification system that comprehensively describes the attributes of any biological gene and its products by using a dynamically updated controlled vocabulary and strictly defined concepts. In this study, significant DE genes between different groups were subjected to GO term enrichment analysis on biological process (BP), cell component (CC), and molecular function (MF) through the standard vocabulary provided by Gene Ontology (http://www.geneontology.org). to calculate the hypergeometric distribution relationship between DE genes and several specific branches in the GO classification to explore which gene function changes may be related to DE genes in different samples. The Kyoto Encyclopedia of Genes and Genomes (KEGG) database is a database that systematically analyzes gene function, and links genomic information and functional information, and the metabolic pathway database is the most widely used. Through the pathway analysis of DE genes, the metabolic pathways enriched in DE genes were found, and which metabolic pathways might lead to differential gene expression between different groups of samples were identified. The GO and pathway analysis used hypergeometric tests to calculate p-values that indicate the importance of the conditional pathway or GO item enrichment in the DE gene. The lower the p-value, the more significant the GO term or pathway is (recommended cutoff value 0.05).
2.8 GEO database verification
The relevant transcriptome data were downloaded from the GEO database of the National Center for Biotechnology Information (NCBI) (http://www.ncbi.nlm.nih.gov/geo/). A total of 12,505 studies were retrieved using the keyword “Tuberculosis”. Select the GSE dataset with the study type of expression profiling by array, organisms of mouse and Homo sapiens, and sample of whole blood or PBMC. The GEO2R tool was used to screen DE genes with the threshold of ∣log2FC∣> 0.5 and adjust the p-value < 0.05. Select the datasets associated with TB diagnosis to screen DE genes compared with the top 10 upregulated or downregulated DE genes in the TB model group vs. the normal group in our profile. Then, select the datasets related to TB treatment to screen DE genes compared with the top 10 DE genes in the high-dose NBXH group vs. the TB model group.
Seven datasets, including GSE54992, GSE48027, GSE98461, GSE83456, GSE62525, GSE34608, and GSE14361, were selected to verify the top 10 upregulated or downregulated DE genes in TB model group vs. normal group. The GSE54992 dataset included nine patients with active tuberculosis (ATB) and six healthy controls (HCs) (Cai et al., 2014). The GSE48027 dataset included 6 MTB-infected mouse models and four uninfected healthy mice (Manca et al., 2013). The GSE98461 dataset included 4 ATB patients and 4 HCs (Pan et al., 2017). The GSE83456 dataset included 45 ATB patients and 61HCs (Blankley et al., 2016). The GSE62525 dataset included 14 ATB patients and 14 HCs (Lee et al., 2016). The GSE34608 dataset included 8 ATB patients and 18 HCs (Maertzdorf et al., 2012). The GSE14316 dataset included 4 MTB-infected mouse models and four uninfected healthy mice (Schreiber et al., 2009).
Four datasets, including GSE54992, GSE62147, GSE48027, and GSE31348, were selected to verify the top 10 upregulated or downregulated DE genes in the high-dose NBXH group vs. TB model group. The GSE54992 dataset included 9 ATB patients before and after 26 weeks of treatment (Cai et al., 2014). The GSE62147 dataset included 26 ATB patients before and after 26 weeks of treatment (Tientcheu et al., 2015). The GSE48027 dataset included 2 MTB-infected mouse models before and after 26 weeks of treatment (Manca et al., 2013). The GSE31348 dataset included 27 ATB patients before and after 26 weeks of treatment (Cliff et al., 2013).
2.9 RT-qPCR validation experiment
About 10 mL of whole blood from 40 initially treated pulmonary TB patients and 39 HCs was collected in heparin sodium anticoagulant tubes and RNAs were extracted for RT-qPCR assay, to verify the DE genes in the TB model group vs. the normal group. The inclusion criteria of the initially treated pulmonary TB patients (18–60 years old) was sputum-culture positive, and/or Xpert-positive, and/or interferon-γ release assay (IGRA)-positive, without anti-TB treatment or with anti-TB treatment for less than 1 month. Then, PBMCs were respectively isolated from five initially treated pulmonary TB patients before and after chemotherapy combined with NBXH treatment, and RNAs were extracted for RT-qPCR assay to verify the DE gene in the high dose NBXH group vs. TB model group. The inclusion criteria were initial active pulmonary TB patients treated with anti-TB chemotherapy combined with NBXH for ≥8 weeks. All PBMCs were stimulated with CFP10-ESAT6 fusion antigen (Shanghai Jin Nuo, China) for 24 h, all RNAs were extracted using the Trizol reagent (Invitrogen, United States), and 1 μg RNA was reverse-transcribed into cDNA using a reverse transcription kit (Takara, Japan). The quantification of cDNA was examined using RT-qPCR on Roche 480 (Roche, Switzerland) with the following procedure: pre-denatured at 95°C for 3 min; 40 cycles of denaturing at 95°C for 10 s, annealing at 60°C for 20 s; extension at 72°C for 1 s. RT-qPCR of each cDNA sample was repeated twice, and the final Ct value was the mean of the two times. The relative expression levels of all genes were measured by the 2−△△Ct method using GAPDH as the internal reference. The sequences of each gene primer are shown in Table 1.
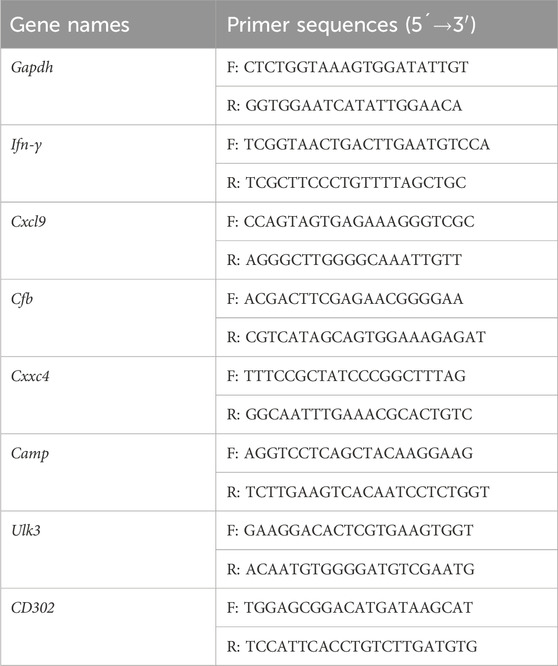
Table 1. The primer sequences for amplification of the DE genes.
2.10 Data analysis
The mouse gene expression profile was analyzed by assessing the Fold Change method, and the threshold of Fold Change values for screening upregulated or downregulated DE genes was set to ≥2, with a p-value < 0.05. The R language was used for hierarchical cluster analysis. GO analysis and pathway analysis were performed using standard enrichment calculation methods. RT-qPCR data were statistically analyzed using GraphPad Prism nine software (California, United States). The t-test was used to compare the relative expression data between ATB patients and HCs, and a p-value of <0.05 was considered statistically significant. The relative expression data of ATB patients before and after treatment were used paired t-test, and p-value < 0.05 was considered to have a statistical difference.
3 Result
3.1 Anti-TB immune responses in mice infected with MTB
3.1.1 DE genes in the TB model group vs. the normal group
The scatter plot, volcano plot, and cluster diagram of DE genes in the TB model group vs. normal group, JHW group vs. TB model group, and each dose of NBXH group vs. TB model group were shown in Supplementary Figure S1. In the TB model group vs. normal group, the total number of upregulated DE genes was 829, while the total number of downregulated DE genes was 505 (Table 2), indicating that the gene expression in mice was significantly disturbed after MTB infection.
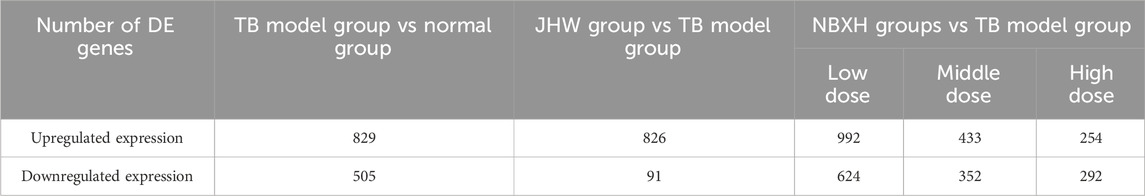
Table 2. The number of DE genes in TB model group vs. normal group, JHW group vs. TB model group, and each NBXH dose group vs. TB model group.
The top 10 upregulated and downregulated DE genes in the TB model group vs. the normal group and their changes in other groups were shown in Tables 3, 4 respectively. The results showed that: 1) In the TB model group vs. normal group, the FC value of the upregulated DE genes was much higher than that of the downregulated DE genes, and the disturbance of gene expression was mainly upregulation. 2) In the TB model group vs. normal group, the upregulated DE genes were all related to host immune defense and cytokine regulation, and the downregulated DE genes involved many functions, including nervous system function regulation, endocrine, immunity, etc., These results indicated that mice infected with MTB mainly upregulated the expression of anti-TB immune-related genes.
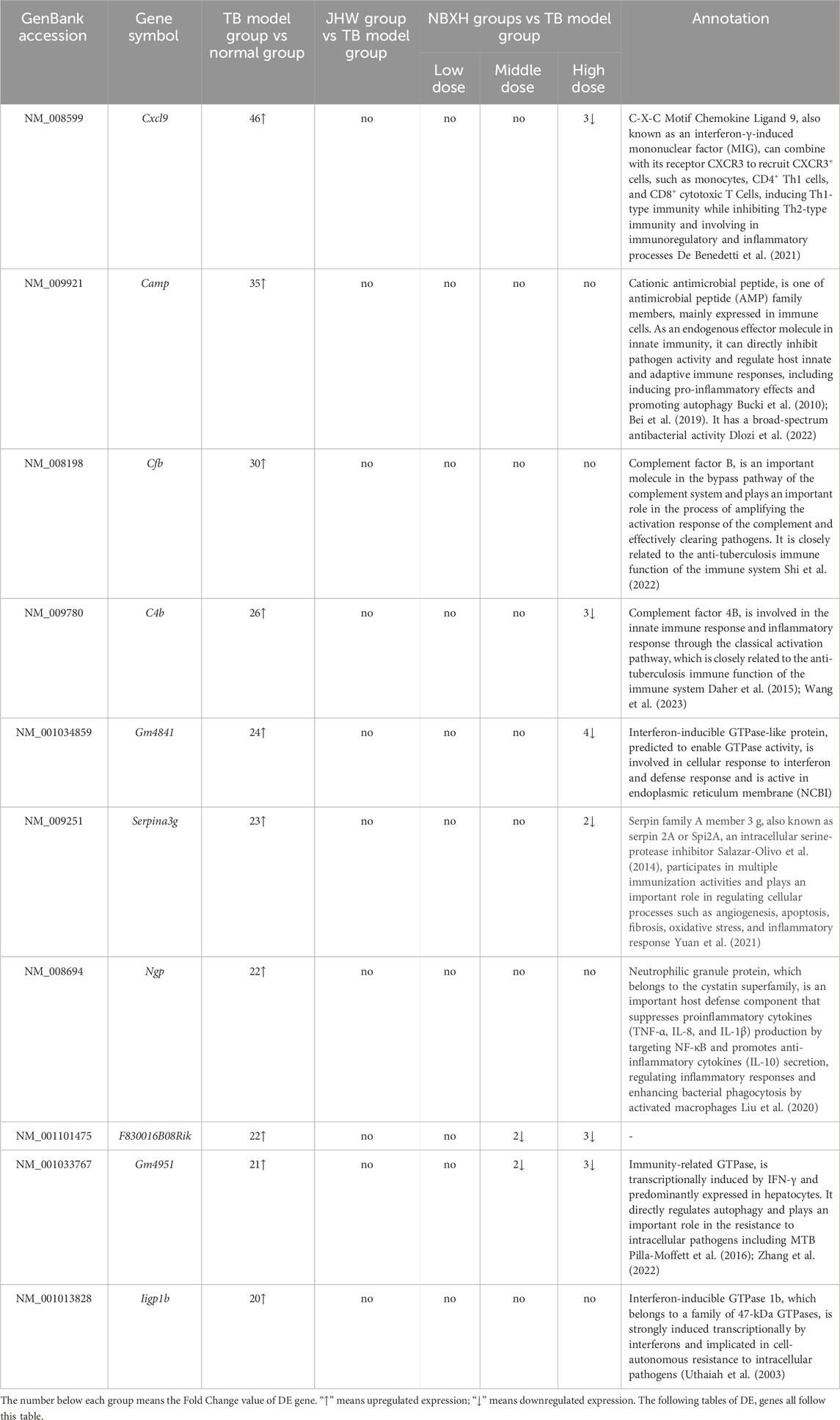
Table 3. The top 10 significantly upregulated DE genes in TB model group vs normal group and their changes in JHW group vs TB model group and each NBXH dose group vs TB model group.
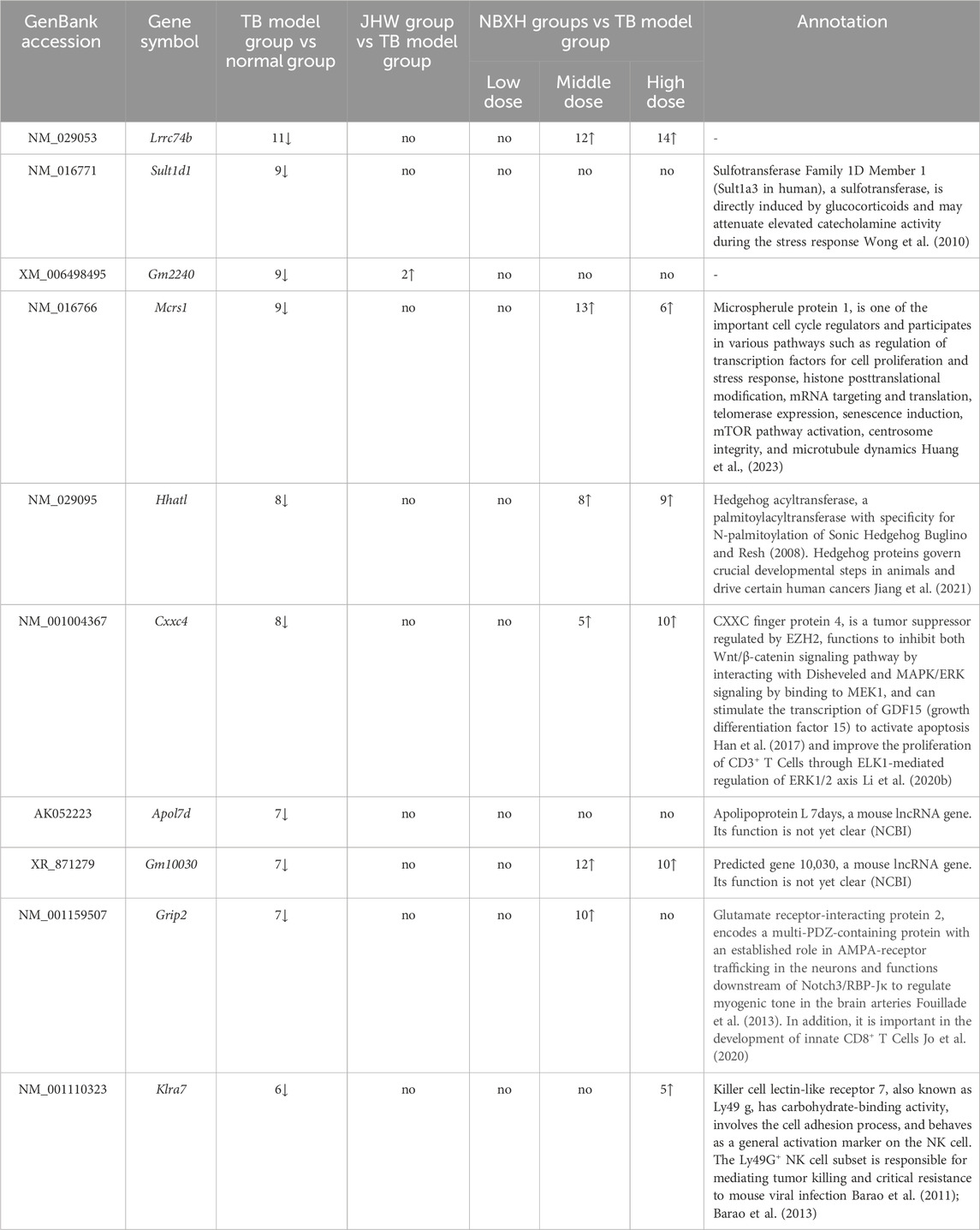
Table 4. The top 10 significantly downregulated DE genes in TB model group vs normal group and their changes in JHW group vs TB model group and eah NBXH dose group vs TB model group.
3.1.2 GO analysis in the TB model group vs. the normal group
GO analysis of the top 10 items of CC, BP, and MF in the TB model group vs. the normal group (Supplementary Figure S2) showed that: 1) Upregulated CC items were mainly associated with the extracellular regions, cell surface, and various membrane components, such as cell periphery, cellular anatomical entity, plasma membrane, cell surface, extracellular region, membrane. Downregulated CC items were mainly related to synapses and cell connection components, such as synapse, cell junction, synaptic membrane, neurofilament, postsynapse, and postsynaptic membrane. 2) Upregulated BP items were mainly related to immune defense responses such as defense response, immune system process, response to external stimulus, response to stress, immune response, response to other organisms, response to external biotic stimulus, defense response to other organisms, response to biotic stimulus, interspecies interaction between organisms. Downregulated BP items involved multiple processes such as animal organ morphogenesis, response to steroid hormone, positive regulation of epithelial to mesenchymal transition, cell adhesion, positive regulation of cell differentiation, biological adhesion, water transport. 3) Upregulated MF items mainly involved various substance binding and enzyme activities, such as protein binding, binding, peptide antigen binding, identical protein binding, peptide binding, amide binding, enzyme inhibitor activity, GTPase activity, protein-containing complex binding, signaling receptor binding. Downregulated MF items mainly involved the transporter activity and substance binding, such as inorganic molecular entity transmembrane transporter activity, cation transmembrane transporter activity, carbohydrate binding, transmembrane transporter activity, MHC protein complex binding, ion transmembrane transporter activity, coreceptor activity, water transmembrane transporter activity, transporter activity, and steroid binding.
3.1.3 Pathway enrichment analysis in the TB model group vs. the normal group
We selected the top 10 upregulated pathways ranked by enrichment score in the TB model group vs. normal group, and then counted the enrichment scores of these pathways in the JHW group vs. TB model group and each dose of NBXH group vs. TB model group (Table 5). Most pathways were related to host immune responses, such as phagosome, antigen processing and presentation, and NOD-like receptor signaling pathway. After summarizing the DE genes involved in each pathway, we found that although these pathways involved various functions, the changed targets in each pathway were certain, mainly related to immune defense responses such as cytokines, complement, and MHC pathways (Figure 2A).
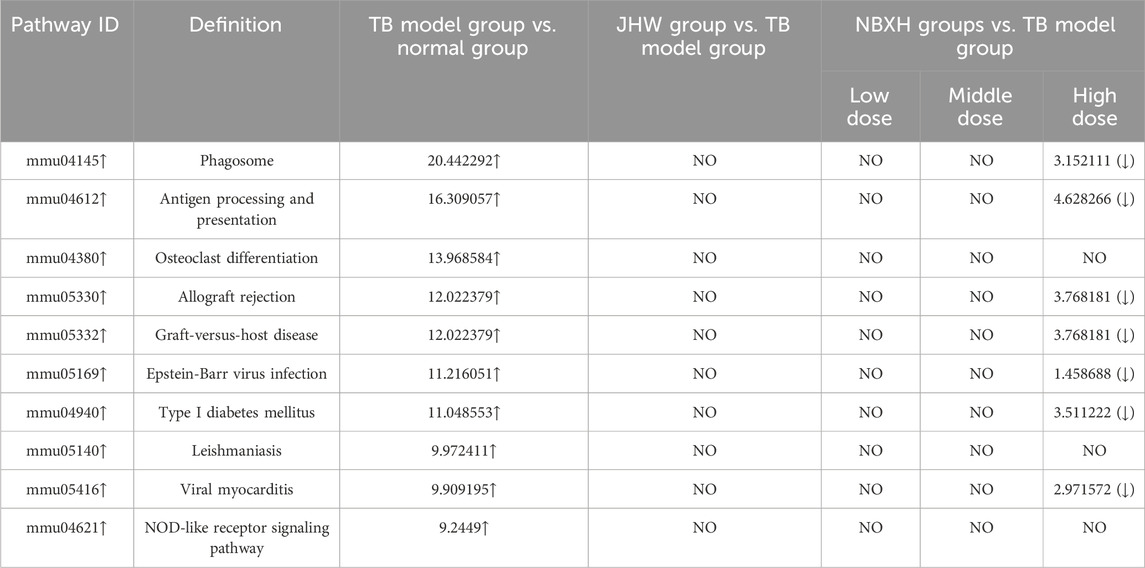
Table 5. The top 10 significantly upregulated pathways in TB model group and their changes in JHW group and various NBXH groups.
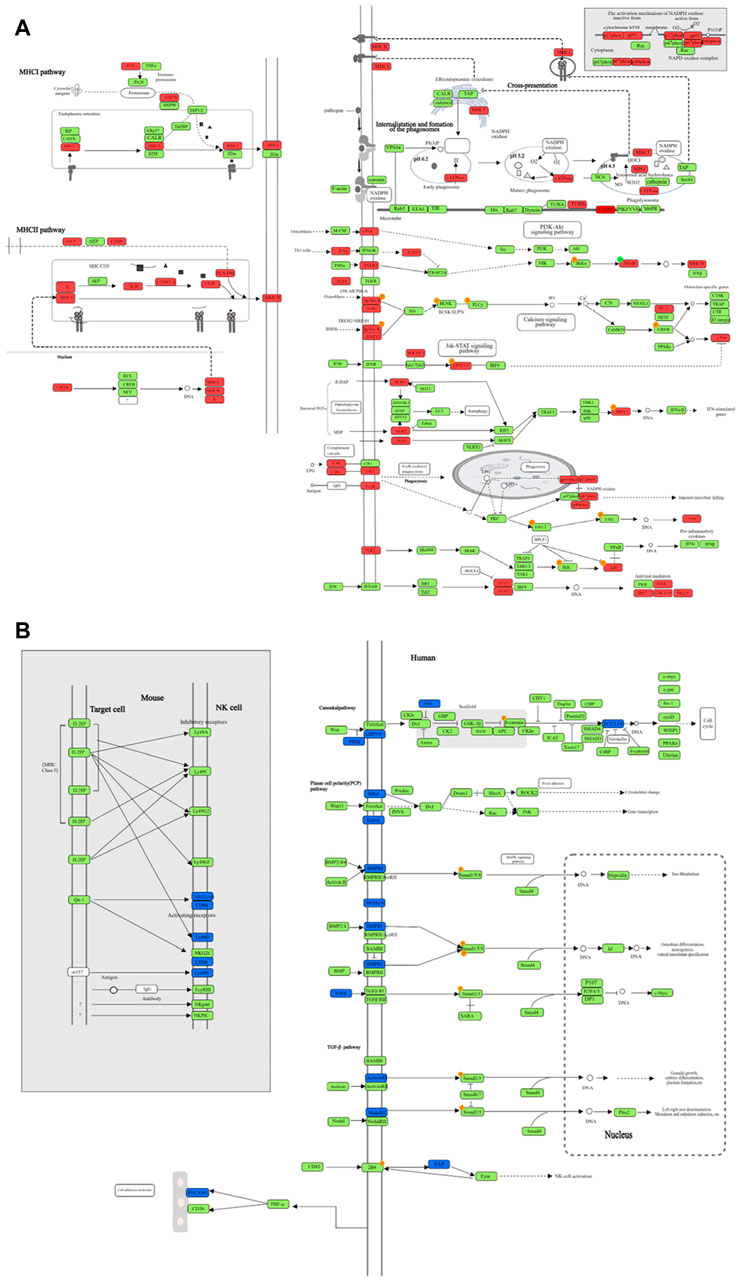
Figure 2. The DE genes in significantly upregulated and downregulated pathways after MTB infection. (A) Each pathway shown in Table 5, and the significantly upregulated DE genes in each pathway after MTB infection; (B) Each pathway shown in Table 6, and the significantly downregulated DE genes in each pathway after MTB infection. Red represents the upregulated DE genes, green represents unchanged genes, and blue represents downregulated DE genes.
In the TB model group vs. normal group, there were only five downregulated pathways (Table 6), and all the pathways involved were related to host immune responses, such as natural killer cell-mediated cytotoxicity, Wnt signaling pathway, and TGF-β signaling pathway. After summarizing the DE genes involved in the five pathways (Figure 2B), we found that the targets with downregulated differential expression mainly presented in NK cell surface receptors, Wnt signaling pathway, and TGF-β signaling pathway.
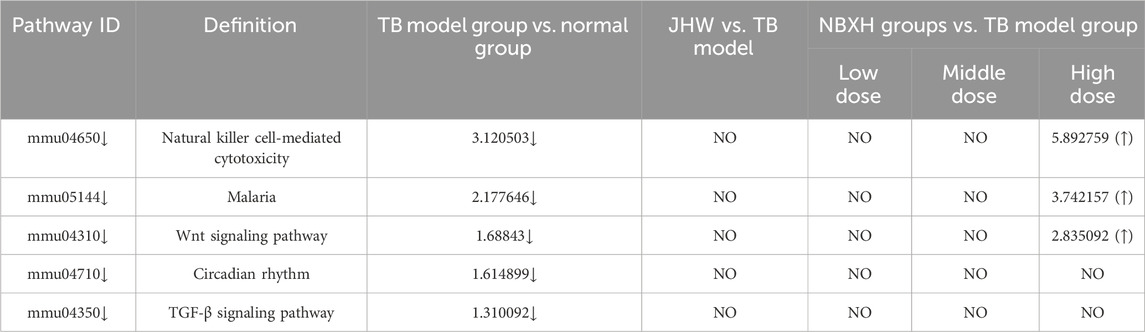
Table 6. The five downregulated pathways in TB model group and their changes in JHW group and various NBXH group.
3.1.4 Verifying the DE genes after MTB infection through GEO database
In this study, significantly upregulated or downregulated DE genes in the TB model group vs. the normal group may also be confirmed in the GSE54992, GSE48027, GSE98461, GSE83456, GSE62525, GSE34608, GSE14361 datasets (Table 7). For example, significantly upregulated DE genes (camp, cxcl9, cfb, gm4951, c4b, gm4841, ngp, serpina3g, and iigp1b) after MTB infection in this study were also significantly upregulated in multiple databases, and significantly downregulated DE genes (sult1d1 and hhatl) after MTB infection were also downregulated in the GSE14316 database. The gene mcrs1 was significantly downregulated in both this study and GSE34608, but upregulated in GSE48027.
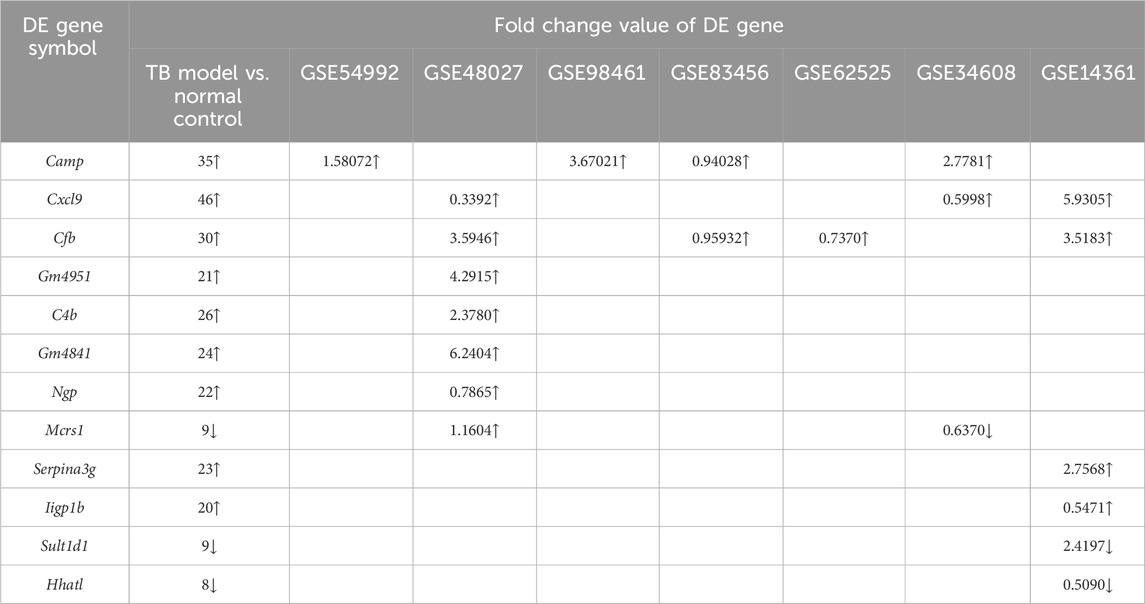
Table 7. Validation of DE genes by GEO database.
3.1.5 Verifying the DE genes after MTB infection through RT-qPCR
The relative transcript expression levels of six genes (cxcl9, camp, cfb, cxxc4, ulk3, and cd302) in the TB model group vs. the normal group, were analyzed in the TB patients and healthy individuals by RT-qPCR to verify the reliability of profile results and ifn-γ was used as a positive control (Figure 3). The results showed that: In initially treated TB patients, the expressions of ifn-γ, cxcl9, and cfb were significantly higher than those of HCs (p < 0.0001, p < 0.0001, p < 0.001), and the expressions of camp, cxxc4, and ulk3 were significantly lower than those of HCs (p < 0.05). However, there was no significant difference in the expression of cd302 was no significance from that of compared to HCs (p > 0.05). The results of cxcl9, cfb, cxxc4, and ulk3 verification were consistent with the results of gene expression profiles, while the results of camp verification were contrary to the results of expression profiles, and the results of cd302 verification showed no significance.
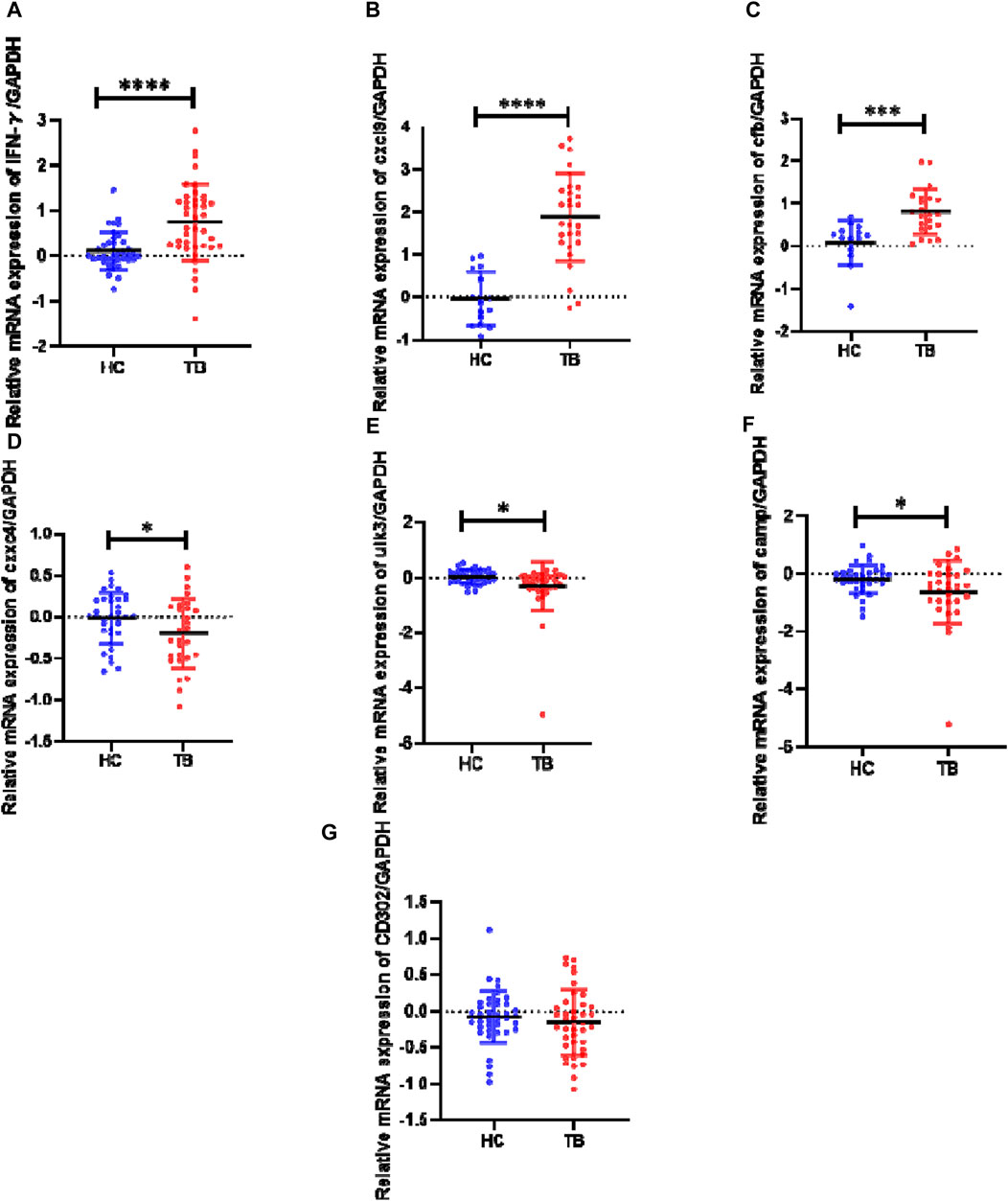
Figure 3. Expression levels of genes ifn-γ, cxcl9, cfb, cxxc4, ulk3, camp, and cd302 in the human PBMC samples as determined by RT-qPCR HC (healthy controls): negative IGRA and without abnormality in lung CT; TB (tuberculosis): initial treatment TB patients with positive IGRA or smear or Xpert. The data between the two groups were tested by Student’s t-test. Results of cytokine relative expression were expressed as a scatter chart, in which (A) means ifn-γ (p-value < 0.0001); (B) means cxcl9 (p-value < 0.0001); (C) means cfb (p-value = 0.0003); (D) means cxxc4 (p-value = 0.0493); (E) means ulk3 (p-value = 0.0227); (F) means camp (p-value = 0.0362); (G) means cd302 (p-value = 0.4306).
3.2 Immune responses in mice after NBXH treatment
3.2.1 DE genes in various dose NBXH groups
After treatment with low, middle, and high doses of NBXH, mice infected with MTB showed significant changes in gene expression in vivo (Table 2), and we found a negative correlation between the therapeutic dose of NBXH and the number of DE genes. Figure 4 reveals the number of overlapped DE genes between each dose of the NBXH group vs. TB model group and TB model group vs. normal group. The results showed that there were 50, 63, and 98 DE genes upregulated in the TB model group vs. normal group but downregulated in the low, middle, and high-dose NBXH groups vs. TB model group, respectively; there were 135, 72, and 95 DE genes downregulated in TB model group vs. normal group but upregulated in the low, middle, and high-dose NBXH groups vs. TB model group, respectively.
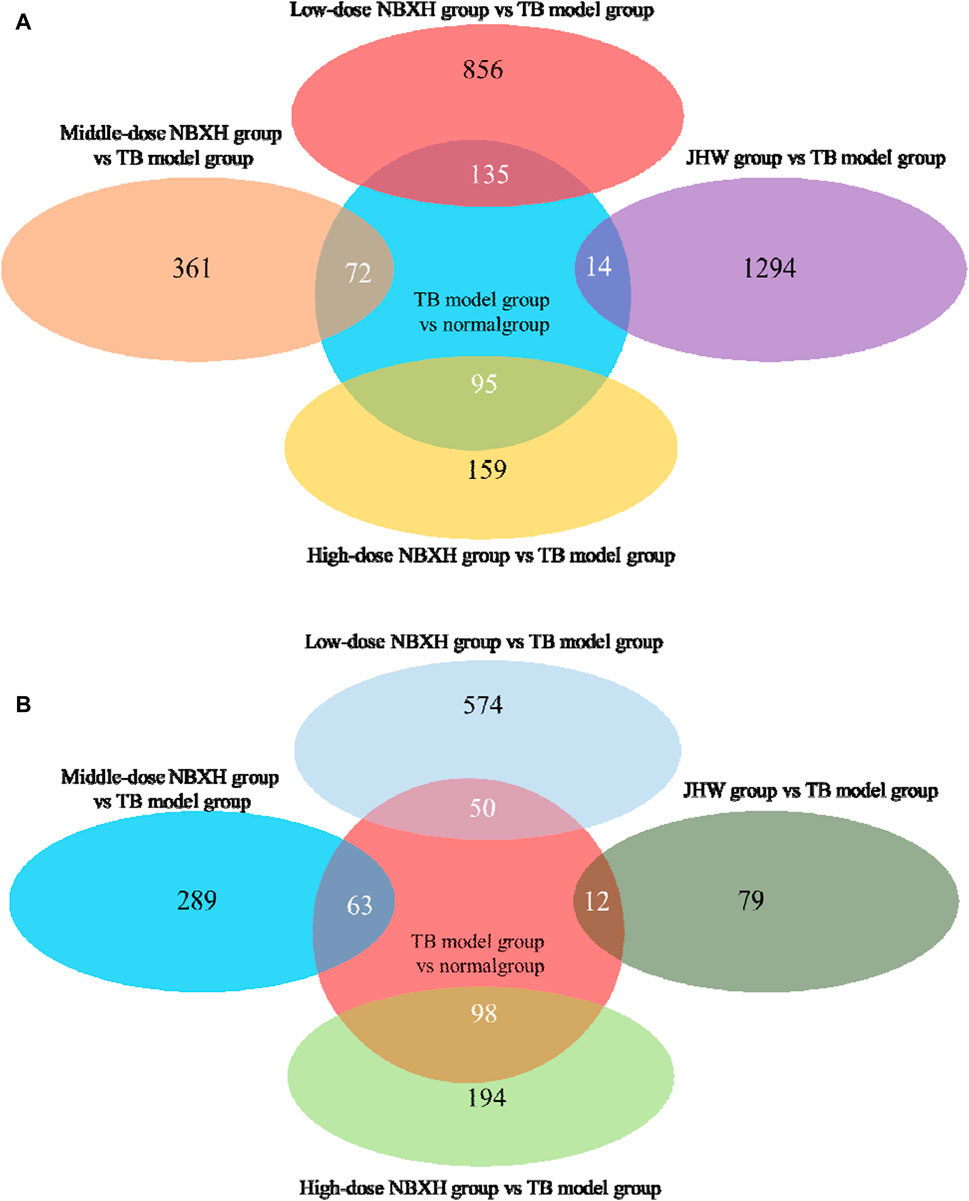
Figure 4. The DE gene Venn diagram between TB model group vs. normal group, JHW group vs. TB model group, and each dose NBXH group vs. TB model group. (A) Downregulated DE genes in the TB model group vs. normal group and upregulated DE genes in JHW group vs. TB model group and each NBXH group vs. TB model group; (B) Upregulated DE genes in the TB model group vs. normal group and downregulated DE genes in JHW group vs. TB model group and each NBXH group vs. TB model group, intersecting parts mean overlapping DE genes.
In addition, the top 10 upregulated and downregulated DE genes in the TB model group vs. normal group and their changes in low, middle, and high-dose NBXH groups vs. TB model group (Tables 3, 4) showed that: 1) The upregulated DE genes in the TB model group vs. normal group became downregulated in high-dose NBXH group vs. TB model group but had no significant difference in low-dose NBXH group vs. TB model group, middle-dose NBXH group vs. TB model group, and JHW group vs. TB model group. 2) The downregulated DE genes in the TB model group vs. normal group had no significant difference in the low-dose NBXH group vs. TB model group and the JHW group vs. TB model group, while most of the downregulated DE genes were significantly upregulated in the middle-dose NBXH group vs. TB model group and high-dose NBXH group vs. TB model group.
Therefore, we focused on the DE gene changes after the treatment with high-dose NBXH. The top 10 DE genes in the high-dose NBXH group vs. TB model group showed (Supplementary Tables S1, S2): 1) Of the top 10 upregulated DE genes in the high-dose NBXH group vs. TB model group, there also were 1, 5, and six upregulated DE genes in the JHW group vs. TB model group, low-dose NBXH group vs. TB model group, middle-dose NBXH group vs. TB model group, respectively. However, 9 DE genes were downregulated in the TB model group vs. the normal group, mainly related to information transmission in the nervous system and regulation of immune system function. 2) Of the top 10 downregulated DE genes in the high-dose NBXH group vs. TB model group, there also were 1, 4, and eight downregulated DE genes in the JHW group vs. TB model group, low-dose NBXH group vs. TB model group, and middle-dose NBXH group vs. TB model group, respectively. However, 6 DE genes were upregulated in the TB model group vs. the normal group, mainly related to nerve and endocrine regulation.
Although the low-dose NBXH group vs. TB model group had the most DE genes, there was no significant recovery effect on gene expression disorders caused by MTB infection. Middle-dose NBXH had a recovery effect on gene expression abnormalities caused by MTB infection, but the number or the FC values of recovered DE genes were not as well as those of high-dose NBXH. Although the high-dose NBXH group had the least number of DE genes, it had a significant corrective effect on most abnormal gene expression and pathways caused by MTB infection.
3.2.2 GO analysis in various dose NBXH groups
The top 10 GO items in low-, middle-high-dose NBXH groups vs. TB model group, and JHW group vs. TB model group (Supplementary Figure S2) showed that: 1) Of the top 10 downregulated GO items in the low-dose, middle-dose, and high-dose NBXH group vs. TB model group, and JHW group vs. TB model group, there were 0, 4, 4, and four upregulated GO items in the TB model group vs. normal group, respectively. Of the top 10 upregulated GO items in the low-dose, middle-dose, and high-dose NBXH groups vs. TB model group, and JHW group vs. TB model group, there were 6, 2, 10, and 6 downregulated GO items in the TB model group vs. normal group, respectively. 2) In the high-dose NBXH group vs. TB model group, the upregulated CC items were mainly related to various membrane components, extracellular regions, and cell surfaces, such as membrane, cell surface, side of membrane, intrinsic component of membrane, and integral component of membrane; while the downregulated CC items were mainly associated with organelles and MHC molecules, such as organelle, MHC protein complex, extracellular region, MHC class I protein complex, MHC class I peptide loading complex, and intracellular organelle. 3) In the high-dose NBXH group vs. TB model group, the upregulated BP items were mainly related to the negative regulatory process and immunity, such as negative regulation of cellular process, cell adhesion, biological adhesion, negative regulation of biomineral tissue development, negative regulation of biomineralization, negative regulation of biological process, natural killer cell-mediated immunity, and regulation of epidermal growth factor-activated receptor activity; while the downregulated BP items involved various processes of occurrence and development, antigen processing and presentation, etc., such as cell differentiation, cellular developmental process, developmental process, anatomical structure development, TAP-independent antigen processing and presentation of endogenous peptide antigen via MHC class I via ER pathway, positive regulation of biological process; canonical Wnt signaling pathway. 4) In the high-dose NBXH group vs. TB model group, the upregulated MF items mainly involved various substance binding and enzyme activities, such as binding, MHC protein complex binding, phosphatidylinositol kinase activity, filamin binding, transferase activity (GTP-Rho binding), and MAP kinase activity; while the downregulated MF items mainly involved various substances binding, such as molecular function regulator, peptide antigen binding, CD8 receptor binding, β-2-microglobulin binding, TAP binding, binding, β-catenin binding, serine-type endopeptidase inhibitor activity, intracellular calcium activated chloride channel activity, and γ-catenin binding.
3.2.3 Pathway enrichment analysis in various dose NBXH groups
The eight upregulated pathways in the high-dose NBXH group vs. TB model group and their changes in other dose NBXH groups vs. TB model group, TB model group vs. normal group, and JHW group vs. TB model group were shown in Supplementary Table S3, most of them were related to host immune response and endocrine such as natural killer cell-mediated cytotoxicity, parathyroid hormone synthesis, secretion and action. After summarizing the DE genes involved in eight pathways (Figure 5A), we found that although these pathways were involved in various functions, the changed genes in each pathway were certain, mainly related to cytokines and NK cell receptors such as CD94 and IFNsR. The top 10 downregulated pathways in the high-dose NBXH group vs. TB model group were shown in Supplementary Table S4, which were also related to host immunity and endocrine, such as antigen processing and presentation and phagosome. After summarizing the DE genes involved in the 10 pathways (Figure 5B), we found that downregulated DE genes were mainly observed in MHC molecular-related pathways.
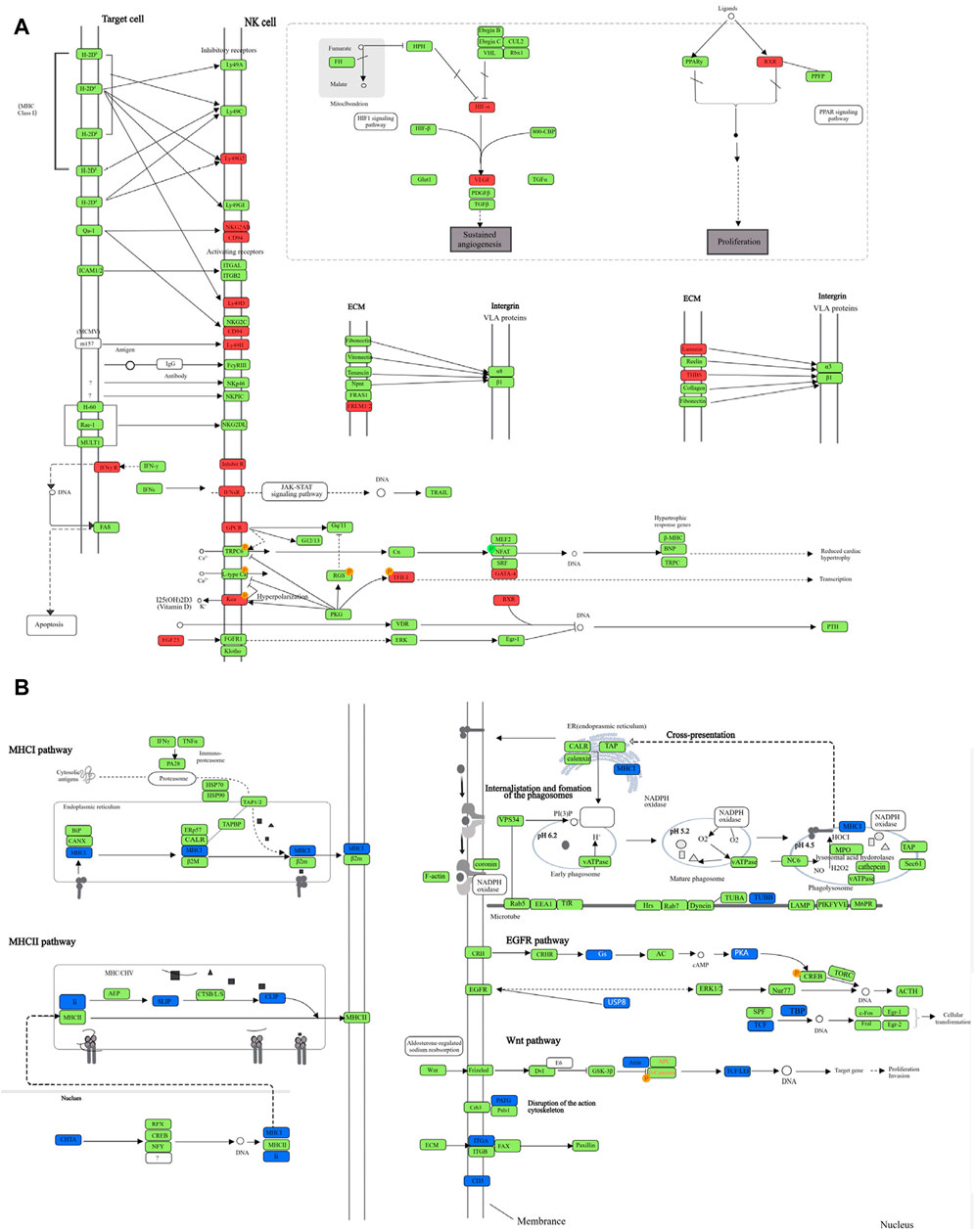
Figure 5. The DE genes in significantly upregulated and downregulated pathways after NBXH high dose group vs. TB model group. (A) The diagram of each pathway shown in Supplementary Table S3, and the significantly upregulated DE genes in each pathway after the high-dose NBXH treatment on mouse TB model; (B) The diagram of each pathway shown in Supplementary Table S4, and the significantly downregulated DE genes in each pathway after the high-dose NBXH treatment on mouse TB model. Red represents the upregulated DE genes, green represents unchanged genes, and blue represents downregulated DE genes.
3.2.4 Verifying treatment-related DE genes through GEO database
Some significantly upregulated or downregulated DE genes in the high-dose NBXH group vs. TB model group were also confirmed in the GSE54992, GSE62147, and GSE48027 datasets (Table 8). For example, DE genes (cd302, gm4841, ina, and cdh2) significantly upregulated after MTB infection were significantly downregulated after high-dose NBXH treatment and chemotherapy in multiple datasets. Among some DE genes significantly downregulated after MTB infection, lrrc74b and ulk3 were significantly upregulated after high-dose NBXH treatment and chemotherapy in both datasets, while nrxn3 was significantly upregulated after high-dose NBXH treatment, but still showed downregulation after chemotherapy in the GSE54992 and GSE62147 datasets.
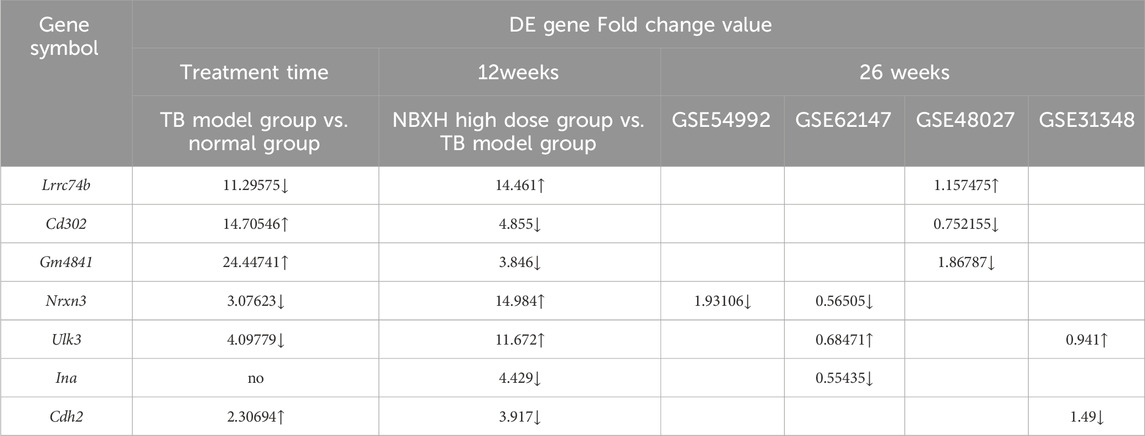
Table 8. Validation of the DE genes in the high-dose NBXH group vs. TB model group by GEO databases.
3.2.5 Verifying treatment-related DE genes through RT-qPCR
Using ifn-γ as the positive control group, DE gene cxcl9, cfb, cxxc4, and ulk3 were further verified by RT-qPCR before and after chemotherapy combined with NBXH treatment in TB patients (Figure 6). The results showed that: the expressions of ifn-γ and ulk3 showed no significant changes before and after treatment (p = 0.8381, p = 0.8888), while the expression of cxcl9 was significantly upregulated (p-value = 0.0188), and the expression of cfb also showed an upregulated trend but without significant difference (p-value = 0.5534), and the expression of cxxc4 was downregulated but also without significant difference (p-value = 0.2022).
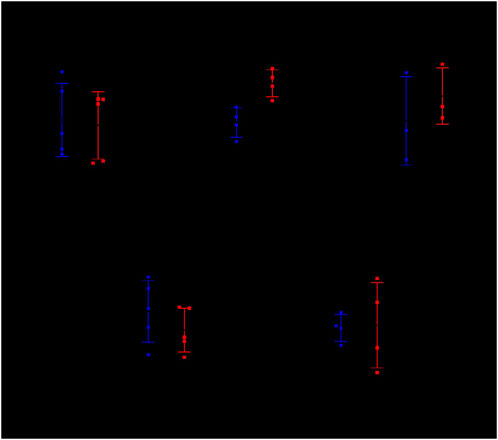
Figure 6. Transcript levels of ifn-γ, cxcl9, cfb, cxxc4, and ulk3 genes in the human PBMCs samples as determined by RT-qPCR.
TB patients before treatment (BT): initially treated pulmonary TB patients with positive IGRA or smear or Xpert; TB patients after treatment (AT): initially treated pulmonary TB patients with positive IGRA or smear or Xpert after treatment for ≥2 months.
The data between the two groups were tested by a Student’s t-test. The results were expressed as a scatter chart, where A means ifn-γ relative expression (p-value = 0.8381); B means cxcl9 relative expression (p-value = 0.0188); C means cfb relative expression (p-value = 0.5534); D means cxxc4 relative expression (p-value = 0.2022); E means ulk3 relative expression (p-value = 0.8888).
4 Discussion
TCM combined with chemotherapeutics in the treatment of TB has achieved significant clinical efficacy. For example, JHW, a Chinese medicine preparation, has been applied to clinics with a positive anti-TB therapeutic effect. Our previous studies have proved that NBXH has significant effects on the treatment of TB and alleviates the side effects of chemotherapy (Liang et al., 2017; Liang et al., 2021). This study attempts to explain the mechanism of the host responses against MTB infection in mice and the mechanisms of TCM preparation NBXH in the early treatment of acute MTB-infected mouse model through the gene expression microarray technology, to provide scientific explanation and verification for TCM.
4.1 Activation of anti-TB immunity in mice after MTB infection
After the mice were infected with MTB 3 months, the top 10 significantly upregulated DE genes in the TB model group vs. normal group were mainly related to anti-TB immune responses (Table 2). Cxcl9, encoding the monokine induced by interferon-γ (MIG), was the most significantly upregulated DE gene. CXCL9 binds to its receptor CXCR3 to recruit CXCR3+ cells, such as monocytes, CD4+ Th1 cells, and CD8+ cytotoxic T Cells, inducing Th1 immunity and inhibiting Th2 immunity (De Benedetti et al., 2021). It participates in immune regulation and inflammatory processes and plays an important role in host anti-TB immunity. Several studies have shown that cxcl9 gene expression and serum levels were significantly upregulated in hosts after MTB infection (Fan et al., 2022; Khalid et al., 2023; Li et al., 2023; Sampath et al., 2023). In this study, both the results of the GEO database and RT-qPCR validation also showed that the expression of the cxcl9 gene was significantly upregulated after MTB infection, and cxcl9 can serve as a potential diagnostic marker of MTB infection. Other studies have shown that cxcl9 gene expression and serum levels were significantly downregulated after effective treatment, which can be used as potential therapeutic monitoring biomarkers (Ambreen et al., 2021; Khalid et al., 2023). The cxcl9 expression in our result was consistent with other study, which means the treatment was effective. However, the opposite result in RT-qPCR assay may indicate that the patient’s condition has not been fully controlled and further treatment is needed. Cationic antibacterial peptide (CAMP), a member of the antimicrobial peptide family (AMP), is mainly expressed in epithelial cells (such as intestine, airway, skin, etc.,) and innate immune cells (such as neutrophils, natural killer cells, mast cells, dendritic cells, monocytes, and macrophages). As an endogenous effector molecule in innate immunity, CAMP can directly inhibit pathogen activity and regulate host innate and adaptive immune responses, including inducing pro-inflammatory effects, promoting autophagy, etc., (Bucki et al., 2010; Chen et al., 2017; Bei et al., 2019). Therefore, CAMP has broad application prospects in anti-TB treatment due to its broad-spectrum antibacterial activity (Dlozi et al., 2022). Our results and the results of 3 GEO datasets indicated that CAMP plays an important role in resisting MTB invasion. However, the opposite result in RT-qPCR assay may be related to the suppression of immune system function in TB patients. In addition to cxcl9 and camp, the significantly upregulated DE genes in the TB model group vs. normal group also included cfb (Shi et al., 2022), c4b (Daher et al., 2015; Wang et al., 2023), and ngp (Kathamuthu et al., 2021), which are closely related to the host’s anti-TB immune function. The complement system is an important component of innate immunity. Complement factor B (CFB) and complement C4b are members of the complement family, playing important roles in mobilizing both innate and adaptive immunity against pathogen invasion (Conigliaro et al., 2019). CFB is activated through the alternative complement pathway (AP) (Matsumoto et al., 1997), then activates the NF-κb inflammatory pathway to secrete inflammatory cytokines and recruit macrophages (Wang et al., 2009). C4b is activated through the classical complement pathway (CP) and lectin pathway (LP). The C1 complex cleaves C4 to produce C4a and C4b, and C4b further activates C3, initiating a series of complement cascade reactions, and forming complement complexes to exert immune function (Werner and Criss, 2023). Our results of the animal experiments, GEO datasets, and RT-qPCR validation all proved that the expressions of cfb and c4b were significantly upregulated after MTB infection, indicating that the host activates innate immune responses and participates in inflammation responses to clear invasive pathogens through the alternative pathway and the classical activation pathway, respectively (Utaiwat et al., 2021). Neutrophilic granule protein (NGP), a member of the cystatin superfamily, is mainly expressed in granulocytes (Moscinski and Hill, 1995). NGP can enhance phagocytosis of macrophages, act as an antagonist of lipopolysaccharide (LPS), activate TLR4-NF-κb signaling pathway to induce IL-10 secretion, inhibit TNF-α and IL-1β secretion (Liu et al., 2020), enhance the host’s immune defense ability, and inhibit inflammatory responses. The GEO validation results also showed that ngp was highly expressed in TB patients to participate in inflammatory responses. Serpina3g, a member of the serine protease inhibitor superfamily, participates in various immune activities. As a target of the JAG-STAT signaling pathway, SERPINA3G plays an important role in inducing GM-CSF synthesis and regulating the function of neutrophils and B lymphocytes, promoting anti-inflammatory, antioxidant, and anti-fibrotic effects (Li et al., 2014). Meanwhile, the expression level of serpina3g has also been reported to be positively correlated with the degree of pulmonary injury induced by chronic obstructive pulmonary disease (COPD) (Uysal and Uzun, 2019). GEO validation results also showed that serpina3g was highly expressed in TB patients, which proved that this gene played an important role in the inflammatory response of TB.
After MTB infection in mice for 3 months, the top 10 significantly downregulated DE genes mainly involved stress responses, cellular processes, and immune responses, etc., (Table 3). Among them, SULT1D1, as a sulfonyltransferase, is mainly expressed in the kidneys and livers of mice. Its main biological function is to regulate the sulfonation metabolism of catecholamines, playing an important role in the inactivation of endogenous dopamine derived compounds (including catecholamines) (Tsoi et al., 2001; Wong et al., 2010). GEO verification results showed that the expression of sult1d1 was downregulated in TB patients, which may reduce the inactivation of catecholamines, thus maintaining the stress response ability of TB patients. CXXC finger protein 4 (CXXC4) is the negative regulatory factor of Wnt/β-catenin signaling pathway and is also able to suppress the activation of NF-κB and p38 mitogen activated protein kinase (MAPK) pathway (Han et al., 2017; Li et al., 2020c), which are involved in the chronic inflammatory responses of TB (Holscher et al., 2020; Poladian et al., 2023). It was clearly confirmed through RT-qPCR validation. CXXC4 deficiency was the host’s desire to activate Wnt/β-Catenin signaling pathway to enhance the activities of the NF-κB and MAPK pathways, which promotes macrophage activation, phagosome maturation, and the release of various cytokines to control MTB growth (Holscher et al., 2020; Poladian et al., 2023). Grip2 is a key regulatory factor in the development of innate CD8+ T Cells (i8Ts) (Jo et al., 2020), which is nonconventional αβT cells that exhibit features of memory phenotype. Downregulation of grip2 expression by MTB infection will affect the differentiation and maturation of CD8+ T Cells.
The results of GO analysis and pathway analysis also confirmed that the NK cell surface receptors, Wnt signal pathway, and TGF-β signal pathway were downregulated in mice after MTB infection, which weakens multiple processes such as adhesion, differentiation, endocrine, and transportation, and also promoted the host’s recognition, processing, and presentation of antigens, as well as immune defense responses. The body was also striving to mobilize the immune system to resist damage caused by MTB invasion. The impact of MTB infection on host gene expression is comprehensive, even disrupting gene expression in nervous and endocrine systems. The neurohumoral regulatory system extensively regulates immune system function. In recent years, more and more researchers have paid attention to the regulatory role of the neurohumoral system on the immune response caused by exogenous pathogen infections (Huang et al., 2019; Malin et al., 2020; Kim et al., 2022). For example, activation of the cholinergic anti-inflammatory pathway (CAP) can limit lung infection (Su et al., 2010), and reduce inflammatory damage, as well as inhibit the NF-κB pathway in splenic macrophages to reduce inflammatory responses while inhibiting the migration of splenic macrophages (Borovikova et al., 2000; Wang et al., 2003). In addition to the vagus nerve, the sympathetic and parasympathetic nerves can also regulate inflammatory responses. Gang Cao et al.’s research showed that neuron-derived neuropeptide Y (NPY) can precisely regulate the inflammatory responses of the spleen, acting as the "interaction language” between the immune system and central nervous system (Yu et al., 2022). Therefore, the DE genes of the nervous and endocrine systems that exhibit significant changes after MTB infection may also be involved in the neuroimmune regulation process, and we will further investigate and clarify it.
4.2 Relationship between NBXH dose and effect
The dose-effect relationship results showed: 1) The dosage change of NBXH influenced the therapeutic effect, and within a certain dosage range, the therapeutic effect will have a positive correlation with the change of NBXH dosage. The dosage of TCM is an extremely important part of TCM prescriptions, which is closely related to clinical efficacy. Chen JQ et al. and Chen YY et al. used metabolomics technology to clarify the dose-effect relationship of TCM (Chen et al., 2020; Chen et al., 2022). Our study firstly revealed the dose-response relationship through transcriptomics, then combined with traditional pharmacological and toxicological studies can provide a basis for determining the clinical dosage of NBXH granules. 2) High-dose NBXH can enhance immune system function, reduce inflammatory damage, and play an anti-TB effect. JHW also had a partial recovery effect on the abnormal gene expression caused by MTB infection, but the recovery effect is not as good as high-dose NBXH. Moreover, the targets and regulatory pathways of the anti-TB effects of JHW and NBXH are different, indicating that these two TCM prescriptions exert anti-TB effects through different mechanisms of action.
4.3 Anti-TB mechanism of NBXH
The most significantly upregulated DE gene in high-dose NBXH group was nrxn3, which encodes a nerve cell adhesion molecule necessary for synaptic formation and maintenance, playing a role in the neuromuscular junction, synaptic formation, maturation, differentiation, and intercellular signaling processes. Therefore, high or low nrxn3 expression is associated with a variety of neurological and psychiatric diseases (Kasem et al., 2018). The lack of NRXN3 can affect synaptic development, synaptic signaling, and neurotransmitter release, and the downregulation of nrxn3 expression has been identified as the highest-risk gene associated with Alzheimer disease (AD) and senescence (Zheng et al., 2018). Meanwhile, nrxn3 is also involved in the occurrence or metastasis of various tumors (Sun et al., 2013). There is currently no literature reporting its role in pathogen infection or host anti-infection. This study found for the first time that nrxn3 expression was downregulated after MTB infection. The current study showed that TB patients had a higher risk of AD than non-TB patients. TB may be associated with an increased risk of AD, and inflammation may play a key role in AD. The GEO verification results showed that nrxn3 expression in TB patients decreased after chemotherapy, but current studies have shown that rifampicin can have therapeutic effects on AD through anti-inflammatory, anti-tau, anti-amyloid, and cholinergic effects (Yulug et al., 2018). High-dose NBXH therapy significantly upregulates nrxn3, which may reduce the risk of AD and is worthy of further exploration. Ulk3, as a feature gene of autophagy, is a key target of the mTOR signaling pathway, and its expression can activate the cellular autophagy process (Gonzalez-Rodriguez et al., 2022), which is an important part of host anti-TB immunity (Lam et al., 2017). The results of GEO and RT-qPCR indicate that both chemotherapy and NBXH treatment can enhance the anti-TB autophagy effect of the body. In this study, it was found for the first time that btbd17, krt72, otop3t, and gm10050 were significantly downregulated after MTB infection, while significantly upregulated after NBXH treatment. There were few reports on these genes, and no TB-related dataset was found in the GEO database, so further research is needed.
There are currently no reports in the field of TB on the top 10 significantly downregulated DE genes after high-dose NBXH treatment, and most genes can also not be retrieved from relevant datasets in the GEO database. Among them, mageb16, cdkn2b, and cdh2 genes are associated with tumors. The mageb16 gene, most significantly downregulated in this study, usually encodes tumor antigens and is also a DE gene or protein in many genomics research results such as gastric cancer (Zaragoza-Huesca et al., 2022), colorectal cancer (Juhari et al., 2021), bladder cancer (Liu et al., 2016), and other tumor diseases, as well as feline abdominal virus infection (Shuid et al., 2015), indicating that it may play an important role in tumorigenesis. Cdkn2b encodes the tumor suppressor gene p15ink4b, which is closely linked to the cdkn2a chromosome and participates in the cell cycle and senescence. Cdkn2b is often inactivated due to deletion, methylation, or mutation in various tumors, leading to tumor progression and being considered an inhibitor of tumors (Jang et al., 2019; Yang et al., 2021; Hjazi et al., 2023). Cdh2, a member of the Cadherin family, regulates many biological processes and has been proven to be closely associated with various cancers (Kourtidis et al., 2017). Cancer patients with high cdh2 expression usually have poorer prognosis, while those with low cdh2 expression have improved prognosis (Ma et al., 2016; Chen et al., 2018). This study found for the first time that MTB infection does not affect mageb16 expression, but cdkn2b and cdh2 expressions are both significantly upregulated. It was suggested that persistent tissue damage caused by long-term chronic inflammation of TB, cell proliferation induced by damage, and cicatrix formed by tissue repair were closely related to the occurrence of lung cancer. In addition to the damage recovery effect, further exploration is needed to determine whether NBXH also has a certain anti-cancer effect. Three genes related to anti-TB immunity, serpinb6a, cd302, and gm4841, were significantly upregulated after MTB infection. Serpinb6a can inhibit cathepsin G (CatG) activity in monocyte and neutrophil-mediated immune inflammatory responses, prevent cell apoptosis, promote cell survival, and regulate inflammatory responses (Burgener et al., 2019). Therefore, downregulation of serpinb6a expression after NBXH treatment can enhance macrophage inflammatory response and apoptosis, which is beneficial for clearing invading MTB and reducing the inflammatory damage caused by neutrophils. CD302, a protein containing a C-type lectin domain, is an important pattern recognition receptor (PRR) in the innate immune system. Its biological function is related to cell adhesion and migration, as well as phagocytosis and endocytosis, and it participates in the host’s resistance to the invasion and infection of various pathogenic microorganisms. It is mainly highly expressed in the liver, lungs, lymph nodes, spleen, and myeloid cells (such as monocytes, granulocytes, macrophages, and DC cells) (Kato et al., 2007; Lo et al., 2016; Peng et al., 2023). After NBXH treatment, the elevated cd302 was downregulated by one-third, indicating that NBXH regulated the host’s anti-TB responses by inhibiting the phagocytic and bactericidal activity of monocytes/macrophages. In addition, gprc5b, which is widely expressed in the pancreas, is an important regulatory factor for pancreatic function, and its high expression leads to β cells being sensitive to the harmful effects of cytokines and increases the apoptosis induced by cytokines, which is related to type II diabetes. The downregulation of cd302 can increase insulin secretion and promote β cell survival, which makes it an attractive target for developing new medicine to treat type II diabetes and its complications (such as diabetes nephropathy) (Soni et al., 2013; Atanes et al., 2018; Tao et al., 2022). NBXH treatment can downregulate gprc5b expression, which is beneficial to prevent TB with diabetes. There are also three genes (svs3a, vmn1r65, and ina) related to molecular activity or cell structure. Currently, there are few research reports, and their role in anti-TB is not yet clear. GEO verification results only have confirmed that ina was significantly downregulated after treatment. Therefore, these genes as targets for NBXH treatment still need further exploration to explain their clinical significance.
The GO analysis and pathway analysis results showed that the antigen processing and presentation process was downregulated by a quarter compared to the TB model group vs. the normal group and NK cell-mediated cytotoxicity was significantly enhanced. The above analysis indicates that NBXH treatment can enhance anti-TB immunity such as autophagy, NK cell function, and apoptosis, which is beneficial for clearing intracellular MTB (Qin et al., 2023). At the same time, NBXH also had a certain inhibitory effect on immune regulatory targets that are overactivated during immune system activation.
In conclusion, MTB infection led to widespread changes in host gene expression and mainly stimulated the host’s anti-TB immune responses. The treatment using high-dose NBXH partially repaired the abnormal gene expression caused by MTB infection, further strengthened the anti-TB immunity in areas such as autophagy and NK cell-mediated cytotoxicity, and had a certain inhibitory effect on overactivated immune regulatory factors to alleviate immune damage. This study is the first to reveal the dose-response relationship of NBXH through transcriptomics, and further elucidate the mechanism of MTB infection and NBXH therapy, providing the scientific basis for TCM treatment of TB.
Data availability statement
The authors confirm that the data supporting the findings of this study are available within the article/Supplementary Material. The transcriptome data is available at the GEO database, accession number: GSE276937. Further enquiries can be directed to the corresponding authors.
Ethics statement
The studies involving humans were approved by the Ethics Committee of the Eighth Medical Center of the PLA General Hospital (309202108101027). The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study. The animal study was approved by The Animal Ethics Committee of the Eighth Medical Center of the PLA General Hospital (309202108101027). The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
NW: Formal Analysis, Methodology, Writing–original draft, Writing–review and editing. QM: Formal Analysis, Methodology, Validation, Writing–original draft. JZ: Methodology, Writing–original draft. JW: Methodology, Writing–original draft. XL: Supervision, Writing–original draft. YL: Data curation, Supervision, Writing–original draft. XW: Funding acquisition, Project administration, Resources, Supervision, Writing–review and editing.
Funding
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphar.2024.1415951/full#supplementary-material
References
Ambreen, A., Khaliq, A., Naqvi, S. Z. H., Tahir, A., Mustafa, M., Chaudhary, S. U., et al. (2021). Host biomarkers for monitoring therapeutic response in extrapulmonary tuberculosis. Cytokine 142, 155499. doi:10.1016/j.cyto.2021.155499
An, H. R., Bai, X. J., Liang, J. Q., Wang, T., Wang, Z. Y., Xue, Y., et al. (2022). The relationship between absolute counts of lymphocyte subsets and clinical features in patients with pulmonary tuberculosis. Clin. Respir. J. 16 (5), 369–379. doi:10.1111/crj.13490
An, H.-R., Wu, X., Wang, Z., Xu, J., Zheng, S., et al. (2013). The clinical characteristics of anti-tuberculosis drug induced liver injury in 2457 hospitalized patients with tuberculosis in China. Afr. J. Pharm. Pharmacol. 7, 710–714. doi:10.5897/AJPP2013.2963
Arora, G., Bothra, A., Prosser, G., Arora, K., and Sajid, A. (2021). Role of post-translational modifications in the acquisition of drug resistance in Mycobacterium tuberculosis. FEBS J. 288 (11), 3375–3393. doi:10.1111/febs.15582
Atanes, P., Ruz-Maldonado, I., Hawkes, R., Liu, B., Persaud, S. J., and Amisten, S. (2018). Identifying signalling pathways regulated by GPRC5B in β-cells by CRISPR-cas9-mediated genome editing. Cell Physiol. Biochem. 45 (2), 656–666. doi:10.1159/000487159
Barao, I., Alvarez, M., Ames, E., Orr, M. T., Stefanski, H. E., Blazar, B. R., et al. (2011). Mouse Ly49G2+ NK cells dominate early responses during both immune reconstitution and activation independently of MHC. Blood 117 (26), 7032–7041. doi:10.1182/blood-2010-11-316653
Barao, I., Wright, P. W., Sungur, C. M., Anderson, S. K., Redelman, D., and Murphy, W. J. (2013). Differential expression of the Ly49G(B6), but not the Ly49G(BALB), receptor isoform during natural killer cell reconstitution after hematopoietic stem cell transplantation. Biol. Blood Marrow Transpl. 19 (10), 1446–1452. doi:10.1016/j.bbmt.2013.07.021
Bei, Y., Pan, L. L., Zhou, Q., Zhao, C., Xie, Y., Wu, C., et al. (2019). Cathelicidin-related antimicrobial peptide protects against myocardial ischemia/reperfusion injury. BMC Med. 17 (1), 42. doi:10.1186/s12916-019-1268-y
Blankley, S., Graham, C. M., Turner, J., Berry, M. P. R., Bloom, C. I., Xu, Z., et al. (2016). The transcriptional signature of active tuberculosis reflects symptom status in extra-pulmonary and pulmonary tuberculosis. PLoS One 11 (10), e0162220. doi:10.1371/journal.pone.0162220
Borovikova, L. V., Ivanova, S., Zhang, M., Yang, H., Botchkina, G. I., Watkins, L. R., et al. (2000). Vagus nerve stimulation attenuates the systemic inflammatory response to endotoxin. Nature 405 (6785), 458–462. doi:10.1038/35013070
Bucki, R., Leszczyńska, K., Namiot, A., and Sokołowski, W. (2010). Cathelicidin LL-37: a multitask antimicrobial peptide. Arch. Immunol. Ther. Exp. Warsz. 58 (1), 15–25. doi:10.1007/s00005-009-0057-2
Buglino, J. A., and Resh, M. D. (2008). Hat is a palmitoylacyltransferase with specificity for N-palmitoylation of Sonic Hedgehog. J. Biol. Chem. 283 (32), 22076–22088. doi:10.1074/jbc.M803901200
Burgener, S. S., Leborgne, N. G. F., Snipas, S. J., Salvesen, G. S., Bird, P. I., and Benarafa, C. (2019). Cathepsin G inhibition by Serpinb1 and Serpinb6 prevents programmed necrosis in neutrophils and monocytes and reduces GSDMD-driven inflammation. Cell Rep. 27 (12), 3646–3656 e5. doi:10.1016/j.celrep.2019.05.065
Cai, L., Li, Z., Guan, X., Cai, K., Wang, L., Liu, J., et al. (2019). The research progress of host genes and tuberculosis susceptibility. Oxid. Med. Cell Longev. 2019, 9273056. doi:10.1155/2019/9273056
Cai, Y., Yang, Q., Tang, Y., Zhang, M., Liu, H., Zhang, G., et al. (2014). Increased complement C1q level marks active disease in human tuberculosis. PLoS One 9 (3), e92340. doi:10.1371/journal.pone.0092340
Chen, J. Q., Chen, Y. Y., Tao, H. J., Pu, Z. J., Shi, X. Q., Zhang, J., et al. (2020b). An integrated metabolomics strategy to reveal dose-effect relationship and therapeutic mechanisms of different efficacy of rhubarb in constipation rats. J. Pharm. Biomed. Anal. 177, 112837. doi:10.1016/j.jpba.2019.112837
Chen, Q., Cai, J., and Jiang, C. (2018). CDH2 expression is of prognostic significance in glioma and predicts the efficacy of temozolomide therapy in patients with glioblastoma. Oncol. Lett. 15 (5), 7415–7422. doi:10.3892/ol.2018.8227
Chen, T., Zhong, F., Yao, C., Chen, J., Xiang, Y., Dong, J., et al. (2020a). A systematic review on traditional uses, sources, phytochemistry, pharmacology, pharmacokinetics, and toxicity of fritillariae cirrhosae bulbus. Evid. Based Complement. Altern. Med. 2020, 1536534. doi:10.1155/2020/1536534
Chen, X., Qi, G., Qin, M., Zou, Y., Zhong, K., Tang, Y., et al. (2017). DNA methylation directly downregulates human cathelicidin antimicrobial peptide gene (CAMP) promoter activity. Oncotarget 8 (17), 27943–27952. doi:10.18632/oncotarget.15847
Chen, Y. Y., Chen, J. Q., Tang, Y. P., Shang, E. X., Zhao, Q., Zou, J. B., et al. (2022). Integrated dose-response metabolomics with therapeutic effects and adverse reactions may demystify the dosage of traditional Chinese medicine. Chin. Med. 17 (1), 130. doi:10.1186/s13020-022-00687-4
Choi, J. H., Jin, S. W., Choi, C. Y., Kim, H. G., Kim, S. J., Lee, H. S., et al. (2017). Saponins from the roots of Platycodon grandiflorum ameliorate high fat diet-induced non-alcoholic steatohepatitis. Biomed. Pharmacother. 86, 205–212. doi:10.1016/j.biopha.2016.11.107
Cliff, J. M., Lee, J. S., Constantinou, N., Cho, J. E., Clark, T. G., Ronacher, K., et al. (2013). Distinct phases of blood gene expression pattern through tuberculosis treatment reflect modulation of the humoral immune response. J. Infect. Dis. 207 (1), 18–29. doi:10.1093/infdis/jis499
Conigliaro, P., Triggianese, P., Ballanti, E., Perricone, C., Perricone, R., and Chimenti, M. S. (2019). Complement, infection, and autoimmunity. Curr. Opin. Rheumatol. 31 (5), 532–541. doi:10.1097/BOR.0000000000000633
Daher, R. K., Stewart, G., Boissinot, M., Boudreau, D. K., and Bergeron, M. G. (2015). Influence of sequence mismatches on the specificity of recombinase polymerase amplification technology. Mol. Cell Probes 29 (2), 116–121. doi:10.1016/j.mcp.2014.11.005
De Benedetti, F., Prencipe, G., Bracaglia, C., Marasco, E., and Grom, A. A. (2021). Targeting interferon-gamma in hyperinflammation: opportunities and challenges. Nat. Rev. Rheumatol. 17 (11), 678–691. doi:10.1038/s41584-021-00694-z
Dlozi, P. N., Gladchuk, A., Crutchley, R. D., Keuler, N., Coetzee, R., and Dube, A. (2022). Cathelicidins and defensins antimicrobial host defense peptides in the treatment of TB and HIV: pharmacogenomic and nanomedicine approaches towards improved therapeutic outcomes. Biomed. Pharmacother. 151, 113189. doi:10.1016/j.biopha.2022.113189
Duan, L. Y., Liang, Y., Gong, W. P., Xue, Y., Mi, J., Wang, J., et al. (2021). Comparative study on the antituberculous effect and mechanism of the traditional Chinese medicines NiuBeiXiaoHe extract and JieHeWan. Mil. Med. Res. 8 (1), 34. doi:10.1186/s40779-021-00324-5
Fan, J., Yang, Y., Wang, L., Shang, X., Zhang, L., Sun, H., et al. (2022). Study on the correlation between interleukin-27 and CXCL10 in pulmonary tuberculosis. J. Immunol. Res. 2022, 2932837. doi:10.1155/2022/2932837
Fouillade, C., Baron-Menguy, C., Domenga-Denier, V., Thibault, C., Takamiya, K., Huganir, R., et al. (2013). Transcriptome analysis for Notch3 target genes identifies Grip2 as a novel regulator of myogenic response in the cerebrovasculature. Arterioscler. Thromb. Vasc. Biol. 33 (1), 76–86. doi:10.1161/ATVBAHA.112.251736
Gao, Q., Yang, M., and Zuo, Z. (2018). Overview of the anti-inflammatory effects, pharmacokinetic properties and clinical efficacies of arctigenin and arctiin from Arctium lappa L. Acta Pharmacol. Sin. 39 (5), 787–801. doi:10.1038/aps.2018.32
Gonzalez-Rodriguez, P., Cheray, M., Keane, L., Engskog-Vlachos, P., and Joseph, B. (2022). ULK3-dependent activation of GLI1 promotes DNMT3A expression upon autophagy induction. Autophagy 18 (12), 2769–2780. doi:10.1080/15548627.2022.2039993
Han, M., Dai, D., Yousafzai, N. A., Wang, F., Wang, H., Zhou, Q., et al. (2017). CXXC4 activates apoptosis through up-regulating GDF15 in gastric cancer. Oncotarget 8 (61), 103557–103567. doi:10.18632/oncotarget.21581
Hjazi, A., Ghaffar, E., Asghar, W., Alauldeen Khalaf, H., Ikram Ullah, M., Mireya Romero-Parra, R., et al. (2023). CDKN2B-AS1 as a novel therapeutic target in cancer: mechanism and clinical perspective. Biochem. Pharmacol. 213, 115627. doi:10.1016/j.bcp.2023.115627
Holscher, C., Gräb, J., Hölscher, A., Müller, A. L., Schäfer, S. C., and Rybniker, J. (2020). Chemical p38 MAP kinase inhibition constrains tissue inflammation and improves antibiotic activity in Mycobacterium tuberculosis-infected mice. Sci. Rep. 10 (1), 13629. doi:10.1038/s41598-020-70184-x
Huang, C. J., Lyu, X., and Kang, J. (2023). The molecular characteristics and functional roles of microspherule protein 1 (MCRS1) in gene expression, cell proliferation, and organismic development. Cell Cycle 22 (6), 619–632. doi:10.1080/15384101.2022.2145816
Huang, Y., Zhao, C., and Su, X. (2019). Neuroimmune regulation of lung infection and inflammation. QJM 112 (7), 483–487. doi:10.1093/qjmed/hcy154
Jang, W., Park, J., Kwon, A., Choi, H., Kim, J., Lee, G. D., et al. (2019). CDKN2B downregulation and other genetic characteristics in T-acute lymphoblastic leukemia. Exp. Mol. Med. 51 (1), 1–15. doi:10.1038/s12276-018-0195-x
Jiang, S., Chen, C. F., Ma, X. P., Wang, M. Y., Wang, W., Xia, Y., et al. (2019). Antibacterial stilbenes from the tubers of Bletilla striata. Fitoterapia 138, 104350. doi:10.1016/j.fitote.2019.104350
Jiang, Y., Benz, T. L., and Long, S. B. (2021). Substrate and product complexes reveal mechanisms of Hedgehog acylation by HHAT. Science 372 (6547), 1215–1219. doi:10.1126/science.abg4998
Jo, Y., Balmer, L., Lee, B., Shim, J. A., Ali, L. A., Morahan, G., et al. (2020). Variants of innate CD8(+) T cells are associated with Grip2 and Klf15 genes. Cell Mol. Immunol. 17 (9), 1007–1009. doi:10.1038/s41423-019-0357-3
Juhari, W. K. W., Ahmad Amin Noordin, K. B., Zakaria, A. D., Rahman, W. F. W. A., Mokhter, W. M. M. W. M., Hassan, M. R. A., et al. (2021). Whole-genome profiles of Malay colorectal cancer patients with intact MMR proteins. Genes (Basel) 12 (9), 1448. doi:10.3390/genes12091448
Kasem, E., Kurihara, T., and Tabuchi, K. (2018). Neurexins and neuropsychiatric disorders. Neurosci. Res. 127, 53–60. doi:10.1016/j.neures.2017.10.012
Kathamuthu, G. R., Moideen, K., Sridhar, R., Baskaran, D., and Babu, S. (2021). Reduced neutrophil granular proteins and post-treatment modulation in tuberculous lymphadenitis. PLoS One 16 (6), e0253534. doi:10.1371/journal.pone.0253534
Kato, M., Khan, S., d'Aniello, E., McDonald, K. J., and Hart, D. N. J. (2007). The novel endocytic and phagocytic C-Type lectin receptor DCL-1/CD302 on macrophages is colocalized with F-actin, suggesting a role in cell adhesion and migration. J. Immunol. 179 (9), 6052–6063. doi:10.4049/jimmunol.179.9.6052
Khalid, H., Pierneef, L., van Hooij, A., Zhou, Z., de Jong, D., Tjon Kon Fat, E., et al. (2023a). Development of lateral flow assays to detect host proteins in cattle for improved diagnosis of bovine tuberculosis. Front. Vet. Sci. 10, 1193332. doi:10.3389/fvets.2023.1193332
Khalid, S., Ambreen, A., Khaliq, A., Ullah, H., Mustafa, M., and Mustafa, T. (2023b). Identification of host biomarkers from dried blood spots for monitoring treatment response in extrapulmonary tuberculosis. Sci. Rep. 13 (1), 599. doi:10.1038/s41598-022-26823-6
Kim, J. K., Park, E. J., and Jo, E. K. (2022). Itaconate, arginine, and gamma-aminobutyric acid: a host metabolite triad protective against mycobacterial infection. Front. Immunol. 13, 832015. doi:10.3389/fimmu.2022.832015
Kourtidis, A., Lu, R., Pence, L. J., and Anastasiadis, P. Z. (2017). A central role for cadherin signaling in cancer. Exp. Cell Res. 358 (1), 78–85. doi:10.1016/j.yexcr.2017.04.006
Lam, A., Prabhu, R., Gross, C. M., Riesenberg, L. A., Singh, V., and Aggarwal, S. (2017). Role of apoptosis and autophagy in tuberculosis. Am. J. Physiol. Lung Cell Mol. Physiol. 313 (2), L218–L229. doi:10.1152/ajplung.00162.2017
Lee, S. W., Wu, L. S. H., Huang, G. M., Huang, K. Y., Lee, T. Y., and Weng, J. T. Y. (2016). Gene expression profiling identifies candidate biomarkers for active and latent tuberculosis. BMC Bioinforma. 17 (Suppl. 1), 3. doi:10.1186/s12859-015-0848-x
Li, H., Ren, W., Liang, Q., Zhang, X., Li, Q., Shang, Y., et al. (2023). A novel chemokine biomarker to distinguish active tuberculosis from latent tuberculosis: a cohort study. QJM 116, 1002–1009. doi:10.1093/qjmed/hcad214
Li, L., Byrne, S. M., Rainville, N., Su, S., Jachimowicz, E., Aucher, A., et al. (2014). Brief report: serpin Spi2A as a novel modulator of hematopoietic progenitor cell formation. Stem Cells 32 (9), 2550–2556. doi:10.1002/stem.1778
Li, P., Ge, D., Hu, F., Chu, J., Chen, X., et al. (2020b). CXXC finger protein 4 inhibits the CDK18-ERK1/2 axis to suppress the immune escape of gastric cancer cells with involvement of ELK1/MIR100HG pathway. J. Cell Mol. Med. 24 (17), 10151–10165. doi:10.1111/jcmm.15625
Li, P., Luo, X., Xie, Y., Hu, F., Chu, J., et al. (2020c). GC-derived EVs enriched with MicroRNA-675-3p contribute to the MAPK/PD-L1-Mediated tumor immune escape by targeting CXXC4. Mol. Ther. Nucleic Acids 22, 615–626. doi:10.1016/j.omtn.2020.08.020
Li, W., and Yang, H. J. (2021). Phenolic constituents from Platycodon grandiflorum root and their anti-inflammatory activity. Molecules 26 (15), 4530. doi:10.3390/molecules26154530
Li, X., Liu, Q., Sun, N., Zhang, B., Shi, C., et al. (2020a). Traditional Chinese medicine combined with western medicine for the treatment of secondary pulmonary tuberculosis: a PRISMA-compliant meta-analysis. Med. Baltim. 99 (16), e19567. doi:10.1097/MD.0000000000019567
Li, Y., Wang, Q., Wei, H. C., Liang, Y. Y., Niu, F. J., Li, K. W., et al. (2022). Fructus arctii: an overview on its traditional uses, pharmacology and phytochemistry. J. Pharm. Pharmacol. 74 (3), 321–336. doi:10.1093/jpp/rgab140
Liang, Y., Gong, W., Wang, X., Zhang, J., Ling, Y., Song, J., et al. (2021). Chinese traditional medicine NiuBeiXiaoHe (NBXH) extracts have the function of antituberculosis and immune recovery in BALB/c mice. J. Immunol. Res. 2021, 6234560. doi:10.1155/2021/6234560
Liang, Y., Wang, X., Song, J., Wang, L., Chen, D., Yang, Y., et al. (2017). Therapeutic effects of traditional Chinese medicine Niubeixiaohe in mouse tuberculosis models. J. Ethnopharmacol. 195, 318–323. doi:10.1016/j.jep.2016.11.037
Liu, K., Tian, L. X., Tang, X., Wang, J., Tang, W. Q., Ma, Z. F., et al. (2020). Neutrophilic granule protein (NGP) attenuates lipopolysaccharide-induced inflammatory responses and enhances phagocytosis of bacteria by macrophages. Cytokine 128, 155001. doi:10.1016/j.cyto.2020.155001
Liu, M., Li, M., Liu, J., Wang, H., Zhong, D., Zhou, H., et al. (2016). Elevated urinary urea by high-protein diet could be one of the inducements of bladder disorders. J. Transl. Med. 14, 53. doi:10.1186/s12967-016-0809-9
Lo, T. H., Silveira, P. A., Fromm, P. D., Verma, N. D., Vu, P. A., Kupresanin, F., et al. (2016). Characterization of the expression and function of the C-type lectin receptor CD302 in mice and humans reveals a role in dendritic cell migration. J. Immunol. 197 (3), 885–898. doi:10.4049/jimmunol.1600259
Ma, T., Zhao, Y., Wei, K., Yao, G., Pan, C., Liu, B., et al. (2016). MicroRNA-124 functions as a tumor suppressor by regulating CDH2 and epithelial-mesenchymal transition in non-small cell lung cancer. Cell Physiol. Biochem. 38 (4), 1563–1574. doi:10.1159/000443097
Maertzdorf, J., Weiner, J., Mollenkopf, H. J., Bauer, T., Prasse, A., et al. (2012). Common patterns and disease-related signatures in tuberculosis and sarcoidosis. Proc. Natl. Acad. Sci. U. S. A. 109 (20), 7853–7858. doi:10.1073/pnas.1121072109
Malin, S. G., Shavva, V. S., Tarnawski, L., and Olofsson, P. S. (2020). Functions of acetylcholine-producing lymphocytes in immunobiology. Curr. Opin. Neurobiol. 62, 115–121. doi:10.1016/j.conb.2020.01.017
Manca, C., Koo, M. S., Peixoto, B., Fallows, D., Kaplan, G., and Subbian, S. (2013). Host targeted activity of pyrazinamide in Mycobacterium tuberculosis infection. PLoS One 8 (8), e74082. doi:10.1371/journal.pone.0074082
Matsumoto, M., Fukuda, W., Circolo, A., Goellner, J., Strauss-Schoenberger, J., Wang, X., et al. (1997). Abrogation of the alternative complement pathway by targeted deletion of murine factor B. Proc. Natl. Acad. Sci. U. S. A. 94 (16), 8720–8725. doi:10.1073/pnas.94.16.8720
Moscinski, L. C., and Hill, B. (1995). Molecular cloning of a novel myeloid granule protein. J. Cell Biochem. 59 (4), 431–442. doi:10.1002/jcb.240590404
Pan, L., Wei, N., Jia, H., Gao, M., Chen, X., Wei, R., et al. (2017). Genome-wide transcriptional profiling identifies potential signatures in discriminating active tuberculosis from latent infection. Oncotarget 8 (68), 112907–112916. doi:10.18632/oncotarget.22889
Peng, J., Li, J., Liang, J., Li, W., Yang, Y., Yang, Y., et al. (2023). A C-type lectin-like receptor CD302 in yellow drum (Nibea albiflora) functioning in antibacterial activity and innate immune signaling. Int. J. Biol. Macromol. 247, 125734. doi:10.1016/j.ijbiomac.2023.125734
Pilla-Moffett, D., Barber, M. F., Taylor, G. A., and Coers, J. (2016). Interferon-Inducible GTPases in host resistance, inflammation and disease. J. Mol. Biol. 428 (17), 3495–3513. doi:10.1016/j.jmb.2016.04.032
Poladian, N., Orujyan, D., Narinyan, W., Oganyan, A. K., Navasardyan, I., Velpuri, P., et al. (2023). Role of NF-κB during Mycobacterium tuberculosis infection. Int. J. Mol. Sci. 24 (2), 1772. doi:10.3390/ijms24021772
Pontali, E., Raviglione, M. C., and Migliori, G. B. (2019). Regimens to treat multidrug-resistant tuberculosis: past, present and future perspectives. Eur. Respir. Rev. 28 (152), 190035. doi:10.1183/16000617.0035-2019
Qin, Y., Wang, Q., and Shi, J. (2023). Immune checkpoint modulating T cells and NK cells response to Mycobacterium tuberculosis infection. Microbiol. Res. 273, 127393. doi:10.1016/j.micres.2023.127393
Salazar-Olivo, L. A., Mejia-Elizondo, R., Alonso-Castro, A. J., Ponce-Noyola, P., Maldonado-Lagunas, V., Melendez-Zajgla, J., et al. (2014). SerpinA3g participates in the antiadipogenesis and insulin-resistance induced by tumor necrosis factor-α in 3T3-F442A cells. Cytokine 69 (2), 180–188. doi:10.1016/j.cyto.2014.05.025
Sampath, P., Rajamanickam, A., Thiruvengadam, K., Natarajan, A. P., Hissar, S., Dhanapal, M., et al. (2023). Plasma chemokines CXCL10 and CXCL9 as potential diagnostic markers of drug-sensitive and drug-resistant tuberculosis. Sci. Rep. 13 (1), 7404. doi:10.1038/s41598-023-34530-z
Schreiber, T., Ehlers, S., Heitmann, L., Rausch, A., Mages, J., Murray, P. J., et al. (2009). Autocrine IL-10 induces hallmarks of alternative activation in macrophages and suppresses antituberculosis effector mechanisms without compromising T cell immunity. J. Immunol. 183 (2), 1301–1312. doi:10.4049/jimmunol.0803567
Sekita, Y., Murakami, K., Yumoto, H., Mizuguchi, H., Amoh, T., Ogino, S., et al. (2016). Anti-bacterial and anti-inflammatory effects of ethanol extract from Houttuynia cordata poultice. Biosci. Biotechnol. Biochem. 80 (6), 1205–1213. doi:10.1080/09168451.2016.1151339
Shi, T., Huang, L., Zhou, Y., and Tian, J. (2022). Role of GBP1 in innate immunity and potential as a tuberculosis biomarker. Sci. Rep. 12 (1), 11097. doi:10.1038/s41598-022-15482-2
Shingnaisui, K., Dey, T., Manna, P., and Kalita, J. (2018). Therapeutic potentials of Houttuynia cordata Thunb. against inflammation and oxidative stress: a review. J. Ethnopharmacol. 220, 35–43. doi:10.1016/j.jep.2018.03.038
Shuid, A. N., Safi, N., Haghani, A., Mehrbod, P., Haron, M. S. R., Tan, S. W., et al. (2015). Apoptosis transcriptional mechanism of feline infectious peritonitis virus infected cells. Apoptosis 20 (11), 1457–1470. doi:10.1007/s10495-015-1172-7
Soni, A., Amisten, S., Rorsman, P., and Salehi, A. (2013). GPRC5B a putative glutamate-receptor candidate is negative modulator of insulin secretion. Biochem. Biophys. Res. Commun. 441 (3), 643–648. doi:10.1016/j.bbrc.2013.10.099
Su, X., Matthay, M. A., and Malik, A. B. (2010). Requisite role of the cholinergic alpha7 nicotinic acetylcholine receptor pathway in suppressing Gram-negative sepsis-induced acute lung inflammatory injury. J. Immunol. 184 (1), 401–410. doi:10.4049/jimmunol.0901808
Sun, H. T., Cheng, S. X., Tu, Y., Li, X. H., and Zhang, S. (2013). FoxQ1 promotes glioma cells proliferation and migration by regulating NRXN3 expression. PLoS One 8 (1), e55693. doi:10.1371/journal.pone.0055693
Tao, Y., Lin, Y., An, L., Li, Y., Han, J., et al. (2022). Knockdown of GPRC5B alleviates the high glucose-induced inflammation and extracellular matrix deposition of podocyte through inhibiting NF-κB pathway. Allergol. Immunopathol. Madr. 50 (2), 142–146. doi:10.15586/aei.v50i2.566
Tientcheu, L. D., Maertzdorf, J., Weiner, J., Adetifa, I. M., Mollenkopf, H. J., Sutherland, J. S., et al. (2015). Differential transcriptomic and metabolic profiles of M. africanum- and M. tuberculosis-infected patients after, but not before, drug treatment. Genes Immun. 16 (5), 347–355. doi:10.1038/gene.2015.21
Tsoi, C., Falany, C. N., Morgenstern, R., and Swedmark, S. (2001). Identification of a new subfamily of sulfotransferases: cloning and characterization of canine SULT1D1. Biochem. J. 356 (Pt 3), 891–897. doi:10.1042/0264-6021:3560891
Utaiwat, S., Senawong, G., Khongsukwiwat, K., Woranam, K., Sattayasai, J., and Senawong, T. (2021). Immunomodulatory potential of the industrialized Houttuynia cordata fermentation product in vitro and in wistar rats. Foods 10 (11), 2582. doi:10.3390/foods10112582
Uthaiah, R. C., Praefcke, G. J. K., Howard, J. C., and Herrmann, C. (2003). IIGP1, an interferon-gamma-inducible 47-kDa GTPase of the mouse, showing cooperative enzymatic activity and GTP-dependent multimerization. J. Biol. Chem. 278 (31), 29336–29343. doi:10.1074/jbc.M211973200
Uysal, P., and Uzun, H. (2019). Relationship between circulating Serpina3g, matrix metalloproteinase-9, and tissue inhibitor of metalloproteinase-1 and -2 with chronic obstructive pulmonary disease severity. Biomolecules 9 (2), 62. doi:10.3390/biom9020062
Wang, D., Yang, J., Du, Q., Li, H., and Wang, S. (2016). The total alkaloid fraction of bulbs of Fritillaria cirrhosa displays anti-inflammatory activity and attenuates acute lung injury. J. Ethnopharmacol. 193, 150–158. doi:10.1016/j.jep.2016.08.009
Wang, H., Yu, M., Ochani, M., Amella, C. A., Tanovic, M., Susarla, S., et al. (2003). Nicotinic acetylcholine receptor alpha7 subunit is an essential regulator of inflammation. Nature 421 (6921), 384–388. doi:10.1038/nature01339
Wang, J., Ohno-Matsui, K., Yoshida, T., Shimada, N., Ichinose, S., Sato, T., et al. (2009). Amyloid-beta up-regulates complement factor B in retinal pigment epithelial cells through cytokines released from recruited macrophages/microglia: another mechanism of complement activation in age-related macular degeneration. J. Cell Physiol. 220 (1), 119–128. doi:10.1002/jcp.21742
Wang, L., Zheng, X., Ma, J., Gu, J., and Sha, W. (2023). Comparative proteomic analysis of exosomes derived from patients infected with non-tuberculous Mycobacterium and Mycobacterium tuberculosis. Microorganisms 11 (9), 2334. doi:10.3390/microorganisms11092334
Werner, L. M., and Criss, A. K. (2023). Diverse functions of C4b-binding protein in health and disease. J. Immunol. 211 (10), 1443–1449. doi:10.4049/jimmunol.2300333
Wong, S., Tan, K., Carey, K. T., Fukushima, A., Tiganis, T., and Cole, T. J. (2010). Glucocorticoids stimulate hepatic and renal catecholamine inactivation by direct rapid induction of the dopamine sulfotransferase Sult1d1. Endocrinology 151 (1), 185–194. doi:10.1210/en.2009-0590
Yang, L., Ma, D. W., Cao, Y. P., Li, D. Z., Zhou, X., Feng, J. F., et al. (2021). PRMT5 functionally associates with EZH2 to promote colorectal cancer progression through epigenetically repressing CDKN2B expression. Theranostics 11 (8), 3742–3759. doi:10.7150/thno.53023
Yu, J., Xiao, K., Chen, X., Deng, L., Zhang, L., Li, Y., et al. (2022). Neuron-derived neuropeptide Y fine-tunes the splenic immune responses. Neuron 110 (8), 1327–1339 e6. doi:10.1016/j.neuron.2022.01.010
Yuan, Q., Wang, S. Q., Zhang, G. T., He, J., Liu, Z. D., Wang, M. R., et al. (2021). Highly expressed of SERPINA3 indicated poor prognosis and involved in immune suppression in glioma. Immun. Inflamm. Dis. 9 (4), 1618–1630. doi:10.1002/iid3.515
Yulug, B., Hanoglu, L., Ozansoy, M., Isık, D., Kilic, U., Kilic, E., et al. (2018). Therapeutic role of rifampicin in Alzheimer's disease. Psychiatry Clin. Neurosci. 72 (3), 152–159. doi:10.1111/pcn.12637
Zaragoza-Huesca, D., Garrido-Rodríguez, P., Jiménez-Fonseca, P., Martínez de Castro, E., Sánchez-Cánovas, M., Visa, L., et al. (2022). Identification of thrombosis-related genes in patients with advanced gastric cancer: data from AGAMENON-SEOM registry. Biomedicines 10 (1), 148. doi:10.3390/biomedicines10010148
Zhai, W., Wei, E., Li, R., Ji, T., Jiang, Y., Wang, X., et al. (2021). Characterization and evaluation of the pro-coagulant and immunomodulatory activities of polysaccharides from Bletilla striata. ACS Omega 6 (1), 656–665. doi:10.1021/acsomega.0c05171
Zhang, C., Ning, D., Pan, J., Chen, C., Gao, C., Ding, Z., et al. (2021). Anti-inflammatory effect fraction of Bletilla striata and its protective effect on LPS-induced acute lung injury. Mediat. Inflamm. 2021, 6684120. doi:10.1155/2021/6684120
Zhang, L., Cui, M., and Chen, S. (2020). Identification of the molecular mechanisms of peimine in the treatment of cough using computational target fishing. Molecules 25 (5), 1105. doi:10.3390/molecules25051105
Zhang, W., Hou, J., Yan, X., Leng, J., Li, R., Zhang, J., et al. (2018). Platycodon grandiflorum saponins ameliorate cisplatin-induced acute nephrotoxicity through the NF-κB-Mediated inflammation and PI3K/Akt/Apoptosis signaling pathways. Nutrients 10 (9), 1328. doi:10.3390/nu10091328
Zhang, Z., Xun, Y., Rong, S., Yan, L., SoRelle, J. A., Li, X., et al. (2022). Loss of immunity-related GTPase GM4951 leads to nonalcoholic fatty liver disease without obesity. Nat. Commun. 13 (1), 4136. doi:10.1038/s41467-022-31812-4
Keywords: tuberculosis, Chinese traditional medicine, NiuBeiXiaoHe, transcriptome, mechanism of action
Citation: Wang N, Ma Q, Zhang J, Wang J, Li X, Liang Y and Wu X (2024) Transcriptomics-based anti-tuberculous mechanism of traditional Chinese polyherbal preparation NiuBeiXiaoHe intermediates. Front. Pharmacol. 15:1415951. doi: 10.3389/fphar.2024.1415951
Received: 17 April 2024; Accepted: 29 July 2024;
Published: 19 September 2024.
Edited by:
Jianping Xie, Southwest University, ChinaCopyright © 2024 Wang, Ma, Zhang, Wang, Li, Liang and Wu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xiaojun Li, eGlhb2p1bndzd0BzaW5hLmNvbQ==; Yan Liang, YW15NTkxOUBzaW5hLmNvbQ==; Xueqiong Wu, eHVlcWlvbmd3dUAxMzkuY29t
†These authors have contributed equally to this work to this study