
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Pharmacol. , 15 June 2023
Sec. Experimental Pharmacology and Drug Discovery
Volume 14 - 2023 | https://doi.org/10.3389/fphar.2023.1152588
This article is part of the Research Topic Drug-ability Strategies for Potential Antimycobacterial Candidate: Opportunities and Challenges View all 5 articles
 Mariana Martins Drumond1,2*
Mariana Martins Drumond1,2* Ana Paula Tapia-Costa3Elisabeth Neumann4
Ana Paula Tapia-Costa3Elisabeth Neumann4 Álvaro Cantini Nunes5Jorge Wanderson Barbosa6Diego E. Kassuha3
Álvaro Cantini Nunes5Jorge Wanderson Barbosa6Diego E. Kassuha3 Pamela Mancha-Agresti2*
Pamela Mancha-Agresti2*Aim: This study aims to verify the antibacterial and antibiofilm action of cell-free spent medium (CFSM) from four lactic acid bacteria with potential probiotic characteristics (Lactiplantibacillus plantarum, Lactobacillus acidophilus, Lactobacillus johnsonii, and Lactobacillus delbrueckii) against two Pseudomonas aeruginosa strains.
Main methods: The minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) of the CFSM, antibacterial activity by analysing the formation of inhibition zones, and inhibition of planktonic cultures were determined. Whether an increase in the concentration of CFSM influenced the growth of pathogenic strains and the anti-adhesive activity of the CFSM in biofilm formation (crystal violet and MTT assays) were determined, which were all corroborated by using scanning electron microscopy.
Key findings: The relationship between the MIC and MBC values showed a bactericidal or bacteriostatic effect for all the cell-free spent media (CFSMs) tested for P. aeruginosa 9027™ and 27853™ strains. The CFSM supplemental doses of 18 or 22%, 20 or 22%, 46 or 48%, and 50 or 54% of L. acidophilus, L. delbrueckii, L. plantarum, and L. johnsonii, respectively, could completely inhibit the growth of both pathogen strains. The antibiofilm activity of the CFSM in three biofilm conditions (pre-coated, co-incubated, and preformed) demonstrated values ranging between 40% and 80% for biofilm inhibition, and similar results were observed for cell viability.
Significance: This work provides strong evidence that the postbiotic derived from different Lactobacilli could be practical as an adjuvant therapy for reducing the use of antibiotics, being a good candidate to overcome the growing challenge of hospital infections due to this pathogen.
The gram-negative opportunistic human pathogen Pseudomonas aeruginosa is responsible for much of the morbidity and mortality in nosocomial settings (Serra et al., 2015), leading to a wide variety of life-threatening acute and chronic infections that compromise the soft tissues, bloodstream, lungs, and urinary tract (Driscoll et al., 2007), specifically in cancer, cystic fibrosis, and immunocompromised patients (Wu and Zhu, 2021). The mortality rates are over 35% for hospital-acquired P. aeruginosa infectious, like ventilator-associated pneumonia or bacteraemia (Lynch et al., 2017), and approximately 75% for sepsis in patients who have more than 40% of their total body surface burned (Atiyeh et al., 2005), the skin and soft tissues being the principal areas infected by P. aeruginosa, which represents 21% of all cases of bacteraemia (Wahab and Rahman, 2013). The colonisation of burned tissues by P. aeruginosa characteristically results in a significantly greater area of infection and thereby postpones or prevents the healing process (Gjødsbøl et al., 2006). Thus, this pathogen causes the highest mortality rate among gram-negative bacteria, proteases, and lipases—known as virulence factors—which are used to degrade host barriers and evade immune mechanisms, allowing bacterial invasion of the bloodstream from the skin and consequently leading to soft tissue infections (Wu et al., 2011).
Pseudomonas aeruginosa is a biofilm-forming bacterium with a high intercellular signalling pathway, the quorum sensing (QS) system; these processes represent bacterial social behaviours. The QS-controlled functions are critical for acute virulence since these processes appear to play an essential role in the development of biofilms (Passador et al., 1993; Parsek and Greenberg, 2005; Atkinson, 2007). Biofilms are considered an aggregation of microorganism communities, attached and growing either on biotic or abiotic surfaces, generating a protection matrix by secreting biochemical substances and forming a physical semipermeable barrier that limits the diffusion of molecules (Hall-Stoodley et al., 2004; Mihai et al., 2018). When compared to planktonic cells, the pathogen microorganisms present in biofilms are highly resistant to antimicrobial agents (Penesyan et al., 2015), and the development of antibiotic resistance by P. aeruginosa has been widely documented. Thus, there is an urgent need to look for alternative strategies to treat chronic, biofilm-mediated infections of P. aeruginosa.
Live probiotics are microorganisms that offer health benefits to the host by direct or indirect interactions between cells or through their released metabolites. Thus, they are suitable for many different approaches (Reid et al., 2011). The safety of live microorganisms persists as a debated issue, especially for susceptible individuals (Ravel et al., 2011; Borges et al., 2014). For this reason, different strategies have been developed to replace the use of live microorganisms, such as the use of killed probiotics called paraprobiotics (Trindade et al., 2021), microbial extracts, and cell-free supernatants (Piqué et al., 2019; Batista et al., 2022). Thus, antimicrobial elements secreted by probiotic microorganisms can be called “postbiotics,” which include non-viable cells that promote health benefits to the host when administered in adequate amounts (Collado et al., 2019); Salminen et al., 2021).
Postbiotics are compounds from several bioactive compounds that are secreted by the action of microorganisms either on food ingredients (e.g., bioactive peptides) or during growth and fermentation in complex microbiological cultures having bioactive soluble factors (Aguilar-Toalá et al., 2018; Collado et al., 2019; Barros et al., 2020; Cuevas-González et al., 2020). Organic acids, short-chain fatty acids, carbohydrates, antimicrobial peptides, enzymes, vitamins, cofactors, immune-signalling compounds, and complex agents have also been described as bioactive metabolites present in postbiotics prepared from lactic acid bacteria (LAB) (Moradi et al., 2019; Rajakovich and Balskus, 2019).
Exciting features of postbiotics, such as no biogenic amine production, a safe structure, being incapable of transferring antibiotic resistance, a defined chemical composition, stability in a broad range of temperatures and pH, and a broad spectrum of antimicrobial activity, make them valuable ingredients to be used in different approaches (Barros et al., 2020; Rad et al., 2021).
In this context, we aimed to develop an interesting and alternative use of the postbiotics of four LAB with potential probiotic characteristics (Lactiplantibacillus plantarum, Lactobacillus acidophilus, Lactobacillus johnsonii, and Lactobacillus delbrueckii) to combat P. aeruginosa and their ability to reduce the formation of biofilms by this pathogenic bacteria.
The bacterial strains used in this study are summarised in Table 1. The four probiotic strains belonging to the Lactobacilli group (L. acidophilus, L. delbrueckii, L. johnsonii, and L. plantarum) were grown in De Man, Rogosa, and Sharpe agar (MRS, neogem®, Rüdersdorf, Germany) for 24 h at 37°C. L. acidophilus and L. plantarum were isolated from a typical fermented Mexican drink; L. delbrueckii was isolated from raw cow’s milk from the culture collection of the Centro de Investigación y Desarrollo en Criotecnología de Alimentos (CIDCA, Facultad de Ciencias Exactas, Universidad Nacional de La Plata, Argentina); and L. johnsonii was isolated from a human vagina and kindly provided by Dr. Regina Nardi Drummond from the Laboratório de Microbiologia Aplicada of the Departamento de Microbiologia at the Universidade Federal de Minas Gerais (UFMG).

TABLE 1. Bacteria strains and precedence.
A single colony from each strain was transferred into the MRS broth under the same incubation conditions for 24 h to obtain the cell-free spent medium (CFSM). Pathogenic P. aeruginosa strains from the American Type Culture Collection (ATCC®) (PA 9027™ and PA 27853™) were cultured in the brain heart infusion (BHI) medium (HiMedia, India) for 16 h at 37°C.
The active overnight culture was diluted in fresh medium, starting from an initial optical density (OD600nm) of 0.04, and incubated at 37°C in microaerobiosis for 24 h. To create this condition, the falcon tubes were filled to the top with the medium without shaking for 24 h at 37°C. The CFSM of each probiotic strain was obtained as previously described by Bayoumi and Griffiths (2012). Briefly, the overnight culture of the four probiotic strains was prepared by centrifugation at 6,000g/4°C for 10 min, and the cells were then removed. The supernatant was filter sterilised with a 0.22 μm pore filter (Kasvi, São José dos Pinhais, Paraná) to remove the residue of viable cells and referred to as CFSM. An aliquot of the CFSM of each probiotic strain was plated on solid MRS medium and incubated at 37°C for 48 h to ensure the absence of viable cells. The CFSM of all probiotic strains was stored at −20°C until use in further assays.
To determine the minimum inhibitory concentration (MIC) of each CFSM, the broth microdilution method was used in 96-well round-bottomed polystyrene microtiter plates (SPL Life Sciences, Pocheon-si, Korea) conducted according to the methodology described by Ruíz et al. (2012), with some modifications. The CFSM was serially diluted in the BHI broth (HiMedia, India) using a two-fold dilution; 100 μL of diluted CFSM was placed in each well and 25 μL of an overnight-grown culture of the indicator strain, P. aeruginosa, was added after adjusting its absorbance to 0.5 at 600 nm optical density equivalent to 3.0 × 1014 colony-forming units (CFU)/mL for PA 27853™ and 14.8 × 1014 CFU/mL for PA 9027™. The plate was incubated at 37°C for 24 h under aerobic conditions. Positive (P. aeruginosa + BHI) and negative (only BHI medium) controls were also included on the plate. The MIC values were defined as the lowest concentration of CFSM that could inhibit the visible growth of the pathogenic bacteria.
To assess the minimum bactericidal concentrations (MBCs), 50 μL from the wells with no apparent growth, and without turbidity, were spread onto the surface of solid BHI agar (1.5%) (BD Difco™ Bacto agar, United States) on Petri dishes and incubated at 37°C for 24 h. These assays were performed as three independent experiments with three replicates per CFSM analysed, and the experiment was conducted at three different times. The sample with the lowest concentration of CFSM that showed no growth on BHI agar was recorded as the MBC. The MIC levels were further used in the following assessment.
The overnight cultures of P. aeruginosa (ATCC® 9027™ and 27853™) were cultured in BHI medium for 16 h at 37°C. A suspension of each bacterium of 0.5 absorbance (OD600nm) was adjusted for use in the agar diffusion method to determine the growth inhibition zone of the tested bacteria, following the methodology as described by Cruz et al. (2001), with some modifications. First, a bottom layer (9 mL) of MRS was prepared in the Petri dish (90 × 60 mm). Second, a top layer of molten and cooled BHI medium (5 mL) was mixed with each tested bacterial suspension, containing 3.0 × 1014 and 14.8 × 1014 CFU/mL for PA 27853™ and 9027™, respectively, and poured onto the bottom layer. One well of 9 mm diameter was prepared in each plate, and 100 μL (1×), 200 μL (2×), and 300 μL (3×) of each pure postbiotic were introduced into each well. The plates were incubated for 24 h at 37°C. A clear inhibition zone of more than 1 mm was formed around the well and measured and recorded as a positive inhibitory result. The measurement was performed from the edge of the formed halo. The experiment was conducted at three different times. To evaluate whether the potential inhibitory effect of the CFSM could be due to its acidic pH, it was neutralised (pH = 7) with NaOH (5 M) (Neon, São Paulo, Brazil), and the assay was repeated twice.
The antibacterial activity of the CFSM for four Lactobacillaceae strains against planktonic cultures of P. aeruginosa (PA 9027™ and PA 27853™) was assessed according to the methodology described by Lin et al. (2015), with some modifications. Briefly, the CFSM was prepared as described previously; the overnight culture of the microbial cells was centrifuged (5,000 rpm/5 min), and the pellets were washed twice with 0.90% NaCl solution (Synthia, Sao Paulo, Brazil). As mentioned above, the cell suspensions were adjusted to 0.5 (OD600nm). Then, 250 μL of each P. aeruginosa suspension was faced with 250 μL each of the CFSM and mixed with 1.5 mL of sterile physiological solution. The control group was formed as follows: 250 μL of microbial P. aeruginosa suspension was faced with 250 μL of MRS medium (the medium where the probiotic bacteria were grown) plus 1.5 mL of sterile physiological solution. The cultures were incubated at 37°C for 24 h and then diluted and plated on a solid BHI medium (HiMedia, India). The plates were incubated (37°C/24 h), and the CFU/mL was determined. This assay was performed as two independent experiments with two replicates per group.
To establish the percentage of CFSM able to inhibit the growth of P. aeruginosa, the growth curve assay was conducted with increased CFSM concentrations. This assay was performed as two independent experiments. Briefly, 200 μL of overnight culture at 0.04 (OD600nm) (8.3 × 109 CFU/mL for PA 9027™ and 7.43 × 109 CFU/mL for PA 27853™) was placed in 96-well polystyrene plates (SPL Life Sciences, Pocheon-si, Korea) as the initial point of the curve, this being the point with 100% of pathogen cells. Then, 98% of the pathogen culture (196 μL) was placed in the next well with 2% (4 μL) of each CFSM; 96% of the pathogen culture and 4% of each CFSM were placed in the next well; and so on, with the final well containing 40% of the pathogen cells and 60% of each CFSM. The microtiter plates were incubated at 37°C for 16 h when the pathogen strains had reached the stationary phase. The optical density was then measured. This assay was performed in triplicate. It is important to highlight that the same experiment was previously carried out with pathogens and increasing MRS broth volumes to evaluate if the MRS had any influence on the growing pathogens.
The anti-adhesion activity of the CFSM for the Lactobacillaceae against P. aeruginosa spp. was performed in pre-coated, co-incubated, and preformed biofilm experiments.
The pre-coated experiments were carried out according to the methodology described by Gudiña et al. (2010). Briefly, 96-well microtiter plates (SPL Life Sciences, Pocheon-si, Korea) were coated with 200 μL of different CFSMs in triplicate, and the microtiter plates were incubated at 37°C for 24 h. This volume was chosen to cover both the bottom and sides of the well. After this time, the CFSM was removed, and plates were washed twice with 100 μL of phosphate buffer saline (PBS, 0.1 M), pH 7.2, to remove the non-adhering supernatant. Then, 200 μL of each 16-h growth culture of P. aeruginosa suspension (1 × 109 CFU/mL) cultured in BHI broth (HiMedia, India) with sucrose (1%) to enhance the formation of densely clustered microcolonies in polystyrene material (LabSynth, São Paulo, Brazil) was added to each well. The microtiter plate was incubated at 37°C for 24 h under sterile conditions to allow cell attachment. To remove the non-adhering cells, the plate was gently washed twice with 200 μL PBS (0.1 M), pH 7.2. Biofilm formation was assessed using the Crystal Violet assay (Newprov, Pinhais, Paraná, Brasil). This methodology was chosen as the reference protocol for detecting biofilm formation by isolated food pathogens (Rodrigues et al., 2010; Dias-Souza, 2013; Marinho et al., 2013). The formed biofilm was fixed for 15 min by adding 300 μL of methanol (99% v/v) (Dinâmica, São Paulo, Brasil) to each well, and the plate was air dried. After that, 300 μL of crystal violet (0.5%) was added and incubated at room temperature for 15 min. Crystal violet was then removed, the wells were washed with 200 μL of distilled water to remove the excess stain, and the plate was air dried. Then, 200 μL of 95% ethanol (Dinâmica, São Paulo, Brasil) was added to the wells, and 100 μL of this distaining solution from each well was transferred to a new plate. The absorbance was then measured at 595 nm using a microplate reader (Multiskan Spectrum, Thermo Scientific). Pathogenic bacterial suspension without CFSM was prepared as the control. The percentage reduction in adherence was calculated using the following equation:
where ODc represents the optical density of the well with CFSM and pathogenic suspension, while ODo represents the optical density of the pathogenic suspension without CFSM (control).
The microtiter plate anti-adhesion assay estimates the percentage reduction of pathogenic bacteria adhesion to the control wells, which was 0% in the absence of Lactobacillaceae CFSM. This analysis was carried out in triplicate, and the mean optical density was taken.
In the co-incubated experiment, the capacity of the CFSMs to inhibit biofilm formation was evaluated. The suspension of the previously activated pathogenic bacteria (P. aeruginosa) was centrifugated at 10,000g for 5 min. The supernatant was discarded, and the pellet was washed thrice with PBS (0.1 M), pH 7.2. The cells were resuspended in BHI broth (HiMedia, India) with sucrose 1% (LabSynth, São Paulo, Brazil) and adjusted to absorbance 0.5 (OD600nm) equivalent to 3.0 × 1014 CFU/mL for PA 27853™ and 14.8 × 1014 CFU/mL for PA 9027™. Then, 100 µL of each bacterial solution was added to the individual well of a sterile flat-bottomed 96-well polystyrene microtiter plate (SPL Life Sciences, Pocheon-si, Korea) with 100 μL of different Lactobacillaceae CFSMs, and the plate was incubated at 37°C for 24 h. For the negative control, only BHI + sucrose 1% medium was added without the bacterial suspension, and the positive control was the bacterial suspension without CFSM. Biofilm formation was assessed using the Crystal Violet assay as described above. The mean absorbance (OD595nm) was used to determine the percentage inhibition of biomass formation according to the following equation:
The effect of the CFSM on biofilm growth and development was evaluated as described by Jadhav et al. (2013). Biofilms were allowed to form for 24 h before adding the CFSM. Biofilm formation was achieved by transferring 100 μL of bacterial culture that was prepared as described above (OD600nm = 0.5) into the wells of a sterile flat-bottomed 96-well polystyrene microtiter plate in triplicates. The microtiter plates were covered and incubated for 24 h at 37°C to allow cell attachment and biofilm formation. Following incubation, 100 μL of each CFSM was added to each well to reach a final volume of 200 µL. The plates were then incubated for 24 h. As the negative control, BHI + sucrose 1% medium was added without bacterial suspension, and the positive control was the bacterial suspension without CFSM. Finally, the biofilms were assessed for biomass attachment using the Crystal Violet assay, as previously mentioned. The Crystal Violet assay was performed as three independent experiments with n = 6 biofilms per CFSM.
To assess the metabolic activity of the biofilm formed, the modified 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT) reduction assay was used according to the study by Schillaci et al. (2008). MTT salt (Sigma Aldrich, São Paulo, Brazil) was dissolved in 0.1 M PBS to give a final concentration of 5 mg/mL. Plates containing preformed biofilm were incubated with 100 μL of each CFSM for 24 h. Then, the medium was gently removed, the plates were air dried, and 100 μL of MTT solution was added to each well and incubated for 3 h at 37°C under sterile conditions. The insoluble purple formazan (obtained by enzymatic hydrolysis of MTT by the dehydrogenase enzyme found in living cells) was further dissolved in 100 μL of dimethyl sulphoxide (DMSO, Sigma Aldrich, Darmstadt, Germany). The absorbance was then measured at an optical density of 570 nm using a microplate reader (Multiskan Spectrum, Thermo Scientific). The MTT assay was performed as three independent experiments with n = 6 biofilms per CFSM.
SEM analysed the effect of each postbiotic on biofilm inhibition. The goal was to use this technique to show biofilm inhibition by the CFSM. We chose only the co-incubated conditions of two P. aeruginosa studies since this is a qualitative experiment. To develop this assay, we used glass slides (0.13–0.17 mm thick, Global Glass, China) following the methodology as described by Marttinen et al. (2013) and Rossoni et al. (2018), with some modifications. The sterilised slides were placed in a 24-well polystyrene plate (SPL Life Sciences, Pocheon-si, Korea), and 400 μL of the overnight bacterial culture that was prepared as described above (OD600nm = 0.5) was added. The plates were incubated in a bacteriological incubator at 37°C for 1 h to promote the initial adhesion of P. aeruginosa onto the discs. Afterwards, 400 μL of each CFSM was added to each well. For the positive control, 400 μL of the MRS medium was used instead of the CFSM; the negative control was BHI + 1% sucrose medium, without slides. The plates were incubated at 37°C for 16 h. After biofilm formation, the wells were washed twice with 0.1 M PBS, and the specimens were fixed with methanol (99%) (Dinâmica, São Paulo, Brasil) for 1 h. The samples were then dehydrated in increasing ethanol (Dinâmica, São Paulo, Brasil) concentration series (10%, 25%, 50%, 75%, and 90%) for 20 min each, followed by immersion in 100% alcohol for 1 h. The plates were kept in a bacteriological incubator at 37°C for 24 h to permit total drying of the specimens. The dried slides were transferred to aluminium stubs and sputter coated with gold for 160 s at 40 mA (Denton Vacuum Desk II, Denton Vacuum LLC, Moorestown, NJ, United States). The samples were examined and imaged using a JEOL JSM-5600 scanning electron microscope (JEOL United States, Inc., Peabody, MA, United States) at the Characterisation Laboratory, Materials Engineering Department, Centro Federal de Educação Tecnológica de Minas Gerais (CEFET-MG). These experiments were performed twice with n = 2 biofilms per group.
The effect of each CFSM on biofilm formation was analysed using the GraphPad Prism 9.1 software. The one-way analysis of variance (ANOVA) method was used for multiple comparisons, followed by Tukey’s post hoc test (parametric data) or by the Kruskal–Wallis test and post-tested by using the Dunn’s test (non-parametric data). All experiments were performed at least in duplicates, and the standard deviation from the mean was calculated. Values were considered significantly different from each other at p < 0.05.
The MIC values of different CFSMs were determined by the microdilution method, and the results varied depending on the pathogenic bacteria and CFSM tested. The MIC and MBC values are presented in Table 2. The MIC values of each CFSM are important to investigate the mechanism of action of these products for their ability to inhibit biofilms. Pure CFSM and its concentrated forms (1×, 2×, and 3×) were used to analyse the planktonic growth and inhibition zones.

TABLE 2. MIC and MBC values.
To evaluate whether the MRS broth of Lactobacillaceae culture interferes with the results, we included a control group with only P. aeruginosa and the MRS medium. The same experiment was conducted using a physiological solution instead of the BHI medium and using the MRS as control. The obtained results showed no interference in P. aeruginosa growth with the medium used to prepare the CFSM, as shown in Figures 1A, B. The four CFSMs analysed presented antibacterial activity against both tested strains of P. aeruginosa. A reduction in the number of CFU/mL of pathogenic cells was observed when they were faced with the CFSMs when compared to the pathogens that faced physiological solution, the control group (100% P. aeruginosa growth means 1.71 × 109 ± 1.44 × 109 CFU/mL for PA 27853™ and 3.05 × 1014 ± 3.18 × 1014 for PA 9027™). The observed reduction, expressed in percentage, to PA 27853™ growth was 99.98% with CFSM from L. acidophilus, 99.47% from L. delbrueckii, 98.19% from L. plantarum, and 98.83% from L. johnsonii; and the observed reduction to PA 9027™ was 99.28% for CFSM from L. acidophilus, 99.14% from L. delbrueckii, 99.66% from L. plantarum, and 99.29% from L. johnsonii. All the samples were statistically different from the control (p < 0.001), yet similar.

FIGURE 1. Growth expressed by P. aeruginosa obtained by counting the CFU/mL in the antibacterial activity in vitro test for 24 h in the presence of Lactobacilli supernatants. The supernatants of four Lactobacilli strains were tested. (A) Graph of 100% growth of PA 2853™ (1.71 ± 1.44 × 109 CFU/mL) and the CFU/mL of PA in the presence of different Lactobacillus supernatant. (B) Graph of 100% growth of PA 9027™ (3.05 ± 3.18 × 1014 CFU/mL) and the CFU/mL of PA in the presence of different CFSMs. Different capital letters refer to significant differences (p < 0.05) between treatments and the control group by ANOVA followed by Tukey’s post hoc test. Different lowercase letters indicate statistically significant differences (p < 0.05) between the tested treatments by ANOVA followed by Tukey’s post hoc test.
The inhibition of P. aeruginosa growth was evaluated in the presence of different percentages of CFSM in the four studied strains at 16 h. An absorbance (OD600nm) of 0.04 or less means that there was no growth or that it was very low. This result obtained for PA 9027™ with 18%, 20%, 46%, and 50% of L. acidophilus, L. delbrueckii CIDCA 133, L. plantarum, and L. johnsonii CFSM, respectively, is shown in Figure 2A. For PA 27853™, 22%, 22%, 48%, and 54% of L. acidophilus, L. delbrueckii CIDCA 133, L. plantarum, and L. johnsonii CFSM, respectively, showed an absorbance (OD600nm) of 0.04 (Figure 2B). These results demonstrate the inhibitory power of different CFSMs with respect to P. aeruginosa growth.
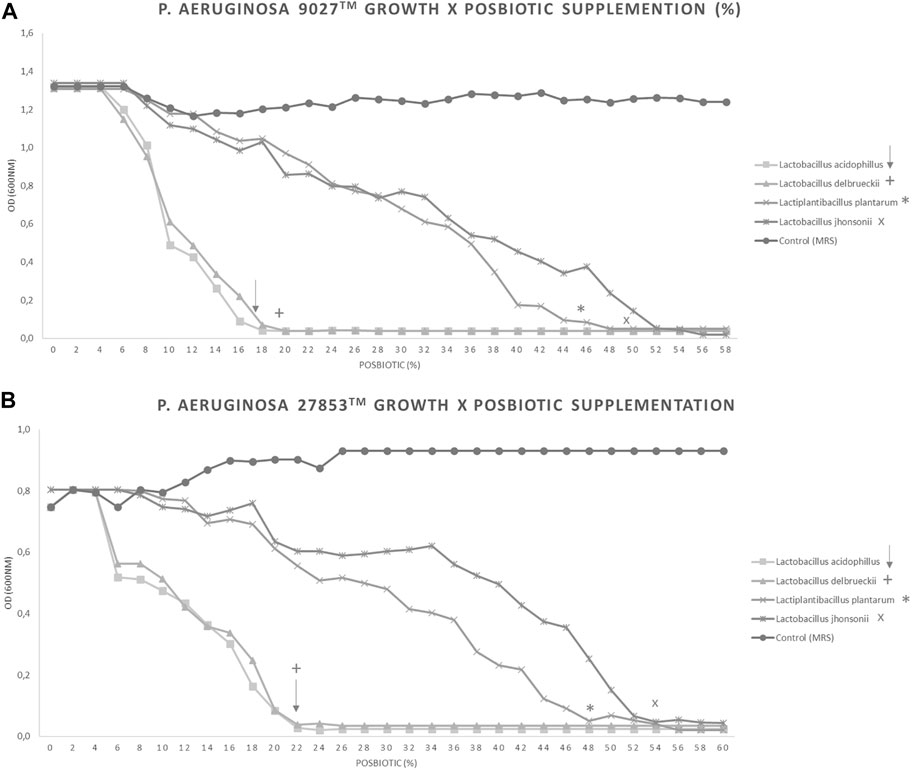
FIGURE 2. . Different percentages of CFSM supplementation can inhibit the growth of two Pseudomonas aeruginosa strains. Increasing amounts of each CFSM were added to the medium containing P. aeruginosa. CFSMs tested against PA 9027™ (A) and PA 27853™ (B). Symbols show the percentage of total inhibition of growth achieved: ↓ CFSM from L. acidophilus, + CFSM from L. delbrueckii, * CFSM from L. plantarum, and × CFSM from L. johnsonii.
The tested CFSMs could inhibit both strains of PA in an inhibition zone ranging between 3.00 and 7.00 mm, as shown in Table 3. Considering the maximum concentration tested (300 μL = 3×), our results demonstrated that the highest antibacterial activity could be attributed to L. acidophilus CFSM, which showed inhibition zones of 5.67 ± 1.15 mm and 7.00 ± 2.64 mm for PA 9027™ and PA 27853™, respectively. Although L. johnsonii CFSM was the postbiotic with a lower inhibition zone, 3.00 ± 1.00 mm and 3.33 ± 1.53 mm for PA 9027™ and 27853™, respectively (Table 3), the inhibition zone was significant.
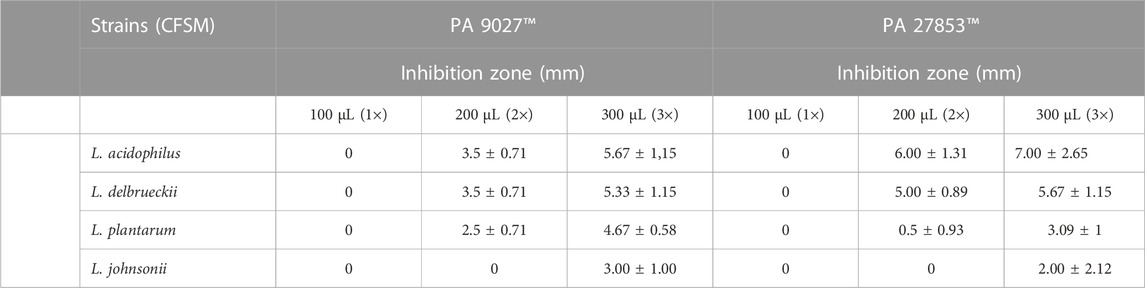
TABLE 3. Antibacterial activity at different concentrations of CFSM against two Pseudomonas aeruginosa strains
To confirm how the CFSM of different Lactobacillaceae could influence the extracellular matrix of P. aeruginosa biofilms, the total biomass was quantified by colourimetric assay using crystal violet. This assay is used as an indicator of cellular adhesion, either for viable or non-viable attached cells (Kouidhi et al., 2010). As PA 27853™ and PA 9027™ have a high ability to produce biofilms, their ability to decrease biofilm formation was tested in three different conditions when assessed at the MIC concentration of CFSM: 1) the initial cell attachment by planktonic cells (pre-coated), 2) co-incubation (planktonic cells associated with CFSM), and 3) on preformed (24 h) biofilms. These assays indicate a positive effect on biofilm inhibition for all CFSMs that were tested for PA 27853™ or PA 9027™. For PA 27853™, the high percentage of inhibition in a pre-coated approach was represented by the CFSM from L. acidophilus (63.457% ± 15.746%), which showed statistical differences (p < 0.05) from the other CFSM (44.742% ± 8.991% for L. delbrueckii, 35.750% ± 13.920% for L. plantarum, and 42.370% ± 9.439% for L. johnsonii) and the control (Figure 3A). The co-incubation experiment exposed over 50% biofilm inhibition by the CFSM of L. delbrueckii (64.352% ± 3.116%), L. plantarum (71.705% ± 2.074%), and L. johnsonii (57.154% ± 2.821%); nevertheless, L. acidophilus showed 34.768% ± 3.537% of biofilm inhibition (Figure 3B). Preformed biofilms were significantly reduced in the presence of CFSM from L. acidophilus (88.070% ± 10.652%) and L. plantarum (88.841% ± 9.304%). Although CFSM from L. delbrueckii (59.140% ± 13.096%) and L. johnsonii (45.822% ± 18.181%) had a lower inhibition of preformed biofilm, it was still significant (Figure 3C). Regarding the crystal violet results of PA 9027™, in the three studied approaches, it was possible to observe that the L. johnsonii CFSM had a high percentage of biofilm inhibition, that is, 84.925% ± 8.701%, 84.872% ± 3.510%, and 75.509% ± 13.227% for pre-coated, co-incubated, and preformed approaches, respectively. Although the other CFSM studies presented a lower percentage of inhibition, they still had a high and significant inhibition rate of up to 62% in the three methodologies used (Figures 3D–F).
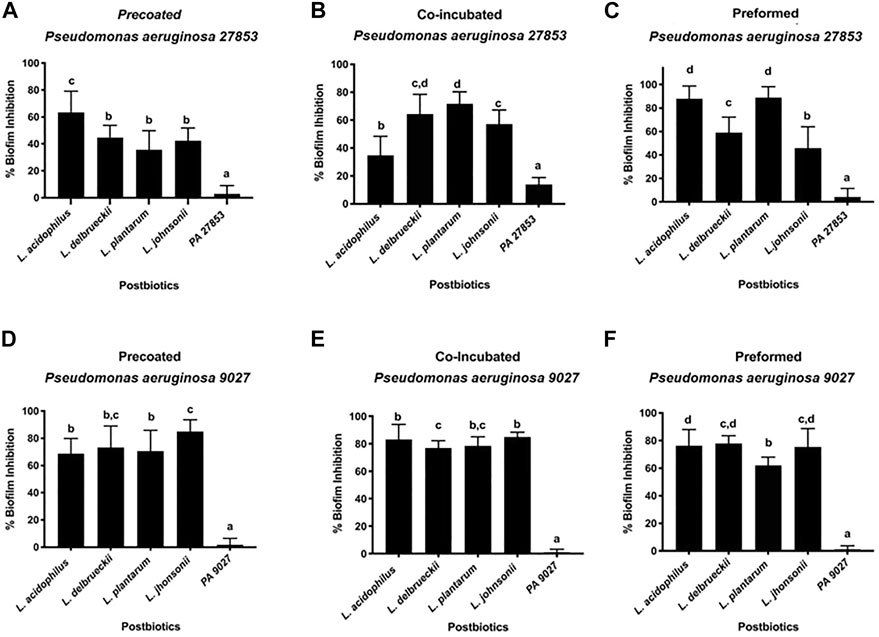
FIGURE 3. Effect of different CFSMs expressed as a percentage of inhibition of biofilm formation, on pre-coated (A, D), co-incubated (B, E), and preformed (C, F) biofilms on two Pseudomonas aeruginosa strains (27853™ - A–C and 9027™ D–F).
The biofilm was also evaluated by SEM analysis, in which either PA 27853™ or PA 9027™ showed strong adherence on the glass slides and a compact island-like biofilm covered by a large amount of dense white network-like structure. By this technique, it is possible to qualitatively verify the reduction in the number of PA cells and also a reduction in the cell’s compaction and a drop in or the absence of the dense white network-like structure, which represents the inhibitory effect of the CFSM in PA biofilm formation (Figure 4) when the control biofilm is compared with the biofilm that is exposed to the CFSM.
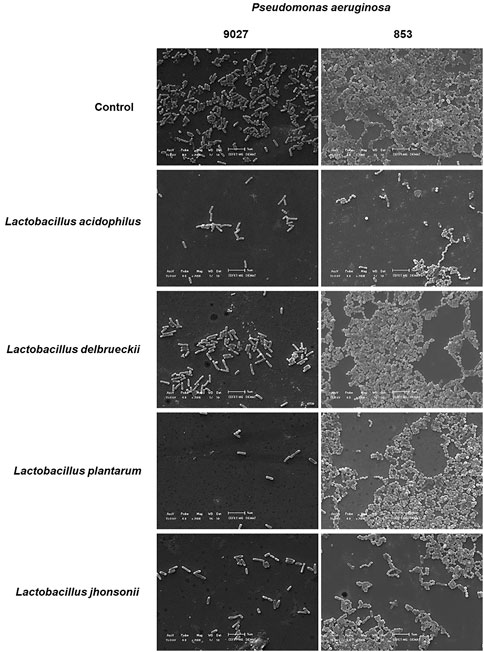
FIGURE 4. SEM of biofilm formation and inhibition in vitro assay. The images of the control groups of PA 27853™ and 9027™ in the BHI medium (sucrose, 10%) present numerous bacterial cells and biofilm formation. The magnification used for the image was ×2,000 using a JEOL JS-5600 scanning electron microscope (JEOL United States, Inc., Peabody, MA, United States) at the Characterisation Laboratory, Materials Engineering Department, Centro Federal de Educação Tecnológica de Minas Gerais (CEFET-MG). It was possible to observe a reduction in the number of PA cells and their aggregation in the CFSM treatments when compared to the respective controls.
The correlation between the Crystal Violet and MTT assay results provides a better understanding of the influence of the different CFSMs tested on biofilm inhibition. The MTT assay is an indicator of cell viability (Krom et al., 2007) because it describes the attached cells' metabolic status. Thus, the MTT assay is performed to confirm the viability of cells present in the different approaches in biofilm formation. The experimental controls, PA 27853™ and PA 9027™, showed increased metabolic activity, as expected, with percentages of up to 80% for both pathogenic strains. The MTT results for PA 27853™ showed cell viability of approximately 55% for all CFSMs tested in the pre-coated biofilm, and there were no significant differences between the different CFSMs; nevertheless, there were substantial differences with the control (Figure 5A). In the co-incubated condition, 33.490% ± 10.867% cell viability was observed in the formed biofilm in the presence of the CFSM for L. delbrueckii, 33.663% ± 10.867% for L. plantarum, 47.836% ± 13.470% for L. acidophilus, and 48.320% ± 12.820% for L. johnsonii, statistically differentiating these from the positive control, which showed a cell viability of approximately 90% (Figure 5B). In the preformed biofilm treated with postbiotics from L. acidophilus, L. delbrueckii, and L. johnsonii, this pathogenic strain presented the lowest cell viability of 5.252% ± 2.735%, 4.399% ± 2.879%, and 9.420% ± 8.145%, respectively. Even though the CFSM from L. johnsonii manifested the highest cell viability (45.904% ± 13.454%), it was still statistically different from the control (Figure 5C).
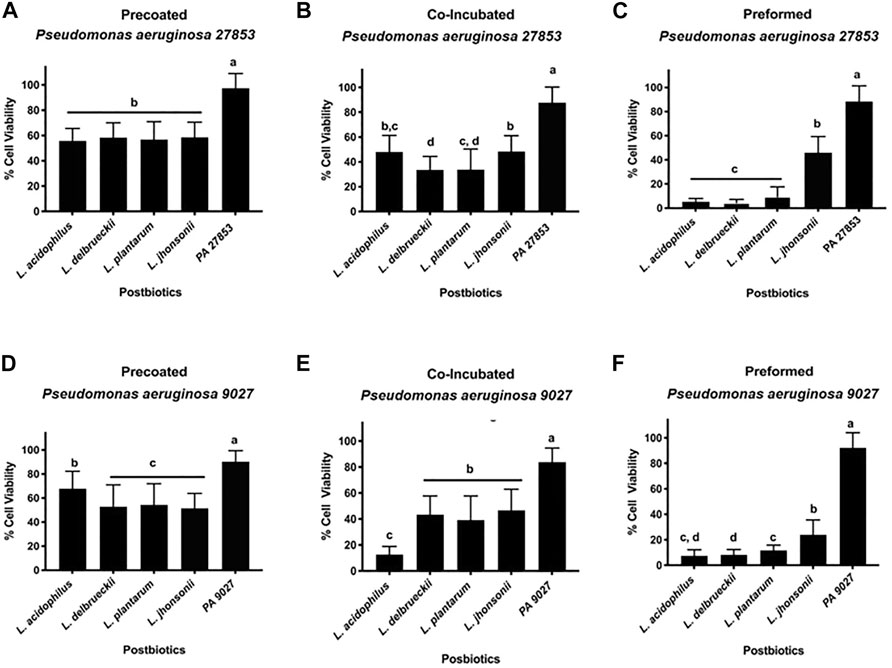
FIGURE 5. (A–F) Effect of CFSMs on the metabolic activity of pre-coated (A, D), co-incubated (B, E) and preformed (C, F) biofilm cells of Pseudomonas aeruginosa 27853™ (A–C) and 9027™ (D–F) incubated for 24 h was determined by MTT performance. Results show that the metabolic activity of CFSM in treated cells was reduced when compared to the untreated control. Different letters (a, b, c, and d) indicate statistically significant differences (p < 0.05) by ANOVA followed by Tukey’s post hoc test.
The cell viability presented with PA 9027™ in the pre-coated biofilm was approximately 50% for the CFSM of L. delbrueckii, L. plantarum, and L. johnsonii without statistical differences between them (p > 0.05). In this approach, L. acidophilus was the postbiotic that showed the highest cell viability (67.715% ± 14.487%); however, it was lower than that of the control, which presented a cell viability of 88.307% ± 12.906%, with a statistical difference of p < 0.05 (Figure 5D). Contrary to this result, for the co-incubation conditions, the CFSM from L. acidophilus presented the lowest cell viability (12.622% ± 6.225%) being the only treatment that was statistically different from the other treatments that showed a cell viability between 39% and 46%; however, these treatments showed statistical differences when compared with the control group (83.771% ± 10.825%) (Figure 5E). Finally, as shown with PA 27853™ for the preformed biofilm, L. johnsonii was the CFSM that presented a high cell viability (23.871% ± 11.768%) when compared with the other CFSMs, which showed a cell viability of approximately 10% (Figure 5F).
Postbiotics have gained more attention in the last decade due to their beneficial actions on the host without the adverse risk of inducing bacteraemia in immunocompromised patients from the delivery of live cells (Gao et al., 2019). Consequently, the works of several research groups have produced satisfactory results for this biotic fraction.
Pseudomonas aeruginosa is one of the most significant bacteria that can cause enormous burden to public health, having the capacity to adapt its genome and physiology throughout chronic infections and develop rapid antibiotic resistance; this pathogen has been listed in the global priority pathogens list by the World Health Organization (2017) and is considered an opportunistic pathogen (Cai et al., 2015). Controlling the growth of pathogens is as crucial as their elimination. Interference in their pathogenic mechanisms to regulate the production of bacterial virulence, such as in biofilm formation through the attenuation of the bacterial QS signalling system, by QS target agents (Jakobsen et al., 2012; Chen et al., 2019) is a necessary approach to delay the development of their resistance (LaSarre and Federle, 2013). For this reason, the present study was conducted to elucidate the antibacterial and antibiofilm potential of four supernatants, free of live cells, produced by four LAB against P. aeruginosa, one of the primary pathogens that effect infection of skin burns, delaying the healing process. Probiotics isolated from different niches display antibacterial effects against gram-negative and gram-positive microorganisms. The interest of researchers in studying the action of various cell-free supernatant probiotics is increasing due to the antibacterial properties assigned to the organic acids produced by these, which reduce the pH of the medium where these grow. In addition, it has been reported that the bioactive compounds released by probiotics, such as bacteriocins and hydrogen peroxide, are responsible for their antimicrobial properties (Noordiana, 2013; Yang et al., 2014; Drider et al., 2016). Thus, several research studies support the potential use of CFSMs and their bioactive compounds as antimicrobials against different pathogens.
Currently, “postbiotics” refer to soluble components with biological activity that could be a safer alternative to whole bacteria (Tsilingiri et al., 2012). The interest in postbiotics has increased in the last 5 years due to their beneficial actions on the host without the adverse risk of inducing bacteraemia, especially in immunocompromised patients and children, due to their capacity to deliver metabolites deprived of live cells (Gao et al., 2019).
The antimicrobial activity of drugs is usually assessed by the determination of the MIC and MBC of the drug in vitro after overnight aerobic incubation. Levison (2004) reported that the MBC is usually the same as the MIC for bactericidal drugs and generally not more than four-fold higher. On the contrary, the MBC of bacteriostatic medicines is multifold times more elevated than their MICs.
Considering the MIC and MBC results obtained for P. aeruginosa, it has been possible to show the bactericidal effect of CFSM from L. acidophilus and L. johnsonii. The other two CFSMs showed a bacteriostatic impact on PA 27853™. When these analyses were performed on PA 9027™, it was possible to attribute the bactericidal effect to the CFSM for L. acidophilus and the bacteriostatic effect to the rest of the CFSMs studied. Thus, all of the CFSMs of the tested Lactobacillaceae strains are potential agents against both P. aeruginosa strains. Our results are in accordance with the study by Saadatzadeh et al. (2013), who reported the powerful inhibitory effect of a cell-free extract of L. casei on P. aeruginosa and other pathogens that they tested, evidencing the anti-pathogenic impact of this biotic.
The antimicrobial susceptibility of PA was evaluated using agar overlay interference tests. It was shown that all Lactobacillaceae grown for 24 h on MRS medium formed a supernatant that exhibited inhibitory activity in the highest concentration tested (3×) for P. aeruginosa. It is essential to highlight that the CFSM of L. acidophilus showed an inhibition zone at a lower concentration for both P. aeruginosa 9027™ and 25853™, denoting the bactericidal effect of this CFSM. Meanwhile, L. delbrueckii presented bacteriostatic behaviour against both the tested strains of PA, showing an inhibition zone at the two highest concentrations tested. The CFSM of L. plantarum and L. johnsonii also showed an inhibition zone at the highest concentration tested. This antibacterial property of the CFSMs might be due to the presence of different organic acids in their composition since they have an acidic pH (of 3.70 for L. acidophilus, 4.2 for L. delbrueckii, 4.7 for L. johnsonii, and 4.7 for L. plantarum), and this antibacterial evidence can also be attributed to ethyl alcohol, short-chain fatty acids such as acetic acid (Bilkova et al., 2011), lactic acid, bacteriocins, hydrogen peroxide, and surfactants, which are some of the secondary metabolites produced by Lactobacilli that have antimicrobial activity (Noordiana, 2013; Yang et al., 2014; Drider et al., 2016; Melo et al., 2016; Plaza-Diaz et al., 2019). Thus, postbiotic compounds can inhibit the growth of several bacterial pathogens by producing antimetabolites. The presence and characterisation of these compounds require future studying such as by using chromatography.
To evaluate if the principal inhibitory effect is due to the presence of acids, the acidic pH of the CFSMs was neutralised, and their antibacterial activity was observed against both the tested P. aeruginosa strains. Our results agree with the results of Shaimaa (2019), who analysed the cell-free supernatant of L. acidophilus ATCC 4356, by agar well diffusion and showed its high antibacterial potential against most of the isolated P. aeruginosa strains; the same was demonstrated by Shokri-Kojori et al. (2018) in their study that analysed different CFSMs of Lactobacillus against P. aeruginosa. Both these works have demonstrated that inhibitory activity against P. aeruginosa strains was not observed after neutralising the acidic pH of the Lactobacillus supernatant. This event could be attributed to the production of antibacterial organic acid molecules, such as lactic, acetic, and formic acids, or possibly to bacteriocins that were active only under acidic conditions (Shokri-Kojori et al., 2018). This affirmation is in accordance with the result of Wala’a and Nibras (2013), where bacteriocins from L. acidophilus were found to exhibit activity against Serratia marcescens at a pH of 4.7, with half of the activity being lost at pH 8, and then the whole activity being lost at other pH values. Another study of L. casei CFSM showed anti-Shigella activity in vitro, which was abolished after neutralisation (Mirnejad et al., 2013). We can conclude that maintaining natural conditions of the CFSM is imporant, at least with regard to antibacterial activity; also, this behaviour is not specific for one specific pathogen, as reported by many researchers.
To better understand the achieved results, we investigated the antibacterial potential of CFSMs in the growth kinetics of both P. aeruginosa strains and observed that a lower percentage of CFSM was used for L. acidophilus and L. delbrueckii CIDCA 133 (18 or 22% and 20 or 22%, respectively) and a higher percentage for L. plantarum and L. johnsonii (46 or 48% and 50 or 54%, respectively). At these obtained percentages, the complete inhibition of growth of either PA 9027™ or PA 27853™ was achieved. These results follow the antimicrobial activity, where we observed the best effect (higher inhibition zones) with the CFSM of L. acidophilus and in decreasing effectivity with the CFSM of L. delbrueckii CIDCA 133, L. plantarum and, lastly, L. johnsonii. Finally, we analysed the effect of each CFSM against planktonic cells, expressed as the percentage of growth reduction, after analysing the CFU/mL. Independent of the CFSM and the PA tested, we observed a significant reduction (more than 90%) in PA growth. Together, these experiments enable us to infer the anti-pseudomonas activity of all these CFSMs.
Our study has demonstrated that the CFSMs from all tested strains have a strong and moderate inhibition of biofilm formation for all the three tested approaches (co-incubation, pre-coated, and preformed) either to PA 9027™ or PA 27853™.
Many virulent factors are correlated with a pathogen’s initial surface adhesion following the production of extracellular polymeric substances that lead to biofilm formation; this is a phenomenon of microorganisms that results in a persistent microbial mass resistant to antimicrobial agents and is related to approximately 80% of human bacterial infections (Limoli et al., 2015). In biofilms, the microbial population is attached to the surface and maintained by the liberation of an adhesive and protective matrix that provides tolerance and resistance to antimicrobials, which has important implications in healthcare (De Vincenti et al., 2018), especially in burn infections by delaying the healing process. An agent is considered antibacterial if it can achieve a minimum of 3-log/99.9% reduction in bacterial growth (French et al., 2006; Levison and Levison, 2009; Balouiri et al., 2016). Based on our results for planktonic cells, it is possible to infer that the CFSMs of L. acidophilus and L. delbrueckii significantly reduced the metabolic activity of PA 27853™, with an average measured 4.5-log10 reduction. These results follow the antibacterial results obtained in the present study, showing the powerful effect of these CFSM against PA 27853™. The same was not observed for PA 9027™ since all CFSMs from the investigated Lactobacillaceae species demonstrated a reduction in the metabolic activity of P. aeruginosa, but only by 2-log10. These results are partially in accordance with the study by Jeyanathan et al. (2021) on whole cells of L. acidophilus, L. plantarum, and L. reuteri that showed a minimal impact on PA growth; however, the CFSMs of L. reuteri and L. plantarum did reveal a reduction in PA growth. Together, this shows the importance of testing each CFSM against each pathogen's microorganism since the effect of the postbiotics is strain dependent, as described for the probiotics.
The CFSMs of different LAB could exhibit other biological activities. Proteins and released exopolysaccharides (EPSs) are some of the biologically active compounds in CFSM (Wang et al., 2020) The transcriptome analyses of the CFSM of L. acidophilus A4 showed that the EPSs present exhibit antibiofilm activity by affecting genes that participate in chemotaxis (cheY), in Curli amyloid fibre production, which is the component of the extracellular matrix produced by Enterobacteriaceae during biofilm formation of enterohaemorrhagic E. coli O157 (Kim et al., 2009). The cheY gene is also associated with P. aeruginosa biofilm (Barken et al., 2008). The capacity of EPS to inhibit the initial autoaggregation and cell attachment of bacterial cells has also been reported to weaken cell surface modifications or reduce cell-to-cell surface interactions (Kanmani et al., 2013). Based on these reports, we quantified the difference in the total biomass of the biofilms and the metabolic activity present in the formed biomass since P. aeruginosa strains are biofilm producers, as shown in our work (positive control, which showed the highest biofilm formation) and according to other reports (Jabalameli et al., 2012; Shokri-Kojori et al., 2018). The biofilm formed by P. aeruginosa in the presence of CFSM exhibited a significant reduction in biomass when compared to the control group for PA 9027™ and PA 27853™. The CFSMs for all analysed Lactobacillaceae showed anti-adhesion activity against PA, which could be observed in both the co-incubation and pre-coated approaches, with the metabolic activity at approximately 50%. In the preformed approach, the anti-adhesion activity was not remarkable, and the cell viability was extremely low (less than 20%), except for L. johnsonii, which showed cell viability of approximately 40%. Considering the behaviour of CFSMs against PA 9027™, our results show the highest biofilm inhibition percentage of all approaches tested for all CFSMs (between 70% and 80%).
Regarding the percentage of cell viability, our results show values lower than 50%, being statistically different from the positive control. In the preformed approach, it was possible to observe the most insufficient metabolic activity for all tested CFSMs. For the CFSM of L. acidophilus, our biofilm inhibition results are in accordance with Shaimaa (2019), who studied the antibacterial and anti-biofilm effect of the CFSM of L. acidophilus ATCC 4356 against several P. aeruginosa strains, showing that these postbiotics significantly inhibited 68.52% of biofilm formation and removed already preformed biofilms with 43.8% activity. SEM verified the reduction in mass biofilm formation by the CFSM.
Thus, the results of the present work, together with other recent reports in the literature regarding the effect of these probiotics against different pathogens, specifically against P. aeruginosa, open new perspectives for biological control agents and could be an exciting approach to replace antibiotics or complement their reduced usage, making these good candidates to overcome the growing challenges of hospital-acquired infections due to this pathogen and other pathogenic microorganisms.
Further studies should address the isolation and characterisation of the bioactive molecules present in the CFSMs studied and the action mechanisms of these postbiotic elements on specific target cells.
The raw data supporting the conclusion of this article will be made available by the authors, without undue reservation.
Conceptualisation: PM-A and MD. Methodology: PM-A, MD, AT-C, and JB. Formal analysis and investigation: PM-A and MD. Original draft writing and manuscript preparation: PM-A. Manuscript reviewing and editing: MD, AN, EN, DK, and PM-A. All authors contributed to the article and have approved the submitted version.
This research was funded by Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG)—grant number APQ-00766-21, Brasil, and by the Rede Mineira de Engenharia de Tecidos e Terapia Celular, FAPEMIG—grant number RED-00570/16.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Aguilar-Toalá, J. E., Garcia-Varela, R., Garcia, H. S., Mata-Haro, V., González-Córdova, A. F., Vallejo-Cordoba, B., et al. (2018). Postbiotics: An evolving term within the functional foods field. Trends Food Sci. Technol. 75, 105–114. doi:10.1016/j.tifs.2018.03.009
Atiyeh, B. S., Gunn, S. W., and Hayek, S. N. (2005). State of the art in burn treatment. World J. Surg. 29, 131–148. doi:10.1007/s00268-004-1082-2
Atkinson, S. (2007). N-acylhomoserine lactones, quorum sensing, and bioflm development in gram-negative bacteria. Wymondham Horiz. Biosci., 95–122.
Balouiri, M., Sadiki, M., and Ibnsouda, S. K. (2016). Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 6, 71–79. doi:10.1016/j.jpha.2015.11.005
Barken, K. B., Pamp, S. J., Yang, L., Gjermansen, M., Bertrand, J. J., Klausen, M., et al. (2008). Roles of type IV pili, flagellum-mediated motility and extracellular DNA in the formation of mature multicellular structures in Pseudomonas aeruginosa biofilms. Environ. Microbiol. 10, 2331–2343. doi:10.1111/j.1462-2920.2008.01658.x
Barros, C. P., Guimarães, J. T., Esmerino, E. A., Duarte, M. C. K., Silva, M. C., Silva, R., et al. (2020). Paraprobiotics and postbiotics: Concepts and potential applications in dairy products. Curr. Opin. Food Sci. 32, 1–8. doi:10.1016/j.cofs.2019.12.003
Batista, V. L., De Jesus, L. C. L., Tavares, L. M., Barroso, F. L. A., Fernandesda, L. J. S., Freitasdos, A. S., et al. (2022). Paraprobiotics and postbiotics of Lactobacillus delbrueckii CIDCA 133 mitigate 5-FU-induced intestinal inflammation. Microorganisms 10, 1418. doi:10.3390/microorganisms10071418
Bayoumi, M. A., and Griffiths, M. W. (2012). In vitro inhibition of expression of virulence genes responsible for colonization and systemic spread of enteric pathogens using Bifidobacterium bifidum secreted molecules. Int. J. Food Microbiol. 156 (3), 255–263. doi:10.1016/j.ijfoodmicro.2012.03.034
Bilkova, A., Kinova Sepova, H., Bukovsky, M., and Bezakova, L. (2011). Antibacterial potential of lactobacilli isolated from a lamb. Vet. Med. (Praha) 56, 319–324. doi:10.17221/1583-VETMED
Borges, S., Silva, J., and Teixeira, P. (2014). The role of lactobacilli and probiotics in maintaining vaginal health. Arch. Gynecol. Obstet. 289, 479–489. doi:10.1007/s00404-013-3064-9
Cai, Z., Liu, Y., Chen, Y., Yam, J., Chew, S., Chua, S., et al. (2015). RpoN regulates virulence factors of Pseudomonas aeruginosa via modulating the PqsR quorum sensing regulator. Int. J. Mol. Sci. 16, 28311–28319. doi:10.3390/ijms161226103
Chen, Y., Chen, S., Li, K., Zhang, Y., Huang, X., Li, T., et al. (2019). Overdosage of balanced protein complexes reduces proliferation rate in aneuploid cells. Cell Syst. 9, 129–142.e5. doi:10.1016/j.cels.2019.06.007
Collado, J., Kalemanov, M., Campelo, F., Bourgoint, C., Thomas, F., Loewith, R., et al. (2019). Tricalbin-mediated contact sites control ER curvature to maintain plasma membrane integrity. Dev. Cell 51, 476–487.e7. doi:10.1016/j.devcel.2019.10.018
Cruz, J. M., Domínguez, J. M., Domínguez, H., and Parajó, J. C. (2001). Antioxidant and antimicrobial effects of extracts from hydrolysates of lignocellulosic materials. J. Agric. Food Chem. 49, 2459–2464. doi:10.1021/jf001237h
Cuevas-González, P. F., Liceaga, A. M., and Aguilar-Toalá, J. E. (2020). Postbiotics and paraprobiotics: From concepts to applications. Food Res. Int. 136, 109502. doi:10.1016/j.foodres.2020.109502
De Vincenti, L., Glasenapp, Y., Cattò, C., Villa, F., Cappitelli, F., and Papenbrock, J. (2018). Hindering the formation and promoting the dispersion of medical biofilms: Non-lethal effects of seagrass extracts. BMC Complement. Altern. Med. 18, 168. doi:10.1186/s12906-018-2232-7
Dias-Souza, M. (2013). Evaluation of Antimicrobial and Anti-biofilm activities of Anacardium occidentale stem bark extract. J. Nat. Prod. 6, 198–205.
Drider, D., Bendali, F., Naghmouchi, K., and Chikindas, M. L. (2016). Bacteriocins: Not only antibacterial agents. Probiotics Antimicrob. Proteins 8, 177–182. doi:10.1007/s12602-016-9223-0
Driscoll, J. A., Brody, S. L., and Kollef, M. H. (2007). The epidemiology, pathogenesis and treatment of Pseudomonas aeruginosa infections. Drugs 67, 351–368. doi:10.2165/00003495-200767030-00003
French, S. D., Cameron, M., Walker, B. F., Reggars, J. W., and Esterman, A. J. (2006). Superficial heat or cold for low back pain. Cochrane Database Syst. Rev. doi:10.1002/14651858.CD004750.pub2
Gao, J., Li, Y., Wan, Y., Hu, T., Liu, L., Yang, S., et al. (2019). A novel postbiotic from Lactobacillus rhamnosus GG with a beneficial effect on intestinal barrier function. Front. Microbiol. 10. doi:10.3389/fmicb.2019.00477
Gjødsbøl, K., Christensen, J. J., Karlsmark, T., Jørgensen, B., Klein, B. M., and Krogfelt, K. A. (2006). Multiple bacterial species reside in chronic wounds: A longitudinal study. Int. Wound J. 3, 225–231. doi:10.1111/j.1742-481X.2006.00159.x
Gudiña, E. J., Rocha, V., Teixeira, J. A., and Rodrigues, L. R. (2010). Antimicrobial and antiadhesive properties of a biosurfactant isolated from Lactobacillus paracasei ssp. paracasei A20. Lett. Appl. Microbiol. 50, 419–424. doi:10.1111/j.1472-765X.2010.02818.x
Hall-Stoodley, L., Costerton, J. W., and Stoodley, P. (2004). Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2, 95–108. doi:10.1038/nrmicro821
Jabalameli, F., Mirsalehian, A., Khoramian, B., Aligholi, M., Khoramrooz, S. S., Asadollahi, P., et al. (2012). Evaluation of biofilm production and characterization of genes encoding type III secretion system among Pseudomonas aeruginosa isolated from burn patients. Burns 38, 1192–1197. doi:10.1016/j.burns.2012.07.030
Jadhav, S., Shah, R., Bhave, M., and Palombo, E. A. (2013). Inhibitory activity of yarrow essential oil on Listeria planktonic cells and biofilms. Food control. 29, 125–130. doi:10.1016/j.foodcont.2012.05.071
Jakobsen, T. H., van Gennip, M., Phipps, R. K., Shanmugham, M. S., Christensen, L. D., Alhede, M., et al. (2012). Ajoene, a sulfur-rich molecule from garlic, inhibits genes controlled by quorum sensing. Antimicrob. Agents Chemother. 56, 2314–2325. doi:10.1128/AAC.05919-11
Jeyanathan, A., Ramalhete, R., Blunn, G., Gibbs, H., Pumilia, C. A., Meckmongkol, T., et al. (2021). Lactobacillus cell-free supernatant as a novel bioagent and biosurfactant against Pseudomonas aeruginosa in the prevention and treatment of orthopedic implant infection. J. Biomed. Mater Res. B Appl. Biomater. 109, 1634–1643. doi:10.1002/jbm.b.34821
Kanmani, P., Suganya, K., kumar, R. S., Yuvaraj, N., Pattukumar, V., Paari, K. A., et al. (2013). Synthesis and functional characterization of antibiofilm exopolysaccharide produced by Enterococcus faecium MC13 isolated from the gut of fish. Appl. Biochem. Biotechnol. 169, 1001–1015. doi:10.1007/s12010-012-0074-1
Kim, Y., oh, S., and Kim, S. H. (2009). Released exopolysaccharide (r-EPS) produced from probiotic bacteria reduce biofilm formation of enterohemorrhagic Escherichia coli O157:H7. Biochem. Biophys. Res. Commun. 379, 324–329. doi:10.1016/j.bbrc.2008.12.053
Kouidhi, B., Zmantar, T., and Bakhrouf, A. (2010). Anti-cariogenic and anti-biofilms activity of Tunisian propolis extract and its potential protective effect against cancer cells proliferation. Anaerobe 16, 566–571. doi:10.1016/j.anaerobe.2010.09.005
Krom, B. P., Cohen, J. B., McElhaney Feser, G. E., and Cihlar, R. L. (2007). Optimized candidal biofilm microtiter assay. J. Microbiol. Methods 68, 421–423. doi:10.1016/j.mimet.2006.08.003
LaSarre, B., and Federle, M. J. (2013). Exploiting quorum sensing to confuse bacterial pathogens. Microbiol. Mol. Biol. Rev. 77, 73–111. doi:10.1128/MMBR.00046-12
Levison, M. E., and Levison, J. H. (2009). Pharmacokinetics and pharmacodynamics of antibacterial agents. Infect. Dis. Clin. North Am. 23, 791–815. doi:10.1016/J.IDC.2009.06.008
Levison, M. E. (2004). Pharmacodynamics of antimicrobial drugs. Infect. Dis. Clin. North Am. 18, 451–465. doi:10.1016/j.idc.2004.04.012
Limoli, D. H., Jones, C. J., and Wozniak, D. J. (2015). Bacterial extracellular polysaccharides in biofilm formation and function. Microbiol. Spectr. 3. doi:10.1128/microbiolspec.MB-0011-2014
Lin, X., Chen, X., Chen, Y., Jiang, W., and Chen, H. (2015). The effect of five probiotic lactobacilli strains on the growth and biofilm formation of Streptococcus mutans. Oral Dis. 21. doi:10.1111/odi.12257
Lynch, J., Zhanel, G., and Clark, N. (2017). Emergence of antimicrobial resistance among Pseudomonas aeruginosa: Implications for therapy. Semin. Respir. Crit. Care Med. 38, 326–345. doi:10.1055/s-0037-1602583
Marinho, A. R., Martins, P. D., Ditmer, E. M., d’Azevedo, P. A., Frazzon, J., Sand, S. T. V. D., et al. (2013). Biofilm formation on polystyrene under different temperatures by antibiotic resistant Enterococcus faecalis and Enterococcus faecium isolated from food. Braz. J. Microbiol. 44, 423–426. doi:10.1590/S1517-83822013005000045
Marttinen, A. M., Haukioja, A. L., Keskin, M., and Söderling, E. M. (2013). Effects of Lactobacillus reuteri PTA 5289 and L. Paracasei DSMZ16671 on the adhesion and biofilm formation of Streptococcus mutans. Curr. Microbiol. 67, 193–199. doi:10.1007/s00284-013-0352-3
Melo, T. A., Dos Santos, T. F., de Almeida, M. E., Junior, L. A. G. F., Andrade, E. F., Rezende, R. P., et al. (2016). Inhibition of Staphylococcus aureus biofilm by Lactobacillus isolated from fine cocoa. BMC Microbiol. 16, 250. doi:10.1186/s12866-016-0871-8
Mihai, M. M., Preda, M., Lungu, I., Gestal, M. C., Popa, M. I., and Holban, A. M. (2018). Nanocoatings for chronic wound repair—modulation of microbial colonization and biofilm formation. Int. J. Mol. Sci. 19, 1179. doi:10.3390/ijms19041179
Mirnejad, R., Vahdati, A. R., Rashidiani, J., Erfani, M., and Piranfar, V. (2013). The antimicrobial effect of Lactobacillus casei culture supernatant against multiple drug resistant clinical isolates of Shigella sonnei and Shigella flexneri in vitro. Iran. Red. Crescent Med. J. 15. doi:10.5812/ircmj.7454
Moradi, M., Mardani, K., and Tajik, H. (2019). Characterization and application of postbiotics of Lactobacillus spp. on Listeria monocytogenes in vitro and in food models. LWT 111, 457–464. doi:10.1016/j.lwt.2019.05.072
Noordiana, N. (2013). Antibacterial agents produced by lactic acid bacteria isolated from Threadfin Salmon and Grass Shrimp. Int. Food Res. J. 20, 117–124.
Parsek, M. R., and Greenberg, E. P. (2005). Sociomicrobiology: The connections between quorum sensing and biofilms. Trends Microbiol. 13, 27–33. doi:10.1016/J.TIM.2004.11.007
Passador, L., Cook, J. M., Gambello, M. J., Rust, L., and Iglewski, B. H. (1993). Expression of Pseudomonas aeruginosa virulence genes requires cell-to-cell communication. Sci. (1979) 260, 1127–1130. doi:10.1126/science.8493556
Penesyan, A., Gillings, M., and Paulsen, I. (2015). Antibiotic Discovery: Combatting bacterial resistance in cells and in biofilm communities. Molecules 20, 5286–5298. doi:10.3390/molecules20045286
Piqué, N., Berlanga, M., and Miñana-Galbis, D. (2019). Health benefits of heat-killed (tyndallized) probiotics: An overview. Int. J. Mol. Sci. 20, 2534. doi:10.3390/ijms20102534
Plaza-Diaz, J., Ruiz-Ojeda, F. J., Gil-Campos, M., and Gil, A. (2019). Mechanisms of action of probiotics. Adv. Nutr. 10, S49–S66. doi:10.1093/advances/nmy063
Rad, A. H., Aghebati-Maleki, L., Kafil, H. S., and Abbasi, A. (2021). Molecular mechanisms of postbiotics in colorectal cancer prevention and treatment. Crit. Rev. Food Sci. Nutr. 61, 1787–1803. doi:10.1080/10408398.2020.1765310
Rajakovich, L. J., and Balskus, E. P. (2019). Metabolic functions of the human gut microbiota: The role of metalloenzymes. Nat. Prod. Rep. 36, 593–625. doi:10.1039/c8np00074c
Ravel, J., Gajer, P., Abdo, Z., Schneider, G. M., Koenig, S. S. K., McCulle, S. L., et al. (2011). Vaginal microbiome of reproductive-age women. Proc. Natl. Acad. Sci. 108, 4680–4687. doi:10.1073/pnas.1002611107
Reid, G., Younes, J. A., Van der Mei, H. C., Gloor, G. B., Knight, R., and Busscher, H. J. (2011). Microbiota restoration: Natural and supplemented recovery of human microbial communities. Nat. Rev. Microbiol. 9, 27–38. doi:10.1038/nrmicro2473
Rodrigues, L. B., Santos, L. R. dos, Tagliari, V. Z., Rizzo, N. N., Trenhago, G., Oliveira, A. P. D., et al. (2010). Quantification of biofilm production on polystyrene by Listeria, Escherichia coli and Staphylococcus aureus isolated from a poultry slaughterhouse. Braz. J. Microbiol. 41, 1082–1085. doi:10.1590/S1517-83822010000400029
Rossoni, R. D., Vellosodos, M. S., de Barros, P. P., Alvarenga, J. A., Santosdos, J. D. S. P., dos, A. C. C., et al. (2018). Inhibitory effect of probiotic Lactobacillus supernatants from the oral cavity on Streptococcus mutans biofilms. Microb. Pathog. 123, 361–367. doi:10.1016/j.micpath.2018.07.032
Ruíz, F. O., Gerbaldo, G., García, M. J., Giordano, W., Pascual, L., and Barberis, I. L. (2012). Synergistic effect between two bacteriocin-like inhibitory substances produced by lactobacilli strains with inhibitory activity for Streptococcus agalactiae. Curr. Microbiol. 64, 349–356. doi:10.1007/s00284-011-0077-0
Saadatzadeh, A., Fazeli, M. R., Jamalifar, H., and Dinarvand, R. (2013). Probiotic properties of lyophilized cell free extract of Lactobacillus casei. Jundishapur J. Nat. Pharm. Prod. 8, 131–137. doi:10.17795/jjnpp-8564
Salminen, S., Collado, M. C., Endo, A., Hill, C., Lebeer, S., Quigley, E. M. M., et al. (2021). The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 18, 649–667. doi:10.1038/s41575-021-00440-6
Schillaci, D., Arizza, V., Dayton, T., Camarda, L., and Stefano, V. D. (2008). In vitro anti-biofilm activity of Boswellia spp. oleogum resin essential oils. Lett. Appl. Microbiol. 47, 433–438. doi:10.1111/j.1472-765X.2008.02469.x
Serra, R., Grande, R., Butrico, L., Rossi, A., Settimio, U. F., Caroleo, B., et al. (2015). Chronic wound infections: The role of Pseudomonas aeruginosa and Staphylococcus aureus. Expert Rev. Anti Infect. Ther. 13, 605–613. doi:10.1586/14787210.2015.1023291
Shaimaa, E. (2019). The effect of Lactobacillus acidophilus as a probiotic against Pseudomonas aeruginosa growth and biofilm formation. Nov. Res. Microbiol. J. 3, 428–439. doi:10.21608/nrmj.2019.44951
Shokri-Kojori, E., Wang, G.-J., Wiers, C. E., Demiral, S. B., Guo, M., Kim, S. W., et al. (2018). β-Amyloid accumulation in the human brain after one night of sleep deprivation. Proc. Natl. Acad. Sci. 115, 4483–4488. doi:10.1073/pnas.1721694115
Trindade, L. M., Torres, L., Matos, I. D., Miranda, V. C., de Jesus, L. C. L., Cavalcante, G., et al. (2021). Paraprobiotic lacticaseibacillus rhamnosus protects intestinal damage in an experimental murine model of mucositis. Probiotics Antimicrob. Proteins. doi:10.1007/s12602-021-09842-z
Tsilingiri, K., Barbosa, T., Penna, G., Caprioli, F., Sonzogni, A., Viale, G., et al. (2012). Probiotic and postbiotic activity in health and disease: Comparison on a novel polarised ex-vivo organ culture model. Gut 61, 1007–1015. doi:10.1136/gutjnl-2011-300971
Wahab, A. A., and Rahman, M. M. (2013). Pseudomonas aeruginosa bacteremia secondary to acute right leg cellulitis: Case of community-acquired infection. EXCLI J. 12, 997–1000.
Wala’a, S., and Nibras, N. M. (2013). M. Partial purification and characterization of bacteriocin produced by Lactobacillus acidophilus. J. Biotech Resear Cent 7, 80–86.
Wang, W., Ma, H., Yu, H., Qin, G., Tan, Z., Wang, Y., et al. (2020). Molecules screening of Lactobacillus plantarum subsp. plantarum with potential probiotic activities for inhibiting ETEC K88 in weaned piglets. Molecules 25, 01–20. doi:10.3390/molecules25194481
World Health Organization (2017). Global priority list of antibiotic-resistant bacteria to guide research, discovery, and development of new antibiotics.
Wu, D. C., Chan, W. W., Metelitsa, A. I., Fiorillo, L., and Lin, A. N. (2011). Pseudomonas skin infection. Am. J. Clin. Dermatol 12, 157–169. doi:10.2165/11539770-000000000-00000
Wu, T., and Zhu, J. (2021). Recent development and optimization of pseudomonas aeruginosa exotoxin immunotoxins in cancer therapeutic applications. Int. Immunopharmacol. 96, 107759. doi:10.1016/j.intimp.2021.107759
Keywords: biofilm, lactic acid bacteria, postbiotics fraction (supernatant), skin pathogens, Pseudomonas aeruginosa
Citation: Drumond MM, Tapia-Costa AP, Neumann E, Nunes ÁC, Barbosa JW, Kassuha DE and Mancha-Agresti P (2023) Cell-free supernatant of probiotic bacteria exerted antibiofilm and antibacterial activities against Pseudomonas aeruginosa: A novel biotic therapy. Front. Pharmacol. 14:1152588. doi: 10.3389/fphar.2023.1152588
Received: 27 January 2023; Accepted: 22 May 2023;
Published: 15 June 2023.
Edited by:
Sunday Oyedemi, University of the Free State, South AfricaReviewed by:
Wenjing Zhao, Shenzhen Campus of Sun Yat-Sen University, ChinaCopyright © 2023 Drumond, Tapia-Costa, Neumann, Nunes, Barbosa, Kassuha and Mancha-Agresti. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mariana Martins Drumond, bW1kcnVtb25kQGNlZmV0bWcuYnI=, bW1kcnVtb25kQGdtYWlsLmNvbQ==; Pamela Mancha-Agresti, cC5tYW5jaGEuYWdyZXN0aUBnbWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.