 Seegehalli M. Anil†
Seegehalli M. Anil† Hadar Peeri
Hadar Peeri Hinanit Koltai
Hinanit Koltai
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Pharmacol., 09 May 2022
Sec. Experimental Pharmacology and Drug Discovery
Volume 13 - 2022 | https://doi.org/10.3389/fphar.2022.908198
This article is part of the Research TopicMedicinal Cannabis: Evolution of Therapeutic Use, Future Approaches and Other ImplicationsView all 12 articles
Inflammation often develops from acute, chronic, or auto-inflammatory disorders that can lead to compromised organ function. Cannabis (Cannabis sativa) has been used to treat inflammation for millennia, but its use in modern medicine is hampered by a lack of scientific knowledge. Previous studies report that cannabis extracts and inflorescence inhibit inflammatory responses in vitro and in pre-clinical and clinical trials. The endocannabinoid system (ECS) is a modulator of immune system activity, and dysregulation of this system is involved in various chronic inflammations. This system includes cannabinoid receptor types 1 and 2 (CB1 and CB2), arachidonic acid-derived endocannabinoids, and enzymes involved in endocannabinoid metabolism. Cannabis produces a large number of phytocannabinoids and numerous other biomolecules such as terpenes and flavonoids. In multiple experimental models, both in vitro and in vivo, several phytocannabinoids, including Δ9-tetrahydrocannabinol (THC), cannabidiol (CBD) and cannabigerol (CBG), exhibit activity against inflammation. These phytocannabinoids may bind to ECS and/or other receptors and ameliorate various inflammatory-related diseases by activating several signaling pathways. Synergy between phytocannabinoids, as well as between phytocannabinoids and terpenes, has been demonstrated. Cannabis activity can be improved by selecting the most active plant ingredients (API) while eliminating parts of the whole extract. Moreover, in the future cannabis components might be combined with pharmaceutical drugs to reduce inflammation.
The immune system consists of both adaptive and innate immunity. Innate immunity is the rapid and non-specific response to pathogens mediated by myeloid cells and natural killer (NK) cells. On the other hand, adaptive immunity is a slower but specific response that generates immunological memory, involving the activation of B and T lymphocytes (Netea et al., 2019). During normal inflammation, innate immunity is activated within minutes to hours as a first line of defense against pathogen infection, followed by the elimination of the threats carried out by both the innate and the adaptive immune responses (Netea et al., 2019). Ending inflammation and returning to homeostasis is a process known as resolution. However, failure to remove the inciting stimulus efficiently can lead to the development of chronic inflammation and progression of tissue damage (Feehan and Gilroy, 2019). This kind of chronic, unresolved inflammation contributes significantly to various pathogeneses, including that of asthma (Pothen et al., 2015), COVID-19 (Effenberger et al., 2021), atherosclerosis (Galkina and Ley, 2009), chronic obstructive pulmonary disease (Sevenoaks and Stockley, 2006), inflammatory bowel disease (Actis et al., 2019), neurodegenerative disease (Perry, 2004), multiple sclerosis (Sá, 2012) and rheumatoid arthritis (Masoumi et al., 2021).
Cannabis (Cannabis sativa) has been used as medicine for the treatment of inflammation for millennia, but its use in modern medicine has been hampered by a lack of scientific knowledge (Ryz et al., 2017). Previous studies reported that cannabis extracts and inflorescence inhibited inflammatory responses in vitro and in pre-clinical and clinical studies. For example, a high-CBD cannabis ethanolic extract reduced the release of skin inflammation mediators in keratinocytes (Sangiovanni et al., 2019). Similarly, a study on a mouse model of colitis showed that oral or intraperitoneally treatment with high-CBD cannabis extract led to a reduction in intestinal inflammation and hypermotility, in contrast to pure CBD treatment at matched doses (Pagano et al., 2016). Moreover, two clinical trials on patients with Crohn’s disease reported that daily treatment with THC-rich cannabis inflorescence had beneficial effects against the disease symptoms with no significant side effects and reduced the need for other medications (Naftali et al., 2011; Naftali et al., 2013). In another clinical trial, daily cannabis treatment was associated with lower levels of pro-inflammatory biomarkers in cerebral fluid (CSF) of HIV patients (Watson et al., 2021).
Great efforts have been made to suppress chronic inflammation. Cannabis and its compounds were shown to have anti-inflammatory activity (see Appendix A for methodology), but to exploit the full potential of cannabis it is important to define the active molecules and understand the cellular and molecular mechanisms that underlie its anti-inflammatory activity.
Monocytes are the major starting entities of inflammation. Once released from bone marrow, monocytes migrate through the blood into various tissues and undergo the tissue-specific maturation required to become inflammatory macrophages that respond to infection, injury, or damage. The various sub-populations of activated macrophages may differ in morphology, release of inflammatory mediators and functional properties, but in inflammation they have three major functions: phagocytosis, antigen presentation and immunomodulation (Fujiwara and Kobayashi, 2005). The process of inflammation is orchestrated via inflammatory mediators. Pro-inflammatory cytokines, such as tumor necrosis factor alpha (TNF-α) and interleukin (IL)-1β are released from activated macrophages in response to infection (Figure 1; Abdulkhaleq et al., 2018). TNF-α and IL-1β act through specific cell membrane-bound receptors and participate in the recruitment of polymorphonuclear neutrophils (PMNs) into the site of infection and their activation (Hackel et al., 2021).
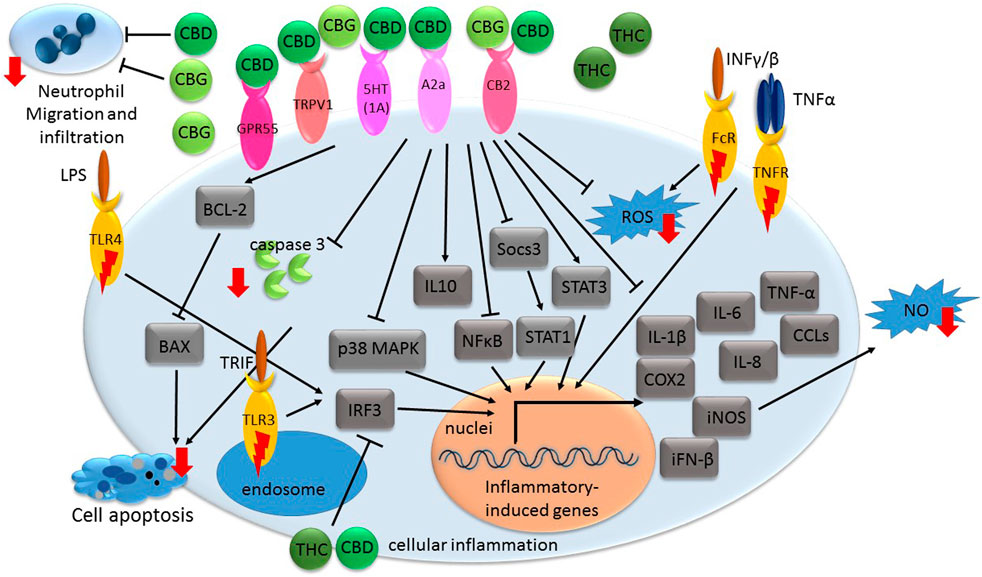
FIGURE 1. A general illustration of some of the signaling pathways suggested being associated with phytocannabinoid-mediated inflammation suppression. Receptors with inflammatory-inducing activity are marked with a red lightning bolt. Other receptors interact with phytocannabinoids to convey anti-inflammatory responses. Genes or proteins are designated in rectangular boxes. Red arrows denote reduction in biological processes or components following cannabinoid treatments. CBD-cannabidiol; CBG-cannabigerol; THC- Δ9-tetrahydrocannabinol; CB- cannabinoid receptor; GPR- G protein-coupled receptor; TRPV- transient receptor potential vanilloid; A2a-adenosine receptor; iFN- interferon; TNF- tumor necrosis factor; CCL- C-C motif chemokine; IL-interleukin; COX-cyclooxygenase; iNOS- nitric oxide synthase; ROS- reactive oxygen species; NO- nitric oxide; MAPK- mitogen-activated protein kinase; LPS- bacterial lipopolysaccharide; NFκB- nuclear factor kappa B; IRF3- regulatory factor 3; FcR- Fc receptor; TNFR- TNF receptor; INF- interferon; 5HT(1A)- serotonin receptor; TLR-toll-like receptor; TRIF- Toll-Interleukin-1 Receptor (TIR)-domain-containing adaptor-inducing interferon-β.
TNF-α facilitates the release of other pro-inflammatory cytokines from immune effector cells, including interferon alpha (IFN-α), interferon gamma (IFN-γ), IL-1β, IL-6, IL-8, Transforming growth factor beta (TGF-β) and chemokines (Silva et al., 2019). Further, in cases of enhanced inflammation, when the cell is stimulated, typically by bacterial lipopolysaccharide (LPS) or pro-inflammatory cytokines, there is induction of inducible nitric oxide synthase (iNOS). An increase in iNOS levels generates significant amounts of nitric oxide (NO) radicals or cyclooxygenase 2 (COX2); COX2 catalyzes the conversion of arachidonic acid to prostaglandins (PGs), prostacyclin and thromboxane A2 (Salvemini et al., 2013; Cinelli et al., 2020).
The signal transduction of inflammatory responses involves several signaling pathways including mitogen-activated protein kinase (MAPK), toll-like receptor (TLR), Janus kinase/signal transducers and activators of transcription (JAK-STAT), and nuclear factor kappa B (NFκB) pathways (Figure 1; Zhao et al., 2021). The activation of these pathways involves a series of phosphorylation events leading to the induction of various anti-apoptotic target genes and the expression of cytokines, chemokines, and adhesion molecules (Taniguchi and Karin, 2018; Fitzgerald and Kagan, 2020). Moreover, during inflammatory processes, reactive oxygen species (ROS) are commonly multiplied and can contribute to host cell and organ damage. Further, intracellular redox changes induced by ROS augment NF-κB activation through the phosphorylation and degradation of IκB by increasing IkB kinase ß (IKK) or Akt kinase activity (Haddad, 2002).
Resolution of inflammation may involve increased production of IL-10, among others. IL-10 is an anti-inflammatory cytokine, which inhibits the release of lipid mediators and pro-inflammatory cytokines (e.g., IL-1β, IL-6, and TNF-α; Figure 1; Panigrahy et al., 2021).
The endocannabinoid system (ECS) is a modulator of multiple physiological activities, including in the nervous, endocrine, immune, blood circulation, gastrointestinal tract and reproductive systems (Di Marzo et al., 1998). Accordingly, dysregulation of the ECS is involved with various pathological conditions, including inflammation among others (Di Marzo and Piscitelli, 2015; Hillard, 2018), whereas therapeutic modulation of ECS activity has beneficial effects on various medical conditions, including those associated with inflammation (Ambrose and Simmons, 2019; Giacobbe et al., 2021). ECS is involved in both innate and adaptive immunity and in several chronic inflammatory diseases (Chiurchiù et al., 2015). ECS includes cannabinoid receptors types 1 and 2 (CB1 and CB2, respectively) and multiple other receptors such as the peroxisome proliferator-activated receptors (PPARs) and ion channels (e.g., the transient receptor potential ankyrin [TRPA] family and the transient receptor potential vanilloid [TRPV] family) (Biringer, 2021). Also included in the ECS are the receptors’ ligand, arachidonic acid derived endocannabinoids, and enzymes for endocannabinoid metabolism (Di Marzo et al., 1998).
Most immune cells express endocannabinoids, the enzymes regulating their biosynthesis and degradation, and endocannabinoid receptors (Chiurchiù et al., 2015). Both CB1 and CB2 are expressed in immune cells, with CB2 being expressed 10–100 times higher than CB1 in these cells (Jean-Gilles et al., 2010; Rahaman and Ganguly, 2021). Moreover, CB receptor activation regulates anti-inflammatory responses. For example, activation of CB2 receptors by its agonist inhibited the release of the pro-inflammatory cytokine IL-12 and IL-23 and enhanced the release of the anti-inflammatory cytokine IL-10 from cultured activated macrophages. This study suggested that the inhibitory effect of CB2 on IL-12 production was mediated by ERK1/2-MAPK (Correa et al., 2009).
In another example, a CB2 receptor agonist reduced in human peripheral blood mononuclear cells LPS-induced ERK1/2 and NF-kB-p65 phosphorylation and release of the pro-inflammatory cytokines TNF-α, IL-1β, IL-6 and IL-8 (Capozzi et al., 2021). A selective/inverse agonist of CB2 induced the differentiation of Th0 cells into regulatory T cells (Treg) cell phenotypes in a naïve CD4+ T lymphocyte population isolated from a mouse spleen. The Treg phenotype is important for suppressing immune response by inhibiting T cell proliferation and cytokine production. The Treg phenotype was induced via P38 phosphorylation and STAT5A activation and was characterized by the expression of FoxP3, TGF-β and IL-10. Accordingly, treatment with this CB2 selective/inverse agonist reduced colitis severity in vivo (Gentili et al., 2019).
Cannabis produces a large number of phytocannabinoids (Hanuš et al., 2016). Phytocannabinoids are aromatic oxygenated hydrocarbons, derived from meroterpenoids with a resorcinyl core structure with isoprenyl, alkyl or aralkyl substitutions. The characteristic alkyl side chain typically contains an odd number of carbon atoms (Hanuš et al., 2016; Gülck and Møller, 2020). They are produced in the plant in their acid form and are decarboxylated to the active form (Gülck and Møller, 2020). Among the phytocannabinoids, Δ9-trans-tetrahydrocannabinols (Δ9-THCs) and cannabidiols (CBDs) are the most abundant (Table 1). Cannabigerol (CBG) in its acid form (CBGA) serves as a core intermediate that diverges to provide the phytocannabinolic acids (Table 1; Hanuš et al., 2016; Tahir et al., 2021).
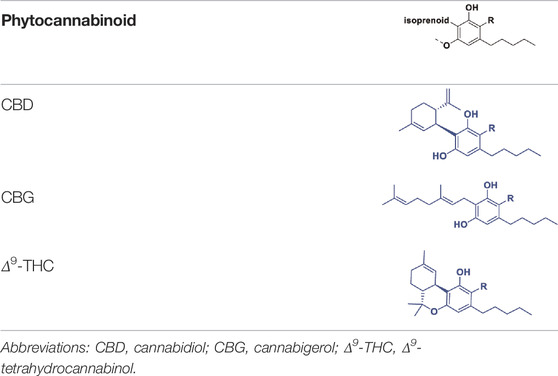
TABLE 1. Representative structures of three major phytocannabinoids.
In addition to phytocannabinoids, cannabis produces a plethora of non-cannabinoid constituents including a vast array of terpenes as the second-largest class of cannabis constituents (El Sohly et al., 2017). Cannabis biosynthesizes flavonoids as well, among them cannflavins, which are prenylated (C5) and geranylated (C10) flavones (Bautista et al., 2021).
CBD was demonstrated in multiple experimental models, in vitro and in vivo, to exert anti-inflammatory activities and ameliorate various inflammatory-related degenerative diseases. The mechanism of this anti-inflammatory activity is, however, not completely understood. CBD treatment of hypoxic-ischemic (HI) immature brains of newborn mice was shown to significantly reduce IL-6, TNF-α, COX-2 and iNOS expression in brain slices. This activity was suggested to be mediated via CB2 and adenosine A2A receptors (Figure 1; Castillo et al., 2010). Likewise, treatments of lipopolysaccharide-treated mice with a low dose of CBD decreased TNF-α production; this effect was abolished in A2A receptor knockout mice and reversed with an A2A adenosine receptor antagonist, supporting the notion that CBD may enhance adenosine signaling (Carrier et al., 2006). Further, in a murine model of acute lung injury, CBD, via the A2A adenosine receptor, significantly reduced leukocyte migration into the lungs and reduced the levels of albumin, TNF-α, IL-6 and other chemokines in bronchoalveolar lavage fluid (Figure 1; Ribeiro et al., 2012). CBD also reduced the activity of myeloperoxidase (MOP, an index of neutrophil infiltration) in lung tissue (Ribeiro et al., 2012).
In newborn pigs with HI brain injury, CBD administration reduced inflammation and prevented the increase in brain IL-1 levels. It also prevented the decrease in the number of viable neurons and the increase of excitotoxicity and oxidative stress. This activity was suggested to be mediated via CB2 and 5HT1A receptors (Figure 1; Pazos et al., 2013). In liver filtrate from mice with acute hepatitis, CBD was shown to trigger Myeloid-derived suppressor cells (MDSCs); these cells are regulators of the immune system that suppress T cell functions. MDSCs induction by CBD was mediated through activation of TRPV1 (Figure 1). CBD also significantly reduced blood levels of IL-2, TNF-α, IFN-γ, IL-6, IL-12, IL-17, MCP-1 and C-C motif chemokine (CCL)-11 in this model (Hegde et al., 2011).
CBD is also a selective antagonist of GPR55, another G protein-coupled receptor present in human macrophages (Figure 1). Pharmacological activation of GPR55 by its selective agonist O-1602 enhanced pro-inflammatory responses in macrophages-derived foam cells associated with a reduction in IL-10 levels and induction in TNF-α levels (Lanuti et al., 2015).
CBD treatment completely inhibited TNF-α production via p38 MAPK pathway (Figure 1) in microglial cells isolated from the retinas of newborn rats treated with endotoxin or LPS for acute ocular inflammation. In addition, LPS-treated rat retinas accumulated macrophages and activated microglia, increased levels of ROS and nitrotyrosine, and activated p38 MAPK and neuronal apoptosis. Treatment with CBD blocked all these effects (El-Remessy et al., 2008).
CBD decreases the production and release of IL-1β, IL-6 and IFN-β from LPS-activated microglial cells of BV-2 mice. CBD reduced the activity of the NF-κB pathway and the levels of IL-1β and IL-6. CBD also decreased Socs3 gene expression; Socs3 is a main negative regulator of STATs. In accordance, CBD treatment up-regulated the STAT3 transcription factor phosphorylation, needed for its activation (Figure 1; Kozela et al., 2010). However, NF-κB and STAT3 are likely to play important and in some cases, overlapping roles in pro-inflammatory and cancer processes (He & Karin, 2011). In contrast, CBD decreased the phosphorylation of the LPS-induced STAT1 transcription factor, a key player in pro-inflammatory processes that are IFN-β-dependent (Kozela et al., 2010).
The anti-inflammatory activity of CBG is less studied than that of CBD. Yet, several studies demonstrated significant anti-inflammatory activity of CBG. For example, CBG treatment was shown to reduce nitric oxide production in macrophages via the CB2 receptor and reduce ROS formation in intestinal epithelial cells and iNOS expression (Figure 1) in the inflamed colons. Treatment with CBG also reduced oedema in colon submucosa. This treatment also reduced the colon weight/length ratio; this ratio is a reliable marker of intestinal inflammation in a murine model of colitis glands (Borrelli et al., 2013). In addition, CBG decreased dinitrobenzene sulfonic acid (DNBS)-induced neutrophil infiltration, as evaluated by MOP activity (Borrelli et al., 2013).
In a study that characterized the anti-inflammatory properties of CBG on human skin cells in vitro, it was demonstrated that CBG treatment reduced ROS levels in human dermal fibroblasts, better than vitamin C. CBG also protected human epidermal keratinocytes by inhibiting pro-inflammatory cytokines that were released following induction using UVA, UVB or Cutibacterium acnes exposure, including TNF-α, IL-1β, IL-6 and IL-8 (Figure 1; Perez et al., 2022). Furthermore, the researchers performed a single-blind clinical study on 20 healthy volunteers with sodium lauryl sulfate (SLS)-induced contact dermatitis and found that topical application of 0.1% CBG serum showed significantly lower trans-epidermal water loss (TEWL) values compared to placebo and untreated sites. Moreover, the CBG serum reduced redness and inflammation following 48 h treatment, and after 2 weeks of application, the skin condition almost returned to baseline levels of visual grade (Perez et al., 2022).
Several studies have described the neuroprotective properties of CBG against inflammation. It was demonstrated that CBG pre-treatment of cultured motor neurons not only reduced the levels of pro-inflammatory cytokines, including IL-1β, TNF-α and IFN-γ (Figure 1), but also inhibited apoptosis in LPS-stimulated macrophages, via suppression of caspase-3 and Bax expression and induction of Bcl-2 levels (Gugliandolo et al., 2018). In addition, in a study that examined the effects of CBG on Huntington’s disease pathology in 3-nitropropionate model in vivo, it was found that treatment with the phytocannabinoid reduced neuronal death by half and significantly attenuated the upregulation of expression of COX-2, iNOS and pro-inflammatory cytokines such as TNF-α and IL-6 (Figure 1; Valdeolivas et al., 2015).
Several experiments suggest that THC has anti-inflammatory effects. For example, topical treatment of THC on DNFB-mediated allergic contact dermatitis in mice revealed that THC effectively decreased myeloid immune cell infiltration and contact allergic ear swelling (Gaffal et al., 2013). These anti-inflammatory effects were evident in both wild-type and CB1/2 receptor-deficient mice suggesting that these activities of THC were not mediated via CB1 or CB2 receptors. In addition, THC reduced the production by epidermal keratinocytes of CCL8 and CCL2 induced by IFNγ and the production of IFNγ by T cells (Figure 1). As a result, in a CB1/2 receptor-independent way, THC limited the recruitment of myeloid immune cells in vitro (Gaffal et al., 2013).
Interestingly, in LPS-induced macrophages, THC (and CBD) attenuated TLR3/4 signaling in a MyD88-independent manner (Fitzpatrick et al., 2020). TLR3 signaling is mediated via a toll-interleukin-1 receptor (TIR)-domain-containing adaptor-inducing interferon-β (TRIF). TLR4-induced expression of regulatory factor 3 (IRF3) activation, and CXCL10 and IFN-β were repressed by the THC and/or CBD (alone or in combination) treatments. However, these phytocannabinoid treatments did not impact TNF-α/CXCL8 expression and TLR4-induced IκB-α degradation. These activities of THC and CBD were independent of the cannabinoid receptors or PPARγ (Figure 1; Fitzpatrick et al., 2020). Finally, THC, dose-dependently, protected against diclofenac-induced gastric inflammation, hemorrhagic streaks and gastric ulcers in male mice, and protected against tissue damage at doses insufficient to cause common cannabinoid side effects (Kinsey and Cole, 2013).
The synergy between phytocannabinoids (Mazuz et al., 2020; Anis et al., 2021) as well as between phytocannabinoids and terpenes (Namdar et al., 2019) has been demonstrated. Pre-clinical evidence suggests an ‘entourage effect’ might be inferred from the superior medical activities of full-spectrum cannabis extracts versus single molecules (Koltai et al., 2019). Furthermore, in some cases a “parasitage effect” might be detected, as there might also be negative molecular interactions in vitro (Namdar et al., 2020).
Indeed, as detailed above, phytocannabinoids are potent anti-inflammatory and immunomodulatory agents and in some cases they act via different signaling pathways. For example, although both THC and CBD decreased inflammation in LPS-activated microglial cells of a BV-2 mouse, they acted through different, although partially overlapping, mechanisms. CBD but not THC inhibited the NF-κB-dependent pathway, yet both CBD and THC regulated the IFNβ pathway activity (Kozela et al., 2010).
In order to take advantage of the synergy and to diminish negative interactions between phytomolecules, activity might be improved by selecting the most active phytomolecules while eliminating parts of the whole extract. This approach was demonstrated in the reduction of inflammation in colon cells and tissues. A THCA-rich fraction from the cannabis strain was shown to have superior activity against inflammation over the crude extract (Nallathambi et al., 2017) suggesting that the selection of active compounds may reduce the presence of inactive compounds or even those that have pro-inflammatory effects.
Moreover, in some cases, the activity of a combination of phytomolecules was found to be superior over that of a single molecule. This was demonstrated in an in vivo study on inflammation, where treatment with CBD combined with cannabis extract overcame the bell-shaped dose-response of purified CBD, suggesting that components found in the extract synergize with CBD to achieve the desired anti-inflammatory action (Gallily et al., 2015). In addition, a phytocannabinoid formulation showed superior activity reducing lung inflammation over the cannabis-derived fraction in vitro. Moreover, this particular phytocannabinoid and CBD formulation had superior activity over CBD alone (Anil et al., 2021).
Cannabis compounds, in some cases via the endocannabinoids system, were shown to affect some of the cornerstones of chronic inflammation. However, in light of the large number of active molecules produced by cannabis and their sometimes-synergistic interactions, there is a need to better specify cannabis-based treatments and the active compounds, while utilizing the synergy identified between cannabis phytomolecules. Thus, even if CBD or THC are considered potentially leading molecules, additional cannabis-derived compounds may be selected for improved activity.
Future approaches for improved usage of cannabis demand the development, transformation and formulation of full-spectrum cannabis extracts into active plant ingredients (APIs) to achieve higher effectivity. This might be done via careful selection of phytomolecules composition (Koltai et al., 2019; Koltai and Namdar, 2020). Notably, selecting only a few compounds for drug formulation may be compatible with modern medicine due to the potential for standardization, and careful dosing of API-based products. Importantly, once the mode of action of phytocannabinoids and that of their combination is known, APIs might be targeted towards specific mechanisms involved with inflammation.
Moreover, it might be that cannabis components can be combined with other pharmaceutical drugs to reduce inflammation. On the one hand, complementary effects might be identified due to different and perhaps complementary modes of action of cannabis compounds and pharmaceutical drugs. For example, THC was shown to reduce gastric inflammation caused by diclofenac, which may facilitate diclofenac’s effective usage against inflammation (Kinsey and Cole, 2013). On the other, CBD and THC were shown to have metabolism-dependent inhibition for Cytochrome P450 (CYP) enzymes. CYPs are responsible for drug metabolism, including detoxication and metabolic activation of xenobiotics (Yamaori et al., 2011). Hence, combined treatment with cannabis and anti-inflammatory drugs should be carefully considered.
SA composed the first draft of the manuscript, HP improved the draft and HK composed the final draft of the manuscript.
SA and HP are research scholars of ARO and HK is an employee of ARO. No external funding supported this manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We thank Zach Dunseth for English editing.
CBD, cannabidiol; CBG, cannabigerol; Δ9‐THC, Δ9‐tetrahydrocannabinol.
Methodology: To reflect on the effect of cannabis and its derived compounds on acute or chronic inflammation and the accumulating knowledge regarding cannabis active compounds and their mode of action, we have conducted a literature review using the following terms: “inflammation “, “acute inflammation”, “chronic inflammation”, “medical use of cannabis”, “therapy” “Cannabis sativa”, “C. sativa”, “cannabis”, “cannabinoids”, “terpenes”, “cannabis oil”, “adverse effects”, “endocannabinoid”, “phytocannabinoid” and “entourage effect”. The search was conducted on general and multidisciplinary research databases for peer-reviewed scientific manuscripts, including PubMed, Google Scholar, Scopus, and Web of Science.
Abdulkhaleq, L. A., Assi, M. A., Abdullah, R., Zamri-Saad, M., Taufiq-Yap, Y. H., and Hezmee, M. N. M. (2018). The Crucial Roles of Inflammatory Mediators in Inflammation: A Review. Vet. World 11 (5), 627–635. doi:10.14202/vetworld.2018.627-635
Actis, G. C., Pellicano, R., Fagoonee, S., and Ribaldone, D. G. (2019). History of Inflammatory Bowel Diseases. J. Clin. Med. 8 (11), 1970. doi:10.3390/jcm8111970
Ambrose, T., and Simmons, A. (2019). Cannabis, Cannabinoids, and the Endocannabinoid System-is There Therapeutic Potential for Inflammatory Bowel Disease? J. Crohns Colitis 13 (4), 525–535. doi:10.1093/ecco-jcc/jjy185
Anil, S. M., Shalev, N., Vinayaka, A. C., Nadarajan, S., Namdar, D., Belausov, E., et al. (2021). Cannabis Compounds Exhibit Anti-inflammatory Activity In Vitro in COVID-19-Related Inflammation in Lung Epithelial Cells and Pro-inflammatory Activity in Macrophages. Sci. Rep. 11 (1), 1462–1514. doi:10.1038/s41598-021-81049-2
Anis, O., Vinayaka, A. C., Shalev, N., Namdar, D., Nadarajan, S., Anil, S. M., et al. (2021). Cannabis-Derived Compounds Cannabichromene and Δ9-Tetrahydrocannabinol Interact and Exhibit Cytotoxic Activity against Urothelial Cell Carcinoma Correlated with Inhibition of Cell Migration and Cytoskeleton Organization. Molecules 26 (2), 465. doi:10.3390/molecules26020465
Bautista, J. L., Yu, S., and Tian, L. (2021). Flavonoids in Cannabis Sativa: Biosynthesis, Bioactivities, and Biotechnology. ACS omega 6 (8), 5119–5123. doi:10.1021/acsomega.1c00318
Biringer, R. G. (2021). Endocannabinoid Signaling Pathways: beyond CB1R and CB2R. J. Cel Commun. Signal. 15 (3), 335–360. doi:10.1007/s12079-021-00622-6
Borrelli, F., Fasolino, I., Romano, B., Capasso, R., Maiello, F., Coppola, D., et al. (2013). Beneficial Effect of the Non-psychotropic Plant Cannabinoid Cannabigerol on Experimental Inflammatory Bowel Disease. Biochem. Pharmacol. 85 (9), 1306–1316. doi:10.1016/j.bcp.2013.01.017
Capozzi, A., Caissutti, D., Mattei, V., Gado, F., Martellucci, S., Longo, A., et al. (2021). Anti-inflammatory Activity of a CB2 Selective Cannabinoid Receptor Agonist: Signaling and Cytokines Release in Blood Mononuclear Cells. Molecules 27 (1), 64. doi:10.3390/molecules27010064
Carrier, E. J., Auchampach, J. A., and Hillard, C. J. (2006). Inhibition of an Equilibrative Nucleoside Transporter by Cannabidiol: a Mechanism of Cannabinoid Immunosuppression. Proc. Natl. Acad. Sci. U S A. 103 (20), 7895–7900. doi:10.1073/pnas.0511232103
Castillo, A., Tolón, M. R., Fernández-Ruiz, J., Romero, J., and Martinez-Orgado, J. (2010). The Neuroprotective Effect of Cannabidiol in an In Vitro Model of Newborn Hypoxic-Ischemic Brain Damage in Mice Is Mediated by CB(2) and Adenosine Receptors. Neurobiol. Dis. 37 (2), 434–440. doi:10.1016/j.nbd.2009.10.023
Chiurchiù, V., Lanuti, M., De Bardi, M., Battistini, L., and Maccarrone, M. (2015). The Differential Characterization of GPR55 Receptor in Human Peripheral Blood Reveals a Distinctive Expression in Monocytes and NK Cells and a Proinflammatory Role in These Innate Cells. Int. Immunol. 27 (3), 153–160. doi:10.1093/intimm/dxu097
Cinelli, M. A., Do, H. T., Miley, G. P., and Silverman, R. B. (2020). Inducible Nitric Oxide Synthase: Regulation, Structure, and Inhibition. Med. Res. Rev. 40 (1), 158–189. doi:10.1002/med.21599
Correa, F., Docagne, F., Mestre, L., Clemente, D., Hernangómez, M., Loría, F., et al. (2009). A Role for CB2 Receptors in Anandamide Signalling Pathways Involved in the Regulation of IL-12 and IL-23 in Microglial Cells. Biochem. Pharmacol. 77 (1), 86–100. doi:10.1016/j.bcp.2008.09.014
Di Marzo, V., and Piscitelli, F. (2015). The Endocannabinoid System and its Modulation by Phytocannabinoids. Neurotherapeutics 12 (4), 692–698. doi:10.1007/s13311-015-0374-6
Di Marzo, V., Melck, D., Bisogno, T., and De Petrocellis, L. (1998). Endocannabinoids: Endogenous Cannabinoid Receptor Ligands with Neuromodulatory Action. Trends Neurosci. 21 (12), 521–528. doi:10.1016/s0166-2236(98)01283-1
Effenberger, M., Grander, C., Grabherr, F., Griesmacher, A., Ploner, T., Hartig, F., et al. (2021). Systemic Inflammation as Fuel for Acute Liver Injury in COVID-19. Dig. Liver Dis. 53 (2), 158–165. doi:10.1016/j.dld.2020.08.004
El Sohly, M. A., Radwan, M. M., Gul, W., Chandra, S., and Galal, A. (2017). “Phytochemistry of Cannabis Sativa L,” in Phytocannabinoids: Unraveling the Complex Chemistry and Pharmacology of Cannabis Sativa. Editors A. D. Kinghorn, H. Falk, S. Gibbons, and J. I. Kobayashi (Cham: Springer International Publishing), 1–36. doi:10.1007/978-3-319-45541-9_1
El-Remessy, A. B., Tang, Y., Zhu, G., Matragoon, S., Khalifa, Y., Liu, E. K., et al. (2008). Neuroprotective Effects of Cannabidiol in Endotoxin-Induced Uveitis: Critical Role of P38 MAPK Activation. Mol. Vis. 14, 2190–2203.
Feehan, K. T., and Gilroy, D. W. (2019). Is Resolution the End of Inflammation? Trends Mol. Med. 25 (3), 198–214. doi:10.1016/j.molmed.2019.01.006
Fitzgerald, K. A., and Kagan, J. C. (2020). Toll-like Receptors and the Control of Immunity. Cell 180 (6), 1044–1066. doi:10.1016/j.cell.2020.02.041
Fitzpatrick, J. M., Minogue, E., Curham, L., Tyrrell, H., Gavigan, P., Hind, W., et al. (2020). MyD88-dependent and -independent Signalling via TLR3 and TLR4 Are Differentially Modulated by Δ9-tetrahydrocannabinol and Cannabidiol in Human Macrophages. J. Neuroimmunol. 343, 577217. doi:10.1016/j.jneuroim.2020.577217
Fujiwara, N., and Kobayashi, K. (2005). Macrophages in Inflammation. Curr. Drug Targets Inflamm. Allergy 4 (3), 281–286. doi:10.2174/1568010054022024
Gaffal, E., Cron, M., Glodde, N., and Tüting, T. (2013). Anti-inflammatory Activity of Topical THC in DNFB-Mediated Mouse Allergic Contact Dermatitis Independent of CB1 and CB2 Receptors. Allergy 68 (8), 994–1000. doi:10.1111/all.12183
Galkina, E., and Ley, K. (2009). Immune and Inflammatory Mechanisms of Atherosclerosis. Annu. Rev. Immunol. 27, 165–197. doi:10.1146/annurev.immunol.021908.132620
Gallily, R., Yekhtin, Z., and Hanuš, L. O. (2015). Overcoming the Bell-Shaped Dose-Response of Cannabidiol by Using Cannabis Extract Enriched in Cannabidiol. Pharmacol. Pharm. 06 (02), 75–85. doi:10.4236/pp.2015.62010
Gentili, M., Ronchetti, S., Ricci, E., Di Paola, R., Gugliandolo, E., Cuzzocrea, S., et al. (2019). Selective CB2 Inverse Agonist JTE907 Drives T Cell Differentiation towards a Treg Cell Phenotype and Ameliorates Inflammation in a Mouse Model of Inflammatory Bowel Disease. Pharmacol. Res. 141, 21–31. doi:10.1016/j.phrs.2018.12.005
Giacobbe, J., Marrocu, A., Di Benedetto, M. G., Pariante, C. M., and Borsini, A. (2021). A Systematic, Integrative Review of the Effects of the Endocannabinoid System on Inflammation and Neurogenesis in Animal Models of Affective Disorders. Brain Behav. Immun. 93, 353–367. doi:10.1016/j.bbi.2020.12.024
Gülck, T., and Møller, B. L. (2020). Phytocannabinoids: Origins and Biosynthesis. Trends Plant Sci. 25 (10), 985–1004. doi:10.1016/j.tplants.2020.05.005
Gugliandolo, A., Pollastro, F., Grassi, G., Bramanti, P., and Mazzon, E. (2018). In Vitro model of Neuroinflammation: Efficacy of Cannabigerol, a Non-psychoactive Cannabinoid. Int. J. Mol. Sci. 19 (7), 1992. doi:10.3390/ijms19071992
Hackel, A., Aksamit, A., Bruderek, K., Lang, S., and Brandau, S. (2021). TNF-α and IL-1β Sensitize Human MSC for IFN-γ Signaling and Enhance Neutrophil Recruitment. Eur. J. Immunol. 51 (2), 319–330. doi:10.1002/eji.201948336
Haddad, J. J. (2002). Redox Regulation of Pro-inflammatory Cytokines and IκB-α/NF-κB Nuclear Translocation and Activation. Biochem. Biophys. Res. Commun. 296 (4), 847–856. doi:10.1016/s0006-291x(02)00947-6
Hanuš, L. O., Meyer, S. M., Munoz, E., Taglialatela-Scafiti, O., and Appendino, G. (2016). Phytocannabinoids: a Unified Critical Inventory. Nat. Prod. Rep. 33, 1357–1392. doi:10.1039/C6NP00074F
He, G., and Karin, M. (2011). NF-κB and STAT3 - Key Players in Liver Inflammation and Cancer. Cell Res. 21 (1), 159–168. doi:10.1038/cr.2010.183
Hegde, V. L., Nagarkatti, P. S., and Nagarkatti, M. (2011). Role of Myeloid-Derived Suppressor Cells in Amelioration of Experimental Autoimmune Hepatitis Following Activation of TRPV1 Receptors by Cannabidiol. PloS one 6 (4), e18281. doi:10.1371/journal.pone.0018281
Hillard, C. J. (2018). Circulating Endocannabinoids: from Whence Do They Come and where Are They Going? Neuropsychopharmacology 43 (1), 155–172. doi:10.1038/npp.2017.130
Jean-Gilles, L., Gran, B., and Constantinescu, C. S. (2010). Interaction between Cytokines, Cannabinoids and the Nervous System. Immunobiology 215, 606–610. doi:10.1016/j.imbio.2009.12.006
Kinsey, S. G., and Cole, E. C. (2013). Acute Δ(9)-tetrahydrocannabinol Blocks Gastric Hemorrhages Induced by the Nonsteroidal Anti-inflammatory Drug Diclofenac Sodium in Mice. Eur. J. Pharmacol. 715 (1-3), 111–116. doi:10.1016/j.ejphar.2013.06.001
Koltai, H., and Namdar, D. (2020). Cannabis Phytomolecule 'Entourage': From Domestication to Medical Use. Trends Plant Sci. 25 (10), 976–984. doi:10.1016/j.tplants.2020.04.007
Koltai, H., Poulin, P., and Namdar, D. (2019). Promoting Cannabis Products to Pharmaceutical Drugs. Eur. J. Pharm. Sci. 132, 118–120. doi:10.1016/j.ejps.2019.02.027
Kozela, E., Pietr, M., Juknat, A., Rimmerman, N., Levy, R., and Vogel, Z. (2010). Cannabinoids Δ9-Tetrahydrocannabinol and Cannabidiol Differentially Inhibit the Lipopolysaccharide-Activated NF-κB and Interferon-β/STAT Proinflammatory Pathways in BV-2 Microglial Cells. J. Biol. Chem. 285 (3), 1616–1626. doi:10.1074/jbc.m109.069294
Lanuti, M., Talamonti, E., Maccarrone, M., and Chiurchiù, V. (2015). Activation of GPR55 Receptors Exacerbates oxLDL-Induced Lipid Accumulation and Inflammatory Responses, while Reducing Cholesterol Efflux from Human Macrophages. PLoS One 10 (5), e0126839. doi:10.1371/journal.pone.0126839
Masoumi, M., Bashiri, H., Khorramdelazad, H., Barzaman, K., Hashemi, N., Sereshki, H. A., et al. (2021). Destructive Roles of Fibroblast-like Synoviocytes in Chronic Inflammation and Joint Damage in Rheumatoid Arthritis. Inflammation 44 (2), 466–479. doi:10.1007/s10753-020-01371-1
Mazuz, M., Tiroler, A., Moyal, L., Hodak, E., Nadarajan, S., Vinayaka, A. C., et al. (2020). Synergistic Cytotoxic Activity of Cannabinoids from Cannabis Sativa against Cutaneous T-Cell Lymphoma (CTCL) In-Vitro and Ex-Vivo. Oncotarget 11, 1141–1156. . doi:10.18632/oncotarget.27528
Naftali, T., Lev, L. B., Yablecovitch, D., Yablekovitz, D., Half, E., and Konikoff, F. M. (2011). Treatment of Crohn's Disease with Cannabis: an Observational Study. Isr. Med. Assoc. J. 13, 455–458.
Naftali, T., Bar-Lev Schleider, L., Dotan, I., Lansky, E. P., Sklerovsky Benjaminov, F., and Konikoff, F. M. (2013). Cannabis Induces a Clinical Response in Patients with Crohn's Disease: a Prospective Placebo-Controlled Study. Clin. Gastroenterol. Hepatol. 11 (10), 1276. doi:10.1016/j.cgh.2013.04.034
Nallathambi, R., Mazuz, M., Ion, A., Selvaraj, G., Weininger, S., Fridlender, M., et al. (2017). Anti-Inflammatory Activity in Colon Models Is Derived from Δ9-Tetrahydrocannabinolic Acid that Interacts with Additional Compounds in Cannabis Extracts. Cannabis Cannabinoid Res. 2 (1), 167–182. doi:10.1089/can.2017.0027
Namdar, D., Voet, H., Ajjampura, V., Nadarajan, S., Mayzlish-Gati, E., Mazuz, M., et al. (2019). Terpenoids and Phytocannabinoids Co-produced in Cannabis Sativa Strains Show Specific Interaction for Cell Cytotoxic Activity. Molecules 24, 3031. doi:10.3390/molecules24173031
Namdar, D., Anis, O., Poulin, P., and Koltai, H. (2020). Chronological Review and Rational and Future Prospects of Cannabis-Based Drug Development. Molecules 25 (20), 4821. doi:10.3390/molecules25204821
Netea, M. G., Schlitzer, A., Placek, K., Joosten, L. A. B., and Schultze, J. L. (2019). Innate and Adaptive Immune Memory: an Evolutionary Continuum in the Host's Response to Pathogens. Cell Host Microbe 25 (1), 13–26. doi:10.1016/j.chom.2018.12.006
Pagano, E., Capasso, R., Piscitelli, F., Romano, B., Parisi, O. A., Finizio, S., et al. (2016). An Orally Active Cannabis Extract with High Content in Cannabidiol Attenuates Chemically-Induced Intestinal Inflammation and Hypermotility in the Mouse. Front. Pharmacol. 7, 341. doi:10.3389/fphar.2016.00341
Panigrahy, D., Gilligan, M. M., Serhan, C. N., and Kashfi, K. (2021). Resolution of Inflammation: An Organizing Principle in Biology and Medicine. Pharmacol. Ther. 227, 107879. doi:10.1016/j.pharmthera.2021.107879
Pazos, M. R., Mohammed, N., Lafuente, H., Santos, M., Martínez-Pinilla, E., Moreno, E., et al. (2013). Mechanisms of Cannabidiol Neuroprotection in Hypoxic-Ischemic Newborn Pigs: Role of 5HT(1A) and CB2 Receptors. Neuropharmacology 71, 282–291. doi:10.1016/j.neuropharm.2013.03.027
Perez, E., Fernandez, J. R., Fitzgerald, C., Rouzard, K., Tamura, M., and Savile, C. (2022). In Vitro and Clinical Evaluation of Cannabigerol (CBG) Produced via Yeast Biosynthesis: a Cannabinoid with a Broad Range of Anti-inflammatory and Skin Health-Boosting Properties. Molecules 27 (2), 491. doi:10.3390/molecules27020491
Perry, V. H. (2004). The Influence of Systemic Inflammation on Inflammation in the Brain: Implications for Chronic Neurodegenerative Disease. Brain Behav. Immun. 18 (5), 407–413. doi:10.1016/j.bbi.2004.01.004
Pothen, J. J., Poynter, M. E., and Bates, J. H. (2015). A Computational Model of Unresolved Allergic Inflammation in Chronic Asthma. Am. J. Physiol. Lung Cel Mol. Physiol. 308 (4), L384–L390. doi:10.1152/ajplung.00268.2014
Rahaman, O., and Ganguly, D. (2021). Endocannabinoids in Immune Regulation and Immunopathologies. Immunology 164 (2), 242–252. doi:10.1111/imm.13378
Ribeiro, A., Ferraz-de-Paula, V., Pinheiro, M. L., Vitoretti, L. B., Mariano-Souza, D. P., Quinteiro-Filho, W. M., et al. (2012). Cannabidiol, a Non-psychotropic Plant-Derived Cannabinoid, Decreases Inflammation in a Murine Model of Acute Lung Injury: Role for the Adenosine A(2A) Receptor. Eur. J. Pharmacol. 678 (1-3), 78–85. doi:10.1016/j.ejphar.2011.12.043
Ryz, N. R., Remillard, D. J., and Russo, E. B. (2017). Cannabis Roots: A Traditional Therapy with Future Potential for Treating Inflammation and Pain. Cannabis Cannabinoid Res. 2 (1), 210–216. doi:10.1089/can.2017.0028
Sá, M. J. (2012). Physiopathology of Symptoms and Signs in Multiple Sclerosis. Arq. Neuropsiquiatr. 70, 733–740. doi:10.1590/s0004-282x2012000900016
Salvemini, D., Kim, S. F., and Mollace, V. (2013). Reciprocal Regulation of the Nitric Oxide and Cyclooxygenase Pathway in Pathophysiology: Relevance and Clinical Implications. Am. J. Physiol. Regul. Integr. Comp. Physiol. 304 (7), R473–R487. doi:10.1152/ajpregu.00355.2012
Sangiovanni, E., Fumagalli, M., Pacchetti, B., Piazza, S., Magnavacca, A., Khalilpour, S., et al. (2019). Cannabis Sativa L. Extract and Cannabidiol Inhibit In Vitro Mediators of Skin Inflammation and Wound Injury. Phytother. Res. 33 (8), 2083–2093. doi:10.1002/ptr.6400
Sevenoaks, M. J., and Stockley, R. A. (2006). Chronic Obstructive Pulmonary Disease, Inflammation and Co-morbidity--a Common Inflammatory Phenotype? Respir. Res. 7 (1), 70–79. doi:10.1186/1465-9921-7-70
Silva, L. B., dos Santos Neto, A. P., Maia, S. M. A. S., dos Santos Guimarães, C., Quidute, I. L., Carvalho, A. d. A. T., et al. (2019). The Role of TNF-α as a Proinflammatory Cytokine in Pathological Processes. Todent J 13 (1), 332–338. doi:10.2174/1874210601913010332
Tahir, M. N., Shahbazi, F., Rondeau-Gagné, S., and Trant, J. F. (2021). The Biosynthesis of the Cannabinoids. J. Cannabis Res. 3 (1), 7–12. doi:10.1186/s42238-021-00062-4
Taniguchi, K., and Karin, M. (2018). NF-κB, Inflammation, Immunity and Cancer: Coming of Age. Nat. Rev. Immunol. 18 (5), 309–324. doi:10.1038/nri.2017.142
Valdeolivas, S., Navarrete, C., Cantarero, I., Bellido, M. L., Muñoz, E., and Sagredo, O. (2015). Neuroprotective Properties of Cannabigerol in Huntington's Disease: Studies in R6/2 Mice and 3-Nitropropionate-Lesioned Mice. Neurotherapeutics 12 (1), 185–199. doi:10.1007/s13311-014-0304-z
Watson, C. W., Campbell, L. M., Sun-Suslow, N., Hong, S., Umlauf, A., Ellis, R. J., et al. (2021). Daily Cannabis Use Is Associated with Lower CNS Inflammation in People with HIV. J. Int. Neuropsychol. Soc. 27 (6), 661–672. doi:10.1017/S1355617720001447
Yamaori, S., Maeda, C., Yamamoto, I., and Watanabe, K. (2011). Differential Inhibition of Human Cytochrome P450 2A6 and 2B6 by Major Phytocannabinoids. Forensic Toxicol. 29 (2), 117–124. doi:10.1007/s11419-011-0112-7
Keywords: inflammation, medicinal cannabis, phytocannabinoids, Δ 9-tetrahydrocannabinol (THC), cannabidiol (CBD), cannabigerol (CBG)
Citation: Anil SM, Peeri H and Koltai H (2022) Medical Cannabis Activity Against Inflammation: Active Compounds and Modes of Action. Front. Pharmacol. 13:908198. doi: 10.3389/fphar.2022.908198
Received: 30 March 2022; Accepted: 08 April 2022;
Published: 09 May 2022.
Edited by:
Francesca Baratta, University of Turin, ItalyReviewed by:
Ethan Budd Russo, CReDO Sciecne, United StatesCopyright © 2022 Anil, Peeri and Koltai. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hinanit Koltai, aGtvbHRhaUBhZ3JpLmdvdi5pbA==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.