
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Pharmacol. , 02 May 2024
Sec. Pharmacogenetics and Pharmacogenomics
Volume 15 - 2024 | https://doi.org/10.3389/fphar.2024.1373007
 Gabriela Echeverría-Garcés1,2
Gabriela Echeverría-Garcés1,2 María José Ramos-Medina3
María José Ramos-Medina3 Rodrigo Vargas2,4
Rodrigo Vargas2,4 Alejandro Cabrera-Andrade5,6Adriana Altamirano-Colina7María Paula Freire7Juliana Montalvo-Guerrero7
Alejandro Cabrera-Andrade5,6Adriana Altamirano-Colina7María Paula Freire7Juliana Montalvo-Guerrero7 Sebastián Rivera-Orellana8Paulina Echeverría-Espinoza7
Sebastián Rivera-Orellana8Paulina Echeverría-Espinoza7 Luis A. Quiñones2,9,10
Luis A. Quiñones2,9,10 Andrés López-Cortés7*
Andrés López-Cortés7*Introduction: Gastric cancer is one of the most prevalent types of cancer worldwide. The World Health Organization (WHO), the International Agency for Research on Cancer (IARC), and the Global Cancer Statistics (GLOBOCAN) reported an age standardized global incidence rate of 9.2 per 100,000 individuals for gastric cancer in 2022, with a mortality rate of 6.1. Despite considerable progress in precision oncology through the efforts of international consortia, understanding the genomic features and their influence on the effectiveness of anti-cancer treatments across diverse ethnic groups remains essential.
Methods: Our study aimed to address this need by conducting integrated in silico analyses to identify actionable genomic alterations in gastric cancer driver genes, assess their impact using deleteriousness scores, and determine allele frequencies across nine global populations: European Finnish, European non-Finnish, Latino, East Asian, South Asian, African, Middle Eastern, Ashkenazi Jewish, and Amish. Furthermore, our goal was to prioritize targeted therapeutic strategies based on pharmacogenomics clinical guidelines, in silico drug prescriptions, and clinical trial data.
Results: Our comprehensive analysis examined 275,634 variants within 60 gastric cancer driver genes from 730,947 exome sequences and 76,215 whole-genome sequences from unrelated individuals, identifying 13,542 annotated and predicted oncogenic variants. We prioritized the most prevalent and deleterious oncogenic variants for subsequent pharmacogenomics testing. Additionally, we discovered actionable genomic alterations in the ARID1A, ATM, BCOR, ERBB2, ERBB3, CDKN2A, KIT, PIK3CA, PTEN, NTRK3, TP53, and CDKN2A genes that could enhance the efficacy of anti-cancer therapies, as suggested by in silico drug prescription analyses, reviews of current pharmacogenomics clinical guidelines, and evaluations of phase III and IV clinical trials targeting gastric cancer driver proteins.
Discussion: These findings underline the urgency of consolidating efforts to devise effective prevention measures, invest in genomic profiling for underrepresented populations, and ensure the inclusion of ethnic minorities in future clinical trials and cancer research in developed countries.
Gastric cancer is a heterogeneous disease originating in the mucus-producing cells lining the stomach interior (Ajani et al., 2017). It involves a wide range of biological components, including hormonal imbalances, ethnic backgrounds, environmental factors, epigenetics, genetic mutations, alterations in protein expression, and modifications in signaling pathways (Hanahan, 2022). According to the World Health Organization (WHO), the International Agency for Research on Cancer (IARC), and the Global Cancer Statistics (GLOBOCAN), the global age-standardized incidence rate for gastric cancer in 2022 was 9.2 per 100,000 individuals, with a mortality rate of 6.1 (Sung et al., 2021).
Since the Human Genome Project began in 1990 until the completion of the human genome sequence in 2022, genomics has played a crucial role in both foundational and translational research (Green et al., 2020; Nurk et al., 2022). Advances in high-throughput technologies, particularly in large-scale DNA sequencing, have improved our understanding of the molecular mechanisms underlying gastric cancer. This progress has been instrumental in identifying cancer driver genes (Kandoth et al., 2013; Lawrence et al., 2014), germline mutations (Lu et al., 2015), cancer driver variants in both coding and non-coding regions (Sjöblom et al., 2006; Tamborero et al., 2013; Porta-Pardo et al., 2017; Rheinbay et al., 2020), druggable proteins (Rubio-Perez et al., 2015), drug resistance mechanisms (Vasan et al., 2019), pharmacogenomics (PGx) clinical guidelines (Quinones et al., 2014; López-Cortés et al., 2020c; 2017; Varela et al., 2021), and the development of artificial intelligence algorithms (López-Cortés et al., 2020a; Jumper et al., 2021).
Genetic variants are critical in driving oncogenesis in gastric cancer, leading to the activation of oncogenes or the inactivation of tumor suppressor genes through mechanisms such as gene regulation disruption, creation of fusion genes with oncogenic properties, and alterations in the genomic architecture that globally affect gene expression (Tan and Yeoh, 2015; Yao et al., 2015).
In recent years, it has become evident that patients with the same cancer type do not uniformly respond to standard treatments (Raguz and Yagüe, 2008; Mansoori et al., 2017). Precision oncology addresses this variability by providing personalized treatment options, including appropriate medications and dosages, considering individual patient needs, their ethnicity, and treatment timing (Garraway et al., 2013; Quinones et al., 2014; Pérez-Villa et al., 2023). Therefore, identifying actionable genomic alterations is a primary goal of cancer research, particularly in the driver gene landscape of gastric cancer, to devise effective therapeutic strategies and PGx clinical guidelines (ICGC/TCGA Pan-Cancer Analysis of Whole Genomes Consortium, 2020).
Tailoring drug development to individual multi-omics profiles can enhance drug efficacy and reduce adverse reactions (López-Cortés et al., 2020c; Pérez-Villa et al., 2023; Ramos-Medina et al., 2024). Despite significant advances in precision oncology through international consortium efforts like The Cancer Genome Atlas (TCGA) and Therapeutically Applicable Research to Generate Effective Treatments (TARGET) (Spratt et al., 2016; Guerrero et al., 2018), the underrepresentation of diverse ethnic backgrounds in many cancer research studies has led to a significant bias toward Caucasians in cancer genomic databases (Guerrero et al., 2018). This bias poses a considerable obstacle to the advancement of PGx and precision oncology, especially in developing regions (Salas-Hernández et al., 2023). To bridge this gap, the primary objective of our study was to conduct an integrated in silico analysis to identify actionable genomic alterations in gastric cancer, evaluate their impact through deleteriousness scores and allele frequencies across nine global populations, and prioritize targeted therapeutic strategies. By integrating these findings with PGx clinical guidelines (Barbarino et al., 2018), in silico drug prescriptions (Tamborero et al., 2018), and clinical trial data (Ochoa et al., 2021), we aim to broaden the scope of precision oncology, ensuring it benefits a more diverse population by encouraging the development of effective and personalized therapeutic interventions.
We conducted an integrative in silico analysis to understand the genomic landscape of gastric cancer across diverse ethnic populations, examine current pharmacogenomics guidelines in clinical practice, analyze in silico drug prescriptions of therapeutic actionable genomic alterations, and evaluate the drugs involved in early-stage and late-stage clinical trials.
The Global Cancer Observatory (https://gco.iarc.fr/) provides a complete assessment of the global burden of cancer. By using the latest version of GLOBOCAN 2022, we identified and ranked the countries worldwide with the highest estimated age-standardized incidence and mortality rates for gastric cancer (Sung et al., 2021).
The intOGen framework (https://www.intogen.org) is a bioinformatics resource that can identify cancer genes and determine how they work in different types of tumors (Martínez-Jiménez et al., 2020). This tool uses seven methods to identify cancer driver genes based on point mutations, namely, dNdScv (Martincorena et al., 2017), CBaSE (Weghorn and Sunyaev, 2017), MutPanning (Dietlein et al., 2020), OncodriveCLUSTL (Arnedo-Pac et al., 2019), HotMAPS (Tokheim et al., 2016), smRegions (Martínez-Jiménez et al., 2020), and OncodriveFML (Mularoni et al., 2016). Therefore, we retrieved 60 gastric cancer driver genes and identified their involvement as oncogenes (Sondka et al., 2018), tumor suppressor genes (Sondka et al., 2018), kinase genes (Manning et al., 2002; Eid et al., 2017), DNA-repair genes (Wood et al., 2001; Lange et al., 2011), RNA-binding proteins (Hentze et al., 2018), cell cycle genes (Bar-Joseph et al., 2008), metastatic genes (Zheng et al., 2018), and cancer immunotherapy genes (Patel et al., 2017).
The process of identifying the oncogenic variome involved two main steps. First, we extracted 275,634 single-nucleotide and insertion/deletion variants from the Genome Aggregation Database (gnomAD v4.0) (https://gnomad.broadinstitute.org/), which belonged to 60 gastric cancer driver genes (Collins et al., 2020; Karczewski et al., 2020; Nurk et al., 2022). We used the complete sequence of a human genome (GRCh38/hg38) as the reference genome (Collins et al., 2020; Karczewski et al., 2020; Nurk et al., 2022). In the second step, we used two methods, OncodriveMUT and boostDM, integrated into the Cancer Genome Interpreter (CGI) platform (https://www.cancergenomeinterpreter.org) to evaluate the tumorigenic potential of the 275,634 extracted genomic variants (Tamborero et al., 2018; Muiños et al., 2021). OncodriveMUT is a rule-based approach that considers various genomic features such as regions depleted by germline variants, gene mechanism of action, gene signals of positive selection, and clusters of somatic mutations, whereas boostDM is a machine learning-based methodology that uses in silico saturation mutagenesis of cancer genes to assess the oncogenic potential of mutations in human tissues. Both methods allowed us to categorize driver variants into annotated (known), predicted, and passenger mutations based on the Catalog of Validated Oncogenic Mutations (Tamborero et al., 2018; Muiños et al., 2021).
Combined Annotation-Dependent Depletion (CADD) version 1.7 (https://cadd.gs.washington.edu/) serves as a machine learning-based tool that scores and classifies genetic variants to support the assessment of clinical observations. As one of the pioneering methods for the genome-wide prioritization of variants across various molecular functions, CADD was developed using more than 60 genomic features. It is capable of assessing the impact of both single-nucleotide variants and insertions/deletions (Kircher et al., 2014; Schubach et al., 2024). This tool uses a method that contrasts natural selection with simulated mutations, enabling the integration of multiple annotations into a single metric. Designed for compatibility with the GRCh38/hg38 human reference genome, CADD facilitates comprehensive genetic analysis (Rentzsch et al., 2019). In this context, we used CADD to assess the deleteriousness of annotated and predicted cancer-causing gene mutations, specifically in gastric cancer driver genes. The deleteriousness of the oncogenic variome was categorized as very high (30–50), high (25–30), medium (15–25), low (10–15), and very low (0–10).
The process of enrichment analysis provides scientists with a carefully selected explanation of sets of genes or proteins obtained from large-scale experiments in omics (López-Cortés et al., 2020b; 2022; 2021). In this study, a functional enrichment analysis was conducted on genes/proteins that drive gastric cancer and carry annotated or predicted oncogenic variants. The analysis was performed using g:Profiler version e101_eg48_p14_baf17f0 (https://biit.cs.ut.ee/gprofiler/gost) (Raudvere et al., 2019) to identify significant annotations (Benjamini–Hochberg false discovery rate (FDR) q < 0.001) related to biological processes in Gene Ontology (GO) (The Gene Ontology Consortium, 2021), signaling pathways in the Kyoto Encyclopedia of Genes and Genomes (KEGG) (Kanehisa and Goto, 2000), signaling pathways in Reactome (Fabregat et al., 2016), pathways in WikiPathways (Slenter et al., 2018), and Human Phenotype Ontology (HPO) (Köhler et al., 2021). Finally, signaling pathways that were significant and relevant to gastric cancer were manually curated and presented using a Manhattan plot.
gnomAD is a collection of genomic sequencing data from various projects conducted globally (Karczewski et al., 2020). The v4.0 dataset focuses on GRCh38/hg38 and includes 730,947 exome sequences and 76,215 whole-genome sequences from unrelated individuals with different ancestral backgrounds. This study evaluated the allele frequencies of the annotated and predicted gastric cancer oncogenic variome in 9 populations, namely, African (n = 37,545), Amish (n = 456), Latino (n = 30,019), Ashkenazi Jewish (n = 14,804), East Asian (n = 22,448), European Finnish (n = 32,026), European non-Finnish (n = 590,031), Middle Eastern (n = 3,031), and South Asian (n = 45,546) (Collins et al., 2020; Karczewski et al., 2020).
The Pharmacogenomics Knowledge Base (PharmGKB) (https://www.pharmgkb.org/) is a database focused on pharmacogenomics. It contains important information about the relationship between genes and drugs, as well as specific guidelines for applying pharmacogenomics in clinical practice (Whirl-Carrillo et al., 2012; Barbarino et al., 2018). The database collects information from various sources, including the National Comprehensive Cancer Network (NCCN), the European Society for Medical Oncology (ESMO), the Clinical Pharmacogenetics Implementation Consortium (CPIC) (Relling and Klein, 2011; Relling et al., 2020), the Canadian Pharmacogenomics Network for Drug Safety (Ross et al., 2010), and the Royal Dutch Association for the Advancement of Pharmacy (Swen et al., 2011). In this context, we retrieved clinical annotations, gene–drug pairs, and genomic variants associated with gastric cancer pharmacogenomics guidelines.
Another CGI approach is in silico drug prescription. This involves identifying therapeutic actionable genomic alterations for drug response in tumors and organizing them based on their clinical importance level (Muiños et al., 2021). The method relies on two databases, the Cancer Biomarker database and the Cancer Bioactivities database, to explore the connection between the oncogenic variome and drug response (Dienstmann et al., 2015; Tamborero et al., 2018). Our study aimed to analyze the druggability of gastric cancer driver proteins carrying annotated and predicted oncogenic variants. This in silico analysis helped us identify the most appropriate treatment strategies based on precision oncology.
The Open Targets Platform, last updated in March 2024 (https://www.targetvalidation.org), provides access to and visualization of potential therapeutic targets and drugs involved in gastric cancer clinical trials (Carvalho-Silva et al., 2019; Ochoa et al., 2021). Additionally, the Drug Repurposing Hub (https://www.broadinstitute.org/drug-repurposing-hub) serves as a bioinformatics resource to identify the mechanisms of action of drugs approved by the US Food and Drug Administration (FDA) (Corsello et al., 2017). To further elucidate the involvement of drugs in clinical trials, we used a Sankey plot, enabling a clearer distinction between drugs associated with early-stage (phases I and II) and late-stage (phases III and IV) clinical trials.
According to the WHO, IARC, and GLOBOCAN 2022, the top 10 countries worldwide with the highest estimated age-standardized incidence rates of gastric cancer per 100,000 inhabitants were Mongolia (35.5), Japan (27.6), South Korea (27.0), Tajikistan (19.4), Iran (19.4), Azerbaijan (16.6), Kyrgyzstan (16.5), Bhutan (15.9), and Belarus (15.3) (Figure 1A; Supplementary Table S1); meanwhile, the top 10 countries worldwide with the highest estimated age-standardized mortality rate were Mongolia (31.5), Tajikistan (16.7), Iran (15.4), Bhutan (14.2), Kyrgyzstan (14.0), Mali (13.5), Azerbaijan (12.8), North Korea (11.4), Belarus (11.3), and Sao Tome and Principe (11.1) (Figure 1B; Supplementary Table S2) (Sung et al., 2021).
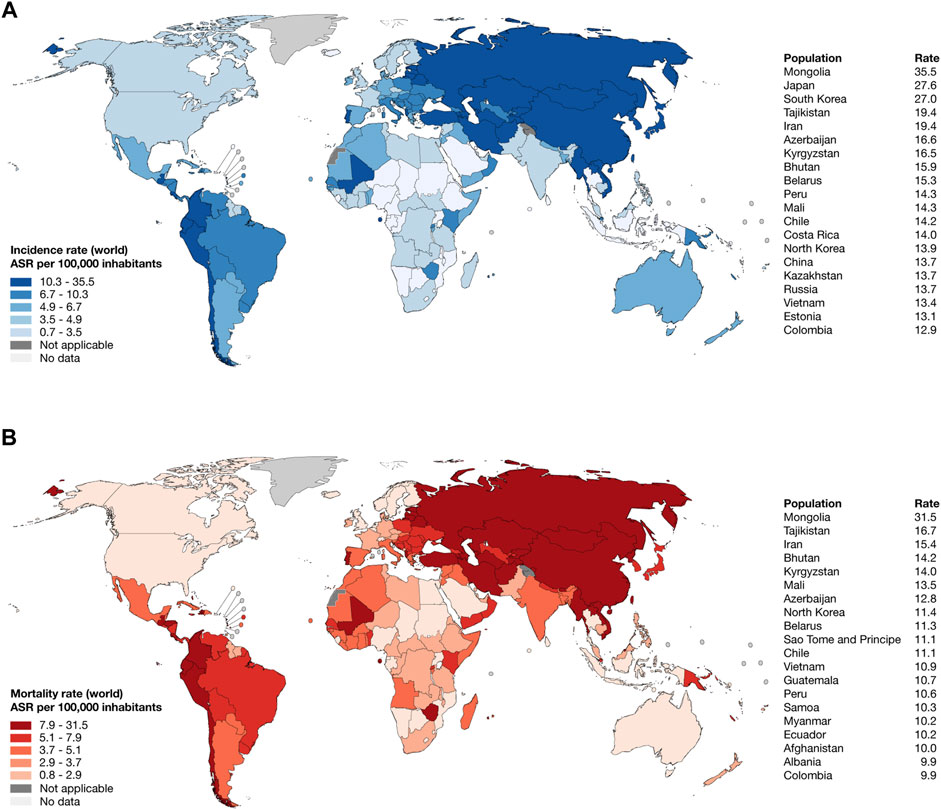
Figure 1. Epidemiology of stomach cancer in 2022. (A) Heatmap and ranking of the estimated age-standardized incidence rate of stomach cancer per 100,000 inhabitants worldwide. (B) Heatmap and ranking of the estimated age-standardized mortality rate of stomach cancer per 100,000 inhabitants worldwide. ASR: age-standardized rate.
We retrieved 60 gastric cancer driver genes from the intOGen framework (Martínez-Jiménez et al., 2020), of which 33 (55%) were tumor suppressor genes (Sondka et al., 2018), 33 (55%) were metastatic genes (Zheng et al., 2018), 15 (25%) were oncogenes (Sondka et al., 2018), 11 (18%) were kinase genes (Manning et al., 2002; Eid et al., 2017), 5 (8%) encoded RNA-binding proteins (Hentze et al., 2018), 4 (7%) were cancer immunotherapy genes (Patel et al., 2017), 2 (3%) were DNA-repair genes (Wood et al., 2001; Lange et al., 2011), and 2 (3%) were cell cycle genes (Bar-Joseph et al., 2008) (Supplementary Table S3).
Figure 2A presents the results of the OncodriveMUT and boostDM analyses used to identify the oncogenic variome of 60 gastric cancer driver genes by using the GRCh38/hg38 human reference genome. After analyzing 275,634 variants, we identified 13,542 oncogenic variants, of which 243 (2%) were annotated and 13,299 (98%) were predicted. The consequence type analysis revealed that 12,380 (91%) were missense variants, 758 (6%) were stop-gained variants, 154 (1%) were splice-donor variants, 153 (1%) were splice-acceptor variants, 77 (0.5%) were splice region variants, and 20 (0.5%) were start-lost variants (Supplementary Table S4). Regarding the deleteriousness score, 1,887 (14%) oncogenic variants had very high CADD scores, 7,321 (54%) oncogenic variants had high CADD scores, and 4,246 (31%) had medium CADD scores. Figure 2B displays the violin plots and ranking of CADD scores of the annotated and predicted oncogenic variome related to gastric cancer driver genes. The mean CADD score of the annotated oncogenic variants was 30.6, and the annotated oncogenic variant with the highest CADD score was ATM rs587779813 (score = 55). The mean CADD score of the predicted oncogenic variants was 26.8, and the predicted oncogenic variant with the highest CADD score was FAT4 rs774644392 (score = 61). The details of the ranking of the 13,542 oncogenic variants are given in Supplementary Table S5. Finally, Figure 2C provides details on the number of annotated and predicted oncogenic variants per gastric cancer driver gene. Genes with the highest number of oncogenic variants were FAT3 (n = 2,019), FAT1 (n = 1,610), and ATM (n = 1,502).
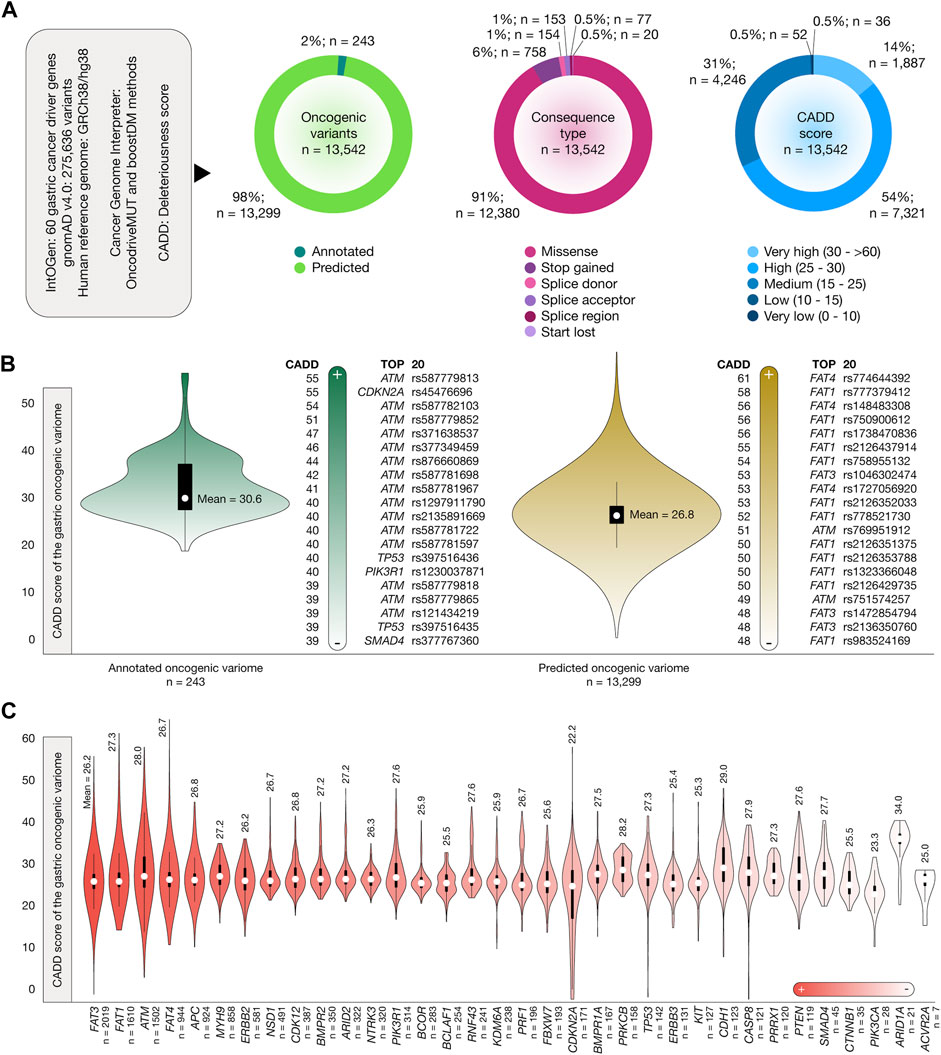
Figure 2. Stomach cancer driver genes, oncogenic variants, and Combined Annotation-Dependent Depletion (CADD) deleteriousness scores. (A) Features of stomach cancer driver genes, oncogenic variants, consequence type, and CADD deleteriousness scores. (B) Bean plots of CADD deleteriousness scores of the gastric oncogenic variome and ranking of annotated and predicted oncogenic variants with the highest CADD deleteriousness scores. (C) Ranking of the gastric cancer driver genes with the highest number of oncogenic variants and their mean CADD deleteriousness scores.
Figure 3A displays a heatmap of the 34 gastric cancer driver genes carrying annotated and predicted oncogenic variants involved in several tumorigenic processes. We identified 21 tumor suppressor genes, 19 metastatic genes, 9 oncogenes, 10 kinome genes, 3 genes encoding RNA-binding proteins, 3 cancer immunotherapy genes, 2 DNA-repair genes, and 1 cell cycle gene. Figure 3B shows a Manhattan plot of the functional enrichment analysis performed on the 34 gastric cancer driver genes with annotated and predicted oncogenic variants by using g:Profiler software (Raudvere et al., 2019). We identified 463 GO biological processes (The Gene Ontology Consortium, 2021), 39 KEGG signaling pathways (Kanehisa and Goto, 2000), 24 Reactome signaling pathways (Fabregat et al., 2016), 36 WikiPathways (Slenter et al., 2018), and 229 HP ontologies (Köhler et al., 2021). Subsequently, some of the most significant (Benjamini–Hochberg FDR q < 0.001) annotations related to gastric cancer were cell surface receptor (GO:0007166), apoptotic (GO:0097190), p53 (KEGG:04115), ErbB (KEGG:04012), Hippo (KEGG:04390), Wnt (KEGG:04310), sphingolipid (KEGG:04071), Rap1 (KEGG:04015), FoxO (KEGG:04068), PI3K-Akt (KEGG:04151), CKAP4 (WP:WP5322), and IL-18 (WP:WP4754) signaling pathways. Lastly, the stomach cancer annotation was significant as a Human Phenotype Ontology (HPO:0012126) (Figure 3C; Supplementary Table S6).
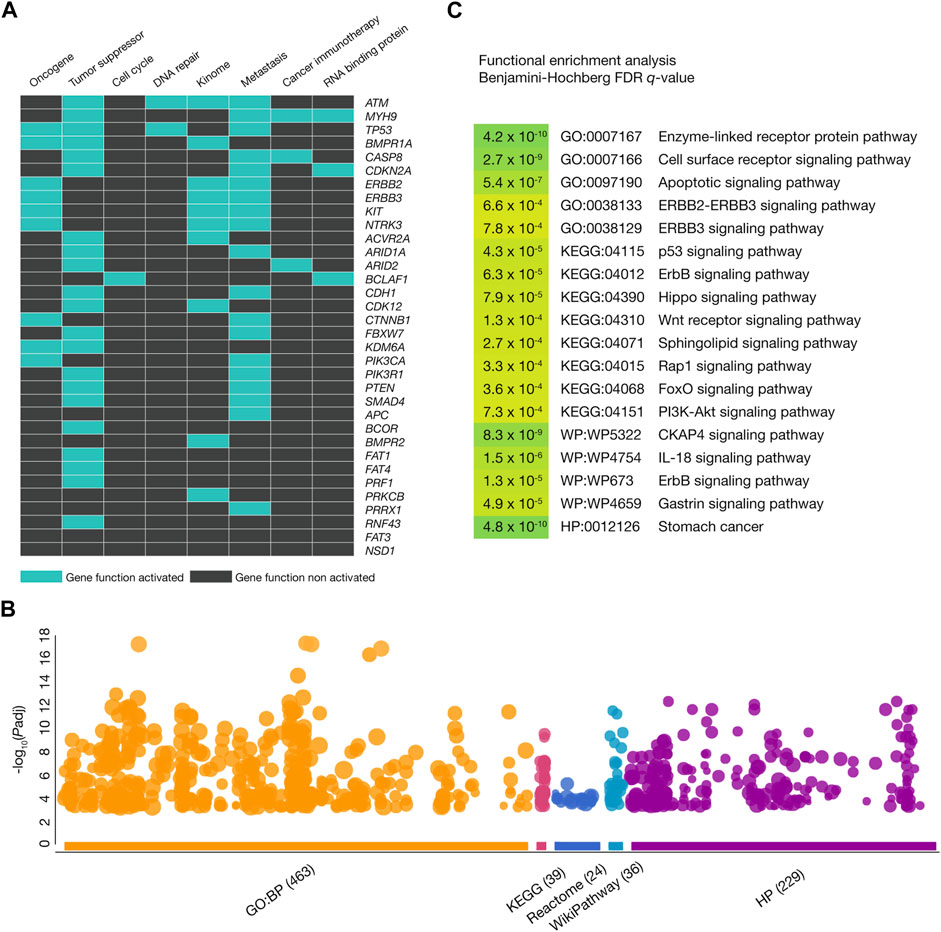
Figure 3. Functional enrichment analysis. (A) Heatmap of gastric cancer driver genes with oncogenic variants being part of oncogenes, tumor suppressor genes, cell cycle genes, DNA-repair genes, kinome, metastatic genes, cancer immunotherapy genes, and genes encoding RNA-binding proteins. (B) Most significant (Benjamini–Hochberg false discovery rate (FDR) q-value <0.001) Gene Ontology (GO) biological processes, Kyoto Encyclopedia of Genes and Genomes (KEGG) signaling pathways, WikiPathways, and Human Phenotype Ontology annotations where the gastric cancer driver genes with oncogenic variants were involved. (C) Manhattan plot of the most significant GO biological processes, KEGG signaling pathways, Reactome signaling pathways, WikiPathways, and Human Phenotype Ontology annotations.
Figure 4 shows scatter plots identifying oncogenic variants with the highest allele frequencies and the most deleterious CADD scores for each human population. The European Finnish and European non-Finnish populations had the highest mean CADD scores (26.9), followed by South Asian (26.6), Latino (26.3), East Asian (26.3), African (26.2), Middle Eastern (25.9), Amish (25.1), and Ashkenazi Jewish (25.0) populations. Worldwide, the top five oncogenic variants with the highest allele frequencies were ERBB2 rs1058808 (0.64196), ERBB2 rs1136201 (0.22083), ATM rs1800054 (0.01102), FAT1 rs111886222 (0.00922), and BCLAF1 rs77469096 (0.00749). The ERBB2 rs1058808 and ERBB2 rs1136201 oncogenic variants displayed the highest allele frequencies in the European Finnish (0.67745; 0.29819), European non-Finnish (0.67423; 0.24314), Latino (0.55676; 0.16603), East Asian (0.43731; 0.12632), South Asian (0.69270; 0.15676), African (0.22469; 0.03998), Middle Eastern (0.69702; 0.13086), Ashkenazi Jewish (0.68386; 0.14177), and Amish (0.77083; 0.27093) populations. Lastly, Figure 4 and Supplementary Table S5 provide a comprehensive ranking of oncogenic variants with the highest allele frequencies and CADD scores per human population.
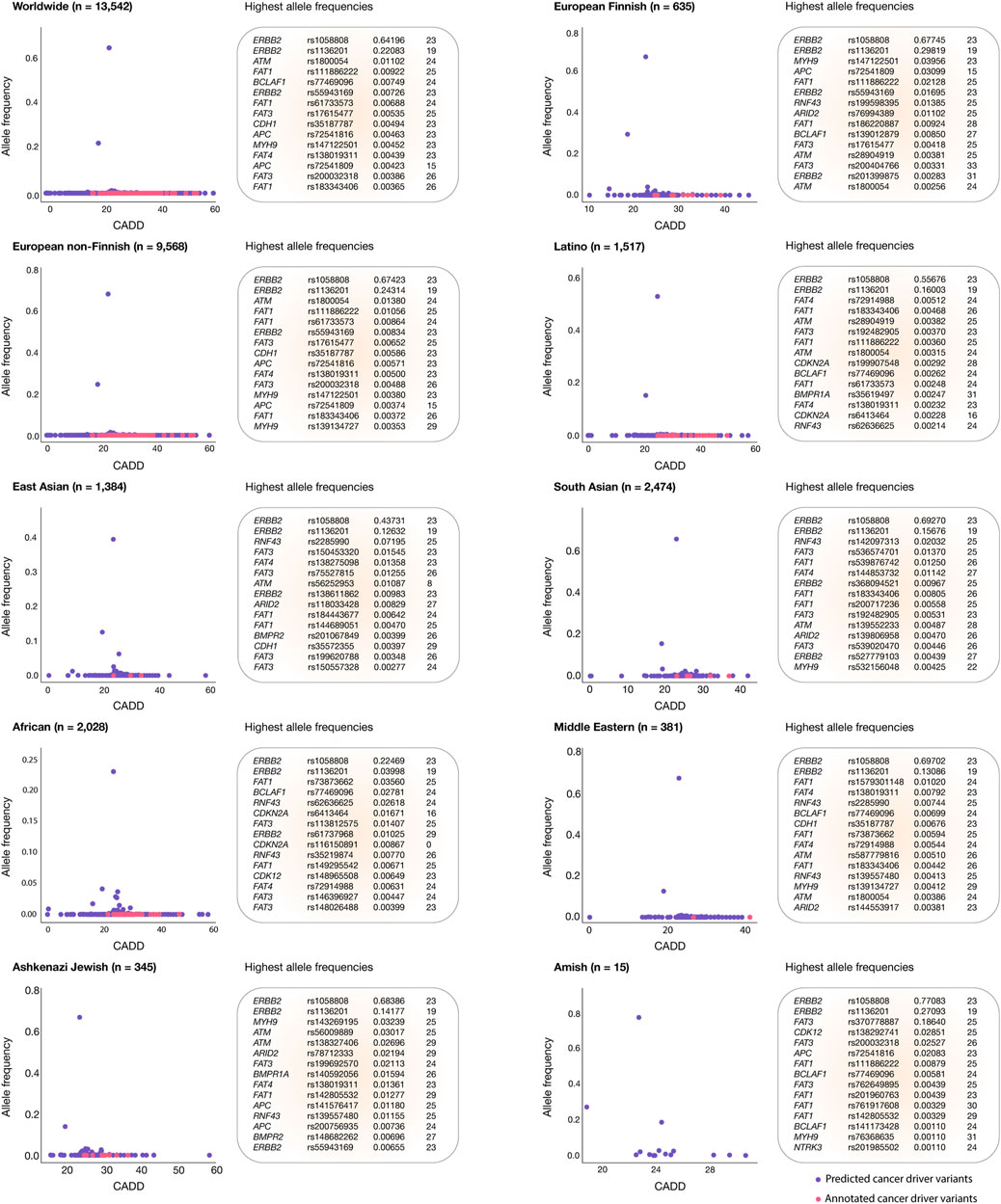
Figure 4. Gastric cancer oncogenic variants with the highest allele frequencies and CADD deleteriousness scores. Scatter plots and ranking of the annotated and predicted oncogenic variants with the highest allele frequencies and CADD deleteriousness scores from the European Finnish, European non-Finnish, Latino, East Asian, South Asian, African, Middle Eastern, Ashkenazi Jewish, and Amish populations.
The PharmGKB compiles clinical guidelines, drug labels, genotype–phenotype correlations, and actionable target–drug associations (Whirl-Carrillo et al., 2012; Barbarino et al., 2018). In relation to gastric cancer, it currently lists 12 clinical annotations that involve 9 genes, 12 variants, and 6 drugs, which include combination therapies. Fluorouracil is effective in patients carrying the EGFR rs2293347, IGFBP3 rs2960436, and IGFBP3 rs2854744 variants. A combination of fluorouracil, platinum compounds, and radiotherapy is effective for patients with the ERCC1 rs2298881, XRCC4 rs2075685, and XRCC4 rs10040363 variants. Epirubicin, fluorouracil, and oxaliplatin have demonstrated efficacy in patients with NQO1 rs1800566 and PON1 rs662 variants. Patients with the VEGFA rs25648 variant respond to a combination of cisplatin, fluorouracil, and oxaliplatin. Cisplatin is effective for patients with ERCC1 rs3212986 and ERCC1 rs11615 variants. Lastly, a regimen combining anthracyclines, cyclophosphamide, doxorubicin, epirubicin, fluorouracil, methotrexate, and oxaliplatin is effective for patients with the NOS rs1799983 variant (Figure 5A; Supplementary Table S7).
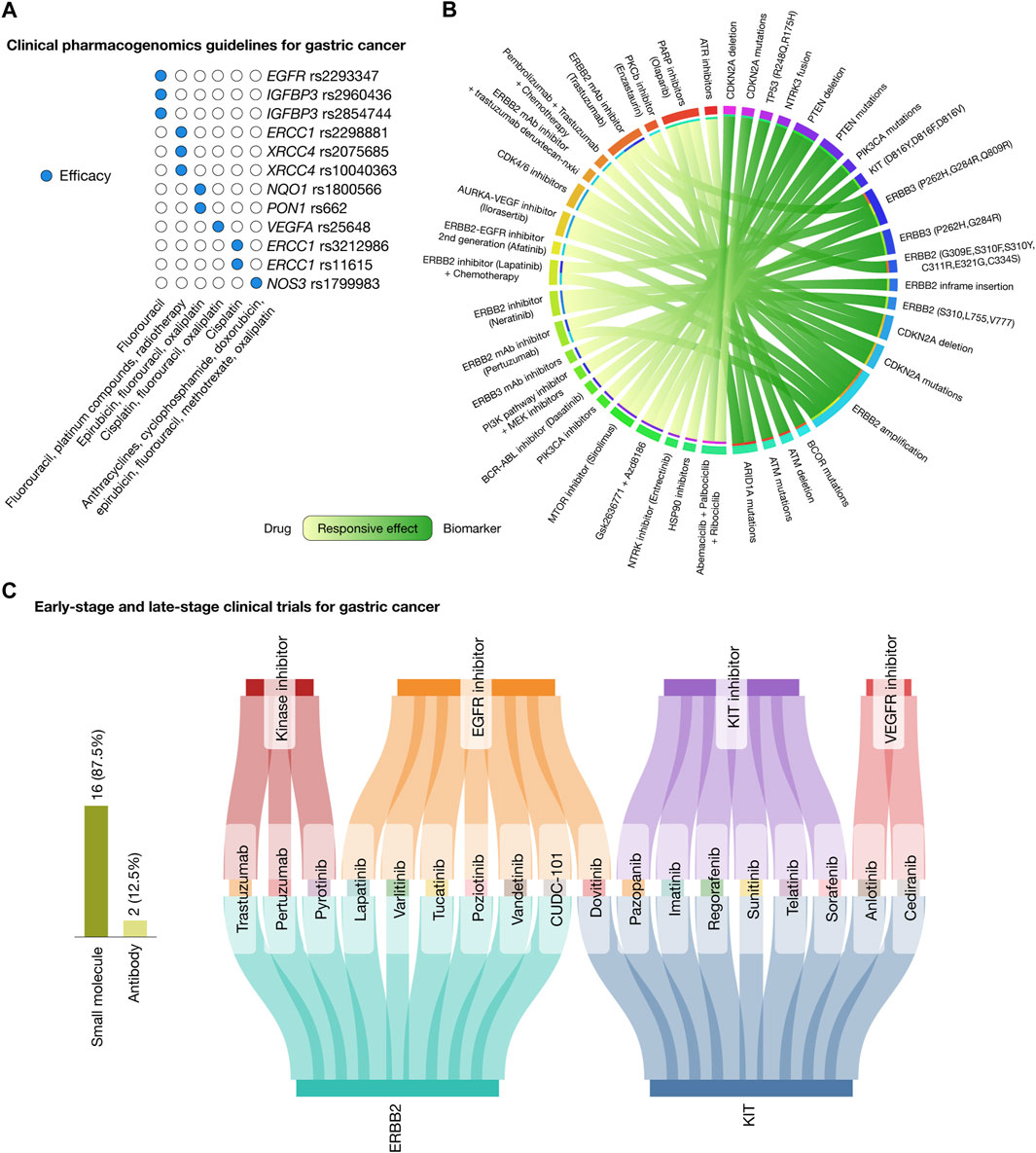
Figure 5. Landscape of therapeutic strategies based on precision oncology. (A) Current clinical pharmacogenomic guidelines for gastric cancer. (B) Circos plot showing in silico drug prescriptions of responsive effects targeting gastric cancer actionable genomic alterations. (C) Sankey plot of early-stage and late-stage clinical trials for gastric cancer connecting therapeutic targets, drugs, and mechanisms of action.
Figure 5B presents a Circos plot identifying potential biomarkers for drug response in gastric cancer treatments according to the Cancer Genome Interpreter and the Cancer Biomarker database (Dienstmann et al., 2015; Tamborero et al., 2018). Patients with ARID1A oncogenic mutations have responsive effects with ATR inhibitors and olaparib; AMT oncogenic mutations and deletions with olaparib; BCOR oncogenic mutations with enzastaurin; ERBB2 amplification with trastuzumab, afatinib, pertuzumab, a combination of lapatinib and chemotherapy, and a combination of pembrolizumab, trastuzumab, and chemotherapy; ERBB2 mutations (G309E, S310F, S310Y, C311R, E321G, and C334S) and ERBB2 inframe insertions (P780GSP, 781GSP, A775YVMA, and G776YVMA) with neratinib; CDKN2A oncogenic mutations and deletions with CDK4/6 inhibitors and ilorasertib; ERBB3 mutations (P262H, G284R, and Q809R) with ERBB3 inhibitors, trastuzumab, pertuzumab, a combination of lapatinib and chemotherapy, and a combination of the PI3K pathway inhibitor and MEK inhibitors; KIT mutations (D816Y, D816F, and D816V) with dasatinib; PIK3CA oncogenic mutations with PIK3CA inhibitors; PTEN oncogenic mutations and deletions with sirolimus, and a combination of Gsk2636771 and Azd8186; NTRK3 fusion with entrectinib; TP53 mutations (R248Q and R175H) with HSP90 inhibitors; and lastly, CDKN2A oncogenic mutations and deletions with a combination of abemaciclib, palbociclib, and ribociclib (Supplementary Table S8) (Dienstmann et al., 2015; Tamborero et al., 2018).
The Open Targets Platform provides insights into the ongoing phase I, II, III, and IV clinical trials focusing on gastric cancer driver proteins (Carvalho-Silva et al., 2019; Ochoa et al., 2021), while the Drug Repurposing Hub details the mechanism of action of the FDA-approved drugs (Corsello et al., 2017). Figure 5C depicts a Sankey plot, representing 27 clinical trial events encompassed by 2 druggable proteins (ERBB2 and KIT), 18 drugs (comprising 84% small molecules and 16% antibodies), and 4 mechanisms of action, namely, kinase inhibitors (trastuzumab, pertuzumab, and pyrotinib), EGFR inhibitors (lapatinib, varlitinib, tucatinib, poziotinib, vandetanib, CUDC-101, and dovitinib), KIT inhibitors (pazopanib, imatinib, regorafenib, sunitinib, telatinib, and sorafenib), and VEGFR inhibitors (anlotinib and cediranib) (Supplementary Table S9).
In the realm of precision oncology for gastric cancer, the focus on personalized therapeutic strategies, tailored to the unique molecular profile of an individual’s tumor, represents a significant shift in cancer treatment paradigms (Liu and Meltzer, 2017). Instead of a one-size-fits-all approach, precision oncology highlights the necessity of a deep understanding of the tumor genetic landscape, recognizing that each patient may exhibit unique targets amenable to specific treatments (Le Tourneau et al., 2019). The use of bioinformatics tools is essential in interpreting the vast amount of data produced by omics technologies, thereby uncovering actionable insights that can guide treatment decisions (Valencia and Hidalgo, 2012).
The importance of precision oncology goes beyond simply identifying molecular targets; it involves integrating these insights into a comprehensive treatment strategy that takes into account the tumor microenvironment, clinical history, genetic predisposition, and lifestyle (Le Tourneau et al., 2019). This strategy facilitates a more refined approach to gastric cancer treatment, transitioning from conventional chemotherapy regimens to targeted therapies that offer greater efficacy and fewer side effects.
However, the adoption of precision oncology encounters significant obstacles, notably in developing countries. These challenges encompass the formulation of extensive PGx clinical guidelines to guide treatment decisions across a culturally and genetically varied patient base. Moreover, there is a pressing need for rigorous cost-effectiveness analysis, suitable regulatory frameworks, and increased gene/drug trials (Quinones et al., 2014; López-Cortés et al., 2017; Salas-Hernández et al., 2023).
Additionally, the lack of representation of minority populations in genomic studies poses a considerable limitation to the universal applicability of precision oncology. The majority of the genomic data available today are derived from Caucasian populations, which distorts our understanding of cancer genetics and constrains the efficacy of targeted therapies across different ethnic groups (Guerrero et al., 2018). Overcoming this discrepancy necessitates a deliberate effort to include more diverse populations in genomic research, alongside investments in genomic testing infrastructure in underrepresented regions.
Our study contributes to this domain by pinpointing actionable genomic alterations in gastric cancer, analyzing allele frequencies across populations worldwide, and optimizing therapeutic strategies in precision oncology. In this context, we analyzed 275,634 single-nucleotide and insertion/deletion variants across 60 recognized gastric cancer driver genes from 730,947 exome sequences and 76,215 whole-genome sequences from unrelated individuals, spanning a broad spectrum of ethnic backgrounds, including African, Amish, Latino, Ashkenazi Jewish, East Asian, European Finnish, European non-Finnish, Middle Eastern, and South Asian. Such a diverse dataset underscores our commitment to inclusivity in genomic research. Our findings highlighted 13,542 oncogenic variants, with a subset showing elevated deleteriousness scores. Through a functional enrichment analysis of the 34 gastric cancer driver genes carrying oncogenic variants, we found biological significance to crucial molecular mechanisms involved in the development of gastric cancer, including apoptotic, ERBB2-ERBB3, p53, Hippo, Wnt, sphingolipid, Rap1, FoxO, PI3K-Akt, CKAP4, IL-18, and gastrin signaling pathways (Lei et al., 2022).
Understanding the impact of oncogenic variants across different ethnic groups is crucial as these variants can significantly influence the susceptibility to gastric cancer (Cordova-Delgado et al., 2021; Gonzalez-Hormazabal et al., 2021; Landeros et al., 2021). This knowledge is essential for prioritizing therapeutic strategies and making informed decisions concerning healthcare economics, public health policies, and global preventive measures. To this end, we calculated the allele frequencies of gastric cancer oncogenic variants across each ethnic group, uncovering notable differences and identifying several predominant oncogenic variants worldwide. In our findings, the European non-Finnish population had 9,568 oncogenic variants with allele frequencies greater than 0; South Asians had 2,474; Africans had 2,028; Latinos had 1,517; East Asians had 1,384; European Finnish had 635; Middle Eastern populations had 381; Ashkenazi Jewish had 345; and the Amish population had 15. Notably, the most widespread oncogenic variants globally were ERBB2 rs1058808 (0.64196) and ERBB2 rs1136201 (0.22083). Among specific ethnic groups, the most prevalent in the European non-Finnish population was ATM rs1800054 (0.01380), in South Asians was RNF43 rs142097313 (0.02032), in Africans was FAT1 rs73873662 (0.03560), in Latinos was FAT4 rs191329848 (0.00512), in East Asians was RNF43 rs2285990 (0.07195), in European Finnish was MYH9 rs147122501 (0.03956), in the Middle Eastern population was FAT1 rs1579301148 (0.01020), in Ashkenazi Jewish was MYH9 rs143269195 (0.03239), and in the Amish population was FAT3 rs370778887 (0.18640). These findings underscore the importance of a comprehensive understanding of the effect of these variants to develop preventive strategies for healthy populations and craft effective therapeutic approaches for patients with gastric cancer (Pirmohamed, 2023).
A primary goal in deciphering the cancer genome is understanding the impact of oncogenic variants on the effectiveness of anti-cancer treatments. By integrating our discoveries with PGx clinical guidelines (Barbarino et al., 2018), in silico drug prescriptions (Tamborero et al., 2018), and data from both early-stage and late-stage clinical trials (Ochoa et al., 2021), we aim to broaden the scope of precision oncology across human populations. This initiative seeks to enhance the detection of oncogenic variants in individuals with gastric cancer and those predisposed to it, thereby facilitating the creation of more personalized treatment plans. In our thorough analysis, we identified 11 key therapeutically actionable targets carrying 3,362 annotated and predicted oncogenic variants with the highest deleteriousness scores and allele frequencies across human populations. These targets exhibited variant distributions across ethnic groups and were correlated with specific treatments. Europeans of non-Finnish descent had 2,406 oncogenic variants, South Asians had 560 variants, Africans had 492 variants, Latinos had 368 variants, East Asians had 318 variants, and Europeans of Finnish descent had 164 variants in the ATM, ERBB2, ERBB3, ARID1A, PTEN, TP53, CDKN2A, NTRK3, PIK3CA, KIT, and BCOR therapeutic targets. Ashkenazi Jews had 90 variants in ATM, ERBB2, BCOR, PTEN, TP53, KIT, NTRK3, ERBB3, CDKN2A, and BCOR. The Middle Eastern population had 89 variants in ATM, ERBB2, ERBB3, BCOR, CDKN2A, PTEN, and NTRK3. Lastly, the Amish population had three variants in the ERBB2 and NTRK3 therapeutic targets.
Regarding responsive treatments analyzed through in silico drug prescriptions, ATR and PARP inhibitors, particularly olaparib, target ARID1A oncogenic mutations and AMT oncogenic mutations and deletions. Treatments for ERBB2 amplifications include trastuzumab, afatinib, pertuzumab, a combination of lapatinib and chemotherapy, and a combination of pembrolizumab, trastuzumab, and chemotherapy. Neratinib is effective for certain ERBB2 mutations (G309E, S310F, S310Y, C311R, E321G, and C334S) and ERBB2 inframe insertions (P780GSP, 781GSP, A775YVMA, and G776YVMA). CDK4/6 inhibitors and ilorasterbin are used for CDKN2A mutations and deletions. For specific ERBB3 mutations (P262H, G284R, and Q809R), treatments include ERBB3 inhibitors, trastuzumab, pertuzumab, combinations of lapatinib and chemotherapy, and combinations of PI3K pathway inhibitors and MEK inhibitors. Dasatinib targets specific KIT mutations (D816Y, D816F, and D816V); PIK3CA inhibitors are used for PIK3CA oncogenic mutations; sirolimus and a combination of Gsk2636771 and Azd8186 target PTEN mutations and deletions; entrectinib targets NTRK3 fusion; HSP90 inhibitors address certain TP53 mutations (R248Q and R175H); and a combination of abemaciclib, palbociclib, and ribociclib targets CDKN2A oncogenic mutations (Dienstmann et al., 2015; Tamborero et al., 2018). Regarding clinical trials, 27 events are investigating 18 potential drugs targeting the ERBB2 and KIT therapeutic targets. Trastuzumab and pertuzumab kinase inhibitors are under evaluation in phase III clinical trials targeting ERBB2 (NCT01774786 and NCT01041404) (Van Cutsem et al., 2015; Tabernero et al., 2023). Meanwhile, other EGFR inhibitors, KIT inhibitors, and VEGFR inhibitors are being evaluated in phases I and II clinical trials. This comprehensive approach, which integrates genomic data analysis with targeted in silico drug prescriptions and clinical trial agents, marks a significant advancement in precision oncology. It paves the way for more effective and personalized treatment options for patients with gastric cancer and underscores the potential for preventive strategies in at-risk populations across different ethnic groups.
Despite the significant advances made in precision oncology for gastric cancer, several limitations persist. The underrepresentation of minority populations in genomic studies restricts the universal applicability of findings as most genomic data are derived from Caucasian populations (Spratt et al., 2016; Guerrero et al., 2018). This disparity distorts our understanding of cancer genomics and limits the effectiveness of targeted therapies across diverse ethnic groups. Furthermore, the implementation of precision oncology, especially in developing countries, faces obstacles such as the need for comprehensive pharmacogenomics clinical guidelines, cost-effectiveness analysis, suitable regulatory frameworks, and increased gene/drug trails. These challenges highlight the urgent need for a more inclusive approach to genomic research and the development of tailored treatment strategies that consider the unique genetic landscape of each patient with cancer.
Looking ahead, it is imperative to broaden the inclusion of diverse populations in genomic studies, alongside investments in genomic testing infrastructure in regions currently underrepresented. Future research should focus on the database of oncogenic variants across varied ethnicities to forge more effective and personalized treatment options for patients with gastric cancer. There lies a promising path in leveraging in silico drug prescriptions and clinical trial data to further refine treatment strategies. The incorporation of these developments with PGx clinical guidelines is crucial for the optimization of therapeutic approaches and the creation of effective preventive strategies for at-risk populations across different ethnic groups. Additionally, enhancing investments in genetics research, particularly in developing regions, and efforts to include ethnic minorities in clinical trials and cancer research will be crucial (Paz-y-Miño et al., 2010; Paz-Y-Miño et al., 2015; Paz-Y-Miño et al., 2016; López-Cortés et al., 2018).
In conclusion, our study marks a significant advancement in applying precision oncology to gastric cancer, underscoring the critical role of genetic diversity in enhancing therapeutic strategies. Through the analysis of a wide range of oncogenic variants across diverse populations, we established a foundation for treatments that are both more equitable and effective. Integrating our discoveries with PGx clinical guidelines, in silico drug prescriptions, and clinical data heralds a new era of personalized cancer care. Nevertheless, overcoming existing limitations and adopting future perspectives require a unified global effort to ensure that precision oncology serves patients globally (Iorio et al., 2016; Pirmohamed, 2023). As we progress toward a more inclusive and personalized approach to cancer treatment, the transformative potential of precision oncology to improve patient care and outcomes becomes increasingly evident.
The original contributions presented in the study are included in the article/Supplementary Material; further inquiries can be directed to the corresponding author.
GE-G: data curation, formal analysis, investigation, methodology, software, and writing–review and editing. MR-M: data curation, formal analysis, investigation, methodology, software, and writing–review and editing. RV: data curation, formal analysis, investigation, methodology, software, and writing–review and editing. AC-A: data curation, formal analysis, investigation, methodology, software, and writing–review and editing. AA-C: data curation, formal analysis, investigation, methodology, software, and writing–review and editing. MF: data curation, formal analysis, investigation, methodology, software, and writing–review and editing. JM-G: data curation, formal analysis, investigation, methodology, software, validation, and writing–review and editing. SR-O: data curation, formal analysis, investigation, methodology, software, and writing–review and editing. PE-E: data curation, formal analysis, investigation, methodology, software, and writing–review and editing. LQ: conceptualization, data curation, formal analysis, investigation, methodology, resources, software, and writing–review and editing. AL-C: conceptualization, data curation, formal analysis, funding acquisition, investigation, methodology, project administration, resources, software, supervision, validation, visualization, writing–original draft, and writing–review and editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. Publication of this article was funded by Universidad de Las Américas, Quito, Ecuador.
This work was supported by the Latin American Society of Pharmacogenomics and Personalized Medicine (SOLFAGEM).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphar.2024.1373007/full#supplementary-material
Ajani, J. A., Lee, J., Sano, T., Janjigian, Y. Y., Fan, D., and Song, S. (2017). Gastric adenocarcinoma. Nat. Rev. Dis. Prim. 3, 17036. doi:10.1038/nrdp.2017.36
Arnedo-Pac, C., Mularoni, L., Muiños, F., Gonzalez-Perez, A., and Lopez-Bigas, N. (2019). OncodriveCLUSTL: a sequence-based clustering method to identify cancer drivers. Bioinformatics 35, 4788–4790. doi:10.1093/bioinformatics/btz501
Barbarino, J. M., Whirl-Carrillo, M., Altman, R. B., and Klein, T. E. (2018). PharmGKB: a worldwide resource for pharmacogenomic information. Wiley Interdiscip. Rev. Syst. Biol. Med. 10, e1417. doi:10.1002/wsbm.1417
Bar-Joseph, Z., Siegfried, Z., Brandeis, M., Brors, B., Lu, Y., Eils, R., et al. (2008). Genome-wide transcriptional analysis of the human cell cycle identifies genes differentially regulated in normal and cancer cells. Proc. Natl. Acad. Sci. U. S. A. 105, 955–960. doi:10.1073/pnas.0704723105
Carvalho-Silva, D., Pierleoni, A., Pignatelli, M., Ong, C., Fumis, L., Karamanis, N., et al. (2019). Open Targets Platform: new developments and updates two years on. Nucleic Acids Res. 47, D1056-D1065–D1065. doi:10.1093/nar/gky1133
Collins, R. L., Brand, H., Karczewski, K. J., Zhao, X., Alföldi, J., Francioli, L. C., et al. (2020). A structural variation reference for medical and population genetics. Nature 581, 444–451. doi:10.1038/s41586-020-2287-8
Cordova-Delgado, M., Bravo, M. L., Cumsille, E., Hill, C. N., Muñoz-Medel, M., Pinto, M. P., et al. (2021). A case-control study of a combination of single nucleotide polymorphisms and clinical parameters to predict clinically relevant toxicity associated with fluoropyrimidine and platinum-based chemotherapy in gastric cancer. BMC Cancer 21, 1030. doi:10.1186/s12885-021-08745-0
Corsello, S. M., Bittker, J. A., Liu, Z., Gould, J., McCarren, P., Hirschman, J. E., et al. (2017). The Drug Repurposing Hub: a next-generation drug library and information resource. Nat. Med. 23, 405–408. doi:10.1038/nm.4306
Dienstmann, R., Jang, I. S., Bot, B., Friend, S., and Guinney, J. (2015). Database of genomic biomarkers for cancer drugs and clinical targetability in solid tumors. Cancer Discov. 5, 118–123. doi:10.1158/2159-8290.CD-14-1118
Dietlein, F., Weghorn, D., Taylor-Weiner, A., Richters, A., Reardon, B., Liu, D., et al. (2020). Identification of cancer driver genes based on nucleotide context. Nat. Genet. 52, 208–218. doi:10.1038/s41588-019-0572-y
Eid, S., Turk, S., Volkamer, A., Rippmann, F., and Fulle, S. (2017). KinMap: a web-based tool for interactive navigation through human kinome data. BMC Bioinforma. 18, 16. doi:10.1186/s12859-016-1433-7
Fabregat, A., Sidiropoulos, K., Garapati, P., Gillespie, M., Hausmann, K., Haw, R., et al. (2016). The reactome pathway knowledgebase. Nucleic Acids Res. 44, D481–D487. doi:10.1093/nar/gkv1351
Garraway, L. A., Verweij, J., and Ballman, K. V. (2013). Precision oncology: an overview. J. Clin. Oncol. 31, 1803–1805. doi:10.1200/JCO.2013.49.4799
Gonzalez-Hormazabal, P., Pelaez, D., Musleh, M., Bustamante, M., Stambuk, J., Pisano, R., et al. (2021). NOD1 rs2075820 (p.E266K) polymorphism is associated with gastric cancer among individuals infected with cagPAI-positive H. pylori. Biol. Res. 54, 13. doi:10.1186/s40659-021-00336-4
Green, E. D., Gunter, C., Biesecker, L. G., Di Francesco, V., Easter, C. L., Feingold, E. A., et al. (2020). Strategic vision for improving human health at the Forefront of Genomics. Nature 586, 683–692. doi:10.1038/s41586-020-2817-4
Guerrero, S., López-Cortés, A., Indacochea, A., García-Cárdenas, J. M., Zambrano, A. K., Cabrera-Andrade, A., et al. (2018). Analysis of racial/ethnic representation in select basic and applied cancer research studies. Sci. Rep. 8, 13978. doi:10.1038/s41598-018-32264-x
Hanahan, D. (2022). Hallmarks of cancer: new dimensions. Cancer Discov. 12, 31–46. doi:10.1158/2159-8290.CD-21-1059
Hentze, M. W., Castello, A., Schwarzl, T., and Preiss, T. (2018). A brave new world of RNA-binding proteins. Nat. Rev. Mol. Cell. Biol. 19, 327–341. doi:10.1038/nrm.2017.130
ICGC/TCGA Pan-Cancer Analysis of Whole Genomes Consortium (2020). Pan-cancer analysis of whole genomes. Nature 578, 82–93. doi:10.1038/s41586-020-1969-6
Iorio, F., Knijnenburg, T. A., Vis, D. J., Bignell, G. R., Menden, M. P., Schubert, M., et al. (2016). A landscape of pharmacogenomic interactions in cancer. Cell. 166, 740–754. doi:10.1016/j.cell.2016.06.017
Jumper, J., Evans, R., Pritzel, A., Green, T., Figurnov, M., Ronneberger, O., et al. (2021). Highly accurate protein structure prediction with AlphaFold. Nature 596, 583–589. doi:10.1038/s41586-021-03819-2
Kandoth, C., McLellan, M. D., Vandin, F., Ye, K., Niu, B., Lu, C., et al. (2013). Mutational landscape and significance across 12 major cancer types. Nature 502, 333–339. doi:10.1038/nature12634
Kanehisa, M., and Goto, S. (2000). KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 28, 27–30. doi:10.1093/nar/28.1.27
Karczewski, K. J., Francioli, L. C., Tiao, G., Cummings, B. B., Alföldi, J., Wang, Q., et al. (2020). The mutational constraint spectrum quantified from variation in 141,456 humans. Nature 581, 434–443. doi:10.1038/s41586-020-2308-7
Kircher, M., Witten, D. M., Jain, P., O’Roak, B. J., Cooper, G. M., and Shendure, J. (2014). A general framework for estimating the relative pathogenicity of human genetic variants. Nat. Genet. 46, 310–315. doi:10.1038/ng.2892
Köhler, S., Gargano, M., Matentzoglu, N., Carmody, L. C., Lewis-Smith, D., Vasilevsky, N. A., et al. (2021). The human phenotype ontology in 2021. Nucleic Acids Res. 49, D1207–D1217. doi:10.1093/nar/gkaa1043
Landeros, N., Corvalan, A. H., Musleh, M., Quiñones, L. A., Varela, N. M., and Gonzalez-Hormazabal, P. (2021). Novel risk associations between microRNA polymorphisms and gastric cancer in a Chilean population. Int. J. Mol. Sci. 23, 467. doi:10.3390/ijms23010467
Lange, S. S., Takata, K., and Wood, R. D. (2011). DNA polymerases and cancer. Nat. Rev. Cancer 11, 96–110. doi:10.1038/nrc2998
Lawrence, M. S., Stojanov, P., Mermel, C. H., Robinson, J. T., Garraway, L. A., Golub, T. R., et al. (2014). Discovery and saturation analysis of cancer genes across 21 tumour types. Nature 505, 495–501. doi:10.1038/nature12912
Lei, Z.-N., Teng, Q.-X., Tian, Q., Chen, W., Xie, Y., Wu, K., et al. (2022). Signaling pathways and therapeutic interventions in gastric cancer. Signal Transduct. Target. Ther. 7, 358. doi:10.1038/s41392-022-01190-w
Le Tourneau, C., Borcoman, E., and Kamal, M. (2019). Molecular profiling in precision medicine oncology. Nat. Med. 25, 711–712. doi:10.1038/s41591-019-0442-2
Liu, X., and Meltzer, S. J. (2017). Gastric cancer in the era of precision medicine. Cell. Mol. Gastroenterol. Hepatol. 3, 348–358. doi:10.1016/j.jcmgh.2017.02.003
López-Cortés, A., Abarca, E., Silva, L., Velastegui, E., León-Sosa, A., Karolys, G., et al. (2021). Identification of key proteins in the signaling crossroads between wound healing and cancer hallmark phenotypes. Sci. Rep. 11, 17245. doi:10.1038/s41598-021-96750-5
López-Cortés, A., Cabrera-Andrade, A., Vázquez-Naya, J. M., Pazos, A., Gonzáles-Díaz, H., Paz-Y-Miño, C., et al. (2020a). Prediction of breast cancer proteins involved in immunotherapy, metastasis, and RNA-binding using molecular descriptors and artificial neural networks. Sci. Rep. 10, 8515. doi:10.1038/s41598-020-65584-y
López-Cortés, A., Guerrero, S., Redal, M. A., Alvarado, A. T., and Quiñones, L. A. (2017). State of art of cancer pharmacogenomics in Latin american populations. Int. J. Mol. Sci. 18, 639. doi:10.3390/ijms18060639
López-Cortés, A., Leone, P. E., Freire-Paspuel, B., Arcos-Villacís, N., Guevara-Ramírez, P., Rosales, F., et al. (2018). Mutational analysis of oncogenic AKT1 gene associated with breast cancer risk in the high altitude Ecuadorian mestizo population. Biomed. Res. Int. 2018, 7463832. doi:10.1155/2018/7463832
López-Cortés, A., Paz-Y-Miño, C., Guerrero, S., Cabrera-Andrade, A., Barigye, S. J., Munteanu, C. R., et al. (2020b). OncoOmics approaches to reveal essential genes in breast cancer: a panoramic view from pathogenesis to precision medicine. Sci. Rep. 10, 5285. doi:10.1038/s41598-020-62279-2
López-Cortés, A., Paz-Y-Miño, C., Guerrero, S., Jaramillo-Koupermann, G., León Cáceres, Á., Intriago-Baldeón, D. P., et al. (2020c). Pharmacogenomics, biomarker network, and allele frequencies in colorectal cancer. Pharmacogenomics J. 20, 136–158. doi:10.1038/s41397-019-0102-4
López-Cortés, A., Prathap, L., Ortiz-Prado, E., Kyriakidis, N. C., León Cáceres, Á., Armendáriz-Castillo, I., et al. (2022). The close interaction between hypoxia-related proteins and metastasis in pancarcinomas. Sci. Rep. 12, 11100. doi:10.1038/s41598-022-15246-y
Lu, C., Xie, M., Wendl, M. C., Wang, J., McLellan, M. D., Leiserson, M. D. M., et al. (2015). Patterns and functional implications of rare germline variants across 12 cancer types. Nat. Commun. 6, 10086. doi:10.1038/ncomms10086
Manning, G., Whyte, D. B., Martinez, R., Hunter, T., and Sudarsanam, S. (2002). The protein kinase complement of the human genome. Science 298, 1912–1934. doi:10.1126/science.1075762
Mansoori, B., Mohammadi, A., Davudian, S., Shirjang, S., and Baradaran, B. (2017). The different mechanisms of cancer drug resistance: a brief review. Adv. Pharm. Bull. 7, 339–348. doi:10.15171/apb.2017.041
Martincorena, I., Raine, K. M., Gerstung, M., Dawson, K. J., Haase, K., Van Loo, P., et al. (2017). Universal patterns of selection in cancer and somatic tissues. Cell. 171, 1029–1041. doi:10.1016/j.cell.2017.09.042
Martínez-Jiménez, F., Muiños, F., Sentís, I., Deu-Pons, J., Reyes-Salazar, I., Arnedo-Pac, C., et al. (2020). A compendium of mutational cancer driver genes. Nat. Rev. Cancer 20, 555–572. doi:10.1038/s41568-020-0290-x
Muiños, F., Martínez-Jiménez, F., Pich, O., Gonzalez-Perez, A., and Lopez-Bigas, N. (2021). In silico saturation mutagenesis of cancer genes. Nature 596, 428–432. doi:10.1038/s41586-021-03771-1
Mularoni, L., Sabarinathan, R., Deu-Pons, J., Gonzalez-Perez, A., and López-Bigas, N. (2016). OncodriveFML: a general framework to identify coding and non-coding regions with cancer driver mutations. Genome Biol. 17, 128. doi:10.1186/s13059-016-0994-0
Nurk, S., Koren, S., Rhie, A., Rautiainen, M., Bzikadze, A. V., Mikheenko, A., et al. (2022). The complete sequence of a human genome. Science 376, 44–53. doi:10.1126/science.abj6987
Ochoa, D., Hercules, A., Carmona, M., Suveges, D., Gonzalez-Uriarte, A., Malangone, C., et al. (2021). Open Targets Platform: supporting systematic drug-target identification and prioritisation. Nucleic Acids Res. 49, D1302–D1310. doi:10.1093/nar/gkaa1027
Patel, S. J., Sanjana, N. E., Kishton, R. J., Eidizadeh, A., Vodnala, S. K., Cam, M., et al. (2017). Identification of essential genes for cancer immunotherapy. Nature 548, 537–542. doi:10.1038/nature23477
Paz-y-Miño, C., López-Cortés, A., Muñoz, M. J., Cabrera, A., Castro, B., and Sánchez, M. E. (2010). Incidence of the L858R and G719S mutations of the epidermal growth factor receptor oncogene in an Ecuadorian population with lung cancer. Cancer Genet. cytogenet. 196, 201–203. doi:10.1016/j.cancergencyto.2009.10.007
Paz-Y-Miño, C., Robles, P., Salazar, C., Leone, P. E., García-Cárdenas, J. M., Naranjo, M., et al. (2016). Positive association of the androgen receptor CAG repeat length polymorphism with the risk of prostate cancer. Mol. Med. Rep. 14, 1791–1798. doi:10.3892/mmr.2016.5414
Paz-Y-Miño, C. A., García-Cárdenas, J. M., López-Cortés, A., Salazar, C., Serrano, M., and Leone, P. E. (2015). Positive association of the cathepsin D ala224val gene polymorphism with the risk of alzheimer’s disease. Am. J. Med. Sci. 350, 296–301. doi:10.1097/MAJ.0000000000000555
Pérez-Villa, A., Echeverría-Garcés, G., Ramos-Medina, M. J., Prathap, L., Martínez-López, M., Ramírez-Sánchez, D., et al. (2023). Integrated multi-omics analysis reveals the molecular interplay between circadian clocks and cancer pathogenesis. Sci. Rep. 13, 14198. doi:10.1038/s41598-023-39401-1
Pirmohamed, M. (2023). Pharmacogenomics: current status and future perspectives. Nat. Rev. Genet. 24, 350–362. doi:10.1038/s41576-022-00572-8
Porta-Pardo, E., Kamburov, A., Tamborero, D., Pons, T., Grases, D., Valencia, A., et al. (2017). Comparison of algorithms for the detection of cancer drivers at subgene resolution. Nat. Methods 14, 782–788. doi:10.1038/nmeth.4364
Quinones, L. A., Lavanderos, M. A., Cayun, J. P., Garcia-Martin, E., Agundez, J. A., Caceres, D. D., et al. (2014). Perception of the usefulness of drug/gene pairs and barriers for pharmacogenomics in Latin America. Curr. Drug Metab. 15, 202–208. doi:10.2174/1389200215666140202220753
Raguz, S., and Yagüe, E. (2008). Resistance to chemotherapy: new treatments and novel insights into an old problem. Br. J. Cancer 99, 387–391. doi:10.1038/sj.bjc.6604510
Ramos-Medina, M. J., Echeverría-Garcés, G., Kyriakidis, N. C., León Cáceres, Á., Ortiz-Prado, E., Bautista, J., et al. (2024). CardiOmics signatures reveal therapeutically actionable targets and drugs for cardiovascular diseases. Heliyon 10, e23682. doi:10.1016/j.heliyon.2023.e23682
Raudvere, U., Kolberg, L., Kuzmin, I., Arak, T., Adler, P., Peterson, H., et al. (2019). g:Profiler: a web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucleic Acids Res. 47, W191-W198–W198. doi:10.1093/nar/gkz369
Relling, M. V., and Klein, T. E. (2011). CPIC: clinical pharmacogenetics implementation consortium of the pharmacogenomics research network. Clin. Pharmacol. Ther. 89, 464–467. doi:10.1038/clpt.2010.279
Relling, M. V., Klein, T. E., Gammal, R. S., Whirl-Carrillo, M., Hoffman, J. M., and Caudle, K. E. (2020). The clinical pharmacogenetics implementation consortium: 10 years later. Clin. Pharmacol. Ther. 107, 171–175. doi:10.1002/cpt.1651
Rentzsch, P., Witten, D., Cooper, G. M., Shendure, J., and Kircher, M. (2019). CADD: predicting the deleteriousness of variants throughout the human genome. Nucleic Acids Res. 47, D886-D894–D894. doi:10.1093/nar/gky1016
Rheinbay, E., Nielsen, M. M., Abascal, F., Wala, J. A., Shapira, O., Tiao, G., et al. (2020). Analyses of non-coding somatic drivers in 2,658 cancer whole genomes. Nature 578, 102–111. doi:10.1038/s41586-020-1965-x
Ross, C. J. D., Visscher, H., Sistonen, J., Brunham, L. R., Pussegoda, K., Loo, T. T., et al. (2010). The Canadian pharmacogenomics network for drug safety: a model for safety pharmacology. Thyroid 20, 681–687. doi:10.1089/thy.2010.1642
Rubio-Perez, C., Tamborero, D., Schroeder, M. P., Antolín, A. A., Deu-Pons, J., Perez-Llamas, C., et al. (2015). In silico prescription of anticancer drugs to cohorts of 28 tumor types reveals targeting opportunities. Cancer Cell. 27, 382–396. doi:10.1016/j.ccell.2015.02.007
Salas-Hernández, A., Galleguillos, M., Carrasco, M., López-Cortés, A., Redal, M. A., Fonseca-Mendoza, D., et al. (2023). An updated examination of the perception of barriers for pharmacogenomics implementation and the usefulness of drug/gene pairs in Latin America and the Caribbean. Front. Pharmacol. 14, 1175737. doi:10.3389/fphar.2023.1175737
Schubach, M., Maass, T., Nazaretyan, L., Röner, S., and Kircher, M. (2024). CADD v1.7: using protein language models, regulatory CNNs and other nucleotide-level scores to improve genome-wide variant predictions. Nucleic Acids Res. 52, D1143–D1154. doi:10.1093/nar/gkad989
Sjöblom, T., Jones, S., Wood, L. D., Parsons, D. W., Lin, J., Barber, T. D., et al. (2006). The consensus coding sequences of human breast and colorectal cancers. Science 314, 268–274. doi:10.1126/science.1133427
Slenter, D. N., Kutmon, M., Hanspers, K., Riutta, A., Windsor, J., Nunes, N., et al. (2018). WikiPathways: a multifaceted pathway database bridging metabolomics to other omics research. Nucleic Acids Res. 46, D661-D667–D667. doi:10.1093/nar/gkx1064
Sondka, Z., Bamford, S., Cole, C. G., Ward, S. A., Dunham, I., and Forbes, S. A. (2018). The COSMIC Cancer Gene Census: describing genetic dysfunction across all human cancers. Nat. Rev. Cancer 18, 696–705. doi:10.1038/s41568-018-0060-1
Spratt, D. E., Chan, T., Waldron, L., Speers, C., Feng, F. Y., Ogunwobi, O. O., et al. (2016). Racial/ethnic disparities in genomic sequencing. JAMA Oncol. 2, 1070–1074. doi:10.1001/jamaoncol.2016.1854
Sung, H., Ferlay, J., Siegel, R. L., Laversanne, M., Soerjomataram, I., Jemal, A., et al. (2021). Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 71, 209–249. doi:10.3322/caac.21660
Swen, J. J., Nijenhuis, M., de Boer, A., Grandia, L., Maitland-van der Zee, A. H., Mulder, H., et al. (2011). Pharmacogenetics: from bench to byte--an update of guidelines. Clin. Pharmacol. Ther. 89, 662–673. doi:10.1038/clpt.2011.34
Tabernero, J., Hoff, P. M., Shen, L., Ohtsu, A., Shah, M. A., Siddiqui, A., et al. (2023). Pertuzumab, trastuzumab, and chemotherapy in HER2-positive gastric/gastroesophageal junction cancer: end-of-study analysis of the JACOB phase III randomized clinical trial. Gastric Cancer 26, 123–131. doi:10.1007/s10120-022-01335-4
Tamborero, D., Gonzalez-Perez, A., Perez-Llamas, C., Deu-Pons, J., Kandoth, C., Reimand, J., et al. (2013). Comprehensive identification of mutational cancer driver genes across 12 tumor types. Sci. Rep. 3, 2650. doi:10.1038/srep02650
Tamborero, D., Rubio-Perez, C., Deu-Pons, J., Schroeder, M. P., Vivancos, A., Rovira, A., et al. (2018). Cancer Genome Interpreter annotates the biological and clinical relevance of tumor alterations. Genome Med. 10, 25. doi:10.1186/s13073-018-0531-8
Tan, P., and Yeoh, K.-G. (2015). Genetics and molecular pathogenesis of gastric adenocarcinoma. Gastroenterology 149, 1153–1162. doi:10.1053/j.gastro.2015.05.059
The Gene Ontology Consortium (2021). The Gene Ontology resource: enriching a GOld mine. Nucleic Acids Res. 49, D325–D334. doi:10.1093/nar/gkaa1113
Tokheim, C., Bhattacharya, R., Niknafs, N., Gygax, D. M., Kim, R., Ryan, M., et al. (2016). Exome-scale discovery of hotspot mutation regions in human cancer using 3D protein structure. Cancer Res. 76, 3719–3731. doi:10.1158/0008-5472.CAN-15-3190
Valencia, A., and Hidalgo, M. (2012). Getting personalized cancer genome analysis into the clinic: the challenges in bioinformatics. Genome Med. 4, 61. doi:10.1186/gm362
Van Cutsem, E., Bang, Y.-J., Feng-Yi, F., Xu, J. M., Lee, K.-W., Jiao, S.-C., et al. (2015). HER2 screening data from ToGA: targeting HER2 in gastric and gastroesophageal junction cancer. Gastric Cancer 18, 476–484. doi:10.1007/s10120-014-0402-y
Varela, N. M., Guevara-Ramírez, P., Acevedo, C., Zambrano, T., Armendáriz-Castillo, I., Guerrero, S., et al. (2021). A new insight for the identification of oncogenic variants in breast and prostate cancers in diverse human populations, with a focus on latinos. Front. Pharmacol. 12, 630658. doi:10.3389/fphar.2021.630658
Vasan, N., Baselga, J., and Hyman, D. M. (2019). A view on drug resistance in cancer. Nature 575, 299–309. doi:10.1038/s41586-019-1730-1
Weghorn, D., and Sunyaev, S. (2017). Bayesian inference of negative and positive selection in human cancers. Nat. Genet. 49, 1785–1788. doi:10.1038/ng.3987
Whirl-Carrillo, M., McDonagh, E. M., Hebert, J. M., Gong, L., Sangkuhl, K., Thorn, C. F., et al. (2012). Pharmacogenomics knowledge for personalized medicine. Clin. Pharmacol. Ther. 92, 414–417. doi:10.1038/clpt.2012.96
Wood, R. D., Mitchell, M., Sgouros, J., and Lindahl, T. (2001). Human DNA repair genes. Science 291, 1284–1289. doi:10.1126/science.1056154
Yao, F., Kausalya, J. P., Sia, Y. Y., Teo, A. S. M., Lee, W. H., Ong, A. G. M., et al. (2015). Recurrent fusion genes in gastric cancer: CLDN18-ARHGAP26 induces loss of epithelial integrity. Cell. Rep. 12, 272–285. doi:10.1016/j.celrep.2015.06.020
Keywords: gastric cancer, precision oncology, ethnic groups, genomic alterations, therapeutic strategies
Citation: Echeverría-Garcés G, Ramos-Medina MJ, Vargas R, Cabrera-Andrade A, Altamirano-Colina A, Freire MP, Montalvo-Guerrero J, Rivera-Orellana S, Echeverría-Espinoza P, Quiñones LA and López-Cortés A (2024) Gastric cancer actionable genomic alterations across diverse populations worldwide and pharmacogenomics strategies based on precision oncology. Front. Pharmacol. 15:1373007. doi: 10.3389/fphar.2024.1373007
Received: 19 January 2024; Accepted: 10 April 2024;
Published: 02 May 2024.
Edited by:
Marcelo Rizzatti Luizon, Federal University of Minas Gerais, BrazilReviewed by:
Andrea Orellana-Manzano, Facultad de Ciencias de la Vida (FCV), EcuadorCopyright © 2024 Echeverría-Garcés, Ramos-Medina, Vargas, Cabrera-Andrade, Altamirano-Colina, Freire, Montalvo-Guerrero, Rivera-Orellana, Echeverría-Espinoza, Quiñones and López-Cortés. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Andrés López-Cortés, YWFsYzg0QGdtYWlsLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.