 Xiaoyin Ye
Xiaoyin Ye Tong Zhang
Tong Zhang Han Han
Han Han- 1School of Traditional Chinese Medicine, Shanghai University of Traditional Chinese Medicine, Shanghai, China
- 2Experiment Center for Teaching and Learning, Shanghai University of Traditional Chinese Medicine, Shanghai, China
The accumulation of bile acids in the liver leads to the development of cholestasis and hepatocyte injury. Nuclear receptors control the synthesis and transport of bile acids in the liver. Among them, the farnesoid X receptor (FXR) is the most common receptor studied in treating cholestasis. The activation of this receptor can reduce the amount of bile acid synthesis and decrease the bile acid content in the liver, alleviating cholestasis. Ursodeoxycholic acid (UDCA) and obeticholic acid (OCA) have a FXR excitatory effect, but the unresponsiveness of some patients and the side effect of pruritus seriously affect the results of UDCA or OCA treatment. The activator of peroxisome proliferator-activated receptor alpha (PPARα) has emerged as a new target for controlling the synthesis and transport of bile acids during cholestasis. Moreover, the anti-inflammatory effect of PPARα can effectively reduce cholestatic liver injury, thereby improving patients’ physiological status. Here, we will focus on the function of PPARα and its involvement in the regulation of bile acid transport and metabolism. In addition, the anti-inflammatory effects of PPARα will be discussed in some detail. Finally, we will discuss the application of PPARα agonists for cholestatic liver disorders.
Introduction
Bile acids are synthesized from cholesterol in the liver, secreted in the bile, and carried to the intestines (Di Ciaula et al., 2017). In other words, bile acids are the products of cholesterol. Hepatocyte dysfunction results in bile acid accumulation in the liver and occurrence of cholestasis. Cholestasis can be caused by the mechanical blockage of the bile duct (Chai et al., 2015), gene defect (Bull and Thompson, 2018), hormonal disorders (Piechota and Jelski, 2020), and drug administration (Gijbels et al., 2019). Generally speaking, cholestasis can be categorized into two types according to the location of the bile flow disturbance: intrahepatic and extrahepatic cholestasis. Alkaline phosphatase (ALP), γ-glutamyl transpeptidase (GGT), 5′-nucleotidase, aspartate aminotransferase, and alanine aminotransferase (ALT) are used as indicators for diagnosing cholestasis (Padda et al., 2011).
Although the clinical manifestations of cholestasis are diverse, all of them have a common feature: excess bile acids accumulate in the liver and cause hepatocyte damage (Halilbasic et al., 2015; Patel and Seetharam, 2016). Meanwhile, nuclear receptors are critical components involved in the regulation of bile acid transporters. They have been used as therapeutic targets for cholestatic liver diseases. The farnesoid X receptor (FXR) is a bile acid-activated transcription factor and is essential for bile acid homeostasis (Keitel et al., 2019). FXR is mainly expressed in the liver and intestine and controls cholestasis by sensing bile acids and regulating them through negative feedback (Forman et al., 1995; Cariello et al., 2018; Ticho et al., 2019). Current treatments are all associated with FXR, including ursodeoxycholic acid (UDCA), which partly affects FXR (Li et al., 2016), and obeticholic acid (OCA), which is a direct FXR agonist. For example, FXR agonists and OCA are recommended to use in patients with primary biliary cholangitis (PBC) (Lindor et al., 2009; Gulamhusein and Hirschfield, 2020). Despite advances in cholestasis treatment, it has unwanted side effects, including pruritus, worsening liver function, headache, and anemia (D'Amato et al., 2021). In the clinic, UDCA is ineffective for most patients, and its narrow application range is only useful to patients with PBC (Leuschner et al., 2000). Meanwhile, OCA may cause pruritus in some patients, preventing further treatment (Younossi et al., 2019). Even worse, some studies also proved that FXR activation may aggravate obstructive cholestasis (Stedman et al., 2006). Therefore, expanding our knowledge of cholestatic liver injury and finding novel nuclear receptors and medical treatments to treat cholestasis without side effects are necessary. In this review, we provide an overview of the function of peroxisome proliferator-activated receptor alpha (PPARα) and its adaptive response to cholestasis. PPARα may serve as a therapeutic target for the treatment of cholestatic disorders.
Burden of cholestasis-exceed bile acid and inflammation
In the pathological process of cholestasis, bile acids accumulate in the liver due to transporter disorders. The symptom may develop into cirrhosis or liver failure and subsequently result in liver transplantation. Bile acids are considered the direct reasons for hepatocyte damage. Chenodeoxycholate is highly toxic and causes hepatocyte injury (Spivey et al., 1993). However, this burden of cholestasis is complex. We have summarized it in Table 1.

TABLE 1. The potential mechanisms of hepatocytes injury during cholestasis.
Intrahepatic cholestasis is a disease that often occurs during pregnancy and is accompanied by pruritus or elevated serum transaminases. At the same time, the bile acid content in the blood also increases. This symptom not only affects pregnant women but also causes complications for newborns, which may lead to their death (Floreani and Gervasi, 2016). Progressive familial intrahepatic cholestasis (PFIC) is a genetic disease caused by a genetic defect. Three main subtypes of PFIC have been identified. Patients show jaundice and pruritus in infancy or early childhood. This will induce a series of poor outcomes including cirrhosis and liver failure (Baker et al., 2019). Newborns usually have neonatal jaundice and even neonatal cholestasis. This may lead to abnormal liver function, liver failure, and even death (Satrom and Gourley, 2016). In conclusion, cholestasis can cause damage to the liver, leading to liver failure and cirrhosis. Clinically, it will be characterized by an increase in the total bile acid content in the blood. If patients cannot get effective treatment, then liver transplant will be the only choice for them.
Meanwhile, it should be noted that inflammation is an important reason for liver injury. Necrosis is more common than apoptosis in the area of cholestatic liver injury, which is a standard inflammatory feature (Woolbright and Jaeschke, 2012). After 6 h of bile duct ligation (BDL), neutrophils will accumulate in the area of necrosis and liver injury (Woolbright et al., 2013). Bile acids kill hepatocytes by activating neutrophils to produce reactive oxygen species. Inhibiting neutrophil function in hepatocytes can reduce oxidative stress and liver injury (Copple et al., 2010). The expression levels of serum inflammatory cytokines, such as tumor necrosis factor-α (TNF-α), interleukin (IL)-1β, and IL-6, are increased in patients with cholestasis, demonstrating that inflammation plays a role in cholestasis (Barak et al., 2009). Corilagin reduces cholestatic liver injury induced by alpha-naphthylisothiocyanate by exerting anti-inflammation effects and decreasing nuclear factor kappa-B (NF-κB) levels (Jin et al., 2013). Stearic acid, a drug with anti-inflammatory potential, also attenuates the pathophysiological changes in cholestasis induced by BDL (Pan et al., 2010). The hepatoprotective effect of stearic acid is associated with anti-inflammatory effects (Pan et al., 2010).
Therefore, we consider that inflammation is an inevitable burden of cholestasis. The mechanism of bile acids in triggering inflammation remains controversial. Early growth response factor-1 (Egr-1), inflammasome, and Toll-like receptors are all related to inflammation caused by bile acid (Figure 1). Three pathways will be described in detail as follows.
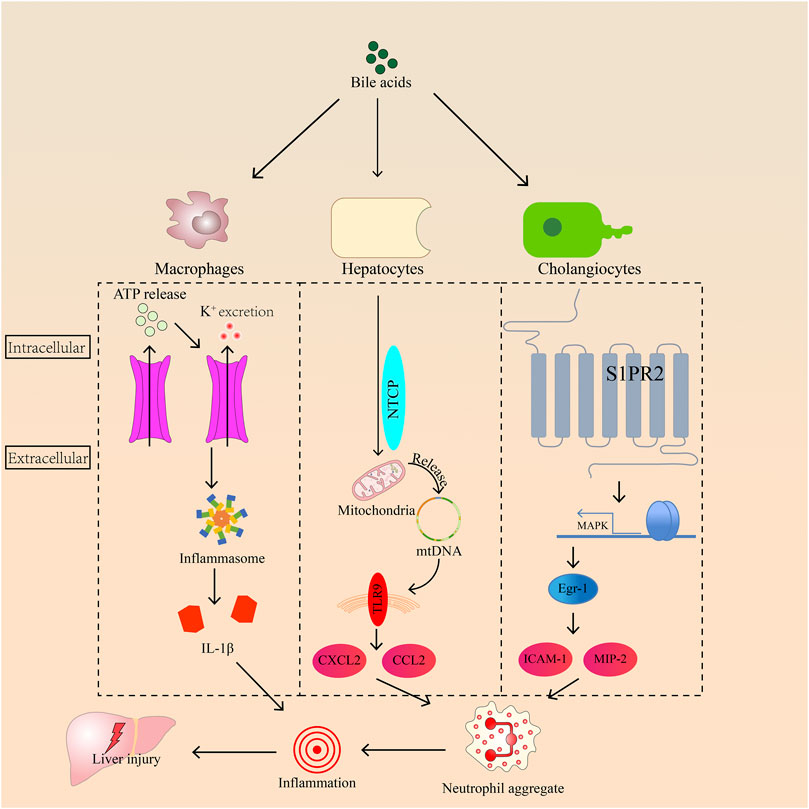
FIGURE 1. The roles of Egr-1, toll-like receptor-9, and inflammasome in regulating inflammation induced by bile acids in hepatocyte.
Egr-1 plays an important role in the development of inflammation during cholestasis. It is activated by bile acids in hepatocytes, and its activation requires mitogen-activated protein kinase (MAPK) signaling. Then, Egr-1 can activate intercellular adhesion molecule 1 (ICAM-1) production, and macrophage inflammatory protein 2 induces neutrophil aggregation, leading to inflammation (Kim et al., 2006; Allen et al., 2010). Sphingosine-1-phosphate receptor 2 (S1PR2), a bile acid sensor, transmits signals to bile acids and Egr-1 through MAPK, resulting in neutrophil accumulation in the liver (Zhang et al., 2019). Inflammasomes consist of an upstream sensor protein of the NOD-like receptor family, an adaptor protein, and the downstream effector protease caspase-1 (de Vasconcelos et al., 2016). The inflammasome is activated by K+ outflow caused by bile acids in macrophages, inducing ATP release (Gong et al., 2016). The autocatalytic cleavage of caspase-1 promotes proinflammatory factor pro-IL-1β and pro-IL-18 maturity when sensor receptors are stimulated by microbial or sterile stressors (Elliott and Sutterwala, 2015). Meanwhile, bile acids activate inflammasomes in inflammatory macrophages (Hao et al., 2017). For example, chenodeoxycholic acid increases the concentrations of IL-1β through the activation of the NLRP3 inflammasome in macrophages (Gong et al., 2016). Toll-like receptors, protective immune sentries, sense pathogen- or damage-associated molecular patterns and trigger gene expression changes that ultimately eradicate invading microbes (Lim and Staudt, 2013). In vitro, high levels of bile acids injure the mitochondria by changing their membrane potential and releasing cytochrome c (Botla et al., 1995). Patients with cholestasis have high bile acid levels in liver cells. This symptom may also cause mitochondrial damage in the hepatocytes, damaging or releasing the mitochondrial genome (mtDNA) from mitochondria. Some studies also proved that cholestasis is often accompanied by mtDNA damage (Tiao et al., 2007; Xu et al., 2012). In hepatocytes, the mitochondria release mtDNA, which can be detected by Toll-like receptor 9. Ultimately, bile acids induce cell necrosis and release C-X-C motif chemokine ligand 2 and chemokine ligand 2 to recruit neutrophils that cause inflammation through damaged mtDNA (Cai et al., 2017).
In addition to causing liver injury, inflammation can aggravate cholestasis by affecting transporters of bile acids in hepatocytes. TNF-α and IL-1 can reduce the mRNA expression levels of bile salt export pump (BSEP) and multidrug resistance-associated protein 2 (MRP2), which control bile acid discharge from the liver (Azeltine et al., 2020). Meanwhile, oxidative stress caused by neutrophils is a mechanism of cell death, and subsequent fibrosis results in cirrhosis and liver failure. Thus, controlling inflammation is as important as regulating the size of the bile acid pool in cholestasis treatment.
Peroxisome proliferator-activated receptor alpha
Peroxisome proliferator-activated receptors (PPARs) belong to the subfamily 1 of the nuclear hormone receptor superfamily of transcription factors (Nuclear Receptors Nomenclature Committee, 1999) and regulate genes important for cell differentiation and various metabolic processes, especially lipid and glucose homeostasis (Grygiel-Gorniak, 2014). PPARs comprise the following three subtypes: PPARα, PPARγ, and PPARδ (also designated as PPARβ). PPARs have the basic structural properties of most nuclear receptors; that is, PPARs consist of four functional domains, namely, A/B, C, D, and E/F. The N-terminal A/B domain contains the ligand-independent activation function 1, which is responsible for PPAR phosphorylation. The conserved central DNA binding domain, also known as the C domain, is composed of two zinc fingers and is responsible for the binding of PPAR to the peroxisome proliferator response element (PPRE) in the promoter of the PPAR target genes. The D domain is a docking site for various cofactors. The E domain is also named the ligand-binding domain (Christofide s et al., 2021). The full transcriptional activity of PPARs requires the binding of cognate lipid ligands and heterodimerization with another nuclear receptor, retinoid-X receptor (RXR) (Gyamfi and Wan, 2009). PPARs stimulate the expression of several genes by binding to specific PPREs through cooperation with retinoid X receptors.
PPARα is expressed in the skeletal muscles, heart, liver, kidney, and brown adipose tissues and is associated with fatty acid catabolism (Han et al., 2020). PPARα agonists can control bile acid homeostasis by inducing metabolic enzymes and inhibiting bile acid synthesis (Ghonem et al., 2015). The expression of PPARα is associated with fatty acid catabolism, and PPARα functions as a lipid sensor and controls energy combustion (Ip et al., 2003; Han et al., 2020). Meanwhile, PPARα plays a vital role in glucose homeostasis and insulin resistance development (Fruchart et al., 2001). Synthetic PPARα agonists have been identified, including fenofibrate, which is one of the most commonly used fibrates in cholestasis treatment trials (Dai et al., 2017); WY-14643; and bezafibrate, which is a pan-PPAR agonist. The natural ligands of PPARα mainly include unsaturated fatty acids, leukotriene B4, and 8-hydroxyeicosatetraenoic acid (Grygiel-Gorniak, 2014). Normally, PPARα agonists are used to treat patients with fatty liver, diabetes, and dyslipidemia (Oscarsson et al., 2018; Zhu et al., 2020). Recently, researchers found that PPARα agonists can improve the condition of patients with primary sclerosing cholangitis (PSC) and PBC (de Vries et al., 2021). Meanwhile, another study had shown that they can be used as a treatment for cholestasis (Honda et al., 2013).
Regulation of bile acid transport and metabolism by peroxisome proliferator-activated receptor alpha
In vivo, the activation of PPARα is associated with the increased hepatobiliary circulation of bile acids, inhibition of hepatic bile acid biosynthesis, and reduction in plasma triglycerides (Zollner et al., 2010). We will discuss the roles of PPARα in bile acid transport and metabolism and its implications for cholestatic disorders (Figure 2).

FIGURE 2. The role of PPARα in reguleation bile acids transport and metabolism.
Activation of peroxisome proliferator-activated receptor alpha by bile acids
As a nuclear receptor that controls lipid and glucose metabolism, PPARα can be influenced by bile acid. In addition to activated bile acids, PPARα is a transcriptional target of bile acid-activated FXR, indicating that it can be indirectly activated by bile acids through FXR (Pineda et al., 2003; Dai et al., 2018; Hua et al., 2019). However, when mice were fed with a bile acid-enriched diet, PPARα was inhibited by bile acids (Sinal et al., 2001). The mRNA levels of targets for PPARα, including CYP4A1, CYP4A3, ACOX, BE, and thiolase, were reduced substantially when using combined cholic acid/WY-14643 diet compared with WY-14643 alone. Moreover, bile acids control intestinal peptide absorption transporter in the gut by inhibiting PPARα (Okamura et al., 2014). Although bile acids can indirectly activate PPARα through FXR, the real relationship between bile acids and PPARα is inhibition.
Regulation of bile acid metabolism
The activation of PPARα by ligands, including WY-14643 and fatty acids, or during fasting induces a range of actions involved in bile acid homeostasis. PPARα detoxification pathways that process harmful bile acids can be mainly divided into two types, namely, hydroxylation by members of the cytochrome P450 subfamily and conjugation by UDP glucuronosyltransferases (UGTs) and sulfotransferases (SULTs). These processes reduce the number of harmful bile acids in the body by inhibiting the generation of bile acids, increasing the polarity of bile acids, and facilitating excretion through bile or urine. Cholesterol 7 alpha-hydroxylase (CYP7A1), a rate-limiting enzyme of bile acid production, is downregulated by PPARα. Treatment with 0.05% ciprofibrate decreased CYP7A1 enzyme activity and mRNA, but these effects were completely abolished in Pparα−/− mice (Patel et al., 2000; Post et al., 2001). Meanwhile, sterol 26-hydroxylase (CYP27A1), the key enzyme of alternate pathways of bile acid synthesis, is also reduced in this case (Post et al., 2001). The inhibition of murine Cyp7a1 gene caused by PPARα activation is sensitive to the human CYP7A1 gene. The differential response of the murine Cyp7a1 and human CYP7A1 gene promoters to PPARα activators is attributable to the additional PPARα/RXRα-binding site in the murine CYP7A1 gene promoter (Cheema and Agellon, 2000). After HepG2 cells were cultured with WY-14643, the level of CYP7A1 transactivation decreased with HNF-4 alpha level. This result suggested that PPARα and agonist repress CYP7A1 by reducing the availability of HNF-4 that would bind the DR-1 sequence, thereby attenuating CYP7A1 transactivation by HNF-4 (Marrapodi and Chiang, 2000). In addition to CYP7A1, PPARα can activate other cytochromes to promote bile acid excretion out of the body. Perfluorodecanoic acid is a persistent organic pollutant with the ability to activate PPARα. Studies found that perfluorodecanoic acid reduced the mRNA level of Cyp7a1, Cyp8b1, and sodium taurocholate cotransporting polypeptide (NTCP) in mice administered with perfluorodecanoic acid. This phenomenon was not found in Pparα-null mice (Luo et al., 2017). The reduction of the above three proteins can reduce the production of bile acid in the liver and reduce the size of the bile acid pool of hepatocytes.
CYP3A4 is the major enzyme expressed in the liver and gut. It is involved in the detoxification, hydrolyzation, and subsequent glucuronidation of bile acids by UGTs (Chen et al., 2014). In vivo, WY-14643 is involved in the induction of CYP3A4 mRNA in the liver but not in the intestine; this finding establishes PPARα as a direct transcriptional regulator of hepatic CYP3A4 (Thomas et al., 2013). Sterol 12α-hydroxylase (CYP8B1) increases the level of cholic acid/chenodeoxycholic acid. Chenodeoxycholic acid is converted into lithocholic acid after 7α-dehydroxylation by coliform flora. Therefore, the activation of CYP8B1 can decrease the production of lithocholic acid to reduce the hepatotoxicity of total bile acids in hepatocytes. The expression of Cyp8b1 decreased in stard10−/− mice after PPARα activity was impaired (Ito et al., 2013). PPARα activation can induce an increase in taurocholic acid level and is related to an increase in CYP8B1 level (Xie et al., 2019). In vitro, WY-14643 treatment increased the relative amount of cholic acid in HepG2 cells by activating CYP8B1 (Shi et al., 2005). PPRE identified in the rat sterol 12 alpha-hydroxylase promoter region in HepG2 cells was activated after WY-14643 treatment (Hunt et al., 2000). Therefore, PPARα can decrease the toxicity of total bile acids by decreasing the level of chenodeoxycholic acid. Other cytochromes, such as cytochrome P450 3a, cytochrome P450 2b, and cytochrome P450 2c, were induced in a dose-dependent manner by gemfibrozil (Shi et al., 2017).
Organic anion-transporting polypeptide (OATP) mediates the Na+-independent transport of organic anions such as sulfobromophthalein and conjugated and unconjugated bile acids to the liver. In vivo, the mRNA expression of Oatp1a1, 1b2, 2a1, and 2b1 in the liver is decreased by PPARα ligands (clofibrate, ciprofibrate, and diethylhexylphthalate) (Cheng et al., 2005). Through this process, the bile acid level in the liver was decreased.
The UGT family is responsible for the transfer of glucuronic acid to other molecules, such as bile acids, and acts as a catalyst. The induction of UGT2B4 by bile acids contributes to a feed-forward reduction of bile acid toxicity (Barbier et al., 2003a). The incubation of human hepatocytes with WY-14643 increases UGT2B4 mRNA levels (Barbier et al., 2003b). These results suggested that the activation of PPARα can reduce bile acid toxicity through UGT2B4. UGT1A1, UGT1A3, UGT1A4, and UGT1A6 are the targets of PPARα in human hepatocytes (Senekeo-Effenberger et al., 2007). SULT catalyzes the sulfation of bile acids, increases its water solubility, and promotes excretion. PPARα participates in the transcriptional regulation of SULT2A1 and SULT2A8 (Fang et al., 2005; Feng et al., 2017).
Organic solute transporter (OST) subunits OSTα and OSTβ facilitate bile acid efflux from the enterocyte into the portal circulation. OSTα/β knockout mice have longer and thicker small intestines and are largely protected against experimental cholestatic liver injury (van de Wiel et al., 2022). However, some researchers found that the level of OSTα/β expression was not changed in Oatp1a1-null BDL mice with increased Pparα expression (Zhang et al., 2012). We speculated that the ability of PPARα to regulate bile acids is not via OSTα/β.
Regulation of bile acid elimination
Multidrug resistance 2 (MDR2, also known as ABCB4) is a multidrug resistance gene located in zone 1, region 2 of the long arm of chromosome 7. It mainly exists in the bile duct membranes of hepatocytes and is expressed in the normal human placenta. The MDR2 gene transfers phospholipids from hepatic lobules to the outer surface of the bile duct membrane (Elferink and Groen, 2002). Abcb4−/− mice displayed progressive liver damage at an early age, and this effect was accompanied by hyperbilirubinemia and an increase in liver enzymes in the plasma (Elferink and Groen, 2002). Human MDR3 and mouse Mdr2 have a high degree of homology, and the p-gp amino acid sequences encoded by them have 90% similarity (Gros et al., 1988). Mdr2 plays an essential role in the secretion of phosphatidylcholine into bile and may be a phospholipid transport protein or phospholipid flippase (Smit et al., 1993). Phospholipids are essential components of the bile and reduce the detergent activity of bile acid micelles, thereby protecting the membranes of cells lining the biliary tree from damage. When Mdr2 is damaged, the amount of phospholipids in the bile ducts becomes insufficient, and liver damage subsequently occurs. PFIC3 is caused by the mutations in the ABCB4 gene (Davit-Spraul et al., 2010). In cholestasis, UDCA may contribute to therapeutic effects by inducing alternative excretory routes for bile acids and other cholephiles through activating ABCB4 (Zollner et al., 2003). Therefore, MDR3 can be a target for partial cholestasis treatment.
Fibrates, the agonists of PPARα, induce the hepatic expression of MDR2 and encode the canalicular phospholipid translocator (Kok et al., 2003). The secretion of phospholipids and cholesterol increased only during high-bile-salt infusions, and no fibrate effects were observed in PPARα−/− mice. The exposure of cultured wild-type mouse hepatocytes to PPARα agonists specifically induced Mdr2 mRNA levels. Thus, PPARα increased the amounts of phospholipids in the canalicular network through Mdr2 in mice. However, given the species-specific nature of the gene, whether PPARα can activate ABCB4 in humans remains unclear. Thus, fenofibrate was used to stimulate human liver cells (Ghonem et al., 2014). Fenofibrate significantly upregulated MDR3 mRNA and protein expression in primary cultured human hepatocytes and stimulated MDR3 promoter activity in HepG2 cells. In silico analysis of the 5′-upstream region of the human MDR3 gene revealed a number of PPREs, showing that PPARα activates MDR3 gene transcription by directly binding to PPRE (Ghonem et al., 2014).
In addition to MDR3, BSEP and the multidrug resistance-associated protein family are associated with PPARα in bile acid homeostasis regulation. BSEP catalyzes the transport of major hydrophobic bile salts, such as taurine and glycine-conjugated cholic acid, across the canalicular membranes of hepatocytes in an ATP-dependent manner. Clofibrate, a PPARα agonist, reduced the total bile acids in mouse livers, but this effect was not observed in PPARα−/− mice. An increase in the mRNA level of BSEP resulted in a reduction of total bile acids in livers (Zhang et al., 2017). Multidrug resistance-associated protein 3 (MRP3) and MRP4 are often adaptively upregulated in cholestasis and can partly alleviate bile acid accumulation in the liver. Thus, the upregulation of MRP3 and MRP4 may be an adjunct to the treatment of cholestasis. PPARα can regulate polysaccharide-resistant proteins (Moffit et al., 2006; Wang et al., 2018). For instance, the mRNA levels of MRP3 and MRP4 increased only in wild-type mice when wild-type and PPARα−/− mice received clofibrate treatment (Moffit et al., 2006). All these proteins were found to be effective in inhibiting intrahepatic bile acid deposition.
Crosstalk with nuclear farnesoid X receptor
FXR is the main regulator of bile acid homeostasis because it transcriptionally drives the modulation of bile acid synthesis, influx, efflux, and detoxification along the enterohepatic axis (Cariello et al., 2018). FXR belongs to the nuclear receptor family and is expressed in the liver; PPARα is activated in the liver by fatty acids during fasting (Pawlak et al., 2015), whereas FXR is activated by bile acid return in the liver during feeding (Preidis et al., 2017). The crosstalk between energy balance, including that between glucose and lipid, has been extensively explored (Preidis et al., 2017). We focus on the interaction between them in terms of bile acid homeostasis.
FXR decreases the rate of bile acid synthesis by activating the small heterodimer partner (SHP), thereby inhibiting CYP7A1, suppressing NTCP, and reducing the rate of bile acid absorption by hepatocytes while promoting the expression of BSEP and expelling bile acids from the liver (Ding et al., 2015). In addition, FXR counteracts liver X receptor in cholesterol and triglyceride metabolism (Kalaan y and Mangelsdorf, 2006). Thus, PPARα and FXR act on common metabolic pathways. The molecular crosstalk between these two nuclear receptors needs to be investigated.
The treatment of HepG2 cells with chenodeoxycholic acid led to a dose-dependent increase in hPPARα mRNA levels (Pineda et al., 2003). The induction of hPPARα expression by bile acids influenced the response of the PPARα target gene CPT-1 to PPARα ligands. This result suggested that an increase in PPARα expression occurs partly through the transcriptional mechanisms of FXR. The discovery of an FXR response element located in the human PPARα promoter further supports this standpoint. Therefore, activating FXR can upregulate the expression of PPARα. However, PPARα was found to have an inhibitory effect in rodents. 1-Naphthyl isocyanate (ANIT), a model drug for cholestasis, inhibits the expression of FXR (Zhang et al., 2020). When the control group and PPARα−/− mice received 0.05% ANIT orally, the levels of Shp and Fxr mRNA doubled in the cholestatic PPARα−/− mice compared with those in the control group (Hua et al., 2019). Similarly, the targets of PPARα were increased in Shp−/− mice (Park et al., 2011). Meanwhile, a study found the relationship between PPARα and FGF15 (Zhou et al., 2014). FGF15/19 is also an important bile acid target gene regulated by FXR to control CYP7A1 upregulation. In the model of inflammatory bowel disease, the accumulation of bile acids in inflamed colon tissues can repress FXR-FGF15 signaling by activating the intestinal PPARα–UGT pathway to eliminate bile acids in the intestine. Treatment with PPARα agonist fenofibrate can decrease the level of serum concentrations of FGF-19 in obesity (Mraz et al., 2011). Thus, the crosstalk between basal PPARα and FXR occurred, and adaptation of bile acid metabolism was inhibited in chronic cholestasis (Hua et al., 2019). In addition to the indirect effects through regulation of bile acids, this crosstalk may be related to PPARα and FXR competing with RXRα (Xie et al., 2019). This research used chenodeoxycholic acid to activate FXR and WY-14643 to active PPARα. The results showed that chenodeoxycholic acid suppressed WY-14643-induced PPARα activation, whereas WY-14643 suppressed chenodeoxycholic acid-induced FXR activation. These suppressive effects were abolished by using HX531, an RXRα inhibitor. The results suggest a crosstalk between PPARα and FXR potentially through RXRα competition. However, PPARα agonists can also cause liver injury (Hedrington and Davis, 2018). Although large randomized human trials have shown little or no hepatocellular abnormalities when fibrates were used alone, signs of hepatotoxicity appeared more noticeably when fibrates were combined with other drugs. In some case reports, liver damage was demonstrated with the treatment of fibrates as indicated by increased aminotransferase levels (Ho et al., 2004; Dohmen et al., 2005). The liver function usually improved after discontinuation of treatment with fibrates. Therefore, the aminotransferase levels should be monitored when fibrates are used to treat disease.
Peroxisome proliferator-activated receptor alpha as a therapeutic target in cholestasis treatment
The mechanism by which PPARα facilitates cholestasis treatment mainly involves the reduction of bile acid pool size in the liver and regulation of damage due to cholestasis (Figure 3). PPARα agonists are usually used for patients who do not respond to UDCA. The compounds under investigation are shown in Table 2. In general, PPARα agonists are usually used as combined medication with UDCA. They are promising drugs for patients with incomplete biochemical responses to UDCA and those with liver fibrosis and dyslipidemia.
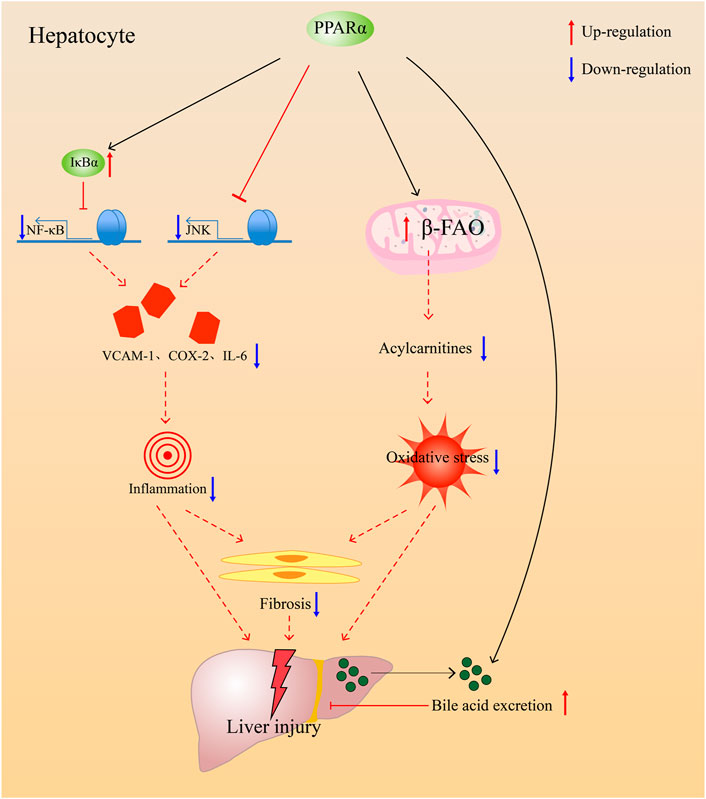
FIGURE 3. PPARα alleviates injury accused by cholestasis through anti-inflammation, anti-oxidative, antifibrosis, and prevent bile acids accumulate in the liver though promoting bile acids excretion.
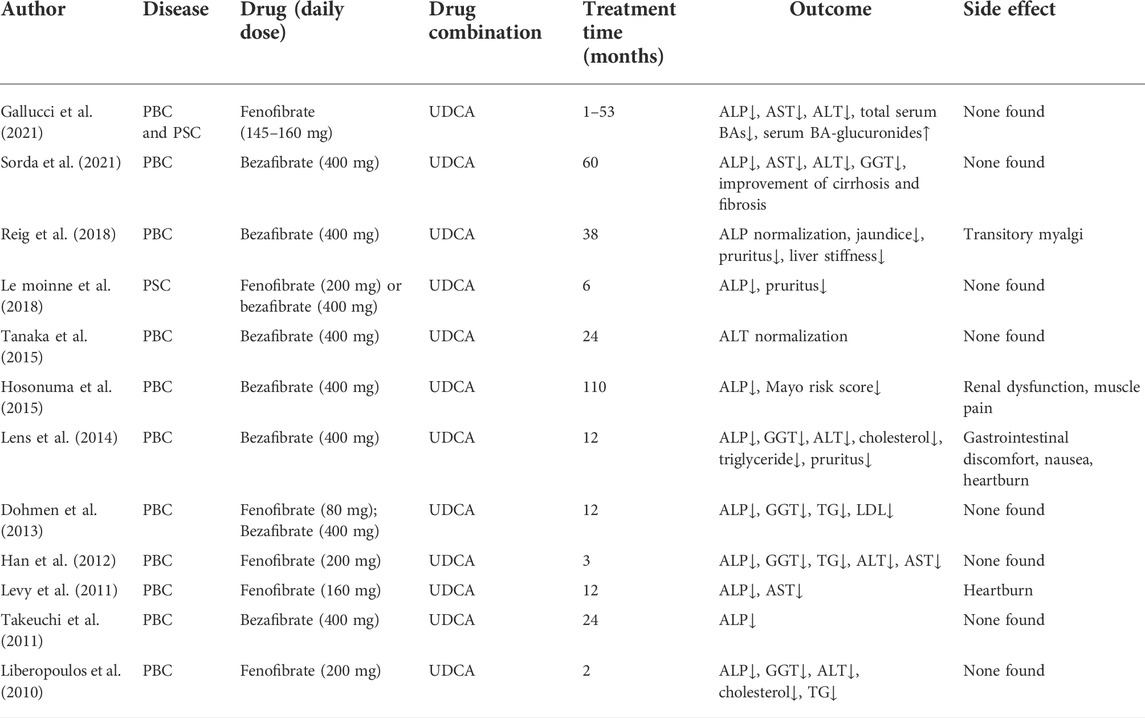
TABLE 2. The agonists of PPARα under investigation.
Anti-cholestasis
In some studies, researchers usually gavage mice with ANIT and lithocholic acid to injure normal hepatocytes or block bile flow with BDL to create a pathological model similar to cholestasis. The transporter-related gene knockout technique has been used in conventional cholestasis models. Fenofibrate, a PPARα agonist, has an effective anti-cholestasis function. In a rat model of BDL, fenofibrate induced a decrease in serum biochemical index and eased hepatocellular damage by increasing PPARα expression within liver cells (Cindoruk et al., 2007). In another rat model of ANIT, the protective effects of fenofibrate against cholestasis-induced hepatic injury depended on PPARα and fenofibrate doses (Dai et al., 2017). When subjected to cholic acid dietary challenge, PPARα−/− mice showed bile acid accumulation in their livers, resulting from decreasing levels of mRNA-encoding transporters, including Abcb11, Abcb4, Abca1, Abcg5, and Abcg8 (Li et al., 2012). In the rat primary biliary cirrhosis model, a novel PPARα/δ dual agonist 5c demonstrated excellent in vivo efficacy (Jiang et al., 2019). Fibrate drugs are PPARα agonists and are mainly used as cholesterol-lowering drugs for patients with elevated triglycerides. Fibrate treatment is effective for PBC patients with incomplete response to UDCA (Cuperus et al., 2014). In patients with UDCA-refractory PBC, additional fibrate treatment is associated with the normalization of ALP, lowered risk of cirrhosis development, and reduced risk of hepatic deterioration (Chung et al., 2019). Fenofibrate, a member of the fibrate family, is a widely used alternative therapy for cardiovascular diseases. It contains high-affinity PPARα agonists (Issemann and Green, 1991). In a pilot study, fenofibrate was added to 22 patients with partial response to UDCA (Han et al., 2012). The results showed that fenofibrate effectively improved the liver biochemical test results in patients who had a partial response to UDCA monotherapy, and no obvious adverse effects were observed in patients who received fenofibrate (Han et al., 2012). Recently, researchers assessed fenofibrate, a PPARα agonist, as a combination therapy drug with UDCA in patients with PBC or PSC who had insufficient biochemical responses to UDCA (Ghonem et al., 2020). The addition of fenofibrate significantly reduced serum ALP levels by 82% vs. those receiving ursodiol monotherapy and normalized serum ALP values by 84% in all patients. Meanwhile, the production of proinflammatory cytokines was suppressed with the addition of fenofibrate. Thus, PPARα agonists can be used in treating cholestatic liver disorders.
Anti-inflammation
Liver injury often accompanies cholestasis and causes cholestasis to further deteriorate into other diseases. Inflammation and oxidative stress are the common causes of liver damage in cholestasis. As an organ of the immune system, the liver induces harmful liver inflammation when suffering from viral infection (Yang et al., 2019). Cholestasis is often accompanied by inflammation characterized by neutrophil infiltration (Wu et al., 2003). Neutrophils are found in the blood and are attracted to the site of inflammation by chemotactic substances when inflammation occurs. ICAM-1 is a protein responsible for the accumulation of neutrophils in the liver (Jaeschke, 1997). In patients with extrahepatic cholestasis, ICAM-1 expression and neutrophil recruitment are upregulated in the liver during extrahepatic cholestasis, which may lead to inflammatory damage to the liver (Gulubova, 1998). Other research also revealed that ANIT drives toxicity toward hepatocytes through neutrophils (Kodali et al., 2006). Thus, the treatment of inflammation is also an important part of the treatment of cholestatic liver injury.
In addition, PPARα can regulate inflammation in the body to regulate lipid and glucose homeostasis (Han et al., 2020). Exercise is a protective factor for lower levels of local inflammatory markers and less myocardial apoptosis, and it seems to be related to the presence of PPARα (Santos et al., 2016). When PPARα−/− mice were treated with proinflammatory substances, they suffered more injury from inflammation due to inflammasome activation or an increase in TNF-α level in the body (Li et al., 2012; Batatinha et al., 2017; Gugliandolo et al., 2019). In addition to this, fibrates had been proved to have the ability to treat neuroinflammation caused by paclitaxel (Caillaud et al., 2021). Therefore, we can conclude that PPARα agonists have a therapeutic effect on inflammation. NF-κB proteins are the key regulators of innate and adaptive immune responses, which can accelerate cell proliferation, inhibit apoptosis, promote cell migration and invasion, and stimulate angiogenesis and metastasis (Taniguchi et al., 2018). NF-κB causes inflammation by promoting ICAM excretion and enabling neutrophils to aggregate. PPARα activator has been reported to have anti-inflammatory properties (Huang et al., 2007; Hennuyer et al., 2016; Huang et al., 2016). WY-14643 can inhibit endotoxin-induced inflammation by suppressing the mRNA expression of IL-6, IL-1β, and TNF-α via controlling the NF-κB pathway (Huang et al., 2016). The same effect has been observed in human epithelial cells (Marx et al., 1999). The absence of PPARα may induce the overexpression of proinflammatory cytokines in LPS stimulus, which can further indicate its effect on anti-inflammation. PPARα treats inflammatory disease by promoting cell autophagy and inhibiting the inflammatory response (Jiao et al., 2016). PPARα induces the expression of the inhibitory protein NF-kappa-B-inhibitor alpha (IkBa) in human aortic smooth muscle cells, as well as in primary human hepatocytes; then, it inhibits NF-κB activation to decrease inflammation (Delerive et al., 2000). Fenofibrate, a PPARα agonist, provides protection against hepatic injury by inhibiting the JNK and NF-κB signaling pathways (Dai et al., 2017). PPARα can promote the inactivation of NF-κB during the inflammatory reaction, and the inhibition can inhibit the inflammatory cascade (Korbecki et al., 2019). In addition to inflammation, bile acids in the liver can cause mitochondrial damage and oxidative stress. PPARα activation by fenofibrate provides protection against liver damage by recovering mitochondrial fatty acid β-oxidation (β-FAO) which impaired by ANIT (Zhao et al., 2017). Therefore, PPARα eliminates oxidative stress by increasing the expression of β-FAO. Moreover, ANIT-induced liver fibrosis was alleviated by fenofibrate through PPARα (Lu et al., 2021), and anti-inflammation and antioxidation may play important roles in antifibrosis (Chung et al., 2018).
This mechanism of PPARα protection against inflammation may offer additional therapeutic opportunities for cholestatic liver diseases. Meanwhile, antifibrosis and antioxidant stress are important to the improvement of liver injury. Recently, PPARα has been found to be related to liver regeneration in mice (Fan et al., 2021). This function of liver regeneration may be a promising way to improve the condition of patients with cholestasis. The therapeutic effect of PPARα on inflammation and the regulation of bile acid homeostasis offers additional therapeutic opportunities for the treatment of cholestatic liver diseases.
Conclusion
With the advanced understanding of the pathology of cholestasis, liver injury has been found to have various causes. Besides bile acid directly damaging hepatocytes, inflammation and oxidative stress can also cause liver injury. Meanwhile, inflammation also affects bile acid transporter proteins. We can speculate that treating inflammation is as important as the regulation of bile acid homeostasis in the therapy of cholestasis. At present, cholestasis is mostly treated by regulating bile acid, and there are certain side effects. The main physiologic function of PPARα is to control glucose metabolism and energy combustion. However, PPARα is involved in the control of bile acid homeostasis, and the treatment of inflammation during cholestasis provides us new perspective to treat this disease. Therefore, finding safe and effective PPARα activators may have important clinical significance for the amelioration of cholestasis (Table 3).
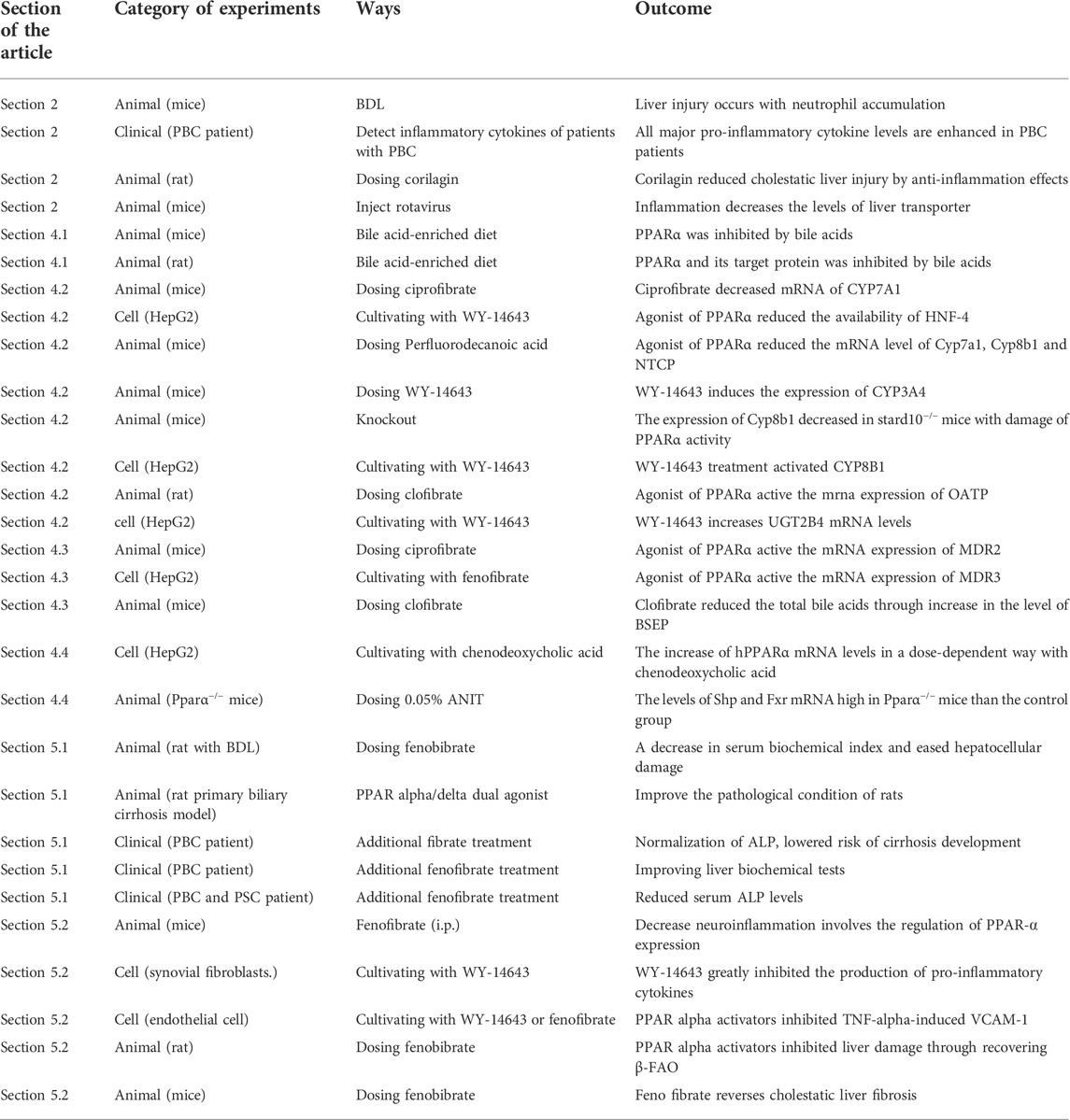
TABLE 3. Category of experiments.
As shown in the scheme (Figure 4), we introduce the ways in which PPARα regulates bile acid homeostasis and reduces liver injury. Through activating bile acids or cholesterol, PPARα can increase the expression of bile acid transporter proteins and bile acid detoxification proteins, including CYP7A1, BSEP, MDR3, MRP2, MRP3, MRP4, CYP3A4, UGTs, and SULTs. Meanwhile, PPARα regulates inflammatory factors such as TNF-α, IL-1β, MCP-1, and MIP-2. It also regulates the activation of neutrophils by inhibiting the expression of JNK and NF-κB. PPARα can also control enzymes to inhabit β-FAO, which may lead to liver injury by means of oxidative stress. In addition, natural PPARα activators are necessary for the treatment of cholestasis as they can suppress hepatocyte apoptosis, necrosis, and fibrosis.
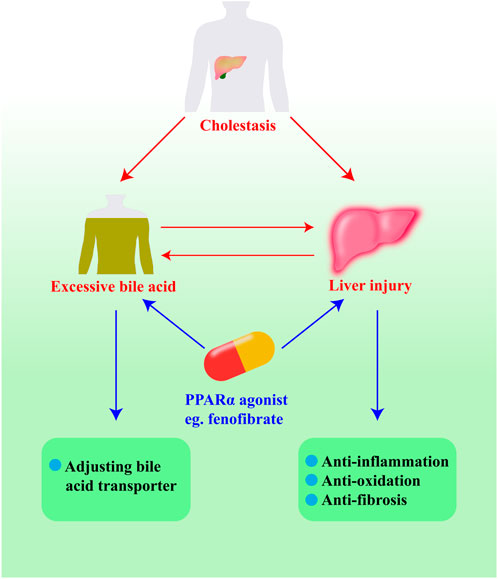
FIGURE 4. The pathways of PPARα agonists protecting against cholestasis.
With the progress of science and technology, there is a deeper understanding of the pathologic mechanism of cholestasis. The comprehensive regulation of bile acids and liver injury undoubtedly plays a role in treating the symptoms and root causes of cholestasis. However, the regulation of PPARα in cholestasis, including the crosstalk of PPARα and FXR, is still unclear. Due to the existence of species specificity, some experimental results may not completely correspond to the findings in clinical settings. Further studies are needed to improve our knowledge behind the PPARα mechanism. At present, fibrates combined with other therapeutic drugs seem to be a possible therapy for cholestatic liver injury in the clinic. PPARα activators are promising in the treatment of cholestasis.
Author contributions
TZ conceptualized and planned the review. XY wrote the manuscript. HH edited the manuscript. All authors read and approved the final manuscript.
Funding
This research was supported by National Natural Science Foundation of China (82173946), Natural Science Foundation of Shanghai (21ZR1460500), National Scientific and Technological Major Special Project of China (2019ZX09201004-002).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
Thanks to all the peer reviewers for their opinions and suggestions.
References
Allen, K., Kim, N. D., Moon, J. O., and Copple, B. L. (2010). Upregulation of early growth response factor-1 by bile acids requires mitogen-activated protein kinase signaling. Toxicol. Appl. Pharmacol. 243 (1), 63–67. doi:10.1016/j.taap.2009.11.013
Azeltine, M. W., Chavez, E. J., Nemec, K. M., Bednarek, J. M., Asokan, R., Balasubramaniyan, N., et al. (2020). Inflammation drives MicroRNAs to limit hepatocyte bile acid transport in murine biliary atresia. J. Surg. Res. 256, 663–672. doi:10.1016/j.jss.2020.07.003
Baker, A., Kerkar, N., Todorova, L., Kamath, B. M., and Houwen, R. (2019). Systematic review of progressive familial intrahepatic cholestasis. Clin. Res. Hepatol. Gastroenterol. 43 (1), 20–36. doi:10.1016/j.clinre.2018.07.010
Barak, V., Selmi, C., Schlesinger, M., Blank, M., Agmon-Levin, N., Kalickman, I., et al. (2009). Serum inflammatory cytokines, complement components, and soluble interleukin 2 receptor in primary biliary cirrhosis. J. Autoimmun. 33 (3-4), 178–182. doi:10.1016/j.jaut.2009.09.010
Barbier, O., Duran-Sandoval, D., Pineda-Torra, I., Kosykh, V., Fruchart, J. C., Staels, B., et al. (2003b). Peroxisome proliferator-activated receptor alpha induces hepatic expression of the human bile acid glucuronidating UDP-glucuronosyltransferase 2B4 enzyme. J. Biol. Chem. 278 (35), 32852–32860. doi:10.1074/jbc.M305361200
Barbier, O., Torra, I. P., Sirvent, A., Claudel, T., Blanquart, C., Duran-Sandoval, D., et al. (2003a). FXR induces the UGT2B4 enzyme in hepatocytes: A potential mechanism of negative feedback control of FXR activity. GASTROENTEROLOGY 124 (7), 1926–1940. doi:10.1016/s0016-5085(03)00388-3
Batatinha, H. A., Lima, E. A., Teixeira, A. A., Souza, C. O., Biondo, L. A., Silveira, L. S., et al. (2017). Association between aerobic exercise and rosiglitazone avoided the NAFLD and liver inflammation exacerbated in PPAR-alpha knockout mice. J. Cell. Physiol. 232 (5), 1008–1019. doi:10.1002/jcp.25440
Botla, R., Spivey, J. R., Aguilar, H., Bronk, S. F., and Gores, G. J. (1995). Ursodeoxycholate (UDCA) inhibits the mitochondrial membrane permeability transition induced by glycochenodeoxycholate: A mechanism of UDCA cytoprotection. J. Pharmacol. Exp. Ther. 272 (2), 930–938.
Bull, L. N., and Thompson, R. J. (2018). Progressive familial intrahepatic cholestasis. Clin. Liver Dis. 22 (4), 657–669. doi:10.1016/j.cld.2018.06.003
Cai, S. Y., Ouyang, X., Chen, Y., Soroka, C. J., Wang, J., Mennone, A., et al. (2017). Bile acids initiate cholestatic liver injury by triggering a hepatocyte-specific inflammatory response. JCI Insight 2 (5), e90780. doi:10.1172/jci.insight.90780
Caillaud, M., Patel, N. H., White, A., Wood, M., Contreras, K. M., Toma, W., et al. (2021). Targeting Peroxisome Proliferator-Activated Receptor-alpha (PPAR- alpha) to reduce paclitaxel-induced peripheral neuropathy. Brain Behav. Immun. 93, 172–185. doi:10.1016/j.bbi.2021.01.004
Cariello, M., Piccinin, E., Garcia-Irigoyen, O., Sabba, C., and Moschetta, A. (2018). Nuclear receptor FXR, bile acids and liver damage: Introducing the progressive familial intrahepatic cholestasis with FXR mutations. Biochim. Biophys. Acta. Mol. Basis Dis. 1864 (4 Pt B), 1308–1318. doi:10.1016/j.bbadis.2017.09.019
Chai, J., Feng, X., Zhang, L., Chen, S., Cheng, Y., He, X., et al. (2015). Hepatic expression of detoxification enzymes is decreased in human obstructive cholestasis due to gallstone biliary obstruction. PLOS ONE 10 (3), e0120055. doi:10.1371/journal.pone.0120055
Cheema, S. K., and Agellon, L. B. (2000). The murine and human cholesterol 7alpha-hydroxylase gene promoters are differentially responsive to regulation by fatty acids mediated via peroxisome proliferator-activated receptor alpha. J. Biol. Chem. 275 (17), 12530–12536. doi:10.1074/jbc.275.17.12530
Chen, J., Zhao, K. N., and Chen, C. (2014). The role of CYP3A4 in the biotransformation of bile acids and therapeutic implication for cholestasis. Ann. Transl. Med. 2 (1), 7. doi:10.3978/j.issn.2305-5839.2013.03.02
Cheng, X., Maher, J., Dieter, M. Z., and Klaassen, C. D. (2005). Regulation of mouse organic anion-transporting polypeptides (Oatps) in liver by prototypical microsomal enzyme inducers that activate distinct transcription factor pathways. Drug Metab. Dispos. 33 (9), 1276–1282. doi:10.1124/dmd.105.003988
Christofides, A., Konstantinidou, E., Jani, C., and Boussiotis, V. A. (2021). The role of peroxisome proliferator-activated receptors (PPAR) in immune responses. Metabolism. 114, 154338. doi:10.1016/j.metabol.2020.154338
Chung, K. W., Lee, E. K., Lee, M. K., Oh, G. T., Yu, B. P., Chung, H. Y., et al. (2018). Impairment of PPARα and the fatty acid oxidation pathway aggravates renal fibrosis during aging. J. Am. Soc. Nephrol. 29 (4), 1223–1237. doi:10.1681/ASN.2017070802
Chung, S. W., Lee, J. H., Kim, M. A., Leem, G., Kim, S. W., Chang, Y., et al. (2019). Additional fibrate treatment in UDCA-refractory PBC patients. Liver Int. 39 (9), 1776–1785. doi:10.1111/liv.14165
Cindoruk, M., Kerem, M., Karakan, T., Salman, B., Akin, O., Alper, M., et al. (2007). Peroxisome proliferators-activated alpha agonist treatment ameliorates hepatic damage in rats with obstructive jaundice: An experimental study. BMC Gastroenterol. 7, 44. doi:10.1186/1471-230X-7-44
Copple, B. L., Jaeschke, H., and Klaassen, C. D. (2010). Oxidative stress and the pathogenesis of cholestasis. Semin. Liver Dis. 30 (2), 195–204. doi:10.1055/s-0030-1253228
Cuperus, F. J., Halilbasic, E., and Trauner, M. (2014). Fibrate treatment for primary biliary cirrhosis. Curr. Opin. Gastroenterol. 30 (3), 279–286. doi:10.1097/MOG.0000000000000056
D'Amato, D., De Vincentis, A., Malinverno, F., Vigano, M., Alvaro, D., Pompili, M., et al. (2021). Real-world experience with obeticholic acid in patients with primary biliary cholangitis. JHEP Rep. 3 (2), 100248. doi:10.1016/j.jhepr.2021.100248
Dai, M., Hua, H., Lin, H., Xu, G., Hu, X., Li, F., et al. (2018). Targeted metabolomics reveals a protective role for basal PPARα in cholestasis induced by α-naphthylisothiocyanate. J. Proteome Res. 17 (4), 1500–1508. doi:10.1021/acs.jproteome.7b00838
Dai, M., Yang, J., Xie, M., Lin, J., Luo, M., Hua, H., et al. (2017). Inhibition of JNK signalling mediates PPARα-dependent protection against intrahepatic cholestasis by fenofibrate. Br. J. Pharmacol. 174 (18), 3000–3017. doi:10.1111/bph.13928
Davit-Spraul, A., Gonzales, E., Baussan, C., and Jacquemin, E. (2010). The spectrum of liver diseases related to ABCB4 gene mutations: Pathophysiology and clinical aspects. Semin. Liver Dis. 30 (2), 134–146. doi:10.1055/s-0030-1253223
de Vasconcelos, N. M., Van Opdenbosch, N., and Lamkanfi, M. (2016). Inflammasomes as polyvalent cell death platforms. Cell. Mol. Life Sci. 73 (11-12), 2335–2347. doi:10.1007/s00018-016-2204-3
de Vries, E., Bolier, R., Goet, J., Pares, A., Verbeek, J., de Vree, M., et al. (2021). Fibrates for itch (FITCH) in fibrosing cholangiopathies: A double-blind, randomized, placebo-controlled trial. Gastroenterology 160 (3), 734–743.e6. e6. doi:10.1053/j.gastro.2020.10.001
Delerive, P., Gervois, P., Fruchart, J. C., and Staels, B. (2000). Induction of IkappaBalpha expression as a mechanism contributing to the anti-inflammatory activities of peroxisome proliferator-activated receptor-alpha activators. J. Biol. Chem. 275 (47), 36703–36707. doi:10.1074/jbc.M004045200
Di Ciaula, A., Garruti, G., Lunardi, B. R., Molina-Molina, E., Bonfrate, L., Wang, D. Q., et al. (2017). Bile acid physiology. Ann. Hepatol. 16 (Suppl. 1), s4–s14. doi:10.5604/01.3001.0010.5493
Ding, L., Yang, L., Wang, Z., and Huang, W. (2015). Bile acid nuclear receptor FXR and digestive system diseases. Acta Pharm. Sin. B 5 (2), 135–144. doi:10.1016/j.apsb.2015.01.004
Dohmen, K., Tanaka, H., and Haruno, M. (2013). Effectiveness of fenofibrate in comparison to bezafibrate for patients with asymptomatic primary biliary cirrhosis. Fukuoka Igaku Zasshi 104 (10), 350–361. doi:10.15017/1398604
Dohmen, K., Wen, C. Y., Nagaoka, S., Yano, K., Abiru, S., Ueki, T., et al. (2005). Fenofibrate-induced liver injury. World J. Gastroenterol. 11 (48), 7702–7703. doi:10.3748/wjg.v11.i48.7702
Elferink, R. O., and Groen, A. K. (2002). Genetic defects in hepatobiliary transport. Biochim. Biophys. Acta 1586 (2), 129–145. doi:10.1016/s0925-4439(01)00103-x
Elliott, E. I., and Sutterwala, F. S. (2015). Initiation and perpetuation of NLRP3 inflammasome activation and assembly. Immunol. Rev. 265 (1), 35–52. doi:10.1111/imr.12286
Fan, S., Gao, Y., Qu, A., Jiang, Y., Li, H., Xie, G., et al. (2021). YAP-TEAD mediates PPAR α–induced hepatomegaly and liver regeneration in mice. Hepatology 75, 74–88. doi:10.1002/hep.32105
Fang, H. L., Strom, S. C., Cai, H., Falany, C. N., Kocarek, T. A., Runge-Morris, M., et al. (2005). Regulation of human hepatic hydroxysteroid sulfotransferase gene expression by the peroxisome proliferator-activated receptor alpha transcription factor. Mol. Pharmacol. 67 (4), 1257–1267. doi:10.1124/mol.104.005389
Feng, L., Yuen, Y. L., Xu, J., Liu, X., Chan, M. Y., Wang, K., et al. (2017). Identification and characterization of a novel PPARα-regulated and 7α-hydroxyl bile acid-preferring cytosolic sulfotransferase mL-STL (Sult2a8). J. Lipid Res. 58 (6), 1114–1131. doi:10.1194/jlr.M074302
Floreani, A., and Gervasi, M. T. (2016). New insights on intrahepatic cholestasis of pregnancy. Clin. Liver Dis. 20 (1), 177–189. doi:10.1016/j.cld.2015.08.010
Forman, B. M., Goode, E., Chen, J., Oro, A. E., Bradley, D. J., Perlmann, T., et al. (1995). Identification of a nuclear receptor that is activated by farnesol metabolites. Cell 81 (5), 687–693. doi:10.1016/0092-8674(95)90530-8
Fruchart, J. C., Staels, B., and Duriez, P. (2001). The role of fibric acids in atherosclerosis. Curr. Atheroscler. Rep. 3 (1), 83–92. doi:10.1007/s11883-001-0015-x
Gallucci, G. M., Trottier, J., Hemme, C., Assis, D. N., Boyer, J. L., Barbier, O., et al. Adjunct fenofibrate up-regulates bile acid glucuronidation and improves treatment response for patients with cholestasis, Hepatol. Commun. 2021, 5(12), 2035–2051.doi: doi:10.1002/hep4.1787
Ghonem, N. S., Ananthanarayanan, M., Soroka, C. J., and Boyer, J. L. (2014). Peroxisome proliferator-activated receptor alpha activates human multidrug resistance transporter 3/ATP-binding cassette protein subfamily B4 transcription and increases rat biliary phosphatidylcholine secretion. Hepatology 59 (3), 1030–1042. doi:10.1002/hep.26894
Ghonem, N. S., Assis, D. N., and Boyer, J. L. (2015). Fibrates and cholestasis. Hepatology 62 (2), 635–643. doi:10.1002/hep.27744
Ghonem, N. S., Auclair, A. M., Hemme, C. L., Gallucci, G. M., de la Rosa, R. R., Boyer, J. L., et al. (2020). Fenofibrate improves liver function and reduces the toxicity of the bile acid pool in patients with primary biliary cholangitis and primary sclerosing cholangitis who are partial responders to ursodiol. Clin. Pharmacol. Ther. 108 (6), 1213–1223. doi:10.1002/cpt.1930
Gijbels, E., Vilas-Boas, V., Deferm, N., Devisscher, L., Jaeschke, H., Annaert, P., et al. (2019). Mechanisms and in vitro models of drug-induced cholestasis. Arch. Toxicol. 93 (5), 1169–1186. doi:10.1007/s00204-019-02437-2
Gong, Z., Zhou, J., Zhao, S., Tian, C., Wang, P., Xu, C., et al. (2016). Chenodeoxycholic acid activates NLRP3 inflammasome and contributes to cholestatic liver fibrosis. Oncotarget 7 (51), 83951–83963. doi:10.18632/oncotarget.13796
Gros, P., Raymond, M., Bell, J., and Housman, D. (1988). Cloning and characterization of a second member of the mouse mdr gene family. Mol. Cell. Biol. 8 (7), 2770–2778. doi:10.1128/mcb.8.7.2770
Grygiel-Gorniak, B. (2014). Peroxisome proliferator-activated receptors and their ligands: Nutritional and clinical implications--a review. Nutr. J. 13, 17. doi:10.1186/1475-2891-13-17
Gugliandolo, E., Fusco, R., Ginestra, G., D'Amico, R., Bisignano, C., Mandalari, G., et al. (2019). Involvement of TLR4 and PPAR-alpha receptors in host response and NLRP3 inflammasome activation, against pulmonary infection with Pseudomonas aeruginosa. SHOCK 51 (2), 221–227. doi:10.1097/SHK.0000000000001137
Gulamhusein, A. F., and Hirschfield, G. M. (2020). Primary biliary cholangitis: Pathogenesis and therapeutic opportunities. Nat. Rev. Gastroenterol. Hepatol. 17 (2), 93–110. doi:10.1038/s41575-019-0226-7
Gulubova, M. V. (1998). Intercellular adhesion molecule-1 (ICAM-1) expression in the liver of patients with extrahepatic cholestasis. Acta Histochem. 100 (1), 59–74. doi:10.1016/s0065-1281(98)80006-8
Gyamfi, M. A., and Wan, Y. J. (2009). Mechanisms of resistance of hepatocyte retinoid X receptor alpha-null mice to WY-14, 643-induced hepatocyte proliferation and cholestasis. J. Biol. Chem. 284 (14), 9321–9330. doi:10.1074/jbc.M808861200
Halilbasic, E., Fuchs, C., Hofer, H., Paumgartner, G., and Trauner, M. (2015). Therapy of primary sclerosing cholangitis--today and tomorrow. Dig. Dis. 33 (Suppl. 2), 149–163. doi:10.1159/000440827
Han, X., Wu, Y., Yang, Q., and Cao, G. (2020). Peroxisome proliferator-activated receptors in the pathogenesis and therapies of liver fibrosis. Pharmacol. Ther. 222, 107791. doi:10.1016/j.pharmthera.2020.107791
Han, X. F., Wang, Q. X., Liu, Y., You, Z. R., Bian, Z. L., Qiu, D. K., et al. (2012). Efficacy of fenofibrate in Chinese patients with primary biliary cirrhosis partially responding to ursodeoxycholic acid therapy. J. Dig. Dis. 13 (4), 219–224. doi:10.1111/j.1751-2980.2012.00574.x
Hao, H., Cao, L., Jiang, C., Che, Y., Zhang, S., Takahashi, S., et al. (2017). Farnesoid X receptor regulation of the NLRP3 inflammasome underlies cholestasis-associated sepsis. Cell Metab. 25 (4), 856–867. e5. doi:10.1016/j.cmet.2017.03.007
Hedrington, M. S., and Davis, S. N. (2018). Peroxisome proliferator-activated receptor alpha-mediated drug toxicity in the liver. Expert Opin. Drug Metab. Toxicol. 14 (7), 671–677. doi:10.1080/17425255.2018.1483337
Hennuyer, N., Duplan, I., Paquet, C., Vanhoutte, J., Woitrain, E., Touche, V., et al. (2016). The novel selective PPARα modulator (SPPARMα) pemafibrate improves dyslipidemia, enhances reverse cholesterol transport and decreases inflammation and atherosclerosis.. ATHEROSCLEROSIS 249, 200–208. doi:10.1016/j.atherosclerosis.2016.03.003
Ho, C. Y., Kuo, T. H., Chen, T. S., Tsay, S. H., Chang, F. Y., Lee, S. D., et al. (2004). Fenofibrate-induced acute cholestatic hepatitis. J. Chin. Med. Assoc. 67 (5), 245–247.
Honda, A., Ikegami, T., Nakamuta, M., Miyazaki, T., Iwamoto, J., Hirayama, T., et al. (2013). Anticholestatic effects of bezafibrate in patients with primary biliary cirrhosis treated with ursodeoxycholic acid. HEPATOLOGY 57 (5), 1931–1941. doi:10.1002/hep.26018
Hosonuma, K., Sato, K., Yamazaki, Y., Yanagisawa, M., Hashizume, H., Horiguchi, N., et al. (2015). A prospective randomized controlled study of long-term combination therapy using ursodeoxycholic acid and bezafibrate in patients with primary biliary cirrhosis and dyslipidemia. Am. J. Gastroenterol. 110 (3), 423–431. doi:10.1038/ajg.2015.20
Hua, H., Dai, M., Luo, Y., Lin, H., Xu, G., Hu, X., et al. (2019). Basal PPARα inhibits bile acid metabolism adaptation in chronic cholestatic model induced by α-naphthylisothiocyanate. Toxicol. Lett. 300, 31–39. doi:10.1016/j.toxlet.2018.10.015
Huang, D., Zhao, Q., Liu, H., Guo, Y., and Xu, H. (2016). PPAR-Alpha agonist WY-14643 inhibits LPS-induced inflammation in synovial fibroblasts via NF-kB pathway. J. Mol. Neurosci. 59 (4), 544–553. doi:10.1007/s12031-016-0775-y
Huang, T. H., Tran, V. H., Roufogalis, B. D., Li, Y., and Gypenoside, X. L. I. X. (2007). Gypenoside XLIX, a naturally occurring PPAR-alpha activator, inhibits cytokine-induced vascular cell adhesion molecule-1 expression and activity in human endothelial cells. Eur. J. Pharmacol. 565 (1-3), 158–165. doi:10.1016/j.ejphar.2007.03.013
Hunt, M. C., Yang, Y. Z., Eggertsen, G., Carneheim, C. M., Gafvels, M., Einarsson, C., et al. (2000). The peroxisome proliferator-activated receptor alpha (PPARalpha) regulates bile acid biosynthesis. J. Biol. Chem. 275 (37), 28947–28953. doi:10.1074/jbc.M002782200
Ip, E., Farrell, G. C., Robertson, G., Hall, P., Kirsch, R., Leclercq, I., et al. (2003). Central role of PPARalpha-dependent hepatic lipid turnover in dietary steatohepatitis in mice. Hepatology 38 (1), 123–132. doi:10.1053/jhep.2003.50307
Issemann, I., and Green, S. (1991). Cloning of novel members of the steroid hormone receptor superfamily. J. Steroid Biochem. Mol. Biol. 40 (1-3), 263–269. doi:10.1016/0960-0760(91)90191-7
Ito, M., Yamanashi, Y., Toyoda, Y., Izumi-Nakaseko, H., Oda, S., Sugiyama, A., et al. (2013). Disruption of Stard10 gene alters the PPARα-mediated bile acid homeostasis.. Biochim. Biophys. Acta 1831 (2), 459–468. doi:10.1016/j.bbalip.2012.11.008
Jaeschke, H. (1997). Cellular adhesion molecules: Regulation and functional significance in the pathogenesis of liver diseases. Am. J. Physiol. 273 (3 Pt 1), G602–G611. doi:10.1152/ajpgi.1997.273.3.G602
Jiang, Z., Liu, X., Yuan, Z., He, H., Wang, J., Zhang, X., et al. (2019). Discovery of a novel selective dual peroxisome proliferator-activated receptor α/δ agonist for the treatment of primary biliary cirrhosis. ACS Med. Chem. Lett. 10 (7), 1068–1073. doi:10.1021/acsmedchemlett.9b00189
Jiao, M. J., Zhou, L., Ren, F., Wang, Y. D., Shen, C., Duan, Z. P., et al. (2016). [Cellular and molecular mechanisms of anti-inflammatory effect of peroxisome proliferator-activated receptor α]. Zhonghua Gan Zang Bing Za Zhi 24 (12), 916–920. doi:10.3760/cma.j.issn.1007-3418.2016.12.008
Jin, F., Cheng, D., Tao, J. Y., Zhang, S. L., Pang, R., Guo, Y. J., et al. (2013). Anti-inflammatory and anti-oxidative effects of corilagin in a rat model of acute cholestasis. BMC Gastroenterol. 13, 79. doi:10.1186/1471-230X-13-79
Kalaany, N. Y., and Mangelsdorf, D. J. (2006). LXRS and FXR: The yin and yang of cholesterol and fat metabolism. Annu. Rev. Physiol. 68, 159–191. doi:10.1146/annurev.physiol.68.033104.152158
Keitel, V., Droge, C., and Haussinger, D. (2019). Targeting FXR in cholestasis. Handb. Exp. Pharmacol. 256, 299–324. doi:10.1007/164_2019_231
Kim, N. D., Moon, J. O., Slitt, A. L., and Copple, B. L. (2006). Early growth response factor-1 is critical for cholestatic liver injury. Toxicol. Sci. 90 (2), 586–595. doi:10.1093/toxsci/kfj111
Kodali, P., Wu, P., Lahiji, P. A., Brown, E. J., and Maher, J. J. (2006). ANIT toxicity toward mouse hepatocytes in vivo is mediated primarily by neutrophils via CD18. Am. J. Physiol. Gastrointest. Liver Physiol. 291 (2), G355–G363. doi:10.1152/ajpgi.00458.2005
Kok, T., Bloks, V. W., Wolters, H., Havinga, R., Jansen, P. L., Staels, B., et al. (2003). Peroxisome proliferator-activated receptor alpha (PPARalpha)-mediated regulation of multidrug resistance 2 (Mdr2) expression and function in mice. Biochem. J. 369 (Pt 3), 539–547. doi:10.1042/BJ20020981
Korbecki, J., Bobinski, R., and Dutka, M. (2019). Self-regulation of the inflammatory response by peroxisome proliferator-activated receptors. Inflamm. Res. 68 (6), 443–458. doi:10.1007/s00011-019-01231-1
Lemoinne, S., Pares, A., Reig, A., Ben, B. K., Kemgang, F. A., Gaouar, F., et al. (2018). Primary sclerosing cholangitis response to the combination of fibrates with ursodeoxycholic acid: French-Spanish experience. Clin. Res. Hepatol. Gastroenterol. 42 (6), 521–528. doi:10.1016/j.clinre.2018.06.009
Lens, S., Leoz, M., Nazal, L., Bruguera, M., and Pares, A. (2014). Bezafibrate normalizes alkaline phosphatase in primary biliary cirrhosis patients with incomplete response to ursodeoxycholic acid. Liver Int. 34 (2), 197–203. doi:10.1111/liv.12290
Leuschner, M., Dietrich, C. F., You, T., Seidl, C., Raedle, J., Herrmann, G., et al. (2000). Characterisation of patients with primary biliary cirrhosis responding to long term ursodeoxycholic acid treatment. Gut 46 (1), 121–126. doi:10.1136/gut.46.1.121
Levy, C., Peter, J. A., Nelson, D. R., Keach, J., Petz, J., Cabrera, R., et al. (2011). Pilot study: Fenofibrate for patients with primary biliary cirrhosis and an incomplete response to ursodeoxycholic acid. Aliment. Pharmacol. Ther. 33 (2), 235–242. doi:10.1111/j.1365-2036.2010.04512.x
Li, F., Patterson, A. D., Krausz, K. W., Tanaka, N., and Gonzalez, F. J. (2012). Metabolomics reveals an essential role for peroxisome proliferator-activated receptor alpha in bile acid homeostasis. J. Lipid Res. 53 (8), 1625–1635. doi:10.1194/jlr.M027433
Li, X., Yuan, Z., Liu, R., Hassan, H. M., Yang, H., Sun, R., et al. (2016). UDCA and CDCA alleviate 17α-ethinylestradiol-induced cholestasis through PKA-AMPK pathways in rats. Toxicol. Appl. Pharmacol. 311, 12–25. doi:10.1016/j.taap.2016.10.011
Liberopoulos, E. N., Florentin, M., Elisaf, M. S., Mikhailidis, D. P., and Tsianos, E. (2010). Fenofibrate in primary biliary cirrhosis: A pilot study. Open cardiovasc. Med. J. 4, 120–126. doi:10.2174/1874192401004010120
Lim, K. H., and Staudt, L. M. (2013). Toll-like receptor signaling. Cold Spring Harb. Perspect. Biol. 5 (1), a011247. doi:10.1101/cshperspect.a011247
Lindor, K. D., Gershwin, M. E., Poupon, R., Kaplan, M., Bergasa, N. V., and Heathcote, E. J. (2009). Primary biliary cirrhosis. HEPATOLOGY 50 (1), 291–308. doi:10.1002/hep.22906
Lu, Z., Li, S., Luo, J., Luo, Y., Dai, M., Zheng, X., et al. (2021). Fenofibrate reverses liver fibrosis in cholestatic mice induced by alpha-naphthylisothiocyanate. PHARMAZIE 76 (2), 103–108. doi:10.1691/ph.2021.0988
Luo, M., Tan, Z., Dai, M., Song, D., Lin, J., Xie, M., et al. (2017). Dual action of peroxisome proliferator-activated receptor alpha in perfluorodecanoic acid-induced hepatotoxicity. Arch. Toxicol. 91 (2), 897–907. doi:10.1007/s00204-016-1779-7
Marrapodi, M., and Chiang, J. Y. (2000). Peroxisome proliferator-activated receptor α (PPARα) and agonist inhibit cholesterol 7α-hydroxylase gene (CYP7A1) transcription. J. Lipid Res. 41 (4), 514–520. doi:10.1016/s0022-2275(20)32398-1
Marx, N., Sukhova, G. K., Collins, T., Libby, P., and Plutzky, J. (1999). PPARalpha activators inhibit cytokine-induced vascular cell adhesion molecule-1 expression in human endothelial cells. CIRCULATION 99 (24), 3125–3131. doi:10.1161/01.cir.99.24.3125
Moffit, J. S., Aleksunes, L. M., Maher, J. M., Scheffer, G. L., Klaassen, C. D., Manautou, J. E., et al. (2006). Induction of hepatic transporters multidrug resistance-associated proteins (Mrp) 3 and 4 by clofibrate is regulated by peroxisome proliferator-activated receptor alpha. J. Pharmacol. Exp. Ther. 317 (2), 537–545. doi:10.1124/jpet.105.093765
Mraz, M., Lacinova, Z., Kavalkova, P., Haluzikova, D., Trachta, P., Drapalova, J., et al. (2011). Serum concentrations of fibroblast growth factor 19 in patients with obesity and type 2 diabetes mellitus: The influence of acute hyperinsulinemia, very-low calorie diet and PPAR-alpha agonist treatment. Physiol. Res. 60 (4), 627–636. doi:10.33549/physiolres.932099
Nuclear Receptors Nomenclature Committee (1999). A unified nomenclature system for the nuclear receptor superfamily. Cell 97 (2), 161–163. doi:10.1016/s0092-8674(00)80726-6
Okamura, A., Koyanagi, S., Dilxiat, A., Kusunose, N., Chen, J. J., Matsunaga, N., et al. (2014). Bile acid-regulated peroxisome proliferator-activated receptor-α (PPARα) activity underlies circadian expression of intestinal peptide absorption transporter PepT1/Slc15a1. J. Biol. Chem. 289 (36), 25296–25305. doi:10.1074/jbc.M114.577023
Oscarsson, J., Onnerhag, K., Riserus, U., Sunden, M., Johansson, L., Jansson, P. A., et al. (2018). Effects of free omega-3 carboxylic acids and fenofibrate on liver fat content in patients with hypertriglyceridemia and non-alcoholic fatty liver disease: A double-blind, randomized, placebo-controlled study. J. Clin. Lipidol. 12 (6), 1390–1403. e4. doi:10.1016/j.jacl.2018.08.003
Padda, M. S., Sanchez, M., Akhtar, A. J., and Boyer, J. L. (2011). Drug-induced cholestasis. Hepatology 53 (4), 1377–1387. doi:10.1002/hep.24229
Pan, P. H., Lin, S. Y., Ou, Y. C., Chen, W. Y., Chuang, Y. H., Yen, Y. J., et al. (2010). Stearic acid attenuates cholestasis-induced liver injury. Biochem. Biophys. Res. Commun. 391 (3), 1537–1542. doi:10.1016/j.bbrc.2009.12.119
Park, Y. J., Kim, S. C., Kim, J., Anakk, S., Lee, J. M., Tseng, H. T., et al. (2011). Dissociation of diabetes and obesity in mice lacking orphan nuclear receptor small heterodimer partner. J. Lipid Res. 52 (12), 2234–2244. doi:10.1194/jlr.M016048
Patel, A., and Seetharam, A. (2016). Primary biliary cholangitis: Disease pathogenesis and implications for established and novel therapeutics. J. Clin. Exp. Hepatol. 6 (4), 311–318. doi:10.1016/j.jceh.2016.10.001
Patel, D. D., Knight, B. L., Soutar, A. K., Gibbons, G. F., and Wade, D. P. (2000). The effect of peroxisome-proliferator-activated receptor-alpha on the activity of the cholesterol 7 alpha-hydroxylase gene. Biochem. J. 351 Pt 3, 747–753. doi:10.1042/bj3510747
Pawlak, M., Lefebvre, P., and Staels, B. (2015). Molecular mechanism of PPARα action and its impact on lipid metabolism, inflammation and fibrosis in non-alcoholic fatty liver disease. J. Hepatol. 62 (3), 720–733. doi:10.1016/j.jhep.2014.10.039
Piechota, J., and Jelski, W. (2020). Intrahepatic cholestasis in pregnancy: Review of the literature. J. Clin. Med. 9 (5), E1361. doi:10.3390/jcm9051361
Pineda, T. I., Claudel, T., Duval, C., Kosykh, V., Fruchart, J. C., Staels, B., et al. (2003). Bile acids induce the expression of the human peroxisome proliferator-activated receptor alpha gene via activation of the farnesoid X receptor. Mol. Endocrinol. 17 (2), 259–272. doi:10.1210/me.2002-0120
Post, S. M., Duez, H., Gervois, P. P., Staels, B., Kuipers, F., Princen, H. M., et al. (2001). Fibrates suppress bile acid synthesis via peroxisome proliferator-activated receptor-alpha-mediated downregulation of cholesterol 7alpha-hydroxylase and sterol 27-hydroxylase expression. Arterioscler. Thromb. Vasc. Biol. 21 (11), 1840–1845. doi:10.1161/hq1101.098228
Preidis, G. A., Kim, K. H., and Moore, D. D. (2017). Nutrient-sensing nuclear receptors PPARα and FXR control liver energy balance. J. Clin. Invest. 127 (4), 1193–1201. doi:10.1172/JCI88893
Reig, A., Sese, P., and Pares, A. (2018). Effects of bezafibrate on outcome and pruritus in primary biliary cholangitis with suboptimal ursodeoxycholic acid response. Am. J. Gastroenterol. 113 (1), 49–55. doi:10.1038/ajg.2017.287
Santos, M. H., Higuchi, M. L., Tucci, P. J., Garavelo, S. M., Reis, M. M., Antonio, E. L., et al. (2016). Previous exercise training increases levels of PPAR-alpha in long-term post-myocardial infarction in rats, which is correlated with better inflammatory response. Clin. (Sao Paulo) 71 (3), 163–168. doi:10.6061/clinics/2016(03)08
Satrom, K., and Gourley, G. (2016). Cholestasis in preterm infants. Clin. Perinatol. 43 (2), 355–373. doi:10.1016/j.clp.2016.01.012
Senekeo-Effenberger, K., Chen, S., Brace-Sinnokrak, E., Bonzo, J. A., Yueh, M. F., Argikar, U., et al. (2007). Expression of the human UGT1 locus in transgenic mice by 4-chloro-6-(2, 3-xylidino)-2-pyrimidinylthioacetic acid (WY-14643) and implications on drug metabolism through peroxisome proliferator-activated receptor alpha activation. Drug Metab. Dispos. 35 (3), 419–427. doi:10.1124/dmd.106.013243
Shi, C., Min, L., Yang, J., Dai, M., Song, D., Hua, H., et al. (2017). Peroxisome proliferator-activated receptor alpha activation suppresses cytochrome P450 induction potential in mice treated with gemfibrozil. Basic Clin. Pharmacol. Toxicol. 121 (3), 169–174. doi:10.1111/bcpt.12794
Shi, R. L., Zhao, C. X., Zhu, H. B., Yang, Y., Wang, S. L., Jiang, L. L., et al. (2005). Relationship between the increase of hepatic D-bifunctional protein activity and bile acid biosynthesis in rats. Zhongguo Yi Xue Ke Xue Yuan Xue Bao. 27 (3), 321–324.
Sinal, C. J., Yoon, M., and Gonzalez, F. J. (2001). Antagonism of the actions of peroxisome proliferator-activated receptor-alpha by bile acids. J. Biol. Chem. 276 (50), 47154–47162. doi:10.1074/jbc.M107000200
Smit, J. J., Schinkel, A. H., Oude, E. R., Groen, A. K., Wagenaar, E., van Deemter, L., et al. (1993). Homozygous disruption of the murine mdr2 P-glycoprotein gene leads to a complete absence of phospholipid from bile and to liver disease. Cell 75 (3), 451–462. doi:10.1016/0092-8674(93)90380-9
Sorda, J. A., Gonzalez, B. E., Barreyro, F. J., Avagnina, A., Carballo, P., Paes, D. L. A., et al. (2021). Bezafibrate therapy in primary biliary cholangitis refractory to ursodeoxycholic acid: A longitudinal study of paired liver biopsies at 5 years of follow up. Aliment. Pharmacol. Ther. 54 (9), 1202–1212. doi:10.1111/apt.16618
Spivey, J. R., Bronk, S. F., and Gores, G. J. (1993). Glycochenodeoxycholate-induced lethal hepatocellular injury in rat hepatocytes. Role of ATP depletion and cytosolic free calcium. J. Clin. Invest. 92 (1), 17–24. doi:10.1172/JCI116546
Stedman, C., Liddle, C., Coulter, S., Sonoda, J., Alvarez, J. G., Evans, R. M., et al. (2006). Benefit of farnesoid X receptor inhibition in obstructive cholestasis. Proc. Natl. Acad. Sci. U. S. A. 103 (30), 11323–11328. doi:10.1073/pnas.0604772103
Takeuchi, Y., Ikeda, F., Fujioka, S., Takaki, T., Osawa, T., Yasunaka, T., et al. (2011). Additive improvement induced by bezafibrate in patients with primary biliary cirrhosis showing refractory response to ursodeoxycholic acid. J. Gastroenterol. Hepatol. 26 (9), 1395–1401. doi:10.1111/j.1440-1746.2011.06737.x
Tanaka, A., Hirohara, J., Nakanuma, Y., Tsubouchi, H., and Takikawa, H. (2015). Biochemical responses to bezafibrate improve long-term outcome in asymptomatic patients with primary biliary cirrhosis refractory to UDCA. J. Gastroenterol. 50 (6), 675–682. doi:10.1007/s00535-014-0998-z
Taniguchi, K., Karin, M., and Nf-kappaB, (2018). NF-κB, inflammation, immunity and cancer: Coming of age. Nat. Rev. Immunol. 18 (5), 309–324. doi:10.1038/nri.2017.142
Thomas, M., Burk, O., Klumpp, B., Kandel, B. A., Damm, G., Weiss, T. S., et al. (2013). Direct transcriptional regulation of human hepatic cytochrome P450 3A4 (CYP3A4) by peroxisome proliferator-activated receptor alpha (PPARα). Mol. Pharmacol. 83 (3), 709–718. doi:10.1124/mol.112.082503
Tiao, M. M., Lin, T. K., Kuo, F. Y., Huang, C. C., Du, Y. Y., Chen, C. L., et al. (2007). Early stage of biliary atresia is associated with significant changes in 8-hydroxydeoxyguanosine and mitochondrial copy number. J. Pediatr. Gastroenterol. Nutr. 45 (3), 329–334. doi:10.1097/MPG.0b013e3180cc2c0f
Ticho, A. L., Malhotra, P., Dudeja, P. K., Gill, R. K., and Alrefai, W. A. (2019). Intestinal absorption of bile acids in health and disease. Compr. Physiol. 10 (1), 21–56. doi:10.1002/cphy.c190007
van de Wiel, S., Porteiro, B., Belt, S. C., Vogels, E., Bolt, I., Vermeulen, J., et al. (2022). Differential and organ-specific functions of organic solute transporter alpha and beta in experimental cholestasis. JHEP Rep. 4 (5), 100463. doi:10.1016/j.jhepr.2022.100463
Wang, L., Ma, L., Lin, Y., Liu, X., Xiao, L., Zhang, Y., et al. (2018). Leflunomide increases hepatic exposure to methotrexate and its metabolite by differentially regulating multidrug resistance-associated protein mrp2/3/4 transporters via peroxisome proliferator-activated receptor alpha activation. Mol. Pharmacol. 93 (6), 563–574. doi:10.1124/mol.117.110593
Woolbright, B. L., Antoine, D. J., Jenkins, R. E., Bajt, M. L., Park, B. K., Jaeschke, H., et al. (2013). Plasma biomarkers of liver injury and inflammation demonstrate a lack of apoptosis during obstructive cholestasis in mice. Toxicol. Appl. Pharmacol. 273 (3), 524–531. doi:10.1016/j.taap.2013.09.023
Woolbright, B. L., and Jaeschke, H. (2012). Novel insight into mechanisms of cholestatic liver injury. World J. Gastroenterol. 18 (36), 4985–4993. doi:10.3748/wjg.v18.i36.4985
Wu, C. T., Eiserich, J. P., Ansari, A. A., Coppel, R. L., Balasubramanian, S., Bowlus, C. L., et al. (2003). Myeloperoxidase-positive inflammatory cells participate in bile duct damage in primary biliary cirrhosis through nitric oxide-mediated reactions. Hepatology 38 (4), 1018–1025. doi:10.1053/jhep.2003.50407
Xie, C., Takahashi, S., Brocker, C. N., He, S., Chen, L., Xie, G., et al. (2019). Hepatocyte peroxisome proliferator-activated receptor alpha regulates bile acid synthesis and transport. Biochim. Biophys. Acta. Mol. Cell Biol. Lipids 1864 (10), 1396–1411. doi:10.1016/j.bbalip.2019.05.014
Xu, S., Chen, Y., Lin, H., Pi, H., Zhang, N., Zhao, C., et al. (2012). Damage to mtDNA in liver injury of patients with extrahepatic cholestasis: The protective effects of mitochondrial transcription factor A. Free Radic. Biol. Med. 52 (9), 1543–1551. doi:10.1016/j.freeradbiomed.2012.01.007
Yang, Y. M., Kim, S. Y., and Seki, E. (2019). Inflammation and liver cancer: Molecular mechanisms and therapeutic targets. Semin. Liver Dis. 39 (1), 26–42. doi:10.1055/s-0038-1676806
Younossi, Z. M., Ratziu, V., Loomba, R., Rinella, M., Anstee, Q. M., Goodman, Z., et al. (2019). Obeticholic acid for the treatment of non-alcoholic steatohepatitis: Interim analysis from a multicentre, randomised, placebo-controlled phase 3 trial. Lancet 394 (10215), 2184–2196. doi:10.1016/S0140-6736(19)33041-7
Zhang, S., Yu, M., Guo, F., Yang, X., Chen, Y., Ma, C., et al. (2020). Rosiglitazone alleviates intrahepatic cholestasis induced by α-naphthylisothiocyanate in mice: The role of circulating 15-deoxy-Δ12, 14 -PGJ2 and Nogo. Br. J. Pharmacol. 177 (5), 1041–1060. doi:10.1111/bph.14886
Zhang, Y., Csanaky, I. L., Cheng, X., Lehman-McKeeman, L. D., and Klaassen, C. D. (2012). Organic anion transporting polypeptide 1a1 null mice are sensitive to cholestatic liver injury. Toxicol. Sci. 127 (2), 451–462. doi:10.1093/toxsci/kfs123
Zhang, Y., Lickteig, A. J., Csanaky, I. L., and Klaassen, C. D. (2017). Editor's highlight: Clofibrate decreases bile acids in livers of male mice by increasing biliary bile acid excretion in a PPARα-dependent manner. Toxicol. Sci. 160 (2), 351–360. doi:10.1093/toxsci/kfx191
Zhang, Y., Lu, Y., Ji, H., and Li, Y. (2019). Anti-inflammatory, anti-oxidative stress and novel therapeutic targets for cholestatic liver injury. Biosci. Trends 13 (1), 23–31. doi:10.5582/bst.2018.01247
Zhao, Q., Yang, R., Wang, J., Hu, D. D., and Li, F. (2017). PPARα activation protects against cholestatic liver injury. Sci. Rep. 7 (1), 9967. doi:10.1038/s41598-017-10524-6
Zhou, X., Cao, L., Jiang, C., Xie, Y., Cheng, X., Krausz, K. W., et al. (2014). PPARα-UGT axis activation represses intestinal FXR-FGF15 feedback signalling and exacerbates experimental colitis.. Nat. Commun 5, 4573. doi:10.1038/ncomms5573
Zhu, L., Hayen, A., and Bell, K. (2020). Legacy effect of fibrate add-on therapy in diabetic patients with dyslipidemia: A secondary analysis of the ACCORDION study. Cardiovasc. Diabetol. 19 (1), 28. doi:10.1186/s12933-020-01002-x
Zollner, G., Fickert, P., Fuchsbichler, A., Silbert, D., Wagner, M., Arbeiter, S., et al. (2003). Role of nuclear bile acid receptor, FXR, in adaptive ABC transporter regulation by cholic and ursodeoxycholic acid in mouse liver, kidney and intestine. J. Hepatol. 39 (4), 480–488. doi:10.1016/s0168-8278(03)00228-9
Keywords: cholestasis, inflammation, liver injury, peroxisome proliferator-activated receptor alpha, therapeutic target
Citation: Ye X, Zhang T and Han H (2022) PPARα: A potential therapeutic target of cholestasis. Front. Pharmacol. 13:916866. doi: 10.3389/fphar.2022.916866
Received: 10 April 2022; Accepted: 29 June 2022;
Published: 18 July 2022.
Edited by:
David Sacerdoti, University of Verona, ItalyReviewed by:
Sagnik Chatterjee, Ferring Pharmaceuticals, DenmarkAnnarosa Floreani, IRCSS Negrar, Italy
Copyright © 2022 Ye, Zhang and Han. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tong Zhang, emhhbmd0ZG1qQGhvdG1haWwuY29t; Han Han, cGFzaGFuaGFuQDEyNi5jb20=