 James N. Bates1†
James N. Bates1† Paulina M. Getsy2
Paulina M. Getsy2 Gregory A. Coffee2
Gregory A. Coffee2 Santhosh M. Baby3†
Santhosh M. Baby3† Peter M. MacFarlane2
Peter M. MacFarlane2 Yee-Hsee Hsieh4
Yee-Hsee Hsieh4 Zackery T. Knauss5
Zackery T. Knauss5 Jason A. Bubier6
Jason A. Bubier6 Devin Mueller5
Devin Mueller5 Stephen J. Lewis2,7,8*
Stephen J. Lewis2,7,8*- 1Department of Anesthesiology, University of Iowa Hospitals and Clinics, Iowa City, IA, United States
- 2Department of Pediatrics, Case Western Reserve University, Cleveland, OH, United States
- 3Section of Biology, Galleon Pharmaceuticals, Inc., Horsham, PA, United States
- 4Division of Pulmonary, Critical Care and Sleep Medicine, Case Western Reserve University, Cleveland, OH, United States
- 5Department of Biological Sciences, Kent State University, Kent, OH, United States
- 6Jackson Laboratories, Bar Harbor, ME, United States
- 7Department of Pharmacology, Case Western Reserve University, Cleveland, OH, United States
- 8Functional Electrical Stimulation Center, Case Western Reserve University, Cleveland, OH, United States
The molecular mechanisms underlying the acquisition of addiction/dependence on morphine may result from the ability of the opioid to diminish the transport of L-cysteine into neurons via inhibition of excitatory amino acid transporter 3 (EAA3). The objective of this study was to determine whether the co-administration of the cell-penetrant L-thiol ester, L-cysteine ethyl ester (L-CYSee), would reduce physical dependence on morphine in male Sprague Dawley rats. Injection of the opioid-receptor antagonist, naloxone HCl (NLX; 1.5 mg/kg, IP), elicited pronounced withdrawal phenomena in rats which received a subcutaneous depot of morphine (150 mg/kg) for 36 h and were receiving a continuous infusion of saline (20 μL/h, IV) via osmotic minipumps for the same 36 h period. The withdrawal phenomena included wet-dog shakes, jumping, rearing, fore-paw licking, 360° circling, writhing, apneas, cardiovascular (pressor and tachycardia) responses, hypothermia, and body weight loss. NLX elicited substantially reduced withdrawal syndrome in rats that received an infusion of L-CYSee (20.8 μmol/kg/h, IV) for 36 h. NLX precipitated a marked withdrawal syndrome in rats that had received subcutaneous depots of morphine (150 mg/kg) for 48 h) and a co-infusion of vehicle. However, the NLX-precipitated withdrawal signs were markedly reduced in morphine (150 mg/kg for 48 h)-treated rats that began receiving an infusion of L-CYSee (20.8 μmol/kg/h, IV) at 36 h. In similar studies to those described previously, neither L-cysteine nor L-serine ethyl ester (both at 20.8 μmol/kg/h, IV) mimicked the effects of L-CYSee. This study demonstrates that 1) L-CYSee attenuates the development of physical dependence on morphine in male rats and 2) prior administration of L-CYSee reverses morphine dependence, most likely by intracellular actions within the brain. The lack of the effect of L-serine ethyl ester (oxygen atom instead of sulfur atom) strongly implicates thiol biochemistry in the efficacy of L-CYSee. Accordingly, L-CYSee and analogs may be a novel class of therapeutics that ameliorate the development of physical dependence on opioids in humans.
Introduction
There are numerous problems faced by clinicians treating patients with opioid use disorder (OUD) and other substance use disorders (SUDs) (McCarberg, 2011; Heberlein et al., 2012; Garcia-Portilla et al., 2014; Hah et al., 2017; Dydyk et al., 2022; 2023; Matson et al., 2022; Horn et al., 2023). With regard to managing pain in the general population, the key issues are how to provide μ-opioid receptor (μ-OR) agonist (e.g., fentanyl)-induced analgesia without eliciting 1) euphoria, 2) physical dependence and/or psychological addiction, 3) hyperalgesia, and 4) any combination of the aforementioned conditions (Benyamin et al., 2008; Volkow et al., 2018; Mercadante et al., 2019; Manhapra, 2022; Preux et al., 2022; Balanaser et al., 2023). With respect to treating moderate-to-severe OUD (DSM-5 terminology for opioid addiction in 10%–20% of people who have liability for SUD), the key issues are 1) how to manage the often severe opioid withdrawal (all current strategies to manage opioid withdrawal are inadequate); 2) how to manage patients who have gone through withdrawal and need medication to block euphoria and/or physical dependence, with the three medications available, naltrexone, buprenorphine, and methadone, all having strengths, but major weaknesses; and 3) how to avoid opioid euphoria and/or physical dependence in patients with moderate-to-severe OUD who currently are sober, but require μ-OR agonist analgesia (Humphreys et al., 2022; Jiménez-Fernández et al., 2022; Torres-Lockhart et al., 2022; Alvarez-Perez et al., 2023; Biancuzzi et al., 2023; Frankeberger et al., 2023). With respect to effectively treating SUD involving opioids and other substances (e.g., alcohol, cannabinoids, benzodiazepines, and psycho-stimulants), in 10%–20% of the population prone to SUD, the important issue is how to provide (yet to be developed) effective therapeutics that will attenuate/block dopamine surge-mediated euphoria of each of these families of brain-reward drugs as an adjunct to treating addictive diseases (Chartoff and Connery, 2014; Stephan and Parsa, 2016; Horsfall and Sprague, 2017; Bechara et al., 2019; Serafini and Zachariou, 2019). With respect to making opioid analgesics safer, we lack drugs that effectively modulate the actions of opioids to improve their analgesic profile. The therapeutics should 1) not interfere with or promote opioid analgesia, 2) prevent the acquisition of physical dependence and psychological addiction to opioids, 3) block opioid-induced respiratory depression (OIRD), or 4) stop the development of hyperalgesia (Benyamin et al., 2008; Morgan and Christie, 2011; Volkow et al., 2018; Mercadante et al., 2019; Manhapra, 2022; Preux et al., 2022; Balanaser et al., 2023).
Trivedi et al. (2014) reported that morphine-induced dependence/addiction may involve redox-based changes in global DNA methylation and retrotransposon transcription via the blockade of excitatory amino acid transporter type 3 (EAA3; also known as EAAC1)-dependent uptake of the amino acid, L-cysteine, into brain neurons. Some of the temporal steps described by Trivedi et al. (2014) (see Figure 5 of Trivedi et al., 2014) and others (Lin et al., 2001; Ikemoto et al., 2002; Mao et al., 2002; Xu et al., 2003; Xu et al., 2006; Christie, 2008; Yang et al., 2008; Wang et al., 2009; Daijo et al., 2011; Gutowicz et al., 2011; Liu et al., 2011; Maze and Nestler, 2011; Lim et al., 2012; Sun et al., 2012; Browne et al., 2020) are as follows: 1) morphine blockade of L-cysteine uptake into neurons by G protein-dependent inhibition of EAA3 activity, 2) resulting decreases in intracellular levels of L-cysteine, L-glutathione, and the methylation index, namely, S-adenosyl-methionine/S-adenosyl-homocysteine (SAM/SAH ratio), 3) decreases in the methylation status of global CpG (regions of DNA where a cytosine nucleotide is followed by a guanine nucleotide in the linear sequence of bases along its 5′ → 3′ direction) and in CpG methylation of long interspersed nuclear element-1 (LINE-1) retrotransposon regulatory regions, and 4) activation of transcription of previously silenced LINE-1 genes. Thus, we hypothesized that co-administration of cell-permeant versions of L-cysteine, such as L-cysteine ethyl ester (L-CYSee) (Goto et al., 1983; Hisadome et al., 1986a; Hisadome et al., 1986b; Hisadome et al., 1988; Servin et al., 1988; Hisadome et al., 1990; Schöneich et al., 1992; Hobbs et al., 1993; Fukui et al., 1994; Ding and Demple, 1998; Galanakis et al., 2004; Mosier-Boss and Lieberman, 2005; Perissinotti et al., 2005; Defonsi Lestard et al., 2013; Mendoza et al., 2013; Arias et al., 2019), may prevent the acquisition of physical dependence on morphine and reverse established dependence on the opioid. Previously, we reported that L-CYSee (Lewis et al., 2022), L-cysteine methyl ester (Getsy et al., 2022a), and other thiolesters and related compounds (Baby et al., 2021; Baby et al., 2021; Gaston et al., 2021; Jenkins et al., 2021; Getsy et al., 2022b; Getsy et al., 2022c; Getsy et al., 2022d; Getsy et al., 2022e; Getsy et al., 2022f) prevent and/or reverse the adverse effects of morphine and fentanyl on ventilatory parameters, arterial blood–gas chemistry (pH, pCO2, pO2, and sO2), and alveolar–arterial gradient (index of alveolar gas exchange in the lungs) in freely-moving rats without compromising opioid-induced analgesia or sedation. We now provide evidence that administration of L-CYSee prevents the acquisition of physical dependence, as measured by markedly fewer withdrawal phenomena in response to administration of the opioid receptor antagonist, naloxone HCl (NLX), in freely-moving male rats, and reverses established dependence. The lack of effect of L-cysteine and L-serine ethyl ester (L-SERee, oxygen atom instead of a sulfur atom as in L-CYSee) in these paradigms suggests that the efficacy of L-CYSee involves its cell penetrability in brain regions vital to the expression of morphine dependence, and points to the vital role of thiol biochemistry in the biological efficacy of L-CYSee.
Materials and methods
Permissions, rats, and surgical procedures
All studies were carried out in strict accordance with the NIH Guide for Care and Use of Laboratory Animals (NIH Publication No. 80-23) revised in 1996 and in strict compliance with the ARRIVE (Animal Research: Reporting of In Vivo Experiments) guidelines (http://www.nc3rs.org.uk/). All protocols involving the use of rats were approved by the Animal Care and Use Committees of Galleon Pharmaceuticals (PC0022), Case Western Reserve University (2015-0025), and the University of Virginia (3642-09-07). Adult male Sprague Dawley rats of approximately 12 weeks of age at the time of study were purchased from Harlan Industries (Madison, WI, United States). The rats were given 5 days to recover from transportation before being subjected to surgeries, as described in this paragraph. (+)-Morphine sulfate was obtained from Baxter Healthcare (Deerfield, IL, United States). L-cysteine hydrochloride monohydrate (L-cysteine HCl) powder (C7880, PubChem Substance ID:24892992), L-cysteine ethyl ester hydrochloride (L-CYSee HCl) powder (C121908, PubChem Substance ID: 24892386), and L-serine ethyl ester hydrochloride (L-SERee HCl) powder (223123; PubChem Substance ID:24853367) were divided into 100 mg amounts under N2 gas and stored at 4°C. Solutions of L-cysteine HCl, L-CYSee HCl, and L-SERee HCl were dissolved in saline and brought to pH 7.2 with 0.1 M NaOH at room temperature immediately before use. Naloxone hydrochloride dihydrate powder (BP548; PubChem Substance ID: 24278050) was obtained from Sigma-Aldrich (St. Louis, MO, USA) and was dissolved in saline at pH 7.2 with 0.1 M NaOH at room temperature immediately before use. On the day of the study, all arterial and venous catheters were flushed with 0.3 mL of phosphate-buffered saline (0.1 M, pH 7.4) 3–4 h before commencement of the study. All studies were done in a quiet room with a relative humidity of 50% ± 2% and room temperature of 21.3°C ± 0.2°C. Every group described in this study contained nine rats. Other than the described surgeries (see the next section), no rat had a prior history of use in any experimental protocol and was used in only one protocol in the present study. The times at which the surgeries were performed will be described herein. Surgery times from the initiation of anesthesia to final closing of all wounds took approximately a) 20 min for the placement of one venous catheter, b) 25 min for the placement of two venous catheters, c) 30 min for the placement of two intravenous and one arterial catheter, and d) 20 min for intravenous catheterization in which the connected osmotic minipump is placed subcutaneously. Two protocols were as follows: 1) Examine the ability of L-CYSee and L-cysteine (behavioral and cardiorespiratory studies), and L-SERee (behavioral study only) to prevent the acquisition of physical dependence on morphine upon 36 h exposure to the opioid and test compounds. The question addressed whether treatment with L-CYSee, for example, diminishes NLX-precipitated withdrawal phenomena; and 2) examine whether the introduction of L-CYSee or L-cysteine (behavioral and cardiorespiratory studies) or L-SERee (behavioral study only) at 36 h of morphine treatment reverses established physical dependence on morphine, as tested after 48 h of morphine treatment. The question addressed for the second protocol is whether co-treatment for 12 h with L-CYSee, for example, reverses existing physical dependence on morphine, as expressed by markedly diminished NLX-precipitated withdrawal phenomena.
Protocols to determine the effects of L-CYSee on physical dependency of morphine and prevention of morphine dependence—36-h studies
A. Behavioral studies: At 2 a.m. on the day of surgery, two groups of rats received a jugular vein catheter (PE-10 connected to PE-50) under 2%–3% isoflurane anesthesia (Henderson et al., 2014; Gaston et al., 2021; Getsy et al., 2022f). The jugular vein catheter was connected to a primed ALZET osmotic minipump (Model 2002; ALZA Corporation, CA, United States) positioned at the back of the neck to allow continuous infusion of the vehicle (20 μL/h, IV), L-cysteine (20.8 μmol/kg/h, IV), L-CYSee (20.8 μmol/kg/h, IV), or L-SERee (20.8, μmol/kg/h, IV), as described previously (Jarrott et al., 1987; Lewis et al., 1988a; Jarrott et al., 1988; Lewis et al., 1989). Physical dependence was induced by a slow-release subcutaneous depot of morphine emulsion (150 mg/kg, SC) injected at the left side of the neck, as described in detail by Fennessy and colleagues (Lee and Fennessy, 1970; Laska and Fennessy, 1976; Laska and Fennessy, 1977; Laska and Fennessy, 1978; Lewis et al., 1988b). In brief, morphine base was precipitated from a solution of (+)-morphine sulfate by titrating to pH 9 with 1 mol/L NaOH. After several distilled water washes, a pure base was collected in a filter funnel and dried. Morphine slow-release emulsion was prepared by suspending a weighed amount of base in liquid paraffin and Arlacel A. This mixture was then emulsified with an equal volume of normal saline, as initially described by Collier et al. (1972). All wounds were sutured closed, and the rats were returned to their warmed home cages. After 35.5 h of morphine administration, the rats were placed in individual opaque plastic boxes; after 30 min of acclimatization, they received an intraperitoneal (IP) injection of NLX (1.5 mg/kg), and behavioral phenomena were scored for 45 min by at least three scorers. The scored phenomena were as follows: jumping behavior—all four paws of the ground—jumps; wet-dog shakes—whole body shakes as if to shed water from fur; rearing behavior—rearing on hind legs—rears; episodes of fore-paw licking—FPL; circling behavior—complete 360° rotation; writhes—full-body contortion; sneezes—episodes of sneezing—abrupt expulsion of air that often disturbed the fine bedding material.
B. Plethysmography ventilatory studies: Groups of rats were prepared as described previously except that the rats received a second catheter into the jugular vein, as described by Getsy et al. (2022f), to give a bolus injection of NLX. After 35 h, rats were placed in individual whole-body plethysmography chambers (Young et al., 2013; Getsy et al., 2022a; Getsy et al., 2022b; Getsy et al., 2022c; Getsy et al., 2022d; Getsy et al., 2022e; Getsy et al., 2022f), and the free end of the exteriorized venous catheter was connected to a swivel assembly housed in the lid of the plethysmography chamber (Young et al., 2013; Getsy et al., 2022a; Getsy et al., 2022b; Getsy et al., 2022c; Getsy et al., 2022d; Getsy et al., 2022e; Getsy et al., 2022f). After a 60-min acclimatization period, the rats were given an intravenous injection of NLX (1.5 mg/kg). Ventilatory parameters including the frequency of breathing, tidal volume, minute ventilation, and non-eupneic breathing indices were recorded (to be reported elsewhere) with the number of apneas (>1.5 s between breaths) reported here.
C. Cardiovascular studies: Groups of rats were prepared as described previously except that the rats received a second catheter into a jugular vein (Getsy et al., 2022f) to administer NLX and a catheter into a femoral artery to continuously record mean arterial blood pressure (MAP) and heart rate (Kanbar et al., 2010; Davisson et al., 2014; Brognara et al., 2016; Gaston et al., 2020). After 35 h, the rats were placed in individual opaque plastic boxes, and the free end of the exteriorized jugular vein catheter was connected to an injection line to deliver NLX. The free end of the arterial line was connected to tubing attached to a computer-coupled pressure transducer (Cabe Lab, Inc.) to record pulsatile arterial blood pressure. After a 60-min acclimatization period, the rats received a bolus injection of NLX (1.5 mg/kg, IV), and MAP and heart rate were recorded continuously for 45 min.
D. Body temperature and body weight studies: Groups of rats without a second jugular catheter were prepared as described in the previous paragraph. After 35 h, the rats were placed in individual opaque plastic boxes, and a thermistor probe, used to record body temperature, was connected to a telethermometer (Yellow Springs Instruments) and inserted 5–6 cm into the rectum and taped to the tail (Kregel et al., 1997). The body weights of the rats and their body temperatures were recorded every 15 min during the acclimatization period to establish accurate baseline values. After the 60-min acclimatization period, the rats received an intraperitoneal injection of NLX (1.5 mg/kg), and body temperatures and body weights were recorded every 15 min for 90 min.
Reversal of morphine dependence—48-h studies
A. Behavioral studies: At 2 p.m. on the day of surgery, two groups of rats received a slow-release subcutaneous depot of morphine emulsion (150 mg/kg, SC) injected at the left side of the neck, as described previously. After 36 h of morphine administration, the rats were anesthetized (2% isoflurane) and received a jugular vein catheter connected to a primed ALZET osmotic minipump positioned at the back of the neck for continuous infusion of the vehicle (20 μL/h, IV), L-cysteine (20.8, μmol/kg/h, IV), L-CYSee (20.8 μmol/kg/h, IV), or L-SERee (20.8, μmol/kg/h, IV), as mentioned previously. All wounds were then sutured closed, and the rats were returned to their warmed home cages. After 11.5h, the rats were placed in individual opaque plastic boxes, and after a 30-min period of acclimatization, the rats received an intraperitoneal injection of NLX (1.5 mg/kg), and behavioral phenomena (as detailed previously) were scored for 45 min by at least three scorers.
B. Plethysmography ventilatory studies: Groups of rats were prepared as described previously, except that the rats received two catheters into the same jugular vein, as described by Getsy et al. (2022f), to allow for the bolus injection of NLX via the catheter not connected to the osmotic minipump. After 47 h, rats were placed in individual whole body plethysmography chambers. The free end of the exteriorized jugular vein catheter was connected to a swivel on the lid of the plethysmography chamber. After 60 min of acclimatization, the rats received a bolus injection of NLX (1.5 mg/kg, IV). Ventilatory parameters and non-eupneic breathing indices were recorded, with the number of apneas (>1.5 s between breaths) reported.
C. Cardiovascular studies: Groups of rats were prepared as previously described, except that the rats received two catheters into the same jugular vein to inject NLX via the catheter not connected to the osmotic minipump, and a catheter into a femoral artery to record MAP and heart rate. After 47 h, the rats were placed in individual opaque plastic boxes, and the free end of the exteriorized jugular vein catheter was connected to an injection line to inject NLX. The free end of the arterial line was connected to tubing attached to a computer-coupled pressure transducer to record pulsatile arterial blood pressure. After a 60-min acclimatization period, the rats received an injection of NLX (1.5 mg/kg, IV), and MAP and heart rate were recorded continuously for 45 min.
D. Body temperature and body weight studies: Groups of rats without a second jugular catheter were prepared, as described previously. After 47 h, the rats were placed in individual opaque plastic boxes, and a thermistor probe, used to record body temperature, was connected to a telethermometer (Yellow Springs Instruments) and inserted 5–6 cm into the rectum and taped to the tail. The body weights of the rats and body temperatures were recorded every 15 min during acclimatization to establish baseline values. After the 60-min acclimatization period, the rats received an intraperitoneal injection of NLX (1.5 mg/kg). Body temperature and weights were recorded every 15 min for 90 min.
Data analyses
The directly recorded and arithmetically derived parameters were statistically analyzed. All data are presented as mean ± SEM and were evaluated using one-way ANOVA, followed by Bonferroni corrections for multiple comparisons between means using the error mean square terms from each ANOVA analysis (Wallenstein et al., 1980; Ludbrook, 1998; McHugh, 2011), as detailed previously (Getsy et al., 2023a; Getsy et al., 2023b). A p < 0.05 value denoted the initial level of statistical significance that was modified according to the number of comparisons between means, as described by Wallenstein et al. (1980). The modified t-statistic is t = (mean group 1—mean group 2)/[s x (1/n1 + 1/n2)1/2], where s2 = the mean square within groups obtained from ANOVA (the square root of this value is used in the modified t-statistic formula), and n1 and n2 are the number of rats in each group under comparison. Based on Bonferroni’s inequality, a conservative critical value for modified t-statistics can be obtained from the tables of t-distribution using a significance level of P/m, where m is the number of comparisons to be made between groups (Winer, 1971). The degrees of freedom are those of the mean square for within group variation from the ANOVA table. The critical Bonferroni value can be approximated from the tables of the normal curve by t* = z + (z + z3)/4n, with n being the degrees of freedom and z being the critical normal curve value for P/m (Wallenstein et al., 1980; Ludbrook, 1998; McHugh, 2011). Wallenstein et al. (1980) first demonstrated that the Bonferroni procedure is preferable for general use because it provides critical values that are lower than those of other procedures when the number of comparisons can be limited and will be slightly larger than those of other procedures if many comparisons are made. Statistical analyses were performed with the aid of GraphPad Prism software (GraphPad Software, Inc., La Jolla, CA). F- and P-statistics associated with the ANOVA analyses of the data in Figures 1–5 are given in the respective figure legends.
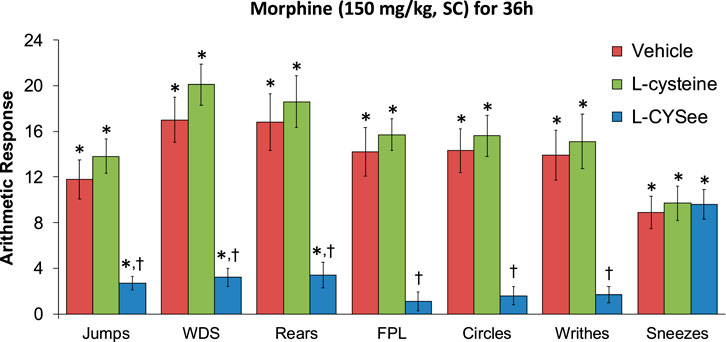
FIGURE 1. Withdrawal behaviors elicited by a bolus injection of NLX (1.5 mg/kg, IP) in rats treated for 36 h with a subcutaneous depot of morphine (150 mg/kg) along with continuous infusion of the vehicle (saline, 20 μL/h, IV) or L-cysteine (20.8 μmol/kg/h, IV) or L-cysteine ethylester (L-CYSee, 20.8 μmol/kg/h, IV). Withdrawal signs: jumps—all four paws off the floor; WDS-wet-dog shakes; rears—rearing on hind legs; FPL—episodes of fore-paw licking; circles-a 360° rotation; writhes-full-body contortion; sneezes-abrupt expulsion of air. The data are presented as mean ± SEM. There were nine rats in each group. Between-group ANOVA statistics: jumps: F2,24 = 18.8, p = 0.00001; WDS: F2,24 = 29.9, p < 0.00001; rears: F2,24 = 16.2, p = 0.00004; FPL: F2,24 = 27.0, p < 0.00001; circles: F2,24 = 24.1, p < 0.00001; writhes: F2,24 = 15.4, p < 0.00005; sneezes: F2,24 = 0.09, p = 0.92. *p < 0.05, significant responses from Pre. †p < 0.05, L-CYSee versus vehicle or L-cysteine.
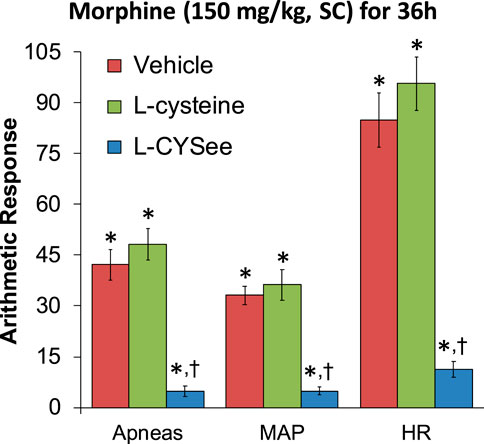
FIGURE 2. Incidence of apneas (>1.5 s between breaths) and increases in mean arterial blood pressure (MAP, mmHg) and heart rate (HR, beats/min) elicited by a bolus injection of NLX (1.5 mg/kg, IV) in rats treated for 36 h with a subcutaneous depot of morphine (150 mg/kg) along with the continuous infusion of vehicle (saline, 20 μL/h, IV), L-cysteine (20.8 μmol/kg/h, IV), or L-cysteine ethyl ester (L-CYSee, 20.8 μmol/kg/h, IV). The data are presented as mean ± SEM. There were nine rats in each group. Between-group ANOVA statistics: MAP: F2,24 = 28.5, p < 0.00001; HR: F2,24 = 47.8, p < 0.00001; apneas: F2,24 = 37.2, p < 0.00001. *p < 0.05, significant responses from Pre. †p < 0.05, L-CYSee versus vehicle or L-cysteine.
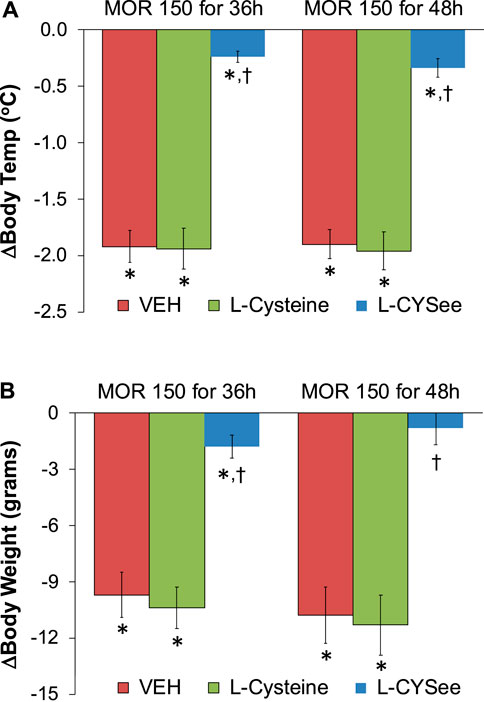
FIGURE 3. Arithmetic changes in body temperatures (A) and body weights (B) elicited by a bolus injection of NLX (1.5 mg/kg, IP) in rats treated for 36 h with a subcutaneous depot of morphine (150 mg/kg) or for 48 h with a subcutaneous depot of morphine (150 mg/kg) with continuous infusion of vehicle (saline, 20 μL/h, IV) or L-cysteine (20.8 μmol/kg/h, IV) or L-CYSee (20.8 μmol/kg/h, IV) for 36 h or beginning after 36 h for 12 h. The data are presented as mean ± SEM. There were nine rats in each group. Between-group ANOVA statistics: body temperature: 36 h: F2,24 = 51.4, p < 0.00001; 48 h: F2,24 = 50.1, p < 0.00001; body weight 36 h: F2,24 = 22.8, <0.00001; 48 h: F2,24 = 18.6, p < 0.00001. *p < 0.05, significant responses from Pre. †p < 0.05, L-CYSee versus vehicle or L-cysteine.
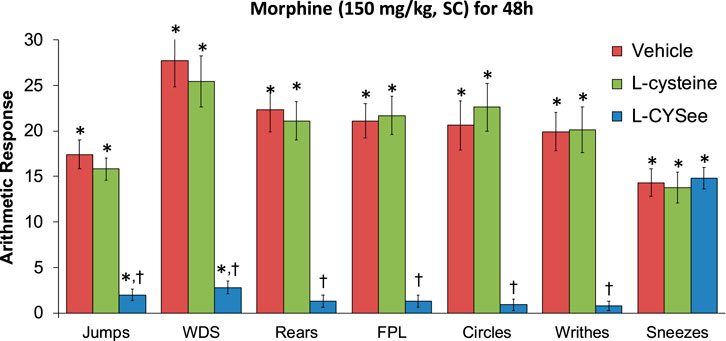
FIGURE 4. Withdrawal behaviors elicited by a bolus injection of NLX (1.5 mg/kg, IP) in rats treated for 48 h with a subcutaneous depot of morphine (150 mg/kg) along with continuous infusion of vehicle (saline, 20 μL/h, IV) or L-cysteine (20.8 μmol/kg/h, IV) or L-cysteine ethylester (L-CYSee, 20.8 μmol/kg/h, IV) that began after 36 h of morphine administration. Withdrawal signs: jumps—all four paws off the floor; WDS-wet-dog shakes; rears—rearing on hind legs; FPL—episodes of fore-paw licking; circles-a 360° rotation; writhes—full-body contortion; sneezes—abrupt expulsion of air. The data are presented as mean ± SEM. There were nine rats in each group. Between-group ANOVA statistics: jumps: F2,24 = 48.8, p < 0.00001; WDS: F2,24 = 33.9, p < 0.00001; rears: F2,24 = 38.7, <0.00001; FPL: F2,24 = 42.3, p < 0.00001; circles: F2,24 = 29.4, p < 0.00001; writhes: F2,24 = 34.0, p < 0.00005; sneezes: F2,24 = 0.11, p = 0.89. *p < 0.05, significant responses from Pre. †p < 0.05, L-CYSee versus vehicle or L-cysteine.
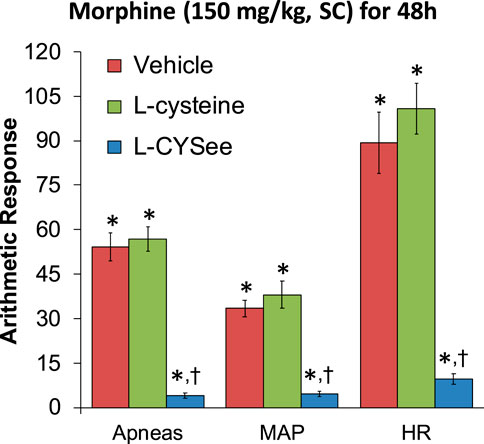
FIGURE 5. Incidence of apneas (>1.5 s between breaths) and increases in mean arterial blood pressure (MAP, mmHg) and heart rate (HR, beats/min) elicited by a bolus injection of NLX (1.5 mg/kg, IV) in rats treated for 48 h with a subcutaneous depot of morphine (150 mg/kg) along with the continuous infusion of vehicle (saline, 20 μL/h, IV), L-cysteine (20.8 μmol/kg/h, IV), or L-cysteine ethyl ester (L-CYSee, 20.8 μmol/kg/h, IV) that began at 36 h of morphine administration for 12 h. The data are presented as mean ± SEM. There were nine rats in each group. Between-group ANOVA statistics: MAP: F2,24 = 33.3, p < 0.00001; HR: F2,24 = 41.8, p < 0.00001; apneas: F2,24 = 69.9, p < 0.00001. *p < 0.05, significant responses from Pre. †p < 0.05, L-CYSee versus vehicle or L-cysteine.
Results
L-CYSee infusion prevents physical dependence on morphine—36-h studies
The behavioral withdrawal phenomena elicited by the injection of NLX (1.5 mg/kg, IP) in rats that were receiving morphine (150 mg/kg, SC), plus a continuous infusion of the vehicle (saline, 20 μL/h, IV) or L-cysteine (20.8 μmol/kg/h, IV) or L-CYSee (20.8 μmol/kg/h, IV) for 36 h are shown in Figure 1. As can be seen, the injection of NLX to rats receiving the infusion of the vehicle elicited substantial increases in jumping behavior (jumps), wet-dog shakes (WDS), rearing behavior (rears), fore-paw licking (FPL), circling behavior (circles), full-body writhing (writhes), and sneezing (sneezes). These responses were similar in magnitude in rats that were receiving an infusion of L-cysteine. In contrast, NLX-precipitated phenomena (except for sneezing) were dramatically diminished in rats that were receiving the continuous infusion of L-CYSee. In addition, the NLX-precipitated withdrawal signs were fully expressed in rats that were receiving an infusion of L-SERee (Table 1). Note that we did not pursue L-SERee studies to see whether it affected the expression of apneic events, and elevations in MAP and heart rate elicited by the injection of NLX.
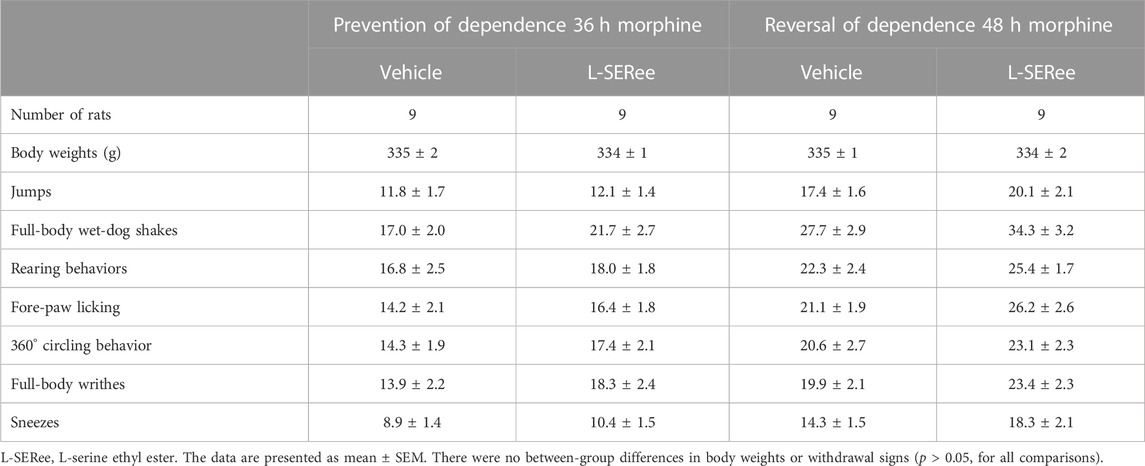
TABLE 1. Naloxone-precipitated withdrawal signs in morphine-treated rats receiving continuous infusion of vehicle or L-serine ethyl ester for 36 h (prevention) or continuous infusion of vehicle or L-serine ethyl ester that began after 36 h of morphine administration and lasted for 12 h (reversal).
The increases in apneic events and elevations in MAP and heart rate, elicited by the injection of NLX (1.5 mg/kg) in rats that were receiving morphine (150 mg/kg, SC) and continuous infusion of vehicle or L-cysteine or L-CYSee are summarized in Figure 2. The injection of NLX in rats that were receiving the vehicle elicited substantial increases in the number of apneas and increases in MAP (sustained hypertension) and in heart rate (HR, tachycardia). These NLX-precipitated events were similar in magnitude in the rats that were receiving the infusion of L-cysteine. In contrast, the NLX-precipitated withdrawal phenomena were markedly reduced in rats receiving continuous infusion of L-CYSee. Actual values for MAP and heart rate before and after injection of NLX in morphine-treated rats receiving infusions of vehicle, L-cysteine, or L-CYSee are shown in Supplementary Table S1. Resting MAP and heart rate values before the injection of NLX were similar in the three groups of rats.
The changes in body temperatures and body weights elicited by the injection of NLX (1.5 mg/kg) in rats that were receiving morphine (150 mg/kg, SC) for 36 h, and continuous infusion of vehicle or L-cysteine or L-CYSee are summarized in Figure 3. The injection of NLX elicited marked decreases in body temperatures and body weights that were similar in magnitude in rats receiving infusion of vehicle or L-cysteine. The NLX-induced responses were markedly smaller in the rats that were receiving an infusion of L-CYSee. Actual body temperatures and body weights before and after injection of NLX in rats receiving morphine for 36 hours with co-infusions of vehicle, L-cysteine, or L-CYSee are shown in Supplementary Table S2. Resting body temperature and body weight values before the injection of NLX were similar in the three groups of rats. After 36 h of morphine treatment, body temperatures were elevated by just over 0.5°C in rats receiving infusions of vehicle or L-cysteine. Body temperature was not increased in rats receiving L-CYSee. The injection of NLX elicited substantial decreases in body temperatures and body weights in the vehicle- or L-cysteine-infusion groups, and much smaller responses in the rats receiving the infusion of L-CYSee.
L-CYSee reversal of physical dependence on morphine—48-h studies
The behavioral withdrawal phenomena elicited by the injection of NLX (1.5 mg/kg, IP) in rats that were receiving morphine (150 mg/kg, SC) for 48 h plus a continuous infusion of the vehicle (saline, 20 μL/h, IV) or L-cysteine (20.8 μmol/kg/h, IV) or L-CYSee (20.8 μmol/kg/h, IV) beginning after 36 h are shown in Figure 4. The injection of NLX into rats receiving the vehicle infusion elicited substantial increases in withdrawal behaviors. These responses were similar in magnitude in rats receiving infusion of L-cysteine, whereas withdrawal phenomena (except for sneezing) were dramatically diminished in rats receiving infusion of L-CYSee for 12 h. NLX-precipitated withdrawal signs in rats receiving an infusion of L-SERee were similar to those receiving infusion of vehicle (Table 1).
The increases in apneic events and elevations in MAP and heart rate, elicited by the injection of NLX (1.5 mg/kg, IP) in rats receiving morphine (150 mg/kg, SC) for 48 h and continuous infusion of vehicle or L-cysteine or L-CYSee beginning at 36 h are shown in Figure 5. NLX elicited substantial increases in apneas and elevations in MAP (hypertension) and in heart rate (HR, tachycardia) in rats receiving infusion of vehicle, and similar responses in rats receiving infusion of L-cysteine. In contrast, NLX-precipitated withdrawal phenomena were markedly reduced in rats receiving L-CYSee infusion. Actual values for MAP and heart rate before and after the injection of NLX in morphine-treated rats receiving infusion of vehicle, L-cysteine, or L-CYSee are shown in Supplementary Table S3. Resting MAP and heart rate values before the injection of NLX were similar in the three groups.
The changes in body temperatures and body weights elicited by the injection of NLX (1.5 mg/kg) in rats receiving morphine (150 mg/kg, SC) and infusion of vehicle or L-cysteine or L-CYSee that began 36 h after morphine administration and lasted for 12 h are summarized in Figure 3. The injection of NLX elicited pronounced decreases in body temperatures and body weights that were similar in magnitude in rats receiving vehicle or L-cysteine. These withdrawal responses were markedly smaller in rats receiving L-CYSee. Full details of the body temperatures and body weight before and after injection of NLX in these morphine-treated rats that were receiving infusions of vehicle, L-cysteine, or L-CYSee are shown in Supplementary Table S4. Resting body temperature and body weight values before the injection of NLX were similar in the three groups of rats. After 48 h of morphine treatment, body temperatures were elevated by just over 0.5°C in rats receiving infusions of vehicle or L-cysteine. The body temperatures were not elevated in rats receiving the infusion of L-CYSee. Body weights were similar in all three groups. The injection of NLX elicited substantial decreases in body temperatures and body weights in the vehicle- or L-cysteine-infusion groups, and much smaller responses in the rats receiving infusion of L-CYSee.
Discussion
The first set of major observations of this study was that co-infusion of the L-thiol ester, L-CYSee, markedly reduced the expression of multiple withdrawal signs (behavioral, cardiorespiratory, body weight loss, and hypothermia) elicited by the injection of the opioid receptor antagonist, NLX, in male Sprague Dawley rats treated for 36 h with slow-release morphine emulsion. The behavioral withdrawal signs indicative of the rats having become physically dependent on morphine, such as jumping, wet-dog shakes, rearing, fore-paw licking, circling, writhing, and sneezing (rapid expulsions of air), as well as decreases in body weight and body temperature, were consistent with previous reports published using this same slow-release morphine model (Lee and Fennessy, 1970; Laska and Fennessy, 1976; Laska and Fennessy, 1977; Laska and Fennessy, 1978; Lewis et al., 1988b), and with a wide variety of other administration protocols used to induce morphine dependence (Hutchinson et al., 2007; Lopez-Gimenez and Milligan, 2010; Morgan and Christie, 2011; Nielsen and Kreek, 2012). The increases in MAP and heart rate elicited by NLX are new findings in our morphine-dependence model, but are in full agreement with evidence that NLX-precipitated withdrawal is associated with hypertension and tachycardia in experimental animals (Buccafusco, 1983; Buccafusco, 1990; Buccafusco et al., 1984; Marshall and Buccafusco, 1985; Dixon and Chang, 1988; Chang and Dixon, 1990; Delle et al., 1990; Baraban et al., 1993) and humans (Newlin et al., 1992; Purssell et al., 1995; Walsh et al., 2003; Levin et al., 2019; Balshaw et al., 2021; Isoardi et al., 2022; Lee et al., 2022). This is due to globalized activation of the sympathetic nervous system. Finally, our finding that NLX elicited a substantial increase in apneic events is new to our morphine-dependence model, but also consistent with findings in rats (Baraban et al., 1993; Baldo, 2022) and humans (Schwarzer et al., 2015; Zamani et al., 2020; Wilson et al., 2023). The inability of L-cysteine to modify the NLX-precipitated withdrawal phenomena certainly suggests that the efficacy of L-CYSee involves the entry of this cell-penetrant L-thiol ester (Goto et al., 1983; Hisadome et al., 1986a; Hisadome et al., 1986b; Hisadome et al., 1988; Servin et al., 1988; Hisadome et al., 1990; Schöneich et al., 1992; Hobbs et al., 1993; Fukui et al., 1994; Ding and Demple, 1998; Galanakis et al., 2004; Mosier-Boss and Lieberman, 2005; Perissinotti et al., 2005; Defonsi Lestard et al., 2013; Mendoza et al., 2013; Arias et al., 2019; Lewis et al., 2022) into neurons involved in the acquisition of physical dependence/addiction on morphine (Laschka et al., 1976a; Laschka et al., 1976b; Laschka and Herz, 1977; Koob, 1987; Saiepour et al., 2001; Glass, 2010; Gardner, 2011; Glass, 2011). Moreover, the inability of L-SERee to prevent the acquisition of physical dependence on morphine indicates that the sulfur atom is vital to the ability of L-CYSee to prevent the intracellular processes within the brain by which morphine induces physical dependence (Deslandes et al., 2002; Gardner, 2011; Koob and Volkow, 2016; Volkow et al., 2019; Koob, 2020; Sakloth et al., 2020).
At present, we do not know how L-CYSee prevents the development of physical dependence on morphine. The mechanisms by which L-thiol esters exert their biological effects are likely to be multi-factorial and possibly include 1) direct binding of L-CYSee to plasma membrane/intracellular proteins, such as ion channels, receptors, and enzymes that alter the activities of the proteins by mechanisms not associated with the changes in the redox status of the proteins; 2) formation of thiol adducts, such as D-glucose-L-cysteine (Wróbel et al., 1997; Szwergold, 2006; Li et al., 2015) and mixed disulfides (Wilcken and Gupta, 1979; Lash and Jones, 1985; Turell, et al., 2013) in the blood; 3) modulation of redox status (e.g., reduction in L-cystine to L-cysteine), and the activity of plasma membrane proteins, such as Kv1.2 K+-channels (Baronas et al., 2017), and after entry into cells, redox modulation of functional intracellular proteins (Bogeski et al., 2011; Bogeski and Niemeyer, 2014; O-Uchi et al., 2014; Gamper and Ooi, 2015; Gao et al., 2017; Garcia et al., 2018); 4) formation of S-thiolated proteins, such as S-cysteinylated, S-cysteinylglycinylated, and S-glutathionylated proteins, in plasma membranes of cells (Winkler et al., 2007; Rossi et al., 2009; Auclair et al., 2013; Belcastro et al., 2017; Ghezzi and Chan, 2017; Bonifácio et al., 2021); 5) conversion of L-CYSee to L-cysteine by membrane-associated esterases (Butterworth et al., 1993; Nishida et al., 1996), which then enters into multiple metabolic pathways including those that generate hydrogen sulfide via the sequential actions of L-cysteine aminotransferase and cystathionine γ-lyase in peripheral and central tissues (Kimura, 2014; Kimura, 2017; Bełtowski, 2019), including the carotid bodies (Prabhakar, 2012); 6) conversion of L-thiol esters to cysteine sulfenic, sulfinic, and sulfonic via cysteine dioxygenase (Yamaguchi and Hosokawa, 1987; Joseph and Maroney, 2007; Stipanuk et al., 2009; Stipanuk et al., 2011); and 7) formation of S-nitroso-L-cysteine, an endogenous S-nitrosothiol (Myers et al., 1990; Bates et al., 1991; Seckler et al., 2020), with many substantial roles in intracellular signaling cascades (Lipton et al., 1993; Foster et al., 2009; Seth and Stamler, 2011; Stomberski et al., 2019; Gaston et al., 2020), including those controlling the cardiorespiratory function (Davisson et al., 1996; Davisson et al., 1997; Ohta et al., 1997; Lipton et al., 2001; Gaston et al., 2006; Lewis et al., 2006; Gaston et al., 2020) and those involved in the attenuation of opioid induced respiratory depression (OIRD) (Getsy et al., 2022a; Getsy et al., 2022b). Any or all of these mechanisms (and possibly those not mentioned) may interact with signaling pathways involved in the acquisition of physical dependence on opioids, such as morphine, and the expression of the NLX-precipitated withdrawal syndrome, including those involving N-methyl D-aspartate (NMDA) glutamatergic receptors (Buccafusco et al., 1995; Herman et al., 1995; Rasmussen, 1995; Noda and Nabeshima, 2004; Glass, 2011; Fluyau et al., 2020), muscarinic receptors (Marshall and Buccafusco, 1987; Holland et al., 1993), corticotropin-releasing factor (CRF) receptor (CRF1) (García-Carmona et al., 2015), tachykinin receptors (Michaud and Couture, 2003), voltage-gated Ca2+-channels (Tokuyama et al., 1995; Dogrul et al., 2002; Esmaeili-Mahani et al., 2008; Alboghobeish et al., 2019), adenylyl cyclase superactivation and opioid receptor phosphorylation (Avido-Reiss et al., 1996; Avido-Reiss et al., 1997; Wang et al., 1999; Eckhardt et al., 2000), oxidative stress (Xu et al., 2006; Mori et al., 2007; Abdel-Zaher et al., 2013; Mansouri et al., 2020; Ward et al., 2020; Houshmand et al., 2021), and the nitric oxide-cGMP signaling cascade (Adams et al., 1993; Cappendijk et al., 1993; Majeed et al., 1994; Vaupel et al., 1995a; Buccafusco et al., 1995; Vaupel et al., 1995b; Herman et al., 1995; Leza et al., 1995; 1996; London et al., 1995; Dambisya and Lee, 1996; Bhatt and Kumar, 2015; Tsakova et al., 2015; Sackner et al., 2019; Gledhill and Babey, 2021). Since L-CYSee blunted the expression of all NLX-precipitated behavioral (except for sneezing), physical (body weight loss and hypothermia), and cardiorespiratory (hypertension, tachycardia, and incidence of apneas) phenomena, it is tempting to assume that L-CYSee interrupts fundamental intracellular processes that are essential for the development of physical dependence on morphine.
The second set of novel findings was that the introduction of L-CYSee infusion 36 h into the morphine administration period appeared to reverse the established physical dependence on the opioid, as assessed at 48 h (i.e., within 12 h of giving continuous infusion of L-CYSee). Specifically, NLX-precipitated behavioral phenomena (except for sneezing), hypertension, tachycardia, apneas, hypothermia, and body weight loss were markedly fewer in the rats that had received L-CYSee for 12 h. Again, the lack of effect of L-cysteine and L-SERee suggests that the intracellular delivery of L-CYSee and its sulfur atom (and associated thiol chemistry) is essential to the ability of the L-thiol ester to reverse the established physical dependence on morphine. Again, the mechanism of action for how L-CYSee reverses physical dependence on morphine is not known, but any of the mechanisms discussed in the previous paragraph, including its potent antioxidant properties, may be involved. The well-known therapeutics and bioactive compounds that reverse established physical dependence include: L-histidine and certain histamine receptor sub-type agonists (Wong and Roberts, 1976), such as melatonin (Raghavendra and Kulkarni SK, 1999; Raghavendra and Kulkarni SK, 2000); the antioxidant, quercetin (Singh et al., 2002; Naidu et al., 2003); the serotonin-reuptake inhibitor, fluoxetine (Singh et al., 2003); the nitric oxide synthase inhibitor, L-NG-nitroarginine methyl ester (Naidu et al., 2003; Sing et al., 2003); inhibitors of Ca2+/calmodulin-dependent protein kinase II (Wang et al., 2003; Tang et al., 2006); the β2-AR antagonist, butoxamine (Liang et al., 2007); adrenomedullin receptor antagonists (Wang et al., 2011); the antipsychotic (dopamine D2 receptor antagonist), haloperidol (Yang et al., 2011); and positive allosteric modulators of AMPA (α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid) glutamatergic receptors (Hu et al., 2018). The mechanism(s) of action for how L-CYSee reverses established physical dependence on morphine is of great clinical relevance, and opens the way for future studies on this and other bioactive L,D-thiol esters and related compounds (Baby et al., 2021; Baby et al., 2021; Gaston et al., 2021; Jenkins et al., 2021; Getsy et al., 2022a; Getsy et al., 2022b; Getsy et al., 2022c; Getsy et al., 2022d; Getsy et al., 2022e; Getsy et al., 2022f; Lewis et al., 2022) with respect to their ability to reverse physical dependence on morphine and other opioids, including heroin and fentanyl.
Many questions remain regarding the potential use of L-thiol esters as therapeutics for key clinical problems associated with opioid analgesia in humans, including: 1) if L-CYSee attenuates/blocks self-administration of opioids in OUD patients, adding it to prescription opioids may result in lower drug abuse or addiction potential; 2) if L-CYSee attenuates or blocks development of physical dependence on opioids, then adding it to prescription opioids will minimize and may potentially eliminate physical dependence in individuals who receive opioids for a long-term basis, such as everyday, all day, for weeks/months; 3) if L-CYSee attenuates/blocks tachyphylaxis to opioid analgesia or hyperalgesia caused by opioids, then the addition of L-CYSee to prescription opioids will maintain their analgesic efficacy over long periods of time, eliminating the development of tolerance, need for escalating doses, and potential complications of hyperalgesia; 4) if L-CYSee has several of the advantageous effects observed in rodents, then adding it to opioid analgesics would multiply the beneficial aspects of the opioids; 5) if L-CYSee prevents the development of physical dependence, and specifically, if it is introduced to an individual with physical dependence and attenuates/blocks opioid withdrawal, it could be used as an outpatient/inpatient medication to manage opioid withdrawal in those who are iatrogenically physically dependent (long-term opioid prescriptions) or those who are addicted and physically dependent; 6) if L-CYSee attenuates/blocks euphoria and/or the development of physiological dependence to opioids, then it would be a good medication for medication-assisted treatment (MAT) and a potentially good drug for harm reduction interventions in people with OUD who are not interested in the psychosocial aspects of counseling and treatment; 7) as some patients with a history of OUD who are currently sober need opioids for the treatment of acute or chronic pain syndromes, this L-thiol ester, if it attenuates or blocks euphoria and physical dependence, could be added to opioid analgesics when given to people with a history of OUD, thereby eliminating the risk of opioid analgesic-precipitating euphoria, drug cravings, and their markedly increased risk of relapse; 8) if L-CYSee attenuates/blocks euphoria from chemically mediated dopamine surges within the ventral tegmentum, nucleus accumbens, or medial prefrontal cortex, where brain rewarding euphoria-producing dopamine surge happens from all drugs of abuse/addiction (Deslandes et al., 2002; Gardner, 2011; Koob and Volkow, 2016; Volkow et al., 2019; Koob, 2020; Sakloth et al., 2020), then it will be useful in the treatment of OUD and other SUDs; and 9) if L-CYSee attenuates/blocks euphoria from chemically mediated dopamine surges, it could be combined with or added to all controlled prescription drugs, resulting in an abuse-resistant or non-abusable form of prescribed opioids, benzodiazepines, and psychostimulants. In relation to point (1), we recently showed that co-administration of the D-isomer, D-cysteine ethyl ester, with fentanyl prevents the development of fentanyl-induced conditioned place preference in both male and female rats (Knauss et al., 2023). Thus, L,D-thiol esters likely reduce the rewarding properties of opioids and reduce their addictive potential.
A final caveat to translating these findings to humans lies in the genetic variation of humans (e.g., 5–6 million SNPs between any two individuals) in comparison to the total variation found in the outbred Sprague Dawley rats tested here (Gileta et al., 2022). This lack of genetic variation in preclinical models is one explanation for the inability of many findings to translate across species (Garner, 2014; Zuberi and Lutz, 2016). The pharmacogenetics of opioids have been studied, and numerous pharmacokinetic molecules, such as COMT, OPRM1, CYP2D6, and ABCB1, have been identified that are useful in clinical application (Owusu Obeng et al., 2017). In animal models, opioid withdrawal specifically has been studied using the C57BL/6J x DBA2 recombinant inbred lines, where a locus on chromosome 16 was identified as affecting the locomotor behavior of naloxone-precipitated withdrawal (Philip et al., 2010). In another population, 129 × C57BL/6 cross-identified loci on chromosomes 1, 5, and 10 were involved in withdrawal jumping frequencies (Kest et al., 2004). Testing the efficacy of drugs using a population of outbred mice containing approximately 45 million segregating SNPs, such as the diversity outbred mice (Saul et al., 2019), will increase the likelihood that the drug will translate within species before traversing across species.
Study limitations
A limitation of the present study is that we have not examined the efficacy of lower doses of L-CYSee to prevent or reverse morphine-induced physical dependence. Finding out the lower limit is key to minimizing potential adverse biological effects that were not monitored in the present study. In particular, we have not determined whether the co-administration of L-CYSee alters the analgesic actions of morphine, although we have reported that L-CYSee (Lewis et al., 2022), L-cysteine methyl ester (Getsy et al., 2022a), and other thiolesters and related compounds (Baby et al., 2021; Baby et al., 2021; Gaston et al., 2021; Jenkins et al., 2021; Getsy et al., 2022b; Getsy et al., 2022c; Getsy et al., 2022d; Getsy et al., 2022e; Getsy et al., 2022f) prevent and/or reverse the actions of morphine and fentanyl on ventilatory parameters, arterial blood–gas chemistry, and alveolar-arterial gradient in freely-moving rats without compromising opioid-induced analgesia or sedation. Synthetic opioids, especially fentanyl, are playing a major and ever-increasing role in the current opioid crisis (Arendt, 2021; Deo et al., (2021), and future studies must determine whether L-CYSee can overcome and reverse physical dependence on fentanyl. A concern about the study protocol regarding the ability of L-CYSee to reverse established morphine dependence, was the short time (12 h) between the surgery to initiate the intravenous infusions (implantation of minipumps under isoflurane anesthesia), and the administration of NLX to precipitate withdrawal. Despite the robust nature of the NLX-induced responses in rats that received intravenous infusions of vehicle, L-cysteine, or L-SERee for 12 h, it is possible that the lingering effects of anesthesia affected the expression of the withdrawal phenomena. Another important limitation of our study is the lack of data about the efficacy of L-CYSee in preventing/reversing physical dependence in female rats. This is especially important because 1) opioids exert qualitatively and quantitatively different pharmacological (e.g., ventilation, analgesia) responses in females compared to males (Dahan et al., 1998; Sarton et al., 1998; Bodnar and Kest, 2010); 2) there are many sex-specific differences in opioid receptor signaling (Bryant et al., 2006; Hosseini et al., 2011); 3) pronounced sex differences in development of opioid hyperalgesia, tolerance and withdrawal (Bodnar and Kest, 2010); and 4) several major behavioral sex differences in the expression and treatment of OUDs (Huhn et al., 2019; Davis et al., 2021; Knouse and Briand, 2021). The lack of understanding about the molecular mechanisms by which L-CYSee affects the acquisition/reversal of morphine dependence is a limitation that needs to be addressed. In addition to potential direct interactions with yet to be defined functional proteins, potential mechanisms of action of L-CYSee may involve 1) direct binding to putative L,D-cysteine-binding protein, such as myristoylated alanine-rich C-kinase substrate (Semenza et al., 2021), 2) interruption of μ-OR-β-arrestin-coupled cell signaling processes to spare the antinociceptive G-protein-dependent actions of morphine (Schmid et al., 2017; Grim et al., 2020), and/or 3) potential conversion of L-CYSee to S-nitroso-L-CYSee or S-nitroso-L-cysteine by S-nitrosylation of the sulfur atom in L-thiol esters via processes requiring nitric oxide synthase (Perissinotti et al., 2005; Hess and Stamler, 2012; Stomberski et al., 2019; Seckler et al., 2020; Seckler et al., 2022), which may act via an intracellular penetrating mechanism(s) (Clancy et al., 2001). To test the possibility that L-CYSee elicits the production of S-nitrosylated versions of the L-thiol ester, we are determining whether intravenous injections of L-CYSee increase the production of S-nitrosylated species in the blood, peripheral tissues, and brain via the use of an ultra-sensitive capacitive sensor (Seckler et al., 2017), and whether such injections of L-CYSee increase the expression of NADPH diaphorase in the brain and peripheral structures, on the basis that NADPH diaphorase is used to visualize free S-nitrosothiols and S-nitrosylated proteins in aldehyde-treated tissue (Seckler et al., 2020). S-nitrosothiols, such as S-nitroso-L-cysteine and S-nitroso-L-glutathione, play important roles in ventilatory control processes in the brainstem, circulating red blood cells, and peripheral structures, such as the carotid bodies (Lipton et al., 2001; Gaston et al., 2006; Palmer et al., 2013; Gaston et al., 2014; Palmer et al., 2015; Gaston et al., 2020). The possibility that the conversion of L-CYSee to S-nitroso-L-CYSee is responsible for the effects observed in this study would add to our understanding of the pharmacology of L–S-nitrosothiols (Davisson et al., 1996; Lewis et al., 1996; Travis et al., 1996; Davisson et al., 1997; Travis et al., 1997; Lewis et al., 2005; Gaston et al., 2006; Lewis et al., 2006; Gaston et al., 2020). Another limitation that certainly needs addressing is our lack of information about the blood and tissue distribution resulting from the infusion of L-CYSee in the presence or absence of morphine, although L-CYSee can be readily detected in plasma, and peripheral and central tissues upon acute administration to naïve rats (Servin et al., 1998). We intend to perform pharmacokinetics analyses of L-CYSee distribution in brain regions involved in the acquisition of opioid dependence, such as the medial prefrontal cortex (Deslandes et al., 2002; Gardner, 2011; Koob and Volkow, 2016; Volkow et al., 2019; Koob, 2020; Sakloth et al., 2020), using our liquid chromatography–mass spectrometry methodology (Altawallbeh et al., 2019).
Conclusion
This study demonstrates that systemic infusion of the membrane-permeable L-thiol ester, L-CYSee, prevents the development of physical dependence on morphine in male Sprague Dawley rats by mechanisms dependent on thiol biochemistry. In addition, this study demonstrates that L-CYSee reverses established dependence on morphine in rats also by thiol-dependent processes. Delineating the exact thiol-dependent signaling pathways will add greatly to our understanding of the processes by which opioids induce dependence, and how the bioactive L-thiol esters exert their effects. Our study was spurred by the ground-breaking work of Trivedi, Deth, and others, which added greatly to our understanding of the mechanisms that opioids cause physical dependence and psychological addiction (Trivedi et al., 2014; Trivedi and Deth, 2015; Trivedi et al., 2015). Their evidence that morphine may cause dependence/addiction by blocking the entry of L-cysteine into neurons inhibiting the EAA3/EAAC1 transporters (Trivedi et al., 2014) prompted our studies with the membrane-permeable, L-thiol ester, L-CYSee. The findings that L-CYSee markedly reduced the majority of NLX-precipitated withdrawal phenomena suggests that decreased levels of L-cysteine entry into cells plays a key role in establishing physical dependence on morphine. The lone withdrawal phenomenon that was not ameliorated by L-CYSee was sneezing, a key feature of the opioid withdrawal response in humans (Ostrea et al., 1975; Specker et al., 1998; Gaalema et al., 2012; Lofwall et al., 2013) and experimental animals (Hendrie, 1985; Liu et al., 2007; Singh et al., 2015). We are currently trying to understand the current state of knowledge about the neural mechanisms responsible for sneezing (Batsel and Lines, 1975; Undem et al., 2000; Li et al., 2021; Ramirez et al., 2022) to see if that can give insights into the signaling pathways that are/are not involved in the actions of L-CYSee. The present findings add to our knowledge about the efficacy of L,D-thiol esters, such as L-CYSee (Lewis et al., 2022) L-GSHee (Jenkins et al., 2021), D-CYSee (Getsy et al., 2022c; Getsy et al., 2022d), D-cystine di(m)ethyl ester (Gaston et al., 2021), and the free radical-superoxide anion scavenger, Tempol (Baby et al., 2021a; Baby et al., 2021b), on the pharmacological actions of opioids.
Data availability statement
The raw data supporting the conclusion of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was approved by the Animal Care and Use Committees of Galleon Pharmaceuticals. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
JmB: conceptualization, data curation, investigation, supervision, validation, visualization, writing–original draft, and writing–review and editing. PG: data curation, formal analysis, investigation, methodology, project administration, validation, visualization, and writing–review and editing. GC: data curation, formal analysis, investigation, methodology, validation, and writing–review and editing. SB: conceptualization, data curation, formal analysis, investigation, project administration, visualization, writing–original draft, and writing–review and editing. PM: conceptualization, data curation, formal Analysis, validation, visualization, writing–original draft, and writing–review and editing. Y-HH: data curation, formal analysis, investigation, validation, visualization, writing–original draft, and writing–review and editing. ZK: data curation, formal analysis, validation, visualization, writing–original draft, and writing–review and editing. JsB: validation, visualization, writing–original draft, and writing–review and editing. DM: data curation, supervision, validation, visualization, writing–original draft, and writing–review and editing. SL: conceptualization, data curation, formal Analysis, funding acquisition, investigation, methodology, project administration, resources, supervision, validation, visualization, writing–original draft, and writing–review and editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. These studies were funded partially by an NIH/NIDA grant (U01DA051373, Optimization of Novel Thiolesters as a Therapeutic Strategy for Combating Opioid Overdoses and Abuse) to SL. In addition, JsB received funding from DA059060 and DA048890. SL received funding from Galleon Pharmaceuticals, Inc who were not involved in study design, collection, analysis, interpretation of data, writing of this article and the decision to submit it for publication.
Acknowledgments
The authors would like to thank the staff at the Animal Care Facilities at the University of Virginia and Case Western Reserve University for their expert and caring technical assistance in helping with the rats and individual experiments (scoring of withdrawal phenomena).
Conflict of interest
Author SB was employed by Galleon Pharmaceuticals, Inc.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphar.2023.1303207/full#supplementary-material
Abbreviations
AMPA, α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid; CRF, corticotropin-releasing factor; EAA3, excitatory amino acid transporter; IP, intraperitoneal; IV, intravenous; L-CYSee, L-cysteine ethyl ester; L-NAC, N-acetyl-L-cysteine; Kv1.2 K+-channels, voltage-gated K+1.2 channels; L-SERee, L-serine ethyl ester; μ-OR, μ-opioid receptor; MAP, mean arterial blood pressure; NMDA, N-methyl D-aspartate; NLX, naloxone hydrochloride; OIRD, opioid-induced respiratory depression; OUD, opioid use disorder; SAM, S-adenosyl-methionine; SAH, S-adenosyl-homocysteine; SC, subcutaneous; SUD, substance use disorder.
References
Abdel-Zaher, A. O., Mostafa, M. G., Farghaly, H. S., Hamdy, M. M., and Abdel-Hady, R. H. (2013). Role of oxidative stress and inducible nitric oxide synthase in morphine-induced tolerance and dependence in mice. Effect of alpha-lipoic acid. Behav. Brain Res. 247, 17–26. doi:10.1016/j.bbr.2013.02.034
Adams, M. L., Kalicki, J. M., Meyer, E. R., and Cicero, T. J. (1993). Inhibition of the morphine withdrawal syndrome by a nitric oxide synthase inhibitor, NG-nitro-L-arginine methyl ester. Life Sci. 52, PL245–PL249. doi:10.1016/0024-3205(93)90472-f
Alboghobeish, S., Naghizadeh, B., Kheirollah, A., Ghorbanzadeh, B., and Mansouri, M. T. (2019). Fluoxetine increases analgesic effects of morphine, prevents development of morphine tolerance and dependence through the modulation of L-type calcium channels expression in mice. Behav. Brain Res. 361, 86–94. doi:10.1016/j.bbr.2018.12.020
Altawallbeh, G., Smith, L., Lewis, S. J., Authier, S., Bujold, K., Gaston, B., et al. (2019). Pharmacokinetic study of Sudaxine in dog plasma using novel LC-MS/MS method. Drug Test. Anal. 11, 403–410. doi:10.1002/dta.2507
Alvarez-Perez, B., Poras, H., and Maldonado, R. (2023). The inhibition of enkephalin catabolism by dual enkephalinase inhibitor: a novel possible therapeutic approach for opioid use disorders. Br. J. Pharmacol. 180, 879–893. doi:10.1111/bph.15656
Arendt, F. (2021). The opioid-overdose crisis and fentanyl: the role of online information seeking via internet search engines. Health Commun. 36, 1148–1154. doi:10.1080/10410236.2020.1748820
Arias, J. M., Díaz, S. B., Ben Altabef, A., and Dupuy, F. G. (2019). Interaction of cysteine and its derivatives with monolayers of dipalmitoylphosphatidylcholine. Colloids Surf. B Biointerfaces 184, 110548. doi:10.1016/j.colsurfb.2019.110548
Auclair, J. R., Brodkin, H. R., D'Aquino, J. A., Petsko, G. A., Ringe, D., and Agar, J. N. (2013). Structural consequences of cysteinylation of Cu/Zn-superoxide dismutase. Biochemistry 52, 6145–6150. doi:10.1021/bi400613h
Avidor-Reiss, T., Nevo, I., Levy, R., Pfeuffer, T., and Vogel, Z. (1996). Chronic opioid treatment induces adenylyl cyclase V superactivation. Involvement of Gbetagamma. J. Biol. Chem. 271, 21309–21315. doi:10.1074/jbc.271.35.21309
Avidor-Reiss, T., Nevo, I., Saya, D., Bayewitch, M., and Vogel, Z. (1997). Opiate-induced adenylyl cyclase superactivation is isozyme-specific. J. Biol. Chem. 272, 5040–5047. doi:10.1074/jbc.272.8.5040
Baby, M. S., Discala, F. G., Gruber, R., Getsy, M. P., Cheng, F., Damron, S. D., et al. (2021a). Tempol reverses the negative effects of morphine on arterial blood-gas chemistry and tissue oxygen saturation in freely-moving rats. Front. Pharmacol. 12, 749084. doi:10.3389/fphar.2021.749084
Baby, S. M., Gruber, R., Discala, J., Puskovic, V., Jose, N., Cheng, F., et al. (2021b). Systemic administration of Tempol attenuates the cardiorespiratory depressant effects of fentanyl. Front. Pharmacol. 12, 690407. doi:10.3389/fphar.2021.690407
Balanaser, M., Carley, M., Baron, R., Finnerup, N. B., Moore, R. A., Rowbotham, M. C., et al. (2023). Combination pharmacotherapy for the treatment of neuropathic pain in adults: systematic review and meta-analysis. Pain 164, 230–251. doi:10.1097/j.pain.0000000000002688
Baldo, B. A. (2022). Current research in pathophysiology of opioid-induced respiratory depression, neonatal opioid withdrawal syndrome, and neonatal antidepressant exposure syndrome. Curr. Res. Toxicol. 3, 100078. doi:10.1016/j.crtox.2022.100078
Balshaw, R., Rowe, A., Cochrane, C. K., Ng, B., Jiang, A., Risi, A., et al. (2021). Comparison of rates of opioid withdrawal symptoms and reversal of opioid toxicity in patients treated with two naloxone dosing regimens: a retrospective cohort study. Clin. Toxicol. (Phila). 59, 38–46. doi:10.1080/15563650.2020.1758325
Baraban, S. C., Stornetta, R. L., and Guyenet, P. G. (1993). Respiratory control of sympathetic nerve activity during naloxone-precipitated morphine withdrawal in rats. J. Pharmacol. Exp. Ther. 265, 89–95.
Baronas, V. A., Yang, R. Y., and Kurata, H. T. (2017). Extracellular redox sensitivity of Kv1.2 potassium channels. Sci. Rep. 7, 9142. doi:10.1038/s41598-017-08718-z
Bates, J. N., Harrison, D. G., Myers, P. R., and Minor, R. L. (1991). EDRF: nitrosylated compound or authentic nitric oxide. Basic Res. Cardiol. 86, 17–26. doi:10.1007/978-3-642-72461-9_3
Batsel, H. L., and Lines, A. J. (1975). Neural mechanisms of sneeze. Am. J. Physiol. 229, 770–776. doi:10.1152/ajplegacy.1975.229.3.770
Bechara, A., Berridge, K. C., Bickel, W. K., Morón, J. A., Williams, S. B., and Stein, J. S. (2019). A neurobehavioral approach to addiction: implications for the opioid epidemic and the psychology of addiction. Psychol. Sci. Public Interest 20, 96–127. doi:10.1177/1529100619860513
Belcastro, E., Gaucher, C., Corti, A., Leroy, P., Lartaud, I., and Pompella, A. (2017). Regulation of protein function by S-nitrosation and S-glutathionylation: processes and targets in cardiovascular pathophysiology. Biol. Chem. 398, 1267–1293. doi:10.1515/hsz-2017-0150
Bełtowski, J. (2019). Synthesis, metabolism, and signaling mechanisms of hydrogen sulfide: an overview. Methods Mol. Biol. 2007, 1–8. doi:10.1007/978-1-4939-9528-8_1
Benyamin, R., Trescot, A. M., Datta, S., Buenaventura, R., Adlaka, R., Sehgal, N., et al. (2008). Opioid complications and side effects. Pain Physician 11 (2 Suppl. l), S105–S120. doi:10.36076/ppj.2008/11/s105
Bhatt, K., and Kumar, A. (2015). Mechanism of morphine addiction by inhibiting the soluble Guanylate Cyclase-Nitric Oxide (sGC-NO) pathway. Math. Biosci. 266, 85–92. doi:10.1016/j.mbs.2015.06.004
Biancuzzi, H., Dal Mas, F., Brescia, V., Campostrini, S., Cascella, M., Cuomo, A., et al. (2023). Opioid misuse: a review of the Main issues, challenges, and strategies. Int. J. Environ. Res. Public Health 19, 11754. doi:10.3390/ijerph191811754
Bodnar, R. J., and Kest, B. (2010). Sex differences in opioid analgesia, hyperalgesia, tolerance and withdrawal: central mechanisms of action and roles of gonadal hormones. Horm. Behav. 58, 72–81. doi:10.1016/j.yhbeh.2009.09.012
Bogeski, I., Kappl, R., Kummerow, C., Gulaboski, R., Hoth, M., and Niemeyer, B. A. (2011). Redox regulation of calcium ion channels: chemical and physiological aspects. Cell Calcium 50, 407–423. doi:10.1016/j.ceca.2011.07.006
Bogeski, I., and Niemeyer, B. A. (2014). Redox regulation of ion channels. Antioxid. Redox Signal. 21, 859–862. doi:10.1089/ars.2014.6019
Bonifácio, V. D. B., Pereira, S. A., Serpa, J., and Vicente, J. B. (2021). Cysteine metabolic circuitries: druggable targets in cancer. Br. J. Cancer 124, 862–879. doi:10.1038/s41416-020-01156-1
Brognara, F., Dias, D. P., Castania, J. A., Fazan, R., Lewis, S. J., and Salgado, H. C. (2016). Cardiovascular responses elicited by continuous versus intermittent electrical stimulation of the aortic depressor nerve in conscious rats. Life Sci. 148, 99–105. doi:10.1016/j.lfs.2016.02.033
Browne, C. J., Godino, A., Salery, M., and Nestler, E. J. (2020). Epigenetic mechanisms of opioid addiction. Biol. Psychiatry 87, 22–33. doi:10.1016/j.biopsych.2019.06.027
Bryant, C. D., Eitan, S., Sinchak, K., Fanselow, M. S., and Evans, C. J. (2006). NMDA receptor antagonism disrupts the development of morphine analgesic tolerance in male, but not female C57BL/6J mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 291, R315–R326. doi:10.1152/ajpregu.00831.2005
Buccafusco, J. J. (1983). Cardiovascular changes during morphine withdrawal in the rat: effects of clonidine. Pharmacol. Biochem. Behav. 18, 209–215. doi:10.1016/0091-3057(83)90365-9
Buccafusco, J. J. (1990). Participation of different brain regions in the anti-narcotic withdrawal action of clonidine in the dependent rat. Brain Res. 513, 8–14. doi:10.1016/0006-8993(90)91083-s
Buccafusco, J. J., Marshall, D. C., and Turner, R. M. (1984). A comparison of the inhibitory effects of clonidine and guanfacine on the behavioral and autonomic components of morphine withdrawal in rats. Life Sci. 35, 1401–1408. doi:10.1016/0024-3205(84)90398-9
Buccafusco, J. J., Terry, A. V., and Shuster, L. (1995). Spinal NMDA receptor - nitric oxide mediation of the expression of morphine withdrawal symptoms in the rat. Brain Res. 679, 189–199. doi:10.1016/0006-8993(95)00203-3
Butterworth, M., Upshall, D. G., and Cohen, G. M. (1993). A novel role for carboxylesterase in the elevation of cellular cysteine by esters of cysteine. Biochem. Pharmacol. 46, 1131–1137. doi:10.1016/0006-2952(93)90460-e
Cappendijk, S. L., de Vries, R., and Dzoljic, M. R. (1993). Inhibitory effect of nitric oxide (NO) synthase inhibitors on naloxone-precipitated withdrawal syndrome in morphine-dependent mice. Neurosci. Lett. 162, 97–100. doi:10.1016/0304-3940(93)90569-7
Chang, A. P., and Dixon, W. R. (1990). Role of plasma catecholamines in eliciting cardiovascular changes seen during naloxone-precipitated withdrawal in conscious, unrestrained morphine-dependent rats. J. Pharmacol. Exp. Ther. 254, 857–863.
Chartoff, E. H., and Connery, H. S. (2014). It's MORe exciting than mu: crosstalk between mu opioid receptors and glutamatergic transmission in the mesolimbic dopamine system. Front. Pharmacol. 5, 116. doi:10.3389/fphar.2014.00116
Christie, M. J. (2008). Cellular neuroadaptations to chronic opioids: tolerance, withdrawal and addiction. Brit J. Pharmacol. 154, 384–396. doi:10.1038/bjp.2008.100
Clancy, R., Cederbaum, A. I., and Stoyanovsky, D. A. (2001). Preparation and properties of S-nitroso-L-cysteine ethyl ester, an intracellular nitrosating agent. J. Med. Chem. 44, 2035–2038. doi:10.1021/jm000463f
Collier, H. O. J., Francis, D. L., and Schneider, C. (1972). Modification of morphine withdrawal by drugs interacting with humoral mechanisms: some contradictions and their interpretation. Nature 237, 220–223. doi:10.1038/237220a0
Dahan, A., Sarton, E., Teppema, L., and Olievier, C. (1998). Sex-related differences in the influence of morphine on ventilatory control in humans. Anesthesiology 88, 903–913. doi:10.1097/00000542-199804000-00009
Daijo, H., Kai, S., Tanaka, T., Wakamatsu, T., Kishimoto, S., Suzuki, K., et al. (2011). Fentanyl activates hypoxia-inducible factor 1 in neuronal SH-SY5Y cells and mice under non-hypoxic conditions in a μ-opioid receptor-dependent manner. Eur. J. Pharmacol. 667, 144–152. doi:10.1016/j.ejphar.2011.06.014
Dambisya, Y. M., and Lee, T. L. (1996). Role of nitric oxide in the induction and expression of morphine tolerance and dependence in mice. Br. J. Pharmacol. 117, 914–918. doi:10.1111/j.1476-5381.1996.tb15280.x
Davis, J. P., Eddie, D., Prindle, J., Dworkin, E. R., Christie, N. C., Saba, S., et al. (2021). Sex differences in factors predicting post-treatment opioid use. Addiction 116, 2116–2126. doi:10.1111/add.15396
Davisson, R. L., Bates, J. N., Johnson, A. K., and Lewis, S. J. (2014). Effects of intracerebroventricular injections of 5-HT on systemic vascular resistances of conscious rats. Microvasc. Res. 95C, 116–123. doi:10.1016/j.mvr.2014.08.002
Davisson, R. L., Travis, M. D., Bates, J. N., Johnson, A. K., and Lewis, S. J. (1997). Stereoselective actions of S-nitrosocysteine in central nervous system of conscious rats. Am. J. Physiol. 272, H2361–H2368. doi:10.1152/ajpheart.1997.272.5.H2361
Davisson, R. L., Travis, M. D., Bates, J. N., and Lewis, S. J. (1996). Hemodynamic effects of L- and D-S-nitrosocysteine in the rat. Stereoselective S-nitrosothiol recognition sites. Circ. Res. 79, 256–262. doi:10.1161/01.res.79.2.256
Defonsi Lestard, M. E., Díaz, S. B., Puiatti, M., Echeverría, G. A., Piro, O. E., Pierini, A. B., et al. (2013). Vibrational and structural behavior of (L)-cysteine ethyl ester hydrochloride in the solid state and in aqueous solution. J. Phys. Chem. A 117, 14243–14252. doi:10.1021/jp409252d
Delle, M., Ricksten, S. E., Häggendal, J., Olsson, K., Skarphedinsson, J. O., and Thorén, P. (1990). Regional changes in sympathetic nerve activity and baroreceptor reflex function and arterial plasma levels of catecholamines, renin and vasopressin during naloxone-precipitated morphine withdrawal in rats. J. Pharmacol. Exp. Ther. 253, 646–654.
Deo, V. S., Gilson, T. P., Kaspar, C., and Singer, M. E. (2021). The fentanyl phase of the opioid epidemic in Cuyahoga County, Ohio, United States. J. Forensic Sci. 66, 926–933. doi:10.1111/1556-4029.14665
Deslandes, P. N., Pache, D. M., and Sewell, R. D. (2002). Drug dependence: neuropharmacology and management. J. Pharm. Pharmacol. 54, 885–895. doi:10.1211/002235702760088992
Ding, H., and Demple, B. (1998). Thiol-mediated disassembly and reassembly of [2Fe-2S] clusters in the redox-regulated transcription factor SoxR. Biochem. 37, 17280–17286. doi:10.1021/bi980532g
Dixon, W. R., and Chang, A. P. (1988). Effect of phentolamine on blood pressure, heart rate and plasma catecholamine levels in conscious, unrestrained morphine dependent rats during naloxone precipitated autonomic withdrawal responses. Proc. West Pharmacol. Soc. 31, 117–119.
Dogrul, A., Zagli, U., and Tulunay, F. C. (2002). The role of T-type calcium channels in morphine analgesia, development of antinociceptive tolerance and dependence to morphine, and morphine abstinence syndrome. Life Sci. 71, 725–734. doi:10.1016/s0024-3205(02)01736-8
Dydyk, A. M., Sizemore, D. C., Haddad, L. M., Lindsay, L., and Porter, B. R. (2023). “NP safe prescribing of controlled substances while avoiding drug diversion,” in StatPearls (Treasure Island (FL): StatPearls Publishing). PMID: 33232099.
Dydyk, A. M., Sizemore, D. C., Smock, W., Dulebohn, S. C., and Porter, B. R. (2022). “Kentucky KASPER and controlled substance prescribing,” in StatPearls (Treasure Island (FL): StatPearls Publishing). PMID: 33620803.
Eckhardt, K., Nevo, I., Levy, R., Mikus, G., Eichelbaum, M., and Vogel, Z. (2000). Morphine-related metabolites differentially activate adenylyl cyclase isozymes after acute and chronic administration. FEBS Lett. 470, 309–314. doi:10.1016/s0014-5793(00)01329-6
Esmaeili-Mahani, S., Fathi, Y., Motamedi, F., Hosseinpanah, F., and Ahmadiani, A. (2008). L-type calcium channel blockade attenuates morphine withdrawal: in vivo interaction between L-type calcium channels and corticosterone. Horm. Behav. 53, 351–357. doi:10.1016/j.yhbeh.2007.10.012
Fluyau, D., Revadigar, N., and Pierre, C. G. (2020). Clinical benefits and risks of N-methyl-d-aspartate receptor antagonists to treat severe opioid use disorder: a systematic review. Drug Alcohol Depend. 208, 107845. doi:10.1016/j.drugalcdep.2020.107845
Foster, M. W., Hess, D. T., and Stamler, J. S. (2009). Protein S-nitrosylation in health and disease: a current perspective. Trends Mol. Med. 15, 391–404. doi:10.1016/j.molmed.2009.06.007
Frankeberger, J., Jarlenski, M., Krans, E. E., Coulter, R. W. S., and Mair, C. (2023). Opioid use disorder and overdose in the first year postpartum: a rapid scoping review and implications for future research. Matern. Child. Health J. 27, 1140–1155. doi:10.1007/s10995-023-03614-7
Fukui, K., Kaneda, M., Takahashi, E., Washio, M., and Doi, K. (1994). Protective effects of sulfhydryl compounds on HOCl-induced intracellular Ca2+ increase in single rat ventricular myocytes. J. Mol. Cell Cardiol. 26, 455–461. doi:10.1006/jmcc.1994.1056
Gaalema, D. E., Scott, T. L., Heil, S. H., Coyle, M. G., Kaltenbach, K., Badger, G. J., et al. (2012). Differences in the profile of neonatal abstinence syndrome signs in methadone-versus buprenorphine-exposed neonates. Addiction 107, 53–62. doi:10.1111/j.1360-0443.2012.04039.x
Galanakis, D., Kourounakis, A. P., Tsiakitzis, K. C., Doulgkeris, C., Rekka, E. A., Gavalas, A., et al. (2004). Synthesis and pharmacological evaluation of amide conjugates of NSAIDs with L-cysteine ethyl ester, combining potent antiinflammatory and antioxidant properties with significantly reduced gastrointestinal toxicity. Bioorg Med. Chem. Lett. 14, 3639–3643. doi:10.1016/j.bmcl.2004.05.025
Gamper, N., and Ooi, L. (2015). Redox and nitric oxide-mediated regulation of sensory neuron ion channel function. Antioxid. Redox Signal 22, 486–504. doi:10.1089/ars.2014.5884
Gao, L., González-Rodríguez, P., Ortega-Sáenz, P., and López-Barneo, J. (2017). Redox signaling in acute oxygen sensing. Redox Biol. 12, 908–915. doi:10.1016/j.redox.2017.04.033
García, I. E., Sánchez, H. A., Martínez, A. D., and Retamal, M. A. (2018). Redox-mediated regulation of connexin proteins; focus on nitric oxide. Biochim. Biophys. Acta Biomembr. 1860, 91–95. doi:10.1016/j.bbamem.2017.10.006
García-Carmona, J. A., Martínez-Laorden, E., Milanés, M. V., and Laorden, M. L. (2015). Sympathetic activity induced by naloxone-precipitated morphine withdrawal is blocked in genetically engineered mice lacking functional CRF1 receptor. Toxicol. Appl. Pharmacol. 283, 42–49. doi:10.1016/j.taap.2015.01.002
Garcia-Portilla, M. P., Bobes-Bascaran, M. T., Bascaran, M. T., Saiz, P. A., and Bobes, J. (2014). Long term outcomes of pharmacological treatments for opioid dependence: does methadone still lead the pack? Br. J. Clin. Pharmacol. 77, 272–284. doi:10.1111/bcp.12031
Gardner, E. L. (2011). Addiction and brain reward and antireward pathways. Adv. Psychosom. Med. 30, 22–60. doi:10.1159/000324065
Garner, J. P. (2014). The significance of meaning: why do over 90% of behavioral neuroscience results fail to translate to humans, and what can we do to fix it? ILAR J. 55, 438–456. doi:10.1093/ilar/ilu047
Gaston, B., Baby, S. M., May, W. J., Young, A. P., Grossfield, A., Bates, J. N., et al. (2021). D-Cystine di(m)ethyl ester reverses the deleterious effects of morphine on ventilation and arterial blood gas chemistry while promoting antinociception. Sci. Rep. 11, 10038. doi:10.1038/s41598-021-89455-2
Gaston, B., May, W. J., Sullivan, S., Yemen, S., Marozkina, N. V., Palmer, L. A., et al. (2014). Essential role of hemoglobin beta-93-cysteine in post-hypoxia facilitation of breathing in conscious mice. J. Appl. Physiol. 116, 1290–1299. doi:10.1152/japplphysiol.01050.2013
Gaston, B., Singel, D., Doctor, A., and Stamler, J. S. (2006). S-nitrosothiol signaling in respiratory biology. Am. J. Respir. Crit. Care Med. 173, 1186–1193. doi:10.1164/rccm.200510-1584PP
Gaston, B., Smith, L., Bosch, J., Seckler, J., Kunze, D., Kiselar, J., et al. (2020). Voltage-gated potassium channel proteins and stereoselective S-nitroso-l-cysteine signaling. JCI Insight 5, e134174. doi:10.1172/jci.insight.134174
Getsy, P. M., Baby, S. M., Gruber, R. B., Gaston, B., Lewis, T. H. J., Grossfield, A., et al. (2022c). S-Nitroso-L-Cysteine stereoselectively blunts the deleterious effects of fentanyl on breathing while augmenting antinociception in freely-moving rats. Front. Pharmacol. 13, 892307. doi:10.3389/fphar.2022.892307
Getsy, P. M., Baby, S. M., May, W. J., Bates, J. N., Ellis, C. R., Feasel, M. G., et al. (2022a). L-cysteine methyl ester overcomes the deleterious effects of morphine on ventilatory parameters and arterial blood-gas chemistry in unanesthetized rats. Front. Pharmacol. 13, 968378. doi:10.3389/fphar.2022.968378
Getsy, P. M., Baby, S. M., May, W. J., Lewis, T. H. J., Bates, J. N., Hsieh, Y. H., et al. (2022b). L-NAC reverses of the adverse effects of fentanyl infusion on ventilation and blood-gas chemistry. Biomed. Pharmacother. 153, 113277. doi:10.1016/j.biopha.2022.113277
Getsy, P. M., Baby, S. M., May, W. J., Young, A. P., Gaston, B., Hodges, M. R., et al. (2022e). D-cysteine ethyl ester reverses the deleterious effects of morphine on breathing and arterial blood-gas chemistry in freely-moving rats. Front. Pharmacol. 13, 883329. doi:10.3389/fphar.2022.883329
Getsy, P. M., Coffee, G. A., and Lewis, S. J. (2023b). Loss of ganglioglomerular nerve input to the carotid body impacts the hypoxic ventilatory response in freely-moving rats. Front. Physiol. 14, 1007043. doi:10.3389/fphys.2023.1007043
Getsy, P. M., Davis, J., Coffee, G. A., Lewis, T. H. J., and Lewis, S. J. (2023a). Hypercapnic signaling influences hypoxic signaling in the control of breathing in C57BL6 mice. J. Appl. Physiol. 134, 1188–1206. doi:10.1152/japplphysiol.00548.2022
Getsy, P. M., Young, A. P., Bates, J. N., Baby, S. M., Seckler, J. M., Grossfield, A., et al. (2022f). S-nitroso-L-cysteine stereoselectively blunts the adverse effects of morphine on breathing and arterial blood gas chemistry while promoting analgesia. Biomed. Pharmacother. 153, 113436. doi:10.1016/j.biopha.2022.113436
Getsy, P. M., Young, A. P., Grossfield, A., Seckler, J. M., Wilson, C. G., Gaston, B., et al. (2022d). D-cysteine ethyl ester and D-cystine dimethyl ester reverse the deleterious effects of morphine on arterial blood-gas chemistry and Alveolar-arterial gradient in anesthetized rats. Respir Physiol Neurobiol 302, 103912. doi:10.1016/j.resp.2022.103912
Ghezzi, P., and Chan, P. (2017). Redox proteomics applied to the thiol secretome. Antioxid. Redox Signal 26, 299–312. doi:10.1089/ars.2016.6732
Gileta, A. F., Fitzpatrick, C. J., Chitre, A. S., St Pierre, C. L., Joyce, E. V., Maguire, R. J., et al. (2022). Genetic characterization of outbred Sprague Dawley rats and utility for genome-wide association studies. PLoS Genet. 18, e1010234. doi:10.1371/journal.pgen.1010234
Giustarini, D., Milzani, A., Dalle-Donne, I., Tsikas, D., and Rossi, R. (2012). N-Acetylcysteine ethyl ester (NACET): a novel lipophilic cell-permeable cysteine derivative with an unusual pharmacokinetic feature and remarkable antioxidant potential. Biochem. Pharmacol. 84, 1522–1533. doi:10.1016/j.bcp.2012.09.010
Glass, M. J. (2010). The role of functional postsynaptic NMDA receptors in the central nucleus of the amygdala in opioid dependence. Vitam. Horm. 82, 145–166. doi:10.1016/S0083-6729(10)82008-4
Glass, M. J. (2011). Opioid dependence and NMDA receptors. ILAR J. 52, 342–351. doi:10.1093/ilar.52.3.342
Gledhill, L. J., and Babey, A. M. (2021). Synthesis of the mechanisms of opioid tolerance: do we still say NO? Cell Mol. Neurobiol. 41, 927–948. doi:10.1007/s10571-021-01065-8
Goto, K., Hisadome, M., Kawazoe, Y., and Tsumagari, T. (1983). Effect of cysteine ethylester hydrochloride (Cystanin) on host defense mechanism]. Nihon Yakurigaku Zasshi 82, 27–35. doi:10.1254/fpj.82.27
Grim, T. W., Acevedo-Canabal, A., and Bohn, L. M. (2020). Toward directing opioid receptor signaling to refine opioid therapeutics. Biol. Psychiatry 87, 15–21. doi:10.1016/j.biopsych.2019.10.020
Gutowicz, M., Kaźmierczak, B., and Barańczyk-Kuźma, A. (2011). The influence of heroin abuse on glutathione-dependent enzymes in human brain. Drug Alcohol Depend. 113, 8–12. doi:10.1016/j.drugalcdep.2010.06.020
Hah, J. M., Bateman, B. T., Ratliff, J., Curtin, C., and Sun, E. (2017). Chronic opioid use after surgery: implications for perioperative management in the face of the opioid epidemic. Anesth. Analg. 125, 1733–1740. doi:10.1213/ANE.0000000000002458
Heberlein, A., Leggio, L., Stichtenoth, D., and Hillemacher, T. (2012). The treatment of alcohol and opioid dependence in pregnant women. Curr. Opin. Psychiatry 25, 559–564. doi:10.1097/YCO.0b013e328358ad36
Hendrie, C. A. (1985). Opiate dependence and withdrawal--a new synthesis? Pharmacol. Biochem. Behav. 23, 863–870. doi:10.1016/0091-3057(85)90084-x
Herman, B. H., Vocci, F., and Bridge, P. (1995). The effects of NMDA receptor antagonists and nitric oxide synthase inhibitors on opioid tolerance and withdrawal. Medication development issues for opiate addiction. Neuropsychopharmacology 13, 269–293. doi:10.1016/0893-133X(95)00140-9
Hess, D. T., and Stamler, J. S. (2012). Regulation by S-nitrosylation of protein post-translational modification. J. Biol. Chem. 287, 4411–4418. doi:10.1074/jbc.R111.285742
Hisadome, M., Fukuda, T., and Terasawa, M. (1990). Effect of cysteine ethylester hydrochloride (Cystanin) on host defense mechanisms (V): potentiation of nitroblue tetrazolium reduction and chemiluminescence in macrophages or leukocytes of mice or rats. Jpn. J. Pharmacol. 53, 57–66. doi:10.1254/jjp.53.57
Hisadome, M., Kimura, Y., Ikegami, K., and Terasawa, M. (1988). Effect of cysteine ethylester hydrochloride (Cystanin) on host defense mechanism (IV): potentiating effects on the function of peritoneal or spleen macrophages. Jpn. J. Pharmacol. 47, 379–385. doi:10.1254/jjp.47.379
Hisadome, M., Nakamura, Y., Okumoto, T., and Ikegami, K. (1986a). Effect of cysteine ethylester hydrochloride (Cystanin) on host defense mechanisms (II): restorative effects on the suppression of antibody production. Nihon Yakurigaku Zasshi 88, 349–354. doi:10.1254/fpj.88.349
Hisadome, M., Nakamura, Y., Okumoto, T., and Ikegami, K. (1986b). Effect of cysteine ethylester hydrochloride (Cystanin) on host defense mechanisms (III): potentiating effects on phagocytosis and nitroblue tetrazolium (NBT) reduction by leukocytes of mice and Guinea pigs. Nihon Yakurigaku Zasshi 88, 369–374. doi:10.1254/fpj.88.369
Hobbs, M. J., Butterworth, M., Cohen, G. M., and Upshall, D. G. (1993). Structure-activity relationships of cysteine esters and their effects on thiol levels in rat lung in vitro. Biochem. Pharmacol. 45, 1605–1612. doi:10.1016/0006-2952(93)90301-c
Holland, L. N., Shuster, L. C., and Buccafusco, J. J. (1993). Role of spinal and supraspinal muscarinic receptors in the expression of morphine withdrawal symptoms in the rat. Neuropharmacology 32, 1387–1395. doi:10.1016/0028-3908(93)90035-2
Horn, D. B., Vu, L., Porter, B. R., and Sarantopoulos, K. (2023). “Responsible controlled substance and opioid prescribing,” in StatPearls (Treasure Island (FL): StatPearls Publishing). PMID: 34283451.
Horsfall, J. T., and Sprague, J. E. (2017). The pharmacology and toxicology of the 'holy trinity. Basic Clin. Pharmacol. Toxicol. 120, 115–119. doi:10.1111/bcpt.12655
Hosseini, M., Taiarani, Z., Hadjzadeh, M. A., Salehabadi, S., Tehranipour, M., and Alaei, H. A. (2011). Different responses of nitric oxide synthase inhibition on morphine-induced antinociception in male and female rats. Pathophysiology 18, 143–149. doi:10.1016/j.pathophys.2010.05.004
Houshmand, G., Pourasghar, M., Shiran, M., Arab Firozjae, A., Goudarzi, M., Manouchehr, F., et al. (2021). Simvastatin prevents morphine antinociceptive tolerance and withdrawal symptoms through antioxidative effect and nitric oxide pathway in mice. Behav. Brain Res. 402, 113104. doi:10.1016/j.bbr.2020.113104
Hu, X., Tian, X., Guo, X., He, Y., Chen, H., Zhou, J., et al. (2018). AMPA receptor positive allosteric modulators attenuate morphine tolerance and dependence. Neuropharmacology 137, 50–58. doi:10.1016/j.neuropharm.2018.04.020
Huhn, A. S., Berry, M. S., and Dunn, K. E. (2019). Review: sex-based differences in treatment outcomes for persons with opioid use disorder. Am. J. Addict. 28, 246–261. doi:10.1111/ajad.12921
Humphreys, K., Shover, C. L., Andrews, C. M., Bohnert, A. S. B., Brandeau, M. L., Caulkins, J. P., et al. (2022). Responding to the opioid crisis in north America and beyond: recommendations of the stanford-lancet commission. Lancet 399, 555–604. doi:10.1016/S0140-6736(21)02252-2
Hutchinson, M. R., Bland, S. T., Johnson, K. W., Rice, K. C., Maier, S. F., and Watkins, L. R. (2007). Opioid-induced glial activation: mechanisms of activation and implications for opioid analgesia, dependence, and reward. ScientificWorldJournal 7, 98–111. doi:10.1100/tsw.2007.230
Ikemoto, M. J., Inoue, K., Akiduki, S., Osugi, T., Imamura, T., Ishida, N., et al. (2002). Identification of addicsin/GTRAP3-18 as a chronic morphine-augmented gene in amygdala. Neuroreport 13, 2079–2084. doi:10.1097/00001756-200211150-00018
Isoardi, K. Z., Parker, L., Harris, K., Rashford, S., and Isbister, G. K. (2022). Acute opioid withdrawal following intramuscular administration of naloxone 1.6 mg: a prospective out-of-hospital series. Ann. Emerg. Med. 80, 120–126. doi:10.1016/j.annemergmed.2022.03.004
Jarrott, B., Lewis, S. J., Doyle, A. E., and Louis, W. J. (1988). Effects of continuous infusions (10 days) and cessation of infusions of clonidine and rilmenidine (S 3341) on cardiovascular and behavioral parameters of spontaneously hypertensive rats. Am. J. Cardiol. 61, 39D-44D–44D. doi:10.1016/0002-9149(88)90463-8
Jarrott, B., Lewis, S. J., Louis, W. J., Maccarrone, C., and Shulkes, A. (1987). Regional brain concentrations of several putative peptide neurotransmitters in normotensive and spontaneously hypertensive rats: effects of continuous (10-day) clonidine infusion. J. Cardiovasc Pharmacol. 10, S14–S21. Suppl 12. doi:10.1097/00005344-198709002-00002
Jenkins, M. W., Khalid, F., Baby, S. M., May, W. J., Young, A. P., Bates, J. N., et al. (2021). Glutathione ethyl ester reverses the deleterious effects of fentanyl on ventilation and arterial blood-gas chemistry while prolonging fentanyl-induced analgesia. Sci. Rep. 11, 6985. doi:10.1038/s41598-021-86458-x
Jiménez-Fernández, B., Calomarde-Gómez, C., López-Lazcano, A., Lligoña, A., Gual, A., and López-Pelayo, H. (2022). Systematic review on the clinical management of chronic pain and comorbid opioid use disorder. Adicciones 0, 1680. doi:10.20882/adicciones.1680
Joseph, C. A., and Maroney, M. J. (2007). Cysteine dioxygenase: structure and mechanism. Chem. Commun. (Camb). 32, 3338–3349. doi:10.1039/b702158e
Kanbar, R., Stornetta, R. L., Cash, D. R., Lewis, S. J., and Guyenet, P. G. (2010). Photostimulation of Phox2b medullary neurons activates cardiorespiratory function in conscious rats. Am. J. Respir. Crit. Care Med. 182, 1184–1194. doi:10.1164/rccm.201001-0047OC
Kest, B., Palmese, C. A., Juni, A., Chesler, E. J., and Mogil, J. S. (2004). Mapping of a quantitative trait locus for morphine withdrawal severity. Mamm. Genome 15, 610–617. doi:10.1007/s00335-004-2367-3
Kimura, H. (2014). The physiological role of hydrogen sulfide and beyond. Nitric Oxide 41, 4–10. doi:10.1016/j.niox.2014.01.002
Kimura, H. (2017). Hydrogen sulfide and polysulfide signaling. Antioxid. Redox Signal. 27, 619–621. doi:10.1089/ars.2017.7076
Knauss, Z. T., Hearn, C. J., Hendryx, N. C., Aboalrob, F. S., Mueller-Figueroa, Y., Damron, D. S., et al. (2023). Fentanyl-induced reward seeking is sex and dose dependent and is prevented by D-cysteine ethylester. Front. Pharmacol. 14, 1241578. doi:10.3389/fphar.2023.1241578
Knouse, M. C., and Briand, L. A. (2021). Behavioral sex differences in cocaine and opioid use disorders: the role of gonadal hormones. Neurosci. Biobehav Rev. 128, 358–366. doi:10.1016/j.neubiorev.2021.06.038
Koob, G. F. (1987). Neural substrates of opioid tolerance and dependence. NIDA Res. Monogr. 76, 46–52.
Koob, G. F. (2020). Neurobiology of opioid addiction: opponent process, hyperkatifeia, and negative reinforcement. Biol. Psychiatry 87, 44–53. doi:10.1016/j.biopsych.2019.05.023
Koob, G. F., and Volkow, N. D. (2016). Neurobiology of addiction: a neurocircuitry analysis. Lancet Psychiatry 3, 760–773. doi:10.1016/S2215-0366(16)00104-8
Kregel, K. C., Kenney, M. J., Massett, M. P., Morgan, D. A., and Lewis, S. J. (1997). Role of nitrosyl factors in the hemodynamic adjustments to heat stress in the rat. Am. J. Physiol. 273, H1537–H1543. doi:10.1152/ajpheart.1997.273.3.H1537
Kularatne, R. N., Bulumulla, C., Catchpole, T., Takacs, A., Christie, A., Stefan, M. C., et al. (2020). Protection of human retinal pigment epithelial cells from oxidative damage using cysteine prodrugs. Free Radic. Biol. Med. 152, 386–394. doi:10.1016/j.freeradbiomed.2020.03.024
Laschka, E., and Herz, A. (1977). Sites of action of morphine involved in the development of physical dependence in rats. III. Autoradiographic studies. III. Autoradiogr. Stud. Psychopharmacol. Berl. 53, 33–37. doi:10.1007/BF00426691
Laschka, E., Herz, A., and Bläsig, J. (1976a). Sites of action of morphine involved in the development of physical dependence in rats. I. Comparison of precipitated morphine withdrawal after intraperitoneal and intraventricular injection of morphine antagonists. Psychopharmacologia 46, 133–139. doi:10.1007/BF00421382
Laschka, E., Teschemacher, H., Mehraein, P., and Herz, A. (1976b). Sites of action of morphine involved in the development of physical dependence in rats. II. Morphine withdrawal precipitated by application of morphine antagonists into restricted parts of the ventricular system and by microinjection into various brain areas. Psychopharmacologia 46, 141–147. doi:10.1007/BF00421383
Lash, L. H., and Jones, D. P. (1985). Distribution of oxidized and reduced forms of glutathione and cysteine in rat plasma. Arch. Biochem. Biophys. 240, 583–592. doi:10.1016/0003-9861(85)90065-7
Laska, F. J., and Fennessy, M. R. (1977). Dissociation of increased 5-hydroxyindoleacetic acid levels and physical dependence: the effects of naloxone. Clin. Exp. Pharmacol. Physiol. 4, 515–523. doi:10.1111/j.1440-1681.1977.tb02681.x
Laska, F. J., and Fennessy, M. R. (1978). Induction of physical dependence on cyclazocine and pentazocine in the rat. Eur. J. Pharmacol. 48, 57–65. doi:10.1016/0014-2999(78)90044-4
Laska, J. F., and Fennessy, M. R. (1976). Physical dependence in the rat induced by slow release morphine: dose-response, time course and brain biogenic amines. Clin. Exp. Pharmacol. Physiol. 3, 587–598. doi:10.1111/j.1440-1681.1976.tb00640.x
Lee, J. R., and Fennessy, M. R. (1970). The relationship between morphine analgesia and the levels of biogenic amines in the mouse brain. Eur. J. Pharmacol. 12, 65–70. doi:10.1016/0014-2999(70)90029-4
Lee, S., Tatla, V., Buga, S., and Pon, D. (2022). Comparing the safety and efficacy of intravenous naloxone administration in opioid-naive and opioid-tolerant hospitalized oncology patients. J. Opioid Manag. 18, 497–502. doi:10.5055/jom.2022.0744
Levin, C. J., Wai, J. M., Jones, J. D., and Comer, S. D. (2019). Changes in cardiac vagal tone as measured by heart rate variability during naloxone-induced opioid withdrawal. Drug Alcohol Depend. 204, 107538. doi:10.1016/j.drugalcdep.2019.06.040
Lewis, S. J., Dunlop, M., and Jarrott, B. (1989). Serum glucose and insulin levels in normotensive (WKY) and spontaneously hypertensive (SH) rats during and after the cessation of continuous (10 day) clonidine infusion. J. Pharm. Pharmacol. 41, 353–355. doi:10.1111/j.2042-7158.1989.tb06474.x
Lewis, S. J., Hoque, A., and Bates, J. N. (2005). Differentiation of L- and D-S-nitrosothiol recognition sites in vivo. J. Cardiovasc Pharmacol. 46, 660–671. doi:10.1097/01.fjc.0000181714.94827.5d
Lewis, S. J., Owen, J. R., and Bates, J. N. (2006). S-nitrosocysteine elicits hemodynamic responses similar to those of the Bezold-Jarisch reflex via activation of stereoselective recognition sites. Eur. J. Pharmacol. 531, 254–258. doi:10.1016/j.ejphar.2005.11.027
Lewis, S. J., Rowe, P., and Jarrott, B. (1988a). Involvement of hypothalamic adrenaline in the clonidine withdrawal syndrome in normotensive and spontaneously hypertensive rats. Clin. Exp. Pharmacol. Physiol. 15, 773–780. doi:10.1111/j.1440-1681.1988.tb01017.x
Lewis, S. J., Svec, J., Fennessy, M. R., and Jarrott, B. (1988b). Lack of involvement of endogenous mu-receptor opioids in the hypothermic effects of clonidine in normotensive and spontaneously hypertensive rats. Neuropharmacology 27, 537–540. doi:10.1016/0028-3908(88)90137-2
Lewis, S. J., Travis, M. D., and Bates, J. N. (1996). Stereoselective S-nitrosocysteine recognition sites in rat brain. Eur. J. Pharmacol. 312, R3–R5. doi:10.1016/0014-2999(96)00607-3
Lewis, T. H. J., May, W. J., Young, A. P., Bates, J. N., Baby, S. M., Getsy, P. M., et al. (2022). The ventilatory depressant actions but not the antinociceptive effects of morphine are blunted in rats receiving intravenous infusion of L-cysteine ethyl ester. Biomed. Pharmacother. 156, 113939. doi:10.1016/j.biopha.2022.113939
Leza, J. C., Lizasoain, I., Cuéllar, B., Moro, M. A., and Lorenzo, P. (1996). Correlation between brain nitric oxide synthase activity and opiate withdrawal. Naunyn Schmiedeb. Arch. Pharmacol. 353, 349–354. doi:10.1007/BF00168639
Leza, J. C., Lizasoain, I., San-Martín-Clark, O., and Lorenzo, P. (1995). Morphine-induced changes in cerebral and cerebellar nitric oxide synthase activity. Eur. J. Pharmacol. 285, 95–98. doi:10.1016/0014-2999(95)00474-y
Li, F., Jiang, H., Shen, X., Yang, W., Guo, C., Wang, Z., et al. (2021). Sneezing reflex is mediated by a peptidergic pathway from nose to brainstem. Cell 184, 3762–3773.e10. doi:10.1016/j.cell.2021.05.017
Li, Y., Su, L., Li, F., Wang, C., Yuan, D., Chen, J., et al. (2015). Acute and sub-chronic toxicity of glucose-cysteine Maillard reaction products in Sprague-Dawley rats. Food Chem. Toxicol. 80, 271–276. doi:10.1016/j.fct.2015.03.021
Liang, D. Y., Shi, X., Li, X., Li, J., and Clark, J. D. (2007). The beta2 adrenergic receptor regulates morphine tolerance and physical dependence. Behav. Brain Res. 181, 118–126. doi:10.1016/j.bbr.2007.03.037
Lim, G., Wang, S., and Mao, J. (2012). cAMP and protein kinase A contribute to the downregulation of spinal glutamate transporters after chronic morphine. Neurosci. Lett. 376, 9–13. doi:10.1016/j.neulet.2004.11.016
Lin, C. I., Orlov, I., Ruggiero, A. M., Dykes-Hoberg, M., Lee, A., Jackson, M., et al. (2001). Modulation of the neuronal glutamate transporter EAAC1 by the interacting protein GTRAP3-18. Nature 410, 84–88. doi:10.1038/35065084
Lipton, A. J., Johnson, A., Macdonald, T., Lieberman, M. W., Gozal, D., and Gaston, B. (2001). S-nitrosothiols signal the ventilatory response to hypoxia. Nature 413, 171–174. doi:10.1038/35093117
Lipton, S. A., Choi, Y. B., Pan, Z. H., Lei, S. Z., Chen, H. S., Sucher, N. J., et al. (1993). A redox-based mechanism for the neuroprotective and neurodestructive effects of nitric oxide and related nitroso-compounds. Nature 364, 626–632. doi:10.1038/364626a0
Liu, L., Fu, Y., He, H., Ji, F., Liu, F., and Cao, M. (2011). Expression of EAAT3 in prefrontal cortex and hippocampus in CPP reinstatement rats induced by morphine. Chin. J. Pathophysiol. 27, 1720–1724.
Liu, Z., Zheng, J. F., Yang, L. Q., Yi, L., and Hu, B. (2007). Effects of sinomenine on NO/nNOS system in cerebellum and spinal cord of morphine-dependent and withdrawal mice. Sheng Li Xue Bao 59, 285–292.
Lofwall, M. R., Babalonis, S., Nuzzo, P. A., Siegel, A., Campbell, C., and Walsh, S. L. (2013). Efficacy of extended-release tramadol for treatment of prescription opioid withdrawal: a two-phase randomized controlled trial. Drug Alcohol Depend. 133, 188–197. doi:10.1016/j.drugalcdep.2013.05.010
London, E. D., Kimes, A. S., and Vaupel, D. B. (1995). Inhibitors of nitric oxide synthase and the opioid withdrawal syndrome. NIDA Res. Monogr. 147, 170–181.
Lopez-Gimenez, J. F., and Milligan, G. (2010). Opioid regulation of mu receptor internalisation: relevance to the development of tolerance and dependence. CNS Neurol. Disord. Drug Targets 9, 616–626. doi:10.2174/187152710793361522
Ludbrook, J. (1998). Multiple comparison procedures updated. Clin. Exp. Pharmacol. Physiol. 25, 1032–1037. doi:10.1111/j.1440-1681.1998.tb02179.x
Majeed, N. H., Przewłocka, B., Machelska, H., and Przewłocki, R. (1994). Inhibition of nitric oxide synthase attenuates the development of morphine tolerance and dependence in mice. Neuropharmacology 33, 189–192. doi:10.1016/0028-3908(94)90006-x
Manhapra, A. (2022). Complex persistent opioid dependence-an opioid-induced chronic pain syndrome. Curr. Treat. Options Oncol. 23, 921–935. doi:10.1007/s11864-022-00985-x
Mansouri, M. T., Naghizadeh, B., Ghorbanzadeh, B., Amirgholami, N., Houshmand, G., and Alboghobeish, S. (2020). Venlafaxine inhibits naloxone-precipitated morphine withdrawal symptoms: role of inflammatory cytokines and nitric oxide. Metab. Brain Dis. 35, 305–313. doi:10.1007/s11011-019-00491-4
Mao, J., Sung, B., Ji, R.-R., and Lim, G. (2002). Chronic morphine induces downregulation of spinal glutamate transporters: implications in morphine tolerance and abnormal pain sensitivity. J. Neurosci. 22, 8312–8323. doi:10.1523/JNEUROSCI.22-18-08312.2002
Marshall, D. C., and Buccafusco, J. J. (1985). Supraspinal and spinal mediation of naloxone-induced morphine withdrawal in rats. Brain Res. 329, 131–142. doi:10.1016/0006-8993(85)90518-9
Marshall, D. C., and Buccafusco, J. J. (1987). Spinal cholinergic neurons and the expression of morphine withdrawal symptoms in the rat. J. Neurosci. 7, 621–628. doi:10.1523/JNEUROSCI.07-03-00621.1987
Matson, P. A., Ridenour, T., Ialongo, N., Spoth, R., Prado, G., Hammond, C. J., et al. (2022). State of the art in substance use prevention and early intervention: applications to pediatric primary care settings. Prev. Sci. 23, 204–211. doi:10.1007/s11121-021-01299-4
Maze, I., and Nestler, E. J. (2011). The epigenetic landscape of addiction. Ann. N. Y. Acad. Sci. 1216, 99–113. doi:10.1111/j.1749-6632.2010.05893.x
McCarberg, B. H. (2011). Pain management in primary care: strategies to mitigate opioid misuse, abuse, and diversion. Postgrad. Med. 123, 119–130. doi:10.3810/pgm.2011.03.2270
McHugh, M. L. (2011). Multiple comparison analysis testing in ANOVA. Biochem. Med. Zagreb. 21, 203–209. doi:10.11613/bm.2011.029
Mendoza, J., Passafaro, R., Baby, S., Young, A. P., Bates, J. N., Gaston, B., et al. (2013). L-Cysteine ethyl ester reverses the deleterious effects of morphine on, arterial blood-gas chemistry in tracheotomized rats. Respir. Physiol. Neurobiol. 189, 136–143. doi:10.1016/j.resp.2013.07.007
Mercadante, S., Arcuri, E., and Santoni, A. (2019). Opioid-induced tolerance and hyperalgesia. CNS Drugs 33, 943–955. doi:10.1007/s40263-019-00660-0
Michaud, N., and Couture, R. (2003). Cardiovascular and behavioural effects induced by naloxone-precipitated morphine withdrawal in rat: characterization with tachykinin antagonists. Neuropeptides 37, 345–354. doi:10.1016/j.npep.2003.09.003
Morgan, M. M., and Christie, M. J. (2011). Analysis of opioid efficacy, tolerance, addiction and dependence from cell culture to human. Br. J. Pharmacol. 164, 1322–1334. doi:10.1111/j.1476-5381.2011.01335.x
Mori, T., Ito, S., Matsubayashi, K., and Sawaguchi, T. (2007). Comparison of nitric oxide synthase inhibitors, phospholipase A2 inhibitor and free radical scavengers as attenuators of opioid withdrawal syndrome. Behav. Pharmacol. 18, 725–729. doi:10.1097/FBP.0b013e3282f18da6
Mosier-Boss, P. A., and Lieberman, S. H. (2005). The role of hydrogen bonding in the selectivity of L-cysteine methyl ester (CYSM) and L-cysteine ethyl ester (CYSE) for chloride ion. Spectrochim. Acta A Mol. Biomol. Spectrosc. 61, 845–854. doi:10.1016/j.saa.2004.06.011
Myers, P. R., Minor, R. L., Guerra, R., Bates, J. N., and Harrison, D. G. (1990). Vasorelaxant properties of the endothelium-derived relaxing factor more closely resemble S-nitrosocysteine than nitric oxide. Nature 345, 161–163. doi:10.1038/345161a0
Naidu, P. S., Singh, A., Joshi, D., and Kulkarni, S. K. (2003). Possible mechanisms of action in quercetin reversal of morphine tolerance and dependence. Addict. Biol. 8, 327–336. doi:10.1080/13556210310001602248
Newlin, D. B., Wong, C. J., and Cheskin, L. J. (1992). Cardiovascular responses to naloxone challenge in opiate-dependent individuals. Pharmacol. Biochem. Behav. 43, 357–360. doi:10.1016/0091-3057(92)90162-9
Nielsen, D. A., and Kreek, M. J. (2012). Common and specific liability to addiction: approaches to association studies of opioid addiction. Drug Alcohol Depend. 123, S33–S41. doi:10.1016/j.drugalcdep.2012.03.026
Nishida, K., Ohta, Y., Ito, M., Nagamura, Y., Kitahara, S., Fujii, K., et al. (1996). Conversion of gamma-glutamylcysteinylethyl ester to glutathione in rat hepatocytes. Biochim. Biophys. Acta 1313, 47–53. doi:10.1016/0167-4889(96)00054-7
Noda, Y., and Nabeshima, T. (2004). Opiate physical dependence and N-methyl-D-aspartate receptors. Eur. J. Pharmacol. 500, 121–128. doi:10.1016/j.ejphar.2004.07.017
Ohta, H., Bates, J. N., Lewis, S. J., and Talman, W. T. (1997). Actions of S-nitrosocysteine in the nucleus tractus solitarii are unrelated to release of nitric oxide. Brain Res. 746, 98–104. doi:10.1016/s0006-8993(96)01188-2
Ostrea, E. M., Chavez, C. J., and Strauss, M. E. (1975). A study of factors that influence the severity of neonatal narcotic withdrawal. Addict. Dis. 2, 187–199.
O-Uchi, J., Ryu, S. Y., Jhun, B. S., Hurst, S., and Sheu, S. S. (2014). Mitochondrial ion channels/transporters as sensors and regulators of cellular redox signaling. Antioxid. Redox Signal 21, 987–1006. doi:10.1089/ars.2013.5681
Owusu Obeng, A., Hamadeh, I., and Smith, M. (2017). Review of opioid pharmacogenetics and considerations for pain management. Pharmacotherapy 37, 1105–1121. doi:10.1002/phar.1986
Palmer, L. A., deRonde, K., Brown-Steinke, K., Gunter, S., Jyothikumar, V., Forbes, M. S., et al. (2015). Hypoxia-induced changes in protein S-nitrosylation in female mouse brainstem. Am. J. Respir. Cell Mol. Biol. 52, 37–45. doi:10.1165/rcmb.2013-0359OC
Palmer, L. A., May, W. J., deRonde, K., Brown-Steinke, K., Bates, J. N., Gaston, B., et al. (2013). Ventilatory responses during and following exposure to a hypoxic challenge in conscious mice deficient or null in S-nitrosoglutathione reductase. Resp. Physiol. Neurobiol. 185, 571–581. doi:10.1016/j.resp.2012.11.009
Perissinotti, L. L., Turjanski, A. G., Estrin, D. A., and Doctorovich, F. (2005). Transnitrosation of nitrosothiols: characterization of an elusive intermediate. J. Am. Chem. Soc. 127, 486–487. doi:10.1021/ja044056v
Philip, V. M., Duvvuru, S., Gomero, B., Ansah, T. A., Blaha, C. D., Cook, M. N., et al. (2010). High-throughput behavioral phenotyping in the expanded panel of BXD recombinant inbred strains. Genes Brain Behav. 9, 129–159. doi:10.1111/j.1601-183X.2009.00540.x
Prabhakar, N. R. (2012). Hydrogen sulfide (H(2)S): a physiologic mediator of carotid body response to hypoxia. Adv. Exp. Med. Biol. 758, 109–113. doi:10.1007/978-94-007-4584-1_15
Preux, C., Bertin, M., Tarot, A., Authier, N., Pinol, N., Brugnon, D., et al. (2022). Prevalence of opioid use disorder among patients with cancer-related pain: a systematic review. J. Clin. Med. 11, 1594. doi:10.3390/jcm11061594
Purssell, R., Godwin, J., Moe, J., Buxton, J., Crabtree, A., Kestler, A., et al. (1995). Nitric oxide synthase inhibitors. Preclinical studies of potential use for treatment of opioid withdrawal. Neuropsychopharmacology 13, 315–322. doi:10.1016/0893-133X(95)00138-4
Raghavendra, V., and Kulkarni, S. K. (1999). Reversal of morphine tolerance and dependence by melatonin: possible role of central and peripheral benzodiazepine receptors. Brain Res. 834, 178–181. doi:10.1016/s0006-8993(99)01520-6
Raghavendra, V., and Kulkarni, S. K. (2000). Possible mechanisms of action in melatonin reversal of morphine tolerance and dependence in mice. Eur. J. Pharmacol. 409, 279–289. doi:10.1016/s0014-2999(00)00849-9
Ramirez, J. M., Vlemincx, E., Baertsch, N. A., and Severs, L. J. (2022). The sigh and related behaviors. Handb. Clin. Neurol. 188, 357–372. doi:10.1016/B978-0-323-91534-2.00015-1
Rasmussen, K. (1995). The role of the locus coeruleus and N-methyl-D-aspartic acid (NMDA) and AMPA receptors in opiate withdrawal. Neuropsychopharmacology 13, 295–300. doi:10.1016/0893-133X(95)00082-O
Rossi, R., Giustarini, D., Milzani, A., and Dalle-Donne, I. (2009). Cysteinylation and homocysteinylation of plasma protein thiols during ageing of healthy human beings. J. Cell Mol. Med. 13, 3131–3140. doi:10.1111/j.1582-4934.2008.00417.x
Sackner, M. A., Lopez, J. R., Banderas, V., and Adams, J. A. (2019). Holistic approach to opioid use disorder: think nitric oxide. J. Opioid Manag. 15, 521–555. doi:10.5055/jom.2019.0543
Saiepour, M. H., Semnanian, S., and Fathollahi, Y. (2001). Occurrence of morphine tolerance and dependence in the nucleus paragigantocellularis neurons. Eur. J. Pharmacol. 411, 85–92. doi:10.1016/s0014-2999(00)00862-1
Sakloth, F., Polizu, C., Bertherat, F., and Zachariou, V. (2020). Regulators of G Protein signaling in analgesia and addiction. Mol. Pharmacol. 98, 739–750. doi:10.1124/mol.119.119206
Sarton, E., Dahan, A., and Teppema, L. (1998). Do sex-related differences exist in the respiratory pharmacology of opioids? Adv. Exp. Med. Biol. 450, 141–145. doi:10.1007/978-1-4757-9077-1_23
Saul, M. C., Philip, V. M., Reinholdt, L. G., Center for Systems Neurogenetics of Addiction, , and Chesler, E. J. (2019). High-diversity mouse populations for complex traits. Trends Genet. 35, 501–514. doi:10.1016/j.tig.2019.04.003
Schmid, C. L., Kennedy, M., Ross, N. C., Lovell, K. M., Yue, Z., Morgenweck, J., et al. (2017). Bias factor and therapeutic window correlate to predict safer opioid analgesics. Cell 171, 1165–1175. doi:10.1016/j.cell.2017.10.035
Schöneich, C., Dillinger, U., von Bruchhausen, F., and Asmus, K. D. (1992). Oxidation of polyunsaturated fatty acids and lipids through thiyl and sulfonyl radicals: reaction kinetics, and influence of oxygen and structure of thiyl radicals. Arch. Biochem. Biophys. 292, 456–467. doi:10.1016/0003-9861(92)90016-p
Schwarzer, A., Aichinger-Hinterhofer, M., Maier, C., Vollert, J., and Walther, J. W. (2015). Sleep-disordered breathing decreases after opioid withdrawal: results of a prospective controlled trial. Pain 156, 2167–2174. doi:10.1097/j.pain.0000000000000279
Seckler, J. M., Grossfield, A., May, W. J., Getsy, P. M., and Lewis, S. J. (2022). Nitrosyl factors play a vital role in the ventilatory depressant effects of fentanyl in unanesthetized rats. Biomed. Pharmacother. 146, 112571. doi:10.1016/j.biopha.2021.112571
Seckler, J. M., Meyer, N. M., Burton, S. T., Bates, J. N., Gaston, B., and Lewis, S. J. (2017). Detection of trace concentrations of S-nitrosothiols by means of a capacitive sensor. PLoS One 12, grime0187149. doi:10.1371/journal.pone.0187149
Seckler, J. M., Shen, J., Lewis, T. H. J., Abdulameer, M. A., Zaman, K., Palmer, L. A., et al. (2020). NADPH diaphorase detects S-nitrosylated proteins in aldehyde-treated biological tissues. Sci. Rep. 10, 21088. doi:10.1038/s41598-020-78107-6
Semenza, E. R., Harraz, M. M., Abramson, E., Malla, A. P., Vasavda, C., Gadalla, M. M., et al. (2021). D-cysteine is an endogenous regulator of neural progenitor cell dynamics in the mammalian brain. Proc. Natl. Acad. Sci. U. S. A. 118, e2110610118. doi:10.1073/pnas.2110610118
Serafini, R. A., and Zachariou, V. (2019). Opioid-galanin receptor heteromers differentiate the dopaminergic effects of morphine and methadone. J. Clin. Invest. 129, 2653–2654. doi:10.1172/JCI128987
Servin, A. L., Goulinet, S., and Renault, H. (1988). Pharmacokinetics of cysteine ethyl ester in rat. Xenobiotica 18, 839–847. doi:10.3109/00498258809041722
Seth, D., and Stamler, J. S. (2011). The SNO-proteome: causation and classifications. Curr. Opin. Chem. Biol. 15, 129–136. doi:10.1016/j.cbpa.2010.10.012
Singh, A., Naidu, P. S., and Kulkarni, S. K. (2002). Quercetin, a bioflavonoid reverses development of tolerance and dependence to morphine. Drug Dev. Res. 57, 167–172. doi:10.1002/ddr.10119
Singh, P., Sharma, B., Gupta, S., and Sharma, B. M. (2015). In vivo and in vitro attenuation of naloxone-precipitated experimental opioid withdrawal syndrome by insulin and selective KATP channel modulator. Psychopharmacol. Berl. 232, 465–475. doi:10.1007/s00213-014-3680-5
Singh, V. P., Jain, N. K., and Kulkarni, S. K. (2003). Fluoxetine suppresses morphine tolerance and dependence: modulation of NO-cGMP/DA/serotoninergic pathways. Methods Find. Exp. Clin. Pharmacol. 25, 273–280. doi:10.1358/mf.2003.25.4.769675
Specker, S., Wananukul, W., Hatsukami, D., Nolin, K., Hooke, L., Kreek, M. J., et al. (1998). Effects of dynorphin A(1-13) on opiate withdrawal in humans. Psychopharmacol. Berl. 137, 326–332. doi:10.1007/s002130050626
Stephan, B. C., and Parsa, F. D. (2016). Avoiding opioids and their harmful side effects in the postoperative patient: exogenous opioids, endogenous endorphins, wellness, mood, and their relation to postoperative pain. Hawaii J. Med. Public Health 75, 63–67.
Stipanuk, M. H., Simmons, C. R., Karplus, P. A., and Dominy, J. E. (2011). Thiol dioxygenases: unique families of cupin proteins. Amino Acids 41, 91–102. doi:10.1007/s00726-010-0518-2
Stipanuk, M. H., Ueki, I., Dominy, J. E., Simmons, C. R., and Hirschberger, L. L. (2009). Cysteine dioxygenase: a robust system for regulation of cellular cysteine levels. Amino Acids 37, 55–63. doi:10.1007/s00726-008-0202-y
Stomberski, C. T., Hess, D. T., and Stamler, J. S. (2019). Protein S-nitrosylation: determinants of specificity and enzymatic regulation of S-Nitrosothiol-Based signaling. Antioxid. Redox Signal 30, 1331–1351. doi:10.1089/ars.2017.7403
Sun, H., Maze, I., Dietz, D. M., Scobie, K. N., Kennedy, P. J., Damez-Werno, D., et al. (2012). Morphine epigenomically regulates behavior through alterations in histone H3 lysine 9 dimethylation in the nucleus accumbens. J. Neurosci. 32, 17454–17464. doi:10.1523/JNEUROSCI.1357-12.2012
Szwergold, B. S. (2006). Alpha-thiolamines such as cysteine and cysteamine act as effective transglycating agents due to formation of irreversible thiazolidine derivatives. Med. Hypotheses 66, 698–707. doi:10.1016/j.mehy.2005.10.029
Tang, L., Shukla, P. K., Wang, L. X., and Wang, Z. J. (2006). Reversal of morphine antinociceptive tolerance and dependence by the acute supraspinal inhibition of Ca2+/calmodulin-dependent protein kinase II. J. Pharmacol. Exp. Ther. 317, 901–909. doi:10.1124/jpet.105.097733
Tokuyama, S., Feng, Y., Wakabayashi, H., and Ho, I. K. (1995). Ca2+ channel blocker, diltiazem, prevents physical dependence and the enhancement of protein kinase C activity by opioid infusion in rats. Eur. J. Pharmacol. 279, 93–98. doi:10.1016/0014-2999(95)00140-g
Torres-Lockhart, K. E., Lu, T. Y., Weimer, M. B., Stein, M. R., and Cunningham, C. O. (2022). Clinical management of opioid withdrawal. Addiction 117, 2540–2550. doi:10.1111/add.15818
Tosi, G. M., Giustarini, D., Franci, L., Minetti, A., Imperatore, F., Caldi, E., et al. (2021). Superior properties of N-acetylcysteine ethyl ester over N-acetyl cysteine to prevent retinal pigment epithelial cells oxidative damage. Int. J. Mol. Sci. 22, 600. doi:10.3390/ijms22020600
Travis, M. D., Davisson, R. L., Bates, J. N., and Lewis, S. J. (1997). Hemodynamic effects of L- and D-S-nitroso-beta,beta-dimethylcysteine in rats. Am. J. Physiol. 273, H1493–H1501. doi:10.1152/ajpheart.1997.273.3.H1493
Travis, M. D., Stoll, L., Bates, J. N., and Lewis, S. J. (1996). L- and D-S-nitroso-beta,beta-dimethylcysteine differentially increase cGMP in cultured vascular smooth muscle cells. Eur. J. Pharmacol. 318, 47–53. doi:10.1016/s0014-2999(96)00719-4
Trivedi, M., Shah, J., Hodgson, N., Byun, H. M., and Deth, R. (2014). Morphine induces redox-based changes in global DNA methylation and retrotransposon transcription by inhibition of excitatory amino acid transporter type 3-mediated cysteine uptake. Mol. Pharmacol. 85, 747–757. doi:10.1124/mol.114.091728
Trivedi, M. S., and Deth, R. (2015). Redox-based epigenetic status in drug addiction: a potential contributor to gene priming and a mechanistic rationale for metabolic intervention. Front. Neurosci. 8, 444. doi:10.3389/fnins.2014.00444
Trivedi, M. S., Hodgson, N. W., Walker, S. J., Trooskens, G., Nair, V., and Deth, R. C. (2015). Epigenetic effects of casein-derived opioid peptides in SH-SY5Y human neuroblastoma cells. Nutr. Metab. (Lond) 12, 54. doi:10.1186/s12986-015-0050-1
Tsakova, A., Surcheva, S., Simeonova, K., Altankova, I., Marinova, T., Usunoff, K., et al. (2015). Nitroxidergic modulation of behavioural, cardiovascular and immune responses, and brain NADPH diaphorase activity upon morphine tolerance/dependence in rats. Biotechnol. Biotechnol. Equip. 29, 92–100. doi:10.1080/13102818.2014.990924
Turell, L., Radi, R., and Alvarez, B. (2013). The thiol pool in human plasma: the central contribution of albumin to redox processes. Free Radic. Biol. Med. 65, 244–253. doi:10.1016/j.freeradbiomed.2013.05.050
Undem, B. J., Kajekar, R., Hunter, D. D., and Myers, A. C. (2000). Neural integration and allergic disease. J. Allergy Clin. Immunol. 106, S213–S220. doi:10.1067/mai.2000.110153
Vaupel, D. B., Kimes, A. S., and London, E. D. (1995a). Nitric oxide synthase inhibitors. Preclinical studies of potential use for treatment of opioid withdrawal. Neuropsychopharmacology 13, 315–322. doi:10.1016/0893-133X(95)00138-4
Vaupel, D. B., Kimes, A. S., and London, E. D. (1995b). Comparison of 7-nitroindazole with other nitric oxide synthase inhibitors as attenuators of opioid withdrawal. Psychopharmacol. Berl. 118, 361–368. doi:10.1007/BF02245935
Volkow, N., Benveniste, H., and McLellan, A. T. (2018). Use and misuse of opioids in chronic pain. Annu. Rev. Med. 69, 451–465. doi:10.1146/annurev-med-011817-044739
Volkow, N. D., Michaelides, M., and Baler, R. (2019). The neuroscience of drug reward and addiction. Physiol. Rev. 99, 2115–2140. doi:10.1152/physrev.00014.2018
Wallenstein, S., Zucker, C. L., and Fleiss, J. L. (1980). Some statistical methods useful in circulation research. Circ. Res. 47, 1–9. doi:10.1161/01.res.47.1.1
Walsh, S. L., Strain, E. C., and Bigelow, G. E. (2003). Evaluation of the effects of lofexidine and clonidine on naloxone-precipitated withdrawal in opioid-dependent humans. Addiction 98, 427–439. doi:10.1046/j.1360-0443.2003.00372.x
Wang, D., Chen, P., Li, Q., Quirion, R., and Hong, Y. (2011). Blockade of adrenomedullin receptors reverses morphine tolerance and its neurochemical mechanisms. Behav. Brain Res. 221, 83–90. doi:10.1016/j.bbr.2011.02.046
Wang, Q., Liu-Chen, L.-Y., and Traynor, J. R. (2009). Differential modulation of μ- and δ-opioid receptor agonists by endogenous RGS4 protein in SH-SY5Y cells. J. Biol. Chem. 284, 18357–18367. doi:10.1074/jbc.M109.015453
Wang, Z., Bilsky, E. J., Wang, D., Porreca, F., and Sadée, W. (1999). 3-Isobutyl-1-methylxanthine inhibits basal mu-opioid receptor phosphorylation and reverses acute morphine tolerance and dependence in mice. Eur. J. Pharmacol. 371, 1–9. doi:10.1016/s0014-2999(99)00131-4
Wang, Z. J., Tang, L., and Xin, L. (2003). Reversal of morphine antinociceptive tolerance by acute spinal inhibition of Ca2+/calmodulin-dependent protein kinase II. Eur. J. Pharmacol. 465, 199–200. doi:10.1016/s0014-2999(03)01484-5
Ward, P., Moss, H. G., Brown, T. R., Kalivas, P., and Jenkins, D. D. (2020). N-acetylcysteine mitigates acute opioid withdrawal behaviors and CNS oxidative stress in neonatal rats. Pediatr. Res. 88, 77–84. doi:10.1038/s41390-019-0728-6
Wilcken, D. E., and Gupta, V. J. (1979). Cysteine-homocysteine mixed disulphide: differing plasma concentrations in normal men and women. Clin. Sci. (Lond). 57, 211–215. doi:10.1042/cs0570211
Wilson, M., Fritz, R., Finlay, M., and Cook, D. J. (2023). Piloting smart home sensors to detect overnight respiratory and withdrawal symptoms in adults prescribed opioids. Pain Manag. Nurs. 24, 4–11. doi:10.1016/j.pmn.2022.08.011
Winkler, A., Kutchan, T. M., and Macheroux, P. (2007). 6-S-cysteinylation of bi-covalently attached FAD in berberine bridge enzyme tunes the redox potential for optimal activity. J. Biol. Chem. 282, 24437–24443. doi:10.1074/jbc.M703642200
Wong, C. L., and Roberts, M. B. (1976). The effects of L-histidine and of specific histamine receptor agonists, on the expression of morphine tolerance and physical dependence in mice. Agents Actions 6, 569–576. doi:10.1007/BF01971571
Wróbel, M., Ubuka, T., Yao, W. B., and Abe, T. (1997). Effect of glucose-cysteine adduct on cysteine desulfuration in Guinea pig tissues. Physiol. Chem. Phys. Med. NMR 29, 11–14.
Xu, B., Wang, Z., Li, G., Li, B., Lin, H., Zheng, R., et al. (2006). Heroin-administered mice involved in oxidative stress and exogenous antioxidant-alleviated withdrawal syndrome. Basic Clin. Pharmacol. Toxicol. 99, 153–161. doi:10.1111/j.1742-7843.2006.pto_461.x
Xu, N.-J., Bao, L., Fan, H.-P., Bao, G.-B., Pu, L., Lu, Y.-J., et al. (2003). Morphine withdrawal increases glutamate uptake and surface expression of glutamate transporter GLT1 at hippocampal synapses. J. Neurosci. 23, 4775–4784. doi:10.1523/JNEUROSCI.23-11-04775.2003
Yamaguchi, K., and Hosokawa, Y. (1987). Cysteine dioxygenase. Methods Enzymol. 143, 395–403. doi:10.1016/0076-6879(87)43069-3
Yang, C., Chen, Y., Tang, L., and Wang, Z. J. (2011). Haloperidol disrupts opioid-antinociceptive tolerance and physical dependence. J. Pharmacol. Exp. Ther. 338, 164–172. doi:10.1124/jpet.110.175539
Yang, L., Wang, S., Sung, B., Lim, G., and Mao, J. (2008). Morphine induces ubiquitin-proteasome activity and glutamate transporter degradation. J. Biol. Chem. 283, 21703–21713. doi:10.1074/jbc.M800809200
Young, A. P., Gruber, R. B., Discala, J. F., May, W. J., Palmer, L. A., Lewis, S. J., et al. (2013). Co-activation of μ- and δ-opioid receptors elicits tolerance to morphine-induced ventilatory depression via generation of peroxynitrite. Resp. Physiol. Neurobiol. 186, 255–264. doi:10.1016/j.resp.2013.02.028
Zamani, N., Buckley, N. A., and Hassanian-Moghaddam, H. (2020). Buprenorphine to reverse respiratory depression from methadone overdose in opioid-dependent patients: a prospective randomized trial. Crit. Care 24, 44. doi:10.1186/s13054-020-2740-y
Keywords: opioids, morphine, naloxone, physical dependence, withdrawal, L-cysteine, L-cysteine ethyl ester, rats
Citation: Bates JN, Getsy PM, Coffee GA, Baby SM, MacFarlane PM, Hsieh Y-H, Knauss ZT, Bubier JA, Mueller D and Lewis SJ (2023) L-cysteine ethyl ester prevents and reverses acquired physical dependence on morphine in male Sprague Dawley rats. Front. Pharmacol. 14:1303207. doi: 10.3389/fphar.2023.1303207
Received: 27 September 2023; Accepted: 31 October 2023;
Published: 04 December 2023.
Edited by:
Francisco Lopez-Munoz, Camilo José Cela University, SpainReviewed by:
Christopher Freet, The Pennsylvania State University, United StatesMarilia Gag Pereira, Federal University of Alfenas, Brazil
Copyright © 2023 Bates, Getsy, Coffee, Baby, MacFarlane, Hsieh, Knauss, Bubier, Mueller and Lewis. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Stephen J. Lewis, c2psNzhAY2FzZS5lZHU=
†Present address: James N. Bates, Chief Medical Officer, Atelerix Life Sciences Inc., Charlottesville, VA, United States
Santhosh M. Baby, Translational Sciences Treatment Discovery, Galvani Bioelectronics, Inc., Collegeville, PA, United States