 Sandra Cotino-Nájera
Sandra Cotino-Nájera Luis A. Herrera
Luis A. Herrera Guadalupe Domínguez-Gómez
Guadalupe Domínguez-Gómez José Díaz-Chávez
José Díaz-Chávez
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Pharmacol. , 10 November 2023
Sec. Pharmacology of Anti-Cancer Drugs
Volume 14 - 2023 | https://doi.org/10.3389/fphar.2023.1287505
One of the primary diseases that cause death worldwide is cancer. Cancer cells can be intrinsically resistant or acquire resistance to therapies and drugs used for cancer treatment through multiple mechanisms of action that favor cell survival and proliferation, becoming one of the leading causes of treatment failure against cancer. A promising strategy to overcome chemoresistance and radioresistance is the co-administration of anticancer agents and natural compounds with anticancer properties, such as the polyphenolic compound resveratrol (RSV). RSV has been reported to be able to sensitize cancer cells to chemotherapeutic agents and radiotherapy, promoting cancer cell death. This review describes the reported molecular mechanisms by which RSV sensitizes tumor cells to radiotherapy and chemotherapy treatment.
Cancer treatment and therapy have improved significantly in recent years, increasing patients’ survival and quality of life. However, cancer remains one of the main diseases with the highest mortality worldwide (Siegel et al., 2022). This poor prognosis in cancer patients is partly due to the adverse effects and complications that limit the patient’s survival and quality of life when using cancer treatments. But the main reason behind the failure of the most used therapies, such as chemotherapy, is cancer cells’ intrinsic or acquired resistance to the drugs. For example, cancer cells can evade the toxicity of drugs by developing resistance to them, which prevents the patient from getting better (Longley and Johnston, 2005; Holohan et al., 2013). On the other hand, radiotherapy is another treatment frequently used in cancer patients. However, the acquisition of resistance of cancer cells to radiotherapy treatment is usually common in patients with glioma, prostate cancer (PCa), and melanoma. Making it an ineffective treatment for this type of cancer (Kma, 2013; Chang et al., 2016).
The molecular mechanisms of intrinsic or acquired resistance of cancer cells are multifactorial; these mechanisms can range from altered expression of transport proteins, increased ability to repair DNA damage or the ability to copy your DNA even with mutagenesis-induced errors caused by the same targeted therapies, high tolerance to stress conditions, defects or evasion of apoptotic processes through senescence, alterations in oncogene/tumor suppressor expression and reprogramming of metabolic pathways (Rebucci and Michiels, 2013; Sarmento-Ribeiro et al., 2019; Cocetta et al., 2021).
Therefore, chemoresistance and radioresistance are challenges in the field of oncology that need to be further explored in order to be avoided. That is why researchers are currently studying new compounds that, combined with standard therapies, can improve their effectiveness, enhance their action, and, in turn, reduce the adverse effects of antineoplastic drugs to obtain better results in cancer treatment. In this sense, the natural compound resveratrol (RSV) is a good candidate for its anticancer properties, especially when combined with other chemotherapeutic drugs. For example, it has been reported that RSV can reduce the risk of multidrug resistance (MDR) through multiple cellular targets involved in carcinogenesis and chemo/radioresistance (Varoni et al., 2016; Ferraz da Costa et al., 2020). This review describes the molecular mechanisms by which RSV achieves its chemo- and radiosensitizing effects in cancer.
Many studies have demonstrated the chemopreventive effects of natural compounds such as curcumin, silymarin, allicin, lycopene, ellagic acid, and RSV. Furthermore, it has been reported that combining these natural compounds with anticancer drugs improves the anticancer activity of the drugs and reduces their side effects (Cragg and Pezzuto, 2016; Catanzaro et al., 2018; Berretta et al., 2020).
RSV (3,4′,5-trihydroxy-trans-stilbene) is found in many plants and foods such as grapes, blueberries, peanuts, berries, cocoa, etc. (Burns et al., 2002; Biesalski, 2007; Ko et al., 2017). It is a phytoalexin produced in plants as a defense mechanism in response to pathogen attacks (fungal or bacterial infections) or environmental stress (such as UV irradiation, metal salts, etc.) (Dercks and Creasy, 1989; Cocetta et al., 2021). RSV can be found in cis or trans-isomeric forms and their glycosides, trans-piceid and cis-piceid (Ali et al., 2010; Varoni et al., 2016; Ferraz da Costa et al., 2020). Since the publication by Jang et al. (Jang et al., 1997), the first article on the anticancer properties of RSV, the field of cancer research has given great interest to this molecule. In addition, a wide variety of beneficial biological effects of RSV have been discovered and explored, including its antioxidant, anti-inflammatory, anticancer, cardio- and neuroprotective activity (Park and Pezzuto, 2015; Kuršvietienė et al., 2016). However, in vivo experimental models have demonstrated that RSV is rapidly metabolized and eliminated, which leads to low bioavailability of the compound. Following oral administration, RSV is absorbed by passive diffusion or via membrane transporters at the intestinal level and is then released into the bloodstream, where it can be detected as an unchanged or metabolized molecule. (Ferraz da Costa et al., 2020). Even though 75% of RSV has been shown to be absorbed orally, only 1% is detected in the blood plasma after all metabolism (Varoni et al., 2016; Chimento et al., 2019; Ferraz da Costa et al., 2020).
To improve the bioavailability of RSV other means of RSV transport have begun to be used to enhance its bioavailability, such as delivering RSV through nanocarriers like nanoparticles or using different strategies, such as combining RSV with other compounds (bio-enhancers) (Kucinska et al., 2014; Santos et al., 2019; De Vries et al., 2021; Baek et al., 2023). For example, in the study of Zhang et al., they developed a nanocarrier of RSV-loaded poly (ε-caprolactone)-poly (ethylene glycol) nanoparticles with an erythrocyte membrane. This system improved RSV’s poor water solubility and helped it escape the control of immune cells, improving its biocompatibility and tumor penetration in vivo models. Furthermore, they demonstrated for the first time that RSV could induce ferroptotic cell death in colorectal cancer by initiating lipid peroxidation and suppressing the expression of SLC7A11 and GPX4 (Zhang et al., 2022b).
Bioactive or bioenhancer compounds have also been used (piperine, quercetin, biflavone ginkgetin) that, in combination with RSV, improve bioavailability, solubility, absorption, and cellular permeability (De Vries et al., 2021; Jaisamut et al., 2021; Vesely et al., 2021).
Even in recent years, different synthetic derivatives of RSV (methoxylated, hydroxylated and halogenated), also known as prodrugs, have been developed to improve the bioavailability of RSV and its biological activities (Nawaz et al., 2017; Ferraz da Costa et al., 2020). Some examples of these are 3,5,4′-tri-O-acetyl-resveratrol (TARES) and resveratrol 3-O-β-D-glucopyranoside (De Vries et al., 2021).
On the other hand, some studies have reported that low daily doses of RSV have potent chemopreventive effects in vivo (Scott et al., 2012), which could be related to RSV conjugates or metabolites. Like many other polyphenols, RSV is metabolized by several enzymes, such as cytochrome P450 superfamily enzymes, sulfotransferases, and UDP-glucuronosyltransferases, to form conjugated (glucuronide and sulfated) metabolites. Unabsorbed polyphenols and their conjugates reach the lower gastrointestinal tract (cecum and colon) and interact with the intestinal microbiota. Dihydroresveratrol (DHR), lunularin (LUN), and 3,4′-dihydroxy-trans-stilbene are RSV metabolites derived from gut microbiota (Li et al., 2022). Interestingly, DHR and LUN have been shown to exert more potent antiproliferative and anti-inflammatory effects in renal and colonic cell lines, and it is suggested that DHR and LUN may contribute significantly to the chemopreventive properties elicited by RSV in the kidney and colon (Li et al., 2022).
RSV has a wide variety of biological activities, such as antioxidant, anti-inflammatory, antiviral, neuroprotective, cardioprotective, immunomodulatory, and anticancer activity (Kotecha et al., 2016; Nawaz et al., 2017; Giordo et al., 2022). A large amount of literature reports the anticancer effects of RSV (Behroozaghdam et al., 2022; Wu et al., 2022; Zucchi et al., 2023).
In fact, RSV exerts its antitumor effects through pleiotropic mechanisms of action (Repossi et al., 2020). Its ability to act on multiple targets has contributed to its usefulness as an anticancer agent (Varoni et al., 2016); in addition, in combination with other therapies (chemotherapeutics and radiotherapy, for example,), its ability to sensitize tumor cells resistant to such therapies has been demonstrated (El-Benhawy et al., 2021; Cheuk et al., 2022; Choi et al., 2022; Komorowska et al., 2022). Likewise, it has been seen that RSV protects healthy cells from the adverse effects of conventional agents (Ivanova et al., 2019). Therefore, its potential utility as an anticancer agent is quite attractive. Some RSV targets involved in the carcinogenesis process are exemplified in the following figure (Figure 1).
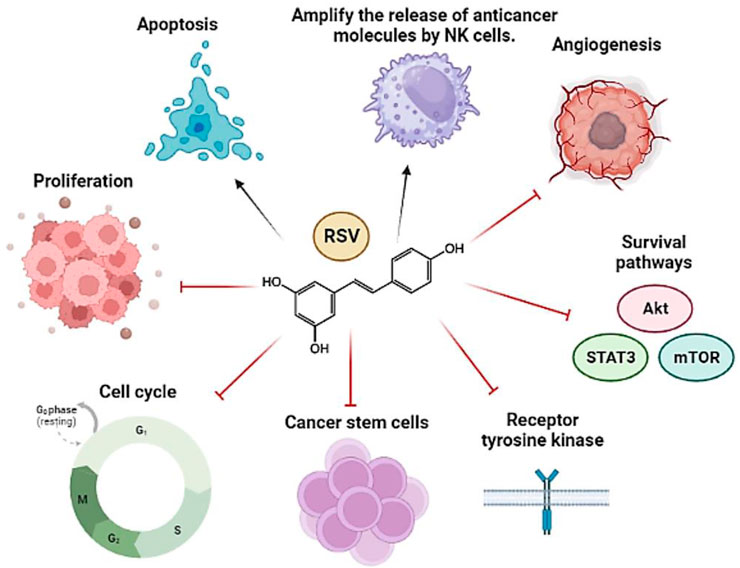
FIGURE 1. General representation of the main pathways and molecular targets affected by RSV in cancer. The arrows refer to promotion or increase, while the hammerhead lines refer to inhibition.
Due to its antioxidant and antimutagenic properties, RSV manages to prevent the onset of carcinogenesis. Likewise, it inhibits tumor growth and promotion by interfering with metabolic pathways such as glucose metabolism (Liu et al., 2018), inhibiting Cyclooxygenase-COX, and reducing the proliferative activity of cancer cells and their metastatic potential (Hu et al., 2016b). Furthermore, as shown by Schneider et al. (Schneider et al., 2001), RSV prevents the formation of colon tumors and reduces small intestine tumors in ApcMin/+ mice by decreasing the expression of genes directly involved in the progression of the cell cycle and cell proliferation (e.g., Cyclin D1 and D2). Moreover, RSV positively regulates genes activating immune cells such as HLA, FasL, and FOXP3, among others (Schneider et al., 2001; Fontenot et al., 2003; Ashrafizadeh et al., 2021).
On the other hand, several studies have shown the antitumor capacity of RSV by interfering with different signaling pathways such as STAT3, PI3K/Akt/mTOR, Wnt, insulin growth factor (IGF), SIRT1/AMPK, etc. (Habibie et al., 2014). In addition, RSV has also been reported to decrease NF-ĸB phosphorylation and acetylation, causing deficiencies in factors involved in tumor invasion and metastasis (I-CAM, AP-1, VEGF). The physical interaction between NF-ĸB and SIRT1 is involved in the anticancer activity of RSV (Buhrmann et al., 2016). SIRT1 inhibits NF-ĸB signaling by deacetylating the p65 subunit of the NF-ĸB complex. Furthermore, SIRT1 stimulates oxidative energy production by activating AMPK, PPARα, and PGC-1α, inhibiting NF-ĸB signaling and suppressing inflammation (Shao et al., 2015; He et al., 2017; Zhao et al., 2017). Also, RSV has been shown to suppress the growth of HCT116 colorectal cancer cells by inhibiting SIRT1-dependent NF-ĸB, in addition to inducing apoptosis in ls174t cells through the induction of the expression of the proapoptotic protein Bax inhibits the anti-apoptotic protein Bcl-2 (B-cell lymphoma 2) (Chen et al., 2009; Buhrmann et al., 2016).
In another critical study, Reagan-Shaw et al. (Reagan-Shaw et al., 2004) revealed that RSV significantly inhibits the induction of epidermal hyperplasia, mediated by exposure to UVB radiation through the decrease in proliferating cell nuclear antigen (PCNA), CDK-2, -4, and -6, as well as Cyclins-D1 and D2 in SKH-1 mouse cells. On the other hand, RSV in HaCaT cells inhibits cell proliferation by inhibiting the PI3K/AKT/mTOR pathway (Fabbrocini et al., 2010; Kisková and Kassayová, 2019). Another study reported that RSV suppresses cell growth and induces apoptosis in Colo16 squamous epidermal cancer cells (SCC) by inactivating Wnt and its target genes (survivin, c-Myc, cyclin D1, and VEGF). In addition, RSV increases the expression of the Wnt signaling inhibitor (Axin2) (Liu et al., 2017).
On the other hand, the role of resveratrol as an epigenetic regulator is very important in its anticancer activity. Let us remember that DNA hypermethylation (catalyzed by specific DNA methyltransferases (DMNT)) and histone deacetylation (mediated by histone deacetylases (HDACs) are key epigenetic mechanisms for the silencing and repression of many genes, including those involved in cell cycle regulation, DNA repair, inflammatory response, and apoptosis. Multiple studies have described the ability of RSV to increase or decrease the methylation of genes involved in tumorigenesis. For example, it has been shown that RSV can decrease the methylation of the promoters of the PTEN and BRCA-1 genes. Likewise, it has been seen that the methylation of the tumor suppressor gene RASSF1A and IL-10 decreases. And RSV can also increase the methylation of oncogenes such as AURKA, CCNB1, and HK2 (Lee et al., 2018). Additionally, RSV has also been involved in the deacetylation of genes such as p53 through SIRT1 (Lee et al., 2018; Rajendran et al., 2022).
Interestingly, it has also been reported that RSV modulates the expression of some miRNAs (short non-coding RNA) and lncRNAs (long non-coding RNA), which regulate the expression of genes involved in the malignant phenotype of cancer (Wang et al., 2019b; Asemi et al., 2023). For example, oncogenic miRNAs such as miR-19, miR-21, and miR-30a-5p have significantly decreased by RSV treatment in glioma (GBM) cells, modifying the expression of their target genes such as p53, PTEN, STAT3, NF-ĸB, COX-2 (Cocetta et al., 2021). On the other hand, lncRNAs have been identified as possible targets of RSV: MEG3 and ST7OT1 in GBM cell lines, U251 and U87, which increase with RSV treatment and induce apoptosis and necrosis of both cell lines. The MEG3 and ST7OT1 lncRNAs act as tumor suppressor genes. Ectopic expression of MEG3 and ST7OT1 inhibits cell proliferation and promotes apoptosis in human GBM cell lines (Wang et al., 2012; Liu et al., 2015). In contrast, RSV has been reported to decrease the expression of the lncRNA MALAT1 in colorectal and gastric cancer cells through the Wnt/β-catenin signaling pathway (Ji et al., 2014; Yang et al., 2018). The inhibition of MALAT1 expression by RSV is relevant as it is involved in the progression and metastasis of various types of cancer, including colorectal, gastric, lung, and hepatocarcinoma (Lai et al., 2012; Gutschner et al., 2013; Ji et al., 2014; Yang et al., 2018).
In a recent study, RSV in both its cis and trans forms was shown to inhibit the activity of the Anoctamin1 (ANO1) channel (a calcium-activated chloride channel, which is involved in the proliferation, migration, and invasion of various types of cancer, including head and neck squamous cell carcinoma, lung cancer, and prostate cancer) (Carter et al., 2014). In addition, they showed how RSV also decreased the expression of ANO1 protein and mRNA in PC-3 prostate cancer cells (Jeon et al., 2023).
Cells constantly struggle with external stress and damage, which can result in mutations or severe cellular alterations if there is no successful repair. Usually, when there is very serious damage, the cell commits suicide to avoid further destruction and to eradicate genetically unstable and dangerous cells. However, if the cell death mechanism is not working properly, “malignant” cells can begin to proliferate, ultimately resulting in a tumor (cancer) (Zaitceva et al., 2021). Cancer remains one of the world’s leading causes of death, generating enormous costs and burdening humanity. The annual number of cancer cases worldwide is projected to increase from 19.3 million in 2020 to 30.2 million in 2040 (UICC Global Cancer Control, 2023).
The main goal of cancer treatment is the elimination of malignant cells through the induction of cell death. However, cancer cells constitute important barriers to clinical therapies due to their heterogeneity and plasticity. Resistance to cell death is one of cancer’s main characteristics, allowing the uncontrolled multiplication of cancer cells (Zaitceva et al., 2021). During the last years, several mechanisms have been described by which cancer cells can avoid cell death and acquire resistance to current treatments (surgery, radiotherapy, chemotherapy, targeted therapy, and immunotherapy). Among these are the overexpression of antiapoptotic proteins (Bcl-2) and the inactivation of p53 (Carneiro and El-Deiry, 2020). Oncologists point out that classical chemotherapy and radiotherapy are already reaching the limits of their effectiveness, so other methods or alternatives are needed to improve their effectiveness against cancer (Papież and Krzyściak, 2021).
In this review, we describe the signaling pathways and cellular mechanisms that lead to the development of chemoresistance and radioresistance in cancer cells.
Below is a table with information regarding the resistance that some types of cancer must radio and chemotherapy. It is worth mentioning that it is not yet known exactly which types of cancer will be resistant or sensitive to the therapies with which they will be treated since, as mentioned above, resistance can also be acquired during the treatment process, and multiple factors are involved in this. The development of resistance (tumor microenvironment, signaling pathways, cell-cell interactions, changes or mutations at the genetic and epigenetic level). However, this table was prepared based on articles and studies where emphasis is placed on the most studied specific types of cancer that tend to present or be more resistant to a specific therapy (Table 1).
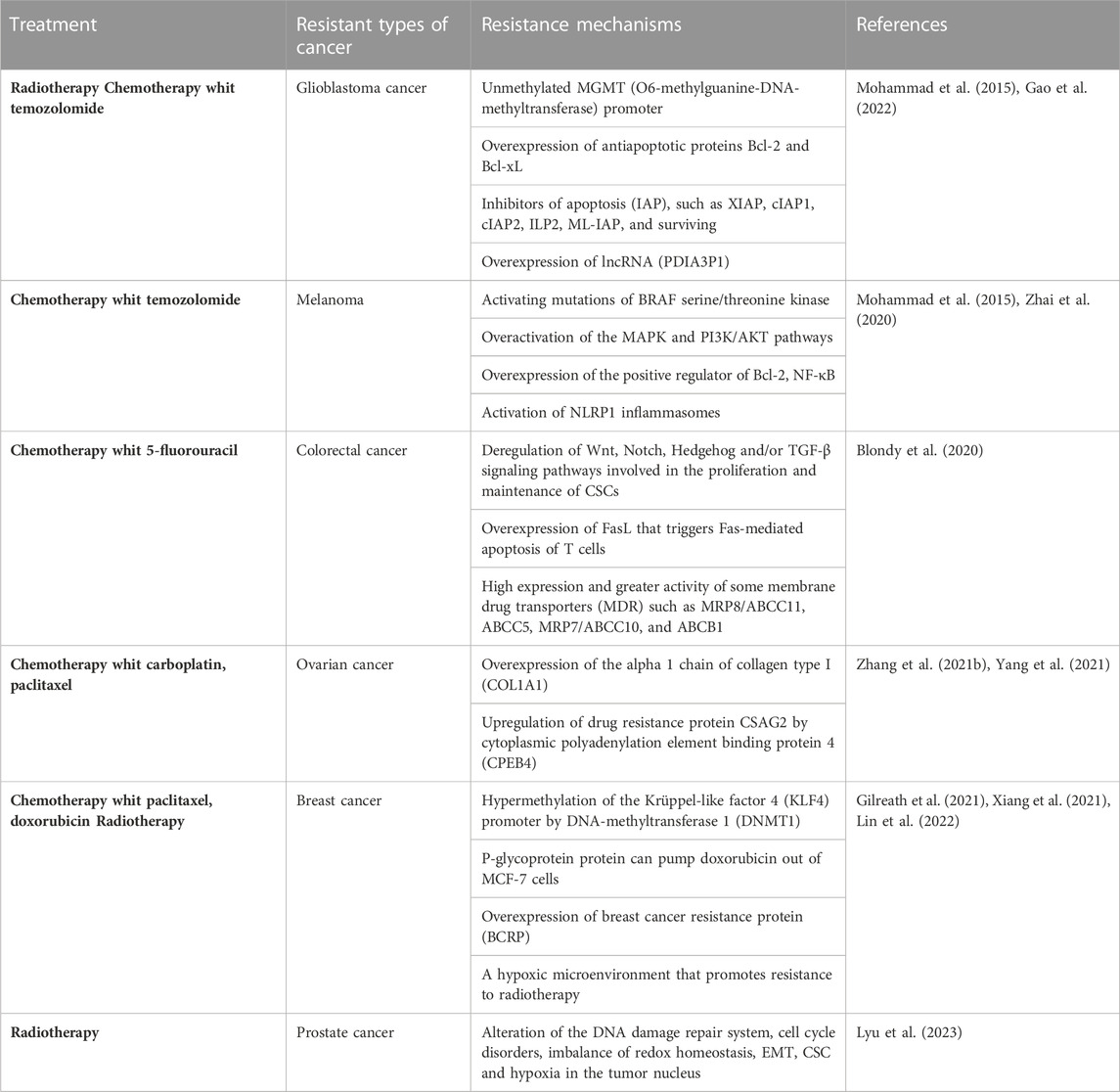
TABLE 1. Mechanisms of resistance to chemo and radiotherapy in cancer.
The characteristics of cancer cells that promote resistance to therapies, currently more described, are the following. Some cancer cells are said to resemble stem cells, defined as cancer stem cells (CSCs); These cells frequently change during tumor progression and after therapeutic exposures, favoring their resistance and progression (Basu et al., 2022). Tumors have also been shown to harbor a population of slow-cycling cells (SCCs) that are not in the proliferative cell cycle and are inherently refractory to antimitotic drugs. However, they can stochastically re-enter the proliferative cell cycle or respond to mitogenic stimuli. Like CSCs, SCCs can evade the immune system and survive cancer treatments, thereby influencing treatment failure, tumor recurrence, and metastasis. Unlike CSCs, SCCs represent a population of transient cells that haphazard go in and out of the G0/G1 phase very quickly (Basu et al., 2022).
On the other hand, a group of cancer cells acquires the ability to resist therapies through anastasis. Anastasis is, in a few words, the arrest of apoptosis and the ability that cells acquire to maintain themselves in a state of senescence after treatment. After this, they preserve and increase their proliferative capacity, making anastasis undesirable during cancer therapy (Zaitceva et al., 2021). Cancer cells enter the anastasis process because some of the mitochondria of these cells remain intact during the apoptosis process; this is by increasing the levels of Bcl-2/Bcl-xL, which inhibit the process of permeabilization of the outer membrane of mitochondria (MOMP). Allowing these mitochondria to repopulate in the cell (Peña-Blanco and García-Sáez, 2018). A limited number of mitochondria undergo MOMP; the amount of cytochrome c released is insufficient to trigger apoptosis but sufficient for sublethal activation of caspase and consequent activation of endonuclease, leading to genome instability (Zaitceva et al., 2021).
On the other hand, polyploid giant cancer cells (PGCC) are found in several types of cancers and have been seen to play an important role in resistance to treatments such as radiotherapy and chemotherapy. These cells are characterized by having multiple nuclei or a single giant nucleus with multiple complete sets of chromosomes. In addition, they contribute to the immortality, invasion, and metastasis of tumors. For example, Was et al. showed how polyploidy develops in response to various genotoxic stresses, such as chemotherapy, radiation, hypoxia, oxidative stress, or environmental factors such as air pollution, ultraviolet light, or hyperthermia (Liu et al., 2022; Was et al., 2022). The general mechanism leading to PGCC formation could be a consequence of endoreplication, which is related to genetic or physical disorders of mitosis, cell fusion, or cell cannibalism. An important evolutionary feature of polyploid cancer cells is the generation of aneuploid clones during depolyploidization, which expands cancer cells’ genetic repertoire, allowing them further development and expansion (Mittal et al., 2017; Liu et al., 2022; Pienta et al., 2022; Was et al., 2022).
Chemoresistance and radioresistance mechanisms in tumor cells can be intrinsic (cells are resistant before treatment) or acquired (resistance develops during treatment). Below, we describe some of the most important mechanisms and signaling pathways (Figure 2), but for a more detailed review, you can consult the works of Fodale et al., 2011; Chang et al., 2016; Mashouri et al., 2019; Ali et al., 2020; Zaitceva et al., 2021; Fodale et al., 2011; Chang et al., 2016; Mashouri et al., 2019; Ali et al., 2020; Zaitceva et al., 2021).
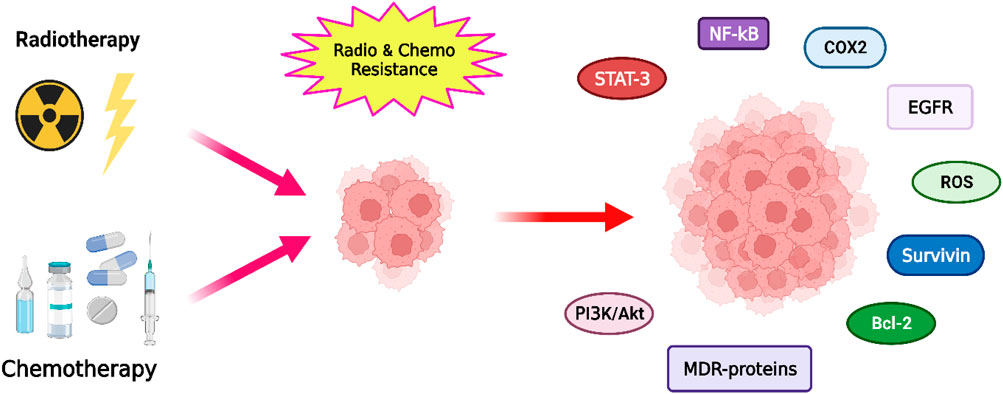
FIGURE 2. Representation of the main pathways, factors and proteins involved in the development of radioresistance and chemoresistance in cancer cells.
Members of the tumor necrosis factor (TNF) superfamily are important inducers of apoptosis, contributing to the death response in tumor cells. However, they are also involved in developing adverse reactions such as resistance to anticancer drugs; these effects are mediated by the production of ROS (Garg and Aggarwal, 2002; Blaser et al., 2016; Cruceriu et al., 2020).
The release of TNF stimulates the activation of the vascular endothelium, release of nitric oxide, recruitment of inflammatory cells, immunoglobulins, complement and alters the permeability of the mitochondrial membrane, favoring the release of cytochrome C and the subsequent activation of caspases, which, in turn, leads to apoptosis. Furthermore, TNF signaling is directly related to the mitochondrial electron transport mechanism and ROS production (Chandel et al., 2001). ROS are an upstream component that activate the process of TNF-induced apoptosis, followed by caspases, mitogen-activated protein kinases (MAPKs), NF-κB, and activation of the transcription factor AP-1 (Garg and Aggarwal, 2002). Oxidative stress is essential in tumor development and cancer therapy (Gorrini et al., 2013). It has been reported that ROS can promote oncogenic mutations and epigenetic changes in cancer cells. These alterations lead to the loss of tumor suppressor genes, accelerated cell metabolism, and altered cell sensitivity to anticancer drugs (Pelicano et al., 2004; Panieri and Santoro, 2016; Priya et al., 2017).
ROS are generated by redox-sensitive pro-survival signaling pathways (oxidation-reduction reactions), which function as intermediates in the transduction of various extracellular signals (Vinod et al., 2013). However, there is a complex intracellular redox network to protect cells against oxidative stress, where several signaling pathways can be activated in the adaptive response of ROS (glucose metabolism, PI3K/Akt pathway, MAPK pathway). These signaling pathways play a critical role in protecting cancer cells against the cytotoxic effects of antineoplastic agents, leading to chemoresistance (Marengo et al., 2019). For example, in chemoresistant epithelial ovarian cancer (EOC) cells, an increase in ROS and apoptosis was observed with the combination of Dactolisib (BEZ235) and cisplatin treatment, which also inhibited the PI3K/Akt/mTOR signaling pathway, reversing epithelial-mesenchymal transition (EMT) and decreasing CSC marker expression compared to cisplatin monotherapy (Deng et al., 2019).
On the other hand, ROS-mediated genotoxic stress has been shown to be involved in NaAsO2 -induced cell cycle arrest, decreased stemness, and chemoresistance of prostate cancer cells (PC-3 and DU145) (Zhang et al., 2021c).
NF-κB is activated by different molecules and signaling pathways, and in turn, this transcription factor modulates the expression of several genes (more than 500 genes) that participate in inflammatory responses, cell differentiation, adaptation to stress, apoptosis, immunity, and one of the main effector pathways that regulate the amount of ROS that leads to cell survival and at the same time to chemoresistance in cancer (Morgan and Liu, 2011; Vinod et al., 2013).
Among the many diseases related to aberrant NF-κB activation, cancer has been the main focus due to the role of NF-κB as a central regulator of the regulation of genes involved in cell survival (Bcl-2, Bcl-xL, inhibitors of apoptosis proteins [IAP] and superoxide dismutase) (Garg et al., 2005) and tumor progression (intercellular adhesion molecule 1 [ICAM1], vascular cell adhesion molecule 1 [VCAM1], leukocyte-endothelial adhesion molecule 1 [PECAM- 1], vascular endothelial growth factor [VEGF], hypoxia-inducible factor [HIF-1], Cyclooxygenase-2 [COX-2], inducible nitric oxide synthase [iNOS], and matrix metalloproteinase [MMP-9]). Furthermore, NF-κB activity is frequently elevated in many tumor types, including leukemia, lymphoma, prostate cancer, breast cancer, colon cancer, melanoma, and head and neck cancer (Garg et al., 2003; Dolcet et al., 2005; Fan et al., 2013; Wu et al., 2017).
NF-κB can be aberrantly activated in cancer by chemotherapeutic agents and ionizing radiation, response to stress, and induced cell death, leading to treatment-induced resistance of tumor cells (Jung and Dritschilo, 2001; Garg et al., 2003; Sampepajung et al., 2021). In vitro and in vivo studies have shown that NF-κB inhibits chemotherapy-induced apoptosis in various tumor types (Bharti and Aggarwal, 2002; Sampepajung et al., 2021). For example, the upregulation of NF-κB-inducible genes has been shown to protect MDA MB-231 breast cancer cells from apoptosis induced by paclitaxel and ionizing radiation (Newton et al., 1999). Similarly, in another study, they treated cell lines of different types of cancer [Hep-3B (liver), AGS (gastric), SiHa (cervical), MCF7 (breast), NTUB1 (bladder), and H460 (lung-non-small cells)] with different chemotherapy regimens (doxorubicin, 5-fluorouracil [5-FU], cisplatin, and paclitaxel), and found a correlation between cell survival with the level of drug-induced NF-κB activity (Chuang et al., 2002). On the other hand, in a study where the inhibitory subunit of NF-κB was transfected, an increase in chemotherapeutic efficacy was observed in vitro and in vivo models of gastrointestinal neoplasms (Cusack et al., 2000).
Another interesting study is that of Park M. et al. (Park et al., 2016); they evaluated the role of protein tyrosine kinase 7 (PTK7) in ESCC resistance to radiotherapy. They observed that PTK7 plays an important role in ESCC radioresistance through activation of the NF-ĸB pathway. In addition, these authors reported an increase in the IAPs, XIAP, and survivin, encoded by NF-ĸB regulated genes, which was associated with radioresistant cells but not in radiosensitive cells; nevertheless, PTK7 knockdown downregulated IAP expression (Park et al., 2016).
COX-2 is an enzyme expressed primarily in response to inflammatory disorders and cancer. It is responsible for mediating the production of prostaglandins and is under clinical investigation as a target for cancer therapy (Milas, 2001; Nakata et al., 2004; Gowda et al., 2017). Over-expression and activity of COX-2 have been associated with more aggressive tumor phenotypes and worse prognosis in patients with breast, colon, head, neck, lung, and pancreatic cancer (Wang et al., 2014; Li et al., 2020). Furthermore, evidence suggests that COX-2 is involved in multiple aspects of carcinogenesis, including tumor growth, metastatic spread, and resistance to various therapies (Harris, 2009; Tong et al., 2018). As a result, some scientists have investigated the usefulness of selective COX-2 inhibitors, such as SC-236 and Celecoxib, in vitro and in vivo to test whether their inhibition can sensitize tumor cells to make treatment more efficient against cancer using chemotherapy and radiotherapy. For example, SC-236 increased the response to radiotherapy in various murine tumor models (you can put here which tumor types) and in a human GBM xenograft in nude mice (Kishi et al., 2000; Petersen et al., 2000). On the other hand, the combination treatment with celecoxib and imatinib resulted in a significant decrease in cell viability and an increase in caspase-3 enzyme activity in HT-29 colon cancer cells (Atari-Hajipirloo et al., 2016).
In another study, celecoxib and afatinib co-treatment inhibited the expression of COX-2 and EGFR, which led to increased sensitization of A549 lung cancer cells to radiotherapy and apoptosis (Zhang et al., 2021a).
The development of resistance to cell death mechanisms, specifical death by apoptosis, is one characteristic that distinguishes tumor cells and plays an important role in developing resistance against anticancer agents.
Bcl-2 is an anti-apoptotic protein that is overexpressed in several solid and hematopoietic tumors, and that also exerts its influence by improving cell survival (Kaufmann and Vaux, 2003), which contributes to resistance to conventional treatments, including chemotherapy and radiotherapy (Deng et al., 2000; Maji et al., 2018). In addition, several studies have shown that Bcl-2 inhibition sensitizes tumor cells to chemotherapy and radiotherapy. For example, transfection with the PTEN gene, which negatively regulates Bcl-2, potentiated the effects of radiation therapy on several prostate cancer cell lines (PC-3-Neo, PC-3-Bcl-2, and LNCaP) (Rosser et al., 2004). Another study that used an antisense oligonucleotide against Bcl-2 observed an increase in apoptosis and greater chemotherapeutic efficacy in a thyroid carcinoma cell line (Kim et al., 2003). These results and others have highlighted Bcl-2 as a potential target for chemosensitization and radiosensitization (Belka and Budach, 2002; Gutiérrez-Puente et al., 2002). For example, in the study carried out by Lu L et al. (Lu et al., 2018), in the MB-468 radioresistant breast cancer cell line, they observed low levels of ROS and higher levels of STAT3 and Bcl-2 proteins; on the other hand, when they added Niclosamide, a potent STAT3 inhibitor, radioresistance was overcome by inhibiting STAT3 and Bcl-2 and inducing ROS (Lu et al., 2018).
On the other hand, the tumor suppressor protein p53 is a key factor in inducing cell cycle arrest, DNA repair, and apoptosis in response to cellular stress. Unfortunately, it is known that in approximately more than 50% of cancerous tumors, p53 is mutated (Levine, 1997). Consequently, there is increased survival and proliferation of cancer cells. In addition, there is evidence suggesting that inactivation of the p53 wild-type protein results in increased chemo-resistance to several chemotherapeutic drugs, including doxorubicin, cisplatin, 5-fluorouracil (5-FU), and etoposide (Ferraz da Costa et al., 2012).
Moreover, the mutant p53 gain of function can induce chemoresistance, increasing drug efflux and metabolism, survival promotion, apoptosis inhibition, upregulation of DNA repair, autophagy suppression, microenvironmental resistance elevation, and the induction of cancer stem cells (He et al., 2017). However, recently, a subset of cancer cells, regardless of their p53 status, exhibited resistance to chemotherapy through the p21 protein. p21 is a transcriptional target of p53 that is induced upon DNA damage and acts to arrest the cell cycle by inhibiting cyclin-dependent kinases (CDKs) (Ashraf et al., 2019).
Hsu et al. Investigate the proliferation-senescence decision in response to chemotherapy and elucidate how early p21 dynamics predict and shape cell fate. They concluded that cells with high or low levels of p21 during doxorubicin treatment are destined to become senescent, while those with an intermediate amount of p21 proliferate after drug washout (Ashraf et al., 2019; Hsu et al., 2019). This data is undoubtedly important for stimulating senescence in the context of cancer therapies.
Survivin is part of the mammalian IAP family, and its main function is to inhibit the apoptosis pathway by blocking the activation of caspases 3, 8, and 9 (Salvesen and Duckett, 2002). Both in vitro and in vivo experiments have shown cancer-inducing properties of surviving (Bao et al., 2002), as well as overexpression in various types of cancer and absence in most normal tissues (Altieri, 2003). Survivin expression has been shown to increase in VEGF-stimulated vascular endothelial cells (O’Connor et al., 2000; Mesri et al., 2001; Tran et al., 2002). In addition, high levels of surviving expression have been associated with a high rate of tumor recurrence, poor overall patient survival, and high tumor resistance to chemotherapy and radiotherapy in several cancers, including lung, breast, colon, stomach, esophagus, pancreas, liver, uterus, and ovary cancer among others (Altieri, 2003). Nestal de Moraes et al. (Nestal de Moraes et al., 2015) demonstrated that the transcription factor FOXM1 upregulates the expression of the anti-apoptotic genes XIAP and Survivin, which contributes to the development of drug resistance and is associated with poor clinical outcomes in breast cancer patients (Nestal de Moraes et al., 2015).
In contrast, survivin inhibition has been reported to sensitize breast cancer cells to paclitaxel, etoposide, doxorubicin, and cisplatin (O’Connor et al., 2000; Wall et al., 2003; Mita et al., 2008; Lyu et al., 2018; Minaiyan et al., 2021); furthermore, survivin inhibition in combination with radiotherapy resulted in a significant decrease in lung cancer cells survival (Lu et al., 2004).
Since the antitumor agent must reach the cancer cell in an adequate concentration to exert its effect, drug uptake or release alterations could also be responsible for the acquisition of chemoresistance (Huang and Sadée, 2006). Transport proteins, also called ATP-dependent multidrug transporters (ABCs), associated with chemoresistance are multidrug resistance proteins (MDR1; P-glycoprotein [P-gp]; MRP1; ABCB1; ABCC1) (Leonard et al., 2003; Crouthamel et al., 2006; Pérez-Gutiérrez et al., 2007), the multidrug resistance-associated protein (MRP1) (Hong et al., 2019), the protein related to lung resistance (LRP) (Schneider et al., 2001; Wang, 2011) and breast cancer resistance protein (BRCP) (Fu et al., 2020). MDR proteins are ATP-binding proteins that regulate P-gp, which are responsible for removing drugs from cells using ATP hydrolysis (Yang et al., 2014).
Many drugs, including daunorubicin (DRN), imatinib, nilotinib, taxol, and doxorubicin, among others, can be expelled from cancer cells that overexpress P-gp and multidrug resistance-associated protein 1 (MRP1) transporter (Gottesman et al., 2002; Kosztyu et al., 2014). For example, in one study, high expression and activity of MRP1 were observed in primary cultures of glioblastoma multiforme biopsies (Quezada et al., 2011). Also, in PC3 and DU145 human prostate cancer cell lines, increased expression of MRP1 in prostate cancer cells is related to resistance to chemotherapy. Similarly, the blockade of MRP1 function by leukotriene receptor antagonists (MK-571 and zafirlukast) led to an intracellular accumulation of the MRP1 substrate and increased sensitivity to cytotoxic drugs (van Brussel and Mickisch, 2003). This result was consistent with the study where they used an NF-ĸB inhibitor to inhibit MDR protein expression, leading to increased apoptosis in prostate cancer cells (Flynn et al., 2003).
On the other hand, elevated levels of ABCB1 have also been shown to be associated with paclitaxel resistance in human osteosarcoma (OS) cell lines, which developed cross-resistance with other ABCB1 substrates, such as doxorubicin, docetaxel, and vincristine (Yang et al., 2014). In addition, the involvement of ABCB1 overexpression in doxorubicin resistance in human OS cells was demonstrated by downregulation or abrogation of ABCB1 expression, which resulted in the restoration of doxorubicin sensitivity (Fanelli et al., 2016; Liu et al., 2016; Serra et al., 2021).
Moreover, the study by Ranibar S et al. (Ranjbar et al., 2019) used compounds derived from 5-oxo-hexahydroquinoline that they named D6, D5, and D3 (which have 3-chlorophenyl, 2,3-dichlorophenyl and 4-chlorophenyl substituents in the C4 position of the 5-oxo-hexahydroquinoline core) these compounds inhibit P-gp, MRP1, and BCRP, respectively; causing a reversal of drug resistance (Doxorubicin, mitoxantrone) at concentrations of 1–10 μM, in human uterine sarcoma cells (MES-SA) sensitive and resistant to drugs with P-gp overexpression.
Protein kinase B (PKB or Akt) is a downstream effector of PI3K and has been described as a mediator of anti-apoptotic signaling in cancer cells. In addition, Akt overexpression has been shown to promote cell cycle progression and tumorigenesis (Zhan and Han, 2004). Akt may contribute to chemoresistance and radioresistance: for example, over-expression of Akt1 has been reported to result in increased resistance of lung cancer cells (describe which cell lines) against a panel of various chemotherapeutic agents (doxorubicin, cisplatin, and mitoxantrone) (Hövelmann et al., 2004). Accumulating evidence to date has suggested that the PI3K/Akt pathway may also be an essential contributor to radioresistance (Zhan and Han, 2004). In this regard, Akt activation in bile duct cancer cells has been shown to be associated with radioresistance, which was demonstrated through indirect inhibition of Akt activation with a PI3K inhibitor (LY294002) (Tanno et al., 2004).
On the other hand, in a recent study, the PI3k/Akt/mTOR pathway was associated with the increase and activation of PDK1, which is associated with radioresistance, motility, and invasiveness of hepatocellular carcinoma. Furthermore, it was observed that pharmacological inhibition of PDK1 in Huh7 cells mediated by BX795 synergistically enhances the radiosensitivity of these cells, increases the apoptotic Bax/Bcl-2 ratio, and abolishes oncogenicity and clonogenicity (Bamodu et al., 2020).
Another study showed that the inactivation of AKT signaling inhibited tumorigenesis and radioresistance mediated by CPNE1 in triple-negative breast cancer cells. Knockdown of CPNE1 also inhibited tumor growth and promoted cell apoptosis in vivo in mouse xenografts (Shao et al., 2020).
In addition, the participation of the PI3K/AKT/mTOR pathway in chemoresistance has also been reported. For example, in the study by Qiu C et al. (Qiu et al., 2020), they show that MNAT1, a cyclin-dependent kinase-activating kinase (CAK) complex, contributes to OS cell resistance to cisplatin via the PI3K/AKT/mTOR. MNAT1 is highly expressed in various types of cancer and is involved in the molecular pathogenesis of cancer and drug resistance (Qiu et al., 2020).
STAT3 is a member of the STAT family of transcription factors that are activated by tyrosine phosphorylation through signaling mediated by receptors such as epidermal growth factor receptor (EGFR), platelet-derived growth factor (PDGF), and cytokines such as interleukin-6 (IL-6) (Akira, 2000; Levy and Darnell, 2002). It is established that IL-6 is produced in an autocrine or paracrine manner and plays an essential role in the malignant progression of various types of cancer, including multiple myeloma (MM), by regulating the growth and survival of tumor cells. The presence of IL-6 leads to constitutive activation of STAT3, resulting in the expression of high levels of the anti-apoptotic protein Bcl-xL (Catlett-Falcone et al., 1999; Bharti et al., 2004). Therefore, STAT3 has various biological functions, including regulation of cell growth, apoptosis, and cell differentiation. STAT3 has also been shown to be permanently active in various human cancers and is required for tumor cell proliferation (Bromberg, 2001). Furthermore, it has been reported that STAT3 can mediate chemoresistance, and its inhibition can sensitize cells to apoptosis. For example, in the study by Bharti et al. (Bharti et al., 2004), inhibition of STAT3 contributed to decreased survival of multiple melanoma cells and sensitized pancreatic cancer cells to apoptosis (Greten et al., 2002). Blockade of STAT3 using various techniques sensitized breast cancer cells (MDA-MB435) to apoptosis induced by taxol and adriamycin (doxorubicin) chemotherapy (Real et al., 2002). Other studies also show that STAT3 inhibition increases radiosensitivity in different tumors such as hepatocellular carcinoma, squamous cell carcinoma of the head and neck, gastric cancer, pancreatic cancer, etc., (Adachi et al., 2012; Bu et al., 2013; Huang et al., 2014; 2016; Lee et al., 2019; You et al., 2019; Wang et al., 2020b).
EGFR is a transmembrane glycoprotein with intrinsic tyrosine kinase activity. By binding with EGF, it regulates a signaling cascade that, in turn, regulates cell growth and proliferation. Similarly, it activates molecular pathways involved in various cellular processes, such as cell differentiation, survival, and transformation (Yarden and Sliwkowski, 2001). EGFR overexpression has been related to more aggressive tumor phenotypes, poor patient prognosis, and lack of response to antitumor therapies (Wang, 2017). For example, increased EGFR expression is associated with increased tumor chemoresistance and radioresistance in tumors, including squamous cell carcinoma, ovarian adenocarcinoma, hepatocarcinoma, glioblastoma, and adenosquamous carcinoma of the cervix (Akimoto et al., 1999; Milas et al., 2000; Nasu et al., 2001; Wang et al., 2020a). On the other hand, various investigators have also reported increased sensitization of tumor cells to radiotherapy through EGFR inhibition in head and neck squamous cell carcinoma (SCC), human colon cancer (GEO), colon cancer, ovarian (OVCAR-3), glioblastoma multiple (Huang et al., 1999; Bianco et al., 2000; Huang and Harari, 2000; Wang et al., 2020a).
Further studies have confirmed that the EGFR/PI3K signaling pathway plays an important role in tumor chemoresistance (Zhang et al., 2019b). showed that p53 sensitized cisplatin-chemoresistant NSCLC (Non-small cell lung cancer) by suppressing the EGFR/PI3K signaling pathway. Similarly, when miR-7 inhibited the EGFR/PI3K signaling pathway, adriamycin sensitivity in breast cancer (MCF-10 and MCF-7/ADR) increased (Huang et al., 2019).
It has also been reported that chemoresistance may be mediated by the glutathione/glutathione S transferase (GSH/GST) system (Zhu et al., 2006). An essential function of GSH is the detoxification of xenobiotics and some endogenous compounds, maintaining intracellular redox balance. These substances are electrophilic and form conjugates with GSH, either spontaneously or enzymatically, in reactions catalyzed by GSH-GSTs (Traverso et al., 2013). Several studies have shown a relationship between the resistance of tumor cells to chemotherapy drugs and an increase in the expression of GSH, GST, and GPx (Chao et al., 1992; Buser et al., 1997).
In contrast, low levels of GSH, GST, and GPx have been found to be associated with favorable clinical features and a good prognosis. In contrast, high GSH and GST activity levels were associated with more aggressive or more advanced disease in tissue samples from women with breast cancer (Buser et al., 1997). In fact, cancer cell lines containing low levels of GSH are much more sensitive to ionizing radiation than cells that overexpress GSH (Meister, 1991).
Increased GSH is an important factor contributing to drug resistance by binding or reacting with drugs, interacting with ROS, preventing protein or DNA damage, or participating in DNA repair processes. For example, GSH depletion and GGT inhibition in melanoma cells significantly increased cytotoxicity through oxidative stress (Benlloch et al., 2005). In addition, cells that overexpress GGT have been shown to be more resistant to hydrogen peroxide and to drugs such as doxorubicin (Hochwald et al., 1997), cisplatin (Godwin et al., 1992), and 5-fluorouracil (McLellan and Wolf, 1999).
Direct or indirect alterations in DNA are the basis of the mechanism of action of many drugs used in cancer therapy. Therefore, increased DNA repair activity compromises the damage induced by chemotherapeutic agents, resulting in chemoresistance (Sakthivel and Hariharan, 2017). Tumor cells have obtained a great capacity to repair damaged DNA through multiple pathways, such as mismatch repair (MMR), base excision repair (BER), nucleotide excision repair (NER), repair by non-homologous end joining (NHEJ), homologous recombination (HR) repair among others. For example, the BER repair pathway is involved in colon cancer resistance to temozolomide chemotherapy (Liu et al., 1999). The “excision repair cross-complement protein 1” (ERCC1), which belongs to the NER repair pathway, has also been reported to be associated with chemoresistance to platinum-based anticancer agents of various tumors, including cancer lung, colon, and breast cancer (Youn et al., 2004).
On the other hand, the Werner syndrome protein (WRN), a DNA helicase vital for the regulation/activation of NHEJ and HR repair, as well as the maintenance of DNA telomere stability, is found to be overexpressed in glioblastoma multiforme cancer cells and is also associated with increased resistance to chemotherapeutic agents, especially cisplatin (Lee et al., 2016). It has also been reported that the ectopic expression of the HOTAIR protein (antisense RNA of the HOX transcript), an important regulator of the transcription factor NF-ĸB, is associated with a more significant DNA damage response to cisplatin treatment and also a greater chemoresistance in ovarian cancer cells (IGROV, OVSAHO, OVMUNA, SKOV3, A2780, HEYC2, A2780-CR5, and OV90) (Özeş et al., 2016). Otherwise, it has been reported that BRCA1, another important protein in homologous recombination repair, activates NF-ĸB in response to topoisomerase inhibitor drugs such as etoposide or camptothecin; NF-ĸB, for its part, transcriptionally activates anti-apoptotic proteins such as Bcl-2 and XIAP (X-linked inhibitor of apoptosis), thus causing chemoresistance (Harte et al., 2014).
The proteasome participates in the degradation of marked proteins that are no longer necessary for the cell or proteins that have suffered some damage or modification by ubiquitination. It also modulates the levels of proapoptotic, antiapoptotic, growth regulatory and stress response factors. Alterations in this proteolytic system are associated with various pathologies, including cancer. Inhibition of proteasome activity results in the upregulation of proapoptotic factors, such as p53, Bax, and Noxa, while reducing the levels of antiapoptotic proteins, such as Bcl-2 and IAP family proteins (McConkey and Zhu, 2008; Vinod et al., 2013).
Proteasome inhibitors have been shown to promote apoptosis in various types of cancer and induce sensitivity to combination chemotherapeutic agents. For example, bortezomib, an inhibitor of the proteasomal pathway, plays an important role in combination chemotherapy with lenalidomide and thalidomide in MM by stimulating the immune system, inhibiting angiogenesis, and sensitizing cancer cells, thereby overcoming chemoresistance (Orlowski, 2004; Vinod et al., 2013; Piechotta et al., 2019).
Hypoxia is a common feature of all solid neoplasms (Rankin and Giaccia, 2016; Chouaib et al., 2017; Najafi et al., 2020); cellular responses to hypoxia are usually regulated by the family of factors hypoxia-inducible factor (HIF) transcriptions (Harris, 2002; Keith and Simon, 2007). HIF is a protein complex formed by a heterodimer consisting of an HIFα subunit and a HIFβ subunit (Li et al., 2009). Under normoxic conditions, the Von Hippel-Lindau tumor suppressor gene product ubiquitinates HIFα and degrades it at the proteasomal pathway, but in hypoxia, the interaction between HIFα and VHL is abolished. As a result, HIFα is stabilized, dimerizes with HIFβ, and then binds to hypoxia-responsive elements in the promoters of hypoxia-regulated genes (Li et al., 2009). The HIF dimer activates gene transcription that modulates cell survival, proliferation, metabolism, and angiogenesis (Harris, 2002; Li et al., 2009; Schwab et al., 2012). Hypoxia is related to a poor prognosis (van den Beucken et al., 2014; Carnero and Lleonart, 2016; Qin et al., 2017); Hypoxia during tumorigenesis can develop by two mechanisms: chronic or acute hypoxia. Chronic hypoxia occurs due to the high proliferation of cancer cells; therefore, they are constantly expelled from the blood vessels. In contrast, acute hypoxia is caused by a temporary cessation of blood flow due to poor tumor vasculature (Brown and Wilson, 2004). Regardless of the mechanism, tumor hypoxia has been widely documented as contributing to resistance to all anticancer therapies, including chemotherapy and radiotherapy (Brown and Wilson, 2004; Mitani et al., 2014; Jeong et al., 2019; Najafi et al., 2020).
Indeed, silencing or inhibition of HIF-1 increases radiation sensitivity in various tumor models. For example, HIF-1 knockdown in human hepatoma cells inhibits proliferation, induces apoptosis, and promotes radiosensitivity in chemically induced hypoxia (Yang et al., 2011; Wang et al., 2019a). In prostate cancer cell lines, the knockdown of HIF-1 by siRNAs induces apoptosis and cell cycle arrest at the G2/M transition, resulting in radiosensitization (Huang et al., 2012, 201). In xenograft tumors with FaDu (Hypopharyngeal carcinoma) and ME180 (Squamous cell carcinoma) cell lines, blockade of the HIF1 response during transient hypoxic stress increases hypoxia, reduces lactate levels, and improves response to high doses of single fraction radiation (Leung et al., 2017; Wang et al., 2019a). In laryngeal carcinoma, the inhibition of HIF-1α and glucose transporter-1 (GLUT1) expression increases radiosensitivity and promotes apoptosis and necrosis (Shen et al., 2017; Wang et al., 2019a).
Many HIF-1-inducible genes, such as VEGF, Glut-1, MDR, IAP3, and Bcl-2, directly or indirectly mediate chemoresistance (Liu et al., 2008; Doktorova et al., 2015). In various types of tumors, such as hepatocellular carcinoma, neuroblastoma, and lung cancer, HIF-1α inhibition re-sensitizes cells to drug treatment; therefore, it is considered a valid target to reduce drug resistance induced by reverse hypoxia (Liu et al., 2008; Sullivan et al., 2008; Huang et al., 2010; Hartwich et al., 2013; Doktorova et al., 2015).
Radiation therapy (XRT) is a cancer treatment that uses high doses of radiation to kill cancer cells. It has been widely used in breast cancer (Moon et al., 2009), prostate cancer (Zietman et al., 2010), carcinoma lung (Machida et al., 2003), medulloblastoma or glioblastoma (Habrand and De Crevoisier, 2001), melanoma (Ivanov et al., 2007), etc.
Ionizing radiation (IR) promotes its effects by inducing DNA damage and activating DNA damage-induced signaling pathways (Kma, 2013). These pathways result in cell cycle arrest or induction of cell death by apoptosis, necrosis, autophagy, or mitotic catastrophe, depending on the total dose (Wang et al., 2006; Suit et al., 2007; Kma, 2013). However, the efficacy of XRT is limited by the radioresistance exhibited by cancer cells (Leone et al., 2008; Ruan et al., 2009; Yang et al., 2012). For instance, prostate cancer is highly resistant to IR (Crook et al., 1995; Hagan et al., 2000). The doses of XRT that are usually used in the treatment of prostate adenocarcinoma are up to 70 Gy and have shown biochemical failure rates of 30% or more, which leads to the need to increase the dose of XRT, which, in turn, results in impotence, rectal and bladder toxicity (Zietman et al., 2010). Likewise, it has been reported that the increase in radiation dose leads to the incidence of skin toxicity in patients undergoing XRT (Weiss and Landauer, 2003). These effects add to some of the side effects of XRT, such as pituitary hormone dysfunction, behavioral problems, and reduced neurogenesis (Baskar et al., 2012; Michaelidesová et al., 2019), which consequently shows diminished therapeutic outcome and poor quality of life for survivors.
Therefore, researchers have focused on finding drugs or compounds that function as effective radiosensitizers, reducing the radiation dose-response threshold for cancer cells with minimal side effects in normal cells (Kma, 2013).
In this context, it has been reported that RSV has radioprotective effects; thanks to its antioxidant properties, this compound acts as a scavenger of free radicals or ROS. In addition, it has been reported that it reduces inflammation by inhibiting IL-8 expression and blocking NF-κB activation (Benitez et al., 2009; Oh et al., 2009). The radioprotective effects of RSV in vivo were possibly first demonstrated in the study by Carsten et al. (Carsten et al., 2008), where it was shown that RSV in combination with IR resulted in a reduction in the frequency of total chromosomal aberrations in mouse bone marrow cells, compared to untreated, RSV-only treated groups hear. In this case, mice were administered RSV at a dose of 100 mg/kg body weight per day, started 2 days before whole body irradiation with 3 Gy (at a dose rate of 1.18 Gy)/min., and analyzed 1 and 30 days after irradiation. It was evident from this observation that RSV possesses a potential radioprotective property (Carsten et al., 2008; Kma, 2013).
On the other hand, several studies have shown that using RSV in combination with radiotherapy increases therapeutic efficacy against cancer (Fang et al., 2012; 2013; Komorowska et al., 2022). The following table exemplifies some studies that have seen the radiosensitizing effect of RSV and the mechanisms involved in it (Table 2).
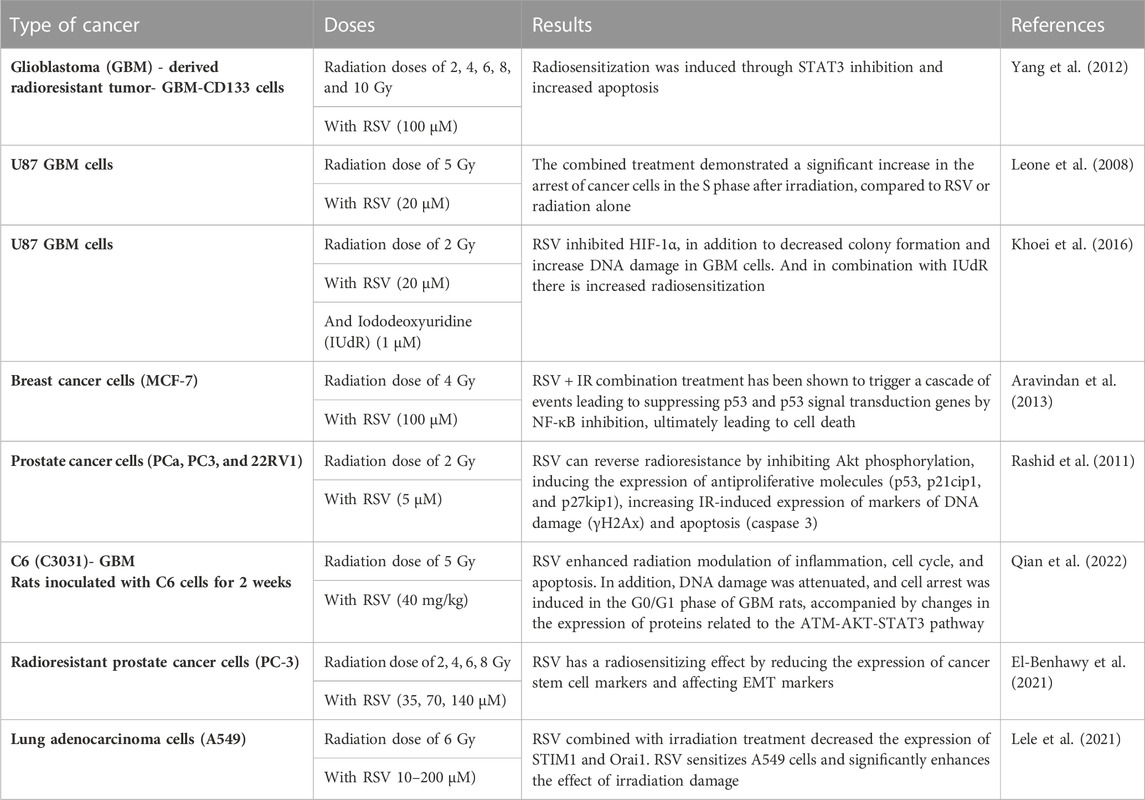
TABLE 2. Radiosensitizing effect of RSV in cancer cells.
In addition to the mechanisms mentioned above, it has been reported that RSV can induce senescence, apoptosis, autophagy, and inhibition of DNA repair, as well as the ability to kill cancer stem cells more efficiently, leading to radiosensitization of cancer cells (Figure 3) (Luo et al., 2013; Wang et al., 2015).
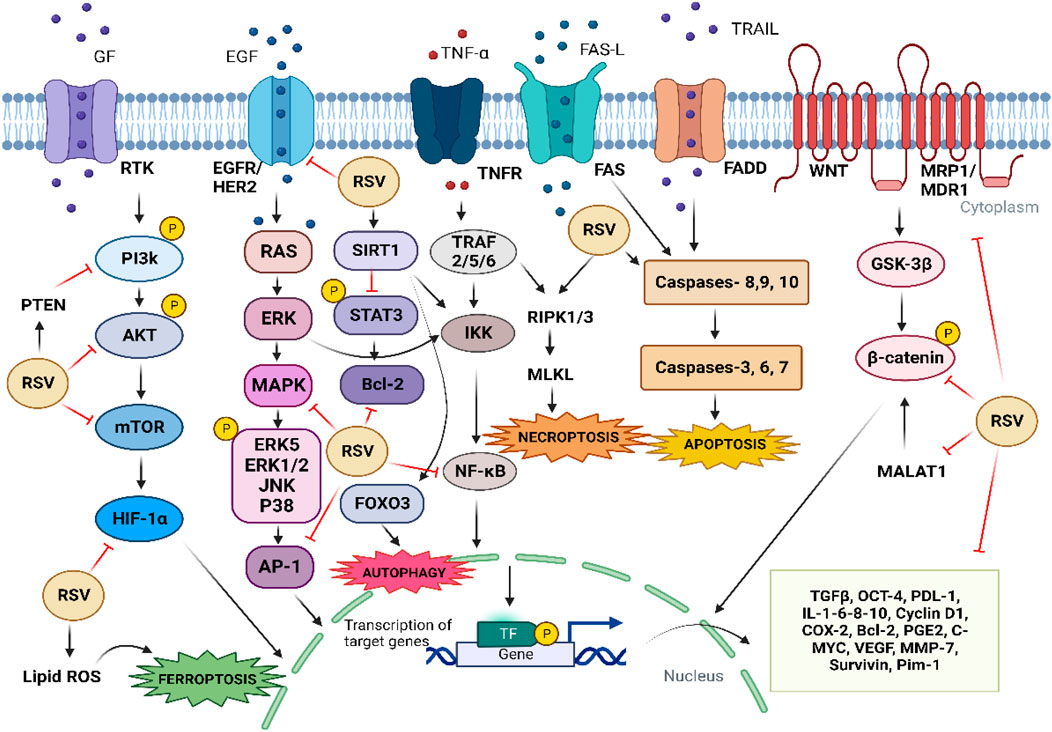
FIGURE 3. Mechanisms by which RSV contributes to chemosensitization and radiosensitization of cancer cells. The arrows refer to activation, while the hammerhead lines refer to inhibition.
Chemosensitization is based on using a drug or compound that enhances the activity of another by influencing one or more resistance mechanisms, making it a valuable strategy to overcome the chemoresistance developed by cancer cells. In addition, it dramatically reduces the adverse effects that occur due to the toxicity of high doses of drugs used in cancer treatment.
In vitro and in vivo studies show that RSV can reverse chemoresistance in tumor cells by modulating apoptosis and downregulation of drug transporters and proteins involved in cancer progression (Figure 3) (Lee et al., 2016).
Paclitaxel is one of the drugs used in chemotherapy to treat different types of cancer, including ovarian, breast, and lung lymphoma, among others. Some side effects after treatment with paclitaxel are anemia, bruising, bleeding, nausea, diarrhea, tingling in the hands and feet, tiredness, hair loss, muscle pain, etc. (Weaver, 2014).
The primary mechanism behind RSV chemosensitization to paclitaxel chemotherapy is the downregulation of Bcl-2 and MDR1/P-gp family members. It has been suggested that RSV-mediated inhibition of the ERK1/2 and AP-1 pathways leads to decreased Bcl-xL in non-Hodgkin’s lymphoma and MM cell lines (Cusack et al., 2000; Jazirehi and Bonavida, 2004).
In another study, when RSV was administered prior to paclitaxel treatment in lung cancer cell lines (A549, EBC-1, Lu65), a significant improvement in the antiproliferative potential of paclitaxel was observed. Furthermore, RSV also caused cell cycle arrest in the G1 and G1/S phases of the cell cycle by inducing the expression of the CDK inhibitors p21/WAF1/CIP1 and p27/KIP1, allowing the dose of paclitaxel required to kill tumor cells to be reduced (Shankar et al., 2007).
Downregulation of survivin is another mechanism by which RSV enhances proliferation-inhibitory effects through S-phase cell cycle arrest and increased apoptosis in neuroblastoma cells treated with paclitaxel, in addition to other drugs such as doxorubicin, cytarabine, taxol, actinomycin, and methotrexate (Fulda and Debatin, 2004). RSV has also been reported to decrease survivin expression in a dose-dependent manner in a multidrug-resistant human non-small cell lung cancer cell line (SPC-A-1/CDDP). Similarly, in a more recent study, RSV was found to increase the sensitivity of renal cells (Caki-1) resistant to paclitaxel by suppressing survivin expression (Min et al., 2019).
Another study demonstrated the synergistic interaction of RSV and paclitaxel in inducing apoptosis in the DBTRG glioblastoma cell line. In this work, they observed that the combination with RSV increases apoptosis markers such as mitochondrial membrane depolarization, ROS levels, and caspase 3 activity in DBTRG cells, compared to treatment with paclitaxel alone. The synergistic effect seems to be mediated by the stimulation and activation of the TRPM2 channel sensitive to mitochondrial oxidative stress, which allows Ca2+ to enter cancer cells, contributing to their death (Öztürk et al., 2019; Cocetta et al., 2021).
Doxorubicin (adriamycin or hydroxydaunorubicin) is a drug (anthracycline-type antibiotic) with antitumor activity produced by Streptococcus peucetius var. caesius; used to treat different types of cancer, such as leukemia, lymphoma, neuroblastoma, sarcoma, Wilms tumor, lung, breast, stomach, ovary, thyroid, and bladder cancer. It can intercalate with DNA, inhibiting DNA and RNA synthesis. It induces a cell cycle arrest during the S phase (Rivankar, 2014). In several studies it has been seen that RSV can sensitize DOX-resistant cancer cells, such as those shown below.
Fenig et al.; found that RSV treatment decreases MRP1 expression in AML cells resistant to doxorubicin. Furthermore, when RSV was administered, the expression of MRP1 decreased, while the cellular uptake of DOX in resistant cells increased. Based on these observations, the authors concluded that RSV might facilitate cellular DOX uptake through the downregulation of MRP1 and that RSV may help to overcome DOX resistance or sensitize AML cells resistant to doxorubicin (Fenig et al., 2004; Gupta et al., 2011). Likewise, RSV chemosensitization to DOX is mediated by inhibition of MDR1/P-gp and Bcl-2 in ovarian cancer cells (OVCAR-3) (Rezk et al., 2006), acute myeloid leukemia (AML-2) (Kweon et al., 2010) and oral squamous cell carcinoma (KBv200) (Quan et al., 2008).
Also, RSV has been shown to increase the chemosensitivity of tumor cells by arresting the cells at different stages of the cell cycle. For example, in DOX-chemoresistant B16 melanoma cells, RSV increased DOX-induced cytotoxicity and decreased cyclin D1 expression. In addition, DOX treatment combined with RSV was associated with an increased cell cycle arrest in the G1/S phase (Gatouillat et al., 2010).
On the other hand, in breast cancer cells (MCF-7/adr) and MDA-MB-231 resistant to DOX, it has been reported that the combination of DOX with RSV inhibits cell growth, promotes apoptosis, and suppresses cell migration (Cocetta et al., 2021). The effect of RSV is linked to the modulation between SIRT1 and β-catenin; RSV was shown to be able to increase SIRT1 (deacetylase) levels and decrease β-catenin expression by ubiquitination, which reversed chemoresistance (Jin et al., 2019). Further experiments showed that RSV treatment significantly increased cellular accumulation of DOX by decreasing the expression levels of the ATP-binding cassette (ABC) transporter genes, MDR1 and MRP1 in MCF-7/adr and MDA-MB-231 cells., as well as in a xenograft model (in vivo) revealing that RSV and DOX treatment in combination significantly inhibits tumor volume (Breast cancer) by 60%, compared to the control group (Kim et al., 2014).
Furthermore, in the study by (Mitani et al., 2014), they investigated the effect of RSV on hypoxia-induced doxorubicin resistance in MCF-7 cells. They were observing how RSV and its derivative 3,5-dihydroxy-4′-methoxy-trans-stilbene, reverse hypoxia-induced doxorubicin resistance at a concentration of 10 μM through the decrease in HIF protein expression -1α and HIF-1 activity activated by hypoxia. Similarly, RSV inhibited the expression of CBR1 induced by hypoxia at the mRNA and protein levels (Mitani et al., 2014).
The combination of DOX and RSV also increases Bax gene expression in HCT116 colon cancer cell lines; furthermore, RSV enhances intracellular DOX uptake by blocking P-gp activity, thereby sensitizing colorectal cancer cells to DOX (Khaleel et al., 2016).
Also, it has been reported that the acquisition of DOX resistance in SGC7901 gastric cancer cells may be due to EMT induced by aberrant activation of Akt, giving cancer cells the ability to overexpress genes related to DOX chemoresistance. This cellular model showed that RSV reverses DOX resistance by suppressing EMT by inhibiting the PI3K/Akt signaling pathway, activating caspase-3-dependent apoptosis. In addition, RSV induced cell cycle arrest by increasing PTEN expression in addition to suppressing cell invasion and N-cadherin expression (Xu et al., 2017).
On the other hand, RSV-inducing chemosensitivity in breast cancer cells (MCF-7) resistant to adriamycin or DOX has also been shown to be dependent on miR-122-5p inhibition. Moreover, inhibition of miR-122-5p showed a significant effect on the regulation of critical anti-apoptotic proteins such as Bcl-2 and cyclin-dependent kinases (CDK2, CDK4, and CDK6) in breast cancer cells (MCF-7) in response to RSV (Zhang et al., 2019a).
In a recent study, Moreira et al. demonstrated that RSV increases the expression of the SIRT1 gene in LoVo cells (derived from metastatic colon adenocarcinoma nodules). SIRT1 negatively regulates the expression of survivin, a major inhibitor of apoptosis and which, as seen earlier in this review, is involved in the resistance of cancer cells to chemotherapeutic therapies. They demonstrated that increased expression of the SIRT1 gene contributed to overcoming resistance to apoptosis in DOX-resistant LoVo colon cancer cells (Moreira et al., 2022). In the study by Xiong Le et al., they prepared nanoparticles with sustained release capacity and targeted IL-13Rα2 to improve its bioavailability from the RSV. These nanoparticles were inserted into an ATC/anaplastic thyroid cancer mouse model, demonstrating that RSV effectively inhibits ATC growth in vivo. And that it can overcome the resistance to DOX and Docetaxel in this model (Xiong et al., 2021).
Temozolomide is a chemotherapy drug that mostly treats brain tumors (GBM, medulloblastomas, neuroblastomas, and sarcomas). Temozolomide comes in capsules and can be used alone or in combination with XRT. It is an alkylating cytostatic agent that, when is activated, forms free radicals capable of causing DNA degradation and even single and double-strand DNA breaks that induce cell cycle arrest in G2/M, which eventually leads to cell apoptosis (Yan et al., 2016). The increase in resistance to TMZ is one of the main reasons for the failure of glioblastoma treatment (Perazzoli et al., 2015; Lee, 2016).
RSV has been shown to enhance the therapeutic efficacy of TMZ in several ways. One proposed mechanism is the reduction of autophagy mediated by an increase in ROS, favoring apoptosis. In the glioblastoma multiforme (GBM) cell line SHG44, TMZ, in combination with RSV, markedly increased the production of ROS, which served as a signal for the activation of AMP-activated protein kinase (AMPK). Subsequently, activated AMPK inhibited mTOR signaling and decreased levels of the anti-apoptotic protein Bcl-2, contributing to the additive antiproliferative effects of combined TMZ and RSV treatment. These results were also confirmed in vivo mouse models (GBM orthotopic xenograft), where the combination of TMZ and RSV treatment induced a reduction in tumor volume and tumor proliferation, which was associated with decreased expression of Ki −67, a proliferation index marker (Yuan et al., 2012). Other studies indicate that GBM-initiating cells (GIC), which display stem cell properties, are involved in tumor resistance to TMZ, and RSV has been shown to enhance GIC sensitivity to TMZ by activating the pATM/pATR/p53 pathway and promoting cancer cell apoptosis. In addition, this work demonstrated that RSV inactivated p-STAT3, promoting the differentiation of glioblastoma-initiating cells (Li et al., 2016). RSV has recently been seen to sensitize glioma cell lines with strong resistance to TMZ through the inhibition of Wnt2 and β-catenin and increased expression of GSK-3β (Yang et al., 2019).
Cisplatin or CDDP (alkylating agent) is a drug widely used in cancer that inhibits DNA synthesis by producing crosslinks within DNA chains known as adducts; its cytotoxic activity is produced by binding to all DNA bases, with a preference for guanine and adenosine bases. It is widely used to treat testicular, ovarian, bladder, head and neck, esophageal, small cell and non-small cell lung, breast, cervical, stomach, prostate, Hodgkin and non-Hodgkin lymphoma, neuroblastomas, sarcomas, MM, melanoma and mesothelioma (Galanski, 2006).
In addition to the high cytotoxicity of cisplatin, the main limitation of the clinical utility of this drug against cancer is the high incidence of chemoresistance (Galluzzi et al., 2012). Below are some studies where RSV sensitizes and improves the effectiveness of treatment with cisplatin.
RSV and cisplatin together show more effective inhibition of non-small lung cancer cell (NSCLC) proliferation and induction of apoptosis than cisplatin treatment alone (Cocetta et al., 2021). In fact, in the study by Ma L et al. (Ma et al., 2015), it was shown that the combination of cisplatin and RSV dramatically improved the efficacy of cisplatin in depolarizing the mitochondrial membrane potential, increasing the release of cytochrome C, as well as decreasing Bcl-2 expression and increased Bax protein in cisplatin-resistant H838 and H520 non-small lung cancer cells, resulting in increased inhibition of proliferation and induction of apoptosis (Ma et al., 2015). Synergistic effects of RSV in combination with cisplatin have also been shown in A549 lung carcinoma cells. The results show that the combination favors autophagy by lessening autophagosome accumulation, AKT phosphorylation, and LC3-II protein levels (Hu et al., 2016a).
The impact of combined RSV and cisplatin treatment has also been tested in hepatocarcinoma cells; Liu and others 2018 showed that RSV-induced chemosensitivity to cisplatin is associated with an imbalance in redox homeostasis that favors DNA damage and apoptosis. The data indicate that RSV can inhibit glutamine metabolism of resistant human hepatocarcinoma cell lines (C3A and SMCC7721), increasing the toxic effect of chemotherapy but not on normal liver cells (Liu et al., 2018; Cocetta et al., 2021).
Another interesting study is that of Weiguo (Zhao et al., 2010), who analyzed the efficacy of RSV with non-small cell lung cancer cells or NSCLC (SPC-A-1/CDDP) resistant to multiple drugs (Paclitaxel, taxol, gefitinib, cisplatin, etc.). Cells were treated with RSV at a concentration of 25, 50, or 100 μM in vitro studies, and tumors were induced in nude mice implanted with SPC-A-1/CDDP cells and fed a special diet containing included RSV at a dose of 1 g/kg/day or 3 g/kg/day. In this work, they evaluated cell proliferation, apoptosis, the distribution of cell cycle phases, the IC50 values of cisplatin, gefitinib, and paclitaxel, the volume of the implanted tumor, and the expression of survivin in mice treated with RSV compared to the control. RSV significantly inhibited the proliferation of NSCLC cells, induced apoptosis, arrested the cell cycle between G0-G1 and S phases or in the G2/M phase, and decreased IC50 values of chemotherapeutic drugs (cisplatin, gefitinib, and paclitaxel) (Gupta et al., 2011). Furthermore, RSV showed antitumor effects in mice, affecting tumor proliferation in a dose- and time-dependent manner; similarly, survivin expression in SPC-A-1/CDDP cells decreased after RSV treatment (Zhao et al., 2010).
Oxaliplatin or OXA (bifunctional alkylating agent) is a type of platinum chemotherapy that binds covalently to DNA and forms platinum-DNA adducts to inhibit DNA replication and transcription (Kelland, 2007). The intrastrand crosslinks formed by OXA can block DNA replication and transcription (Woynarowski et al., 2000). OXA is used to treat colorectal cancer and, in some cases, throat cancer (esophagus) (Kelland, 2007). However, the development of resistance to OXA in vitro and in vivo in colon cancer has been reported (Hsu et al., 2018).
Kaminski et al. (Kaminski et al., 2014) investigated the effect of RSV on the antitumor activity of oxaliplatin in the resistant colon cancer cell line Caco-2 and its possible involvement in the inflammatory response. The results showed that the combined treatment synergistically inhibits cell growth and induces apoptosis with caspase-3 activation, PARP cleavage, and mitochondrial membrane depolarization (Cocetta et al., 2021). In addition, primary macrophages derived from human monocytes were seeded and differentiated to add later supernatants of tumor cells (Caco-2) treated with RSV/OXA and the co-cultures were maintained for 24 h, noting how the co-treatment prevents the immunosuppression of the co-cultured macrophages, making them potentially tumoricidal (Kaminski et al., 2014).
In addition, RSV prevents OXA-induced neuronal damage and peripheral neuropathic pain (CIPNP), which is a common and devastating side effect of cancer therapy. Avoiding the upregulation of NFκB, TNFα, ATF3, and c-fos, increasing the expression of Nrf2, NQO-1, HO-1, and the redox-sensitive deacetylase SIRT1 (Recalde et al., 2020).
5-FU is a drug that inhibits the activity of thymidylate synthetase, the enzyme responsible for producing Thymidine, acting as an antimetabolite, inducing cell cycle arrest, and promoting apoptosis (Longley et al., 2003). 5-FU has shown the most significant impact in treating colorectal cancer (CRC), although it has also been used in treating breast, stomach, pancreas, and certain types of skin cancer (Gu et al., 2019). Despite its great advantages, the clinical application of 5-FU is limited due to the development of resistance of cancer cells. In fact, there is increasing evidence showing that cancer stem cells (CSC) present in the tumor microenvironment (TME) are the main ones responsible for resistance to 5-FU (Yang et al., 2015; Das et al., 2020).
Several works have shown the efficacy of RSV in potentiating the cytotoxic effect of 5-FU (Moutabian et al., 2022). A study conducted in colorectal cancer cells (HCT116 and DLD1) showed that the combined treatment increases cell cycle arrest and decreases the proliferation and migration of colorectal cancer cells by inhibiting the PI3K/Akt signaling pathway. Likewise, RSV showed anti-inflammatory effects by inhibiting pSTAT3 and NFκB proteins (Chung et al., 2018). The combination of RSV and 5-FU also inhibits the proliferation and migration of B16 murine melanoma cells by decreasing the levels of AMPK, COX-2, VASP, and VEGF, compared to the compounds alone (RSV and 5-FU) (Lee et al., 2015). In addition, Buhrmann and others 2018 demonstrated that RSV could reduce TNF-β-induced survival and migration of resistant HCT116 colorectal cancer cells by promoting 5-FU sensitization (Buhrmann et al., 2018).
Recently, Brockmueller et al. demonstrated in tumor microenvironments of 5-FU-resistant HCT-116 and HCT-116R colorectal cancer (CRC) cells with 3D alginate and monolayer cultures how RSV increased the sensitivity of CRC cells to 5-FU by reducing vitality, migration, proliferation, angiogenesis, invasion, and epithelial-mesenchymal transition (Brockmueller et al., 2023).
Gemcitabine (2′, 2′-difluoro 2′-deoxycytidine) is a drug that induces cancer cell death by inhibiting ribonucleotide reductase, an enzyme necessary for the synthesis of deoxyribonucleotides, in addition to inhibiting DNA polymerase. Gemcitabine inhibits DNA synthesis; the cells cannot divide properly and die. Gemcitabine is used to treat different types of cancer, including carcinoma of the bladder, pancreas, oral squamous cell carcinoma, non-small cell lung, ovary, and breast (Mini et al., 2006).
Similarly, it has been shown that cytochrome p450 1b1 (Cyp1b1) is overexpressed in many neoplasms and plays an important role in developing resistance to chemotherapy. Interestingly, RSV has been reported to downregulate Cyp1b1, thereby increasing apoptosis induced by antimetabolites such as 5-FU and gemcitabine in gemcitabine-chemoresistant cholangiocarcinoma (Mz-ChA-1, HuCC-T1, CCLP1, and SG231) tumor models. Mitomycin C and 5-FU, although the precise mechanism of RSV-mediated chemosensitization associated with Cyp1b1 inhibition is unknown (Frampton et al., 2010). However, it has been shown in an orthotopic mouse model of human pancreatic cancer that RSV enhanced the antitumor activity of gemcitabine, and this was associated with decreased expression of Bcl-2, Bcl-xL, COX-2, cyclin D1, MMP-9 and VEGF (Harikumar et al., 2010).
Docetaxel is an antineoplastic drug that stabilizes microtubules, inhibiting their polymerization, which causes cell death by interruption of mitosis. Docetaxel is used to treat different types of cancer, including breast cancer, prostate, non-small cell lung cancer (NSCLC), stomach, ovary, bladder, soft tissue sarcoma, melanoma, and head and neck cancer (Rodríguez Carranza, 2015). Docetaxel resistance in breast cancer cells (SK-BR-3, MCF7, MDA-MB-231, and T47D) is associated with HER2 expression.
HER-2 is a receptor of the epidermal growth factor family involved in cell growth and development. In addition, it promotes the recruitment of several proteins, which lead to the activation of signal transduction cascades such as: the PI3K/AKT/mTOR and RAF/MEK/ERK pathways. Overexpression of this receptor contributes to the progression and survival of breast cancer (Fink and Chipuk, 2013; Li and Li, 2013; Cocetta et al., 2021). Interestingly, RSV treatment inhibits HER-2 activation through docetaxel-induced blockade of MAPK and Akt signaling, as well as survival signaling pathways activated by HER-2, enhancing the sensitization of breast cancer cells (SK-BR-3 and MDA-MB-231) to docetaxel (Vinod et al., 2015).
In a recent study, EGF-conjugated hybrid lipid polymer nanoparticles (LPN) were fabricated to co-deliver docetaxel (DTX) and RSV in non-small cell lung cancer (NSCLC). In vitro and in vivo studies demonstrated that EGF DTX/RSV LPNs have significant synergistic effects, the best tumor inhibition capacity, and the lowest systemic toxicity. These results suggest that EGF DTX/RSV LPNs may be a promising strategy for treating and chemosensitization NSCLC (Song et al., 2018). Similarly, Zhang et al. designed PEGylated nanoliposomes to co-deliver Docetaxel and RSV in Balb/c nude mice bearing prostate cancer (PC3), demonstrating the efficiency of the treatment as a whole (Zhang et al., 2022a). Other recent studies of RSV as a chemo-sensitizer are summarized in Table 3.
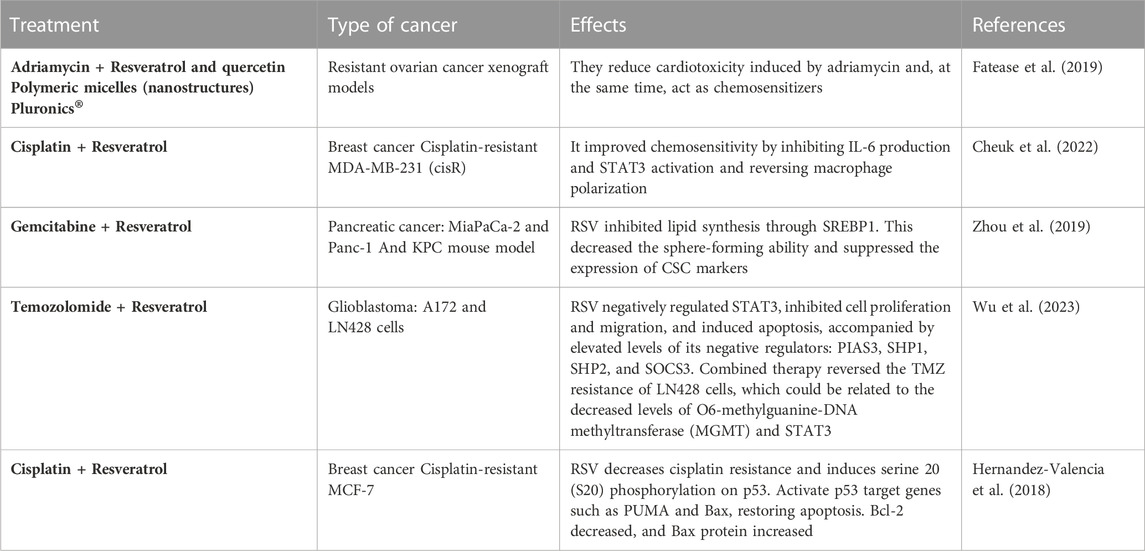
TABLE 3. Chemo-sensitizing effects of resveratrol.
In the following figure, you can visualize in a more general way the events that RSV regulates to carry out the chemo and radiosensitization of cancer cells (Figure 4).
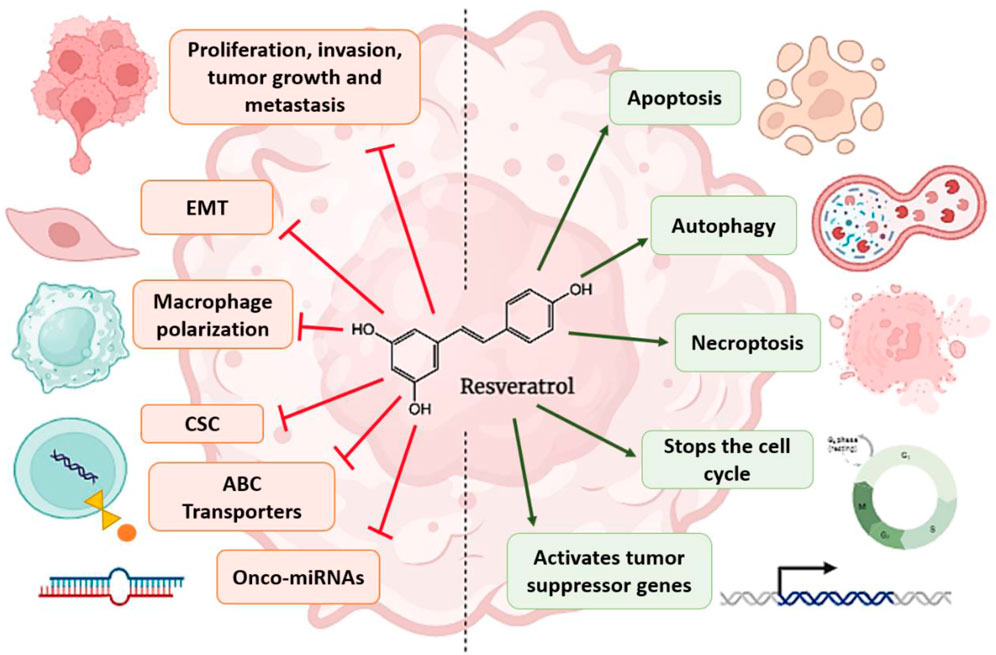
FIGURE 4. Factors regulated by RSV that lead to cancer chemo and radiosensitization. The arrows refer to promotion or increase, while the hammerhead lines refer to inhibition.
The acquisition of chemoresistance and radioresistance remains one of the main problems in the treatment of cancer patients. The fact that tumor cells develop multiple resistance mechanisms and that more than one mechanism can operate simultaneously complicates the success of anticancer treatments such as chemotherapy and radiotherapy.
In this review, we talk about the molecular mechanisms that lead to chemo- and radioresistance by cancer cells, and in addition, we try to concisely describe how RSV, in conjunction with treatments (radiotherapy and chemotherapy), manages to sensitize cancer cells, making them less resistant and favoring the effectiveness of the treatments. In addition, this article also summarizes the general effects of RSV treatment on cancer in an easy way to understand and shows the most recent studies that have addressed the issue of chemo- and radiosensitization of cancer cells by RSV. The RSV has the ability to modify and affect many molecular mechanisms that cause sensitization of cancer cells. Some of the most important mechanisms involved in sensitization we find transmembrane transport (decrease in drug transport proteins), regulation of the cell cycle (arrest of the cycle in the G1/S phases), decreased cell proliferation, activation of different types of cell death (apoptosis, necrosis, autophagy), inhibition of transcriptional factors such as NF-kB, blockage of DNA repair, reduction of inflammation due to the inhibition of COX-2, reduces the formation of CSCs by inhibiting oncogenic genes and onco-miRNAs, inhibition of epithelial-mesenchymal transition, activation of tumor suppressor genes, among others.
Based on everything analyzed and studied in this review, we conclude that RSV is undoubtedly an excellent candidate to be used as a complementary treatment to chemotherapy and radiotherapy since all the mechanisms and cellular targets that RSV regulates clearly favor the sensitization of cancer cells to these medications. This would improve the success rate of the treatments and undoubtedly improve the patients’ quality of life by reducing the treatment period.
By looking at the signaling pathways and mechanisms that are affected or mediated by RSV in a tumor environment, we believe that RSV may be an ideal candidate as a complementary treatment to chemotherapy and radiotherapy. Although, there is a problem with the clinical use of RSV, and we are talking about its bioavailability. Many researchers have taken on the task of ending this barrier to use RSV more appropriately and efficiently. Today, many research works, such as those addressed in Sharifi-Rad’s article (Sharifi-Rad et al., 2021), show how targeted molecular therapy, specifically nanotechnology, shows to be a useful tool in improving the bioavailability of RSV.
Studies have even been carried out where co-encapsulation of RSV with other drugs, such as those used in chemotherapy, is proposed as in the works (Song et al., 2018; Zhang et al., 2022a) where they encapsulated RSV and DOX in nanoparticles of lactic-co-glycolic acid (PLGA) and hybrid lipid polymer (LPN) nanoparticles. This coencapsulation not only improved the half-life of DOX and RSV but also increased the concentrations of both molecules within the tumor, reducing the toxicity of DOX in healthy tissue and increasing the efficacy of DOX in overcoming the resistance of cancer cells (Cocetta et al., 2021). Therefore, these results suggest that nanoformulations protect and improve RSV’s stability and bioavailability, ensuring greater treatment efficiency. In addition, in conjunction with other drugs and therapies, it could even evade the resistance of cancer cells. However, further studies in vivo are needed to optimize nanocarrier and lipocarrier delivery systems.
Interestingly, new RSV analogues have also been reported to have increased bioavailability and have also been shown to possess anti-cancer properties (Ferraz da Costa et al., 2020). However, more studies are needed to understand its action and mechanisms of chemosensitization and radiosensitization.
To take part in the Resource Identification Initiative, please use the corresponding catalog number and RRID in your current manuscript. For more information about the project and for steps on how to search for an RRID, please click here.
Life Science Identifiers (LSIDs) for ZOOBANK registered names or nomenclatural acts should be listed in the manuscript before the keywords with the following format:
urn:lsid:<Authority>:<Namespace>:<ObjectID>[:<Version>].
SC-N: Conceptualization, Investigation, Writing–original draft, Writing–review and editing. LH: Conceptualization, Writing–review and editing, Supervision. GD-G: Conceptualization, Supervision, Writing–original draft, Writing–review and editing. JD-C: Conceptualization, Funding acquisition, Writing–original draft, Writing–review and editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. Details of all funding sources should be provided, including grant numbers if applicable. Please ensure to add all necessary funding information, as after publication this is no longer possible.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Adachi, M., Cui, C., Dodge, C. T., Bhayani, M. K., and Lai, S. Y. (2012). Targeting STAT3 inhibits growth and enhances radiosensitivity in head and neck squamous cell carcinoma. Oral Oncol. 48, 1220–1226. doi:10.1016/j.oraloncology.2012.06.006
Akimoto, T., Hunter, N. R., Buchmiller, L., Mason, K., Ang, K. K., and Milas, L. (1999). Inverse relationship between epidermal growth factor receptor expression and radiocurability of murine carcinomas. Clin. Cancer Res. 5, 2884–2890.
Akira, S. (2000). Roles of STAT3 defined by tissue-specific gene targeting. Oncogene 19, 2607–2611. doi:10.1038/sj.onc.1203478
Ali, K., Maltese, F., Choi, Y. H., and Verpoorte, R. (2010). Metabolic constituents of grapevine and grape-derived products. Phytochem. Rev. 9, 357–378. doi:10.1007/s11101-009-9158-0
Ali, M. Y., Oliva, C. R., Noman, A. S. M., Allen, B. G., Goswami, P. C., Zakharia, Y., et al. (2020). Radioresistance in glioblastoma and the development of radiosensitizers. Cancers 12, 2511. doi:10.3390/cancers12092511
Altieri, D. C. (2003). Validating survivin as a cancer therapeutic target. Nat. Rev. Cancer 3, 46–54. doi:10.1038/nrc968
Aravindan, S., Natarajan, M., Herman, T. S., Awasthi, V., and Aravindan, N. (2013). Molecular basis of ‘hypoxic’ breast cancer cell radio-sensitization: phytochemicals converge on radiation induced Rel signaling. Radiat. Oncol. 8, 46. doi:10.1186/1748-717X-8-46
Asemi, R., Rajabpoor Nikoo, N., Asemi, Z., Shafabakhsh, R., Hajijafari, M., Sharifi, M., et al. (2023). Modulation of long non-coding RNAs by resveratrol as a potential therapeutic approach in cancer: a comprehensive review. Pathology - Res. Pract. 246, 154507. doi:10.1016/j.prp.2023.154507
Ashraf, H. M., Moser, J., and Spencer, S. L. (2019). Senescence evasion in chemotherapy: a sweet spot for p21. Cell 178, 267–269. doi:10.1016/j.cell.2019.06.025
Ashrafizadeh, M., Taeb, S., Haghi-Aminjan, H., Afrashi, S., Moloudi, K., Musa, A. E., et al. (2021). Resveratrol as an enhancer of apoptosis in cancer: a mechanistic review. ACAMC 21, 2327–2336. doi:10.2174/1871520620666201020160348
Atari-Hajipirloo, S., Nikanfar, S., Heydari, A., Noori, F., and Kheradmand, F. (2016). The effect of celecoxib and its combination with imatinib on human HT-29 colorectal cancer cells: involvement of COX-2, Caspase-3, VEGF and NF-κB genes expression. Cell Mol. Biol. (Noisy-le-grand) 62, 68–74.
Baek, Y., Jeong, E. W., and Lee, H. G. (2023). Encapsulation of resveratrol within size-controlled nanoliposomes: impact on solubility, stability, cellular permeability, and oral bioavailability. Colloids Surfaces B Biointerfaces 224, 113205. doi:10.1016/j.colsurfb.2023.113205
Bamodu, O. A., Chang, H.-L., Ong, J.-R., Lee, W.-H., Yeh, C.-T., and Tsai, J.-T. (2020). Elevated PDK1 expression drives PI3K/AKT/MTOR signaling promotes radiation-resistant and dedifferentiated phenotype of hepatocellular carcinoma. Cells 9, 746. doi:10.3390/cells9030746
Bao, R., Connolly, D. C., Murphy, M., Green, J., Weinstein, J. K., Pisarcik, D. A., et al. (2002). Activation of cancer-specific gene expression by the survivin promoter. J. Natl. Cancer Inst. 94, 522–528. doi:10.1093/jnci/94.7.522
Baskar, R., Lee, K. A., Yeo, R., and Yeoh, K.-W. (2012). Cancer and radiation therapy: current advances and future directions. Int. J. Med. Sci. 9, 193–199. doi:10.7150/ijms.3635
Basu, S., Dong, Y., Kumar, R., Jeter, C., and Tang, D. G. (2022). Slow-cycling (dormant) cancer cells in therapy resistance, cancer relapse and metastasis. Seminars Cancer Biol. 78, 90–103. doi:10.1016/j.semcancer.2021.04.021
Behroozaghdam, M., Dehghani, M., Zabolian, A., Kamali, D., Javanshir, S., Hasani Sadi, F., et al. (2022). Resveratrol in breast cancer treatment: from cellular effects to molecular mechanisms of action. Cell. Mol. Life Sci. 79, 539. doi:10.1007/s00018-022-04551-4
Belka, C., and Budach, W. (2002). Anti-apoptotic Bcl-2 proteins: structure, function and relevance for radiation biology. Int. J. Radiat. Biol. 78, 643–658. doi:10.1080/09553000210137680
Benitez, D. A., Hermoso, M. A., Pozo-Guisado, E., Fernández-Salguero, P. M., and Castellón, E. A. (2009). Regulation of cell survival by resveratrol involves inhibition of NFκB-regulated gene expression in prostate cancer cells. Prostate 69, 1045–1054. doi:10.1002/pros.20953
Benlloch, M., Ortega, A., Ferrer, P., Segarra, R., Obrador, E., Asensi, M., et al. (2005). Acceleration of glutathione efflux and inhibition of gamma-glutamyltranspeptidase sensitize metastatic B16 melanoma cells to endothelium-induced cytotoxicity. J. Biol. Chem. 280, 6950–6959. doi:10.1074/jbc.M408531200
Berretta, M., Quagliariello, V., Maurea, N., Di Francia, R., Sharifi, S., Facchini, G., et al. (2020). Multiple effects of ascorbic acid against chronic diseases: updated evidence from preclinical and clinical studies. Antioxidants 9, 1182. doi:10.3390/antiox9121182
Bharti, A. C., and Aggarwal, B. B. (2002). Nuclear factor-kappa B and cancer: its role in prevention and therapy. Biochem. Pharmacol. 64, 883–888. doi:10.1016/s0006-2952(02)01154-1
Bharti, A. C., Shishodia, S., Reuben, J. M., Weber, D., Alexanian, R., Raj-Vadhan, S., et al. (2004). Nuclear factor-kappaB and STAT3 are constitutively active in CD138+ cells derived from multiple myeloma patients, and suppression of these transcription factors leads to apoptosis. Blood 103, 3175–3184. doi:10.1182/blood-2003-06-2151
Bianco, C., Bianco, R., Tortora, G., Damiano, V., Guerrieri, P., Montemaggi, P., et al. (2000). Antitumor activity of combined treatment of human cancer cells with ionizing radiation and anti-epidermal growth factor receptor monoclonal antibody C225 plus type I protein kinase A antisense oligonucleotide. Clin. Cancer Res. 6, 4343–4350.
Biesalski, H. K. (2007). Polyphenols and inflammation: basic interactions. Curr. Opin. Clin. Nutr. Metabolic Care 10, 724–728. doi:10.1097/MCO.0b013e3282f0cef2
Blaser, H., Dostert, C., Mak, T. W., and Brenner, D. (2016). TNF and ROS crosstalk in inflammation. Trends Cell Biol. 26, 249–261. doi:10.1016/j.tcb.2015.12.002
Blondy, S., David, V., Verdier, M., Mathonnet, M., Perraud, A., and Christou, N. (2020). 5-Fluorouracil resistance mechanisms in colorectal cancer: from classical pathways to promising processes. Cancer Sci. 111, 3142–3154. doi:10.1111/cas.14532
Brockmueller, A., Girisa, S., Kunnumakkara, A. B., and Shakibaei, M. (2023). Resveratrol modulates chemosensitisation to 5-FU via β1-integrin/HIF-1α Axis in CRC tumor microenvironment. IJMS 24, 4988. doi:10.3390/ijms24054988
Bromberg, J. F. (2001). Activation of STAT proteins and growth control. Bioessays 23, 161–169. doi:10.1002/1521-1878(200102)23:2<161::AID-BIES1023>3.0.CO;2-0
Brown, J. M., and Wilson, W. R. (2004). Exploiting tumour hypoxia in cancer treatment. Nat. Rev. Cancer 4, 437–447. doi:10.1038/nrc1367
Bu, X., Zhao, C., Wang, W., and Zhang, N. (2013). GRIM-19 inhibits the STAT3 signaling pathway and sensitizes gastric cancer cells to radiation. Gene 512, 198–205. doi:10.1016/j.gene.2012.10.057
Buhrmann, C., Shayan, P., Popper, B., Goel, A., and Shakibaei, M. (2016). Sirt1 is required for resveratrol-mediated chemopreventive effects in colorectal cancer cells. Nutrients 8, 145. doi:10.3390/nu8030145
Buhrmann, C., Yazdi, M., Popper, B., Shayan, P., Goel, A., Aggarwal, B., et al. (2018). Resveratrol chemosensitizes TNF-β-induced survival of 5-FU-treated colorectal cancer cells. Nutrients 10, 888. doi:10.3390/nu10070888
Burns, J., Yokota, T., Ashihara, H., Lean, M. E. J., and Crozier, A. (2002). Plant foods and herbal sources of resveratrol. J. Agric. Food Chem. 50, 3337–3340. doi:10.1021/jf0112973
Buser, K., Joncourt, F., Altermatt, H.-J., Bacchi, M., Oberli, A., and Cerny, Th. (1997). Breast cancer: pretreatment drug resistance parameters (GSH-system, ATase, P-glycoprotein) in tumor tissue and their correlation with clinical and prognostic characteristics. Ann. Oncol. 8, 335–341. doi:10.1023/A:1008202723066
Carneiro, B. A., and El-Deiry, W. S. (2020). Targeting apoptosis in cancer therapy. Nat. Rev. Clin. Oncol. 17, 395–417. doi:10.1038/s41571-020-0341-y
Carnero, A., and Lleonart, M. (2016). The hypoxic microenvironment: a determinant of cancer stem cell evolution. Bioessays 38 (1), S65–S74. doi:10.1002/bies.201670911
Carsten, R. E., Bachand, A. M., Bailey, S. M., and Ullrich, R. L. (2008). Resveratrol reduces radiation-induced chromosome aberration frequencies in mouse bone marrow cells. Radiat. Res. 169, 633–638. doi:10.1667/RR1190.1
Carter, L. G., D’Orazio, J. A., and Pearson, K. J. (2014). Resveratrol and cancer: focus on in vivo evidence. Endocrine-Related Cancer 21, R209–R225. doi:10.1530/ERC-13-0171
Catanzaro, D., Gabbia, D., Cocetta, V., Biagi, M., Ragazzi, E., Montopoli, M., et al. (2018). Silybin counteracts doxorubicin resistance by inhibiting GLUT1 expression. Fitoterapia 124, 42–48. doi:10.1016/j.fitote.2017.10.007
Catlett-Falcone, R., Landowski, T. H., Oshiro, M. M., Turkson, J., Levitzki, A., Savino, R., et al. (1999). Constitutive activation of Stat3 signaling confers resistance to apoptosis in human U266 myeloma cells. Immunity 10, 105–115. doi:10.1016/s1074-7613(00)80011-4
Chandel, N. S., Schumacker, P. T., and Arch, R. H. (2001). Reactive oxygen species are downstream products of TRAF-mediated signal transduction. J. Biol. Chem. 276, 42728–42736. doi:10.1074/jbc.M103074200
Chang, L., Graham, P., Hao, J., Ni, J., Deng, J., Bucci, J., et al. (2016). Cancer stem cells and signaling pathways in radioresistance. Oncotarget 7, 11002–11017. doi:10.18632/oncotarget.6760
Chao, C. C., Huang, Y. T., Ma, C. M., Chou, W. Y., and Lin-Chao, S. (1992). Overexpression of glutathione S-transferase and elevation of thiol pools in a multidrug-resistant human colon cancer cell line. Mol. Pharmacol. 41, 69–75.
Chen, J., Dong, X., and Guo, X. (2009). Inhibitory effect of resveratrol on the growth of human colon cancer ls174t cells and its subcutaneously transplanted tumor in nude mice and the mechanism of action. Zhonghua Zhong Liu Za Zhi 31, 15–19.
Cheuk, I. W., Chen, J., Siu, M., Ho, J. C., Lam, S. S., Shin, V. Y., et al. (2022). Resveratrol enhanced chemosensitivity by reversing macrophage polarization in breast cancer. Clin. Transl. Oncol. 24, 854–863. doi:10.1007/s12094-021-02731-5
Chimento, A., De Amicis, F., Sirianni, R., Sinicropi, M. S., Puoci, F., Casaburi, I., et al. (2019). Progress to improve oral bioavailability and beneficial effects of resveratrol. Int. J. Mol. Sci. 20, 1381. doi:10.3390/ijms20061381
Choi, C. Y., Lim, S.-C., Lee, T.-B., and Han, S. I. (2022). Molecular basis of resveratrol-induced resensitization of acquired drug-resistant cancer cells. Nutrients 14, 699. doi:10.3390/nu14030699
Chouaib, S., Noman, M. Z., Kosmatopoulos, K., and Curran, M. A. (2017). Hypoxic stress: obstacles and opportunities for innovative immunotherapy of cancer. Oncogene 36, 439–445. doi:10.1038/onc.2016.225
Chuang, S.-E., Yeh, P.-Y., Lu, Y.-S., Lai, G.-M., Liao, C.-M., Gao, M., et al. (2002). Basal levels and patterns of anticancer drug-induced activation of nuclear factor-kappaB (NF-kappaB), and its attenuation by tamoxifen, dexamethasone, and curcumin in carcinoma cells. Biochem. Pharmacol. 63, 1709–1716. doi:10.1016/S0006-2952(02)00931-0
Chung, S. S., Dutta, P., Austin, D., Wang, P., Awad, A., and Vadgama, J. V. (2018). Combination of resveratrol and 5-flurouracil enhanced anti-telomerase activity and apoptosis by inhibiting STAT3 and Akt signaling pathways in human colorectal cancer cells. Oncotarget 9, 32943–32957. doi:10.18632/oncotarget.25993
Cocetta, V., Quagliariello, V., Fiorica, F., Berretta, M., and Montopoli, M. (2021). Resveratrol as chemosensitizer agent: state of art and future perspectives. IJMS 22, 2049. doi:10.3390/ijms22042049
Cragg, G. M., and Pezzuto, J. M. (2016). Natural products as a vital source for the discovery of cancer chemotherapeutic and chemopreventive agents. Med. Princ. Pract. 25, 41–59. doi:10.1159/000443404
Crook, J. M., Perry, G. A., Robertson, S., and Esche, B. A. (1995). Routine prostate biopsies following radiotherapy for prostate cancer: results for 226 patients. Urology 45, 624–631. doi:10.1016/S0090-4295(99)80054-5
Crouthamel, M. H., Wu, D., Yang, Z., and Ho, R. J. Y. (2006). A novel MDR1 G1199T variant alters drug resistance and efflux transport activity of P-glycoprotein in recombinant Hek cells. J. Pharm. Sci. 95, 2767–2777. doi:10.1002/jps.20743
Cruceriu, D., Baldasici, O., Balacescu, O., and Berindan-Neagoe, I. (2020). The dual role of tumor necrosis factor-alpha (TNF-α) in breast cancer: molecular insights and therapeutic approaches. Cell Oncol. 43, 1–18. doi:10.1007/s13402-019-00489-1
Cusack, J. C., Liu, R., and Baldwin, A. S. (2000). Inducible chemoresistance to 7-ethyl-10-[4-(1-piperidino)-1-piperidino]-carbonyloxycamptothe cin (CPT-11) in colorectal cancer cells and a xenograft model is overcome by inhibition of nuclear factor-kappaB activation. Cancer Res. 60, 2323–2330.
Das, P. K., Islam, F., and Lam, A. K. (2020). The roles of cancer stem cells and therapy resistance in colorectal carcinoma. Cells 9, 1392. doi:10.3390/cells9061392
Deng, J., Bai, X., Feng, X., Ni, J., Beretov, J., Graham, P., et al. (2019). Inhibition of PI3K/Akt/mTOR signaling pathway alleviates ovarian cancer chemoresistance through reversing epithelial-mesenchymal transition and decreasing cancer stem cell marker expression. BMC Cancer 19, 618. doi:10.1186/s12885-019-5824-9
Deng, X., Kornblau, S. M., Ruvolo, P. P., and May, W. S. (2000). Regulation of Bcl2 phosphorylation and potential significance for leukemic cell chemoresistance. JNCI Monogr. 2000, 30–37. doi:10.1093/oxfordjournals.jncimonographs.a024254
Dercks, W., and Creasy, L. L. (1989). The significance of stilbene phytoalexins in the Plasmopara viticola-grapevine interaction. Physiological Mol. Plant Pathology 34, 189–202. doi:10.1016/0885-5765(89)90043-X
De Vries, K., Strydom, M., and Steenkamp, V. (2021). A brief updated review of advances to enhance resveratrol’s bioavailability. Molecules 26, 4367. doi:10.3390/molecules26144367
Doktorova, H., Hrabeta, J., Khalil, M. A., and Eckschlager, T. (2015). Hypoxia-induced chemoresistance in cancer cells: the role of not only HIF-1. Biomed. Pap. Med. Fac. Univ. Palacky. Olomouc Czech Repub. 159, 166–177. doi:10.5507/bp.2015.025
Dolcet, X., Llobet, D., Pallares, J., and Matias-Guiu, X. (2005). NF-kB in development and progression of human cancer. Virchows Arch. 446, 475–482. doi:10.1007/s00428-005-1264-9
El-Benhawy, S. A., Morsi, M. I., Fahmy, E. I., Soula, M. A., Khalil, F. A., and Arab, A. (2021). Role of resveratrol as radiosensitizer by targeting cancer stem cells in radioresistant prostate cancer cells (PC-3). Asian Pac J. Cancer Prev. 22, 3823–3837. doi:10.31557/APJCP.2021.22.12.3823
Fabbrocini, G., Kisslinger, A., Iannelli, P., Vitale, N., Procaccini, C., Sparaneo, G., et al. (2010). Resveratrol regulates p66Shc activation in HaCaT cells: on the interaction between resveratrol and p66Shc. Exp. Dermatol. 19, 895–903. doi:10.1111/j.1600-0625.2010.01117.x
Fan, Y., Mao, R., and Yang, J. (2013). NF-κB and STAT3 signaling pathways collaboratively link inflammation to cancer. Protein Cell 4, 176–185. doi:10.1007/s13238-013-2084-3
Fanelli, M., Hattinger, C. M., Vella, S., Tavanti, E., Michelacci, F., Gudeman, B., et al. (2016). Targeting ABCB1 and ABCC1 with their specific inhibitor CBT-1® can overcome drug resistance in osteosarcoma. Curr. Cancer Drug Targets 16, 261–274. doi:10.2174/1568009616666151106120434
Fang, Y., Bradley, M. J., Cook, K. M., Herrick, E. J., and Nicholl, M. B. (2013). A potential role for resveratrol as a radiation sensitizer for melanoma treatment. J. Surg. Res. 183, 645–653. doi:10.1016/j.jss.2013.02.037
Fang, Y., DeMarco, V. G., and Nicholl, M. B. (2012). Resveratrol enhances radiation sensitivity in prostate cancer by inhibiting cell proliferation and promoting cell senescence and apoptosis. Cancer Sci. 103, 1090–1098. doi:10.1111/j.1349-7006.2012.02272.x
Fatease, A. A., Shah, V., Nguyen, D. X., Cote, B., LeBlanc, N., Rao, D. A., et al. (2019). Chemosensitization and mitigation of Adriamycin-induced cardiotoxicity using combinational polymeric micelles for co-delivery of quercetin/resveratrol and resveratrol/curcumin in ovarian cancer. Nanomedicine Nanotechnol. Biol. Med. 19, 39–48. doi:10.1016/j.nano.2019.03.011
Fenig, E., Nordenberg, J., Beery, E., Sulkes, J., and Wasserman, L. (2004). Combined effect of aloe-emodin and chemotherapeutic agents on the proliferation of an adherent variant cell line of Merkel cell carcinoma. Oncol. Rep. 11, 213–217. doi:10.3892/or.11.1.213
Ferraz da Costa, D. C., Casanova, F. A., Quarti, J., Malheiros, M. S., Sanches, D., Dos Santos, P. S., et al. (2012). Transient transfection of a wild-type p53 gene triggers resveratrol-induced apoptosis in cancer cells. PLoS One 7, e48746. doi:10.1371/journal.pone.0048746
Ferraz da Costa, D. C., Pereira Rangel, L., Martins-Dinis, M. M. D. da C., Ferretti, G. D. da S., Ferreira, V. F., and Silva, J. L. (2020). Anticancer potential of resveratrol, β-lapachone and their analogues. Molecules 25, 893. doi:10.3390/molecules25040893
Fink, M. Y., and Chipuk, J. E. (2013). Survival of HER2-positive breast cancer cells: receptor signaling to apoptotic control centers. Genes & Cancer 4, 187–195. doi:10.1177/1947601913488598
Flynn, V., Ramanitharan, A., Moparty, K., Davis, R., Sikka, S., Agrawal, K. C., et al. (2003). Adenovirus-mediated inhibition of NF-κB confers chemo-sensitization and apoptosis in prostate cancer cells. Int. J. Oncol. 23, 317–323. doi:10.3892/ijo.23.2.317
Fodale, V., Pierobon, M., Liotta, L., and Petricoin, E. (2011). Mechanism of cell adaptation: when and how do cancer cells develop chemoresistance? Cancer J. 17, 89–95. doi:10.1097/PPO.0b013e318212dd3d
Fontenot, J. D., Gavin, M. A., and Rudensky, A. Y. (2003). Foxp3 programs the development and function of CD4+CD25+ regulatory T cells. Nat. Immunol. 4, 330–336. doi:10.1038/ni904
Frampton, G. A., Lazcano, E. A., Li, H., Mohamad, A., and DeMorrow, S. (2010). Resveratrol enhances the sensitivity of cholangiocarcinoma to chemotherapeutic agents. Lab. Investig. 90, 1325–1338. doi:10.1038/labinvest.2010.99
Fu, Y., Sun, X., Gu, Z., and Zhuang, Z. (2020). Connexin 43 modulates the cellular resistance to paclitaxel via targeting β-tubulin in triple-negative breast cancer. Onco Targets Ther. 13, 5323–5335. doi:10.2147/OTT.S229076
Fulda, S., and Debatin, K.-M. (2004). Sensitization for anticancer drug-induced apoptosis by the chemopreventive agent resveratrol. Oncogene 23, 6702–6711. doi:10.1038/sj.onc.1207630
Galanski, M. (2006). Recent developments in the field of anticancer platinum complexes. PRA 1, 285–295. doi:10.2174/157489206777442287
Galluzzi, L., Senovilla, L., Vitale, I., Michels, J., Martins, I., Kepp, O., et al. (2012). Molecular mechanisms of cisplatin resistance. Oncogene 31, 1869–1883. doi:10.1038/onc.2011.384
Gao, Z., Xu, J., Fan, Y., Qi, Y., Wang, S., Zhao, S., et al. (2022). PDIA3P1 promotes Temozolomide resistance in glioblastoma by inhibiting C/EBPβ degradation to facilitate proneural-to-mesenchymal transition. J. Exp. Clin. Cancer Res. 41, 223. doi:10.1186/s13046-022-02431-0
Garg, A. K., and Aggarwal, B. B. (2002). Reactive oxygen intermediates in TNF signaling. Mol. Immunol. 39, 509–517. doi:10.1016/S0161-5890(02)00207-9
Garg, A. K., Buchholz, T. A., and Aggarwal, B. B. (2005). Chemosensitization and radiosensitization of tumors by plant polyphenols. Antioxidants Redox Signal. 7, 1630–1647. doi:10.1089/ars.2005.7.1630
Garg, A. K., Hortobagyi, G. N., Aggarwal, B. B., Sahin, A. A., and Buchholz, T. A. (2003). Nuclear factor-kappa B as a predictor of treatment response in breast cancer. Curr. Opin. Oncol. 15, 405–411. doi:10.1097/00001622-200311000-00001
Gatouillat, G., Balasse, E., Joseph-Pietras, D., Morjani, H., and Madoulet, C. (2010). Resveratrol induces cell-cycle disruption and apoptosis in chemoresistant B16 melanoma. J. Cell. Biochem. 110, 893–902. doi:10.1002/jcb.22601
Gilreath, C., Boerma, M., Qin, Z., Hudson, M. K., and Wang, S. (2021). The hypoxic microenvironment of breast cancer cells promotes resistance in radiation therapy. Front. Oncol. 10, 629422. doi:10.3389/fonc.2020.629422
Giordo, R., Wehbe, Z., Posadino, A. M., Erre, G. L., Eid, A. H., Mangoni, A. A., et al. (2022). Disease-associated regulation of non-coding RNAs by resveratrol: molecular insights and therapeutic applications. Front. Cell Dev. Biol. 10, 894305. doi:10.3389/fcell.2022.894305
Godwin, A. K., Meister, A., O’Dwyer, P. J., Huang, C. S., Hamilton, T. C., and Anderson, M. E. (1992). High resistance to cisplatin in human ovarian cancer cell lines is associated with marked increase of glutathione synthesis. Proc. Natl. Acad. Sci. U.S.A. 89, 3070–3074. doi:10.1073/pnas.89.7.3070
Gorrini, C., Harris, I. S., and Mak, T. W. (2013). Modulation of oxidative stress as an anticancer strategy. Nat. Rev. Drug Discov. 12, 931–947. doi:10.1038/nrd4002
Gottesman, M. M., Fojo, T., and Bates, S. E. (2002). Multidrug resistance in cancer: role of ATP-dependent transporters. Nat. Rev. Cancer 2, 48–58. doi:10.1038/nrc706
Gowda, R., Sharma, A., and Robertson, G. P. (2017). Synergistic inhibitory effects of Celecoxib and Plumbagin on melanoma tumor growth. Cancer Lett. 385, 243–250. doi:10.1016/j.canlet.2016.10.016
Greten, F. R., Weber, C. K., Greten, T. F., Schneider, G., Wagner, M., Adler, G., et al. (2002). Stat3 and NF-kappaB activation prevents apoptosis in pancreatic carcinogenesis. Gastroenterology 123, 2052–2063. doi:10.1053/gast.2002.37075
Gu, J., Li, Z., Zhou, J., Sun, Z., and Bai, C. (2019). Response prediction to oxaliplatin plus 5-fluorouracil chemotherapy in patients with colorectal cancer using a four-protein immunohistochemical model. Oncol. Lett. 18, 2091–2101. doi:10.3892/ol.2019.10474
Gupta, S. C., Kannappan, R., Reuter, S., Kim, J. H., and Aggarwal, B. B. (2011). Chemosensitization of tumors by resveratrol: resveratrol and cancer. Ann. N. Y. Acad. Sci. 1215, 150–160. doi:10.1111/j.1749-6632.2010.05852.x
Gutiérrez-Puente, Y., Zapata-Benavides, P., Tari, A. M., and López-Berestein, G. (2002). Bcl-2-related antisense therapy. Semin. Oncol. 29, 71–76. doi:10.1053/sonc.2002.34058
Gutschner, T., Hämmerle, M., Eißmann, M., Hsu, J., Kim, Y., Hung, G., et al. (2013). The noncoding RNA MALAT1 is a critical regulator of the metastasis phenotype of lung cancer cells. Cancer Res. 73, 1180–1189. doi:10.1158/0008-5472.CAN-12-2850
Habibie, H., Yokoyama, S., Abdelhamed, S., Awale, S., Sakurai, H., Hayakawa, Y., et al. (2014). Survivin suppression through STAT3/β-catenin is essential for resveratrol-induced melanoma apoptosis. Int. J. Oncol. 45, 895–901. doi:10.3892/ijo.2014.2480
Habrand, J. L., and De Crevoisier, R. (2001). Radiation therapy in the management of childhood brain tumors. Childs Nerv. Syst. 17, 121–133. doi:10.1007/s003810000365
Hagan, M., Wang, L., Hanley, J. R., Park, J. S., and Dent, P. (2000). Ionizing radiation-induced mitogen-activated protein (MAP) kinase activation in DU145 prostate carcinoma cells: MAP kinase inhibition enhances radiation-induced cell killing and G 2/M-phase arrest. Radiat. Res. 153, 371–383. doi:10.1667/0033-7587(2000)153[0371:IRIMAP]2.0.CO;2
Harikumar, K. B., Kunnumakkara, A. B., Sethi, G., Diagaradjane, P., Anand, P., Pandey, M. K., et al. (2010). Resveratrol, a multitargeted agent, can enhance antitumor activity of gemcitabine in vitro and in orthotopic mouse model of human pancreatic cancer. Int. J. Cancer, NA-NA 127, 257–268. doi:10.1002/ijc.25041
Harris, A. L. (2002). Hypoxia-a key regulatory factor in tumour growth. Nat. Rev. Cancer 2, 38–47. doi:10.1038/nrc704
Harris, R. E. (2009). Cyclooxygenase-2 (cox-2) blockade in the chemoprevention of cancers of the colon, breast, prostate, and lung. Inflammopharmacology 17, 55–67. doi:10.1007/s10787-009-8049-8
Harte, M. T., Gorski, J. J., Savage, K. I., Purcell, J. W., Barros, E. M., Burn, P. M., et al. (2014). NF-κB is a critical mediator of BRCA1-induced chemoresistance. Oncogene 33, 713–723. doi:10.1038/onc.2013.10
Hartwich, J., Orr, W. S., Ng, C. Y., Spence, Y., Morton, C., and Davidoff, A. M. (2013). HIF-1α activation mediates resistance to anti-angiogenic therapy in neuroblastoma xenografts. J. Pediatr. Surg. 48, 39–46. doi:10.1016/j.jpedsurg.2012.10.016
He, Q., Li, Z., Wang, Y., Hou, Y., Li, L., and Zhao, J. (2017). Resveratrol alleviates cerebral ischemia/reperfusion injury in rats by inhibiting NLRP3 inflammasome activation through Sirt1-dependent autophagy induction. Int. Immunopharmacol. 50, 208–215. doi:10.1016/j.intimp.2017.06.029
Hernandez-Valencia, J., Garcia-Villa, E., Arenas-Hernandez, A., Garcia-Mena, J., Diaz-Chavez, J., and Gariglio, P. (2018). Induction of p53 phosphorylation at serine 20 by resveratrol is required to activate p53 target genes, restoring apoptosis in MCF-7 cells resistant to cisplatin. Nutrients 10, 1148. doi:10.3390/nu10091148
Hochwald, S. N., Rose, D. M., Brennan, M. F., and Burt, M. E. (1997). Elevation of glutathione and related enzyme activities in high-grade and metastatic extremity soft tissue sarcoma. Ann. Surg. Oncol. 4, 303–309. doi:10.1007/BF02303579
Holohan, C., Van Schaeybroeck, S., Longley, D. B., and Johnston, P. G. (2013). Cancer drug resistance: an evolving paradigm. Nat. Rev. Cancer 13, 714–726. doi:10.1038/nrc3599
Hong, T., Ding, J., and Li, W. (2019). miR-7 reverses breast cancer resistance to chemotherapy by targeting MRP1 and BCL2. OTT 12, 11097–11105. doi:10.2147/OTT.S213780
Hövelmann, S., Beckers, T. L., and Schmidt, M. (2004). Molecular alterations in apoptotic pathways after PKB/Akt-mediated chemoresistance in NCI H460 cells. Br. J. Cancer 90, 2370–2377. doi:10.1038/sj.bjc.6601876
Hsu, C.-H., Altschuler, S. J., and Wu, L. F. (2019). Patterns of early p21 dynamics determine proliferation-senescence cell fate after chemotherapy. Cell 178, 361–373. doi:10.1016/j.cell.2019.05.041
Hsu, H., Chen, M., Baskaran, R., Lin, Y., Day, C. H., Lin, Y., et al. (2018). Oxaliplatin resistance in colorectal cancer cells is mediated via activation of ABCG2 to alleviate ER stress induced apoptosis. J. Cell. Physiology 233, 5458–5467. doi:10.1002/jcp.26406
Hu, S., Li, X., Xu, R., Ye, L., Kong, H., Zeng, X., et al. (2016a). The synergistic effect of resveratrol in combination with cisplatin on apoptosis via modulating autophagy in A549 cells. ABBS 48, 528–535. doi:10.1093/abbs/gmw026
Hu, Y.-Q., Wang, J., and Wu, J.-H. (2016b). Administration of resveratrol enhances cell-cycle arrest followed by apoptosis in DMBA-induced skin carcinogenesis in male Wistar rats. Eur. Rev. Med. Pharmacol. Sci. 20, 2935–2946.
Huang, C.-Y., Tai, W.-T., Hsieh, C.-Y., Hsu, W.-M., Lai, Y.-J., Chen, L.-J., et al. (2014). A sorafenib derivative and novel SHP-1 agonist, SC-59, acts synergistically with radiotherapy in hepatocellular carcinoma cells through inhibition of STAT3. Cancer Lett. 349, 136–143. doi:10.1016/j.canlet.2014.04.006
Huang, C.-Y., Tai, W.-T., Wu, S.-Y., Shih, C.-T., Chen, M.-H., Tsai, M.-H., et al. (2016). Dovitinib acts as a novel radiosensitizer in hepatocellular carcinoma by targeting SHP-1/STAT3 signaling. Int. J. Radiat. Oncol. Biol. Phys. 95, 761–771. doi:10.1016/j.ijrobp.2016.01.016
Huang, L., Ao, Q., Zhang, Q., Yang, X., Xing, H., Li, F., et al. (2010). Hypoxia induced paclitaxel resistance in human ovarian cancers via hypoxia-inducible factor 1alpha. J. Cancer Res. Clin. Oncol. 136, 447–456. doi:10.1007/s00432-009-0675-4
Huang, Q., Wu, Y.-Y., Xing, S.-J., and Yu, Z.-W. (2019). Effect of miR-7 on resistance of breast cancer cells to adriamycin via regulating EGFR/PI3K signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 23, 5285–5292. doi:10.26355/eurrev_201906_18195
Huang, S. M., Bock, J. M., and Harari, P. M. (1999). Epidermal growth factor receptor blockade with C225 modulates proliferation, apoptosis, and radiosensitivity in squamous cell carcinomas of the head and neck. Cancer Res. 59, 1935–1940.
Huang, S. M., and Harari, P. M. (2000). Modulation of radiation response after epidermal growth factor receptor blockade in squamous cell carcinomas: inhibition of damage repair, cell cycle kinetics, and tumor angiogenesis. Clin. Cancer Res. 6, 2166–2174.
Huang, Y., and Sadée, W. (2006). Membrane transporters and channels in chemoresistance and -sensitivity of tumor cells. Cancer Lett. 239, 168–182. doi:10.1016/j.canlet.2005.07.032
Huang, Y., Yu, J., Yan, C., Hou, J., Pu, J., Zhang, G., et al. (2012). Effect of small interfering RNA targeting hypoxia-inducible factor-1α on radiosensitivity of PC3 cell line. Urology 79, 744.e17–744.e24. doi:10.1016/j.urology.2011.10.024
Ivanov, V. N., Zhou, H., and Hei, T. K. (2007). Sequential treatment by ionizing radiation and sodium arsenite dramatically accelerates TRAIL-mediated apoptosis of human melanoma cells. Cancer Res. 67, 5397–5407. doi:10.1158/0008-5472.CAN-07-0551
Ivanova, D., Zhelev, Z., Semkova, S., Aoki, I., and Bakalova, R. (2019). Resveratrol modulates the redox-status and cytotoxicity of anticancer drugs by sensitizing leukemic lymphocytes and protecting normal lymphocytes. Anticancer Res. 39, 3745–3755. doi:10.21873/anticanres.13523
Jaisamut, P., Wanna, S., Limsuwan, S., Chusri, S., Wiwattanawongsa, K., and Wiwattanapatapee, R. (2021). Enhanced oral bioavailability and improved biological activities of a quercetin/resveratrol combination using a liquid self-microemulsifying drug delivery system. Planta Med. 87, 336–346. doi:10.1055/a-1270-7606
Jang, M., Cai, L., Udeani, G. O., Slowing, K. V., Thomas, C. F., Beecher, C. W. W., et al. (1997). Cancer chemopreventive activity of resveratrol, a natural product derived from grapes. Science 275, 218–220. doi:10.1126/science.275.5297.218
Jazirehi, A. R., and Bonavida, B. (2004). Resveratrol modifies the expression of apoptotic regulatory proteins and sensitizes non-Hodgkin’s lymphoma and multiple myeloma cell lines to paclitaxel-induced apoptosis. Mol. Cancer Ther. 3, 71–84. doi:10.1158/1535-7163.71.3.1
Jeon, D., Jo, M., Lee, Y., Park, S.-H., Phan, H. T. L., Nam, J. H., et al. (2023). Inhibition of ANO1 by cis- and trans-resveratrol and their anticancer activity in human prostate cancer PC-3 cells. IJMS 24, 1186. doi:10.3390/ijms24021186
Jeong, H., Kim, S., Hong, B.-J., Lee, C.-J., Kim, Y.-E., Bok, S., et al. (2019). Tumor-associated macrophages enhance tumor hypoxia and aerobic glycolysis. Cancer Res. 79, 795–806. doi:10.1158/0008-5472.CAN-18-2545
Ji, Q., Zhang, L., Liu, X., Zhou, L., Wang, W., Han, Z., et al. (2014). Long non-coding RNA MALAT1 promotes tumour growth and metastasis in colorectal cancer through binding to SFPQ and releasing oncogene PTBP2 from SFPQ/PTBP2 complex. Br. J. Cancer 111, 736–748. doi:10.1038/bjc.2014.383
Jin, X., Wei, Y., Liu, Y., Lu, X., Ding, F., Wang, J., et al. (2019). Resveratrol promotes sensitization to Doxorubicin by inhibiting epithelial-mesenchymal transition and modulating SIRT1/β-catenin signaling pathway in breast cancer. Cancer Med. 8, 1246–1257. doi:10.1002/cam4.1993
Jung, M., and Dritschilo, A. (2001). NF-κB signaling pathway as a target for human tumor radiosensitization. Seminars Radiat. Oncol. 11, 346–351. doi:10.1053/srao.2001.26034
Kaminski, B. M., Weigert, A., Scherzberg, M.-C., Ley, S., Gilbert, B., Brecht, K., et al. (2014). Resveratrol-induced potentiation of the antitumor effects of oxaliplatin is accompanied by an altered cytokine profile of human monocyte-derived macrophages. Apoptosis 19, 1136–1147. doi:10.1007/s10495-014-0988-x
Kaufmann, S. H., and Vaux, D. L. (2003). Alterations in the apoptotic machinery and their potential role in anticancer drug resistance. Oncogene 22, 7414–7430. doi:10.1038/sj.onc.1206945
Keith, B., and Simon, M. C. (2007). Hypoxia-inducible factors, stem cells, and cancer. Cell 129, 465–472. doi:10.1016/j.cell.2007.04.019
Kelland, L. (2007). The resurgence of platinum-based cancer chemotherapy. Nat. Rev. Cancer 7, 573–584. doi:10.1038/nrc2167
Khaleel, S. A., Al-Abd, A. M., Ali, A. A., and Abdel-Naim, A. B. (2016). Didox and resveratrol sensitize colorectal cancer cells to doxorubicin via activating apoptosis and ameliorating P-glycoprotein activity. Sci. Rep. 6, 36855. doi:10.1038/srep36855
Khoei, S., Shoja, M., Mostaar, A., and Faeghi, F. (2016). Effects of resveratrol and methoxyamine on the radiosensitivity of iododeoxyuridine in U87MG glioblastoma cell line. Exp. Biol. Med. (Maywood) 241, 1229–1236. doi:10.1177/1535370215622583
Kim, R., Tanabe, K., Uchida, Y., Emi, M., and Toge, T. (2003). Effect of Bcl-2 antisense oligonucleotide on drug-sensitivity in association with apoptosis in undifferentiated thyroid carcinoma. Int. J. Mol. Med. 11, 799–804. doi:10.3892/ijmm.11.6.799
Kim, T. H., Shin, Y. J., Won, A. J., Lee, B. M., Choi, W. S., Jung, J. H., et al. (2014). Resveratrol enhances chemosensitivity of doxorubicin in multidrug-resistant human breast cancer cells via increased cellular influx of doxorubicin. Biochimica Biophysica Acta (BBA) - General Subj. 1840, 615–625. doi:10.1016/j.bbagen.2013.10.023
Kishi, K., Petersen, S., Petersen, C., Hunter, N., Mason, K., Masferrer, J. L., et al. (2000). Preferential enhancement of tumor radioresponse by a cyclooxygenase-2 inhibitor. Cancer Res. 60, 1326–1331.
Kisková, T., and Kassayová, M. (2019). Resveratrol action on lipid metabolism in cancer. IJMS 20, 2704. doi:10.3390/ijms20112704
Kma, L. (2013). Synergistic effect of resveratrol and radiotherapy in control of cancers. Asian Pac. J. Cancer Prev. 14, 6197–6208. doi:10.7314/APJCP.2013.14.11.6197
Ko, J.-H., Sethi, G., Um, J.-Y., Shanmugam, M. K., Arfuso, F., Kumar, A. P., et al. (2017). The role of resveratrol in cancer therapy. IJMS 18, 2589. doi:10.3390/ijms18122589
Komorowska, D., Radzik, T., Kalenik, S., and Rodacka, A. (2022). Natural radiosensitizers in radiotherapy: cancer treatment by combining ionizing radiation with resveratrol. IJMS 23, 10627. doi:10.3390/ijms231810627
Kosztyu, P., Bukvova, R., Dolezel, P., and Mlejnek, P. (2014). Resistance to daunorubicin, imatinib, or nilotinib depends on expression levels of ABCB1 and ABCG2 in human leukemia cells. Chem. Biol. Interact. 219, 203–210. doi:10.1016/j.cbi.2014.06.009
Kotecha, R., Takami, A., and Espinoza, J. L. (2016). Dietary phytochemicals and cancer chemoprevention: a review of the clinical evidence. Oncotarget 7, 52517–52529. doi:10.18632/oncotarget.9593
Kucinska, M., Piotrowska, H., Luczak, M. W., Mikula-Pietrasik, J., Ksiazek, K., Wozniak, M., et al. (2014). Effects of hydroxylated resveratrol analogs on oxidative stress and cancer cells death in human acute T cell leukemia cell line: prooxidative potential of hydroxylated resveratrol analogs. Chemico-Biological Interact. 209, 96–110. doi:10.1016/j.cbi.2013.12.009
Kuršvietienė, L., Stanevičienė, I., Mongirdienė, A., and Bernatonienė, J. (2016). Multiplicity of effects and health benefits of resveratrol. Medicina 52, 148–155. doi:10.1016/j.medici.2016.03.003
Kweon, S. H., Song, J. H., and Kim, T. S. (2010). Resveratrol-mediated reversal of doxorubicin resistance in acute myeloid leukemia cells via downregulation of MRP1 expression. Biochem. Biophysical Res. Commun. 395, 104–110. doi:10.1016/j.bbrc.2010.03.147
Lai, M., Yang, Z., Zhou, L., Zhu, Q., Xie, H., Zhang, F., et al. (2012). Long non-coding RNA MALAT-1 overexpression predicts tumor recurrence of hepatocellular carcinoma after liver transplantation. Med. Oncol. 29, 1810–1816. doi:10.1007/s12032-011-0004-z
Lee, J. H., Kim, C., Ko, J.-H., Jung, Y. Y., Jung, S. H., Kim, E., et al. (2019). Casticin inhibits growth and enhances ionizing radiation-induced apoptosis through the suppression of STAT3 signaling cascade. J. Cell Biochem. 120, 9787–9798. doi:10.1002/jcb.28259
Lee, P., Chiou, Y., Ho, C., and Pan, M. (2018). Chemoprevention by resveratrol and pterostilbene: targeting on epigenetic regulation. BioFactors 44, 26–35. doi:10.1002/biof.1401
Lee, S. H., Koo, B. S., Park, S. Y., and Kim, Y. M. (2015). Anti-angiogenic effects of resveratrol in combination with 5-fluorouracil on B16 murine melanoma cells. Mol. Med. Rep. 12, 2777–2783. doi:10.3892/mmr.2015.3675
Lee, S. Y. (2016). Temozolomide resistance in glioblastoma multiforme. Genes & Dis. 3, 198–210. doi:10.1016/j.gendis.2016.04.007
Lee, Y.-J., Lee, G. J., Yi, S. S., Heo, S.-H., Park, C.-R., Nam, H.-S., et al. (2016). Cisplatin and resveratrol induce apoptosis and autophagy following oxidative stress in malignant mesothelioma cells. Food Chem. Toxicol. 97, 96–107. doi:10.1016/j.fct.2016.08.033
Lele, W., Lei, L., and Liting, Q. (2021). Resveratrol sensitizes A549 cells to irradiation damage via suppression of store-operated calcium entry with Orai1 and STIM1 downregulation. Exp. Ther. Med. 21, 587. doi:10.3892/etm.2021.10019
Leonard, G. D., Fojo, T., and Bates, S. E. (2003). The role of ABC transporters in clinical practice. Oncologist 8, 411–424. doi:10.1634/theoncologist.8-5-411
Leone, S., Fiore, M., Lauro, M. G., Pino, S., Cornetta, T., and Cozzi, R. (2008). Resveratrol and X rays affect gap junction intercellular communications in human glioblastoma cells. Mol. Carcinog. 47, 587–598. doi:10.1002/mc.20416
Leung, E., Cairns, R. A., Chaudary, N., Vellanki, R. N., Kalliomaki, T., Moriyama, E. H., et al. (2017). Metabolic targeting of HIF-dependent glycolysis reduces lactate, increases oxygen consumption and enhances response to high-dose single-fraction radiotherapy in hypoxic solid tumors. BMC Cancer 17, 418. doi:10.1186/s12885-017-3402-6
Levine, A. J. (1997). p53, the cellular gatekeeper for growth and division. Cell 88, 323–331. doi:10.1016/s0092-8674(00)81871-1
Levy, D. E., and Darnell, J. E. (2002). Stats: transcriptional control and biological impact. Nat. Rev. Mol. Cell Biol. 3, 651–662. doi:10.1038/nrm909
Li, H., Liu, Y., Jiao, Y., Guo, A., Xu, X., Qu, X., et al. (2016). Resveratrol sensitizes glioblastoma-initiating cells to temozolomide by inducing cell apoptosis and promoting differentiation. Oncol. Rep. 35, 343–351. doi:10.3892/or.2015.4346
Li, F., Han, Y., Wu, X., Cao, X., Gao, Z., Sun, Y., et al. (2022). Gut microbiota-derived resveratrol metabolites, dihydroresveratrol and lunularin, significantly contribute to the biological activities of resveratrol. Front. Nutr. 9, 912591. doi:10.3389/fnut.2022.912591
Li, S., Jiang, M., Wang, L., and Yu, S. (2020). Combined chemotherapy with cyclooxygenase-2 (COX-2) inhibitors in treating human cancers: recent advancement. Biomed. Pharmacother. 129, 110389. doi:10.1016/j.biopha.2020.110389
Li, S. G., and Li, L. (2013). Targeted therapy in HER2-positive breast cancer. Biomed. Rep. 1, 499–505. doi:10.3892/br.2013.95
Li, Z., Bao, S., Wu, Q., Wang, H., Eyler, C., Sathornsumetee, S., et al. (2009). Hypoxia-inducible factors regulate tumorigenic capacity of glioma stem cells. Cancer Cell 15, 501–513. doi:10.1016/j.ccr.2009.03.018
Lin, X., Kong, D., and Chen, Z.-S. (2022). Editorial: chemo-radiation-resistance in cancer therapy. Front. Pharmacol. 13, 904063. doi:10.3389/fphar.2022.904063
Liu, J., Erenpreisa, J., and Sikora, E. (2022). Polyploid giant cancer cells: an emerging new field of cancer biology. Seminars Cancer Biol. 81, 1–4. doi:10.1016/j.semcancer.2021.10.006
Liu, L., Ning, X., Sun, L., Zhang, H., Shi, Y., Guo, C., et al. (2008). Hypoxia-inducible factor-1 alpha contributes to hypoxia-induced chemoresistance in gastric cancer. Cancer Sci. 99, 121–128. doi:10.1111/j.1349-7006.2007.00643.x
Liu, L., Taverna, P., Whitacre, C. M., Chatterjee, S., and Gerson, S. L. (1999). Pharmacologic disruption of base excision repair sensitizes mismatch repair-deficient and -proficient colon cancer cells to methylating agents. Clin. Cancer Res. 5, 2908–2917.
Liu, Q., Sun, S., Yu, W., Jiang, J., Zhuo, F., Qiu, G., et al. (2015). Altered expression of long non-coding RNAs during genotoxic stress-induced cell death in human glioma cells. J. Neurooncol 122, 283–292. doi:10.1007/s11060-015-1718-0
Liu, T., Li, Z., Zhang, Q., De Amorim Bernstein, K., Lozano-Calderon, S., Choy, E., et al. (2016). Targeting ABCB1 (MDR1) in multi-drug resistant osteosarcoma cells using the CRISPR-Cas9 system to reverse drug resistance. Oncotarget 7, 83502–83513. doi:10.18632/oncotarget.13148
Liu, Z., Peng, Q., Li, Y., and Gao, Y. (2018). Resveratrol enhances cisplatin-induced apoptosis in human hepatoma cells via glutamine metabolism inhibition. BMB Rep. 51, 474–479. doi:10.5483/BMBRep.2018.51.9.114
Liu, Z.-L., Li, H., Liu, J., Wu, M.-L., Chen, X.-Y., Liu, L.-H., et al. (2017). Inactivated Wnt signaling in resveratrol-treated epidermal squamous cancer cells and its biological implication. Oncol. Lett. 14, 2239–2243. doi:10.3892/ol.2017.6458
Longley, D., and Johnston, P. (2005). Molecular mechanisms of drug resistance. J. Pathol. 205, 275–292. doi:10.1002/path.1706
Longley, D. B., Harkin, D. P., and Johnston, P. G. (2003). 5-Fluorouracil: mechanisms of action and clinical strategies. Nat. Rev. Cancer 3, 330–338. doi:10.1038/nrc1074
Lu, B., Mu, Y., Cao, C., Zeng, F., Schneider, S., Tan, J., et al. (2004). Survivin as a therapeutic target for radiation sensitization in lung cancer. Cancer Res. 64, 2840–2845. doi:10.1158/0008-5472.can-03-3547
Lu, L., Dong, J., Wang, L., Xia, Q., Zhang, D., Kim, H., et al. (2018). Activation of STAT3 and Bcl-2 and reduction of reactive oxygen species (ROS) promote radioresistance in breast cancer and overcome of radioresistance with niclosamide. Oncogene 37, 5292–5304. doi:10.1038/s41388-018-0340-y
Luo, H., Wang, L., Schulte, B. A., Yang, A., Tang, S., and Wang, G. Y. (2013). Resveratrol enhances ionizing radiation-induced premature senescence in lung cancer cells. Int. J. Oncol. 43, 1999–2006. doi:10.3892/ijo.2013.2141
Lyu, F., Shang, S.-Y., Gao, X.-S., Ma, M.-W., Xie, M., Ren, X.-Y., et al. (2023). Uncovering the secrets of prostate cancer’s radiotherapy resistance: advances in mechanism research. Biomedicines 11, 1628. doi:10.3390/biomedicines11061628
Lyu, H., Wang, S., Huang, J., Wang, B., He, Z., and Liu, B. (2018). Survivin-targeting miR-542-3p overcomes HER3 signaling-induced chemoresistance and enhances the antitumor activity of paclitaxel against HER2-overexpressing breast cancer. Cancer Lett. 420, 97–108. doi:10.1016/j.canlet.2018.01.065
Ma, L., Li, W., Wang, R., Nan, Y., Wang, Q., Liu, W., et al. (2015). Resveratrol enhanced anticancer effects of cisplatin on non-small cell lung cancer cell lines by inducing mitochondrial dysfunction and cell apoptosis. Int. J. Oncol. 47, 1460–1468. doi:10.3892/ijo.2015.3124
Machida, H., Matsumoto, Y., Shirai, M., and Kubota, N. (2003). Geldanamycin, an inhibitor of Hsp90, sensitizes human tumour cells to radiation. Int. J. Radiat. Biol. 79, 973–980. doi:10.1080/09553000310001626135
Maji, S., Panda, S., Samal, S. K., Shriwas, O., Rath, R., Pellecchia, M., et al. (2018). “Bcl-2 antiapoptotic family proteins and chemoresistance in cancer,” in Advances in cancer research (Amsterdam, Netherlands: Elsevier), 37–75. doi:10.1016/bs.acr.2017.11.001
Marengo, B., Garbarino, O., Speciale, A., Monteleone, L., Traverso, N., and Domenicotti, C. (2019). MYC expression and metabolic redox changes in cancer cells: a synergy able to induce chemoresistance. Oxidative Med. Cell. Longev. 2019, 7346492–7346499. doi:10.1155/2019/7346492
Mashouri, L., Yousefi, H., Aref, A. R., Ahadi, A. mohammad, Molaei, F., and Alahari, S. K. (2019). Exosomes: composition, biogenesis, and mechanisms in cancer metastasis and drug resistance. Mol. Cancer 18, 75. doi:10.1186/s12943-019-0991-5
McConkey, D. J., and Zhu, K. (2008). Mechanisms of proteasome inhibitor action and resistance in cancer. Drug Resist Updat 11, 164–179. doi:10.1016/j.drup.2008.08.002
McLellan, L. I., and Wolf, C. R. (1999). Glutathione and glutathione-dependent enzymes in cancer drug resistance. Drug Resist Updat 2, 153–164. doi:10.1054/drup.1999.0083
Meister, A. (1991). Glutathione deficiency produced by inhibition of its synthesis, and its reversal; Applications in research and therapy. Pharmacol. Ther. 51, 155–194. doi:10.1016/0163-7258(91)90076-X
Mesri, M., Morales-Ruiz, M., Ackermann, E. J., Bennett, C. F., Pober, J. S., Sessa, W. C., et al. (2001). Suppression of vascular endothelial growth factor-mediated endothelial cell protection by survivin targeting. Am. J. Pathology 158, 1757–1765. doi:10.1016/S0002-9440(10)64131-4
Michaelidesová, A., Konířová, J., Bartůněk, P., and Zíková, M. (2019). Effects of radiation therapy on neural stem cells. Genes 10, 640. doi:10.3390/genes10090640
Milas, L. (2001). Cyclooxygenase-2 (COX-2) enzyme inhibitors as potential enhancers of tumor radioresponse. Semin. Radiat. Oncol. 11, 290–299. doi:10.1053/srao.2001.26018
Milas, L., Mason, K., Hunter, N., Petersen, S., Yamakawa, M., Ang, K., et al. (2000). In vivo enhancement of tumor radioresponse by C225 antiepidermal growth factor receptor antibody. Clin. Cancer Res. 6, 701–708.
Min, Z., Wei, C. L., Ping, G. J., Ying, W., Dan, Z., Jie, K. J., et al. (2019). Resveratrol enhances chemosensitivity of renal cell carcinoma to paclitaxel. Front. Biosci. 24, 1452–1461. doi:10.2741/4790
Minaiyan, G., Shafiee, F., and Akbari, V. (2021). Survivin promoter-driven DFF40 gene expression sensitizes melanoma cancer cells to chemotherapy. Int. J. Toxicol. 40, 380–387. doi:10.1177/10915818211014170
Mini, E., Nobili, S., Caciagli, B., Landini, I., and Mazzei, T. (2006). Cellular pharmacology of gemcitabine. Ann. Oncol. 17, v7–v12. doi:10.1093/annonc/mdj941
Mita, A. C., Mita, M. M., Nawrocki, S. T., and Giles, F. J. (2008). Survivin: key regulator of mitosis and apoptosis and novel target for cancer therapeutics. Clin. Cancer Res. 14, 5000–5005. doi:10.1158/1078-0432.CCR-08-0746
Mitani, T., Ito, Y., Harada, N., Nakano, Y., Inui, H., Ashida, H., et al. (2014). Resveratrol reduces the hypoxia-induced resistance to doxorubicin in breast cancer cells. J. Nutr. Sci. Vitaminol. (Tokyo) 60, 122–128. doi:10.3177/jnsv.60.122
Mittal, K., Donthamsetty, S., Kaur, R., Yang, C., Gupta, M. V., Reid, M. D., et al. (2017). Multinucleated polyploidy drives resistance to Docetaxel chemotherapy in prostate cancer. Br. J. Cancer 116, 1186–1194. doi:10.1038/bjc.2017.78
Mohammad, R. M., Muqbil, I., Lowe, L., Yedjou, C., Hsu, H.-Y., Lin, L.-T., et al. (2015). Broad targeting of resistance to apoptosis in cancer. Seminars Cancer Biol. 35, S78-S103–S103. doi:10.1016/j.semcancer.2015.03.001
Moon, S. H., Shin, K. H., Kim, T. H., Yoon, M., Park, S., Lee, D.-H., et al. (2009). Dosimetric comparison of four different external beam partial breast irradiation techniques: three-dimensional conformal radiotherapy, intensity-modulated radiotherapy, helical tomotherapy, and proton beam therapy. Radiother. Oncol. 90, 66–73. doi:10.1016/j.radonc.2008.09.027
Moreira, H., Szyjka, A., Grzesik, J., Pelc, K., Żuk, M., Kulma, A., et al. (2022). Celastrol and resveratrol modulate SIRT genes expression and exert anticancer activity in colon cancer cells and cancer stem-like cells. Cancers 14, 1372. doi:10.3390/cancers14061372
Morgan, M. J., and Liu, Z. (2011). Crosstalk of reactive oxygen species and NF-κB signaling. Cell Res. 21, 103–115. doi:10.1038/cr.2010.178
Moutabian, H., Majdaeen, M., Ghahramani-Asl, R., Yadollahi, M., Gharepapagh, E., Ataei, G., et al. (2022). A systematic review of the therapeutic effects of resveratrol in combination with 5-fluorouracil during colorectal cancer treatment: with a special focus on the oxidant, apoptotic, and anti-inflammatory activities. Cancer Cell Int. 22, 142. doi:10.1186/s12935-022-02561-7
Najafi, M., Farhood, B., Mortezaee, K., Kharazinejad, E., Majidpoor, J., and Ahadi, R. (2020). Hypoxia in solid tumors: a key promoter of cancer stem cell (CSC) resistance. J. Cancer Res. Clin. Oncol. 146, 19–31. doi:10.1007/s00432-019-03080-1
Nakata, E., Mason, K. A., Hunter, N., Husain, A., Raju, U., Liao, Z., et al. (2004). Potentiation of tumor response to radiation or chemoradiation by selective cyclooxygenase-2 enzyme inhibitors. Int. J. Radiat. Oncol. Biol. Phys. 58, 369–375. doi:10.1016/j.ijrobp.2003.09.061
Nasu, S., Ang, K. K., Fan, Z., and Milas, L. (2001). C225 antiepidermal growth factor receptor antibody enhances tumor radiocurability. Int. J. Radiat. Oncol. Biol. Phys. 51, 474–477. doi:10.1016/s0360-3016(01)01671-6
Nawaz, W., Zhou, Z., Deng, S., Ma, X., Ma, X., Li, C., et al. (2017). Therapeutic versatility of resveratrol derivatives. Nutrients 9, 1188. doi:10.3390/nu9111188
Nestal de Moraes, G., Delbue, D., Silva, K. L., Robaina, M. C., Khongkow, P., Gomes, A. R., et al. (2015). FOXM1 targets XIAP and Survivin to modulate breast cancer survival and chemoresistance. Cell Signal 27, 2496–2505. doi:10.1016/j.cellsig.2015.09.013
Newton, T. R., Patel, N. M., Bhat-Nakshatri, P., Stauss, C. R., Goulet, R. J., and Nakshatri, H. (1999). Negative regulation of transactivation function but not DNA binding of NF-kappaB and AP-1 by IkappaBbeta1 in breast cancer cells. J. Biol. Chem. 274, 18827–18835. doi:10.1074/jbc.274.26.18827
O’Connor, D. S., Schechner, J. S., Adida, C., Mesri, M., Rothermel, A. L., Li, F., et al. (2000). Control of apoptosis during angiogenesis by survivin expression in endothelial cells. Am. J. Pathol. 156, 393–398. doi:10.1016/S0002-9440(10)64742-6
Oh, Y.-C., Kang, O.-H., Choi, J.-G., Chae, H.-S., Lee, Y.-S., Brice, O.-O., et al. (2009). Anti-inflammatory effect of resveratrol by inhibition of IL-8 production in LPS-induced THP-1 cells. Am. J. Chin. Med. 37, 1203–1214. doi:10.1142/S0192415X09007600
Orlowski, R. Z. (2004). Bortezomib in combination with other therapies for the treatment of multiple myeloma. J. Natl. Compr. Canc Netw. 2 (Suppl. 4), S16–S20.
Özeş, A. R., Miller, D. F., Özeş, O. N., Fang, F., Liu, Y., Matei, D., et al. (2016). NF-κB-HOTAIR axis links DNA damage response, chemoresistance and cellular senescence in ovarian cancer. Oncogene 35, 5350–5361. doi:10.1038/onc.2016.75
Öztürk, Y., Günaydın, C., Yalçın, F., Nazıroğlu, M., and Braidy, N. (2019). Resveratrol enhances apoptotic and oxidant effects of paclitaxel through TRPM2 channel activation in DBTRG glioblastoma cells. Oxidative Med. Cell. Longev. 2019, 1–13. doi:10.1155/2019/4619865
Panieri, E., and Santoro, M. M. (2016). ROS homeostasis and metabolism: a dangerous liason in cancer cells. Cell Death Dis. 7, e2253. doi:10.1038/cddis.2016.105
Papież, M. A., and Krzyściak, W. (2021). Biological therapies in the treatment of cancer—update and new directions. IJMS 22, 11694. doi:10.3390/ijms222111694
Park, E.-J., and Pezzuto, J. M. (2015). The pharmacology of resveratrol in animals and humans. Biochimica Biophysica Acta (BBA) - Mol. Basis Dis. 1852, 1071–1113. doi:10.1016/j.bbadis.2015.01.014
Park, M., Yoon, H., Kang, M. C., Kwon, J., and Lee, H. W. (2016). PTK7 regulates radioresistance through nuclear factor-kappa B in esophageal squamous cell carcinoma. Tumor Biol. 37, 14217–14224. doi:10.1007/s13277-016-5288-3
Pelicano, H., Carney, D., and Huang, P. (2004). ROS stress in cancer cells and therapeutic implications. Drug Resist. Updat. 7, 97–110. doi:10.1016/j.drup.2004.01.004
Peña-Blanco, A., and García-Sáez, A. J. (2018). Bax, Bak and beyond — mitochondrial performance in apoptosis. FEBS J. 285, 416–431. doi:10.1111/febs.14186
Perazzoli, G., Prados, J., Ortiz, R., Caba, O., Cabeza, L., Berdasco, M., et al. (2015). Temozolomide resistance in glioblastoma cell lines: implication of MGMT, MMR, P-glycoprotein and CD133 expression. PLoS ONE 10, e0140131. doi:10.1371/journal.pone.0140131
Pérez-Gutiérrez, S., González-Cámpora, R., Amérigo-Navarro, J., Beato-Moreno, A., Sánchez-León, M., Pareja Megía, J. M., et al. (2007). Expression of P-glycoprotein and metallothionein in gastrointestinal stromal tumor and leiomyosarcomas. Clinical implications. Pathol. Oncol. Res. 13, 203–208. doi:10.1007/BF02893500
Petersen, C., Petersen, S., Milas, L., Lang, F. F., and Tofilon, P. J. (2000). Enhancement of intrinsic tumor cell radiosensitivity induced by a selective cyclooxygenase-2 inhibitor. Clin. Cancer Res. 6, 2513–2520.
Piechotta, V., Jakob, T., Langer, P., Monsef, I., Scheid, C., Estcourt, L. J., et al. (2019). Multiple drug combinations of bortezomib, lenalidomide, and thalidomide for first-line treatment in adults with transplant-ineligible multiple myeloma: a network meta-analysis. Cochrane Database Syst. Rev. 2019, CD013487. doi:10.1002/14651858.CD013487
Pienta, K. J., Hammarlund, E. U., Austin, R. H., Axelrod, R., Brown, J. S., and Amend, S. R. (2022). Cancer cells employ an evolutionarily conserved polyploidization program to resist therapy. Seminars Cancer Biol. 81, 145–159. doi:10.1016/j.semcancer.2020.11.016
Priya, L. B., Baskaran, R., Huang, C.-Y., and Padma, V. V. (2017). Neferine ameliorates cardiomyoblast apoptosis induced by doxorubicin: possible role in modulating NADPH oxidase/ROS-mediated NFκB redox signaling cascade. Sci. Rep. 7, 12283. doi:10.1038/s41598-017-12060-9
Qian, L., Mao, L., Mo, W., Wang, R., and Zhang, Y. (2022). Resveratrol enhances the radiosensitivity by inducing DNA damage and antitumor immunity in a glioblastoma rat model under 3 T MRI monitoring. J. Oncol. 2022, 9672773–9672813. doi:10.1155/2022/9672773
Qin, J., Liu, Y., Lu, Y., Liu, M., Li, M., Li, J., et al. (2017). Hypoxia-inducible factor 1 alpha promotes cancer stem cells-like properties in human ovarian cancer cells by upregulating SIRT1 expression. Sci. Rep. 7, 10592. doi:10.1038/s41598-017-09244-8
Qiu, C., Su, W., Shen, N., Qi, X., Wu, X., Wang, K., et al. (2020). MNAT1 promotes proliferation and the chemo-resistance of osteosarcoma cell to cisplatin through regulating PI3K/Akt/mTOR pathway. BMC Cancer 20, 1187. doi:10.1186/s12885-020-07687-3
Quan, F., Pan, C., Ma, Q., Zhang, S., and Yan, L. (2008). Reversal effect of resveratrol on multidrug resistance in KBv200 cell line. Biomed. Pharmacother. 62, 622–629. doi:10.1016/j.biopha.2008.07.089
Quezada, C., Peigñan, L., Segura, R., Riquelme, F., Melo, R., Rojas, Z. D., et al. (2011). Study of resistance to chemotherapy mediated by ABC transporters in biopsies of glioblastoma multiforme. Rev. Med. Chil. 139, 415–424.
Rajendran, P., Abdelsalam, S. A., Renu, K., Veeraraghavan, V., Ben Ammar, R., and Ahmed, E. A. (2022). Polyphenols as potent epigenetics agents for cancer. IJMS 23, 11712. doi:10.3390/ijms231911712
Ranjbar, S., Khonkarn, R., Moreno, A., Baubichon-Cortay, H., Miri, R., Khoshneviszadeh, M., et al. (2019). 5-Oxo-hexahydroquinoline derivatives as modulators of P-gp, MRP1 and BCRP transporters to overcome multidrug resistance in cancer cells. Toxicol. Appl. Pharmacol. 362, 136–149. doi:10.1016/j.taap.2018.10.025
Rankin, E. B., and Giaccia, A. J. (2016). Hypoxic control of metastasis. Science 352, 175–180. doi:10.1126/science.aaf4405
Rashid, A., Liu, C., Sanli, T., Tsiani, E., Singh, G., Bristow, R. G., et al. (2011). Resveratrol enhances prostate cancer cell response to ionizing radiation. Modulation of the AMPK, Akt and mTOR pathways. Radiat. Oncol. 6, 144. doi:10.1186/1748-717X-6-144
Reagan-Shaw, S., Afaq, F., Aziz, M. H., and Ahmad, N. (2004). Modulations of critical cell cycle regulatory events during chemoprevention of ultraviolet B-mediated responses by resveratrol in SKH-1 hairless mouse skin. Oncogene 23, 5151–5160. doi:10.1038/sj.onc.1207666
Real, P. J., Sierra, A., De Juan, A., Segovia, J. C., Lopez-Vega, J. M., and Fernandez-Luna, J. L. (2002). Resistance to chemotherapy via Stat3-dependent overexpression of Bcl-2 in metastatic breast cancer cells. Oncogene 21, 7611–7618. doi:10.1038/sj.onc.1206004
Rebucci, M., and Michiels, C. (2013). Molecular aspects of cancer cell resistance to chemotherapy. Biochem. Pharmacol. 85, 1219–1226. doi:10.1016/j.bcp.2013.02.017
Recalde, M. D., Miguel, C. A., Noya-Riobó, M. V., González, S. L., Villar, M. J., and Coronel, M. F. (2020). Resveratrol exerts anti-oxidant and anti-inflammatory actions and prevents oxaliplatin-induced mechanical and thermal allodynia. Brain Res. 1748, 147079. doi:10.1016/j.brainres.2020.147079
Repossi, G., Das, U. N., and Eynard, A. R. (2020). Molecular basis of the beneficial actions of resveratrol. Archives Med. Res. 51, 105–114. doi:10.1016/j.arcmed.2020.01.010
Rezk, Y. A., Balulad, S. S., Keller, R. S., and Bennett, J. A. (2006). Use of Resveratrol to improve the effectiveness of cisplatin and doxorubicin: study in human gynecologic cancer cell lines and in rodent heart. Am. J. Obstetrics Gynecol. 194, e23–e26. doi:10.1016/j.ajog.2005.11.030
Rivankar, S. (2014). An overview of doxorubicin formulations in cancer therapy. J. Cancer Res. Ther. 10, 853–858. doi:10.4103/0973-1482.139267
Rodríguez Carranza, R. (2015). “Doxorrubicina: antineoplásicos,” in Vademécum académico de Medicamentos (New York, NY: McGraw-Hill Education). Available at: https://accessmedicina.mhmedical.com/content.aspx?bookid=1552§ionid=90369748 (Accessed January 9, 2023).
Rosser, C. J., Tanaka, M., Pisters, L. L., Tanaka, N., Levy, L. B., Hoover, D. C., et al. (2004). Adenoviral-mediated PTEN transgene expression sensitizes Bcl-2-expressing prostate cancer cells to radiation. Cancer Gene Ther. 11, 273–279. doi:10.1038/sj.cgt.7700673
Ruan, K., Song, G., and Ouyang, G. (2009). Role of hypoxia in the hallmarks of human cancer. J. Cell. Biochem. 107, 1053–1062. doi:10.1002/jcb.22214
Sakthivel, K. M., and Hariharan, S. (2017). Regulatory players of DNA damage repair mechanisms: role in Cancer Chemoresistance. Biomed. Pharmacother. 93, 1238–1245. doi:10.1016/j.biopha.2017.07.035
Salvesen, G. S., and Duckett, C. S. (2002). IAP proteins: blocking the road to death’s door. Nat. Rev. Mol. Cell Biol. 3, 401–410. doi:10.1038/nrm830
Sampepajung, E., Hamdani, W., Sampepajung, D., and Prihantono, P. (2021). Overexpression of NF-kB as a predictor of neoadjuvant chemotherapy response in breast cancer. BD 40, S45–S53. doi:10.3233/BD-219007
Santos, A. C., Pereira, I., Pereira-Silva, M., Ferreira, L., Caldas, M., Collado-González, M., et al. (2019). Nanotechnology-based formulations for resveratrol delivery: effects on resveratrol in vivo bioavailability and bioactivity. Colloids Surfaces B Biointerfaces 180, 127–140. doi:10.1016/j.colsurfb.2019.04.030
Sarmento-Ribeiro, A. B., Scorilas, A., Gonçalves, A. C., Efferth, T., and Trougakos, I. P. (2019). The emergence of drug resistance to targeted cancer therapies: clinical evidence. Drug Resist. Updat. 47, 100646. doi:10.1016/j.drup.2019.100646
Schneider, J., Gonzalez-Roces, S., Pollán, M., Lucas, R., Tejerina, A., Martin, M., et al. (2001). Expression of LRP and MDR1 in locally advanced breast cancer predicts axillary node invasion at the time of rescue mastectomy after induction chemotherapy. Breast Cancer Res. 3, 183–191. doi:10.1186/bcr293
Schwab, L. P., Peacock, D. L., Majumdar, D., Ingels, J. F., Jensen, L. C., Smith, K. D., et al. (2012). Hypoxia-inducible factor 1α promotes primary tumor growth and tumor-initiating cell activity in breast cancer. Breast Cancer Res. 14, R6. doi:10.1186/bcr3087
Scott, E., Steward, W. P., Gescher, A. J., and Brown, K. (2012). Resveratrol in human cancer chemoprevention - choosing the ‘right’ dose. Mol. Nutr. Food Res. 56, 7–13. doi:10.1002/mnfr.201100400
Serra, M., Hattinger, C. M., Pasello, M., Casotti, C., Fantoni, L., Riganti, C., et al. (2021). Impact of ABC transporters in osteosarcoma and ewing’s sarcoma: which are involved in chemoresistance and which are not? Cells 10, 2461. doi:10.3390/cells10092461
Shankar, S., Singh, G., and Srivastava, R. K. (2007). Chemoprevention by resveratrol: molecular mechanisms and therapeutic potential. Front. Biosci. 12, 4839–4854. doi:10.2741/2432
Shao, B.-Z., Xu, Z.-Q., Han, B.-Z., Su, D.-F., and Liu, C. (2015). NLRP3 inflammasome and its inhibitors: a review. Front. Pharmacol. 6, 262. doi:10.3389/fphar.2015.00262
Shao, Z., Ma, X., Zhang, Y., Sun, Y., Lv, W., He, K., et al. (2020). CPNE1 predicts poor prognosis and promotes tumorigenesis and radioresistance via the AKT singling pathway in triple-negative breast cancer. Mol. Carcinog. 59, 533–544. doi:10.1002/mc.23177
Sharifi-Rad, J., Quispe, C., Mukazhanova, Z., Knut, E., Turgumbayeva, A., Kipchakbayeva, A., et al. (2021). Resveratrol-based nanoformulations as an emerging therapeutic strategy for cancer. Front. Mol. Biosci. 8, 649395. doi:10.3389/fmolb.2021.649395
Shen, L.-F., Zhao, X., Zhou, S.-H., Lu, Z.-J., Zhao, K., Fan, J., et al. (2017). In vivo evaluation of the effects of simultaneous inhibition of GLUT-1 and HIF-1α by antisense oligodeoxynucleotides on the radiosensitivity of laryngeal carcinoma using micro 18F-FDG PET/CT. Oncotarget 8, 34709–34726. doi:10.18632/oncotarget.16671
Siegel, R. L., Miller, K. D., Fuchs, H. E., and Jemal, A. (2022). Cancer statistics, 2022. CA A Cancer J. Clin. 72, 7–33. doi:10.3322/caac.21708
Song, Z., Shi, Y., Han, Q., and Dai, G. (2018). Endothelial growth factor receptor-targeted and reactive oxygen species-responsive lung cancer therapy by docetaxel and resveratrol encapsulated lipid-polymer hybrid nanoparticles. Biomed. Pharmacother. 105, 18–26. doi:10.1016/j.biopha.2018.05.095
Suit, H., Goldberg, S., Niemierko, A., Ancukiewicz, M., Hall, E., Goitein, M., et al. (2007). Secondary carcinogenesis in patients treated with radiation: a review of data on radiation-induced cancers in human, non-human primate, canine and rodent subjects. Radiat. Res. 167, 12–42. doi:10.1667/RR0527.1
Sullivan, R., Paré, G. C., Frederiksen, L. J., Semenza, G. L., and Graham, C. H. (2008). Hypoxia-induced resistance to anticancer drugs is associated with decreased senescence and requires hypoxia-inducible factor-1 activity. Mol. Cancer Ther. 7, 1961–1973. doi:10.1158/1535-7163.MCT-08-0198
Tanno, S., Yanagawa, N., Habiro, A., Koizumi, K., Nakano, Y., Osanai, M., et al. (2004). Serine/threonine kinase AKT is frequently activated in human bile duct cancer and is associated with increased radioresistance. Cancer Res. 64, 3486–3490. doi:10.1158/0008-5472.CAN-03-1788
Tong, D., Liu, Q., Wang, L., Xie, Q., Pang, J., Huang, Y., et al. (2018). The roles of the COX2/PGE2/EP axis in therapeutic resistance. Cancer Metastasis Rev. 37, 355–368. doi:10.1007/s10555-018-9752-y
Tran, J., Master, Z., Yu, J. L., Rak, J., Dumont, D. J., and Kerbel, R. S. (2002). A role for survivin in chemoresistance of endothelial cells mediated by VEGF. Proc. Natl. Acad. Sci. U. S. A. 99, 4349–4354. doi:10.1073/pnas.072586399
Traverso, N., Ricciarelli, R., Nitti, M., Marengo, B., Furfaro, A. L., Pronzato, M. A., et al. (2013). Role of glutathione in cancer progression and chemoresistance. Oxidative Med. Cell. Longev. 2013, 972913–973010. doi:10.1155/2013/972913
UICC Global Cancer Control (2023). Global cancer observatory. Available at: https://gco.iarc.fr/tomorrow/en/dataviz/isotype.
van Brussel, J. P., and Mickisch, G. H. J. (2003). Multidrug resistance in prostate cancer. Onkologie 26, 175–181. doi:10.1159/000071510
van den Beucken, T., Koch, E., Chu, K., Rupaimoole, R., Prickaerts, P., Adriaens, M., et al. (2014). Hypoxia promotes stem cell phenotypes and poor prognosis through epigenetic regulation of DICER. Nat. Commun. 5, 5203. doi:10.1038/ncomms6203
Varoni, E. M., Lo Faro, A. F., Sharifi-Rad, J., and Iriti, M. (2016). Anticancer molecular mechanisms of resveratrol. Front. Nutr. 3, 8. doi:10.3389/fnut.2016.00008
Vesely, O., Baldovska, S., and Kolesarova, A. (2021). Enhancing bioavailability of nutraceutically used resveratrol and other stilbenoids. Nutrients 13, 3095. doi:10.3390/nu13093095
Vinod, B. S., Maliekal, T. T., and Anto, R. J. (2013). Phytochemicals as chemosensitizers: from molecular mechanism to clinical significance. Antioxidants Redox Signal. 18, 1307–1348. doi:10.1089/ars.2012.4573
Vinod, B. S., Nair, H. H., Vijayakurup, V., Shabna, A., Shah, S., Krishna, A., et al. (2015). Resveratrol chemosensitizes HER-2-overexpressing breast cancer cells to docetaxel chemoresistance by inhibiting docetaxel-mediated activation of HER-2–Akt axis. Cell Death Discov. 1, 15061. doi:10.1038/cddiscovery.2015.61
Wall, N. R., O’Connor, D. S., Plescia, J., Pommier, Y., and Altieri, D. C. (2003). Suppression of survivin phosphorylation on Thr34 by flavopiridol enhances tumor cell apoptosis. Cancer Res. 63, 230–235.
Wang, C., Chen, L., Hou, X., Li, Z., Kabra, N., Ma, Y., et al. (2006). Interactions between E2F1 and SirT1 regulate apoptotic response to DNA damage. Nat. Cell Biol. 8, 1025–1031. doi:10.1038/ncb1468
Wang, D., Guo, X.-Z., Li, H.-Y., Zhao, J.-J., Shao, X.-D., and Wu, C.-Y. (2014). Prognostic significance of cyclooxygenase-2 protein in pancreatic cancer: a meta-analysis. Tumour Biol. 35, 10301–10307. doi:10.1007/s13277-014-2260-y
Wang, H., Jiang, H., Van De Gucht, M., and De Ridder, M. (2019a). Hypoxic radioresistance: can ROS Be the key to overcome it? Cancers 11, 112. doi:10.3390/cancers11010112
Wang, L., Long, L., Wang, W., and Liang, Z. (2015). Resveratrol, a potential radiation sensitizer for glioma stem cells both in vitro and in vivo. J. Pharmacol. Sci. 129, 216–225. doi:10.1016/j.jphs.2015.11.001
Wang, M., Jiang, S., Yu, F., Zhou, L., and Wang, K. (2019b). Noncoding RNAs as molecular targets of resveratrol underlying its anticancer effects. J. Agric. Food Chem. 67, 4709–4719. doi:10.1021/acs.jafc.9b01667
Wang, P., Ren, Z., and Sun, P. (2012). Overexpression of the long non-coding RNA MEG3 impairs in vitro glioma cell proliferation. J. Cell. Biochem. 113, 1868–1874. doi:10.1002/jcb.24055
Wang, P., Yan, Q., Liao, B., Zhao, L., Xiong, S., Wang, J., et al. (2020a). The HIF1α/HIF2α-miR210-3p network regulates glioblastoma cell proliferation, dedifferentiation and chemoresistance through EGF under hypoxic conditions. Cell Death Dis. 11, 992. doi:10.1038/s41419-020-03150-0
Wang, Q., Zhang, J., Zhang, L., Zhao, L., Fan, S., Yang, Z., et al. (2011). Expression of P-gp, MRP, LRP, GST-π and TopoIIα and intrinsic resistance in human lung cancer cell lines. Oncol. Rep. 26, 1081–1089. doi:10.3892/or.2011.1405
Wang, X., Zhang, X., Qiu, C., and Yang, N. (2020b). STAT3 contributes to radioresistance in cancer. Front. Oncol. 10, 1120. doi:10.3389/fonc.2020.01120
Wang, Z. (2017). ErbB receptors and cancer. Methods Mol. Biol. 1652, 3–35. doi:10.1007/978-1-4939-7219-7_1
Was, H., Borkowska, A., Olszewska, A., Klemba, A., Marciniak, M., Synowiec, A., et al. (2022). Polyploidy formation in cancer cells: how a Trojan horse is born. Seminars Cancer Biol. 81, 24–36. doi:10.1016/j.semcancer.2021.03.003
Weaver, B. A. (2014). How Taxol/paclitaxel kills cancer cells. MBoC 25, 2677–2681. doi:10.1091/mbc.e14-04-0916
Weiss, J. F., and Landauer, M. R. (2003). Protection against ionizing radiation by antioxidant nutrients and phytochemicals. Toxicology 189, 1–20. doi:10.1016/S0300-483X(03)00149-5
Woynarowski, J. M., Faivre, S., Herzig, M. C. S., Arnett, B., Chapman, W. G., Trevino, A. V., et al. (2000). Oxaliplatin-induced damage of cellular DNA. Mol. Pharmacol. 58, 920–927. doi:10.1124/mol.58.5.920
Wu, M., Song, D., Li, H., Ahmad, N., Xu, H., Yang, X., et al. (2023). Resveratrol enhances temozolomide efficacy in glioblastoma cells through downregulated MGMT and negative regulators-related STAT3 inactivation. IJMS 24, 9453. doi:10.3390/ijms24119453
Wu, S.-L., Li, Y.-J., Liao, K., Shi, L., Zhang, N., Liu, S., et al. (2017). 2-Methoxyestradiol inhibits the proliferation and migration and reduces the radioresistance of nasopharyngeal carcinoma CNE-2 stem cells via NF-κB/HIF-1 signaling pathway inactivation and EMT reversal. Oncol. Rep. 37, 793–802. doi:10.3892/or.2016.5319
Wu, S.-X., Xiong, R.-G., Huang, S.-Y., Zhou, D.-D., Saimaiti, A., Zhao, C.-N., et al. (2022). Effects and mechanisms of resveratrol for prevention and management of cancers: an updated review. Crit. Rev. Food Sci. Nutr., 1–19. doi:10.1080/10408398.2022.2101428
Xiang, F., Zhu, Z., Zhang, M., Wang, J., Chen, Z., Li, X., et al. (2021). 3,3’-Diindolylmethane enhances paclitaxel sensitivity by suppressing DNMT1-mediated KLF4 methylation in breast cancer. Front. Oncol. 11, 627856. doi:10.3389/fonc.2021.627856
Xiong, L., Lin, X.-M., Nie, J.-H., Ye, H.-S., and Liu, J. (2021). Resveratrol and its nanoparticle suppress doxorubicin/docetaxel-resistant anaplastic thyroid cancer cells in vitro and in vivo. Nanotheranostics 5, 143–154. doi:10.7150/ntno.53844
Xu, J., Liu, D., Niu, H., Zhu, G., Xu, Y., Ye, D., et al. (2017). Resveratrol reverses Doxorubicin resistance by inhibiting epithelial-mesenchymal transition (EMT) through modulating PTEN/Akt signaling pathway in gastric cancer. J. Exp. Clin. Cancer Res. 36, 19. doi:10.1186/s13046-016-0487-8
Yan, Y., Xu, Z., Dai, S., Qian, L., Sun, L., and Gong, Z. (2016). Targeting autophagy to sensitive glioma to temozolomide treatment. J. Exp. Clin. Cancer Res. 35, 23. doi:10.1186/s13046-016-0303-5
Yang, F., Zhao, Z., Cai, S., Ling, L., Hong, L., Tao, L., et al. (2021). Detailed molecular mechanism and potential drugs for COL1A1 in carboplatin-resistant ovarian cancer. Front. Oncol. 10, 576565. doi:10.3389/fonc.2020.576565
Yang, H., Li, X., Zhou, Y., Ban, X., Zeng, T., Li, L., et al. (2015). Stemness and chemotherapeutic drug resistance induced by EIF5A2 overexpression in esophageal squamous cell carcinoma. Oncotarget 6, 26079–26089. doi:10.18632/oncotarget.4581
Yang, H., Wang, J., Bu, X., Yang, B., Wang, B., Hu, S., et al. (2019). Resveratrol restores sensitivity of glioma cells to temozolamide through inhibiting the activation of Wnt signaling pathway. J. Cell. Physiology 234, 6783–6800. doi:10.1002/jcp.27409
Yang, W., Sun, T., Cao, J., and Fan, S. (2011). Hypoxia-inducible factor-1α downregulation by small interfering RNA inhibits proliferation, induces apoptosis, and enhances radiosensitivity in chemical hypoxic human hepatoma SMMC-7721 cells. Cancer Biother Radiopharm. 26, 565–571. doi:10.1089/cbr.2011.0955
Yang, X., Yang, P., Shen, J., Osaka, E., Choy, E., Cote, G., et al. (2014). Prevention of multidrug resistance (MDR) in osteosarcoma by NSC23925. Br. J. Cancer 110, 2896–2904. doi:10.1038/bjc.2014.254
Yang, Y.-P., Chang, Y.-L., Huang, P.-I., Chiou, G.-Y., Tseng, L.-M., Chiou, S.-H., et al. (2012). Resveratrol suppresses tumorigenicity and enhances radiosensitivity in primary glioblastoma tumor initiating cells by inhibiting the STAT3 axis. J. Cell. Physiol. 227, 976–993. doi:10.1002/jcp.22806
Yang, Z., Xie, Q., Chen, Z., Ni, H., Xia, L., Zhao, Q., et al. (2018). Resveratrol suppresses the invasion and migration of human gastric cancer cells via inhibition of MALAT1-mediated epithelial-to-mesenchymal transition. Exp. Ther. Med. 17, 1569–1578. doi:10.3892/etm.2018.7142
Yarden, Y., and Sliwkowski, M. X. (2001). Untangling the ErbB signalling network. Nat. Rev. Mol. Cell Biol. 2, 127–137. doi:10.1038/35052073
You, Y., Wang, Q., Li, H., Ma, Y., Deng, Y., Ye, Z., et al. (2019). Zoledronic acid exhibits radio-sensitizing activity in human pancreatic cancer cells via inactivation of STAT3/NF-κB signaling. Onco Targets Ther. 12, 4323–4330. doi:10.2147/OTT.S202516
Youn, C.-K., Kim, M.-H., Cho, H.-J., Kim, H.-B., Chang, I.-Y., Chung, M.-H., et al. (2004). Oncogenic H-Ras up-regulates expression of ERCC1 to protect cells from platinum-based anticancer agents. Cancer Res. 64, 4849–4857. doi:10.1158/0008-5472.CAN-04-0348
Yuan, Y., Xue, X., Guo, R., Sun, X., and Hu, G. (2012). Resveratrol enhances the antitumor effects of temozolomide in glioblastoma via ROS-dependent AMPK-TSC-mTOR signaling pathway. CNS Neurosci. Ther. 18, 536–546. doi:10.1111/j.1755-5949.2012.00319.x
Zaitceva, V., Kopeina, G. S., and Zhivotovsky, B. (2021). Anastasis: return journey from cell death. Cancers 13, 3671. doi:10.3390/cancers13153671
Zhai, Z., Samson, J. M., Yamauchi, T., Vaddi, P. K., Matsumoto, Y., Dinarello, C. A., et al. (2020). Inflammasome sensor NLRP1 confers acquired drug resistance to temozolomide in human melanoma. Cancers 12, 2518. doi:10.3390/cancers12092518
Zhan, M., and Han, Z. C. (2004). Phosphatidylinositide 3-kinase/AKT in radiation responses. Histol. Histopathol. 19, 915–923. doi:10.14670/HH-19.915
Zhang, L., Lin, Z., Chen, Y., Gao, D., Wang, P., Lin, Y., et al. (2022a). Co-delivery of Docetaxel and Resveratrol by liposomes synergistically boosts antitumor efficiency against prostate cancer. Eur. J. Pharm. Sci. 174, 106199. doi:10.1016/j.ejps.2022.106199
Zhang, P., Song, E., Jiang, M., and Song, Y. (2021a). Celecoxib and Afatinib synergistic enhance radiotherapy sensitivity on human non-small cell lung cancer A549 cells. Int. J. Radiat. Biol. 97, 170–178. doi:10.1080/09553002.2021.1846817
Zhang, W., Jiang, H., Chen, Y., and Ren, F. (2019a). Resveratrol chemosensitizes adriamycin-resistant breast cancer cells by modulating miR-122-5p. J Cell. Biochem. 120, 16283–16292. doi:10.1002/jcb.28910
Zhang, Y., Gan, H., Zhao, F., Ma, X., Xie, X., Huang, R., et al. (2021b). CPEB4-Promoted paclitaxel resistance in ovarian cancer in vitro relies on translational regulation of CSAG2. Front. Pharmacol. 11, 600994. doi:10.3389/fphar.2020.600994
Zhang, Y., Han, C. Y., Duan, F. G., Fan, X.-X., Yao, X.-J., Parks, R. J., et al. (2019b). p53 sensitizes chemoresistant non-small cell lung cancer via elevation of reactive oxygen species and suppression of EGFR/PI3K/AKT signaling. Cancer Cell Int. 19, 188. doi:10.1186/s12935-019-0910-2
Zhang, Z., Ji, Y., Hu, N., Yu, Q., Zhang, X., Li, J., et al. (2022b). Ferroptosis-induced anticancer effect of resveratrol with a biomimetic nano-delivery system in colorectal cancer treatment. Asian J. Pharm. Sci. 17, 751–766. doi:10.1016/j.ajps.2022.07.006
Zhang, Z.-H., Hong, Q., Zhang, Z.-C., Xing, W.-Y., Xu, S., Tian, Q.-X., et al. (2021c). ROS-mediated genotoxic stress is involved in NaAsO2-induced cell cycle arrest, stemness enhancement and chemoresistance of prostate cancer cells in a p53-independent manner. Ecotoxicol. Environ. Saf. 208, 111436. doi:10.1016/j.ecoenv.2020.111436
Zhao, H., Chen, S., Gao, K., Zhou, Z., Wang, C., Shen, Z., et al. (2017). Resveratrol protects against spinal cord injury by activating autophagy and inhibiting apoptosis mediated by the SIRT1/AMPK signaling pathway. Neuroscience 348, 241–251. doi:10.1016/j.neuroscience.2017.02.027
Zhao, W., Bao, P., Qi, H., and You, H. (2010). Resveratrol down-regulates survivin and induces apoptosis in human multidrug-resistant SPC-A-1/CDDP cells. Oncol. Rep. 23, 279–286. doi:10.3892/or_00000634
Zhou, C., Qian, W., Ma, J., Cheng, L., Jiang, Z., Yan, B., et al. (2019). Resveratrol enhances the chemotherapeutic response and reverses the stemness induced by gemcitabine in pancreatic cancer cells via targeting SREBP 1. Cell Prolif. 52, e12514. doi:10.1111/cpr.12514
Zhu, B., Liu, G. T., Zhao, Y. M., Wu, R. S., and Strada, S. J. (2006). Chemosensitizing multiple drug resistance of human carcinoma by bicyclol involves attenuated P-glycoprotein, GST-P and Bcl-2. Cancer Biol. Ther. 5, 536–543. doi:10.4161/cbt.5.5.2655
Zietman, A. L., Bae, K., Slater, J. D., Shipley, W. U., Efstathiou, J. A., Coen, J. J., et al. (2010). Randomized trial comparing conventional-dose with high-dose conformal radiation therapy in early-stage adenocarcinoma of the prostate: long-term results from proton radiation oncology group/american college of radiology 95-09. J. Clin. Oncol. 28, 1106–1111. doi:10.1200/JCO.2009.25.8475
Keywords: radioresistance, chemoresistance, resveratrol, cancer, molecular mechanisms
Citation: Cotino-Nájera S, Herrera LA, Domínguez-Gómez G and Díaz-Chávez J (2023) Molecular mechanisms of resveratrol as chemo and radiosensitizer in cancer. Front. Pharmacol. 14:1287505. doi: 10.3389/fphar.2023.1287505
Received: 01 September 2023; Accepted: 30 October 2023;
Published: 10 November 2023.
Edited by:
Qingbin Cui, University of Toledo College of Medicine and Life Sciences, United StatesReviewed by:
Xiaolin Qian, Southern Research Institute, United StatesCopyright © 2023 Cotino-Nájera, Herrera, Domínguez-Gómez and Díaz-Chávez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Guadalupe Domínguez-Gómez, ZGd1YWRhbHVwZWlzYWJlbEB5YWhvby5jb20=; José Díaz-Chávez, amRpYXpjaGF2ZXowM0BnbWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.