
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Pharmacol. , 04 April 2024
Sec. Experimental Pharmacology and Drug Discovery
Volume 15 - 2024 | https://doi.org/10.3389/fphar.2024.1388747
 Huaxue Huang1,2,3,4†
Huaxue Huang1,2,3,4† Zhi Peng3,4†Shuang Zhan3,4Wei Li3,4Dai Liu3,4Sirui Huang3,4
Zhi Peng3,4†Shuang Zhan3,4Wei Li3,4Dai Liu3,4Sirui Huang3,4 Yizhun Zhu1*‡
Yizhun Zhu1*‡ Wei Wang1,2*‡
Wei Wang1,2*‡Siraitia grosvenorii (Swingle) C. Jeffrey (S. grosvenorii), a perennial indigenous liana from the Cucurbitaceae family, has historically played a significant role in southern China’s traditional remedies for various ailments. Its dual classification by the Chinese Ministry of Health for both medicinal and food utility underscores its has the potential of versatile applications. Recent research has shed light on the chemical composition, pharmacological effects, and toxicity of S. grosvenorii. Its active ingredients include triterpenoids, flavonoids, amino acids, volatile oils, polysaccharides, minerals, vitamins, and other microconstituents. Apart from being a natural sweetener, S. grosvenorii has been found to have numerous pharmacological effects, including alleviating cough and phlegm, preventing dental caries, exerting anti-inflammatory and anti-allergic effects, anti-aging and anti-oxidative, hypoglycemic, lipid-lowering, anti-depression, anti-fatigue, anti-schizophrenic, anti-Parkinson, anti-fibrotic, and anti-tumor activities. Despite its versatile potential, there is still a lack of systematic research on S. grosvenorii to date. This paper aims to address this gap by providing an overview of the main active components, pharmacological efficacy, toxicity, current status of development and application, development dilemmas, and strategies for intensive exploitation and utilization of S. grosvenorii. This paper aims to serve as a guide for researchers and practitioners committed to exploiting the biological resources of S. grosvenorii and further exploring its interdisciplinary potential.
Siraitia grosvenorii (Swingle) C. Jeffrey (S. grosvenorii), stands as an indigenous perennial liana hailing from China within the botanical cohort Cucurbitaceae. Its fruits are commonly known as Luo Han Guo (LHG) or monk fruits (Figure 1). It has an extremely sweet fruit taste and has enjoyed extensive prominence across the southern China since ancient times as a local therapeutic medicine for ailments encompassing the common cold, pharyngitis, and minor gastrointestinal afflictions (Kinghora et al., 1986; Zhang and Li, 2011). Therefore, it was listed by the Chinese Ministry of Health in 1987 in the dual-use category for medicinal and food utility (Li et al., 2014).

Figure 1. (A) The flowers, stems, and leaves of S. grosvenorii. (B) Fresh fruit of S. grosvenorii. (C) Dry fruit of S. grosvenorii.
The chemical composition of S. grosvenorii primarily consists of triterpenoid saponins, flavonoids, amino acids, volatile oils, polysaccharides, minerals, vitamins, and other microconstituents. The main active ingredient in S. grosvenorii is mogroside (Mog), which accounts for approximately 3.8% of its content. Mogs are triterpene glucosides, and belong to the cucurbitacinane-type compound category (Jiang et al., 2020; Wei et al., 2023), including Mog III, Mog IV, Mog V, Mog VI, etc., of which Mog V is the main sweet component. Research has shown that extracts obtained from S. grosvenorii, as well as its various parts including fruit, root, seed, flower, and leaf, possess beneficial properties. These include the ability to alleviate cough and eliminating phlegm (Liu et al., 2007), mitigate inflammation (Liu et al., 2021b; Liu et al., 2022), regulate blood glucose and lipid levels (Qi et al., 2008), exhibit antioxidant (Zhu et al., 2020) and anti-tumor (Haung et al., 2023) attributes, prevent tooth decay (Zhou et al., 2022a), inhibit fibrosis (Liu et al., 2021a), anti-aging (Nie et al., 2019), among other benefits. Mechanistically, S. grosvenorii could regulate physiological activities such as oxidative stress, fatty acid oxidation, and apoptosis, and participate in the alleviation of a variety of diseases, including but not limited to lung, liver, kidney, and brain diseases.
S. grosvenorii is a highly valuable economic product that harbors various active ingredients with numerous functions, making it a promising raw material for diverse applications. Currently, the primary application of S. grosvenorii is as a sweetener. Although the application of S. grosvenorii is promising, there are several issues that need to be addressed, including insufficient product development, low extraction purity, and limited exploration of pharmacological mechanisms. This overview provides a systematic examination of the bioactive constituents, pharmacological effects, toxicological effects, resource development status, practical applications, and strategies for intensive exploitation and utilization of S. grosvenorii. It aims to deepen public understanding of this valuable traditional Chinese medicine and provide new perspectives for further investigation into its functional activities and in-depth development of its applications. This comprehensive review seeks to offer new ideas and reference materials to advance the development and utilization of S. grosvenorii.
No less than 131 triterpenoids, 31 flavonoids, 27 amino acids, 19 volatiles, 6 polysaccharides, 19 minerals, and 4 vitamins have been identified in S. grosvenorii (Pandey and Chauhan, 2019, p.; Wu et al., 2022a). Among them, cucurbitanes are the most dominant structural types in S. grosvenorii, belonging to the tetracyclic triterpenoids.
The triterpenoid compounds found in S. grosvenorii are primarily derivatives of cucurbitane-type tetracyclic triterpenoids, and a variety of Mogs have been isolated and characterized. The total content of Mogs in the fruit of S. grosvenorii are approximately 3.8% (Jiang et al., 2020; Wei et al., 2023) (Table 1) (Figures 2–4). Among these Mog compounds, Mog V had the highest content, accounting for about 0.8%–1.3% (w/w) of the fruit’s composition (Li et al., 2014). It possesses an incredibly sweet taste, with a sweetness potency surpassing that of sucrose by a factor of 425 when present at a concentration of 1/10000 (Li et al., 2014). All Mog compounds contain mogrol, [10-cucurbit-5-ene-3,11,24R,25-tetraol], attached to different numbers of glucose units. Categorization of mogrol, according to the conformational changes in the presence or absence of carbonyl group at C-7 position, hydroxyl group at C-11 position, and methyl group at C-25 position, categorized into five groups (Wu et al., 2022a).
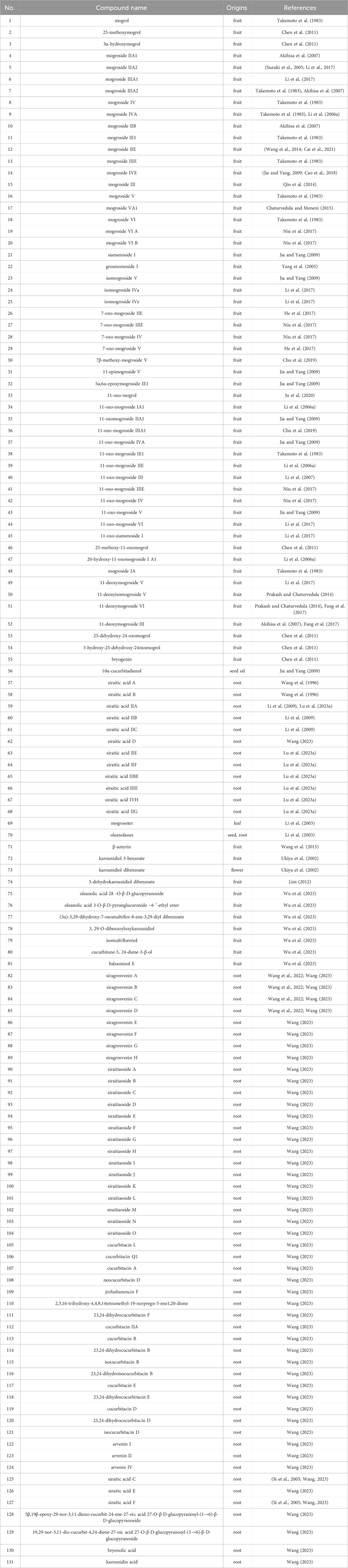
Table 1. The main triterpenoids (3.8%) from S. grosvenorii.
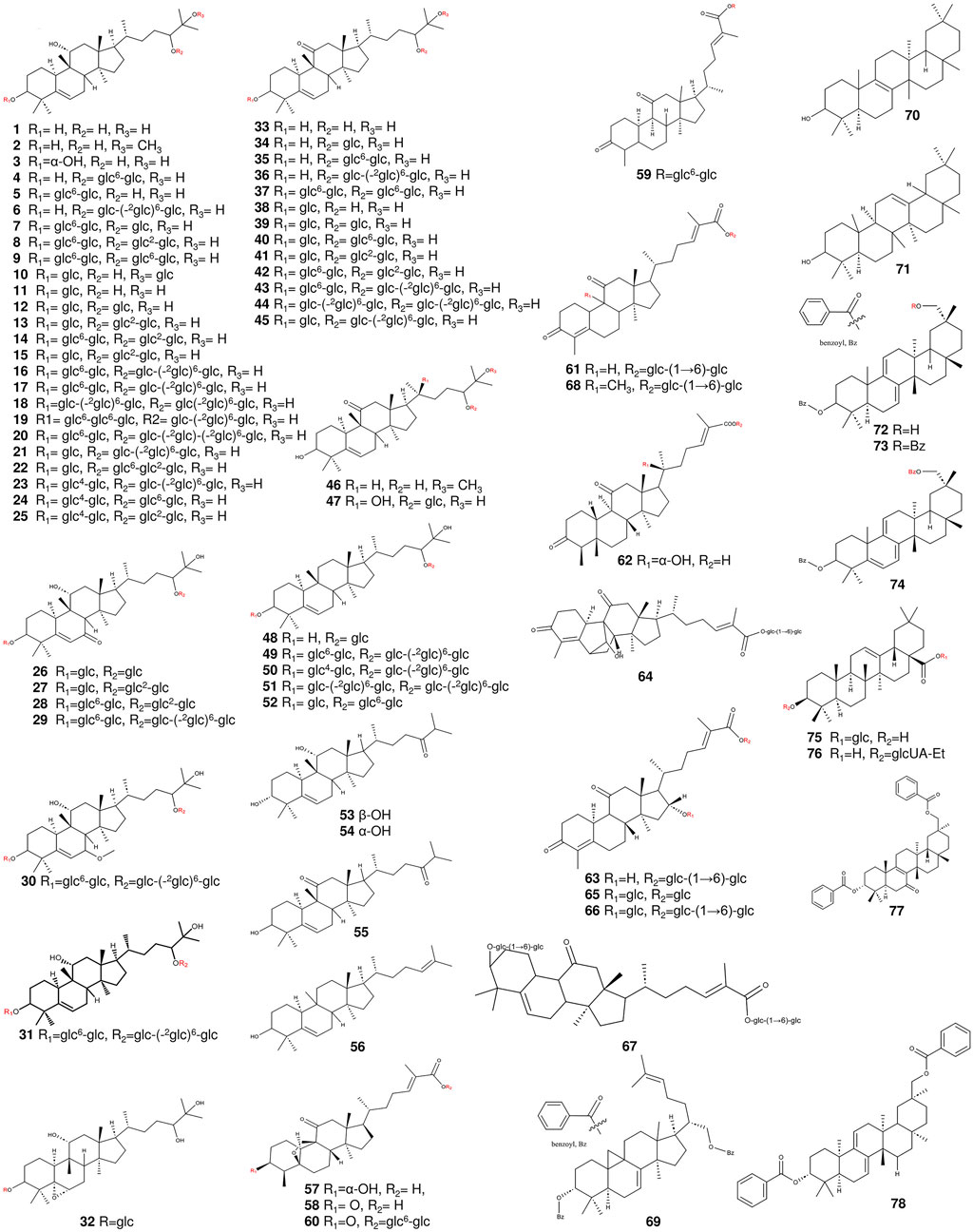
Figure 2. The structures of triterpenoids (compounds 1–78) from S. grosvenorii.

Figure 3. The structures of triterpenoids (compounds 79–107) from S. grosvenorii.
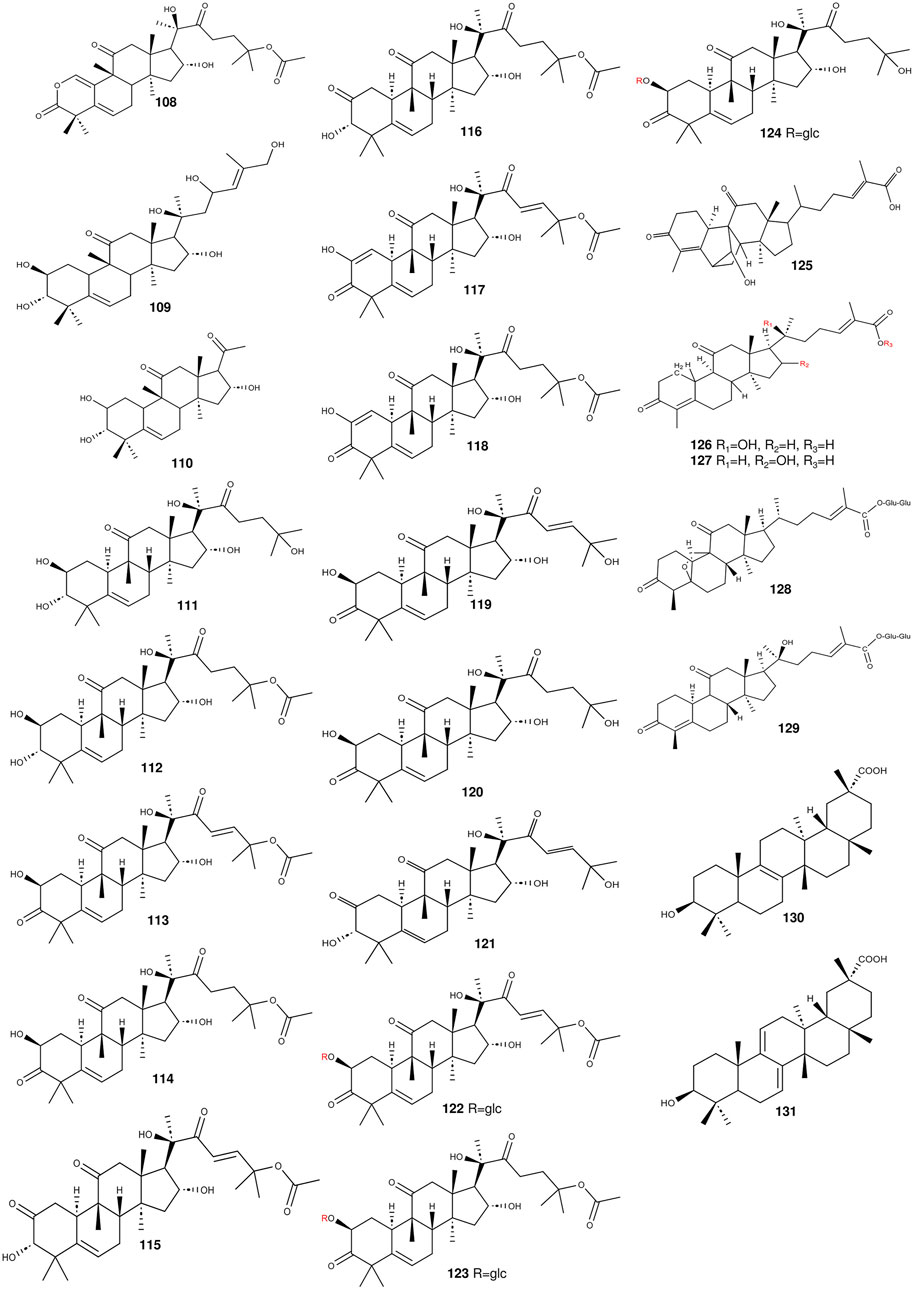
Figure 4. The structures of triterpenoids (compounds 108–131) from S. grosvenorii.
There are studies show that the sweetness of 3β-hydroxy-cucurbit-5-ene derivative glycosides depends on the count and arrangement of glucose units, the oxygen function group at position C-11, and the side chain hydroxylation (Kasai et al., 1988). Glycosides possessing at least three glucose units are endowed with sweetness. For instance, Mog IIE, Mog IIIE, and Mog III with fewer than three glucose units, lack sweet taste (Kasai et al., 1988). Whereas as Siamenoside I, containing 5 glucose units, standing as the most intensely sweet triterpenoid glycoside isolated to date (Li et al., 2014). However, the relationship between glucose unit number and sweetness is not necessarily proportional. For example, despite having the same number of glucose units as Siamenoside I, Mog IVA and Mog IVE are not as sweet (Wu et al., 2022a). Intriguingly, recent study discloses that the glycosyltransferase UGT94-289-3 can catalyze the glycosylation of bitter Mog IIE and III during fruit post-ripening to produce sweet Mog containing 4 to 6 glucose units (Cui et al., 2023). Under optimized catalytic conditions, the glycosyltransferase UGT94-289-3 can effectuate the conversion of 95% of Mog into sweet Mog using Mog III as substrate (Cui et al., 2023). Another pivotal determinant of sweetness is the oxygen functional group at the C-11 position of the aglycone moiet. Mog IVA and Mog IVE containing 11α-hydroxyl glycosides tasted sweet, in stark contrast to compounds with 11β-hydroxyl glycosides that were tasteless, and compounds containing 11-oxides and their dehydrogenated derivatives taste bitter (Pawar et al., 2013). Sweetness is further influenced by side chain hydroxylation. For example, the bitter 11-oxo-mogrosides (11-O-Mog) can be sweetened by hydroxylation of the side-chain double bond with osmium tetroxide (Wu et al., 2022a).
It merits attention that the content of Mogs within the fruit undergoes variations corresponding to its developmental stages. During the initial 30 days post-pollination, Mog IIE predominates, gradually yielding to elevated concentrations of Mog III between 30 and 55 days. Subsequent phases from 56 to 70 days witness the emergence of Mog IV, Mog IVA, and Mog IVE, accompanied a change in fruit flavor from bitter to sweet. Mog V was produced at 70 days, peaking in its sweetness around 85 days post-pollination (Shivani et al., 2021).
Flavonoids are a branch of a class of polyphenols with the structure of benzo-γ-pyrrodone (Dixon et al., 1983). Among the compounds scrutinized, grosvenorine emerged as the principal flavonoid glycoside of the fruits of S. grosvenorii (Wu et al., 2022a) (Table 2) (Figure 5). Notably, among grosvenorine metabolites, were kaempferol and quercetin, emerging as the predominant flavonoids within S. grosvenorii (Duan et al., 2023). In S. grosvenorii, flavonol glycoside content varied with time. Commencing at day 40 and culminating at day 50, a rapid augmentation in flavonol glycosides was manifest. It peaks around day 50, declines after day 60, and eventually stabilizes to around day 20 (Li et al., 2014).
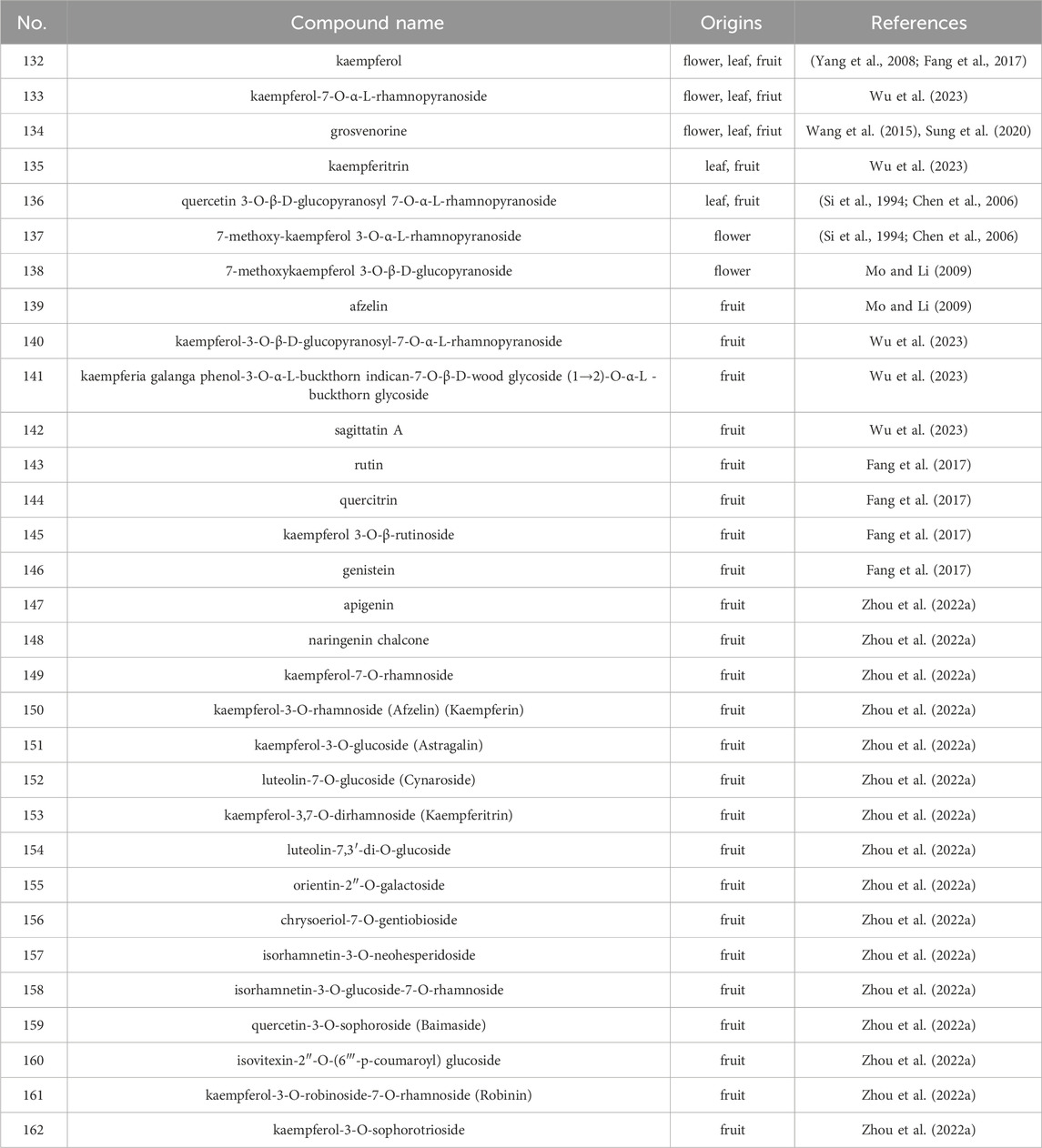
Table 2. The main flavonoids from S. grosvenorii.
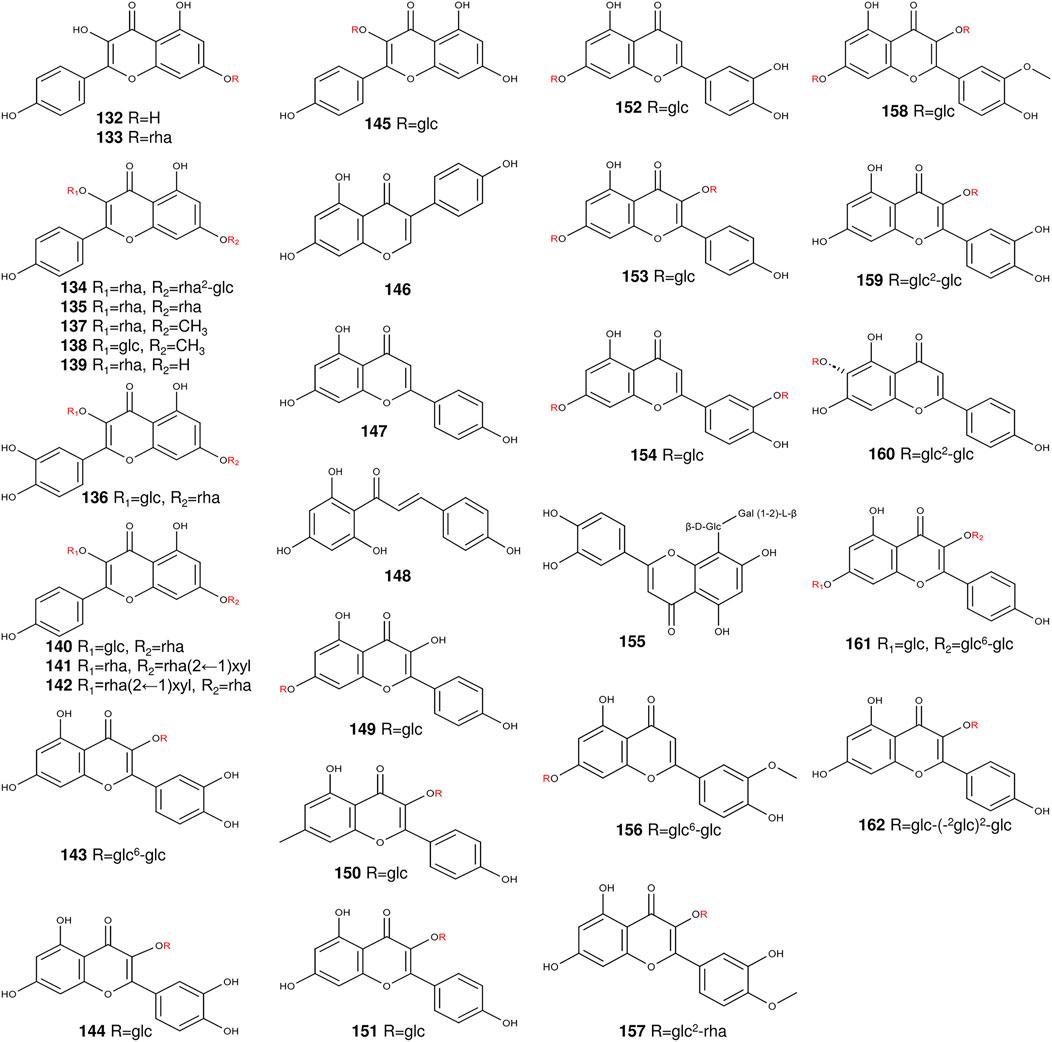
Figure 5. The structures of flavonoids from S. grosvenorii.
Free amino acids, pivotal contributors to protein synthesis, cell signaling, metabolism, physiology, and health. In the pursuit of comprehending amino acid distributions within S. grosvenorii, investigations have delved into the protein content of both fresh and dried fruits (Table 3) (Figure 6), revealing ranges of 8.67%–13.35% and 7.1%–7.8%, respectively. However, the efficiency of amino acid extraction, especially the yield of the ultrasonic methodologies coupled with chromatographic separation, was only 6.54% (Liu et al., 2011). Of the 23 amino acids identified in S. grosvenorii, 19 were significantly increased after Nano-selenium treatment. Including histidine (108.7%), tryptophan (104.5%), serine (87.4%), glycine (85.9%), lysine (67.5%), tyrosine (78.4%), aspartic acid (65.2%), phenylalanine (60.1%), valine (62.0%), glutamic acid (53.4%) and proline (52.5%) (Zhou et al., 2022a). Furthermore, discerning the preponderant amino acids in S. grosvenorii, glutamic acid and aspartic acid emerged as the most abundant constituents, underscoring their pivotal roles in the botanical matrix (Duan et al., 2023).
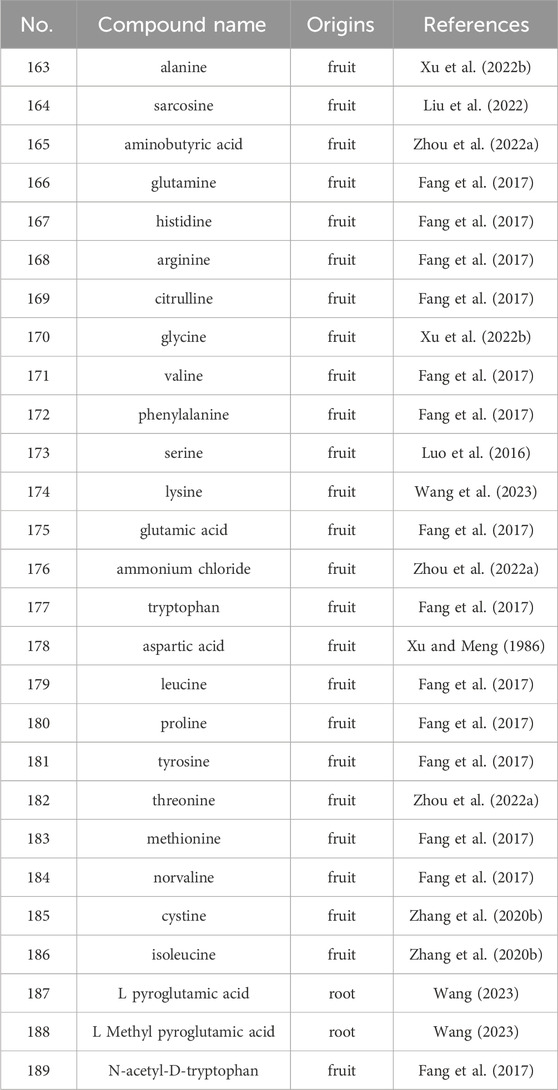
Table 3. The main amino acids from S. grosvenorii.

Figure 6. The structures of amino acids from S. grosvenorii.
The seed oil of S. grosvenorii contains large amounts of fatty aldehydes such as fagni aldehyde, valeraldehyde, hexanal, and nonanal (Li et al., 2014). Predominantly, within the seed kernel of S. grosvenorii, squalene is the principal constituent of the seed oil. Concentrations of essential oils varied considerably between dried and freshly harvested fruits, with the former showing a significant increase in magnitude, ranging from about 0.2% to 0.3%, while the latter was only 0.03% (Li et al., 2014). In the fruits and roots of S. grosvenorii, no less than 17 fatty acids have been found (Table 4) (Figure 7). The highest contents in dried fruit essential oil were palmitic acid (n-Hexadecanoic acid) (45.609%) and linoleic acid (9, 12-octadecadienoic acid) (36.151%), conversely surpassed by butyl 2-butyrate (20.80%) and 2-heptyl alcohol (13.86%) as the prevailing constituents within the essential oil of the fresh fruit (Li et al., 2014). In the existing literature, there are very few descriptions of essential oils in S. grosvenorii.
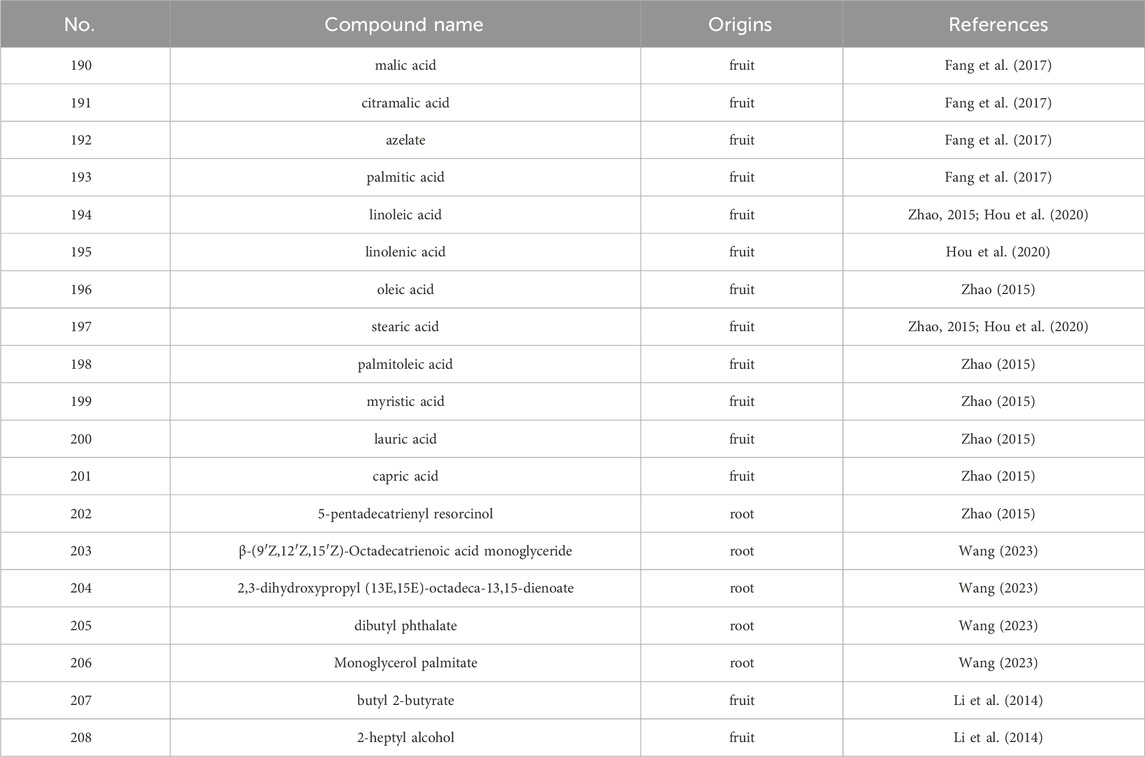
Table 4. The main volatile oils from S. grosvenorii.
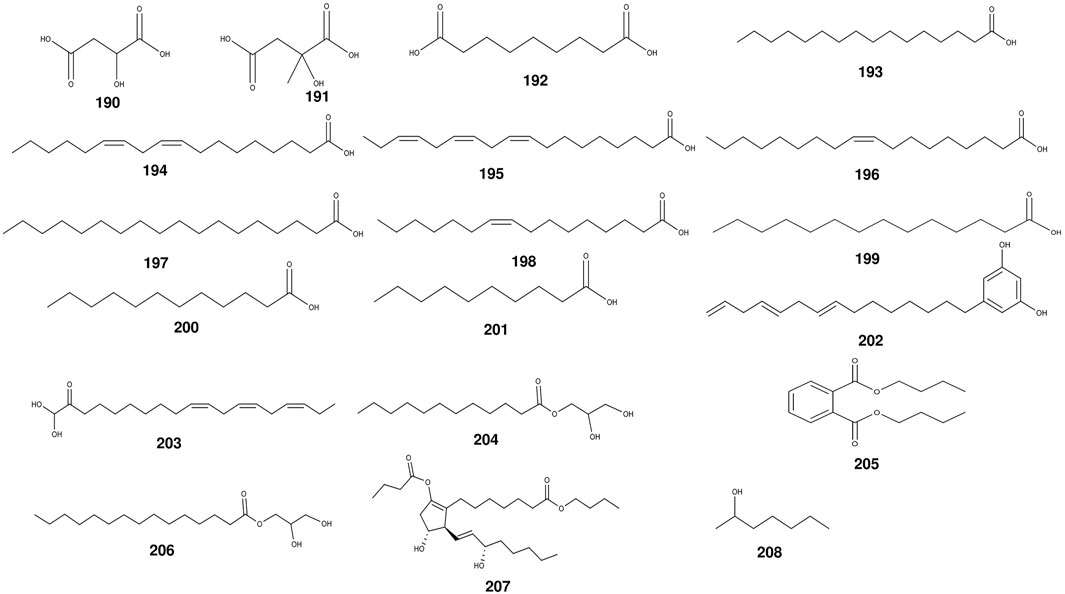
Figure 7. The structures of volatile oils from S. grosvenorii.
Polysaccharides represent polymeric carbohydrate macromolecules composed of long chain monosaccharide units linked by glycosidic bonds. The content of polysaccharide was the highest in the S. grosvenorii flesh (7.55%) (Table 5), while the content of polysaccharide in the seed was the lowest (3.12%). The content of total sugar in S. grosvenorii fruit ranged from 25.17% to 38.31%, encompassing reducible saccharides accounting for values ranging from 16.11% to 32.74% (Li et al., 2014). S. grosvenorii polysaccharide (SGP), possessing a molecular mass of 1.93 × 10³ kDa, characterized by the constituent monomers comprising α-L-arabinose, α-L-arabinose, α-D-mannose, α-D-glucose, α-D-galactose, glucuronic acid, and galacturonic acid, configured in a stoichiometric ratio of 1:1.92:3.98:7.63:1.85:7.348 (Zhu et al., 2020). In addition, a variety of polysaccharides have been isolated and purified from S. grosvenorii residues, including SGPS1 (Chen et al., 2003), SGPS2 (Chen et al., 2003), SGP-1 (Gong et al., 2022), SGP-1-1 (Gong et al., 2022), SGP-2 (Gong et al., 2022), and SGP-3 (Gong et al., 2022). SGP-S1 and SGP-S2 were the first two polysaccharides obtained from S. grosvenorii. SGP-1, SGP-2 and SGP-3 are obtained by first dissolving SGP in distilled water, then gradient elution in distilled water, 0.05, 0.10, 0.15 and 0.20 mol/L Nacl solution, and then further concentrating and dialysis the purified fractions (Gong et al., 2022). Among them, SGP-1 has the strongest antioxidant activity (Gong et al., 2022). Furthermore, SGP-1, identified as a glucomannan derivative originating from the dried fruit of S. grosvenorii, is discerned by a molecular weight measuring 19.037 kDa, composed of residues including 4)-β-D-Glcp-(1→4)-, α-D-Glcp-(1→4)-, and 4)-Manp-(1 distributed in a molar proportion of 4.90:1:2.56 (Gong et al., 2022). Notably, SGP-1-1 composed of arabinose, ribose, galacturonic acid, galactose, mannose, and glucose at the molar ratio of 1.00:1.72: 2.24: 3.64: 3.89: 22.77 (Gong et al., 2021).
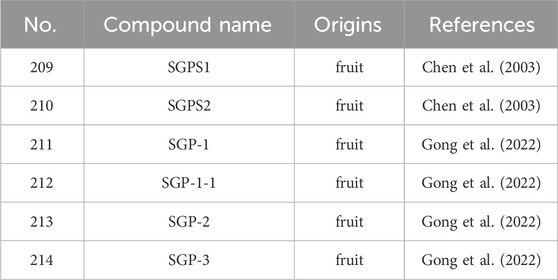
Table 5. The main polysaccharides from S. grosvenorii.
S. grosvenorii is endowed with a spectrum of 19 minerals (Table 6), notably including but not limited to potassium (K), calcium (Ca), phosphorus (P), magnesium (Mg), iron (Fe), zinc (Zn), manganese (Mn), aluminum (Al), copper (Cu), lead (Pb), cadmium (Cd), and selenium (Se). Moreover, the fruit contains high levels of K, Ca, Mg and Se (Duan et al., 2023).
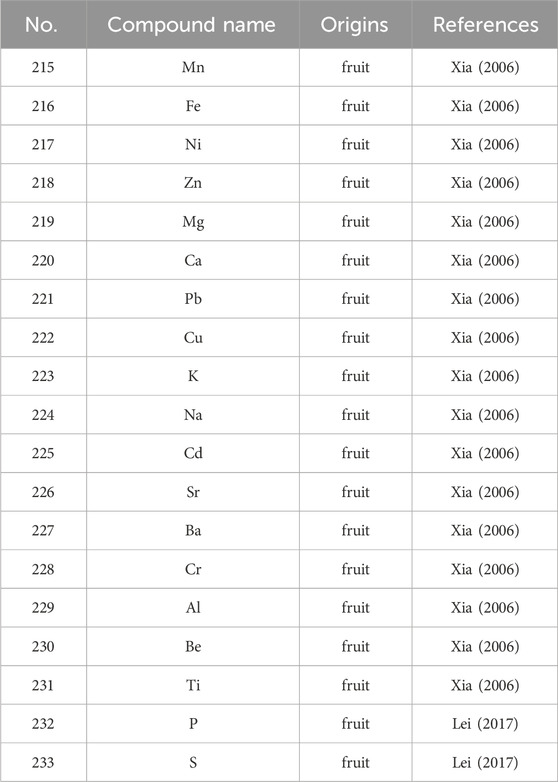
Table 6. The main minerals from S. grosvenorii.
S. grosvenorii, there are also some vitamins such as Thiamine (Vitamin B1), Riboflavin (Vitamin B2), and Ascrobic acid (Vitamin C) (Table 7). Among them, the content of Vitamin B1 in every 100 g of fresh fruit is 338 mg, and Vitamin B2 is 123 mg. The amount of vitamin C detected in S. grosvenorii varies greatly depending on the variety, form, source, growth stage, maturity, differences in drying equipment, and process conditions used by suppliers. Remarkably, this concentration exceed that discerned within botanical counterparts such as citrus, apple, pear, grape, and persimmon. The content of vitamin C in S. grosvenorii fresh fruit (ranging between 339 and 461 mg per 100g) outpaces its dried fruit (ranging between 24.6 and 38.7 mg per 100g) by a factor exceeding tenfold (Ji, 2016). Vitamin C tends to degrade at high temperatures. Therefore, in order to minimize damage to the inherent vitamin C of LHG, it is preferable to preserve LHG under low-temperature vacuum drying conditions (Ji, 2016). In addition, a new strategy to protect LHG, is through the use of microwave heating. The technique involves applying microwave power of 80 W, with a time parameter of 30 min and a microwave interval ratio of 1:1 (Zhou et al., 2018b). This method not only extends the storage life of LHG, but also minimizes the loss of water and vitamin C content.
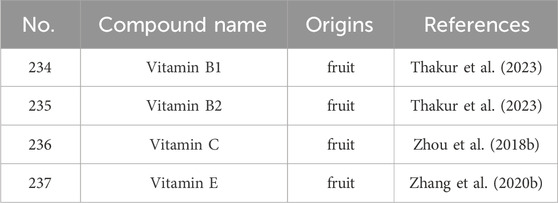
Table 7. The main vitamins from S. grosvenorii.
S. grosvenorii and its root, stem, and leaf also contains wooden fat element, phenolic acids, anthraquinones, alkaloids, sterols, fatty acid, furfural, aromatic, diterpene lactone, hine ring, brain glycosides, and ceramide compounds (Wu et al., 2023).
S. grosvenorii has been extensively documented in various traditional Chinese medicine references such as The Dictionary of Traditional Chinese Medicine, National Compendium of Materia Medica, Lingnan Materia Medica, Color Atlas of Chinese Herbal Medicine, Chinese Herbal Medicine and Chinese Pharmacopoeia. These references highlight the established role of S. grosvenorii in traditional Chinese medicine for removing lung heat, resolving phlegm turbidity, addressing paramesoteric swelling, mitigating cough, alleviating sore throat, addressing hoarseness, chronic laryngitis, chronic bronchitis, and other ailments afflicting the respiratory system (Ji, 2016). Traditionally, traditional Chinese medicine believes that S. grosvenorii is cool in nature, sweet in taste, and meridian distribution in the lung and large intestine (2005).
Given the wide range of documented medicinal and food values of S. grosvenorii, increasing attention has been given to studying its pharmacological effects and the mechanisms of diseases treatment. In this paper, the pharmacological effects of S. grosvenorii are summarized as follows (Table 8).
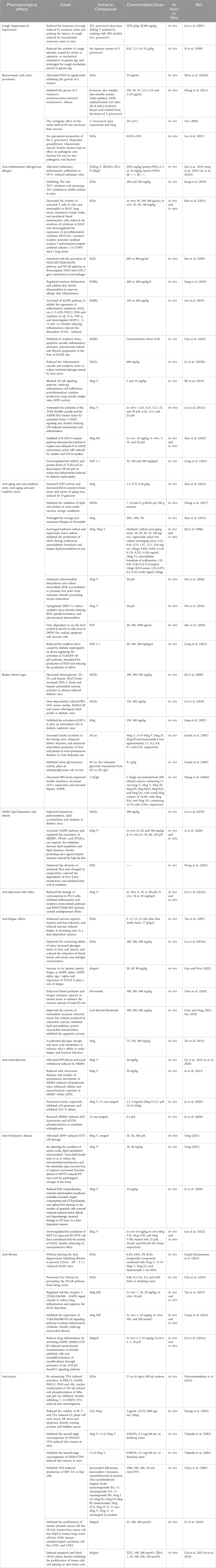
Table 8. The pharmacological effects of S. grosvenorii.
Cough is a sudden, often involuntary discharge of air from the lungs with a characteristic, easily recognizable sound (Farzan, 2008). It has the function of defending against harmful substances in the respiratory tract and maintaining airway patency by clearing excessive airway secretions. Expectoration or sputum refers to the act of coughing and ejecting substances produced in the respiratory tract (Farzan, 2008).
S. grosvenorii has been used in southern China for more than 300 years as a herbal medicine to clear the throat and moisten the throat. There was a screening experiments for the anti-tussive active ingredients of S. grosvenorii have been carried out. Liu et al. conducted an experiment in which they concentrated the crude powder of S. grosvenorii to a concentration of 1.5 g/mL of raw drug. The raw drug was then extracted with water and the resulting filtrate was eluted with water, 50% ethanol, and 95% ethanol on AB-8 macroporous adsorbent resin. This process yielded three parts of S. grosvenorii: a water-soluble part, a 50% ethanol-soluble part, and a 95% ethanol-soluble part (Liu et al., 2007). The 50% ethanol-soluble part was further purified by D-280 anion exchange resin to remove alcoholic flavor, and the resulting effluent was concentrated to obtain the Mog crude product (Liu et al., 2007). After repeated purification on an Octadecylsilyl column, the purity of Mog V was found to be over 94% (Liu et al., 2007). In a mice coughing experiment conducted using S. grosvenorii decoction, it was found that the herb had a certain cough-suppressing effect, but a larger dose (50 g/kg of raw drug) was required to achieve this effect. However, when Mog V (75 mg/kg) was isolated from the effective extracted part (50% alcohol washed) of S. grosvenorii, it showed a significant antitussive effect, with an inhibition rate of 52.93%, indicating a certain quantitative effect relationship (Liu et al., 2007). Another experiment involving phenol red excretion in mice showed that while 50 g/kg of S. grosvenorii raw drug did not have a significant expectorant effect, Mog V at a dose of 150 mg/kg significantly increased phenol red excretion in mice (Liu et al., 2007). Another experiment using guinea pigs, which have a more similar cough reflex to humans, showed that the aqueous extract of S. grosvenorii significantly reduced the number of cough episodes or prolonged cough latency induced by citrate or capsaicin, and inhibited cough induced by mechanical stimulation in guinea pigs (Li et al., 2008). This again indicates the antitussive effect of S. grosvenorii.
The above experimental results indicate that Mog V is the main active component of S. grosvenorii, and the activity of the monomer after isolation and purification is more potent than that of the total aqueous decoction.
Postoperative sore throat (POST), constitutes a prevalent sequel following tracheal intubation and subsequent extubation conducted within the milieu of general anesthesia (El-Boghdadly et al., 2016). The underpinning etiological factor resides within the constrained immunological competence exhibited by the patient throughout the surgical procedure. This physiological limitation manifests as a propitious milieu for the atypical proliferation of oropharyngeal bacterial consortia. Consequent to this microbial overgrowth, deleterious sequelae ensue, comprising injurious ramifications for the oral mucosal integrity, culminating in ulcerative lesions and acute pharyngitis (Scuderi, 2010). It has been showed that adding 10 mg/mL S. grosvenorii extracts (SGEs) to brain heart infusion-sheep blood agar substrate alleviates POST by significantly inhibiting the growth of Streptococcus mutans (S. mutans) (Zhou et al., 2022b). The Glucosyltransferase (GTF) enzyme derived from S. mutans constitutes a pivotal virulence determinant in the etiology of dental caries. β-amyrin, aloe emodin, aloe-emodin acetate, 5α,8α-epidioxy-24(R)-methylcholesta-6,22-dien-3β-ol and p-hydroxyl benzyl acid isolated from the leaves of S. grosvenorii, have demonstrated notable antibacterial efficacy (Zheng et al., 2011). Notably, among them, aloe emodin has strongest antibacterial effect against growth of S. mutans, A. actinomycetemcomitans, F. nucleatum and C. albicans (Zheng et al., 2011). Conversely, the derivative aloe-emodin acetate exhibited diminished inhibitory effects on S. mutans, A. actinomycetemcomitans, and C. albicans when compared to aloe emodin, and exhibited no growth suppression of F. nucleatum (Zheng et al., 2011). p-Hydroxyl benzyl acid demonstrated pronounced inhibition against S. mutans, F. nucleatum and C. albicans, but did not influence A. actinomycetemcomitans (Zheng et al., 2011). 5,8-epidioxy-24(R)methylcholesta-6,22-dien-3-ol had moderate activities against S. mutans, A. actinomycetemcomitans, F. nucleatum and C. albicans (Zheng et al., 2011). β-amyrin exhibited only marginal inhibition against S. mutans and F. nucleatum, whereas aloe emodin demonstrated moderate activities analogous to aloe-emodin acetate against C. albicans. Notably, aloe emodin exhibited significantly more potent antibacterial action against S. mutans, A. actinomycetemcomitans, and F. nucleatum when juxtaposed with aloe-emodin acetate (Zheng et al., 2011).
Caries and periodontitis stand as emblematic oral diseases in humans (Jorn et al., 2005). The main cariogenic bacteria in the oral cavity are S. mutans (Hamada et al., 1984). The pathogenic cascade orchestrated by S. mutans in caries development involves a tripartite sequence. Firstly, the bacterium adheres to the acquired dental membrane in a sucrose independent manner. Subsequently, adhesion to tooth surfaces, contingent upon sucrose, is facilitated through GTF mediation. Lastly, the substrate is metabolized to produce acid (Daboor et al., 2015). Conspicuous investigations have illuminated an intriguing facet of S. grosvenorii, namely its role as a natural sweetener with diminished cariogenic potential. Experimental endeavors conducted in vitro have distinctly portrayed the subdued growth rate and adherence of S. mutans in culture mediums enriched with Mogs, in stark contrast to their sucrose-rich counterparts. The culture mediums abundant in Mogs have been observed to markedly curtail the proliferation and propagation of S. mutans. Further empirical inquiries conducted in vivo have unequivocally indicated a diminished cariogenic impact of concentrated S. grosvenorii juice upon the molars of SD rats when compared with sucrose (Yan, 2009).
While mouthwashes have been harnessed to deter the adhesion of oral microorganisms onto dental biofilms and tooth surfaces, concurrently manifesting antibacterial attributes to thwart the inception of dental complications, synthetic mouthwash formulations encompass an amalgam of compounds preservatives, artificial colors, and taste correction agents. Long-term use of such chemical-laden mouthwashes could lead to gustatory disorders, discoloration of oral hard and soft tissues, allergic reactions, and even oral carcinoma (McCullough and Farah, 2008). Furthermore, the prolonged utilization of chemical mouthwashes might potentially precipitate the eradication of oral microbial communities, thus impinging upon the nitrate-nitrite-NO pathway within salivary secretions, potentially engendering the emergence of cardiovascular maladies and septicemia (Blot, 2021). Remarkably, studies have found that S. grosvenorii alone demonstrated no discernible impact on 11 pathogens in the oral cavity, the natural extract mixture of S. grosvenorii, Platycodon grandiflorum, Chaenomeles sinensis Koehne, enzyme salt, xylitol, mint, green tea, lemon, propolis, silicon dioxide and magnesium stearate exhibited notable anti-inflammatory attributes, evinced by its capacity to inhibit lipopolysaccharide (LPS)-induced NO production and foster the viability of RAW 264.7 cells (Lee et al., 2021). More importantly, natural extracts mixed in proper proportions had an effect on pathogenic oral bacteria but not on non-pathogenic oral bacteria (Lee et al., 2021).
Collectively, these findings indicate that both the fruits and leaves of S. grosvenorii hold promise in terms of their antimicrobial properties. Additionally, various active compounds isolated from the leaves exhibit inhibitory effects on different bacterial species. Notably, Mogs demonstrate preventive efficacy against tooth decay. Combining natural extracts from S. grosvenorii with other natural extracts in appropriate proportions has a good prospect of replacing traditional chemical-based mouthwash formulations, and is worthy of further exploration and consideration.
In recent years, the anti-inflammatory effects of S. grosvenorii have been supported by a large number of experimental data. So far, it has been confirmed that S. grosvenorii has therapeutic effects on the inflammation of the lungs, kidneys, skin, intestines, bones and joints, liver, pancreas and nervous system.
Several studies have provided evidence of the effectiveness of SGEs in reducing lung inflammation. Experiments conducted on both in vivo models of lung inflammation induced by LPS have shown that treatment with SGEs significantly affects the activation of mitogen-activated protein kinase (MAPK)—nuclear factor-κB (NF-κB) in human bronchial epithelial cell line (BEAS-2B) when exposed to LPS. Consequently, this modulation inhibits the production of inflammatory mediators (Kim et al., 2023). Moreover, in a mouse model of chronic obstructive pulmonary disease (COPD) induced by cigarette smoke extract (CSE) and LPS, SGEs have been found to decrease the number of activated T cells, B cells, and neutrophils, as well as reduce inflammatory factor infiltration in various areas like bronchoalveolar lavage fluid (BALF), lung tissue, mesenteric lymph nodes, and peripheral blood mononuclear cells. Furthermore, SGEs downregulate the expression levels of proinflammatory cytokine MUC5AC, transient receptor potential vanilla-like receptor 1, and transient receptor potential fixing protein 1 in lung tissue (Kim et al., 2023). The anti-inflammatory mechanism of SGEs appears to involve inhibiting the activation of NF-κB by interfering with the activation of inosine phosphate 3 kinase (PI3K)/protein kinase B (Akt)/kappa B kinase inhibitor (IKK) and MAPK pathway (Pan et al., 2009). Ultimately, this effect leads to a downregulation of the expression of inflammatory inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) genes in macrophages (Pan et al., 2009).
Studies have validated the effectiveness of S. grosvenorii residue extracts (SGREs) in addressing lung inflammation, skin inflammation, osteoarthritis (OA), and nonalcoholic steatohepatitis (NASH). SGREs have been found to suppress the expression of Th2 and Th17 cytokines while boosting the levels of Th1 cytokines (Kim et al., 2023). In a NC/Nga murine model treated with Dermatophagoides farinae mite antigen extract, SGREs (comprising grosvenorine, kaempferitrin, and Mogs) at doses of 200 and 400 mg/kg exhibited superior improvement in allergic skin inflammation compared to 5 mg/kg dexamethasone. This effect was achieved through the regulation of immune dysfunction and resolution of skin barrier abnormalities (Sung et al., 2020). SGREs exhibit potential as a bioactive treatment for OA. They not only help improve weight distribution on the back foot, thereby reducing discomfort associated with OA, but also have the ability to inhibit various inflammatory mediators such as iNOS, COX-2, 5-lipoxygenase (5-LOX), prostaglandin E2 (PGE2), and leukotriene B4 (LTB4) through the activation of the MAPK/NF-kB pathway. Additionally, they can impact the expression of inflammatory cytokines (including interleukin (IL)-1β, IL-6, tumor necrosis factor (TNF)-α) and cartilage-degrading enzymes (including matrix metalloproteinases (MMP)-1, -2, -9, and -13) (Lee et al., 2023). In a male Hsd: SD rat model of choline-deficient, methionine-deficient, L-amino acid-defined (CDAA)-induced NASH, oral concentrations of SGREs greater than 0.2% regulate the NF-κB pathway by regulating CD44 expression. At the same time, oxidative stress, apoptotic cascade, inflammatory processes, precancerous lesions, and fibrotic progression in the rat liver environment were also inhibited, again demonstrating the inherent anti-inflammation and antioxidant properties of SGREs (Uno et al., 2023).
Mogroside-rich extracts (MGEs) exhibits the potential to mitigate intestinal impairment resulting from thermal stress by ameliorating the cascades of inflammation and oxidative stress, as indicated by previous investigations (Lu et al., 2023b). This finding suggests that MGEs may have the potential to mitigate the consequences of increased thermal stress associated with global climate change.
Mog V has been shown to reduce lung and neuroinflammation. A study established an affinity chromatography strategy to evaluate the anti-asthmatic effects of SGEs using immobilized toxomatine 3 acetylcholine receptor (M3R). The results showed that the flavor components in SGEs that bind to this receptor are Mog V and 11-O-Mog V. The anti-asthmatic effects of these ingredients when used in concert were more significant than SGEs alone (Jia et al., 2019). These findings collectively underscore the promise of Mog V and 11-O-Mog V as candidates for functional food additives. Subsequent studies revealed 93 pathways associated with ovalbumin (OVA)-induced pneumonia through transcriptomic and proteomic analysis. Notably, Mog V effectively inhibited the activation of NF-κB and JAK-STAT pathways, subsequently inhibiting Igha, Ighg1, NF-κB, Jak1 and Stat1 activation to exert discernible anti-inflammatory effects (Song et al., 2019; Dou et al., 2022). A metabolomic analysis was performed using liquid chromatography-mass spectrometry (LC-MS) to elucidate the regulatory role of Mog V in a mouse model of asthma. The results showed that Mog V is involved in regulating six key pathways in asthmatic mice, encompassing vitamin B6 metabolism, taurine and hypotaurine metabolism, ascorbic acid and aldarate metabolism, histidine metabolism, pentose and glucuronic acid interconversion, and the citric acid cycle (TCA cycle). By conducting these experiments, it was observed that the administration of 50 mg/kg Mog V demonstrated similar effects to the positive control, 50 mg/kg Suhuang Zhike Jiaonang, in reducing the presence of biochemical factors and pulmonary inflammatory infiltration in mice with OVA-induced asthma (Liu et al., 2021b). In addition, Mog V was shown to relieve inflammation in a mouse model of LPS-induced acute lung injury. Pretreatment with Mog V at doses of 5 or 10 mg/kg resulted in the attenuation of inflammatory cell invasion, reduction in pro-inflammatory cytokine production, decreased lung wet/dry weight ratio, inhibition of myeloperoxidase activity, and mitigation of lung histopathological changes. These effects were achieved through the blocking of the NF-κB signaling pathway (Shi et al., 2014). In addition, Mog V demonstrated a suppressive effect on the expression of iNOS and COX-2 (Shi et al., 2014). However, Mog V was slightly less effective than dexamethasone (2 mg/kg) (Shi et al., 2014). Mog V also has a lessening effect on neuroinflammation. One evidence is that Mog V exhibits the potential to attenuate the activation of the TLR4-MyD88 cascade and the AMPK/AKT-nuclear factor erythroid2-related factor 2 (Nrf2) signaling axis, thereby mitigating the produce of proinflammatory mediators (namely, TNF-α and IL-1β) within microglial populations, the innate immune effector cells of the central nervous system (CNS). In addition, the secretion of IL-18, IL-6, COX-2, iNOS and high mobility group box 1 (HMGB1) was also inhibited by Mog V, thereby inhibiting LPS neurotoxicity (Liu et al., 2021c).
Mog IIE is one of the main bioactive components in the immature fruit of S. grosvenorii. Studies have shown that it inhibits the IL-9/IL-9 receptor pathway, Thus, the activities of trypsin and cathepsin B in pancreatic acinar cell line AR42J and primary acinar cells induced by nucleoprotein and LPS were attenuated (Xiao et al., 2020). Moreover, this modulation occurs in a dose- and time-dependent manner (Xiao et al., 2020). Therefore, Mog IIE is considered as a potential candidate for ameliorating pancreatitis.
S. grosvenorii polysaccharide SGP-1-1 can reduce the expression of IL-6 and TNF-α cytokines to treat the inflammation associated with diabetic nephropathy. This effect was mediated by down-regulating the mRNA and protein levels of Toll-like receptor 4 (TLR4) and its downstream protein kinase NF-κB p65 (Gong et al., 2021).
In summary, S. grosvenorii has a wide range of anti-inflammatory activities, and its extracts (SGEs), residue extracts (SGREs), MGEs, Mog V, 11-O-Mog V, Mog IIE, and polysaccharides have all demonstrated anti-inflammatory effects. Mog V, the most abundant compound in S. grosvenorii, has been especially effective in treating both respiratory and central system inflammation. The active compounds in S. grosvenorii have been shown to inhibit inflammation by interfering with the activation of the following pathways, including but not limited to the following: ① PI3K/Akt/IKK/NF-κB; ② MAPK/NF-κB pathway; ③ TLR4-MyD88 cascade; ④ AMPK/AKT-Nrf2 signaling axis; ⑤ JAK-STAT pathway. Moving forward, further research is needed to fully elucidate the specific mechanisms underlying S. grosvenorii’s anti-inflammatory properties.
Oxidative stress is the excessive production of reactive oxygen species (ROS) such as superoxide anion free radicals, hydroxyl free radicals and hydrogen peroxide in the body when the body is subjected to various harmful stimuli, which breaks the balance between oxidation and antioxidant systems in the body, leading to tissue damage, aging and a variety of diseases (Golden et al., 2002; Kregel and Zhang, 2007). As an important antioxidant enzyme in the body, superoxide dismutase (SOD) can convert superoxide anion free radicals into hydrogen peroxide and oxygen ions, thereby reducing lipid peroxidation (McCord and Edeas, 2005). Malondialdehyde (MDA) is one of free radicals in the body cause lipid peroxidation product, its content can be used to judge the body indirectly the extent of lipid peroxidation damage and aging (Barrera et al., 2018).
D-galactose is a widely used reagent to study the antioxidant aging effect in vivo. Drosophila has similar aging genes to humans, so it is often used in experimental studies of aging life span. Some studies have shown that Mog can increase SOD activity and reduce MDA content in brain tissue and serum of aging mice induced by D-galactose (Xiao et al., 2014). In addition, it can significantly prolong drosophila life expectancy and the highest life (Xiao et al., 2014). This suggests that S. grosvenorii has a role in delaying aging. Another study explored the mechanism by which S. grosvenorii delayed aging. The results of free radical scavenging experiments and autooxidative hemolysis experiments of rat erythrocytes showed that SGEs had a strong ability to scavenge hydroxyl free radicals and superoxide anion free radicals, and could effectively inhibit the production of MDA in the process of autooxidative hemolysis of rat erythrocytes and liver lipid peroxidation in a dose-dependent manner (Qi et al., 2006). It is worth noting that the important antioxidant active ingredient in SGEs may be Mog V (Qi et al., 2006).
In vitro maturation (IVM) of oocytes is one of the most important steps in the in vitro production (IVP) of human and livestock embryos. Addition of Mog V to IVM medium has demonstrated a substantial augmentation of oocyte maturation, attributable to its facilitative role in first polar body extrusion by enhancing mitochondrial biosynthesis as well as reducing intracellular ROS accumulation (Nie et al., 2020). After 44 h of in vitro maturation, Mog V added to the first polar body of oocytes, sustained 24 h in vitro, has exhibited commendable proficiency in upholding conventional oocyte morphology and nascent embryonic developmental competence. Mog V also led to a reduction in ROS levels, spindle formation, and anomalous chromosomal disposition (Nie et al., 2019). What is noteworthy is that Mog V may improve oocyte quality by upregulating silencing message regulator 1 (SIRT1) to reduce oxidative stress during in vitro aging (Nie et al., 2019).
An innovative polysaccharide, denoted as SGP and characterized by a molecular weight of 1.93 × 103 KDa, has been successfully extracted from residual components of S. grosvenorii. This compound has evinced robust in vitro antioxidant activity, particularly in terms of its proficiency in scavenging DPPH radicals. Moreover, SGP has demonstrated dose-dependent mitigation of ROS levels, apoptotic occurrences, and the proportion of necrotic cells in PC12 cells (a rat adrenal medullary pheochromocytoma cell line) (Zhu et al., 2020). In addition, SGP-1-1 stimulates the production of SOD and reduces the production of MDA by down-regulating the activation of TLR4/NF-κB p65 pathway to reduce oxidative stress caused by diabetic nephropathy (Gong et al., 2021).
Moving on to the realm of edibles, MGEs has emerged as a promising natural antioxidant for the preservation of quality in meat and meat products. The study confirmed the role of MGEs as inherently natural antioxidant in the preparation of dried mince. The natural antioxidants in MGEs not only have a protective effect against lipid and protein oxidation under vacuum storage conditions, but also maintain the reddish-brown character of dried minced meat (Cheng et al., 2017).
Taken together, existing studies indicate that the active substances in S. grosvenorii responsible for its anti-aging effects are Mog V and polysaccharides. The anti-aging mechanism of S. grosvenorii may be related to maintaining the balance of free radical metabolism by improving the activity of antioxidant enzymes in the body, removing excessive free radicals in the body, and reducing the lipid peroxidation of biofilm caused by free radicals, thereby protecting the normal metabolism of the body. In the future, there is a need to further verify whether other active components of S. grosvenorii possess anti-aging properties.
Diabetes mellitus, a pervasive chronic ailment, manifests as an endocrine and metabolic disorder characterized by hyperglycemia, late microvascular and macrovascular complications, of which diabetic nephropathy emerges as a profoundly detrimental consequence (Kaul et al., 2013). SGEs, MGEs, and multiple Mogs may inhibit diabetes-induced hyperglycemia and contribute to the prevention of diabetic complications associated with oxidative stress and hyperlipidemia.
The potential of SGEs to manifest anti-diabetic traits on spontaneously diabetic Goto-Kakizaki rats has been underscored by augmented fasting state insulin secretion, enhanced renal functionality, and fortified hepatic and plasma antioxidant attributes (Suzuki et al., 2007).
An experiment conducted on mice with alloxan-induced diabetic revealed that administering MGEs orally at different doses (100, 300, and 500 mg/kg) over a 4-week period led to significant reductions in blood glucose levels. Additionally, this treatment resulted in decreased levels of serum total cholesterol (TC), triglycerides (TG), and hepatic MDA, while increasing serum high-density lipoprotein cholesterol (HDL-C) levels and restoring hepatic antioxidant enzyme activities. Among these doses, the group receiving MGEs at 100 mg/kg exhibited the most pronounced hypoglycemic, hypolipidemic, and antioxidant effects, akin to the impact of Xiao Ke Wan-pills (at a dose of 894 mg/kg) (Qi et al., 2008). Another study demonstrated that diabetic mice given 150 or 300 mg/kg of MGEs for 5 consecutive weeks experienced a dose-dependent reduction in fasting blood glucose (FBG), glycated serum protein (GSP), serum insulin, homeostasis model assessment of insulin resistance (HOMA-IR), and serum atherogenic lipid profiles without any notable changes in their body weights or energy intake (Liu et al., 2019).
Mogs can prevent the development of diabetic nephropathy through antioxidant effects associated with inhibition the activation of heme oxygenase-1 (HO-1) and has no toxic effects in normal mice (Song et al., 2007). Moreover, S. grosvenorii crude extract (SG-ex) and its main sweet-tasting ingredient, monosodium glutamate (MSG), Mog V, as well as some minor ingredients such as Mog IV, siamenoside I, and Mog III exert anti-hyperglycemic effects in rats through the inhibition of wheat glycoenzyme activity (Suzuki et al., 2005). Professor Li Xiaobo’s team is very accomplished in the research on the anti-blood sugar of S. grosvenorii. They have developed a highly sensitive and rapid ultra-performance liquid chromatography/quadrupole time-of-flight mass spectrometry (UPLC-Q-TOF/MS) method, n conjunction with the Metabolynx™ software and mass defect filtering, to detect and characterize metabolites in the plasma, urine, bile, and feces of healthy rats and rats with type 2 diabetes mellitus (T2DM). Their study found that the main metabolic transformations of Mog V were achieved through dehydrogenation, deoxidation, oxidation, and isomerization (Zhou et al., 2018a). Additionally, they isolated low-polar Siraitia grosvenorii glycosides (L-SGgly), containing 1-3 glucose residues, from the 50% ethanol extract. L-SGgly contained 11-oxo-mog V, Mog V, Mog III, Mog IIE, Mog IIIA1, Mog IIA1, and Mog IA1, with the total content of Mogs being 54.4%, including 15.7% Mog IIA1 and 12.6% Mog IA1 (Zhang et al., 2020a). Compared to other parts of the LHG extract, L-SGgly exhibited promising effects on obese T2DM rats, resulting in a significant reduction in FBG levels and an improvement in insulin resistance. The anti-diabetic activity of L-SGgly may be attributed to increased glucagon-like peptide-1 (GLP-1) levels and activation of hepatic AMPK in T2DM rats (Zhang et al., 2020a). Furthermore, Li Xiaobo’s study suggests that the gut microbiota and their metabolites, such as Elusimicrobium, Lachnospiraceae_UCG-004, acetate, butyrate, and 1β-hydroxycholic acid, are potential dominant bacteria and biomarkers for SGEs and L-SGgly in reducing blood glucose and insulin resistance in T2DM rats (Zhang et al., 2021).
To sum up, S. grosvenorii is a promising dietary supplement for blood sugar control, especially for diabetics with a sweet tooth. Because it can both address diabetics’ desire for sweetness and help lower blood sugar. The hypoglycemic mechanism of S. grosvenorii appears to be linked to its ability to combat oxidative stress, inhibit the activity of wheat glycoenzyme, and activate AMPK in the liver. However, the specific mechanisms responsible for the hypoglycemic effects of S. grosvenorii are not yet fully understood and require further investigation.
S. grosvenorii exhibits promising potential in mitigating non-alcoholic fatty liver disease. Activation of the 5′ AMP-activated protein kinase (AMPK) signaling pathway within the hepatic milieu of diabetic murine models displayed a discernible MGEs dose-dependent pattern. Persistent administration of a MGEs at a concentration of 300 mg/kg over a span of 5 weeks yielded noteworthy amelioration. This was evident in the amelioration of hepatocyte polymorphism, mitigation of lipid accrual, and attenuation of steatosis, with restorative trends approximating basal states, as elucidated in the scholarly discourse (Liu et al., 2019). Mog V can improve the imbalance between lipid acquisition (de novo synthesis of lipids) and lipid clearance (lipolysis and fatty acid oxidation) by activating AMPK and subsequently exerting downstream regulation of SREBP1, PPAR-c and PPAR-α, thereby improving the liver steatosis induced by high fat diet in mice (Liu et al., 2019). Furthermore, the introduction of Nanoselenium S. grosvenorii (NSG) demonstrated multifaceted effects. NSG can not only enhance the diversity of intestinal flora and change its composition, but also significantly reduce the degradation of liver lysine metabolism and inhibit fatty acid β-oxidation, thus having a positive effect on obesity (Wang et al., 2023).
In summation, these discerning findings underscore the potential of S. grosvenorii and its derivative NSG as therapeutic agents to counteract non-alcoholic fatty liver disease and obesity-related pathways. Further exploration is needed to determine whether S. grosvenorii regulates pathways beyond AMPK and to unveil more specific regulatory mechanisms.
Recent investigations have unveiled the involvement of S. grosvenorii in the modulation of neurological disorders. Mog V has demonstrated a safeguarding capacity against corticosterone-induced damage in PC12 cells (Liu et al., 2023a). Furthermore, administration of Mog V has exhibited the potential to mitigate depressive manifestations within the chronic unpredictable mild stress (CUMS) depression model mice. Mechanistically, this intervention exerts an antidepressant influence by suppressing pathways related to inflammation and oxidative stress, along with the brain-derived neurotrophic factor (BDNF)/tyrosine kinase receptor B (TrkB)/AKT pathway (Liu et al., 2023a). These compelling revelations introduce innovative perspectives for the identification of emerging strategies in the realm of antidepressant interventions.
Single-session exhaustive exercise as well as repeated exhaustive exercise have been shown to elicit a decrease in the enzymatic activities of SOD, catalase (CAT), and glutathione peroxidase (GSH-PX). Simultaneously, an elevation in MDA and calcium ion (Ca2+) levels has been observed. This intricate interplay suggests that the potential anti-fatigue mechanism exhibited by S. grosvenorii and its extracts could be attributed to the suppression of oxidative stress and lipid peroxidation pathways.
SGEs have been found to dose-dependently enhance the physiological functions of Kunming mice during incremental load training. These enhancements encompass aspects such as exercise capacity, hypoxia and heat tolerance, and the attenuation of exercise-induced fatigue. The optimal dose was 15.0g/k·w·d. Notably, surpassing this dose threshold led to a diminishing impact on the mice’s physiological improvements (Yao et al., 2007). One important observation has been made regarding the administration of SGEs by gavage at different doses (100 mg/kg, 200 mg/kg, and 400 mg/kg) for 28 consecutive days. Evidently, such intervention imparts an extension to the swimming capacity of murine subjects. Concurrently, augmented hepatic and muscular glycogen reservoirs are discerned, accompanied by a concomitant attenuation of both blood lactic acid and serum urea nitrogen concentrations. Notably, the increased endurance of physical fatigue in mice showed a clear SGEs dose-dependent effect (Liu et al., 2013a).
Mogrol, an end product of Mog metabolism, has been shown to exert anti-fatigue effects by augmenting the relative protein expression of phosphorylated AMP-activated protein kinase alpha (p-AMPKα), AMPKα, peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α), and mitochondrial transcription factor A (TFAM) within the skeletal muscle of fatigued rats (Guo and Wen, 2022). Flavonoids have been documented to enhance the exercise capacity of male SD rats by fostering augmented blood perfusion and oxygen transport capabilities within the muscle tissue (Chen et al., 2020).
Leaf-derived flavonoids display a multifaceted anti-fatigue effect, encompassing the elevation of antioxidant enzyme activities, effective scavenging of excessive free radicals arising from exhaustive exercise, curbing lipid peroxidation, safeguarding myocardial mitochondria (Xie, 2018), and restraining apoptotic processes (Guo and Peng, 2012).
Mogs exhibit a pronounced capacity to bolster anti-fatigue potential and hypoxia tolerance in mice. These benefits can be attributed to heightened glycogen storage and accelerated lactate metabolism, pivotal factors contributing to improved endurance (Xia et al., 2012).
In summation, these studies demonstrate the potential utility of SGEs, mogrol, leaf-derived flavone, and Mogs, in combating fatigue and enhancing endurance in S. grosvenorii. The anti-fatigue effects demonstrated by S. grosvenorii and its derivatives manifest through a spectrum of mechanisms, including modulation of oxidative stress, enhancement of mitochondrial resilience, augmentation of vascular supply, and orchestrated protein expression. However, further studies are needed to reveal the specific molecules targeted by these active ingredients.
Schizophrenia is a kind of the signs and symptoms of unknown origin, main characteristic is showed signs of mental illness (Insel, 2010). MK801 to glutamate-N-methyl-D-aspartate (NMDA) receptor non-competitive antagonist, is currently the animal models of common tools used to construct the schizophrenia (Insel, 2010). Prevent prepulse inhibition (PPI) reflects the preattentional regulation of midbrain information filtering by the forebrain neural circuit. Decreased PPI means that the individual’s filtering mechanism of external environment information is decreased, which leads to information input overload and damage to the attention allocation process (Insel, 2010). Therefore, reducing PPI deficiency can be used as one of the indicators for improving psychiatric symptoms in the treatment of schizophrenia.
S. grosvenorii demonstrates a capacity for the modulation of schizophrenia. Noteworthy in this context is the efficacy of Mog V, has a alleviating effect on PPI deficits and social withdrawal induced by MK801(Ju et al., 2015; Ju et al., 2020). In addition, Mog V treatment reduced the total distance of movement and the number of spontaneous movements in MK801-induced schizophrenic mice (Ju et al., 2015). Moreover, Mog V has demonstrated a propensity to enhance cellular and neurochemical responses to MK801 within the confines of the medial prefrontal cortex (mPFC), as evidenced by empirical observations (Ju et al., 2020). Mog V, in concert with its metabolite 11-oxo-mogrol, serves as a preventive measure against MK-801-driven neuronal injury, largely attributed to its facilitation of neurite outgrowth and concurrent inhibition of cellular apoptosis, alongside the suppression of [Ca2+]i release (Ju et al., 2020). In addition, 11-oxo-mogrol regulates schizophrenia by reversing MK801-induced inactivation of AKT and phosphorylation of mammalian target of rapamycin (mTOR) (Ju et al., 2020).
PD is the second most common neurodegenerative disease in the world, which is common in middle-aged and elderly people. Its clinical manifestations are mainly bradykinesia, muscle stiffness and resting tremor, as well as non-motor symptoms such as anxiety, depression and cognitive impairment. Its main pathological feature is the degenerative loss of dopaminergic neurons in the substantia nigra region of the brain (Kalia and Lang, 2015).
Both Mog V and mogrol could significantly protect SY5Y cells from MPP+ -induced injury in vitro (Tang, 2021). In vivo experiments showed that Mog V could effectively improve the motor deficits and pathological changes of the brain in MPTP-induced PD mice, and had a significant protective effect on neurons in the substantia nigra region of mice. To the little mouse nigra district organization, according to the results of test of metabonomics in significant differences in metabolic substances after the treatment has 2 - D - hydrogen sphingosine, n-acetyl - L - glutamic acid, trimethylamine oxide, hexadecanoic acid, 9 - oleic aldehydes and 1, 11-11 of carbon dibasic acid. The recovered metabolic pathways were mainly focused on lipid and sphingolipid metabolism and amino acid synthesis. This suggests that Mog V plays a neuroprotective role mainly by regulating amino acid synthesis, lipid metabolism and gut-brain axis homeostasis, thereby alleviating mitochondrial dysfunction and substantia nigra neuronal loss (Tang, 2021). A recent investigation has exhibited that the administration of a 10 mg/kg dose of Mog V yielded a marked reversal of motor deficits and the consequential dopaminergic neuronal impairment triggered by rotenone in mice with PD (Luo et al., 2022). The mechanism may be that Mog V reduces overproduction of ROS in a dose-dependent manner, restores mitochondrial membrane potential, increases oxygen consumption rate and adenosine triphosphate (ATP) production, leading to a decrease in the number of apoptotic cells (Luo et al., 2022). Moreover, it is discerned that Mog V exhibits a safeguarding influence against PD, demonstrable in both the substantia nigra of mouse and SH-SY5Y cell line, by orchestrating a downregulation of SIRT3 acetylation (Luo et al., 2022). Furthermore, Mog V appears to intercede the injury-induced activity of the pivotal mitochondrial antioxidant enzyme, SOD2, thereby fortifying its neuroprotective impact (Luo et al., 2022).
Fibrosis-related diseases such as NASH, idiopathic pulmonary fibrosis (IPF), and myocardial fibrosis (MF) are all characterized by excessive deposition of extracellular matrix (ECM) (Jellis et al., 2010; Lederer and Martinez, 2018; Sheka et al., 2020). Fibrosis is the final pathological process of many diseases, however, only a limited number of drugs have shown effectiveness in reversing this condition.
The role of SGEs in the treatment of liver fibrosis has been proven. A study established a rat nutritional model of NASH by long-term feeding of CDAA to SD rats, and overcame dietary antagonism by feeding mice a high-fat diet lacking choline, low methionine, L-amino acid restriction, and free of trans fatty acids (CDAA-HF-T (-)). The majority of the phenotypic and mechanistic traits of human NASH are replicated in these models, including the rapid induction of fibrosis and the proliferation of lesions in the liver. SGEs prevent NASH in CDAA-HF-T (-) mice by inhibiting fibrosis without changing steatosis (Suzuki-Kemuriyama et al., 2021). It is possible that SGEs prevent liver fibrosis by preventing the NF-κB pathway from being active (Uno et al., 2023).
Within S. grosvenorii, active compounds with anti-fibrotic properties include the bitter components Mog IIIE and morgol. Mog IIIE has been found to reduce lung inflammation and inhibit ECM deposition by modulating the TLR4/MyD88-MAPK signaling cascade. In fact, at a dosage of 20 mg/kg, Mog IIIE was found to be more effective than Prednisone at 6.5 mg/kg in inhibiting pulmonary fibrosis (Tao et al., 2017). Additionally, research has demonstrated that Mog IIIE can reduce liver fibrosis by inhibiting the TLR4/MyD88/NF-κB signaling pathway (Yanan et al., 2023). Mogrol isolated from S. grosvenorii can not only reduce lung inflammation by activating AMPK, but also inhibit TGF-β1-induced mesenchymal transformation of alveolar epithelial cells and transdifferentiation of myofibroblasts through activation of the TGF-β1/Smad2/3 signaling pathway, thus exerting antifibrotic effects (Liu et al., 2021a). Notably, a dosage of 10 mg/kg of mogrol demonstrated better inhibition of bleomycin-induced pulmonary fibrosis compared to 40 mg/kg of nintedanib (Liu et al., 2021a).
Interestingly, it remains unclear from existing literature whether Mog V, which has the highest content among these components, possesses anti-fibrotic potential. It is worth noting that S. grosvenorii, may potentially contribute to the treatment, alleviation, or even reversal of fibrosis in the future. Before this can be done, further research and studies are needed to explore its potential therapeutic benefits in this regard.
Prostate cancer and bladder cancer stand as the prevailing malignancies within the domain of urology. Despite the array of available therapeutic interventions for these conditions, their efficacy remains suboptimal. The therapeutic effects of SGEs and multiple Mogs on different cancers have been revealed several times.
Mechanistically, pretreatment of murine skin with SGEs exhibited attenuation of TPA-induced activation of key signaling molecules, including extracellular signal-regulated kinase (ERK)1/2, p38 MAPK, c-Jun N-terminal kinase (JNK)1/2, PI3K, and Akt. This orchestrated reduction curtailed TPA-induced nuclear translocation of NF-κB subunits and phosphorylation of IkBa and p65, thereby culminating in a substantial impediment of 7,12-DMBA-TPA-evoked cutaneous tumor genesis (Weerawatanakorn et al., 2014).
LLE and Mog (LLE is a liquid form with Mog; and Mog is a highly purified Mog powder) can significantly reduce the viability of PC-3 (human prostate cancer cells) and T24 cells (human bladder transitional cell cancer cells), elicited a pronounced induction of G1 phase cell cycle arrest, induce endoplasmic reticulum stress and apoptosis, and thus treat prostate cancer and bladder cancer (Haung et al., 2023). Mog V and 11-oxo-Mog V showed a significant inhibitory effect on the two-stage carcinogenesis test of mouse skin tumors induced by peroxynitrite (ONOO2) as an initiator and TPA as a promoter (Takasaki et al., 2003). Moreover, 11-oxo-Mog V showcased substantial inhibition against two-stage skin carcinogenesis induced by 7,12-dimethylbenz [α]-anthracene (DMBA) and TPA (Takasaki et al., 2003), further accentuating its potential.
Furthermore, there is a investigation unveiled that within S. grosvenorii, 18 triterpenoids and 11-oxo-mogrol exhibited the capacity to curtail the generation of Epstein-Barr virus early antigen (EBV-EA) within Raji cells, incited by TPA (Ukiya et al., 2002). Researchers have tested whether mogrol has an effect on the proliferation of human prostate cancer cell line DU145, human liver cancer cell line HepG2, human lung cancer cell line A549, human nasopharyngeal carcinoma cell lines CNE1 and CNE2. The results showed that mogrol had obvious inhibitory effect on the proliferation of the above cells, but the inhibitory effect on the proliferation of human nasopharyngeal carcinoma CNE1 cells was the strongest. Mogrol has been shown to exert anti-tumor effects by inducing apoptosis (Fu et al., 2016) and G0/G1 cell cycle arrest (Liu et al., 2015), thereby inhibiting the proliferation of tumor cells.
The aforementioned studies have demonstrated that Mog V, 11-oxo-mog V, triterpenoids, 11-oxo-mogrol, and mogrol derived from S. grosvenorii exhibit anti-tumor effects. These effects are likely mediated by reducing cell viability, inducing G1-phase cell-cycle arrest, and triggering endoplasmic reticulum stress and apoptosis.
Although S. grosvenorii is traditionally known for its expectorant and cough suppressant properties, the above studies all suggest that S. grosvenorii has much more pharmacological effects than that. Among its various constituents, Mog V has been extensively studied, yet it remains uncertain whether any particular pharmacological effect outweighs the others. Furthermore, trace components like Mog IIIE and SGP-1-1 have also exhibited promising anti-fibrotic and anti-inflammatory effects. Despite the discovery of various pharmacological effects of S. grosvenorii and its promising efficacy in treating different diseases, it is important to note that these findings are primarily based on animal and cellular experiments. Therefore, it is crucial to conduct sufficient clinical studies to validate the therapeutic effects of S. grosvenorii. Furthermore, the majority of experiments conducted utilized either total Mogs or SGEs. However, there remains a lack of clarity regarding which specific monomeric substance is responsible for exerting distinct pharmacological effects. It is essential to determine which monomer exhibits the highest pharmacological activity and whether any synergistic or antagonistic effects exist among these monomers. Additionally, it is also necessary to compare the therapeutic effect of a single monomer of S. grosvenorii with a positive control. Moreover, it is crucial to gain a comprehensive understanding of the precise mechanisms and target molecules involved in regulating diseases. To address these critical questions, further experimental data will be necessary in future studies.
Although S. grosvenorii has been consumed for more than 300 years, elucidation of its toxicity is necessary before its large-scale use in food and medicine.
At present, some research teams have used acute toxicity test, long toxicity test, bone marrow micronucleus test, sperm abnormality test and reproductive and measuring lymphatic organ index to scrutinized the toxicological ramifications of S. grosvenorii and its extracts. Some researchers administered 3 g/mL of aqueous extract of S. grosvenorii to Kunming mice by gavage at a dose of 120 g/kg of dried fruit. After administration, although the mice’s intake of water and food was significantly reduced, this reduction was transient and did not cause lasting health effects. In the following 2 weeks, the mice maintained normal behavioral tendencies, sensory responses, fur quality, and intake of nutrients and water. Furthermore, the mice showed no obvious signs of toxicity, did not die, and the structural integrity of major organs was not compromised (Zhang et al., 2011). Bone marrow micronucleus frequencies, sperm abnormality coefficients, and reproductive and lymphoid organ indices were also virtually identical to those of untreated mice (Zhang et al., 2011).
Despite these experimental evidence, also cannot easily think of S. grosvenorii no pathogenic potential harm to the human body. Noteworthy, is the observation from a distinct study elucidating that while acute toxicity evaluations of Mogs (total glycosides ≥80%) in male Kunming mice proved non-deleterious, escalated doses (≥5 g/kg) elicited discernible, albeit modest, effects on micronucleus frequency and sperm aberrations (Liu et al., 2013b). In addition, some research teams did a screening experiment on the antitussive active ingredients of S. grosvenorii and found that when the dose of S. grosvenorii water decoction reached 50 g/kg of crude drug, some animals were in a poorer condition than control group mice (Liu et al., 2007), so the possibility of adverse stimulation of larger doses of S. grosvenorii could not be excluded.
Hence, whether harnessed for therapeutic interventions or harnessed as an innate nutritive sweetening agent, the possible genotoxic propensity of S. grosvenorii at high doses should be considered with caution.
S. grosvenorii is exclusively grown in China (He et al., 2012). With special requirements on ecological conditions, its distribution is limited to certain regions in provinces such as Guangxi, Guangdong, Hunan, Jiangxi, with Guangxi accounting for approximately 80% of the cultivated area in China (Li and Lan, 2019). Currently, the primary focus of cultivation for S. grosvenorii revolves around the green fruit variety, followed by La Jiang fruit and long beach fruit (Tang, 2023b). The planting of S. grosvenorii typically occurs through the use of cutting seedings, which takes place annually from mid-March to mid-April (Tang, 2023b). Prior to planting, it is necessary to construct a shed (Tang, 2023b). Once the S. grosvenorii seedlings reach a height of 15 cm, the main vine is carefully chosen and transferred into the shed for pruning (Tang, 2023b). Artificial pollination becomes imperative after flowering (Tang, 2023b). Traditional cultivation of S. grosvenorii faced challenges including low yield, low seedling survival rate, and susceptibility to pests and diseases. However, advancements in technology have successfully addressed these issues, resulting in a significant increase in the yield of S. grosvenorii seedlings from less than 20% to over 90% (Li and Lan, 2019). The new varieties of S. grosvenorii histocultured seedlings have many excellent characteristics such as no virus, good disease resistance, and strong adaptability. The conquest of this technical problem has greatly reduced the cost of labor and agricultural materials. Overcoming this technical obstacle has not only reduced labor and agricultural costs but has also expanded the cultivation areas beyond mountainous regions (Li and Lan, 2019). Guilin City, known for being the largest cultivation area for S. grosvenorii, recorded a cultivation area of 225,500 mu and an LHG output of 2,252 million in 2022 (Qin et al., 2023).
Traditionally, S. grosvenorii is used as a traditional Chinese medicine for the treatment of lung fire and dry cough, sore throat and loss of voice, and intestinal dryness and constipation (Jiang et al., 2015). Currently, its primary application lies in its use as a natural sweetener. In 1995, the United States Food and Drug Administration (FDA) approved the application of Mog in food products, and after obtaining the United States FDA Generally Recognized as Safe (GRAS) certification in 2011, S. grosvenorii extracts/processed products have successfully entered United States market (Pawar et al., 2013). In China, Mog was approved as a food additive at the 17th meeting of the National Food Additives Committee in 1996, and can be used as a sweetener to partially or completely replace sucrose (Chen et al., 2022), The official implementation of the “Food Additives of Mog” in 2017 further clarified its application in Chinese market (Chen et al., 2022). Several other countries, including Japan, South Korea, and the United Kingdom, also permit the use of Mog as a food additive (Chen et al., 2022).
Nowadays, Mog substances are widely used in the food industry. From 2016 to 2020, the amount of Mogs utilized in the United States market has steadily increased, with the global Mog market expected to exceed USD 1 billion, growing at a compound annual growth rate of over 40% from its approximate value of USD 250 million in 2016 (Chen et al., 2022). In the food industry, LHG is employed to produce a variety of consumables, including LHG-fermented wine, LHG cake (Hong et al., 2021; Liu et al., 2023b), LHG preserved fruit, and LHG fruit compound beverages (Li et al., 2006a).
S. grosvenorii has a certain presence in the healthcare industries. In the pharmaceutical field, products like Luo Han Guo Paoteng Tablet and Luo Han Guo yanhou Tablet have been developed to provide relief from pharyngeal discomfort (Jin et al., 1997; Jiang et al., 2011; Chen et al., 2015). Luo Han Guo Lvcha Granules are used to boost immunity, Luo Han Guo Sydney Cream and Jinyinhua Luo Han Guo Lozenges are used to treat sore throats (Zhang et al., 2017). Additionally, when combined with ingredients like hawthorn, yam, and tangerini peel, LHG can be used to create a health drink that strengthens the stomach and aids digestion (Sun, 2022). In China and Japan, Mog extract has been approved as a healthy alternative to sugar for obese and diabetic patients (Gong et al., 2019).
In recent years, the livestock, tobacco and cosmetics industries also witnesses S. grosvenorii’s influence. As its residual components find utility in enhancing feed digestibility, augmenting poultry growth, and ameliorating slaughter performance, evident in the supplementation of Guangxi hemp chicken diets with S. grosvenorii residues (Xu et al., 2022a; Wang et al., 2023; Xiao et al., 2023). Furthermore, incorporating S. grosvenorii flavoring into smoking products not only enhances the sensory experience but also helps protect the health of smokers by reducing harmful pulmonary consequences, alleviating throat irritation, and even aiding smoking cessation (Li et al., 2006a; Zhang et al., 2018; Wu et al., 2022b). Moreover, there are many patented products of S. grosvenorii that find applications in the cosmetic industry due to their skin whitening and moisturizing effects (Zhang and Zhang, 2011; Yang, 2022).
Although the application scope of S. grosvenorii covers a wide range of food, drugs, cosmetics, health products, animal feed and other fields have broad prospects. A problem that cannot be ignored is that S. grosvenorii extracts/processed products are still cold in the Chinese market where they originate. There are many challenges that need to be solved in order to further popularize it in the market.
The development and widespread use of S. grosvenorii as a sweetener and functional product in the market is facing five urgent challenges.
Firstly, the supply of raw materials of S. grosvenorii is insufficient, and the production level of S. grosvenorii extract products is not high. S. grosvenorii extract products are made from the natural S. grosvenorii plant, which has distinct cyclical, regional, and seasonal characteristics in terms of planting, harvesting, and acquisition. The availability of raw materials is influenced by various factors such as geographical limitations and climate changes, making it vulnerable to price fluctuations and an unstable supply (Zhao, 2023). S. grosvenorii is a crop that requires ample sunlight during its growth process, but prolonged exposure to intense light and low temperatures can hinder its growth. As a result, the cultivation area for S. grosvenorii is limited (Tang, 2023a). Currently, most S. grosvenorii cultivation is carried out by individual farmers without proper scientific planning (Tang, 2023a), leading to issues like blindly expanding cultivation trends and inconsistent production standards. Additionally, the lack of effective supervision in S. grosvenorii cultivation has prevented standardized operations, deviating from the intended goals. These problems directly hinder the transformation of the S. grosvenorii industry’s advantages into economic benefits.
Secondly, intense competition exists among sweetener products. The natural sweeteners market is currently flooded with a wide range of competing products, and unfortunately, Mog faces certain drawbacks regarding its price, stability, and taste when compared to other popular high-frequency sweeteners. The extraction cost of Mog is relatively high, leading to end-product prices that are higher than similar alternatives (Zhao, 2023). In addition, the traditional processing methods used in the original production areas of S. grosvenorii, such as firewood baking, have resulted in a distinct roasted taste that is not easily recognized by consumers. Moreover, the purity of available Mogs in the market is generally low, ranging from 25% to 50%, and this low purity level is associated with an unpleasant taste (Huang et al., 2021). To mask its distinctive odor, Mog is often combined with other sweeteners like erythritol, further increasing costs. As a result, in recent years, major food and beverage companies have shown a preference for Steviol glycosides, another natural high-frequency sweetener, instead of S. grosvenorii (Zhao, 2023).
Thirdly, there is limited diversity in the range of S. grosvenorii end products. Mog is primarily used as a sweetener, resulting in its predominant existence as an additive in the market. Moreover, the history of using Mog as a sweetener is relatively short, and research on its pharmacological function and toxicology is still in its early stages. Therefore, the number and variety of end products containing S. grosvenorii as the main ingredient are still insufficient. Between 2014 and 2018, although SGEs were used in approximately 2,507 food, beverage, dairy, and other products worldwide, the number of food and beverage products containing SGEs in 2018 was actually lower than in 2014 (Li and Lan, 2019). Due to the high cost of SGE and its lack of competitiveness in terms of price, it faces challenges in achieving rapid growth both domestically and internationally.
Fourthly, the market recognition of S. grosvenorii products is relatively low. As a specialty cash crop, consumers are not yet familiar with the efficacy of S. grosvenorii, and the lack of branding and market education has resulted in a limited understanding and trust in its characteristics, functions, and advantages. This lack of awareness makes it difficult to generate consumer demand and preference for S. grosvenorii products. Despite the abundance of literature supporting the pharmacological efficacy of S. grosvenorii, insufficient publicity and popularization efforts have created the impression among consumers that there is no difference between Mog as a sweetener and other natural sweeteners.
Fifthly, the marketing channels for S. grosvenorii products are relatively narrow. Most S. grosvenorii products are currently marketed through direct sales channels, resulting in high operating and management expenses and significant labor costs. This has led to increased prices for the extract products, which were already at a disadvantage when compared to similar alternatives (Zhao, 2023). Furthermore, the lack of promotional efforts by distributors has severely restricted the speed of S. grosvenorii’s end products’ rollout. Although in recent years, live broadcasting has emerged as a new sales method, some sellers have tried to promote S. grosvenorii products through this platform. However, due to the low market awareness of SGEs products, this sales method has not proven as effective as expected, despite its convenience.
In order to further expand both domestic and overseas markets for S. grosvenorii as a sweetener or functional product, it is crucial to implement effective strategies and methods for its intensive development and utilization.
While the existing varieties of S. grosvenorii have addressed issues such as low survival rates, pests, diseases, and low yield, there are still certain challenges that distinguish it from Stevia, another natural sweetener plant. These include a lower content of effective components, germplasm degeneration, mixing, and notable problems related to continuous cropping. These factors contribute to the degradation of the soil’s ecological environment. Therefore, it is crucial to continue in-depth research to develop new S. grosvenorii varieties that offer high yield and quality, strong resistance, adaptability, and reduced planting and management costs. This will involve optimizing the regional and seasonal layouts of S. grosvenorii varieties to enhance both yield and quality, as well as extending the availability of fresh fruits in the market.
Moving forward, it is essential to standardize the planting mode for S. grosvenorii, while also accurately estimating market demand and avoiding blind production. Improving the intensity of planting will help to prevent the waste of raw materials and reduce planting costs. To achieve this, it is crucial to propose unified production and technical standards.
Looking ahead, it is crucial to focus on the development and application of S. grosvenorii as a natural sweetener in response to the growing global demand for sugar reduction and increased health consciousness. This can be achieved through extensive research on the development and utilization of the S. grosvenorii serie of natural sweetener products, with a particular emphasis on higher purity MGE. Expanding the application scope of Mog products and establishing a comprehensive support system will be key. Furthermore, there should be efforts to improve the processing techniques of existing S. grosvenorii products, aiming to reduce processing costs and develop efficient and labor-saving technological equipment. Streamlining the production process to achieve continuous and automated production will be crucial, ultimately optimizing human labor in the process.
As a dual-use traditional Chinese medicine, the application of medicines/healthcare products related to the pharmacological activities of S. grosvenorii, such as anti-inflammatory, antioxidant, anti-tumor, anti-fatigue, etc., has yet to be further researched. The pace of research should be accelerated to comprehensively develop the value of medicinal constituents of S. grosvenorii, and to obtain new products with the special efficacy of S. grosvenorii, so as to satisfy the differentiated needs of the market.
In the future, it is vital to increase the cultural promotion of S. grosvenorii in order to enhance consumer awareness and understanding. Various strategies can be employed to achieve this, such as organizing S. grosvenorii cultural festivals, exhibitions, and establishing offline counters. Leveraging both traditional paper media and modern digital channels, public relations and publicity initiatives can be implemented. The focus of these efforts should center around highlighting the medicinal value of S. grosvenorii and its natural, healthy advantages as a sweetener. By emphasizing these key points to deepen consumer impressions of S. grosvenorii and its extracted products, ensuring that they are well-informed and more receptive to its benefits.
In general, price plays a pivotal role in consumers’ decision-making process when it comes to purchasing a product. As a result, it becomes crucial to align the pricing of S. grosvenorii products with the prevailing market environment. This necessitates a thorough evaluation and adjustment of prices to ensure competitiveness. Moreover, it is imperative to establish a robust marketing system for physical stores to enhance the efficiency of product rollout. Simultaneously, optimizing online sales channels is equally important. By leveraging e-commerce platforms, social media, and various online networks, to effectively reach new customer segments, thereby capturing untapped market opportunities.
Based on our comprehensive understanding of S. grosvenorii, we now present a forward-looking analysis of the future direction of S. grosvenorii production in 2030. To ensure the sustainability of plant resources, the planting scale and area for S. grosvenorii will be expanded. Furthermore, the processing techniques will be optimized, yield and quality will be improved, freshness duration of LHG will be extended, and stability of the S. grosvenorii supply chain will be enhanced. In addition, the industry standards and norms will be established, a highly regarded and popular S. grosvenorii brand will emergence. The industrial production of high purity Mogs and the masking of the unique smell of Mogs will be realized. These developments will expand the market for S. grosvenorii as a sweetener. Since 2010, there has been a steady increase in the number of publications and citations on S. grosvenorii, particularly focusing on its health effects and biosynthesis of Mogs (Yeung, 2023). However, at present, there is limited clinical trial evidence regarding the pharmacological effects of S. grosvenorii. Therefore, it is reasonable to expect that by 2030, there will be a substantial amount of clinical data evaluating the overall pharmacological effects and toxicity of S. grosvenorii. This will shed light on the biological effects of the main chemical components within total Mogs of S. grosvenorii, as well as their interactions with other compounds.
Based on the existing research work, this review comprehensively summarizes the chemical composition, pharmacological effects, toxicology, status of resources development, and applications of S. grosvenorii. Additionally, this review offers strategies and methods of intensive development and utilization of S. grosvenorii. It is hoped that this paper will inspire future cultivatiors, researchers and resource developers of S. grosvenorii, driving wider applications in the fields of food, medicine, health products, agricultural products, and cosmetics.
HH: Conceptualization, Methodology, Writing–review and editing. ZP: Conceptualization, Methodology, Writing–original draft, Writing–review and editing. SZ: Formal Analysis, Writing–review and editing. WL: Formal Analysis, Writing–review and editing. DL: Formal Analysis, Writing–review and editing. SH: Investigation, Writing–review and editing. YZ: Funding acquisition, Project administration, Resources, Validation, Writing–review and editing. WW: Funding acquisition, Project administration, Resources, Supervision, Writing–review and editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. Financial support of this research was provided by “Development, Application and Demonstration of High-value Innovation Chain of Luo Han Guo, a Traditional Chinese Medicine and Food Ingredient” (which belonging to “the proposed project of Hunan Province High-tech Industry Science and Technology Innovation Leadership Program 2021–2022”, grant no. 2021GK4018) and “Hunan Natural Sweetener Engineering Technology Research Center” (grant no. 2020TP2004). The funder was not involved in the study design, collection, analysis, interpretation of data, the writing of this article, or the decision to submit it for publication.
Authors HH, ZP, SZ, WL, DL, SH were employed by Hunan Huacheng Biotech, Inc.
Authors HH, ZP, SZ, WL, DL, SH were employed by Hunan Natural Sweetener Engineering Technology Research Center.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Akihisa, T., Hayakawa, Y., Tokuda, H., Banno, N., Shimizu, N., Suzuki, T., et al. (2007). Cucurbitane glycosides from the fruits of Siraitia grosvenorii and their inhibitory effects on Epstein-Barr virus activation. J. Nat. Prod. 70, 783–788. doi:10.1021/np068074x
Barrera, G., Pizzimenti, S., Daga, M., Dianzani, C., Arcaro, A., Cetrangolo, G. P., et al. (2018). Lipid peroxidation-derived aldehydes, 4-hydroxynonenal and malondialdehyde in aging-related disorders. Antioxidants 7, 102. doi:10.3390/antiox7080102
Blot, S. (2021). Antiseptic mouthwash, the nitrate–nitrite–nitric oxide pathway, and hospital mortality: a hypothesis generating review. Intensive Care Med. 47, 28–38. doi:10.1007/s00134-020-06276-z
Cai, X., He, L., Zhou, G., Li, S., and Liao, X. (2021). Mogroside IIe ameliorates cardiomyopathy by suppressing cardiomyocyte apoptosis in a type 2 diabetic model. Front. Pharmacol. 12, 650193. doi:10.3389/fphar.2021.650193
Cao, F., Zhang, Y., Li, W., Shimizu, K., Xie, H., and Zhang, C. (2018). Mogroside IVE attenuates experimental liver fibrosis in mice and inhibits HSC activation through downregulating TLR4-mediated pathways. Int. Immunopharmacol. 55, 183–192. doi:10.1016/j.intimp.2017.12.023
Chaturvedula, V. S. P., and Meneni, S. R. (2015). A new cucurbitane glycoside from siraitia grosvenorii. Nat. Product. Commun. 10, 1934578X1501000–1523. doi:10.1177/1934578X1501000908
Chen, B., Chen, W., Zhang, Y. M., and Wu, W. (2015). Studies on the technics of preparing effervescent tablets of Siraitia grosvenorii. Acta Med. Sin., 101–103.
Chen, G., Mo, W. B., and Li, G. F. (2020). Effect of rhubarb flavone on the expression of Flk-1 and bFGF in skeletal muscle of exercise rats. J. Basic Chin. Med. 26, 45–48.
Chen, M. M., Liu, G. G., He, A. L., and Huang, H. X. (2022). Analysis and prospect of patent strategy of Chinese monk fruit industry. Sci-Tech Dev. Enterp., 10–14.
Chen, Q. B., Chen, H. Y., Li, J., Yi, X. H., and Su, X. J. (2003). The determination of the molecular weight of Siraitia grosvenorii polysaccharides by HPLC. Chin. Tradit. Herb. Med. 34, 1075–1076.
Chen, Q. B., Yang, J. X., Chen, Z. Q., Yi, X. H., and Wei, Z. B. (2006). Separation, purification and identification of flavonol glycoside from momordica grosvenori leafs. Guangxi Sci., 35–36.
Chen, X., Zhuang, J., Liu, J., Lei, M., Ma, L., Chen, J., et al. (2011). Potential AMPK activators of cucurbitane triterpenoids from Siraitia grosvenorii Swingle. Bioorg. Med. Chem. 19, 5776–5781. doi:10.1016/j.bmc.2011.08.030
Cheng, J.-R., Liu, X.-M., Zhang, Y.-S., Zhang, Y.-H., Chen, Z.-Y., Tang, D.-B., et al. (2017). Protective effects of Momordica grosvenori extract against lipid and protein oxidation-induced damage in dried minced pork slices. Meat Sci. 133, 26–35. doi:10.1016/j.meatsci.2017.04.238
Chu, D., Yaseen, A., Wang, L., Chen, B., Wang, M., Hu, W., et al. (2019). Two new cucurbitane glycosides from the fruits of Siraitia grosvenori. Chem. Pharm. Bull. 67, 721–724. doi:10.1248/cpb.c19-00210
Cui, S., Zang, Y., Xie, L., Mo, C., Su, J., Jia, X., et al. (2023). Post-ripening and key glycosyltransferase catalysis to promote sweet mogrosides accumulation of siraitia grosvenorii fruits. Molecules 28, 4697. doi:10.3390/molecules28124697
Daboor, S., Syed, F., Masood, S., Al-Azab, M., Nori, E., and Nori, M. E. (2015). A review on Streptococcus Mutans with its diseases dental caries, dental plaque and endocarditis. Indian J. Microbiol. Res. 2 (2), 76–82.
Dixon, R. A., Dey, P. M., and Lamb, C. J. (1983). “Phytoalexins: enzymology and molecular biology,” in Advances in enzymology - and related areas of molecular biology. Editor A. Meister (USA: Wiley), 1–136. doi:10.1002/9780470123010.ch1
Dou, T., Wang, J., Liu, Y., Jia, J., Zhou, L., Liu, G., et al. (2022). A combined transcriptomic and proteomic approach to reveal the effect of mogroside V on OVA-induced pulmonary inflammation in mice. Front. Immunol. 13, 800143. doi:10.3389/fimmu.2022.800143
Duan, J., Zhu, D., Zheng, X., Ju, Y., Wang, F., Sun, Y., et al. (2023). Siraitia grosvenorii (swingle) C. Jeffrey: research progress of its active components, pharmacological effects, and extraction methods. Foods 12, 1373. doi:10.3390/foods12071373
El-Boghdadly, K., Bailey, C. R., and Wiles, M. D. (2016). Postoperative sore throat: a systematic review. Anaesthesia 71, 706–717. doi:10.1111/anae.13438
Fang, C., Wang, Q., Liu, X., and Xu, G. (2017). Metabolic profiling analysis of Siraitia grosvenorii revealed different characteristics of green fruit and saccharified yellow fruit. J. Pharm. Biomed. Anal. 145, 158–168. doi:10.1016/j.jpba.2017.06.046
Fu, Y. X., Wang, L., and Li, D. P. (2016). Activity and mechanism of anticancer properties of mogrol. Guihaia Jounal 36, 1369–1375. doi:10.11931/guihaia.gxzw201502025
Golden, T. R., Hinerfeld, D. A., and Melov, S. (2002). Oxidative stress and aging: beyond correlation. Aging Cell 1, 117–123. doi:10.1046/j.1474-9728.2002.00015.x
Gong, P., Cui, D., Guo, Y., Wang, M., Wang, Z., Huang, Z., et al. (2021). A novel polysaccharide obtained from Siraitia grosvenorii alleviates inflammatory responses in a diabetic nephropathy mouse model via the TLR4-NF-κB pathway. Food Funct. 12, 9054–9065. doi:10.1039/d1fo01182k
Gong, P., Guo, Y., Chen, X., Cui, D., Wang, M., Yang, W., et al. (2022). Structural characteristics, antioxidant and hypoglycemic activities of polysaccharide from siraitia grosvenorii. Molecules 27, 4192. doi:10.3390/molecules27134192
Gong, X., Chen, N., Ren, K., Jia, J., Wei, K., Zhang, L., et al. (2019). The fruits of siraitia grosvenorii: a review of a Chinese food-medicine. Front. Pharmacol. 10, 1400. doi:10.3389/fphar.2019.01400
Guo, W. Q., and Wen, Z. L. (2022). Anti-exercise fatigue effect of active components of medicinal plant siraitia grosvenorii (swingle). Mol. Plant Breed. 20, 7923–7930.
Guo, Y. J., and Peng, F. L. (2012). Effects of flavones on antioxidant enzymes and apoptosis in heart of exhauted exercise rats. J. Guangxi Normal Univ. Sci. Ed. 30, 143–147. doi:10.16088/j.issn.1001-6600.2012.02.018
Hamada, S., Koga, T., and Ooshima, T. (1984). Virulence factors of Streptococcus mutans and dental caries prevention. J. Dent. Res. 63, 407–411. doi:10.1177/00220345840630031001
Haung, R., Saji, A., Choudhury, M., and Konno, S. (2023). Potential anticancer effect of bioactive extract of monk fruit (<i&gt;Siraitia grosvenori&lt;/i&gt;) on human prostate and bladder cancer cells. J. Cancer Ther. 14, 211–224. doi:10.4236/jct.2023.145019
He, W. P., Liu, L. J., He, C. M., Li, A. Y., and Li, X. M. (2017). Study on low glycemic index (ci) food and pharmacological effects of S. grosvenorii. J. Light. Ind. 27.
He, W. P., Zhu, X. Y., and He, C. W. (2012). Research progress in application of Momordica grosvenori swingle fruit and problems in product development. Sci. Technol. Food Industry 33, 400–402. doi:10.13386/j.issn1002-0306.2012.11.101
Hong, W. L., Xu, J. Q., Sang, Z. Y., Cao, M., and Dai, Z. Q. (2021). Study on optimization of mousse cake process formula by responsesurface methodology. 32–36.
Hou, D. Y., Hui, R. H., Li, T. C., Diao, Q. P., and Qi, Y. Q. (2020). Comparative analysis of fatty acids extracted from siraitia grosvenorii by two methods. J. Anshan Normal Univ. 22, 30–33.
Huang, H. X., Liu, G. G., He, A. L., Huang, J., and Xiong, Y. (2021). Development status and trend of high-purity mogroside industry. Sci-Tech Dev. Enterp., 34–36+39.
Jellis, C., Martin, J., Narula, J., and Marwick, T. H. (2010). Assessment of nonischemic myocardial fibrosis. J. Am. Coll. Cardiol. 56, 89–97. doi:10.1016/j.jacc.2010.02.047
Ji, L. (2016). Effects of different drying and processing methods on vitamin C, color, phenolics, antioxidant activity. mogroside V Luo Han Guo (Siraitia Grosvenorii) drink.
Jia, X., Liu, J., Shi, B., Liang, Q., Gao, J., Feng, G., et al. (2019). Screening bioactive compounds of siraitia grosvenorii by immobilized β2-adrenergic receptor chromatography and druggability evaluation. Front. Pharmacol. 10, 915. doi:10.3389/fphar.2019.00915
Jia, Z., and Yang, X. (2009). A minor, sweet cucurbitane glycoside from Siraitia grosvenorii. Nat. Prod. Commun. 4, 1934578X0900400–772. doi:10.1177/1934578x0900400606
Jiang, C. X., Chen, J. G., and Luo, T. (2015). Nutritional value analysis of medicinal and edible plant. J. Anhui Agric. Sci. 43, 282–284. doi:10.3969/j.issn.0517-6611.2015.11.105
Jiang, J. L., Liang, J., and Yang, Y. Y. (2020). Research progress on pharmacological and toxicological effects of mogroside. Mod. Prev. Med. 47, 2246–2248.
Jiang, S. C., He, L. M., Wu, Y., and Luo, C. R. (2011). An additive for smoked Siraitia grosvenorii, their preparation and Application: CN Patent 101633866.
Jin, C. H., Jiang, X. L., Hong, T., Wang, Y. J., Xu, T. H., and Zhang, D. M. (1997). Pharmacological studies of luohanguo yanhou tablet. J. Chin. Med.Mate, 574–577. doi:10.13863/j.issn1001-4454.1997.11.017
Jorn, A., Bruce, J. P., Lauren N, S., Ingar, O., and Floyd E, D. (2005). Defining the normal bacterial flora of the oral cavity. J. Clin. Microbiol. 13, 5721–5732. doi:10.1128/JCM.43.11.5721-5732.2005
Ju, P., Ding, W., Chen, J., Cheng, Y., Yang, B., Huang, L., et al. (2020). The protective effects of Mogroside V and its metabolite 11-oxo-mogrol of intestinal microbiota against MK801-induced neuronal damages. Psychopharmacology 237, 1011–1026. doi:10.1007/s00213-019-05431-9
Ju, P. J., Ding, W. H., Li, X. B., and Wang, M. Y. (2015). Effect of mogroside V on schizophrenia-like behavior induced by dizocilpine in mice. J. Clin. Psychiatry 25, 231–233.
Kalia, L. V., and Lang, A. E. (2015). Parkinson’s disease. Lancet 386, 896–912. doi:10.1016/S0140-6736(14)61393-3
Kasai, R., Matsumoto, K., Nie, R.-L., Zhou, J., and Tanaka, O. (1988). Glycosides from Chinese medicinal plant, Hemsleya panacis-scandens, and structure-taste relationship to cucurbitane glycosides. Chem. Pharm. Bull. 36, 234–243. doi:10.1248/cpb.36.234
Kaul, K., Tarr, J. M., Ahmad, S. I., Kohner, E. M., and Chibber, R. (2013). “Introduction to diabetes mellitus,” in Diabetes. Editor S. I. Ahmad (New York: Springer), 1–11. doi:10.1007/978-1-4614-5441-0_1
Kim, M.-S., Kim, D.-S., Yuk, H. J., Kim, S.-H., Yang, W.-K., Park, G. D., et al. (2023). Siraitia grosvenorii extract attenuates airway inflammation in a murine model of chronic obstructive pulmonary disease induced by cigarette smoke and lipopolysaccharide. Nutrients 15, 468. doi:10.3390/nu15020468
Kinghora, A. D., Soejarto, D. D., and Inglett, G. E. (1986). Sweetening agents of plant origin. Crit. Rev. Plant Sci. 4, 79–120. doi:10.1080/07352688609382220
Kregel, K. C., and Zhang, H. J. (2007). An integrated view of oxidative stress in aging: basic mechanisms, functional effects, and pathological considerations. Am. J. Physiology-Regulatory, Integr. Comp. Physiology 292, R18–R36. doi:10.1152/ajpregu.00327.2006
Lederer, D. J., and Martinez, F. J. (2018). Idiopathic pulmonary fibrosis. N. Engl. J. Med. 378, 1811–1823. doi:10.1056/NEJMra1705751
Lee, S.-H., Kim, W.-H., Ju, K.-W., Lee, M.-S., Kim, H.-S., Lee, J.-H., et al. (2021). Antibacterial and anti-inflammatory potential of mouthwash composition based on natural extracts. Appl. Sci. 11, 4227. doi:10.3390/app11094227
Lee, Y. M., Kim, M., Yuk, H. J., Kim, S.-H., and Kim, D.-S. (2023). Siraitia grosvenorii residual extract inhibits inflammation in RAW264.7 macrophages and attenuates osteoarthritis progression in a rat model. Nutrients 15, 1417. doi:10.3390/nu15061417
Lei, C. H. (2017). The method of microwave digestion-ICP-OES for the determination of 11 trace elements in S. grosvenorii was established. Light Industry Sci. Technol. 33, 7–8.
Li, C., Lin, L.-M., Sui, F., Wang, Z.-M., Huo, H.-R., Dai, L., et al. (2014). Chemistry and pharmacology of Siraitia grosvenorii: a review. Chin. J. Nat. Med. 12, 89–102. doi:10.1016/S1875-5364(14)60015-7
Li, D., Ikeda, T., Matsuoka, N., Nohara, T., Zhang, H., Sakamoto, T., et al. (2006a). Cucurbitane glycosides from unripe fruits of lo han kuo (siraitia grosvenori). Chem. Pharm. Bull. 54, 1425–1428. doi:10.1248/cpb.54.1425
Li, D., Ikeda, T., Nohara, T., Liu, J., Wen, Y., Sakamoto, T., et al. (2007). Cucurbitane glycosides from unripe fruits of Siraitia grosvenori. Chem. Pharm. Bull. 55, 1082–1086. doi:10.1248/cpb.55.1082
Li, D.-P., El-Aasr, M., Ikeda, T., Ogata, M., Miyashita, H., Yoshimitsu, H., et al. (2009). Two new cucurbitane-type glycosides obtained from roots of Siraitia grosvenori S<small>WINGLE</small>. Chem. Pharm. Bull. 57, 870–872. doi:10.1248/cpb.57.870
Li, F., Yang, F., Liu, X., Wang, L., Chen, B., Li, L., et al. (2017). Cucurbitane glycosides from the fruit of Siraitia grosvenori and their effects on glucose uptake in human HepG2 cells in vitro. Food Chem. 228, 567–573. doi:10.1016/j.foodchem.2017.02.018
Li, J., Li, P. B., and Yuan, G. J. (2008). The antitussive effect of water extrat of grosvenor momordica fruit. J. Hainan Med. Univ., 16–18. doi:10.13210/j.cnki.jhmu.2008.01.005
Li, L., Zheng, W., Wang, C., Qi, J., and Li, H. (2020). Mogroside V protects against hepatic steatosis in mice on a high-fat diet and LO2 cells treated with free fatty acids via AMPK activation. Evidence-Based Complementary Altern. Med. 2020, e7826874. doi:10.1155/2020/7826874
Li, S., Wang, H. S., and Zhang, G. Y. (2003). The analysis of seed oil from Siraitia grosvenorii. Guangxi Med. J. 25, 850–852.
Li, X. Y., and Lan, F. S. (2019). Development prospect of Siraitia grosvenorii industry in China. J. Guangxi Agric. 34, 30–33.
Lim, T. K. (2012). Edible medicinal and non-medicinal plants: volume 2, fruits. Dordrecht: Springer Netherlands. doi:10.1007/978-94-007-1764-0
Liu, B., Yang, J., Hao, J., Xie, H., Shimizu, K., Li, R., et al. (2021a). Natural product mogrol attenuates bleomycin-induced pulmonary fibrosis development through promoting AMPK activation. J. Funct. Foods 77, 104280. doi:10.1016/j.jff.2020.104280
Liu, C., Cai, T. Y., Zhao, X. M., Ma, L. Q., Dou, D. Q., and Sun, Y. X. (2015). Apoptosis effect of Siraitia grosvenorii extracts on lung cancer cells A549 andts mechanisms. Chin. Pharmacol. Bull. 31, 1310–1314.
Liu, D.-D., Ji, X.-W., and Li, R.-W. (2013a). Effects of siraitia grosvenorii fruits extracts on physical fatigue in mice. Iran. J. Pharm. Res. 12, 115–121. Available at: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3813191/ (Accessed July 18, 2023).
Liu, H., Du, Y., Liu, L. L., Liu, Q. S., Mao, H. H., and Cheng, Y. (2023a). Anti-depression-like effect of Mogroside V is related to the inhibition of inflammatory and oxidative stress pathways. Eur. J. Pharmacol. 955, 175828. doi:10.1016/j.ejphar.2023.175828
Liu, H., Qi, X., Yu, K., Lu, A., Lin, K., Zhu, J., et al. (2019). AMPK activation is involved in hypoglycemic and hypolipidemic activities of mogroside-rich extract from Siraitia grosvenorii (Swingle) fruits on high-fat diet/streptozotocin-induced diabetic mice. Food & Funct. 10, 151–162. doi:10.1039/C8FO01486H
Liu, J., Rong, L., Liu, C., and Rong, Y. H. (2011). Optimization of extraction conditions of active constituents from siraitia grosvenorii. AMR 291–294, 2523–2528. doi:10.4028/www.scientific.net/AMR.291-294.2523
Liu, J. N., Xue, C. F., and Si, C. Y. (2023b). Extraction of mogroside and its effect on the quality of soft europeanbread. J. Minnan Normal Univ. Sci. 36, 68–78.
Liu, M. S., Zhang, H., Li, X. B., and Qin, Y. Y. (2013b). The research of mogrosides on male mice’s genotoxicity. Chin. J. Birth Health & Hered. 21, 140–142.
Liu, T., Wang, X. H., Li, C., Zhang, Y., Wu, G. L., Li, D. C., et al. (2007). Study on the Antitussive,Expectorant and antispasmodic effects of SaponinV from momordica grosvenorii. Chin. Pharm. J., 1534–1536+1590.
Liu, Y., Wang, J., Dou, T., Zhou, L., Guan, X., Liu, G., et al. (2022). The liver metabolic features of Mogroside V compared to Siraitia grosvenorii fruit extract in allergic pneumonia mice. Mol. Immunol. 145, 80–87. doi:10.1016/j.molimm.2022.03.008
Liu, Y., Wang, J., Guan, X., Yu, D., Huangfu, M., Dou, T., et al. (2021b). Mogroside V reduce OVA-induced pulmonary inflammation based on lung and serum metabolomics. Phytomedicine 91, 153682. doi:10.1016/j.phymed.2021.153682
Liu, Y., Zhang, B., Liu, J., Qiao, C., Xue, N., Lv, H., et al. (2021c). Mogroside V alleviates lipopolysaccharide-induced neuroinflammation via inhibition of TLR4-MyD88 and activation of AKT/AMPK-Nrf2 signaling pathway. Evidence-Based Complementary Altern. Med. 2021, e5521519. doi:10.1155/2021/5521519
Lu, F., Sun, J., Jiang, X., Song, J., Yan, X., Teng, Q., et al. (2023a). Identification and isolation of α-glucosidase inhibitors from siraitia grosvenorii roots using bio-affinity ultrafiltration and comprehensive chromatography. Int. J. Mol. Sci. 24, 10178. doi:10.3390/ijms241210178
Lu, R., Hu, J., Liu, X., Yu, L., Hu, J., Jiang, H., et al. (2023b). Mogroside-rich extract from Siraitia grosvenorii fruits protects against heat stress-induced intestinal damage by ameliorating oxidative stress and inflammation in mice. Food Funct. 14, 1238–1247. doi:10.1039/d2fo02179j
Luo, H., Peng, C., Xu, X., Peng, Y., Shi, F., Li, Q., et al. (2022). The protective effects of mogroside V against neuronal damages by attenuating mitochondrial dysfunction via upregulating Sirtuin3. Mol. Neurobiol. 59, 2068–2084. doi:10.1007/s12035-021-02689-z
Luo, Z., Qiu, F., Zhang, K., Qin, X., Guo, Y., Shi, H., et al. (2016). In vitro AMPK activating effect and in vivo pharmacokinetics of mogroside V, a cucurbitane-type triterpenoid from Siraitia grosvenorii fruits. RSC Adv. 6, 7034–7041. doi:10.1039/C5RA23275A
McCord, J. M., and Edeas, M. A. (2005). SOD, oxidative stress and human pathologies: a brief history and a future vision. Biomed. Pharmacother. 59, 139–142. doi:10.1016/j.biopha.2005.03.005
McCullough, M., and Farah, C. (2008). The role of alcohol in oral carcinogenesis with particular reference to alcohol-containing mouthwashes. Aust. Dent. J. 53, 302–305. doi:10.1111/j.1834-7819.2008.00070.x
Mo, L. L., and Li, D. P. (2009). Antioxidant activity of flavonol glycosides of Siraitia grosvenorii flower. Mod. Food Sci. Technol. 25, 484–486.
National pharmacopoeia committee (2005). The pharmacopoeia of the People’s Republic of China (a) [S]. Beijing: chemical industry press.
Nie, J., Sui, L., Zhang, H., Zhang, H., Yan, K., Yang, X., et al. (2019). Mogroside V protects porcine oocytes from in vitro ageing by reducing oxidative stress through SIRT1 upregulation. Aging (Albany NY) 11, 8362–8373. doi:10.18632/aging.102324
Nie, J., Yan, K., Sui, L., Zhang, H., Zhang, H., Yang, X., et al. (2020). Mogroside V improves porcine oocyte in vitro maturation and subsequent embryonic development. Theriogenology 141, 35–40. doi:10.1016/j.theriogenology.2019.09.010
Niu, B., Ke, C.-Q., Li, B.-H., Li, Y., Yi, Y., Luo, Y., et al. (2017). Cucurbitane glucosides from the crude extract of siraitia grosvenorii with moderate effects on PGC-1α promoter activity. J. Nat. Prod. 80, 1428–1435. doi:10.1021/acs.jnatprod.6b01086
Pan, M.-H., Yang, J.-R., Tsai, M.-L., Sang, S., and Ho, C.-T. (2009). Anti-inflammatory effect of Momordica grosvenori Swingle extract through suppressed LPS-induced upregulation of iNOS and COX-2 in murine macrophages. J. Funct. Foods 1, 145–152. doi:10.1016/j.jff.2009.01.003
Pandey, A. K., and Chauhan, O. P. (2019). Monk fruit (Siraitia grosvenorii) - health aspects and food applications. 17, 191–198.
Pawar, R. S., Krynitsky, A. J., and Rader, J. I. (2013). Sweeteners from plants--with emphasis on Stevia rebaudiana (bertoni) and siraitia grosvenorii (swingle). Anal. Bioanal. Chem. 405, 4397–4407. doi:10.1007/s00216-012-6693-0
Prakash, I., and Chaturvedula, V. S. P. (2014). Additional new minor cucurbitane glycosides from Siraitia grosvenorii. Molecules 19, 3669–3680. doi:10.3390/molecules19033669
Qi, X. Y., Chen, W. J., Zhang, L. Q., Shan, X. F., and Song, Y. F. (2006). Study on the inhibitory effects of natural sweetner mogrosides on radical and lipid peroxidation. Sci. Agric. Sin. 39, 382–388.
Qi, X.-Y., Chen, W.-J., Zhang, L.-Q., and Xie, B.-J. (2008). Mogrosides extract from Siraitia grosvenori scavenges free radicals in vitro and lowers oxidative stress, serum glucose, and lipid levels in alloxan-induced diabetic mice. Nutr. Res. 28, 278–284. doi:10.1016/j.nutres.2008.02.008
Qin, L. Y., Liu, J., Liu, J., and He, Y. (2023). A Luohanguo “save the market war” quietly started. Guilin Dly. 007.
Qin, Q. F., Lan, X. Q., Wen, Q. W., Liu, T., Lu, D., Wei, H. Y., et al. (2014). Determination of mogroside V in fufang luohanguo Zhike granule by HPLC-ELSD. 31, 850–852. doi:10.13748/j.cnki.issn1007-7693.2014.07.020
Scuderi, P. E. (2010). Postoperative sore throat: more answers than questions. Anesth. Analgesia 111, 831–832. doi:10.1213/ANE.0b013e3181ee85c7
Sheka, A. C., Adeyi, O., Thompson, J., Hameed, B., Crawford, P. A., and Ikramuddin, S. (2020). Nonalcoholic steatohepatitis: a review. JAMA 323, 1175–1183. doi:10.1001/jama.2020.2298
Shi, D., Zheng, M., Wang, Y., Liu, C., and Chen, S. (2014). Protective effects and mechanisms of mogroside V on LPS-induced acute lung injury in mice. Pharm. Biol. 52, 729–734. doi:10.3109/13880209.2013.867451
Shivani, null, Thakur, B. K., Mallikarjun, C. P., Mahajan, M., Kapoor, P., Malhotra, J., et al. (2021). Introduction, adaptation and characterization of monk fruit (Siraitia grosvenorii): a non-caloric new natural sweetener. Sci. Rep. 11, 6205. doi:10.1038/s41598-021-85689-2
Si, J. Y., Chen, D. H., Chang, Q., and Shen, L. G. (1994). Isolation and structure determination of flavonol glycosides from the fresh fruits of siraitia grosvenorii. Acta Pharm. Sin. B 29, 158–160.
Si, J.-Y., Chen, D.-H., and Tu, G.-Z. (2005). Siraitic acid F, a new nor-cucurbitacin with novel skeleton, from the roots of Siraitia grosvenorii. J. Asian Nat. Prod. Res. 7, 37–41. doi:10.1080/10286020310001609010
Song, F., Qi, X., Chen, W., Jia, W., Yao, P., Nussler, A. K., et al. (2007). Effect of Momordica grosvenori on oxidative stress pathways in renal mitochondria of normal and alloxan-induced diabetic mice. Involvement of heme oxygenase-1. Eur. J. Nutr. 46, 61–69. doi:10.1007/s00394-006-0632-9
Song, J., Qian, B., Pan, C., Lv, F., Wang, H., Gao, Y., et al. (2019). Protective activity of mogroside V against ovalbumin-induced experimental allergic asthma in Kunming mice. J. Food Biochem. 43, e12973. doi:10.1111/jfbc.12973
Sun, L. P. (2022). Application of response surface methodology in the development of momordica grosvenori stomach strengthening dietary beverage. Farm Prod. Process., 35–40. doi:10.16693/j.cnki.1671-9646(X).2022.04.008
Sung, Y.-Y., Kim, S.-H., Yuk, H. J., Yang, W.-K., Lee, Y. M., Son, E., et al. (2019). Siraitia grosvenorii residual extract attenuates ovalbumin-induced lung inflammation by down-regulating IL-4, IL-5, IL-13, IL-17, and MUC5AC expression in mice. Phytomedicine 61, 152835. doi:10.1016/j.phymed.2019.152835
Sung, Y.-Y., Yuk, H.-J., Yang, W.-K., Kim, S.-H., and Kim, D.-S. (2020). Siraitia grosvenorii residual extract attenuates atopic dermatitis by regulating immune dysfunction and skin barrier abnormality. Nutrients 12, 3638. doi:10.3390/nu12123638
Suzuki, Y. A., Murata, Y., Inui, H., Sugiura, M., and Nakano, Y. (2005). Triterpene glycosides of Siraitia grosvenori inhibit rat intestinal maltase and suppress the rise in blood glucose level after a single oral administration of maltose in rats. J. Agric. Food Chem. 53, 2941–2946. doi:10.1021/jf0478105
Suzuki, Y. A., Tomoda, M., Murata, Y., Inui, H., Sugiura, M., and Nakano, Y. (2007). Antidiabetic effect of long-term supplementation with Siraitia grosvenori on the spontaneously diabetic Goto–Kakizaki rat. Br. J. Nutr. 97, 770–775. doi:10.1017/S0007114507381300
Suzuki-Kemuriyama, N., Abe, A., Nakane, S., Uno, K., Ogawa, S., Watanabe, A., et al. (2021). Extract of Siraitia grosvenorii (Luo Han Guo) protects against hepatic fibrosis in mice on a choline-deficient, methionine-lowered, L-amino acid-defined, high-fat diet without trans fatty acids. Fundam. Toxicol. Sci. 8, 135–145. doi:10.2131/fts.8.135
Takasaki, M., Konoshima, T., Murata, Y., Sugiura, M., Nishino, H., Tokuda, H., et al. (2003). Anticarcinogenic activity of natural sweeteners, cucurbitane glycosides, from Momordica grosvenori. Cancer Lett. 198, 37–42. doi:10.1016/S0304-3835(03)00285-4
Takemoto, T., Arihara, S., Nakajima, T., and Okuhira, M. (1983). Studies on the constituents of fructus Momordicae. II. Structure of sapogenin. Yakugaku Zasshi 103, 1155–1166. doi:10.1248/yakushi1947.103.11_1155
Tang, Q. (2021). Mogroside on Parkinson’s disease model mice nerve protective effect and mechanism of research. Minzu Univ. China.
Tang, S. F. (2023a). Analysis on the status quo of industry development and green quality cultivation technology of Siraitia grosvenorii. Seed Sci. Technol. 41, 38–40. doi:10.19904/j.cnki.cn14-1160/s.2023.15.012
Tang, W. Y. (2023b). Research on high quality and high yield cultivation technology of S. grosvenorii. Hebei Agric. Mach. 03, 97–99.
Tao, L., Yang, J., Cao, F., Xie, H., Zhang, M., Gong, Y., et al. (2017). Mogroside IIIE, a novel anti-fibrotic compound, reduces pulmonary fibrosis through toll-like receptor 4 pathways. J. Pharmacol. Exp. Ther. 361, 268–279. doi:10.1124/jpet.116.239137
Thakur, K., Partap, M., Kumar, P., Sharma, R., and Warghat, A. R. (2023). Understandings of bioactive composition, molecular regulation, and biotechnological interventions in the development and usage of specialized metabolites as health-promoting substances in Siraitia grosvenorii (Swingle) C. Jeffrey. J. Food Compos. Analysis 116, 105070. doi:10.1016/j.jfca.2022.105070
Ukiya, M., Akihisa, T., Tokuda, H., Toriumi, M., Mukainaka, T., Banno, N., et al. (2002). Inhibitory effects of cucurbitane glycosides and other triterpenoids from the fruit of Momordica grosvenori on Epstein−Barr virus early antigen induced by tumor promoter 12- O -Tetradecanoylphorbol-13-acetate. J. Agric. Food Chem. 50, 6710–6715. doi:10.1021/jf0206320
Uno, K., Miyajima, K., Ogawa, S., Suzuki-Kemuriyama, N., and Nakae, D. (2023). Effects of Siraitia grosvenorii extract on nonalcoholic steatohepatitis-like lesions in Sprague Dawley rats fed a choline-deficient, methionine-lowered, l-amino acid-defined diet. J. Toxicol. Pathol. 36, 1–10. doi:10.1293/tox.2022-0043
Wang, H., Ma, G., Wang, H., Li, L., Dong, A., Liu, H., et al. (2022). Novel triterpenoid alkaloids with their potential cytotoxic activity from the roots of siraitia grosvenorii. Front. Chem. 10, 885487. doi:10.3389/fchem.2022.885487
Wang, H. J. (2023). Research on the antitumor active constituents from the Roots of Siraitia grosvenorii. Peking. Union Med. Coll. doi:10.27648/d.cnki.gzxhu.2022.000283
Wang, L., Yang, Z., Lu, F., Liu, J., Song, Y., and Li, D. (2014). Cucurbitane glycosides derived from mogroside IIE: structure-taste relationships, antioxidant activity, and acute toxicity. Molecules 19, 12676–12689. doi:10.3390/molecules190812676
Wang, M., Xing, S., Luu, T., Fan, M., and Li, X. (2015). The gastrointestinal tract metabolism and pharmacological activities of grosvenorine, a major and characteristic flavonoid in the fruits of Siraitia grosvenorii. Chem. Biodivers. 12, 1652–1664. doi:10.1002/cbdv.201400397
Wang, X., Lu, W., Chen, J., Gong, M., Li, Y., Lu, D., et al. (1996). Studies on the chemical contituents of root of Luohanguo Siraitia grosvenori. Zhong cao yao = Chinese traditional and herbal drugs.
Wang, Y., Sun, W., Yan, S., Meng, Z., Jia, M., Tian, S., et al. (2023). A new strategy to alleviate the obesity induced by endocrine disruptors-A unique lysine metabolic pathway of nanoselenium Siraitia grosvenorii to repair gut microbiota and resist obesity. Food Chem. Toxicol. 175, 113737. doi:10.1016/j.fct.2023.113737
Weerawatanakorn, M., Yang, J.-R., Tsai, M.-L., Lai, C.-S., Ho, C.-T., and Pan, M.-H. (2014). Inhibitory effects of Momordica grosvenori Swingle extracts on 12-O-tetradecanoylphorbol 13-acetate-induced skin inflammation and tumor promotion in mouse skin. Food Funct. 5, 257–264. doi:10.1039/C3FO60332F
Wei, B. Q., Gao, X. Y., Liu, Y. X., Xu, H. M., and Wang, Y. C. (2023). Research progress on function and application of mogroside. Sci. Technol. Food Industry 44, 434–441. doi:10.13386/j.issn1002-0306.2022070323
Wu, J., Jian, Y., Wang, H., Huang, H., Gong, L., Liu, G., et al. (2022a). A review of the phytochemistry and pharmacology of the fruit of siraitia grosvenorii (swingle): a traditional Chinese medicinal food. Molecules 27, 6618. doi:10.3390/molecules27196618
Wu, J. J., Huang, H. X., Li, W. C., Wang, B., Gong, L. M., Sheng, W. B., et al. (2023). Study on chemical constituents and anti-inflammatory activity of the pomace of Siraitia grosvenorii. Nat. Prod. Res. Dev. 35, 1124–1134. doi:10.16333/1.1001-6880.2023.7.003
Wu, Y., Xu, H. N., Xue, Y., Liu, Q. B., Liu, H., Li, Z. H., et al. (2022b). Fabrication of monodisperse gold-copper nanocubes and AuCu-cuprous sulfide heterodimers by a step-wise polyol reduction. Hubei Agric. Sci. 61, 136–145. doi:10.1016/j.jcis.2022.06.120
Xia, X., Zhong, Z. G., Lin, C. Y., and Zhu, X. Y. (2012). Anti-fatigue and hypoxia tolerance effect of mogrosides in mice. Chin. J. Exp. Traditional Med. Formulae 18, 198–201. doi:10.13422/j.cnki.syfjx.2012.17.044
Xia, Y. (2006). Supercritical fluid extraction of mogrosides from Siraitia grosvenorii. McGill University. Available at: https://escholarship.mcgill.ca/concern/theses/n583z025x.
Xiao, G., Chen, Z., and Li, W. N. (2014). Anti-aging effect of mogroside. Chin. J. gerontology 34, 4263–4265.
Xiao, J., Huang, K., Lin, H., Xia, Z., Zhang, J., Li, D., et al. (2020). Mogroside IIE inhibits digestive enzymes via suppression of interleukin 9/interleukin 9 receptor signalling in acute pancreatitis. Front. Pharmacol. 11, 859. doi:10.3389/fphar.2020.00859
Xiao, Z. Z., Teng, S. H., Zhou, J. H., Feng, X. Q., Huang, L. L., Jin, F., et al. (2023). Study on application of Siraitia grosvenorii residue added into diets in Guangxi Ma Chicken. China feed., 158–161. doi:10.15906/j.cnki.cn11-2975/s.2022080019-06
Xie, Y. F. (2018). Effects of Siraitia grosvenorii leaf flavonoids on myocardial mitochondria in rats undergoing exhaustive exercise. Sci. Technol. Inf. 16, 227–228. doi:10.16661/j.cnki.1672-3791.2018.12.227
Xu, J. J., Huang, X. C., Wei, G. C., Cheng, Z. J., Han, W., Yang, D. H., et al. (2022a). Effects of mogroside on growth performance, blood physiological and biochemical indexes, slaughter performance and meat quality of Hu sheep. Mod. Anim. Husb., 29–33.
Xu, W. K., and Meng, L. S. (1986). Determination of protein content of Siraitia grosvenorii. Guihaia, 295–296.
Xu, Y., Liu, S., Bian, L., Li, Z., Luo, C., Chen, Y., et al. (2022b). Engineering of a UDP-glycosyltransferase for the efficient whole-cell biosynthesis of siamenoside I in Escherichia coli. J. Agric. Food Chem. 70, 1601–1609. doi:10.1021/acs.jafc.1c07699
Yan, Z. (2009). Evaluations of antioxidative ability and lipid metabolism mediation as well as caries prevention for Luo Han Kuo. Shanghai Jiaot. Univ.
Yanan, S., Bohan, L., Shuaifeng, S., Wendan, T., Ma, Z., and Wei, L. (2023). Inhibition of Mogroside IIIE on isoproterenol-induced myocardial fibrosis through the TLR4/MyD88/NF-κB signaling pathway. Iran. J. Basic Med. Sci. 26, 114–120. doi:10.22038/IJBMS.2022.67908.14848
Yang, X. W., Zhang, J. Y., and Qian, Z. M. (2008). New natural saponins form fruits of Momordica grosvenorii. Chin. Tradit. Herb. Drugs 39, 810–813.
Yang, X. W., Zhang, J. Y., and Qian, Z. M. (2005). Grosmomoside I, a new cucurbitane triterpenoid glycoside from fruits of momordica grosvenori. Chin. Tradit. Herb. Drugs, 9–14.
Yao, J. W., Tang, H., Shen, W. H., Li, Y. X., Ding, X. S., and Li, X. J. (2007). The observation on impacts of the different dosage of Luo Han Guo on physiological function in mice by train of increasing intensity. Liaoning Sport Sci. Technol.
Yeung, A. W. K. (2023). Bibliometric analysis on the literature of monk fruit extract and mogrosides as sweeteners. Front. Nutr. 10, 1253255. doi:10.3389/fnut.2023.1253255
Zhang, H., Li, X. H., Huang, D. R., Zeng, Y., Yang, J., Wang, S. C., et al. (2011). Effects of grosvenor momordica fruit on bone marrow cell micronucleus andsperm morphology of male mice. Chin. J. Tissue Eng. Res. 15, 5249–5252+2.
Zhang, H., and Li, X. Y. (2011). Research advance of pharmacological effects and toxicity of siraitia grosvenorii (swingle) C. Jeffrey. Chin. Agric. Sci. Bull. 27, 430–433+2.
Zhang, Q. L., Huang, J., Wu, Z. H., and Pi, F. J. (2017). Research overview of pharmacology and application development of Siraitia grosvenorii. J. Pharm. Res. 36, 164–165. doi:10.13506/j.cnki.jpr.2017.03.011
Zhang, S. M., Shi, X. X., Wei, X. L., Zhu, Y. M., and Su, H. J. (2018). Study on extraction technology of siraitia grosvenorii by reflux method andthe application of extract in cigarette. Flavour Fragr. Cosmet., 36–39.
Zhang, Y., Peng, Y., Zhao, L., Zhou, G., and Li, X. (2021). Regulating the gut microbiota and SCFAs in the faeces of T2DM rats should be one of antidiabetic mechanisms of mogrosides in the fruits of Siraitia grosvenorii. J. Ethnopharmacol. 274, 114033. doi:10.1016/j.jep.2021.114033
Zhang, Y. L., Zhou, G. S., Peng, Y., Wang, M. Y., and Li, X. B. (2020a). Anti-hyperglycemic and anti-hyperlipidemic effects of a special fraction of Luohanguo extract on obese T2DM rats. J. Ethnopharmacol. 30, 112273. doi:10.1016/j.jep.2019.112273
Zhang, Y. Z., Xu, G. P., Zhou, L. W., Teng, Q. M., He, W., Zhang, Z. F., et al. (2020b). Analysis the nutrient compositions of Siraitia grosvenorii in the hilly region at north Guangxi. Hubei Agric. Sci. 59, 151–154.
Zhang, Z. T., and Zhang, T. T. (2011). Medicine for improving male sperm quantity and vitality, preparation and preparation method: CN Patent 07737056 A.
Zhao, X. L. (2015). Research progress in physiologically active ingredients of siraitia grosvenorii. Packag. food Mach. 33, 54–57.
Zhao, Z. Y. (2023). Research on the marketing strategy optimization of monk fruit extract products of L company. Guangxi Norm. Univ.
Zheng, Y., Huang, W., Yoo, J.-G., Ebersole, J. L., and Huang, C. B. (2011). Antibacterial compounds from Siraitia grosvenorii leaves. Nat. Prod. Res. 25, 890–897. doi:10.1080/14786419.2010.490212
Zhou, C., Zhang, J., Wu, Y., Cheng, H., Pang, Q., Xiao, Y., et al. (2022a). Metabolomic analysis on the mechanism of nanoselenium biofortification improving the siraitia grosvenorii nutritional and health value. Foods 11, 3019. doi:10.3390/foods11193019
Zhou, G., Zhang, Y., Li, Y., Wang, M., and Li, X. (2018a). The metabolism of a natural product mogroside V, in healthy and type 2 diabetic rats. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 1079, 25–33. doi:10.1016/j.jchromb.2018.02.002
Zhou, X., Reheman, A., Kang, Z., Long, A., and Wang, T. (2022b). Traditional Chinese medicine compounds containing Lonicera japonica, Chrysanthemum morifolium, and siraitia grosvenorii inhibits the growth of Streptococcus mutans. Evid. Based Complement. Altern. Med. 2022, 5802343. doi:10.1155/2022/5802343
Zhou, Z., Huang, G., and Li, G. (2018b). Microwave drying of Siraitia grosvenorii and its effect on the Vitamin C content. China: Atlantis Press, 1725–1729. doi:10.2991/iceesd-18.2018.311
Zhu, Y.-M., Pan, L.-C., Zhang, L.-J., Yin, Y., Zhu, Z.-Y., Sun, H.-Q., et al. (2020). Chemical structure and antioxidant activity of a polysaccharide from Siraitia grosvenorii. Int. J. Biol. Macromol. 165, 1900–1910. doi:10.1016/j.ijbiomac.2020.10.127

Keywords: Siraitia grosvenorii, mogrosides, chemical compounds, pharmacological effects, toxicological effects, resource development status
Citation: Huang H, Peng Z, Zhan S, Li W, Liu D, Huang S, Zhu Y and Wang W (2024) A comprehensive review of Siraitia grosvenorii (Swingle) C. Jeffrey: chemical composition, pharmacology, toxicology, status of resources development, and applications. Front. Pharmacol. 15:1388747. doi: 10.3389/fphar.2024.1388747
Received: 20 February 2024; Accepted: 21 March 2024;
Published: 04 April 2024.
Edited by:
Hua Li, Air Force Medical University, ChinaReviewed by:
Puyang Gong, Southwest Minzu University, ChinaCopyright © 2024 Huang, Peng, Zhan, Li, Liu, Huang, Zhu and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yizhun Zhu, eXp6aHVAbXVzdGVkdS5tbw==, emh1eXpAZnVkYW4uZWR1LmNu; Wei Wang, d2FuZ3dlaTQwMkBob3RtYWlsLmNvbQ==
†These authors have contributed equally to this work and share first authorship
‡These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.