 Ma Ying-Rui1†
Ma Ying-Rui1† Shi Rong
Shi Rong Zhou Qian-Mei
Zhou Qian-Mei- 1Institute of Interdisciplinary Integrative Medicine Research, Shanghai University of Traditional Chinese Medicine, Shanghai, China
- 2Shuguang Hospital Affiliated to Shanghai University of Traditional Chinese Medicine, Shanghai, China
- 3Shanghai Institute of Stem Cell Research and Clinical Translation, Shanghai, China
Breast cancer has a high occurrence rate globally and its treatment has demonstrated clinical efficacy with the use of systemic chemotherapy and immune checkpoint blockade. Insufficient cytotoxic T lymphocyte infiltration and the accumulation of immunosuppressive cells within tumours are the primary factors responsible for the inadequate clinical effectiveness of breast cancer treatment. The stimulator of interferon genes (STING) represents a pivotal protein in the innate immune response. Upon activation, STING triggers the activation and enhancement of innate and adaptive immune functions, resulting in therapeutic benefits for malignant tumours. The STING signalling pathway in breast cancer is influenced by various factors such as deoxyribonucleic acid damage response, tumour immune microenvironment, and mitochondrial function. The use of STING agonists is gaining momentum in breast cancer research. This review provides a comprehensive overview of the cyclic guanosine monophosphate-adenosine monophosphate synthase-STING pathway, its agonists, and the latest findings related to their application in breast cancer.
1 Introduction
Breast cancer is a prevalent malignancy and ranks as the second leading cause of mortality among women, following lung cancer (Sung et al., 2021; Siegel et al., 2022; Xia et al., 2022). It poses a significant risk to women’s health and negatively affects their quality of life. The increasing incidence and mortality rates associated with this disease exert a substantial financial burden on the healthcare industry, highlighting the need for innovative and effective therapeutic interventions. Current treatment modalities for breast cancer include surgery, radiotherapy, chemotherapy, and endocrine therapy, with paclitaxel, platinum, anthracyclines, and capecitabine being the primary chemotherapy agents used (Kerr et al., 2022). Immunotherapy has also emerged as a valuable approach for treating breast cancer, as it yields improved treatment outcomes (Emens, 2018).
The cyclic guanosine monophosphate (GMP)-adenosine monophosphate (AMP) synthase (cGAS) and stimulator of interferon genes (STING) (cGAS-STING) signalling pathway has become a prominent subject of interest in cancer immunotherapy. This pathway detects deoxyribonucleic acid (DNA) in the cytoplasm and stimulates the production of immune factors such as type I interferon (IFN) (IFN-I), initiating a cascade of immune responses with an anti-tumour effect (Deng et al., 2014). Moreover, it enhances the immune response when paclitaxel and platinum-based agents are administered in oncological chemotherapy (Della Corte et al., 2020; Hu Y. et al., 2021; Cao et al., 2022). Particularly, paclitaxel has been closely associated with the activation of the cGAS-STING pathway in breast cancer treatment (Zhu C. et al., 2022; Qiu et al., 2022), suggesting that targeting this pathway offers a novel approach for treating breast cancer. This review focuses on the cGAS-STING pathway and its current implications in DNA repair, the tumour microenvironment (TME), and mitochondrial function in breast cancer. Additionally, it examines existing therapies and emerging targets for treating this disease.
2 The cGAS-STING pathway
Innate immunity serves as the first line of defence against foreign genetic material and plays a crucial role in tumour-induced immune responses. The primary DNA sensor that triggers the innate immune response is cGAS, a protein found throughout the cell that plays crucial roles in anti-tumour immunity, autophagy, cellular senescence, defence against microbial infection, and autoimmune and inflammatory diseases (Zhang et al., 2020; Zhang et al., 2021). Upon DNA recognition, cGAS activates STING protein, which detects cyclic dinucleotides from the endoplasmic reticulum (ER) membrane (Zhu Z. et al., 2022). The cGAS-STING signalling pathway comprises three main phases: double-stranded DNA detection, intracellular signalling, and immune response activation (Yu et al., 2022). Through these stages, the cGAS-STING pathway becomes a central link between immunity and cancer. These details are discussed below.
The DNA sensor cGAS, belonging to the nucleotidyl transferase family, catalyses 2′3′-cyclic GMP-AMP (cGAMP) formation. cGAMP induces conformational changes and oligomerisation of the STING protein. As a result, the activated STING forms tetramers and translocates from the ER to the Golgi apparatus (Dobbs et al., 2015). Subsequently, the activated protein recruits and activates TANK-binding kinase 1 (TBK1) and IFN regulatory factor 3 (IRF3), resulting in the expression of various antiviral genes (Zhang et al., 2020). Among these genes are IFNs-I, which significantly influence the therapeutic efficacy of several anti-cancer drugs, including immunotherapies (Civril et al., 2013; Sun et al., 2013; Zitvogel et al., 2015; Zhou et al., 2020). IFN-α and IFN-β are the most common type I IFNs, promoting the activation and proliferation of cytotoxic T lymphocytes, natural killer (NK) cells, dendritic cells (DCs), and B cells, thereby bridging the innate and acquired immune response (Zhou et al., 2022). The STING protein also activates components of the nuclear factor kappa B (NF-κB) signalling pathway; however, the specific activation mechanism remains unresolved (Burdette et al., 2011; Hopfner and Hornung, 2020). Furthermore, the cGAS-STING signalling pathway induces an anti-tumour immune response by sensing the DNA damage response (DDR) and stimulating the innate immune response within tumours (Jiang et al., 2021).
The STING gene is expressed in various cell types, and studies focused on tumours, it is frequently observed that STING signalling is suppressed. The inhibition is attributed to loss-of-function mutations or epigenetic silencing of the cGAS/STING promoter region (Konno et al., 2018). Apart from transcriptional regulation, different types of modifications could also affect the function of STING. The primary forms of modifications include polyubiquitination and phosphorylation, followed by palmitoylation, sumoylation, oxidation, nitro-alkylation, carbonylation, and disulphide bond formation (Zhang et al., 2022a). Within the cGAS-STING pathway, STING translocation from the ER to the Golgi apparatus is crucial for the activation of the STING signalling pathway. Additionally, it has been reported that the binding of the ER calcium sensor stromal interaction molecule 1 to STING specifically retains STING in the ER, preventing its translocation to the Golgi apparatus. This in turn prevents STING activation and blocks the STING cascade response (Guerini, 2022). Furthermore, the presence of a key factor that terminates STING signalling at the Golgi apparatus, namely, articulin complex 1, facilitates the sorting of phosphorylated STING into lattice-protein-coated transport vesicles for delivery to the endolysosomal system, resulting in its degradation and the termination of STING-dependent immune activation (Liu Y. et al., 2022).
Targeting the cGAS-STING pathway is an emerging therapeutic approach for various cancers due to compelling evidence indicating its activation in the TME elicits potent anti-cancer effects. However, it is crucial to note that the activated cGAS-STING pathway might also exhibit pro-cancer functions under specific conditions. For instance, the cGAS-STING-mediated IFN-I response and the IFN-I-associated senescence could promote tumour initiation by producing various protumourigenic cytokines (Boukhaled et al., 2021). Additionally, the STING protein induces interleukin (IL)-35 production, which activates regulatory B cell functions but simultaneously inhibits NK cell responses (Li et al., 2022). These contrasting roles suggest that the STING pathway operates through diverse mechanisms that require thorough investigation to develop effective targeted therapies.
3 DDR and the cGAS-STING pathway in breast cancer
Genomic instability is a prominent feature that promotes the malignant transformation of cancer (Alhmoud et al., 2020). Defects in DDR and increased replication stress are critical events that promote the clonal evolution of cancer cells by promoting genetic alterations such as gene copy number changes, chromosomal rearrangements, and gene mutations, thereby facilitating tumour progression (Pilié et al., 2019). Notably, DDR deficiency plays a pivotal role in determining tumour immunogenicity, and growing evidence support the notion that DDR-targeted therapy increases anti-tumour immune response (Chabanon et al., 2021). Additionally, cytotoxic drugs that target the DDR pathway are employed as anti-cancer therapies, as this pathway governs many mechanisms underlying tumour cell resistance and sensitivity to cytotoxic radiotherapy (Jiang et al., 2021). For instance, cisplatin, a first-line chemotherapeutic agent for various malignant tumours including breast (Zhu Y. et al., 2022), ovarian (Yang et al., 2022), head and neck (Kwon et al., 2021), lung (Fennell et al., 2016), and bladder (Herr and Soloway, 2022) cancers, exerts its anti-cancer effects by inducing DDR in cancer cells (Dasari and Tchounwou, 2014; Tchounwou et al., 2021).
cGAS activation can occur through two distinct mechanisms: the accumulation of DNA in the cytoplasm and micronuclei or prolonged auto-activation of the DDR signalling (Ragu et al., 2020). For instance, cGAS detects fragmented DNA produced by DDR, leading to changes in cGAMP, activating the cGAS-STING pathway and initiating an immune response (Harding et al., 2017; Reisländer et al., 2020). Consequently, DNA released into the cytoplasm after a DDR event serves as a crucial cGAS trigger, and the absence of cGAS decreases DDR signalling (Banerjee et al., 2021). Blocking DNA replication and repair affects genome integrity and activates the cGAS-STING signalling cascade (Chabanon et al., 2019). Moreover, mutations in DDR-related genes, which impair their function, result in increased expression of genes associated with the cGAS-STING pathway in non-small cell lung cancer (Della Corte et al., 2022). Similarly, pharmacological inhibition of polyadenosine diphosphate-ribose polymerase (PARP) and checkpoint kinase 1 (CHK1) in small-cell lung cancer results in the inhibition of the DDR pathway and activates the cGAS-STING pathway, evoking an anti-tumour immune response (Sen et al., 2019). These findings suggest the need for further investigation into the mechanisms by which DDR activates the cGAS-STING pathway.
Patients with breast cancer often exhibit alterations in DDR genes. For instance, approximately 10.7% of female patients carry deleterious mutations in cancer susceptibility genes, with 6.1% attributed to breast cancer gene (BRCA) 1/2% and 4.6% involving other susceptibility genes such as checkpoint kinase 2, ataxia-telangiectasia mutated (ATM), BRCA1 interacting helicase 1, partner and localiser of BRCA2, phosphatase and tensin homolog, nibrin, RAD51C, RAD51D, mutS homolog 6, and PMS1 homolog 2, mismatch repair system component (Tung et al., 2016). BRCA1 and BRCA2, in particular, play a crucial role in homologous recombination-mediated DNA repair (Krishnan et al., 2021). Germline defects in these genes can contribute to DDR dysfunction in breast cancer, thereby activating the cGAS/STING signalling pathway and eliciting an immune response (Parkes et al., 2017). Therefore, inhibiting DNA repair and promoting DDR progression, which activates the cGAS-STING pathway, present promising avenues for cancer therapy.
DDR-targeted therapies are emerging as promising strategies for treating breast cancer, particularly triple-negative breast cancer, wherein overexpression of DNA repair proteins, such as PARP1 and replication protein A, might alter the sensitivity to chemotherapy and DDR inhibitors (Lee et al., 2020). For instance, IFI16 has demonstrated anti-tumour effects in triple-negative breast cancer by inducing STING-mediated IFN-I production (Ka et al., 2021; Huang et al., 2022). However, the DDR-induced cGAS-STING-mediated IL-6 -signal transducer and activator of transcription 3 pathway in triple-negative breast cancer has been associated with reduced patient survival (Vasiyani et al., 2022). These contrasting findings suggest that the DDR-induced cGAS-STING signalling pathway plays a bidirectional regulatory role in breast cancer, highlighting the importance of investigating the mechanisms that control its directionality in this disease.
Therefore, inhibiting DNA repair and promoting DDR progression, which activates the cGAS-STING pathway, represents a promising avenue for cancer therapy. Other drugs, such as PARP1 inhibitors, have already gained approval for treating breast and ovarian cancers, demonstrating remarkable efficacy (Staniszewska et al., 2022; Tian et al., 2022). Similarly, inhibitors targeting DDR-related genes, such as DNA-dependent protein kinase, catalytic subunit, ATM, ataxia telangiectasia and Rad3-related protein, CHK1, and WEE1, exhibit promising anti-cancer effects (Wengner et al., 2020). Furthermore, paclitaxel activates cGAS by affecting cell mitosis, inducing cGAS-STING pathway-dependent IFN-I responses (Hu Y. et al., 2021).
4 Tumour immune microenvironment and the cGAS-STING pathway in breast cancer
The TME encompasses various components, including the vasculature, extracellular matrix, and non-carcinoma cells, which play crucial roles in tumour initiation, progression, invasion, and metastasis (Bahrami et al., 2018). Significantly, the TME has garnered substantial attention in cancer therapy research due to the potential anti-cancer effects associated with activating the cGAS-STING pathway in the TME (Li and Bakhoum, 2022). In breast cancer, targeting the TME holds great promise and has demonstrated excellent therapeutic outcomes (Mehraj et al., 2021; Zheng et al., 2022). The TME in breast cancer exhibits variable cellular composition and structural characteristics, serving as a central regulator of tumour progression (Danenberg et al., 2022). Immune-activating cells within the TME include tumour-infiltrating lymphocytes, NK cells, and dendritic cells, while immune-suppressing cells comprise T regulatory cells, tumour-associated macrophages, and myeloid-derived suppressor cells. The breast cancer stroma comprises cancer-associated fibroblasts, vascular endothelial cells, and mesenchymal stromal cells (Wilson et al., 2022). Targeting specific cells within the TME, such as eosinophils, tumour-associated macrophages, cancer-associated fibroblasts, tumour-infiltrating lymphocytes, and regulatory CD4+/CD8+ T cells, can enhance anti-tumour immunity in breast cancer (Li Y. et al., 2020; Li D. et al., 2020; Wu et al., 2020; Grisaru-Tal et al., 2021; Soongsathitanon et al., 2021; Sun et al., 2021). Targeting tumour-associated macrophages has demonstrated significant alleviation of chemotherapy resistance in breast cancer.
Evidence suggests that elevated levels of tumour-infiltrating lymphocytes within the TME play a crucial role in treating breast cancer (Ahn et al., 2020). Differential analysis of tumour compartments has revealed that patients with triple-negative breast cancer responsive to chemotherapy exhibit high STING protein levels (Kulasinghe et al., 2021), indicating its presence in the TME of breast cancer and its potential as a treatment target. While chimeric antigen receptor T (CAR-T) cells are a type of cell treatments for treating haematological malignancies, their effectiveness against solid tumours is limited (Tchou et al., 2017). However, when the cGAS-STING pathway is activated within the breast cancer TME, T helper/IL-17-producing CD8+ T -generated CAR-T cells show increased persistence in the TME and enhanced tumour control (Xu et al., 2021). The STING protein induces the production of IFN-β by intra-tumoural DCs, which initiates and recruits T cells into the TME (Foote et al., 2017). Nanoparticles loaded with STING agonists activate the cGAS-STING signalling pathway within the TME, resulting in IFN-β production and the activation of antigen-presenting cells, thereby stimulating the activation of tumour-reactive cytotoxic T cells (Covarrubias et al., 2022). Consequently, STING agonists hold significant promise as a therapy for reshaping the immunosuppressive TME (Wehbe et al., 2021), as they can reverse its immunosuppressive nature and sensitise breast cancer to immunotherapy (Chen et al., 2020; Zhang L. et al., 2022; Shen et al., 2022).
5 Mitochondrial function and the cGAS-STING pathway in breast cancer
Mitochondrial and nuclear DNA leaking into the cytoplasm activate the cGAS-STING signalling in addition to foreign DNA (Hopfner and Hornung, 2020). Due to its location in the ER, particularly in the ER-mitochondria-associated membrane, STING protein has an inherent advantage in detecting mitochondrial stress responses (Smith, 2020). Mitochondria serve as bioenergetic, biosynthetic, and signalling organelles with crucial roles in regulating innate and adaptive immunity (Weinberg et al., 2015), particularly in processes that lead to apoptosis. In mitochondria-mediated apoptosis, activating pro-apoptotic proteins B-cell leukaemia/lymphoma 2 protein (Bcl-2) antagonist killer 1 (BAK)/Bcl-2-associated X protein (BAX) results in mitochondrial outer membrane permeabilisation, thereby inducing caspase activation and cell death (Lohard et al., 2020). Since mitochondria possess their DNA, BAK/BAX-mediated mitochondrial damage triggers the release of mitochondrial DNA (mtDNA). Consequently, the cGAS-STING-mediated cytoplasmic DNA sensing pathway identifies mtDNA and initiates apoptosis (White et al., 2014; McArthur et al., 2018; Chang S. et al., 2022). Mitochondrial inner membrane permeabilisation enables the release of mtDNA into the cytoplasm and activates the cGAS-STING signalling pathway (Riley et al., 2018).
Targeting the cGAS-STING-associated mitochondrial apoptotic pathway is emerging as a novel therapeutic approach for breast cancer. For instance, cyclic di-AMP (c-di-AMP), an analogue of cGAMP, activates the cGAS-STING pathway and induces mitochondria-mediated apoptosis in oestrogen receptor-negative breast cancer cells (Vasiyani et al., 2021). Eribulin, a microtubule-targeting agent, promotes cGAS-STING signalling expression in triple-negative breast cancer cells by facilitating the cytoplasmic accumulation of mtDNA, IFN-β production, and downstream interferon-stimulated genes (Fermaintt et al., 2021). ATM inhibition enhances the effectiveness of immune checkpoint blockade treatment in breast cancer by facilitating the cytoplasmic leakage of mtDNA and the activation of the cGAS-STING pathway (Hu M. et al., 2021).
Mitochondrial reactive oxygen species (ROS) are a vital source of endogenous ROS. In malignant cells, mitochondria exhibit ROS overproduction, which could promote cancer development by altering gene expression and participating in signalling pathways (Yang et al., 2016; Zhao et al., 2016). Thus, ROS has emerged as a target for anti-tumour therapy. In colorectal cancer, SUMO-specific proteinase 3 detects oxidative stress and promotes the STING-mediated DC-initiated anti-tumour immune response (Hu Z. et al., 2021). Mitochondrial lon, a chaperone protein, induces ROS, which could lead to mtDNA damage, activate IFN signalling via the cGAS-STING-TBK1 axis, and promote programmed death ligand 1 -mediated immune escape (Cheng et al., 2020). Sinularin differentially upregulates ROS and causes oxidative DNA damage in breast cancer cells, potentially activating the cGAS-STING pathway (Huang et al., 2018). STING protein could also act as an upstream regulator of ROS and influence the transcriptional program of ROS metabolism (Hayman et al., 2021). However, the understanding of the relationship between ROS and the cGAS-STING pathway in breast cancer is limited, and further investigation is necessary to uncover new treatment options.
6 Anti-cancer effects of STING agonists
Given the significant potential of the cGAS-STING signalling pathway in anti-tumour therapy, the development of STING agonists has received considerable attention. One prominent drug used as a STING agonist in preclinical studies is 5, 6-dimethylxanthenone-4-acetic acid (DMXAA, ASA404, Vadimezan), a flavonoid compound. Initially employed as a tumour vascular disruptor for anti-cancer treatment, DMXAA was later found to activate the cGAS-STING signalling in mouse models (Baguley, 2003; Daei Farshchi Adli et al., 2018). In combination with specific cancer treatment drugs, such as paclitaxel, it has demonstrated favourable efficacy in patients with intermediate to advanced non-small-cell lung cancer (McKeage et al., 2009). However, when combined with others, such as platinum-based drugs, it has demonstrated negligible effect on the outcomes of patients with triple-negative breast cancer (Lara et al., 2011). Despite its considerable potential in mouse models, DMXAA has proven unsuccessful in human clinical studies, possibly due to its inability to induce the STING signalling pathway in humans (Conlon et al., 2013). Indeed, molecular dynamics simulations revealed that dynamic structural differences between human and mouse STING proteins cause differential sensitivity to DMXAA (Shih et al., 2018). While DMXAA has demonstrated promising performance in mouse tumour models, it has laid a foundation for synthesising new derivatives that hold greater promise for cancer treatment (Hou et al., 2020; Gobbi et al., 2021).
Cyclic dinucleotides, including cGAMP and bacterial messengers c-di-AMP and cyclic di-GMP (c-di-GMP), represent valuable STING agonists (Wang et al., 2020). These compounds serve as natural ligands for the STING protein and play a crucial role in activating STING protein after cGAS-mediated cytoplasmic DNA recognition (Li and Chen, 2018). They exhibit high potential in cancer therapy. For instance, c-di-AMP induces breast cancer cell apoptosis via the cGAS-STING pathway activation and regulation (Vasiyani et al., 2021). Moreover, in a mouse model of bladder cancer, bacillus Calmette–Guérin overexpressing c-di-AMP improves anti-tumour effects through a STING-dependent pathway (Singh et al., 2022). Nanoparticles co-synthesised with c-di-AMP and the immunomodulatory trace element manganese significantly improved the therapeutic efficacy of STING-mediated combined radioimmunotherapy (Wang et al., 2022). Furthermore, c-di-GMP-activated STING demonstrates promising immunotherapeutic efficacy in breast cancer (Chandra et al., 2014), whereas c-di-GMP-loaded peptide nanotubes enhance immunotherapy for melanoma (Zhang et al., 2022c). As an endogenous member of the cGAS-STING signalling pathway, cGAMP exhibits significant potential as a STING agonist in anti-tumour therapy, as CAR-T cells generated using cGAMP display enhanced anti-tumour capacity in breast cancer (Xu et al., 2021; Su et al., 2022). Similarly, cGAMP enhances the anti-tumour activity of CAR-NK cells in pancreatic cancer (Da et al., 2022). These findings suggest that the effects observed with these natural agonists are dependent on the cGAS-STING pathway activation.
Clinical drug development of synthetic cyclic dinucleotides as STING agonists is underway. For instance, ADU-S100 (MIW815) activates the cGAS-STING pathway and demonstrates good tolerability in patients with advanced/metastatic solid tumours or lymphomas (Meric-Bernstam et al., 2022a). Combined treatment with ADU-S100 and the programmed cell death protein 1 (PD-1) inhibitor spartalizumab also shows a favourable safety profile in patients with advanced/metastatic solid tumours or lymphoma (Meric-Bernstam et al., 2022b). Additionally, ADU-S100 in combination with PD-1/cyclooxygenase-2 blockade suppresses peritoneal dissemination of colon cancer and elicits durable tumour immunity in colon cancer (Lee et al., 2021). Another synthetic STING agonist, MK-1454, demonstrates potent anti-tumour activity in pre-clinical trials and is currently in clinical development, showing encouraging efficacy (Harrington et al., 2018; Chang W. et al., 2022). Several other synthetic agonists, including SB11285, BMS-986301, MK-2118, GSK3745417, E7766, SNX281, SYNB 1891, TAK-676, and BI-STING, are undergoing clinical trials and studies (Challa et al., 2017; Amouzegar et al., 2021). Furthermore, several synthetic agonists such as ML-RR-S2-cGAMP, ML-RR-S2-CDG, 3′3′-cyclic AIMP, GSK532, and JNJ-4412, although not yet in the clinical research stage, hold promise for the field of cancer treatment (Corrales et al., 2015; Thomsen et al., 2020; Amouzegar et al., 2021).
In addition to the aforementioned STING agonists, there is a subset of drugs that could be used for cancer therapy by activating the cGAS-STING pathway. For instance, E7766 is a macrocyclic bridging STING agonist with high anti-tumour activity in a mouse model of liver metastases and is also considered a clinical candidate (Kim et al., 2021). Moreover, MSA-2 is a compound that binds human and mouse STING proteins, and when combined with anti-PD-1 antibodies, it inhibits tumour growth and improves survival rates (Pan et al., 2020). A novel STING agonist, MSA-1, activates STING proteins in humans and mice and can be combined with PD-1-binding inhibitors to improve anti-PD-1 resistance (Perera et al., 2022). Several other drugs, including SR-717, amidobenzimidazole, STACT-TREX1, and MV-626, are currently under investigation as STING agonists (Chin et al., 2020; Jiang et al., 2020; Liu X. et al., 2022).
7 Conclusion
Breast cancer poses a significant threat to women’s lives; however, the activation of the cGAS-STING pathway, which triggers an immune response, offers promising prospects for its treatment. Within the cytoplasm, the cGAS protein detects DNA and activates the STING protein, resulting in the activation of TBK1, IRF3, IFN-I, and NF-κB to produce a series of immune responses. STING activation is closely associated with DDR, and breast cancer often exhibits DDR-related gene alterations, particularly in BRCA1 or BRCA2 genes. Targeted DDR therapy has emerged as a potential therapeutic approach for breast cancer, with PARP-1 inhibitors serving as an approved example. The TME controls tumour progression, and breast cancer TME exhibits distinct characteristics. The presence of the STING protein in the breast cancer TME has been observed, influencing tumour progression. Consequently, activating the protein within the TME presents an opportunity to reshape its immunosuppressive nature. Mitochondria, which possess their DNA, are essential organelles that can activate the cGAS-STING pathway under certain conditions. Targeting the cGAS-STING-associated mitochondrial apoptotic pathway and mitochondrial ROS provides a novel avenue for breast cancer treatment. Given the promising potential of the activated cGAS-STING signalling pathway in anti-tumour therapy, various STING agonists are being developed as anti-cancer drugs. Examples include DMXAA, c-di-AMP, c-di-GMP, cGAMP, ADU-S100, MK-1454, SB11285, BMS-986301, E7766, MSA-1, and MSA-2 (Figure 1).
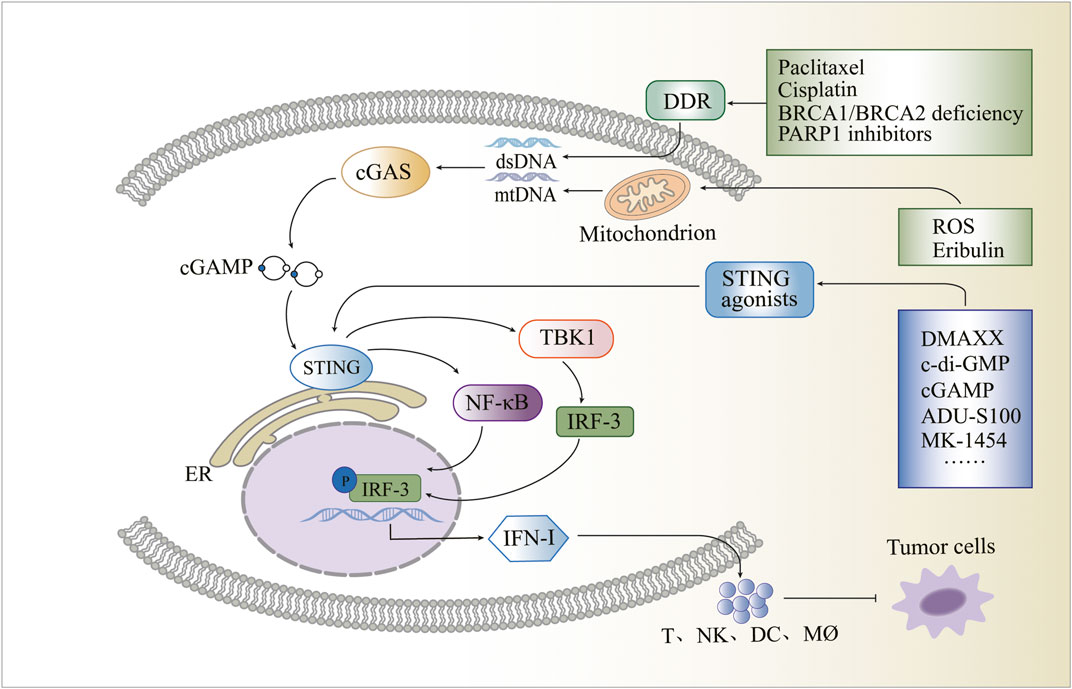
FIGURE 1. Schematic overview of the cyclic guanosine monophosphate-adenosine monophosphate synthase and stimulator of interferon genes pathway and its influencing factors.
The activation of the STING signalling pathway serves as an innate immune sensing mechanism that results in IFN-I production within the TME. This stimulation activates immune cells within the TME, initiating an anti-tumour immune response. Consequently, STING agonists hold great promise as immunotherapeutic drugs. However, STING agonists do not exhibit inhibitory effects on all types of tumours. Currently, the STING signalling pathway is over-activated in tumours with low antigenicity, tumours that release a significant amount of DNA from the cytoplasm due to exposure to potent carcinogens, and tumours with chromosomal instability (CIN) phenotype, which might promote tumour growth and metastasis. Therefore, when considering the use of STING agonists in clinical settings, it is crucial to take into account factors such as the tumour type, antigenicity, and inflammatory microenvironment. It is necessary to understand the CIN status of the tumour and the STING basic activation level.
Understanding the molecular mechanism of STING agonists, as well as identifying and screening tumour-predictive biomarkers suitable for predicting the response to STING agonists, are critical aspects in elucidating the therapeutic potential of these agents. Additionally, selecting appropriate tumour types and optimising treatment dosages are essential for enhancing the efficacy of STING agonists while minimising adverse reactions. Therefore, the development of clinical applications for STING agonists holds great promise. The side effects and indications of STING agonists need to be further confirmed to ensure the safety and efficacy of the treatment. This can be achieved through the development of novel drug delivery systems, which can be combined with other anti-tumour therapies, such as radiotherapy, chemotherapy, targeted therapy, and immunotherapy. While there might be challenges in the research and development of drugs targeting the cGAS-STING pathway, further research on the molecular mechanisms underlying the upstream and downstream pathways and the development of drug delivery systems will pave the way for new targets for anti-tumour research.
Research into the cGAS-STING pathway has undoubtedly expanded the potential for cancer treatment. While the effects of STING activation in combating cancer might vary, its ability to stimulate the immune response holds significant promise for novel therapeutic interventions. Therefore, further research is warranted to fully comprehend the diverse effects of STING activation in cancer. Particularly, the encouraging outcomes observed in using the cGAS-STING pathway for breast cancer treatment highlight its potential for further advancements. Consequently, targeting the STING protein represents a viable approach to potentially enhance future treatment outcomes in this context. This article summarized the regulation of STING and the influencing factors of cGAS-STING pathway. It further enriched the molecular mechanisms of breast cancer. It can provide some research ideas for the follow-up research on STING, and also provide experimental basis for clinical treatment of breast cancer.
Author contributions
MY-R wrote the manuscript. BB-F organized and summarized the literature. LD searched the literature. SR and ZQ-M made comments on the article and revised the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was financially supported by Shanghai Traditional Chinese Medicine High-level Talents Leading Program Project, Science and Technology Commission of Shanghai Municipality Project (22ZR1446900 and 21ZR1447800) and Peak Disciplines (Type IV) of Institutions of Higher Learning in Shanghai.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ahn, S., Chung, Y. R., Seo, A. N., Kim, M., Woo, J. W., and Park, S. Y. (2020). Changes and prognostic values of tumor-infiltrating lymphocyte subsets after primary systemic therapy in breast cancer. PLoS One 15, e0233037. doi:10.1371/journal.pone.0233037
Alhmoud, J. F., Woolley, J. F., Al Moustafa, A. E., and Malki, M. I. (2020). DNA damage/repair management in cancers. Cancers (Basel) 12, 1050. doi:10.3390/cancers12041050
Amouzegar, A., Chelvanambi, M., Filderman, J. N., Storkus, W. J., and Luke, J. J. (2021). STING agonists as cancer therapeutics. Cancers (Basel) 13, 2695. doi:10.3390/cancers13112695
Baguley, B. C. (2003). Antivascular therapy of cancer: Dmxaa. Lancet Oncol. 4, 141–148. doi:10.1016/s1470-2045(03)01018-0
Bahrami, A., Hassanian, S. M., Khazaei, M., Hasanzadeh, M., Shahidsales, S., Maftouh, M., et al. (2018). The therapeutic potential of targeting tumor microenvironment in breast cancer: Rational strategies and recent progress. J. Cell Biochem. 119, 111–122. doi:10.1002/jcb.26183
Banerjee, D., Langberg, K., Abbas, S., Odermatt, E., Yerramothu, P., Volaric, M., et al. (2021). A non-canonical, interferon-independent signaling activity of cGAMP triggers DNA damage response signaling. Nat. Commun. 12, 6207. doi:10.1038/s41467-021-26240-9
Boukhaled, G. M., Harding, S., and Brooks, D. G. (2021). Opposing roles of type I interferons in cancer immunity. Annu. Rev. Pathol. 16, 167–198. doi:10.1146/annurev-pathol-031920-093932
Burdette, D. L., Monroe, K. M., Sotelo-Troha, K., Iwig, J. S., Eckert, B., Hyodo, M., et al. (2011). STING is a direct innate immune sensor of cyclic di-GMP. Nature 478, 515–518. doi:10.1038/nature10429
Cao, L., Tian, H., Fang, M., Xu, Z., Tang, D., Chen, J., et al. (2022). Activating cGAS-STING pathway with ROS-responsive nanoparticles delivering a hybrid prodrug for enhanced chemo-immunotherapy. Biomaterials 290, 121856. doi:10.1016/j.biomaterials.2022.121856
Chabanon, R. M., Muirhead, G., Krastev, D. B., Adam, J., Morel, D., Garrido, M., et al. (2019). PARP inhibition enhances tumor cell-intrinsic immunity in ERCC1-deficient non-small cell lung cancer. J. Clin. Invest. 129, 1211–1228. doi:10.1172/jci123319
Chabanon, R. M., Rouanne, M., Lord, C. J., Soria, J. C., Pasero, P., and Postel-Vinay, S. (2021). Targeting the DNA damage response in immuno-oncology: Developments and opportunities. Nat. Rev. Cancer 21, 701–717. doi:10.1038/s41568-021-00386-6
Challa, S. V., Zhou, S., Sheri, A., Padmanabhan, S., Meher, G., Gimi, R., et al. (2017). Preclinical studies of SB 11285, a novel STING agonist for immuno-oncology. J. Clin. Oncol. 35, e14616. doi:10.1200/JCO.2017.35.15_suppl.e14616
Chandra, D., Quispe-Tintaya, W., Jahangir, A., Asafu-Adjei, D., Ramos, I., Sintim, H. O., et al. (2014). STING ligand c-di-GMP improves cancer vaccination against metastatic breast cancer. Cancer Immunol. Res. 2, 901–910. doi:10.1158/2326-6066.Cir-13-0123
Chang, S., Singh, L., Thaker, K., Abedi, S., Singh, M. K., Patel, T. H., et al. (2022a). Altered retrograde signaling patterns in breast cancer cells cybrids with H and J mitochondrial DNA haplogroups. Int. J. Mol. Sci. 23, 6687. doi:10.3390/ijms23126687
Chang, W., Altman, M. D., Lesburg, C. A., Perera, S. A., Piesvaux, J. A., Schroeder, G. K., et al. (2022b). Discovery of MK-1454: A potent cyclic dinucleotide stimulator of interferon genes agonist for the treatment of cancer. J. Med. Chem. 65, 5675–5689. doi:10.1021/acs.jmedchem.1c02197
Chen, Y. P., Xu, L., Tang, T. W., Chen, C. H., Zheng, Q. H., Liu, T. P., et al. (2020). STING activator c-di-GMP-Loaded mesoporous silica nanoparticles enhance immunotherapy against breast cancer. ACS Appl. Mater Interfaces 12, 56741–56752. doi:10.1021/acsami.0c16728
Cheng, A. N., Cheng, L. C., Kuo, C. L., Lo, Y. K., Chou, H. Y., Chen, C. H., et al. (2020). Mitochondrial Lon-induced mtDNA leakage contributes to PD-L1-mediated immunoescape via STING-IFN signaling and extracellular vesicles. J. Immunother. Cancer 8, e001372. doi:10.1136/jitc-2020-001372
Chin, E. N., Yu, C., Vartabedian, V. F., Jia, Y., Kumar, M., Gamo, A. M., et al. (2020). Antitumor activity of a systemic STING-activating non-nucleotide cGAMP mimetic. Science 369, 993–999. doi:10.1126/science.abb4255
Civril, F., Deimling, T., de Oliveira Mann, C. C., Ablasser, A., Moldt, M., Witte, G., et al. (2013). Structural mechanism of cytosolic DNA sensing by cGAS. Nature 498, 332–337. doi:10.1038/nature12305
Conlon, J., Burdette, D. L., Sharma, S., Bhat, N., Thompson, M., Jiang, Z., et al. (2013). Mouse, but not human STING, binds and signals in response to the vascular disrupting agent 5,6-dimethylxanthenone-4-acetic acid. J. Immunol. 190, 5216–5225. doi:10.4049/jimmunol.1300097
Corrales, L., Glickman, L. H., McWhirter, S. M., Kanne, D. B., Sivick, K. E., Katibah, G. E., et al. (2015). Direct activation of STING in the tumor microenvironment leads to potent and systemic tumor regression and immunity. Cell Rep. 11, 1018–1030. doi:10.1016/j.celrep.2015.04.031
Covarrubias, G., Moon, T. J., Loutrianakis, G., Sims, H. M., Umapathy, M. P., Lorkowski, M. E., et al. (2022). Comparison of the uptake of untargeted and targeted immunostimulatory nanoparticles by immune cells in the microenvironment of metastatic breast cancer. J. Mater Chem. B 10, 224–235. doi:10.1039/d1tb02256c
Da, Y., Liu, Y., Hu, Y., Liu, W., Ma, J., Lu, N., et al. (2022). STING agonist cGAMP enhances anti-tumor activity of CAR-NK cells against pancreatic cancer. Oncoimmunology 11, 2054105. doi:10.1080/2162402x.2022.2054105
Daei Farshchi Adli, A., Jahanban-Esfahlan, R., Seidi, K., Samandari-Rad, S., and Zarghami, N. (2018). An overview on Vadimezan (DMXAA): The vascular disrupting agent. Chem. Biol. Drug Des. 91, 996–1006. doi:10.1111/cbdd.13166
Danenberg, E., Bardwell, H., Zanotelli, V. R. T., Provenzano, E., Chin, S. F., Rueda, O. M., et al. (2022). Breast tumor microenvironment structures are associated with genomic features and clinical outcome. Nat. Genet. 54, 660–669. doi:10.1038/s41588-022-01041-y
Dasari, S., and Tchounwou, P. B. (2014). Cisplatin in cancer therapy: Molecular mechanisms of action. Eur. J. Pharmacol. 740, 364–378. doi:10.1016/j.ejphar.2014.07.025
Della Corte, C. M., Fasano, M., Ciaramella, V., Cimmino, F., Cardnell, R., Gay, C. M., et al. (2022). Anti-tumor activity of cetuximab plus avelumab in non-small cell lung cancer patients involves innate immunity activation: Findings from the CAVE-lung trial. J. Exp. Clin. Cancer Res. 41, 109. doi:10.1186/s13046-022-02332-2
Della Corte, C. M., Sen, T., Gay, C. M., Ramkumar, K., Diao, L., Cardnell, R. J., et al. (2020). STING pathway expression identifies NSCLC with an immune-responsive phenotype. J. Thorac. Oncol. 15, 777–791. doi:10.1016/j.jtho.2020.01.009
Deng, L., Liang, H., Xu, M., Yang, X., Burnette, B., Arina, A., et al. (2014). STING-dependent cytosolic DNA sensing promotes radiation-induced type I interferon-dependent antitumor immunity in immunogenic tumors. Immunity 41, 843–852. doi:10.1016/j.immuni.2014.10.019
Dobbs, N., Burnaevskiy, N., Chen, D., Gonugunta, V. K., Alto, N. M., and Yan, N. (2015). STING activation by translocation from the ER is associated with infection and autoinflammatory disease. Cell Host Microbe 18, 157–168. doi:10.1016/j.chom.2015.07.001
Kim, D. S., Endo, A., Fang, F. G., Huang, K. C., Bao, X., Choi, H. W., et al. (2021). E7766, a macrocycle-bridged stimulator of interferon genes (STING) agonist with potent pan-genotypic activity, ChemMedChem 16, 1740–1743. doi:10.1002/cmdc.202100068
Emens, L. A. (2018). Breast cancer immunotherapy: Facts and hopes. Clin. Cancer Res. 24, 511–520. doi:10.1158/1078-0432.Ccr-16-3001
Fennell, D. A., Summers, Y., Cadranel, J., Benepal, T., Christoph, D. C., Lal, R., et al. (2016). Cisplatin in the modern era: The backbone of first-line chemotherapy for non-small cell lung cancer. Cancer Treat. Rev. 44, 42–50. doi:10.1016/j.ctrv.2016.01.003
Fermaintt, C. S., Takahashi-Ruiz, L., Liang, H., Mooberry, S. L., and Risinger, A. L. (2021). Eribulin activates the cGAS-STING pathway via the cytoplasmic accumulation of mitochondrial DNA. Mol. Pharmacol. 100, 309–318. doi:10.1124/molpharm.121.000297
Foote, J. B., Kok, M., Leatherman, J. M., Armstrong, T. D., Marcinkowski, B. C., Ojalvo, L. S., et al. (2017). A STING agonist given with OX40 receptor and PD-L1 modulators primes immunity and reduces tumor growth in tolerized mice. Cancer Immunol. Res. 5, 468–479. doi:10.1158/2326-6066.Cir-16-0284
Gobbi, S., Belluti, F., Rampa, A., and Bisi, A. (2021). Flavonoid-inspired vascular disrupting agents: Exploring flavone-8-acetic acid and derivatives in the new century. Molecules 26, 4228. doi:10.3390/molecules26144228
Grisaru-Tal, S., Dulberg, S., Beck, L., Zhang, C., Itan, M., Hediyeh-Zadeh, S., et al. (2021). Metastasis-entrained eosinophils enhance lymphocyte-mediated antitumor immunity. Cancer Res. 81, 5555–5571. doi:10.1158/0008-5472.Can-21-0839
Guerini, D. (2022). STING agonists/antagonists: Their potential as therapeutics and future developments. Cells 11, 1159. doi:10.3390/cells11071159
Harding, S. M., Benci, J. L., Irianto, J., Discher, D. E., Minn, A. J., and Greenberg, R. A. (2017). Mitotic progression following DNA damage enables pattern recognition within micronuclei. Nature 548, 466–470. doi:10.1038/nature23470
Harrington, K. J., Brody, J., Ingham, M., Strauss, J., Cemerski, S., Wang, M., et al. (2018). Preliminary results of the first-in-human (FIH) study of MK-1454, an agonist of stimulator of interferon genes (STING), as monotherapy or in combination with pembrolizumab (pembro) in patients with advanced solid tumors or lymphomas. Ann. Oncol. 29, viii712. doi:10.1093/annonc/mdy424.015
Hayman, T. J., Baro, M., MacNeil, T., Phoomak, C., Aung, T. N., Cui, W., et al. (2021). STING enhances cell death through regulation of reactive oxygen species and DNA damage. Nat. Commun. 12, 2327. doi:10.1038/s41467-021-22572-8
Herr, H., and Soloway, M. (2022). Cisplatin, neoadjuvant chemotherapy and bladder cancer. Urology 159, 2–5. doi:10.1016/j.urology.2021.10.017
Hopfner, K. P., and Hornung, V. (2020). Molecular mechanisms and cellular functions of cGAS-STING signalling. Nat. Rev. Mol. Cell Biol. 21, 501–521. doi:10.1038/s41580-020-0244-x
Hou, S., Lan, X. J., Li, W., Yan, X. L., Chang, J. J., Yang, X. H., et al. (2020). Design, synthesis and biological evaluation of acridone analogues as novel STING receptor agonists. Bioorg Chem. 95, 103556. doi:10.1016/j.bioorg.2019.103556
Hu, M., Zhou, M., Bao, X., Pan, D., Jiao, M., Liu, X., et al. (2021b). ATM inhibition enhances cancer immunotherapy by promoting mtDNA leakage and cGAS/STING activation. J. Clin. Invest. 131, e139333. doi:10.1172/jci139333
Hu, Y., Manasrah, B. K., McGregor, S. M., Lera, R. F., Norman, R. X., Tucker, J. B., et al. (2021a). Paclitaxel induces micronucleation and activates pro-inflammatory cGAS-STING signaling in triple-negative breast cancer. Mol. Cancer Ther. 20, 2553–2567. doi:10.1158/1535-7163.Mct-21-0195
Hu, Z., Teng, X. L., Zhang, T., Yu, X., Ding, R., Yi, J., et al. (2021c). SENP3 senses oxidative stress to facilitate STING-dependent dendritic cell antitumor function. Mol. Cell 81, 940–952.e5. doi:10.1016/j.molcel.2020.12.024
Huang, H. W., Tang, J. Y., Ou-Yang, F., Wang, H. R., Guan, P. Y., Huang, C. Y., et al. (2018). Sinularin selectively kills breast cancer cells showing G2/M arrest, apoptosis, and oxidative DNA damage. Molecules 23, 849. doi:10.3390/molecules23040849
Huang, J. L., Chang, Y. T., Hong, Z. Y., and Lin, C. S. (2022). Targeting DNA damage response and immune checkpoint for anticancer therapy. Int. J. Mol. Sci. 23, 3238. doi:10.3390/ijms23063238
Jiang, M., Chen, P., Wang, L., Li, W., Chen, B., Liu, Y., et al. (2020). cGAS-STING, an important pathway in cancer immunotherapy. J. Hematol. Oncol. 13, 81. doi:10.1186/s13045-020-00916-z
Jiang, M., Jia, K., Wang, L., Li, W., Chen, B., Liu, Y., et al. (2021). Alterations of DNA damage response pathway: Biomarker and therapeutic strategy for cancer immunotherapy. Acta Pharm. Sin. B 11, 2983–2994. doi:10.1016/j.apsb.2021.01.003
Ka, N. L., Lim, G. Y., Hwang, S., Kim, S. S., and Lee, M. O. (2021). IFI16 inhibits DNA repair that potentiates type-I interferon-induced antitumor effects in triple negative breast cancer. Cell Rep. 37, 110138. doi:10.1016/j.celrep.2021.110138
Kerr, A. J., Dodwell, D., McGale, P., Holt, F., Duane, F., Mannu, G., et al. (2022). Adjuvant and neoadjuvant breast cancer treatments: A systematic review of their effects on mortality. Cancer Treat. Rev. 105, 102375. doi:10.1016/j.ctrv.2022.102375
Konno, H., Yamauchi, S., Berglund, A., Putney, R. M., Mulé, J. J., and Barber, G. N. (2018). Suppression of STING signaling through epigenetic silencing and missense mutation impedes DNA damage mediated cytokine production. Oncogene 37, 2037–2051. doi:10.1038/s41388-017-0120-0
Krishnan, R., Patel, P. S., and Hakem, R. (2021). BRCA1 and metastasis: Outcome of defective DNA repair. Cancers (Basel) 14, 108. doi:10.3390/cancers14010108
Kulasinghe, A., Monkman, J., Shah, E. T., Matigian, N., Adams, M. N., and O'Byrne, K. (2021). Spatial profiling identifies prognostic features of response to adjuvant therapy in triple negative breast cancer (TNBC). Front. Oncol. 11, 798296. doi:10.3389/fonc.2021.798296
Kwon, M., Nam, G. H., Jung, H., Kim, S. A., Kim, S., Choi, Y., et al. (2021). Statin in combination with cisplatin makes favorable tumor-immune microenvironment for immunotherapy of head and neck squamous cell carcinoma. Cancer Lett. 522, 198–210. doi:10.1016/j.canlet.2021.09.029
Lara, P. N., Douillard, J. Y., Nakagawa, K., von Pawel, J., McKeage, M. J., Albert, I., et al. (2011). Randomized phase III placebo-controlled trial of carboplatin and paclitaxel with or without the vascular disrupting agent vadimezan (ASA404) in advanced non-small-cell lung cancer. J. Clin. Oncol. 29, 2965–2971. doi:10.1200/jco.2011.35.0660
Lee, K. J., Mann, E., Wright, G., Piett, C. G., Nagel, Z. D., and Gassman, N. R. (2020). Exploiting DNA repair defects in triple negative breast cancer to improve cell killing. Ther. Adv. Med. Oncol. 12, 1758835920958354. doi:10.1177/1758835920958354
Lee, S. J., Yang, H., Kim, W. R., Lee, Y. S., Lee, W. S., Kong, S. J., et al. (2021). STING activation normalizes the intraperitoneal vascular-immune microenvironment and suppresses peritoneal carcinomatosis of colon cancer. J. Immunother. Cancer 9, e002195. doi:10.1136/jitc-2020-002195
Li, D., Ji, H., Niu, X., Yin, L., Wang, Y., Gu, Y., et al. (2020b). Tumor-associated macrophages secrete CC-chemokine ligand 2 and induce tamoxifen resistance by activating PI3K/Akt/mTOR in breast cancer. Cancer Sci. 111, 47–58. doi:10.1111/cas.14230
Li, J., and Bakhoum, S. F. (2022). The pleiotropic roles of cGAS-STING signaling in the tumor microenvironment. J. Mol. Cell Biol. 14, mjac019. doi:10.1093/jmcb/mjac019
Li, S., Mirlekar, B., Johnson, B. M., Brickey, W. J., Wrobel, J. A., Yang, N., et al. (2022). STING-induced regulatory B cells compromise NK function in cancer immunity. Nature 610, 373–380. doi:10.1038/s41586-022-05254-3
Li, T., and Chen, Z. J. (2018). The cGAS-cGAMP-STING pathway connects DNA damage to inflammation, senescence, and cancer. J. Exp. Med. 215, 1287–1299. doi:10.1084/jem.20180139
Li, Y., Hodge, J., Liu, Q., Wang, J., Wang, Y., Evans, T. D., et al. (2020a). TFEB is a master regulator of tumor-associated macrophages in breast cancer. J. Immunother. Cancer 8, e000543. doi:10.1136/jitc-2020-000543
Liu, X., Wang, M., Yang, M., Sun, H., Wang, B., Pan, X., et al. (2022b). Structure-activity relationship study of amidobenzimidazole derivatives as stimulator of interferon genes (STING) agonists. Eur. J. Med. Chem. 246, 114943. doi:10.1016/j.ejmech.2022.114943
Liu, Y., Xu, P., Rivara, S., Liu, C., Ricci, J., Ren, X., et al. (2022a). Clathrin-associated AP-1 controls termination of STING signalling. Nature 610, 761–767. doi:10.1038/s41586-022-05354-0
Lohard, S., Bourgeois, N., Maillet, L., Gautier, F., Fétiveau, A., Lasla, H., et al. (2020). STING-dependent paracriny shapes apoptotic priming of breast tumors in response to anti-mitotic treatment. Nat. Commun. 11, 259. doi:10.1038/s41467-019-13689-y
McArthur, K., Whitehead, L. W., Heddleston, J. M., Li, L., Padman, B. S., Oorschot, V., et al. (2018). BAK/BAX macropores facilitate mitochondrial herniation and mtDNA efflux during apoptosis. Science 359, eaao6047. doi:10.1126/science.aao6047
McKeage, M. J., Reck, M., Jameson, M. B., Rosenthal, M. A., Gibbs, D., Mainwaring, P. N., et al. (2009). Phase II study of ASA404 (vadimezan, 5,6-dimethylxanthenone-4-acetic acid/DMXAA) 1800mg/m(2) combined with carboplatin and paclitaxel in previously untreated advanced non-small cell lung cancer. Lung Cancer 65, 192–197. doi:10.1016/j.lungcan.2009.03.027
Mehraj, U., Ganai, R. A., Macha, M. A., Hamid, A., Zargar, M. A., Bhat, A. A., et al. (2021). The tumor microenvironment as driver of stemness and therapeutic resistance in breast cancer: New challenges and therapeutic opportunities. Cell Oncol. (Dordr) 44, 1209–1229. doi:10.1007/s13402-021-00634-9
Meric-Bernstam, F., Sweis, R. F., Hodi, F. S., Messersmith, W. A., Andtbacka, R. H. I., Ingham, M., et al. (2022a). Phase I dose-escalation trial of MIW815 (ADU-S100), an intratumoral STING agonist, in patients with advanced/metastatic solid tumors or lymphomas. Clin. Cancer Res. 28, 677–688. doi:10.1158/1078-0432.Ccr-21-1963
Meric-Bernstam, F., Sweis, R. F., Kasper, S., Hamid, O., Bhatia, S., Dummer, R., et al. (2022b). Combination of the STING agonist MIW815 (ADU-S100) and PD-1 inhibitor spartalizumab in advanced/metastatic solid tumors or lymphomas: An open-label, multicenter, phase ib study. Clin. Cancer Res. 29, 110–121. doi:10.1158/1078-0432.Ccr-22-2235
Pan, B. S., Perera, S. A., Piesvaux, J. A., Presland, J. P., Schroeder, G. K., Cumming, J. N., et al. (2020). An orally available non-nucleotide STING agonist with antitumor activity. Science 369, eaba6098. doi:10.1126/science.aba6098
Parkes, E. E., Walker, S. M., Taggart, L. E., McCabe, N., Knight, L. A., Wilkinson, R., et al. (2017). Activation of STING-dependent innate immune signaling by S-Phase-Specific DNA damage in breast cancer. J. Natl. Cancer Inst. 109, djw199. doi:10.1093/jnci/djw199
Perera, S. A., Kopinja, J. E., Ma, Y., Muise, E. S., Laskey, J., Chakravarthy, K., et al. (2022). STimulator of INterferon genes agonism accelerates antitumor activity in poorly immunogenic tumors. Mol. Cancer Ther. 21, 282–293. doi:10.1158/1535-7163.Mct-21-0136
Pilié, P. G., Tang, C., Mills, G. B., and Yap, T. A. (2019). State-of-the-art strategies for targeting the DNA damage response in cancer. Nat. Rev. Clin. Oncol. 16, 81–104. doi:10.1038/s41571-018-0114-z
Qiu, X., Qu, Y., Guo, B., Zheng, H., Meng, F., and Zhong, Z. (2022). Micellar paclitaxel boosts ICD and chemo-immunotherapy of metastatic triple negative breast cancer. J. Control Release 341, 498–510. doi:10.1016/j.jconrel.2021.12.002
Ragu, S., Matos-Rodrigues, G., and Lopez, B. S. (2020). Replication stress, DNA damage, inflammatory cytokines and innate immune response. Genes (Basel) 11, 409. doi:10.3390/genes11040409
Reisländer, T., Groelly, F. J., and Tarsounas, M. (2020). DNA damage and cancer immunotherapy: A STING in the tale. Mol. Cell 80, 21–28. doi:10.1016/j.molcel.2020.07.026
Riley, J. S., Quarato, G., Cloix, C., Lopez, J., O'Prey, J., Pearson, M., et al. (2018). Mitochondrial inner membrane permeabilisation enables mtDNA release during apoptosis. Embo J. 37, e99238. doi:10.15252/embj.201899238
Sen, T., Rodriguez, B. L., Chen, L., Corte, C. M. D., Morikawa, N., Fujimoto, J., et al. (2019). Targeting DNA damage response promotes antitumor immunity through STING-mediated T-cell activation in small cell lung cancer. Cancer Discov. 9, 646–661. doi:10.1158/2159-8290.Cd-18-1020
Shen, M., Chen, C., Guo, Q., Wang, Q., Liao, J., Wang, L., et al. (2022). Systemic delivery of mPEG-masked trispecific T-cell nanoengagers in synergy with STING agonists overcomes immunotherapy resistance in TNBC and generates a vaccination effect. Adv. Sci. (Weinh) 9, e2203523. doi:10.1002/advs.202203523
Shih, A. Y., Damm-Ganamet, K. L., and Mirzadegan, T. (2018). Dynamic structural differences between human and mouse STING lead to differing sensitivity to DMXAA. Biophys. J. 114, 32–39. doi:10.1016/j.bpj.2017.10.027
Siegel, R. L., Miller, K. D., Fuchs, H. E., and Jemal, A. (2022). Cancer statistics, 2022. CA Cancer J. Clin. 72, 7–33. doi:10.3322/caac.21708
Singh, A. K., Praharaj, M., Lombardo, K. A., Yoshida, T., Matoso, A., Baras, A. S., et al. (2022). Re-engineered BCG overexpressing cyclic di-AMP augments trained immunity and exhibits improved efficacy against bladder cancer. Nat. Commun. 13, 878. doi:10.1038/s41467-022-28509-z
Smith, J. A. (2020). STING, the endoplasmic reticulum, and mitochondria: Is three a crowd or a conversation? Front. Immunol. 11, 611347. doi:10.3389/fimmu.2020.611347
Soongsathitanon, J., Jamjuntra, P., Sumransub, N., Yangngam, S., De la Fuente, M., Landskron, G., et al. (2021). Crosstalk between tumor-infiltrating immune cells and cancer-associated fibroblasts in tumor growth and immunosuppression of breast cancer. J. Immunol. Res. 2021, 8840066. doi:10.1155/2021/8840066
Staniszewska, A. D., Armenia, J., King, M., Michaloglou, C., Reddy, A., Singh, M., et al. (2022). PARP inhibition is a modulator of anti-tumor immune response in BRCA-deficient tumors. Oncoimmunology 11, 2083755. doi:10.1080/2162402x.2022.2083755
Su, M., Zheng, J., Gan, L., Zhao, Y., Fu, Y., and Chen, Q. (2022). Second messenger 2'3'-cyclic GMP-AMP (2'3'-cGAMP): Synthesis, transmission, and degradation. Biochem. Pharmacol. 198, 114934. doi:10.1016/j.bcp.2022.114934
Sun, L., Wu, J., Du, F., Chen, X., and Chen, Z. J. (2013). Cyclic GMP-AMP synthase is a cytosolic DNA sensor that activates the type I interferon pathway. Science 339, 786–791. doi:10.1126/science.1232458
Sun, P., Zhang, X., Wang, R. J., Ma, Q. Y., Xu, L., Wang, Y., et al. (2021). PI3Kα inhibitor CYH33 triggers antitumor immunity in murine breast cancer by activating CD8(+)T cells and promoting fatty acid metabolism. J. Immunother. Cancer 9, e003093. doi:10.1136/jitc-2021-003093
Sung, H., Ferlay, J., Siegel, R. L., Laversanne, M., Soerjomataram, I., Jemal, A., et al. (2021). Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 71, 209–249. doi:10.3322/caac.21660
Tchou, J., Zhao, Y., Levine, B. L., Zhang, P. J., Davis, M. M., Melenhorst, J. J., et al. (2017). Safety and efficacy of intratumoral injections of chimeric antigen receptor (CAR) T cells in metastatic breast cancer. Cancer Immunol. Res. 5, 1152–1161. doi:10.1158/2326-6066.Cir-17-0189
Tchounwou, P. B., Dasari, S., Noubissi, F. K., Ray, P., and Kumar, S. (2021). Advances in our understanding of the molecular mechanisms of action of cisplatin in cancer therapy. J. Exp. Pharmacol. 13, 303–328. doi:10.2147/jep.S267383
Thomsen, M. K., Skouboe, M. K., Boularan, C., Vernejoul, F., Lioux, T., Leknes, S. L., et al. (2020). The cGAS-STING pathway is a therapeutic target in a preclinical model of hepatocellular carcinoma. Oncogene 39, 1652–1664. doi:10.1038/s41388-019-1108-8
Tian, C., Wei, Y., Li, J., Huang, Z., Wang, Q., Lin, Y., et al. (2022). A novel CDK4/6 and PARP dual inhibitor ZC-22 effectively suppresses tumor growth and improves the response to cisplatin treatment in breast and ovarian cancer. Int. J. Mol. Sci. 23, 2892. doi:10.3390/ijms23052892
Tung, N., Lin, N. U., Kidd, J., Allen, B. A., Singh, N., Wenstrup, R. J., et al. (2016). Frequency of germline mutations in 25 cancer susceptibility genes in a sequential series of patients with breast cancer. J. Clin. Oncol. 34, 1460–1468. doi:10.1200/jco.2015.65.0747
Vasiyani, H., Mane, M., Rana, K., Shinde, A., Roy, M., Singh, J., et al. (2022). DNA damage induces STING mediated IL-6-STAT3 survival pathway in triple-negative breast cancer cells and decreased survival of breast cancer patients. Apoptosis 27, 961–978. doi:10.1007/s10495-022-01763-8
Vasiyani, H., Shinde, A., Roy, M., Mane, M., Singh, K., Singh, J., et al. (2021). The analog of cGAMP, c-di-AMP, activates STING mediated cell death pathway in estrogen-receptor negative breast cancer cells. Apoptosis 26, 293–306. doi:10.1007/s10495-021-01669-x
Wang, D., Nie, T., Huang, C., Chen, Z., Ma, X., Fang, W., et al. (2022). Metal-cyclic dinucleotide nanomodulator-stimulated STING signaling for strengthened radioimmunotherapy of large tumor. Small 18, e2203227. doi:10.1002/smll.202203227
Wang, Y., Luo, J., Alu, A., Han, X., Wei, Y., and Wei, X. (2020). cGAS-STING pathway in cancer biotherapy. Mol. Cancer 19, 136. doi:10.1186/s12943-020-01247-w
Wehbe, M., Wang-Bishop, L., Becker, K. W., Shae, D., Baljon, J. J., He, X., et al. (2021). Nanoparticle delivery improves the pharmacokinetic properties of cyclic dinucleotide STING agonists to open a therapeutic window for intravenous administration. J. Control Release 330, 1118–1129. doi:10.1016/j.jconrel.2020.11.017
Weinberg, S. E., Sena, L. A., and Chandel, N. S. (2015). Mitochondria in the regulation of innate and adaptive immunity. Immunity 42, 406–417. doi:10.1016/j.immuni.2015.02.002
Wengner, A. M., Scholz, A., and Haendler, B. (2020). Targeting DNA damage response in prostate and breast cancer. Int. J. Mol. Sci. 21, 8273. doi:10.3390/ijms21218273
White, M. J., McArthur, K., Metcalf, D., Lane, R. M., Cambier, J. C., Herold, M. J., et al. (2014). Apoptotic caspases suppress mtDNA-induced STING-mediated type I IFN production. Cell 159, 1549–1562. doi:10.1016/j.cell.2014.11.036
Wilson, B. E., Gorrini, C., and Cescon, D. W. (2022). Breast cancer immune microenvironment: From pre-clinical models to clinical therapies. Breast Cancer Res. Treat. 191, 257–267. doi:10.1007/s10549-021-06431-0
Wu, H. J., Hao, M., Yeo, S. K., and Guan, J. L. (2020). FAK signaling in cancer-associated fibroblasts promotes breast cancer cell migration and metastasis by exosomal miRNAs-mediated intercellular communication. Oncogene 39, 2539–2549. doi:10.1038/s41388-020-1162-2
Xia, C., Dong, X., Li, H., Cao, M., Sun, D., He, S., et al. (2022). Cancer statistics in China and United States, 2022: Profiles, trends, and determinants. Chin. Med. J. Engl. 135, 584–590. doi:10.1097/cm9.0000000000002108
Xu, N., Palmer, D. C., Robeson, A. C., Shou, P., Bommiasamy, H., Laurie, S. J., et al. (2021). STING agonist promotes CAR T cell trafficking and persistence in breast cancer. J. Exp. Med. 218, e20200844. doi:10.1084/jem.20200844
Yang, L., Xie, H. J., Li, Y. Y., Wang, X., Liu, X. X., and Mai, J. (2022). Molecular mechanisms of platinum-based chemotherapy resistance in ovarian cancer (Review). Oncol. Rep. 47, 82. doi:10.3892/or.2022.8293
Yang, Y., Karakhanova, S., Hartwig, W., D'Haese, J. G., Philippov, P. P., Werner, J., et al. (2016). Mitochondria and mitochondrial ROS in cancer: Novel targets for anticancer therapy. J. Cell Physiol. 231, 2570–2581. doi:10.1002/jcp.25349
Yu, Y., Liu, J., Liu, C., Liu, R., Liu, L., Yu, Z., et al. (2022). Post-translational modifications of cGAS-STING: A critical switch for immune regulation. Cells 11, 3043. doi:10.3390/cells11193043
Zhang, K., Wang, S., Gou, H., Zhang, J., and Li, C. (2021). Crosstalk between autophagy and the cGAS-STING signaling pathway in type I interferon production. Front. Cell Dev. Biol. 9, 748485. doi:10.3389/fcell.2021.748485
Zhang, L., Zhao, J., Hu, X., Wang, C., Jia, Y., Zhu, C., et al. (2022b). A peritumorally injected immunomodulating adjuvant elicits robust and safe metalloimmunotherapy against solid tumors. Adv. Mater 34, e2206915. doi:10.1002/adma.202206915
Zhang, X., Bai, X. C., and Chen, Z. J. (2020). Structures and mechanisms in the cGAS-STING innate immunity pathway. Immunity 53, 43–53. doi:10.1016/j.immuni.2020.05.013
Zhang, Z., Liu, J., Xiao, M., Zhang, Q., Liu, Z., Liu, M., et al. (2022c). Peptide nanotube loaded with a STING agonist, c-di-GMP, enhance cancer immunotherapy against melanoma. Nano Res. 16, 5206–5215. doi:10.1007/s12274-022-5102-z
Zhang, Z., Zhou, H., Ouyang, X., Dong, Y., Sarapultsev, A., Luo, S., et al. (2022a). Multifaceted functions of STING in human health and disease: From molecular mechanism to targeted strategy. Signal Transduct. Target Ther. 7, 394. doi:10.1038/s41392-022-01252-z
Zhao, Y., Qu, T., Wang, P., Li, X., Qiang, J., Xia, Z., et al. (2016). Unravelling the relationship between macroautophagy and mitochondrial ROS in cancer therapy. Apoptosis 21, 517–531. doi:10.1007/s10495-016-1236-3
Zheng, S., Zou, Y., Tang, Y., Yang, A., Liang, J. Y., Wu, L., et al. (2022). Landscape of cancer-associated fibroblasts identifies the secreted biglycan as a protumor and immunosuppressive factor in triple-negative breast cancer. Oncoimmunology 11, 2020984. doi:10.1080/2162402x.2021.2020984
Zhou, J., Ventura, C. J., Fang, R. H., and Zhang, L. (2022). Nanodelivery of STING agonists against cancer and infectious diseases. Mol. Asp. Med. 83, 101007. doi:10.1016/j.mam.2021.101007
Zhou, L., Zhang, Y., Wang, Y., Zhang, M., Sun, W., Dai, T., et al. (2020). A dual role of type I interferons in antitumor immunity. Adv. Biosyst. 4, e1900237. doi:10.1002/adbi.201900237
Zhu, C., Ma, Q., Gong, L., Di, S., Gong, J., Wang, Y., et al. (2022a). Manganese-based multifunctional nanoplatform for dual-modal imaging and synergistic therapy of breast cancer. Acta Biomater. 141, 429–439. doi:10.1016/j.actbio.2022.01.019
Zhu, Y., Hu, Y., Tang, C., Guan, X., and Zhang, W. (2022c). Platinum-based systematic therapy in triple-negative breast cancer. Biochim. Biophys. Acta Rev. Cancer 1877, 188678. doi:10.1016/j.bbcan.2022.188678
Zhu, Z., McGray, A. J. R., Jiang, W., Lu, B., Kalinski, P., and Guo, Z. S. (2022b). Improving cancer immunotherapy by rationally combining oncolytic virus with modulators targeting key signaling pathways. Mol. Cancer 21, 196. doi:10.1186/s12943-022-01664-z
Keywords: STING, DNA damage response, tumour immune microenvironment, mitochondrial function, STING agonists
Citation: Ying-Rui M, Bu-Fan B, Deng L, Rong S and Qian-Mei Z (2023) Targeting the stimulator of interferon genes (STING) in breast cancer. Front. Pharmacol. 14:1199152. doi: 10.3389/fphar.2023.1199152
Received: 03 April 2023; Accepted: 21 June 2023;
Published: 28 June 2023.
Edited by:
Daiqing Liao, University of Florida, United StatesReviewed by:
Xiawei Cheng, East China University of Science and Technology, ChinaXiaotong Sun, Yantai Yuhuangding Hospital, China
Copyright © 2023 Ying-Rui, Bu-Fan, Deng, Rong and Qian-Mei. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zhou Qian-Mei, dGF6aG91QDE2My5jb20=
†These authors have contributed equally to this work