 Min Cao
Min Cao Ping Huang
Ping Huang Lun-shan Xu
Lun-shan Xu Yi-hua Zhang
Yi-hua Zhang- Department of Neurosurgery, Daping Hospital, Army Medical University, Chongqing, China
Pituitary neuroendocrine tumors (PitNETs) are a special class of tumors of the central nervous system that are closely related to metabolism, endocrine functions, and immunity. In this study, network pharmacology was used to explore the metabolites and pharmacological mechanisms of PitNET regulation by gut microbiota. The metabolites of the gut microbiota were obtained from the gutMGene database, and the targets related to the metabolites and PitNETs were determined using public databases. A total of 208 metabolites were mined from the gutMGene database; 1,192 metabolite targets were screened from the similarity ensemble approach database; and 2,303 PitNET-related targets were screened from the GeneCards database. From these, 392 overlapping targets were screened between the metabolite and PitNET-related targets, and the intersection between these overlapping and gutMGene database targets (223 targets) were obtained as the core targets (43 targets). Using the protein–protein interaction (PPI) network analysis, Kyoto encyclopedia of genes and genomes (KEGG) signaling pathway and metabolic pathway analysis, CXCL8 was obtained as a hub target, tryptophan metabolism was found to be a key metabolic pathway, and IL-17 signaling was screened as the key KEGG signaling pathway. In addition, molecular docking analysis of the active metabolites and target were performed, and the results showed that baicalin, baicalein, and compound K had good binding activities with CXCL8. We also describe the potential mechanisms for treating PitNETs using the information on the microbiota (Bifidobacterium adolescentis), signaling pathway (IL-17), target (CXCL8), and metabolites (baicalin, baicalein, and compound K); we expect that these will provide a scientific basis for further study.
1 Introduction
Pituitary neuroendocrine tumors (PitNETs) constitute a special class of tumors of the central nervous system that are closely related to metabolism, endocrine functions, and immunity (Nie et al., 2023). A recent epidemiological study showed that PitNETs affect more than 5% of the global population (Wang et al., 2023a). Intestinal flora interact with the central nervous system via the gut–brain axis, which includes the vagus nerve, enteric nervous system, immune system, and microbial metabolites. Gut microbiota are closely related to the functions of the central nervous system as well as the endocrine and neuroendocrine systems (Leistner and Menke, 2020; Tafet and Nemeroff, 2020; Wang et al., 2023b; Berding et al., 2021; Longo et al., 2023), but their relationships with PitNETs are not completely clear.
Research has shown that the gut microbiomes of patients with PitNETs are significantly different from those of healthy subjects. Compared with the control group, Clostridium innocuum was enriched while Oscillibacter sp.57_20 and Fusobacterium mortiferum populations were poorer in patients with invasive and non-invasive PitNETs (Hu et al., 2022). Another study showed that there were differences in the structures and quantities of intestinal flora among growth-hormone-secreting PitNET patients, non-functional PitNET patients, and healthy controls. In the mouse model, following transplantation of fecal microflora, the intestinal flora of growth-hormone-secreting PitNETs patients promoted the growth of tumor (Nie et al., 2022). Another study showed that compared with healthy controls, growth-hormone-secreting PitNETs not only reduce the alpha diversity of the intestinal flora but also change the beta diversity significantly (Lin et al., 2022). Among the metabolites, the short-chain fatty acid butyrate was shown to enhance growth hormone secretion in the rat anterior pituitary cells through activation of the G-protein-coupled receptors GPR41 and GPR43 (Miletta et al., 2014). These studies indicate that there are potential relationships among PitNETs, intestinal flora, and metabolites. Although some of these relationships have been reported in literature, their specific mechanisms and action pathways need to be clarified through further research. Network pharmacology can comprehensively consider the interactions between metabolites and multiple targets, thereby offering more insights into the comprehensive effect mechanisms (Oh et al., 2023).
In the present study, the key targets, signaling pathways, metabolites, and microbiota that regulate PitNETs were identified via network pharmacology, and their relationships were revealed. First, the gutMGene v1.0 database (http://bio-annotation.cn/gutmgene/) (Cheng et al., 2022) was used to retrieve the metabolites and targets related to the gut microbiota; then, the metabolite-related and PitNET-related targets were determined through public databases. Second, the overlapping targets between the metabolite-related and PitNET-related targets were obtained via Venn diagram; these overlapping targets were further intersected with the targets obtained from the gutMGene database via a Venn diagram to determine the final common targets. These final overlapping targets were considered as the core targets. Third, the protein–protein interaction (PPI) network and Kyoto encyclopedia of genes and genomes (KEGG) pathway enrichment diagram were constructed using the core targets; the metabolic pathways were then analyzed using the corresponding metabolites of the core targets to identify the key signaling pathways and targets in PitNETs. Once the key target was identified, molecular docking analysis was used to predict the potential metabolites that acted on the key target. Finally, the relationships among the identified microbiota, signaling pathways, targets, and metabolites were examined to reveal their roles in PitNETs. Therefore, this study is expected to be an important reference for further experimental research and regulation of metabolites with regard to PitNETs. The steps in the workflow of this study are shown in Figure 1A.
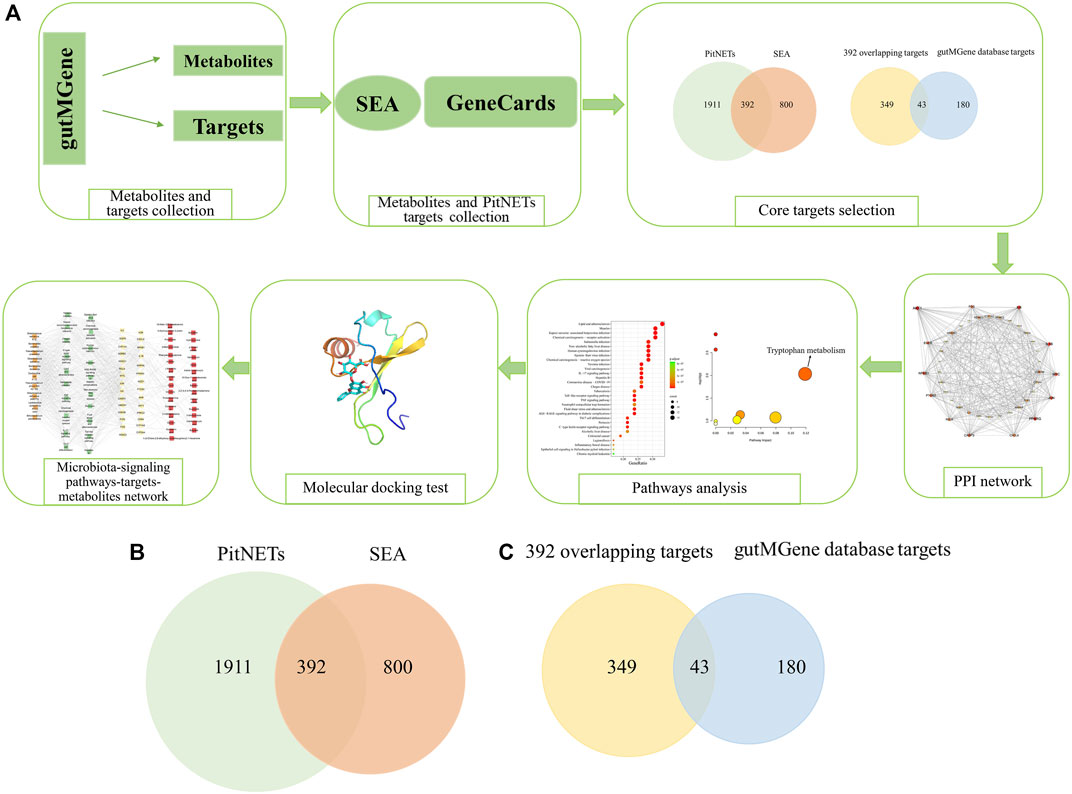
Figure 1. (A) Workflow of the steps in this study. (B) Overlapping targets (392) between metabolite targets predicted by the similarity ensemble approach (SEA) database and PitNET-related targets. (C) Identification of 43 core targets between the 392 overlapping targets and gutMGene database (223 targets).
2 Methods
2.1 Data collection and target acquisition
The metabolites and targets used in the analyses were retrieved from the gutMGene v1.0 database. The related targets of the metabolites were predicted using the similarity ensemble approach (SEA) (https://sea.bkslab.org/) (Keiser et al., 2007) database. The disease targets were obtained from the GeneCards database (https://www.genecards.org/) (Safran et al., 2010), and targets that were larger than the median were selected for subsequent analyses. The intersection of metabolite targets and disease targets were obtained by Venn analysis. Finally, the core targets were obtained from the intersection of all targets from the three databases; we believe that these core targets are important factors in PitNETs.
2.2 Protein–protein interaction network
The core targets were mapped to the PPI network to identify their interactions. The STRING database (https://string-db.org/) was used to perform the PPI analysis.
2.3 KEGG pathway and gene ontology (GO) analyses
R package (Yu et al., 2012) was used to perform KEGG pathway enrichment (Draghici et al., 2007) and GO analysis to analyze targets functions (Luo et al., 2017). Furthermore, the interactions between the key regulatory pathways were analyzed using R software.
2.4 Metabolic pathway analysis
MetaboAnalyst 6.0 (https://www.metaboanalyst.ca/) was used for the metabolic pathway analysis. Here, 31 metabolites corresponding to the 43 core targets were input into the online website for the pathway analysis.
2.5 Molecular docking test
The structured data files of the metabolites were acquired from the PubChem database (https://pubchem.ncbi.nlm.nih.gov/), and the target protein structure was downloaded from the PDB database (https://www1.rcsb.org/). The ligands and protein needed for molecular docking analysis were prepared using AutoDock, and the molecular docking was carried out using Vina in PyRx. The docking results were finally visualized using PyMOL software.
2.6 Construction of the microbiota-signaling pathways-targets-metabolites network
Based on the KEGG enrichment results of the 43 core targets, the top-20 significant signaling pathways were determined as the main signaling pathways. Then, the microbiota and metabolites that were directly related to these top-20 pathways’ targets were selected using the gutMGene database. We chose the top-10 results based on the degree of microbiota to construct the microbiota–signaling pathways–targets–metabolites network, which was then visualized using Cytoscape 3.10.1 (Shannon et al., 2003).
3 Results
3.1 Core targets of the gut microbial metabolites
A total of 208 metabolites were obtained from the gutMGene database. The targets related to these metabolites were identified using the SEA database, through which 1,192 metabolite targets were found. A total of 392 overlapping targets were then identified between the 1,192 metabolite targets and 2,303 PitNET-related targets, as shown in Figure 1B. Finally, the intersection between these 392 overlapping targets and 223 targets from the gutMGene database yielded 43 core targets that were used in the follow-up analyses (Figure 1C) (Supplementary Table S1).
3.2 Protein–protein interaction network analysis
The 43 core targets were used in the PPI network analysis to highlight the important proteins. The protein AKT1 has the highest degree of 31, indicating that it may be an important target related to PitNETs. To date, many studies have shown that AKT1 is closely related to PitNETs (Huang et al., 2020; Dworakowska et al., 2009). Therefore, our findings are consistent with those reported in previous studies. In the present study, the top-13 proteins based on degree are AKT1 (31), IL1B (29), IL6 (29), PPARG (29), JUN (27), MYC (26), NFKB1 (26), CASP3 (25), EGFR (24), PTGS2 (24), FOS (23), CXCL8 (22), and RELA (22) (Figure 2), indicating that these proteins may be related to PitNETs.
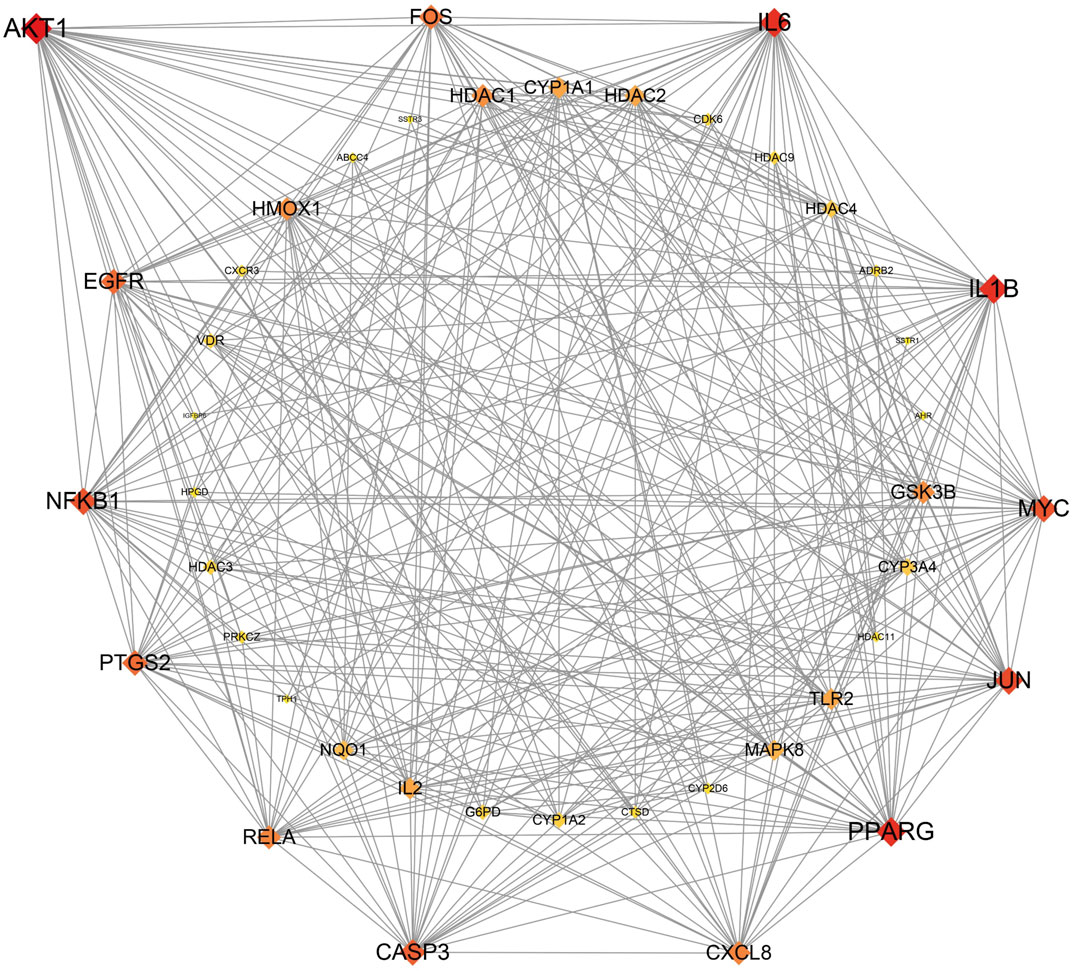
Figure 2. Protein–protein interaction network in this study.
3.3 Signaling pathway and GO enrichment analyses
The 43 core targets were used in the KEGG (Figure 3A) and GO (Figure 3B) enrichment analyses using R programming language. The KEGG results showed that these pathways related to these targets mainly involved inflammation, immune responses, infection, and cancer. The IL-17 signaling pathway is related to inflammation (Amatya et al., 2017), and inflammation has been observed to be closely related to the development of PitNETs (Wang et al., 2021; Chen et al., 2015); and the IL-17 signaling pathway (has04657) has a significant enrichment effect. Studies have shown that IL-17 is a proinflammatory cytokine that promotes the growth and progression of cancer by activating various signaling pathways (Inthanon et al., 2023). In addition, the serum IL-17 levels in patients with invasive PitNETs are significantly higher than those in patients with non-invasive PitNETs (Qiu et al., 2011); the median serum IL-17A level in patients with PitNETs is higher than that in the control group (Glebauskiene et al., 2018). This suggests that the IL-17 signaling pathway may play an important role in the development and invasive behaviors of PitNETs. The GO enrichment analysis results showed that biological processes were related to immune and inflammatory responses, oxidative stress, antioxidant responses, and cell proliferation and differentiation. We further analyzed the interactions between the top-20 significant regulatory pathways (Figure 3C), whose interaction degrees were all equal to 19. Furthermore, 31 metabolites corresponding to the 43 core targets were input into MetaboAnalyst 6.0 for the metabolic pathway analysis (Figure 3D), and pathway impacts exceeding 0.1 were selected as the potential metabolic pathways. In this study, only one potential metabolic pathway of tryptophan metabolism was observed to be enriched. A recent study shown that tryptophan metabolism was closely related to tumor growth (Liu et al., 2023). Based on the above results and published literature, it was found that tryptophan metabolism was related to the protein CXCL8. Tryptophan metabolizing enzymes can accelerate tumor progression by upregulating CXCL8 (Zhao et al., 2021), which is also a chemokine in the IL-17 signaling pathway, and CXCL8 has a high degree value in PPI network; hence, we speculate that CXCL8 could be a potential target in the treatment of PitNETs.
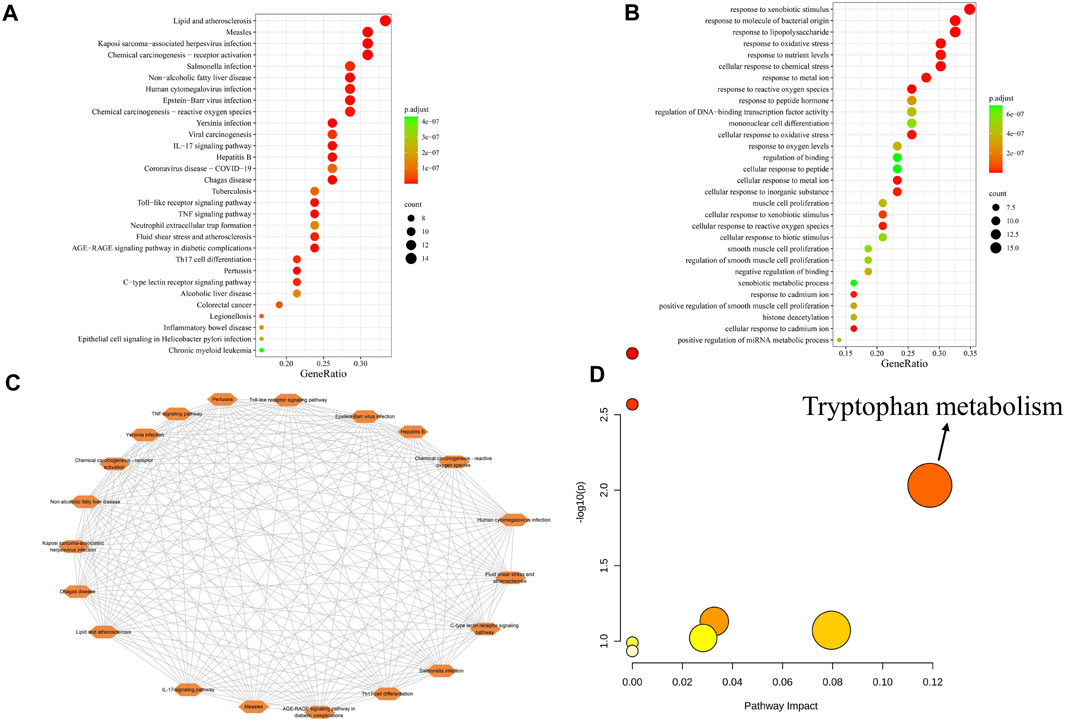
Figure 3. (A) KEGG enrichment analysis of the 43 core targets. (B) GOBP enrichment analysis of the 43 core targets. (C) Pathway interaction diagram. (D) Metabolic pathway analysis of the 31 metabolites identified from the 43 core targets. The colors of the dots represent their p-values from the pathway enrichment analyses; color variation from yellow to red indicates that the p-value is decreasing. The size of the dot represents the pathway impact value from the pathway topology analysis, where the larger the dot, the greater the pathway impact.
3.4 Molecular docking analysis
Molecular docking was used to evaluate the binding energies of CXCL8 (PDB ID: 6WZM) with six potentially related metabolites from the gutMGene database. These metabolites included succinate; acetate; 1-piperazinepentanamide, N-((1S,2R)-2,3-dihydro-2-hydroxy-1H-inden-1-yl)-2-(((1,1-dimethylethyl)amino)carbonyl)-4-(furo(2,3-b)pyridin-5-ylmethyl)-gamma-hydroxy-alpha-(phenylmethyl)-, (alphaR, gammaS,2S)- (also known as compound K); butyrate; baicalein; and baicalin. When the binding energy for docking is less than −5 kcal/mol, it indicates that there is good binding activity between the protein and metabolite. The results showed that the binding energy of CXCL8 with baicalin was −7.0 kcal/mol (Figure 4A), that with baicalein was −6.3 kcal/mol (Figure 4B), and that with compound K was −5.8 kcal/mol (Figure 4C) (Supplementary Table S2). The binding energies of CXCL8 with the other three metabolites exceeded −5 kcal/mol. These results show that baicalin, baicalein, and compound K may be potential metabolites affecting PitNETs.
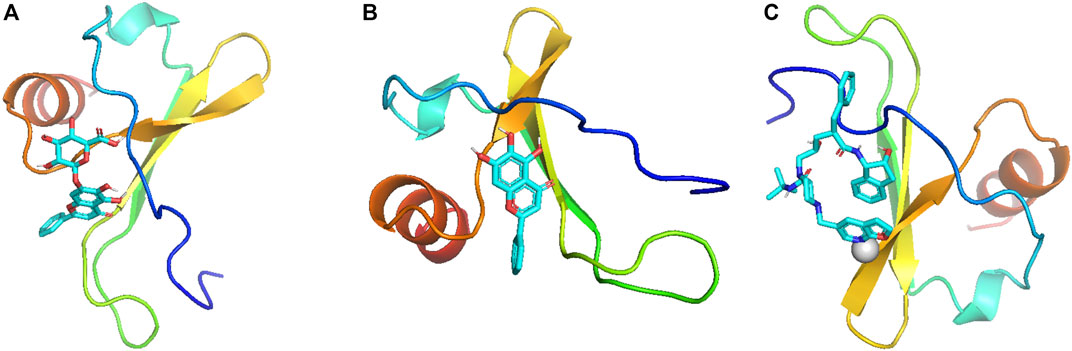
Figure 4. Molecular docking visualization of (A) baicalin–CXCL8, (B) baicalein–CXCL8, and (C) compound-K–CXCL8.
3.5 Analysis of the microbiota-signaling pathways-targets-metabolites network
The microbiota and metabolites directly related to the top-20 significant signaling pathway targets were selected from the gutMGene database. In this study, we only selected the top-10 results based on the degree of microbiota for subsequent analyses. The microbiota–signaling pathways–targets–metabolites network consisted of 89 nodes (10 microbiota, 20 signaling pathways, 29 targets, and 30 metabolites) and 479 edges (Figure 5A). To show the relationships between the signaling pathways and targets more clearly, we indicate the relationships between the signaling pathways and targets in the network separately (Figure 5B). Here, the orange boxes represent the gut microbiota, green boxes represent the signaling pathways, yellow boxes represent the targets, and red boxes represent the metabolites. We found that these top-10 microbiota had the same interaction degrees: Bifidobacterium adolescentis, Escherichia coli K-12, Streptococcus salivarius K12, Streptococcus salivarius JIM8772, Lactobacillus acidophilus ATCC 4357, Faecalibacterium prausnitzii, Streptococcus salivarius, Bacteroides distasonis, Bacteroides vulgatus, and Faecalibacterium prausnitzii A2-165. Studies have found that B. adolescentis displays distinct anti-inflammatory effects (Guo et al., 2019) and can suppress tumorigenesis (Lin et al., 2023; Chen et al., 2023). Previous studies have also shown that inflammation is closely related to the development of PitNETs (Wang et al., 2021; Chen et al., 2015). Therefore, we speculate that B. adolescentis may be an important microbe related to PitNETs. These findings, along with the results of our previous research, show that tryptophan metabolism could be a potential metabolic pathway and could be related to CXCL8. The molecular docking analysis results show that baicalin, baicalein, and compound K have good binding activities with CXCL8 that participates in the IL-17 signaling pathway; furthermore, the IL-17 signaling pathway is related to inflammation, and B. adolescentis plays an important role in inflammatory response. Thus, in the microbiota–signaling pathways–targets–metabolites network, we found that baicalin, baicalein, and compound K were potentially related to CXCL8, the IL-17 signaling pathway, and B. adolescentis. These results show that these six elements could be used as potential markers in the treatment of PitNETs.
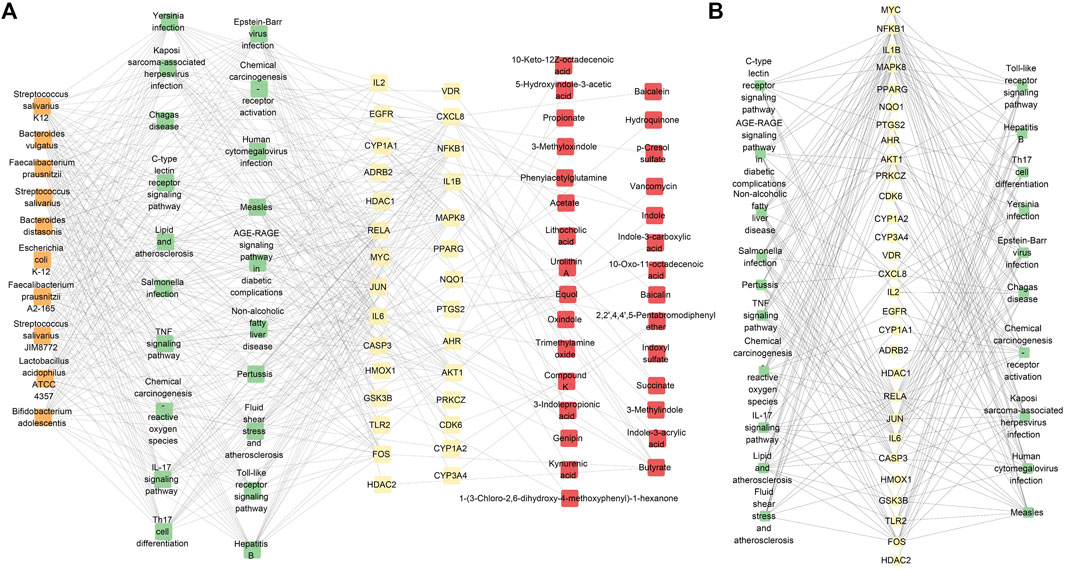
Figure 5. (A) Proposed microbiota-signaling pathways-targets-metabolites network. To show the relationships between the microbiota and pathways more clearly, only the top 10 microbiota are shown based on their interaction degrees. (B) Network showing the signaling pathways and targets.
4 Discussion
PitNETs are commonly divided into functional and non-functional PitNETs. Functional PitNETs can be further divided into various types based on the secretion of hormones, among which prolactinoma is the most common, followed by growth hormone and adrenocorticotropic hormone. We screened the 43 core targets related to PitNETs from the gutMGene v1.0 database and noted that 31 key metabolites corresponding to the 43 core targets were enriched in the tryptophan metabolism pathway. A prior study reported that prolactinoma was related to tryptophan metabolism (Zhou et al., 2015). The change in amino acid metabolism has been considered as a characteristic of tumor cells, which significantly impact the tumor cells and the immune regulation mechanisms in the tumor microenvironment (Xu et al., 2021). Tryptophan is an essential amino acid and a precursor to serotonin; serotonin is a key neurotransmitter in the enteric system and central nervous system. Gut microbiota can potentially affect the brain and behaviors by controlling the availability of the serotonin precursor tryptophan. Gut microbiota also indirectly controlled the hypothalamic–pituitary–adrenal axis (Clarke et al., 2014). This suggests that tryptophan metabolism may play a key role in the development of PitNETs and may be a key metabolic pathway in the regulation of PitNETs.
KEGG enrichment analysis of the 43 core targets showed that IL-17 was a key signaling pathway in the regulation of PitNETs; the IL-17 signaling pathway also plays a key role in inflammation-related diseases (Bunte and Beikler, 2019; Chang and Dong, 2011). IL-17 signaling is known to promote gut barrier immunity via regulation of microbes as well as drive tumor growth (Chandra et al., 2024). Inflammation is closely related to the development of PitNETs (Wang et al., 2021; Chen et al., 2015). Studies have shown that the serum IL-17 levels in patients with invasive PitNETs are significantly higher than those in patients with non-invasive PitNETs (Qiu et al., 2011); furthermore, the median serum IL-17A level in patients with PitNETs is higher than that in the control group (Glebauskiene et al., 2018). CXCL8 participated in the IL-17 signaling pathway (Jain, 2022) and is known to be closely related to tryptophan metabolism. The conversion of tryptophan into kynurenine has been shown to be related to inflammation (Gustafsson et al., 2020), which indicates that CXCL8 may be a potential target for PitNET treatment. Chronic inflammation is an important factor in the occurrence and development of tumors (Coussens and Werb, 2002; Liu et al., 2016). CXCL8 is a PitNET-derived cytokine that plays an important role in the tumor microenvironment (Marques et al., 2019) to regulate tumor proliferation, invasion, and migration in an autocrine or a paracrine manner. CXCL8 can be integrated with multiple intracellular signaling pathways to produce synergistic effects (Liu et al., 2016); it is important for the activation and transport of inflammatory mediators as well as progression and metastasis of tumors (Ha et al., 2017).
B. adolescentis may be an important microbe related to PitNETs. A recent study showed that B. adolescentis produces the microbial metabolite hypaphorine, which inhibits inflammatory responses and hepatic oxidative stress (Qin et al., 2024). Furthermore, dietary supplementation with B. adolescentis has been shown to augment tightening of the intestinal barrier, dampened inflammatory (Roberts et al., 2020) and B.adolescentis can suppress tumorigenesis (Lin et al., 2023; Chen et al., 2023). Preincubation of HT-29 cells with B. adolescentis FRP 61 was shown to significantly inhibit CXCL8 secretion (Carey and Kostrzynska, 2013).
Herein, we found that baicalin, baicalein, and compound K have good binding activities with CXCL8. Baicalin is an important flavonoid isolated from Scutellaria baicalensis Georgi and exhibits important anti-inflammatory, anti-infection, and antitumor functions (Zhao et al., 2020). Baicalein is an important medicinal flavonoid derivative of S. baicalensis Georgi that has anti-inflammatory and anticancer properties (Gupta et al., 2022), in addition to being able to regulate the tumor microenvironment (Wang et al., 2024). Compound K is a rare protopanaxadiol type of ginsenoside that inhibits tumor growth by inducing tumor apoptosis and tumor cell differentiation; it also regulates the tumor microenvironment by inhibiting tumor-angiogenesis-related proteins (Liu et al., 2022). And compound K can be used to treat chronic inflammatory diseases (Tian et al., 2023). These findings indicate that baicalin, baicalein, and compound K could be promising effectors in the treatment of PitNETs.
Thus, this study shows that B. adolescentis may be an important microbe related to PitNETs and that tryptophan metabolism may be a key metabolic pathway in PitNETs treatment and it is closely related to CXCL8. In addition, the results suggest that baicalin, baicalein, and compound K may affect the IL-17 signaling pathway in PitNETs through CXCL8. These findings provide important viewpoints for further research.
As a new interdisciplinary research method, network pharmacology shows great potential in revealing the actions mechanisms, targets, and biological effects of metabolites; therefore, it has significance for comprehensively and deeply understanding the interactions between metabolites and organisms. However, there are some limitations and challenges to using this approach, including imperfect information in the databases used, incomplete data, and the need to verify the accuracies of the target predictions. Thus, in the future, we intend to further verify our current findings through experiments.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material; further inquiries can be directed to the corresponding authors.
Author contributions
MC: Conceptualization, Data curation, Investigation, Software, Supervision, Writing–original draft and Writing–review and editing. PH: Data curation, Investigation and Writing–review and editing. L-SX: Data curation, Supervision, Writing–original draft and Writing–review and editing. Y-HZ: Investigation, Software, Supervision, Funding acquisition, Writing–original draft and Writing–review and editing.
Funding
The authors declare that financial support was received for the research, authorship, and/or publication of this article. This project was sponsored by the Science and Technology Innovation Enhancement Project of Army Medical University (STIEP) (Grant no. 2019XLC3046), Natural Science Foundation Project of Chongqing, Chongqing Science and Technology Commission (Grant no. CSTB2023NSCQ-MSX0848), and Chongqing Sports Science Research Project (Grant B2024100).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphar.2024.1403864/full#supplementary-material
References
Amatya, N., Garg, A. V., and Gaffen, S. L. (2017). IL-17 signaling: the Yin and the Yang. Trends Immunol. 38 (5), 310–322. doi:10.1016/j.it.2017.01.006
Berding, K., Vlckova, K., Marx, W., Schellekens, H., Stanton, C., Clarke, G., et al. (2021). Diet and the microbiota-gut-brain axis: sowing the seeds of good mental health. Adv. Nutr. 12 (4), 1239–1285. doi:10.1093/advances/nmaa181
Bunte, K., and Beikler, T. (2019). Th17 cells and the IL-23/IL-17 axis in the pathogenesis of periodontitis and immune-mediated inflammatory diseases. Int. J. Mol. Sci. 20 (14), 3394. doi:10.3390/ijms20143394
Carey, C. M., and Kostrzynska, M. (2013). Lactic acid bacteria and bifidobacteria attenuate the proinflammatory response in intestinal epithelial cells induced by Salmonella enterica serovar Typhimurium. Can. J. Microbiol. 59 (1), 9–17. doi:10.1139/cjm-2012-0446
Chandra, V., Li, L., Le Roux, O., Zhang, Y., Howell, R. M., Rupani, D. N., et al. (2024). Gut epithelial interleukin-17 receptor a signaling can modulate distant tumors growth through microbial regulation. Cancer Cell 42 (1), 85–100.e6. doi:10.1016/j.ccell.2023.12.006
Chang, S. H., and Dong, C. (2011). Signaling of interleukin-17 family cytokines in immunity and inflammation. Cell Signal 23 (7), 1069–1075. doi:10.1016/j.cellsig.2010.11.022
Chen, S., Fan, L., Lin, Y., Qi, Y., Xu, C., Ge, Q., et al. (2023). Bifidobacterium adolescentis orchestrates CD143(+) cancer-associated fibroblasts to suppress colorectal tumorigenesis by Wnt signaling-regulated GAS1. Cancer Commun. (Lond.) 43 (9), 1027–1047. doi:10.1002/cac2.12469
Chen, Z., Li, Z., Chang, Y., Ma, L., Xu, W., Li, M., et al. (2015). Relationship between NF-κB, MMP-9, and MICA expression in pituitary adenomas reveals a new mechanism of pituitary adenomas immune escape. Neurosci. Lett. 597, 77–83. doi:10.1016/j.neulet.2015.04.025
Cheng, L., Qi, C., Yang, H., Lu, M., Cai, Y., Fu, T., et al. (2022). gutMGene: a comprehensive database for target genes of gut microbes and microbial metabolites. Nucl. Acids Res. 50 (D1), D795–d800. doi:10.1093/nar/gkab786
Clarke, G., Stilling, R. M., Kennedy, P. J., Stanton, C., Cryan, J. F., and Dinan, T. G. (2014). Minireview: gut microbiota: the neglected endocrine organ. Mol. Endocrinol. 28 (8), 1221–1238. doi:10.1210/me.2014-1108
Coussens, L. M., and Werb, Z. (2002). Inflammation and cancer. Nature 420 (6917), 860–867. doi:10.1038/nature01322
Draghici, S., Khatri, P., Tarca, A. L., Amin, K., Done, A., Voichita, C., et al. (2007). A systems biology approach for pathway level analysis. Genome Res. 17 (10), 1537–1545. doi:10.1101/gr.6202607
Dworakowska, D., Wlodek, E., Leontiou, C. A., Igreja, S., Cakir, M., Teng, M., et al. (2009). Activation of RAF/MEK/ERK and PI3K/AKT/mTOR pathways in pituitary adenomas and their effects on downstream effectors. Endocr. Relat. Cancer 16 (4), 1329–1338. doi:10.1677/ERC-09-0101
Glebauskiene, B., Liutkeviciene, R., Vilkeviciute, A., Gudinaviciene, I., Rocyte, A., Simonaviciute, D., et al. (2018). Association of Ki-67 labelling index and IL-17A with pituitary adenoma. Biomed. Res. Int. 2018, 7490585. doi:10.1155/2018/7490585
Guo, Y., Xie, J. P., Deng, K., Li, X., Yuan, Y., Xuan, Q., et al. (2019). Prophylactic effects of Bifidobacterium adolescentis on anxiety and depression-like phenotypes after chronic stress: a role of the gut microbiota-inflammation axis. Front. Behav. Neurosci. 13, 126. doi:10.3389/fnbeh.2019.00126
Gupta, S., Buttar, H. S., Kaur, G., and Tuli, H. S. (2022). Baicalein: promising therapeutic applications with special reference to published patents. Pharm. Pat. Anal. 11 (1), 23–32. doi:10.4155/ppa-2021-0027
Gustafsson, A., Prgomet, Z., Jankovskaja, S., Ruzgas, T., Engblom, J., Ohlsson, L., et al. (2020). Effect of IFN-γ on the kynurenine/tryptophan ratio in monolayer-cultured keratinocytes and a 3D reconstructed human epidermis model. J. Dermatol. Sci. 99 (3), 177–184. doi:10.1016/j.jdermsci.2020.07.005
Ha, H., Debnath, B., and Neamati, N. (2017). Role of the CXCL8-CXCR1/2 axis in cancer and inflammatory diseases. Theranostics 7 (6), 1543–1588. doi:10.7150/thno.15625
Hu, J., Yang, J., Chen, L., Meng, X., Zhang, X., Li, W., et al. (2022). Alterations of the gut microbiome in patients with pituitary adenoma. Pathol. Oncol. Res. 28, 1610402. doi:10.3389/pore.2022.1610402
Huang, N., Zhao, G., Yang, Q., Tan, J., Tan, Y., Zhang, J., et al. (2020). Intracellular and extracellular S100A9 trigger epithelial-mesenchymal transition and promote the invasive phenotype of pituitary adenoma through activation of AKT1. Aging (Albany NY) 12 (22), 23114–23128. doi:10.18632/aging.104072
Inthanon, S., Dejkriengkraikul, P., and Yodkeeree, S. (2023). Notopterol suppresses IL-17-induced proliferation and invasion of A549 lung adenocarcinoma cells via modulation of STAT3, NF-κB, and AP-1 activation. Int. J. Mol. Sci. 24 (20), 15057. doi:10.3390/ijms242015057
Jain, S. (2022). Role of interleukin-17 signaling pathway in the interaction between multiple sclerosis and acute myocardial infarction. Mult. Scler. Relat. Disord. 58, 103515. doi:10.1016/j.msard.2022.103515
Keiser, M. J., Roth, B. L., Armbruster, B. N., Ernsberger, P., Irwin, J. J., and Shoichet, B. K. (2007). Relating protein pharmacology by ligand chemistry. Nat. Biotechnol. 25 (2), 197–206. doi:10.1038/nbt1284
Leistner, C., and Menke, A. (2020). Hypothalamic-pituitary-adrenal axis and stress. Handb. Clin. Neurol. 175, 55–64. doi:10.1016/B978-0-444-64123-6.00004-7
Lin, B., Wang, M., Gao, R., Ye, Z., Yu, Y., He, W., et al. (2022). Characteristics of gut microbiota in patients with GH-secreting pituitary adenoma. Microbiol. Spectr. 10 (1), e0042521. doi:10.1128/spectrum.00425-21
Lin, Y., Fan, L., Qi, Y., Xu, C., Jia, D., Jiang, Y., et al. (2023). Bifidobacterium adolescentis induces Decorin(+) macrophages via TLR2 to suppress colorectal carcinogenesis. J. Exp. Clin. Cancer Res. 42 (1), 172. doi:10.1186/s13046-023-02746-6
Liu, D., Liang, C. H., Huang, B., Zhuang, X., Cui, W., Yang, L., et al. (2023). Tryptophan metabolism acts as a new anti-ferroptotic pathway to mediate tumor growth. Adv. Sci. (Weinh) 10 (6), e2204006. doi:10.1002/advs.202204006
Liu, J., Wang, Y., Yu, Z., Lv, G., Huang, X., Lin, H., et al. (2022). Functional mechanism of ginsenoside compound K on tumor growth and metastasis. Integr. Cancer Ther. 21, 15347354221101203. doi:10.1177/15347354221101203
Liu, Q., Li, A., Tian, Y., Wu, J. D., Liu, Y., Li, T., et al. (2016). The CXCL8-CXCR1/2 pathways in cancer. Cytokine Growth Factor Rev. 31, 61–71. doi:10.1016/j.cytogfr.2016.08.002
Longo, S., Rizza, S., and Federici, M. (2023). Microbiota-gut-brain axis: relationships among the vagus nerve, gut microbiota, obesity, and diabetes. Acta Diabetol. 60 (8), 1007–1017. doi:10.1007/s00592-023-02088-x
Luo, W., Pant, G., Bhavnasi, Y. K., and Blanchard, S. G. (2017). Pathview Web: user friendly pathway visualization and data integration. Nucl. Acids Res. 45 (W1), W501–W508. doi:10.1093/nar/gkx372
Marques, P., Barry, S., Carlsen, E., Collier, D., Ronaldson, A., Awad, S., et al. (2019). Chemokines modulate the tumour microenvironment in pituitary neuroendocrine tumours. Acta Neuropathol. Commun. 7 (1), 172. doi:10.1186/s40478-019-0830-3
Miletta, M. C., Petkovic, V., Eblé, A., Ammann, R. A., Flück, C. E., and Mullis, P. E. (2014). Butyrate increases intracellular calcium levels and enhances growth hormone release from rat anterior pituitary cells via the G-protein-coupled receptors GPR41 and 43. PLoS One 9 (10), e107388. doi:10.1371/journal.pone.0107388
Nie, D., Fang, Q., Cheng, J., Li, B., Li, M., Wang, H., et al. (2022). The intestinal flora of patients with GHPA affects the growth and the expression of PD-L1 of tumor. Cancer Immunol. Immunother. 71 (5), 1233–1245. doi:10.1007/s00262-021-03080-6
Nie, D., Li, C., and Zhang, Y. (2023). PitNETs and the gut microbiota: potential connections, future directions. Front. Endocrinol. (Lausanne) 14, 1255911. doi:10.3389/fendo.2023.1255911
Oh, K. K., Choi, I., Gupta, H., Raja, G., Sharma, S. P., Won, S. M., et al. (2023). New insight into gut microbiota-derived metabolites to enhance liver regeneration via network pharmacology study. Artif. Cells Nanomed. Biotechnol. 51 (1), 1–12. doi:10.1080/21691401.2022.2155661
Qin, P., Li, Y., Su, Y., Wang, Z., Wu, R., Liang, X., et al. (2024). Bifidobacterium adolescentis-derived hypaphorine alleviates acetaminophen hepatotoxicity by promoting hepatic Cry1 expression. J. Transl. Med. 22 (1), 525. doi:10.1186/s12967-024-05312-6
Qiu, L., He, D., Fan, X., Li, Z., Liao, C., Zhu, Y., et al. (2011). The expression of interleukin (IL)-17 and IL-17 receptor and MMP-9 in human pituitary adenomas. Pituitary 14 (3), 266–275. doi:10.1007/s11102-011-0292-5
Roberts, J. L., Liu, G., Darby, T. M., Fernandes, L. M., Diaz-Hernandez, M. E., Jones, R. M., et al. (2020). Bifidobacterium adolescentis supplementation attenuates fracture-induced systemic sequelae. Biomed. Pharmacother. 132, 110831. doi:10.1016/j.biopha.2020.110831
Safran, M., Dalah, I., Alexander, J., Rosen, N., Iny Stein, T., Shmoish, M., et al. (2010). GeneCards version 3: the human gene integrator. Database (Oxford) 2010, baq020. doi:10.1093/database/baq020
Shannon, P., Markiel, A., Ozier, O., Baliga, N. S., Wang, J. T., Ramage, D., et al. (2003). Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 13 (11), 2498–2504. doi:10.1101/gr.1239303
Tafet, G. E., and Nemeroff, C. B. (2020). Pharmacological treatment of anxiety disorders: the role of the HPA Axis. Front. Psychiatry 11, 443. doi:10.3389/fpsyt.2020.00443
Tian, Y., Feng, X., Zhou, Z., Qin, S., Chen, S., Zhao, J., et al. (2023). Ginsenoside compound K ameliorates osteoarthritis by inhibiting the chondrocyte endoplasmic reticulum stress-mediated IRE1α-TXNIP-NLRP3 axis and pyroptosis. J. Agr. Food Chem. 71 (3), 1499–1509. doi:10.1021/acs.jafc.2c06134
Wang, L., Wei, C., Wang, Y., Huang, N., Zhang, T., Dai, Y., et al. (2023a). Identification of the enhancer RNAs related to tumorgenesis of pituitary neuroendocrine tumors. Front. Endocrinol. (Lausanne) 14, 1149997. doi:10.3389/fendo.2023.1149997
Wang, Q., Yang, Q., and Liu, X. (2023b). The microbiota-gut-brain axis and neurodevelopmental disorders. Protein Cell 14 (10), 762–775. doi:10.1093/procel/pwad026
Wang, R., Wang, C., Lu, L., Yuan, F., and He, F. (2024). Baicalin and baicalein in modulating tumor microenvironment for cancer treatment: a comprehensive review with future perspectives. Pharmacol. Res. 199, 107032. doi:10.1016/j.phrs.2023.107032
Wang, X., Ma, L., Ding, Q. Y., Zhang, W. Y., Chen, Y. G., Wu, J. H., et al. (2021). Microglial NLRP3 inflammasome activation-mediated inflammation promotes prolactinoma development. Endocr. Relat. Cancer 28 (7), 433–448. doi:10.1530/ERC-21-0137
Xu, Y., Zhang, H., Sun, Q., Geng, R., Yuan, F., Liu, B., et al. (2021). Immunomodulatory effects of tryptophan metabolism in the glioma tumor microenvironment. Front. Immunol. 12, 730289. doi:10.3389/fimmu.2021.730289
Yu, G., Wang, L. G., Han, Y., and He, Q. Y. (2012). clusterProfiler: an R package for comparing biological themes among gene clusters. Omics J. Integr. Biol. 16 (5), 284–287. doi:10.1089/omi.2011.0118
Zhao, H., Li, C., Li, L., Liu, J., Gao, Y., Mu, K., et al. (2020). Baicalin alleviates bleomycin-induced pulmonary fibrosis and fibroblast proliferation in rats via the PI3K/AKT signaling pathway. Mol. Med. Rep. 21 (6), 2321–2334. doi:10.3892/mmr.2020.11046
Zhao, Y., Sun, J., Li, Y., Zhou, X., Zhai, W., Wu, Y., et al. (2021). Tryptophan 2,3-dioxygenase 2 controls M2 macrophages polarization to promote esophageal squamous cell carcinoma progression via AKT/GSK3β/IL-8 signaling pathway. Acta Pharm. Sin. B 11 (9), 2835–2849. doi:10.1016/j.apsb.2021.03.009
Keywords: pituitary neuroendocrine tumors, gut microbiota, metabolites, tryptophan metabolism, CXCL8
Citation: Cao M, Huang P, Xu L-s and Zhang Y-h (2024) Analysis of gut microbiota-derived metabolites regulating pituitary neuroendocrine tumors through network pharmacology. Front. Pharmacol. 15:1403864. doi: 10.3389/fphar.2024.1403864
Received: 11 May 2024; Accepted: 15 August 2024;
Published: 04 September 2024.
Edited by:
Antonella Smeriglio, University of Messina, ItalyReviewed by:
Krystyna Pawlak, Medical University of Bialystok, PolandEnchen Zhou, University of California, San Diego, United States
Copyright © 2024 Cao, Huang, Xu and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yi-hua Zhang, emhhbmd5aWh1YTY2NkB0bW11LmVkdS5jbg==; Lun-shan Xu, eHVsdW5zaGFuQHRtbXUuZWR1LmNu