 Juliana Correa-Barbosa1
Juliana Correa-Barbosa1 Daniele Ferreira Sodré2
Daniele Ferreira Sodré2 Pedro Henrique Costa Nascimento2
Pedro Henrique Costa Nascimento2 Maria Fâni Dolabela1,2*
Maria Fâni Dolabela1,2*- 1 Pharmaceutical Science Post-graduation Programx, Federal University of Pará, Belém, Pará, Brazil
- 2Faculty of Pharmacy, Federal University of Pará, Belém, Brazil
Neglected diseases (NDs) are treated with a less varied range of drugs, with high cost and toxicity, which makes the search for therapeutic alternatives important. In this context, plants, such as those from the genus Zanthoxylum, can be promising due to active substances in their composition. This study evaluates the potential of species from this genus to treat NDs. Initially, a protocol was developed to carry out a systematic review approved by Prospero (CRD42020200438). The databases PubMed, BVS, Scopus, Science Direct, and Web of Science were used with the following keywords: “zanthoxylum,” “xanthoxylums,” “fagaras,” “leishmaniasis,” “chagas disease,” “malaria,” and “African trypanosomiasis.” Two independent evaluators analyzed the title and abstract of 166 articles, and 122 were excluded due to duplicity or for not meeting the inclusion criteria. From the 44 selected articles, results of in vitro/in vivo tests were extracted. In vitro studies showed that Z. rhoifolium, through the alkaloid nitidine, was active against Plasmodium (IC50 <1 μg/ml) and Leishmania (IC50 <8 μg/ml), and selective for both (>10 and >30, respectively). For Chagas disease, the promising species (IC50 <2 μg/ml) were Z. naranjillo and Z. minutiflorum, and for sleeping sickness, the species Z. zanthoxyloides (IC50 <4 μg/ml) stood out. In the in vivo analysis, the most promising species were Z. rhoifolium and Z. chiloperone. In summary, the species Z. rhoifolium, Z. naranjillo, Z. minutiflorum, Z. zanthoxyloides, and Z. chiloperone are promising sources of active molecules for the treatment of NDs.
1 Introduction
Neglected diseases (NDs), also called neglected tropical diseases (NTDs), are highly prevalent infectious conditions marked by a high degree of morbidity, mainly in the poorest and most vulnerable populations. These occur in developing countries, mostly in Africa, Asia, and the Americas (Souza, 2010; Sousadas et al., 2019).
Several NDs are caused by protozoa: malaria is caused by Plasmodium; leishmaniasis, by Leishmania; Chagas disease, by Trypanosoma cruzi, and sleeping sickness is caused by Trypanosoma brucei. Plasmodium falciparum, P. vivax, P. ovale, P. malariae, and P. knowlesi infect vertebrate erythrocytes and can cause severe and non-severe malaria (Bannister and Sherman, 2009). In the genus Leishmania, there are 30 species that infect mammals, 21 of them affect humans, and transmission occurs through the bite of female sandflies (Bates, 2007).
The species Trypanosoma cruzi and its different strains are responsible for causing Chagas disease, which is transmitted by vectors (80% of cases), blood transfusion (5–20% of cases; Fidalgo et al., 2018), and orally (Ferreira et al., 2014). Trypanosoma brucei is the causative agent of the African sleeping sickness, which occurs in 36 countries in sub-Saharan Africa, the poorest region in the world (Matthews, 2005).
For the treatment of Chagas disease, only two drugs are available, and they can cause severe adverse events (BRASIL, 2018). Also, in leishmaniasis, the number of drugs available for its treatment is limited (Comandolli-Wyrepkowski et al., 2020), while for malaria there are limitations of drugs to treat the hepatic form and resistant strains of P. falciparum (ANVISA, 2020).
Moreover, the treatment of diseases caused by protozoa has a high therapeutic cost, low adherence to treatment, and high inefficiency, since protozoa have developed resistance to the available drugs (Belloze, 2013), damaging to public health. In this sense, it is urgently necessary to search for new therapeutic alternatives, drugs that can cure such illnesses at a low cost with high levels of effectiveness. Thus, new drugs are needed for diseases caused by protozoa, and some studies have already highlighted the use of plants as a source of antiprotozoal agents (Ohashi et al., 2018).
Different studies have evaluated the biological activity of medicinal species or their toxicity, generating isolated information. The systematic review allows the integration of these results and demonstrates their therapeutic potential (Goodman et al., 2016; Melo-Neto et al., 2016). A genus that already has some chemical, pharmacological, and toxicity studies is Zanthoxylum (Dofuor et al., 2019b; Da Silva et al., 2019). From this genus (Zanthoxylum), there are reports of pharmacological evaluation of extracts (Adia et al., 2016), fractions (Alam and Najam us Saqib, 2017), and isolated substances (Bouquet et al., 2012) confirmed by in vitro/in vivo studies. The alkaloid class is the most commonly described, with some studies proving its antiprotozoal activity.
The hypothesized mechanism of action of the alkaloid class is as follows: cytoskeletal blockage or depolymerization (Fernandes, 2017), direct binding to the heme group of hemoglobin, inhibition of vacuolar phospholipase, protein synthesis inhibition, and interaction with DNA (França et al., 2008). Therefore, this study was carried out to compile and evaluate the in vitro/in vivo activities of the genus Zanthoxylum against diseases caused by protozoa.
2 Methods
2.1 Selection criteria and search strategies
For the development of this study, all original indexed articles, written in English, Portuguese, and Spanish, which reported data about the activities of extracts, fractions, and isolated compounds from the genus Zanthoxylum against Leishmania, Plasmodium, Trypanosoma cruzi, and Trypanosoma brucei on preclinical experiments (in vitro and/or in vivo) were included. Review articles, book chapters, case studies, and articles about genus activity against protozoan vectors and activities of synthesized compounds were excluded (Figure 1). Initially, a protocol was developed to carry out the systematic review, and this was approved by Prospero (CRD42020200438).
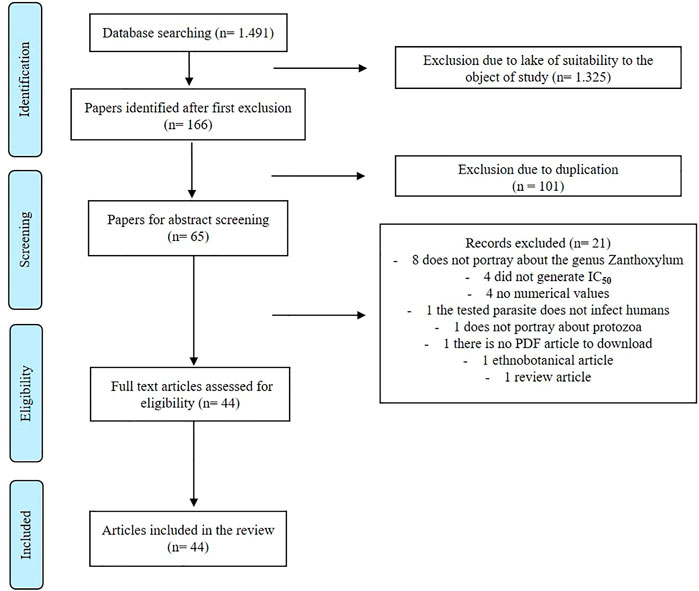
FIGURE 1. Article selection process.
The electronic databases PubMed, Virtual Health Library (VHL), Scopus, Science Direct, and Web of Science, all free access databases, were used to search for articles. The search descriptors were combinations of terms found in the titles of medical subjects (Mesh) and Descriptors in Health Sciences (DeCs). The date of the last search was May 2020.
In PubMed (pubmed.ncbi.nlm.nih.gov), there was mainly North American literature, and the descriptors used were as follows: (zanthoxylum) OR (zanthoxylums)) OR (xanthoxylum)) OR (xanthoxylums)) OR (fagaras, Zanthoxylum)) AND ((((Leishmaniasis) OR (Chagas disease)) OR (malaria)) OR (African trypanosomiasis)). In the BVS (bvsalud.org), there was a predominance of literature from Latin America when the following terms were used: (((leishmaniose) OR (doença de chagas) OR (malaria) OR (tripanossomíase Africana))) AND (tw: ((zanthoxylum)) OR (xanthoxylum).
In the Scopus database (scopus.com), which includes several knowledge areas, we used the following descriptors: (zanthoxylum OR xanthoxylum OR “fagaras, Zanthoxylum”) AND ALL (leishmaniasis OR “Leishmania Infection” OR “Chagas disease”" OR “Trypanosoma cruzi Infection” OR malaria OR “Plasmodium Infection” OR “African trypanosomiasis” OR “African Sleeping Sickness").
Operated by the Anglo-Dutch publisher Elsevier, Science Direct (sciencedirect.com) was also included in the research and the descriptors were as follows: (zanthoxylum OR xanthoxylum) AND (leishmaniasis OR Leishmania Infection OR Chagas disease OR Trypanosoma cruzi Infection OR malaria OR Plasmodium infection OR African trypanosomiasis). Finally, we included Web of Science (webofscience.com) as a multidisciplinary base and the research adopted the descriptors (zanthoxylum OR xanthoxylum OR fagaras, Zanthoxylum”) AND (leishmaniasis OR chagas disease” OR malaria OR african trypanosomiasis").
The articles were selected by two independent examiners, based on the reading of the title and abstract, with a third final examiner. Potentially eligible articles were read in full.
2.2 Bias risk
Because the selected studies may present risks of bias, two instruments were used: the guidelines for pre-clinical in vitro reports on dental materials, developed by Faggion et al. (2012) modified to meet the criteria of in vitro studies (Table 1), and SYRCLE, developed by Hooijmans et al. (2014) to assess the risk of bias of in vivo studies (Table 2). The studies were analyzed in pairs by two examiners, with a third evaluator when there was disagreement.
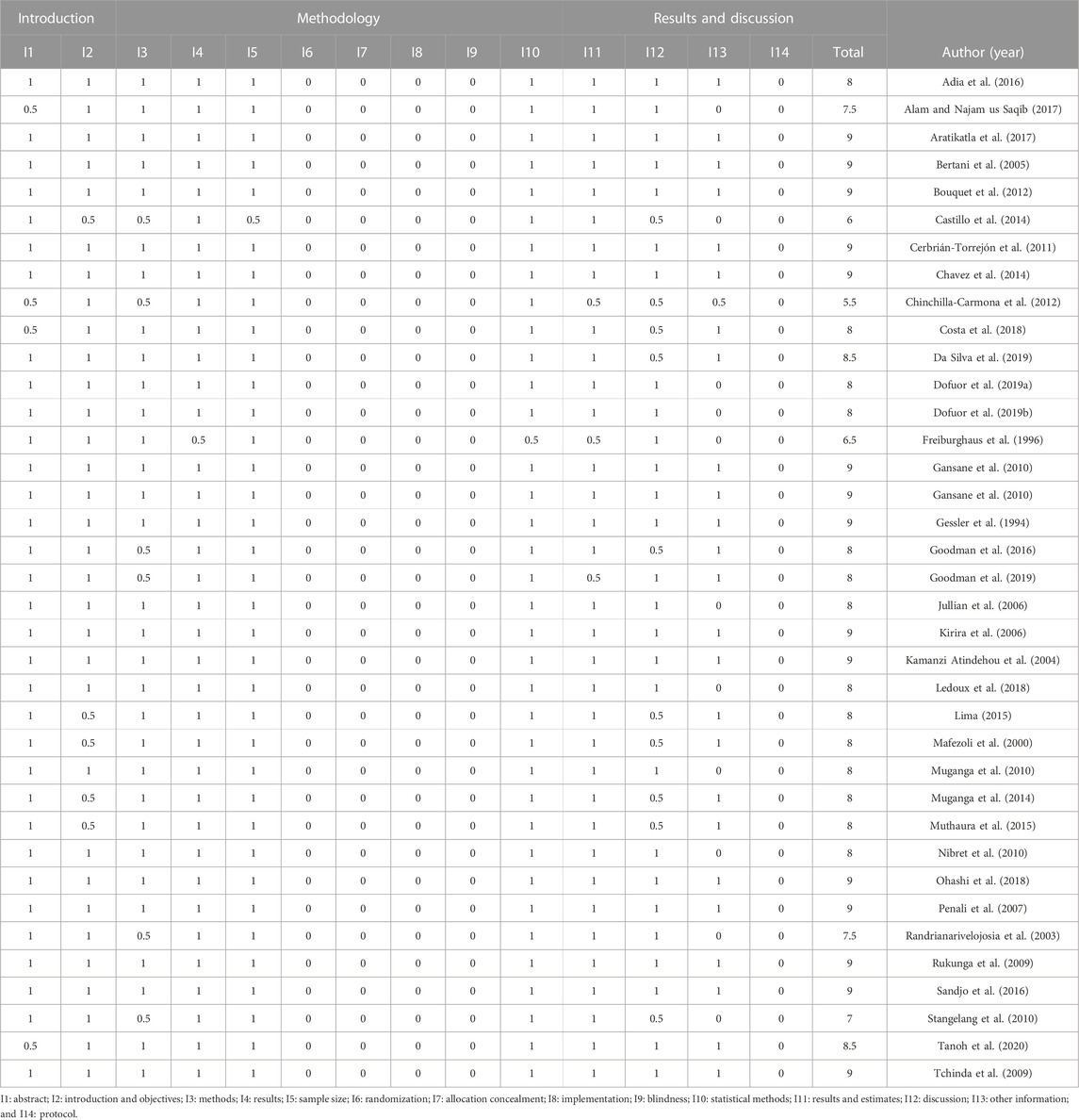
TABLE 1. Results of in vitro study evaluations.
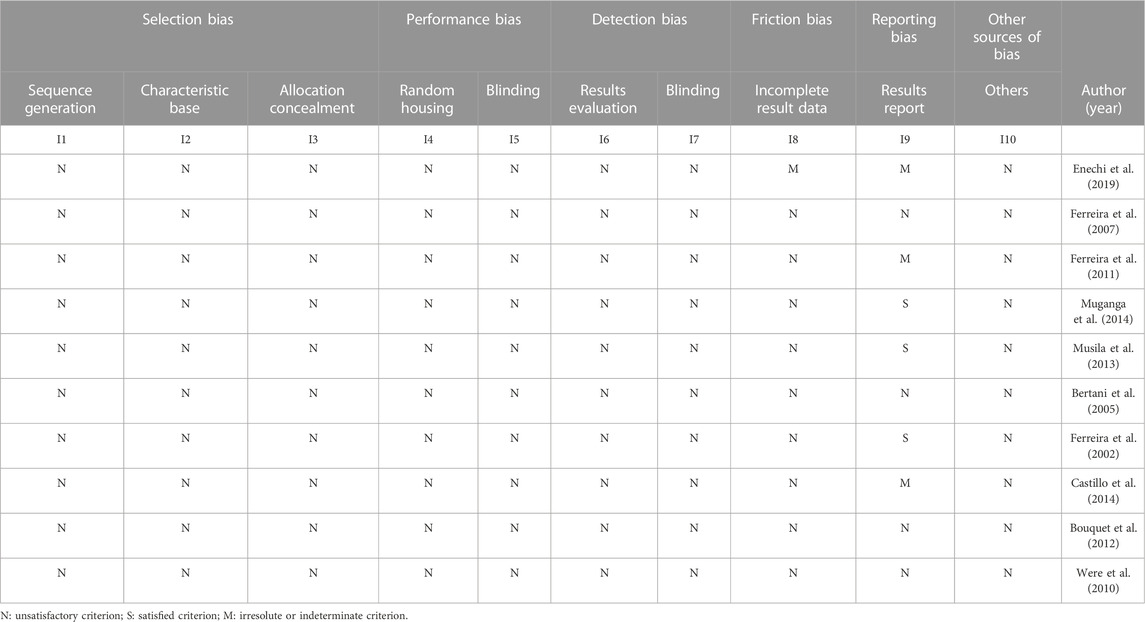
TABLE 2. Results of in vivo studies.
Based on the material developed by Faggion et al. (2012), the risk of bias was classified into numbers, with the value one representing a low risk of bias; 0.5, potential risk of bias; and 0, risk of bias, as shown in Table 1. This instrument was chosen because it is an easy-to-handle tool with a high intra-examiner agreement. The analysis was based on the following aspects: 1) abstract; 2) introduction, background, and objectives; 3) methods and intervention; 4) results; 5) sample size; 6) randomization: sequence generation; 7) allocation concealment; 8) implementation; 9) blinding (10) statistical methods; 11) results and estimates 12) discussion; 13) other information (financing); and 14) protocol.
The tool developed by Hooijmans et al. (2014) was created specifically for intervention studies in animals and is based on the Cochrane tool. The risk of bias was classified into specific characters, where N (not relevant to the item); S (showed relevance to the item); and M (presented medium relevance to the item). The analysis was based on the aspects (1; 2; 3) selection bias; (4; 5) performance bias (6; 7) detection bias; 8) friction bias; 9) random bias; and (10) other sources of bias (Table 2).
Regarding the activity classification, the parameters of Dolabela et al. (2015) for the genus Plasmodium and those of Mota et al. (2015) for the genus Leishmania were used, and the following parameters were adopted to evaluate the activity against Trypanosoma cruzi and Trypanosoma brucei: IC50 < 10 μg/ml, good activity; IC50 of 10–50 μg/ml, moderate activity; IC50 50–100 μg/ml, low activity; and IC50 > 100 μg/ml, inactive. For the selectivity index (IS), the one calculated from the ratio between cytotoxicity for macrophages (CC50) and activity against amastigotes (IC50) was used. IS values >20.0 indicate that the sample tested was more toxic to the parasite than to the host cell. IS values <20.0 demonstrate toxicity of the compound, adapted as described by Don and Ioset (2014).
2.3 Data extraction
Variables of interest for in vitro studies (first author, year of study, genus, species, part of the plant used, 50% inhibitory concentration—IC50, evolutionary form, protozoan species, 50% cytotoxic concentration—CC50, cell lineage, selectivity index, and activity classification) were transferred to tables in Word (Tables 3, 5, 7, 9).
Variables of interest for in vivo studies (first author, year of study, genus, species, part of the plant used, % parasitemia, % chemosuppression, % inhibition of the protozoan, protozoan species, animal species, 50% cytotoxic concentration (CC50), cell lineage, and selectivity index) were transferred to tables in Word (Tables 4, 6, 8).
3 Results
3.1 Selection of articles
In the search, 1,491 articles were identified, of which only 166 met the inclusion criteria. In the screening phase, the authors evaluated the titles of these 166 articles. Of these, 101 articles were excluded due to duplicity, and therefore, 65 articles were chosen for reading. After analyzing the abstracts, eight articles were excluded because they did not study the genus Zanthoxylum, four because they did not generate the IC50, four because they did not present numerical values, one because the tested parasite did not infect humans, one because it did not portray protozoa, one because it was not in a document available for reading without open access, one for being ethnobotanical article, and one review article, totaling 21 articles excluded. Thus, 44 original articles were included in this systematic review (Figure 1).
In the introduction and initial part of the methodology, all articles met the verification completely or partially, noting that the first items are in accordance with the methodology adapted from Faggion Jr et al. (2012). However, the verification in the final part of the methodology is totally different from the initial one, as none of the articles meet the guideline verification criteria, and this is a warning as to how the methodologies of these in vitro articles are being managed (Table 1).
However, based on the results and discussion, it is possible to notice again the improvement in the verification of these articles, and they fully or partially meet this verification. However, in the last item (protocol), none of the articles meet the verification criteria. This means that the articles did not expose their protocols, which hinders the reproducibility of this work (Table 1).
In the end, to have a quality overview of these articles, a score was generated according to the items the article presented during the methodology verification, and after that, we verified that the studies by Aratikatla et al. (2017), Bertani et al. (2005), Bouquet et al. (2012), Cebrián-Torrejón et al. (2011), Enciso et al. (2014), Gansane et al. (2010), Gessler et al. (1994), Kirira et al. (2006), Kamanzi Atindehou et al. (2004), Ohashi et al. (2018), Penali et al. (2007), Rukunga et al. (2009), Sandjo et al. (2016), and Tchinda et al. (2009) and obtained the highest score (score 9 out of 14) among the 37 articles evaluated by this methodology (Table 1).
None of the articles evaluated by Hooijmans et al. (2014) met the satisfaction criteria in selection, performance, and detection biases. Only the studies by Bertani et al. (2005), Musila et al. (2013), and Muganga et al. (2014) showed satisfaction in the assessment used in the reporting bias (Table 2).
3.3 Data collection
An extensive literature review was carried out to identify whether extracts, fractions, and pure substances from the genus Zanthoxylum presented in vitro and in vivo activity against protozoa of the genus Plasmodium, Leishmania, and Trypanosoma. The results can be seen in Tables 3, 4, 5, 6, 7, 8, 9.
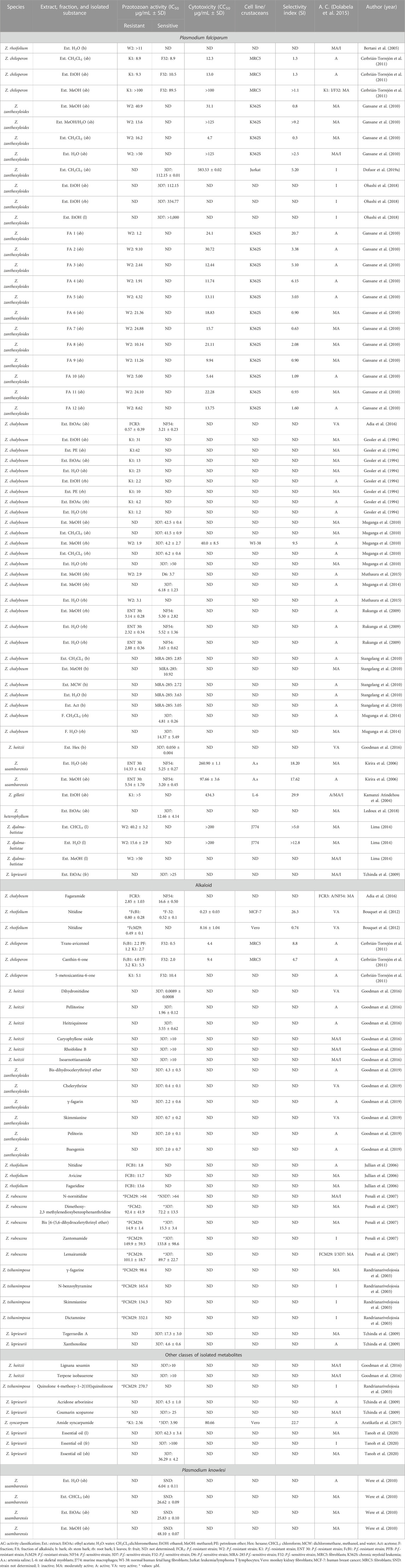
TABLE 3. In vitro antiparasitic activity against Plasmodium.
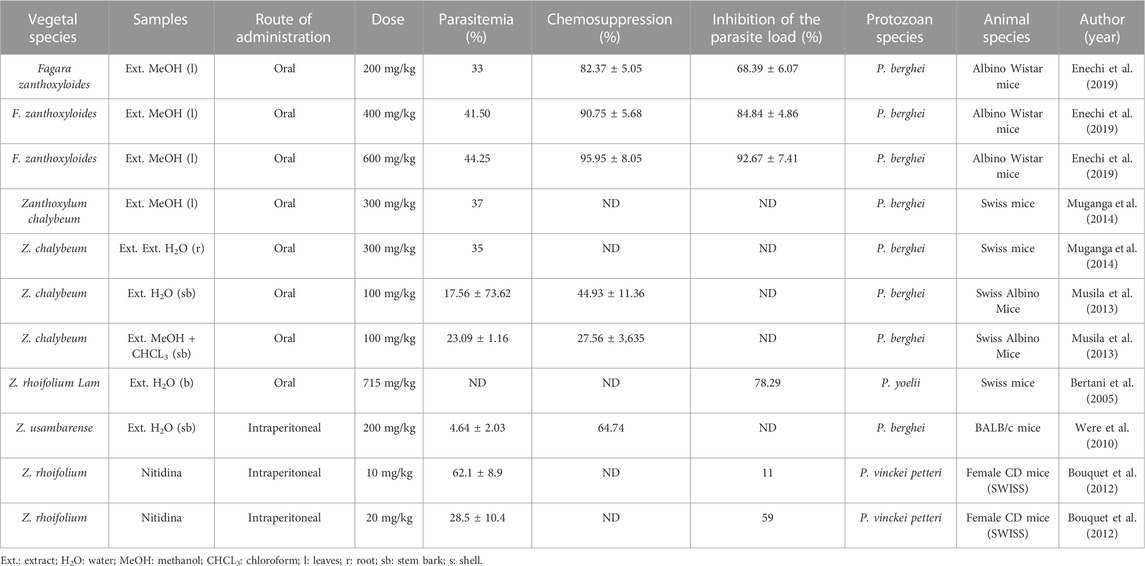
TABLE 4. In vivo antiparasitic activity against Plasmodium.
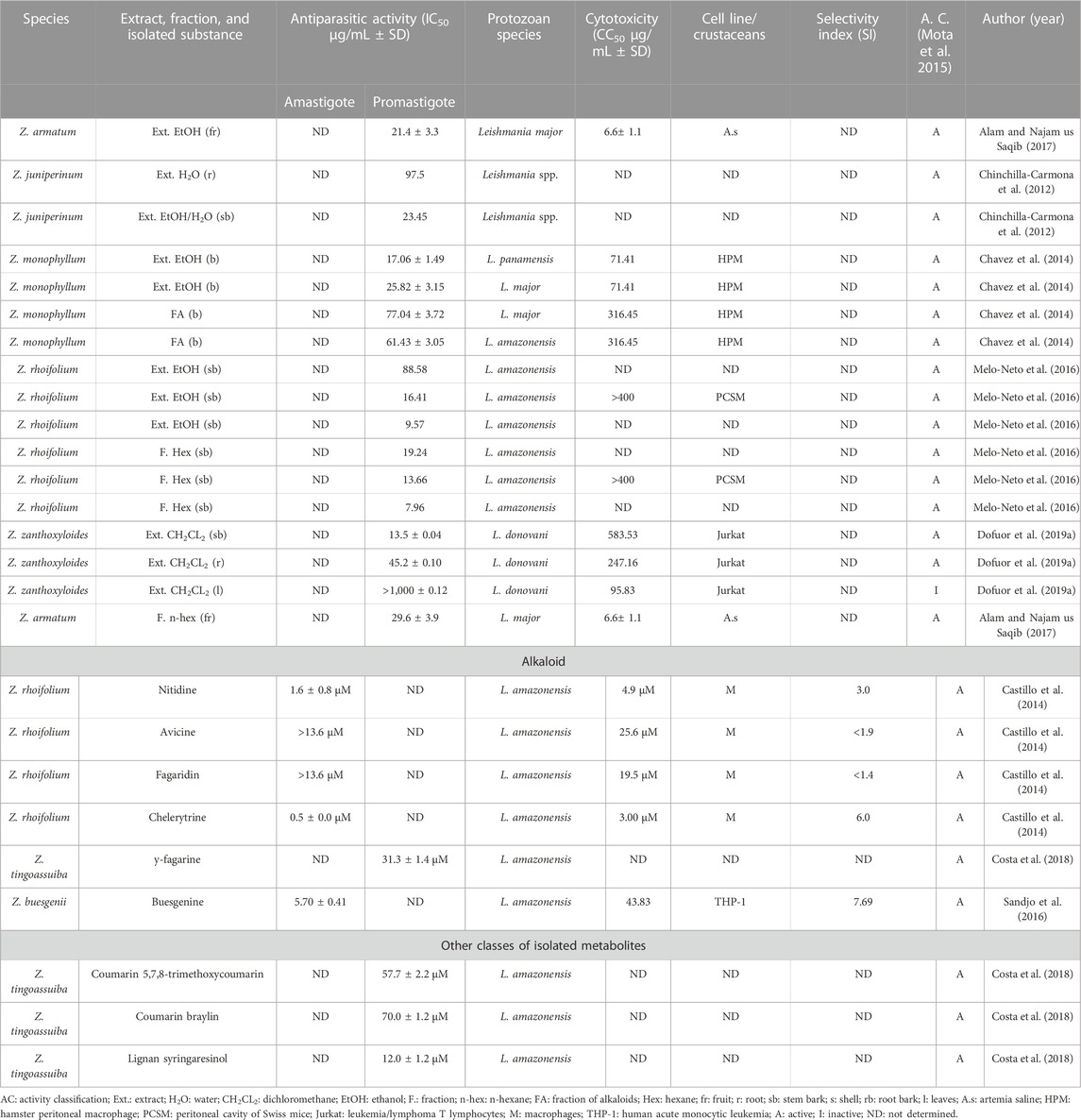
TABLE 5. In vitro antiparasitic activity against the genus Leishmania.
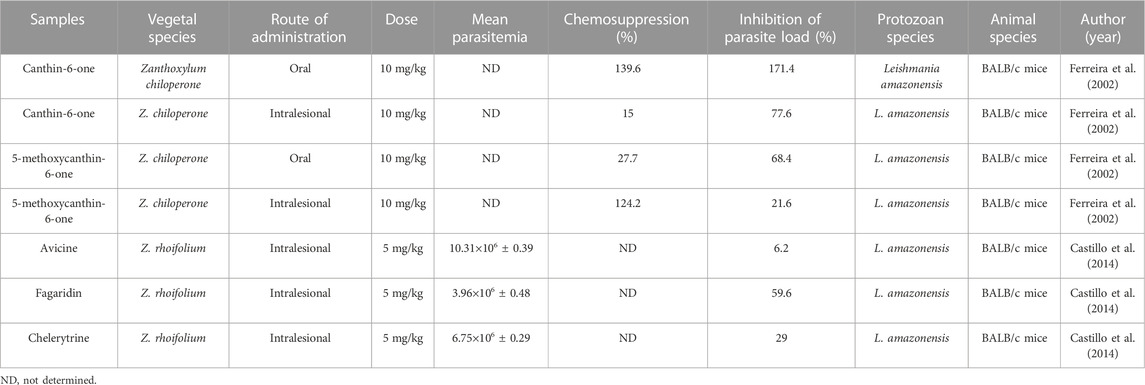
TABLE 6. In vivo antiparasitic activity against Leishmania.
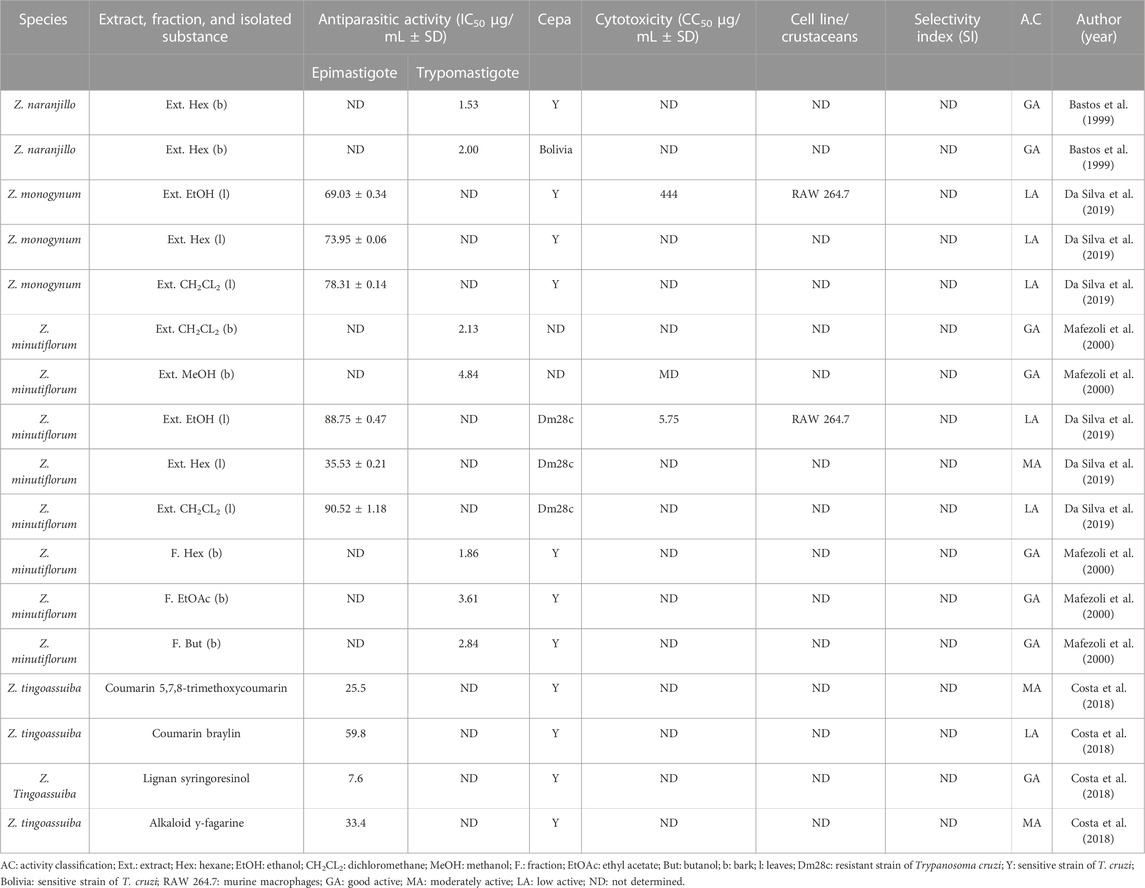
TABLE 7. In vitro antiparasitic activity against Trypanosoma cruzi.
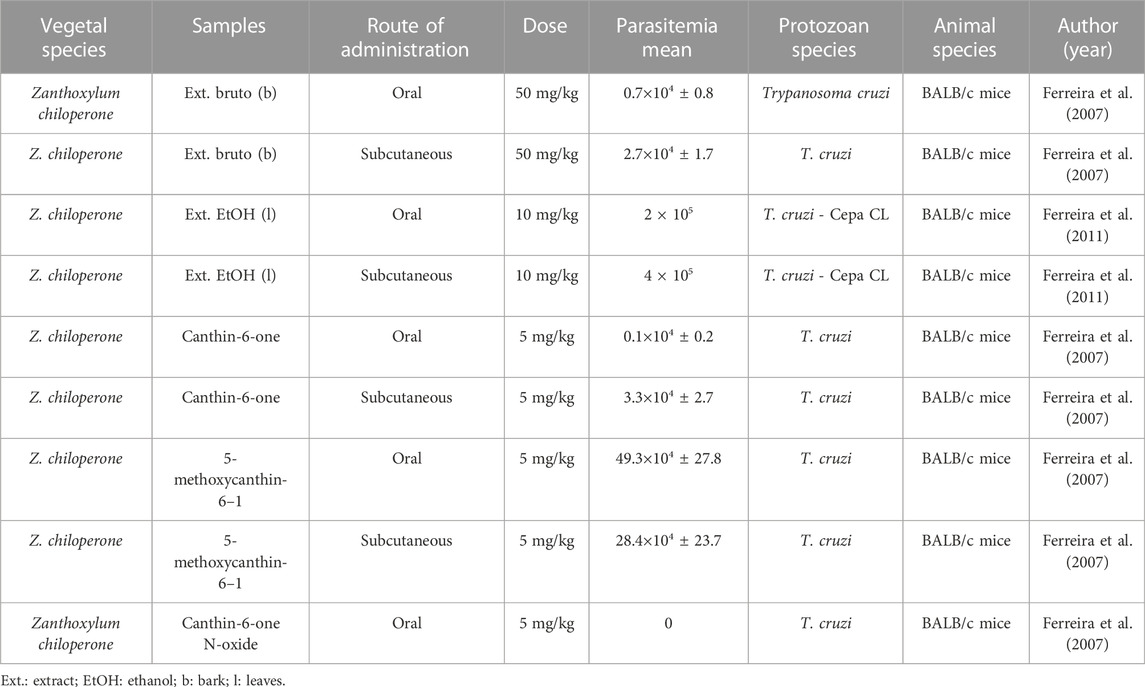
TABLE 8. In vivo antiparasitic activity against Trypanosoma cruzi.
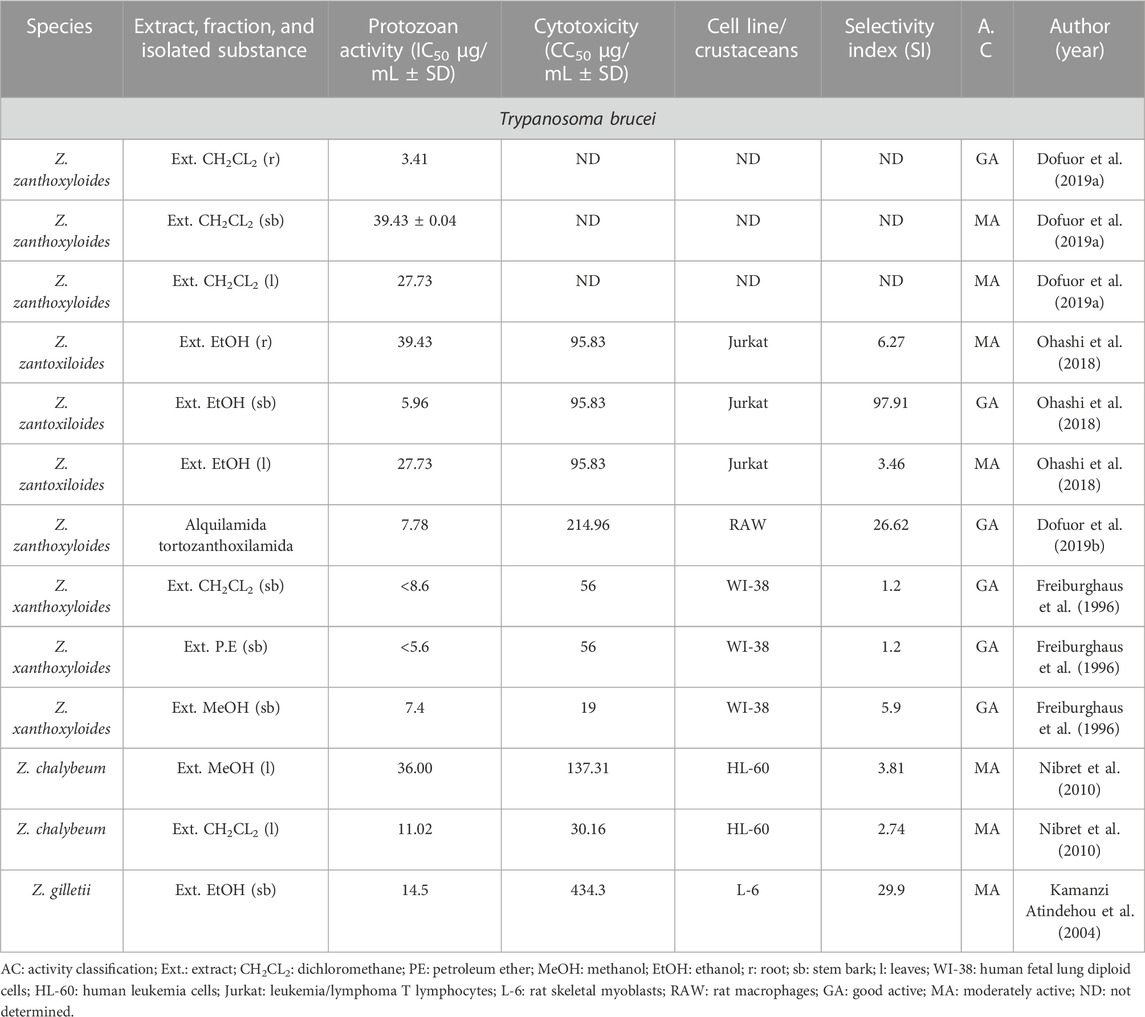
TABLE 9. In vitro antiparasitic activity against Trypanosoma brucei.
3.4 Plasmodium
One study evaluated the antiplasmodic activity against 12 alkaloid fractions from the bark of the Z. zanthoxyloides stem against resistant strains of P. falciparum. Fractions 1–5, 10, and 12 were active and non-cytotoxic to chronic myeloid leukemia strain (Table 3; Gansane et al., 2010). The ethyl acetate extract from the stem bark of Zanthoxylum chalybeum was active against sensitive and resistant strains of P. falciparum (Table 3; Adia et al., 2016).
Ethanolic ethyl acetate and aqueous extracts from the Z. chalybeum root bark showed activity against the same resistant strains (Table 3; Gessler et al., 1994). In addition, the aqueous extract from the Z. chalybeum root bark was active against resistant strains of the parasite, and the methanol extract was active against resistant and sensitive strains (Table 3; Muthaura et al., 2015).
From Z. rhoifolium, one alkaloid was isolated, and the alkaloid nitidine was considered active against the resistant strain of the P. falciparum (Table 3; Jullian et al., 2006). The alkaloid nitidine isolated from Z. rhoifolium was active against two resistant strains and a sensitive strain of P. falciparum, and it was not cytotoxic against monkey kidney fibroblast cells (Table 3; Bouquet et al., 2012).
The hexane extract from the bark of Z. heitzii was active on sensitive strains of P. falciparum. The alkaloids dihydronitidine, pellitorine, heitziquinone, caryophyllene oxide, rhoifoline B, and isoarnottianamide were isolated from the same species. From them, dihydronitidine and pellitorine showed activity against P. falciparum strains.
Finally, for in vitro studies, the syncarpamide, isolated from Z. syncarpum, was active in resistant and sensitive strains of P. falciparum, being non-cytotoxic against the normal monkey kidney fibroblast cells with high SI (Table 3; Aratikatla et al., 2017).
Regarding in vivo studies, percentage parasitemia and percentage chemosuppression were determined using the formula % parasitemia = (total number of parasite cells/total number of cells) x 100% and chemosuppression = [(negative control parasitemia) − (parasitemia with drug)]/negative control parasitemia (Hilou et al., 2006). After oral treatment with 200 mg/kg of methanol extract from Fagara zanthoxyloides in albino Wistar mice infected with Plasmodium berghei, the level of parasitemia was 33%, chemosuppression was 82.37%, and the inhibition rate of the parasite load was 68.39%. After treatment with a dose of 400 mg/kg of the same extract, the level of parasitemia was 41.50%, chemosuppression was 90.75%, and the inhibition rate of the parasite load was 84.84%. After treatment with the highest dose (600 mg/kg) of the extract, the parasitemia was 44.15%, with a chemosuppression of 95.95%, and inhibition of the parasite load of 92.67%. Furthermore, the non-lethality of the extract was confirmed, even at the highest dosage (5,000 mg/kg), having a CC50 of 28.21 ± 1.30 μg/ml (Table 4; Enechi et al., 2019).
An oral treatment with 100 mg/kg of aqueous and methanol + chloroform extract obtained from the bark of the Zanthoxylum chalybeum stem was administered to Swiss albino mice infected with Plasmodium berghei; parasitemia of 17.56% and chemosuppression of 44.93% were found after treatment, and parasitemia of 23.09% and chemosuppression of 27.56% after treatment with methanol + chloroform extract were also found. The two extracts were not cytotoxic (CC50: 268.28 and 25.78 μg/ml, respectively) at a dosage >1,000 μg/ml (Table 4), and for each specific blood smear for a given mouse, four magnification fields were observed, and the number of parasitized cells and the total number of cells in the magnification field were recorded. The data obtained were used to determine the percentage parasitemia and percentage chemosuppression in each mouse (Musila et al., 2013).
Finally, the inhibition of the parasite load in Swiss mice infected with P. yoelii after oral treatment with 715 mg/kg of the aqueous extract from Z. rhoifolium was 78.29% (Table 4; Bertani et al., 2005). In this study, there was no calculation of parasitemia.
3.5 Leishmania
In in vitro studies, the ethanolic extract and the hexane fraction from the bark of the Z. rhoifolium stem showed activity against Leishmania amazonensis promastigotes in 24, 48, and 72 h of treatment (Table 5; Melo-Neto et al., 2016). In the same study, the alkaloids chelerythrin and nitidine, isolated from Z. rhoifolium, showed activity against L. amazonensis amastigotes (Table 5; Castillo et al., 2014).
The dichloromethane extract from the bark of the Z. zanthoxyloides stem showed activity against L. donovani promastigotes; it was not cytotoxic in Jurkat cells (Table 5; Dofuor et al., 2019a).
The ethanolic extract and the alkaloid fraction from the bark of Z. monophyllum showed activity against L. major and L. panamensis promastigotes (Table 5; Enciso et al., 2014). Moreover, the ethanolic extract and the n-hexane fraction from Z. armatum fruits showed activity when tested against Leishmania major promastigotes (Table 5; Alam and Najam us Saqib, 2017).
The hydroethanolic and the aqueous extracts from the bark of Z. juniperinum showed activity against the promastigote form of Leishmania spp. (Table 5; CHINCHILLA-(Chinchilla-Carmona et al., 2012). In addition, the alkaloid buesgenine isolated from Z. buesgenii showed activity against amastigotes of L. amazonensis (Table 5; Sandjo et al., 2016). Also, the syringoresinol lignan, isolated from Z. tingoassuiba, was active against L. amazonensis promastigotes (Table 5; Costa et al., 2018).
Regarding in vivo studies, only isolated compounds of the genus were tested against Leishmania. The antileishmanial activity of canthin-6-one and 5-methoxycanthin-6-one isolated from Zanthoxylum chiloperone was evaluated in BALB/c mice infected with Leishmania amazonensis strains at a dose of 10 mg/kg by different routes (oral and intralesional). After oral treatment with canthin-6-one and 5-methoxycanthin-6-one, the parasite load inhibition rate was 171.4% and 68.4%, respectively, and chemosuppression of 139.6% and 27.7%, respectively. After treatment via the intralesional route, the parasitic inhibition rate was 77.6% and 21.6%, respectively, and the level of chemosuppression was 15% and 124.2%. It did not present toxicity, and its lethal dose was >400 mg/kg intraperitoneally (Table 6; Ferreira et al., 2002).
The alkaloids fagaridin and chelerythrine isolated from Zanthoxylum rhoifolium were also evaluated to assess their antileishmanial activity in BALB/c mice infected with L. amazonensis strains treated with a dose of 5 mg/kg by intralesional injection. After treatment, the parasitemia was 3.96×106 ± 0.48 for fagaridin and 6.75×106 ± 0.29 for chelerythrine; the inhibition rate of the parasite load was 59.6% and 29%, respectively (Table 6; Castillo et al., 2014). In none of the in vivo studies was there an analysis of parasitemia in percentage (%), only the mean was presented.
3.6 Trypanosoma cruzi
The hexane extract from the bark of Zanthoxylum naranjillo showed high activity against different strains of Trypanosoma cruzi trypomastigotes (Table 7; Bastos et al., 1999), followed by the hexane fraction of Z. minutiflorum with activity against T. cruzi trypomastigotes (Table 7; Mafezoli et al., 2000). The lignan compound syringoresinol isolated from the species Z. tingoassuiba was the isolated compound that showed the strongest activity against T. cruzi epimastigotes (Table 7; Costa et al., 2018). No studies examined compound activity against cells in the amastigote phase.
In in vivo studies, the crude extract of the stem of Zanthoxylum chiloperone showed the strongest antitrypanosome activity, being evaluated at a dose of 50 mg/kg orally and subcutaneously in BALB/c mice infected with Trypanosoma cruzi. After oral treatment, a parasitemia of 0.7×104 ± 0.8 was found, and a parasitemia of 2.7×104 ± 1.7 was found after subcutaneous administration (Table 8; Ferreira et al., 2007).
In addition, the alkaloid canthin-6-one, also isolated from the species Zanthoxylum chiloperone, was tested in BALB/c mice infected with T. cruzi strains at a dose of 5 mg/kg, orally and subcutaneously. After exposure to oral treatment, the parasitemia was 0.1×104 ± 0.2, and after subcutaneous treatment, the parasitemia was 3.3×104 ± 2.7, representing the compound with the strongest in vivo activity (Table 8; Ferreira et al., 2007).
3.7 Trypanosoma brucei
The dichloromethane extract obtained from the root of Z. zanthoxyloides showed the strongest activity against T. brucei (Table 9; Dofuor et al., 2019a). Moreover, the alkylamide tortozanthoxylamide, isolated from Z. zanthoxyloides, was the only isolated compound of the genus that was found in tests against T. brucei showing activity against the protozoan, moderate cytotoxicity in rat macrophages, and high SI (Table 9; Dofuor et al., 2019b).
4 Discussion
This systematic review provides information on four types of diseases caused by protozoa and the therapeutic potential of species from the genus Zanthoxylum. Original articles reporting data on preclinical experiments (in vitro and/or in vivo) with extracts, fractions, and isolated compounds of the genus Zanthoxylum against protozoa were included in the review. Two instruments were used to minimize the risk of bias: the guideline, developed by Faggion et al. (2012) for pre-clinical in vitro studies, and SYRCLE, developed by Hooijmans et al. (2014) to evaluate the in vivo studies. It is noteworthy that the analysis was performed by two independent evaluators, with the addition of a third when necessary, ensuring robustness and veracity of the analyzed data.
The treatment for diseases caused by protozoa, such as malaria, leishmaniasis, Chagas disease, and sleeping sickness, has several adverse effects, high toxicity, parasite resistance, and a reduced number of antiprotozoal drugs (Ohashi et al., 2018). Therefore, it is essential to search for new therapeutic alternatives from other sources, such as medicinal plants. Thus, we emphasize the importance of this review as a beginning in the analysis of possible promising species for the treatment of the diseases under study.
The lack of standard protocols for the evaluation of in vitro and in vivo results made it difficult to interpret the results; however, to minimize such factors, the activity classification the parameters of Dolabela et al. (2015) for the genus Plasmodium and those of Mota et al. (2015) for the genus Leishmania were used, and the following parameters were adopted to evaluate the activity against Trypanosoma cruzi and Trypanosoma brucei: IC50 < 10 μg/ml, good activity; IC50 of 10–50 μg/ml, moderate activity; IC50 50–100 μg/ml, low activity; and IC50 > 100 μg/ml, inactive.
Initially, the in vitro and in vivo antimalarial activity of Zanthoxylum was evaluated. Only 10 species were evaluated in vitro, using sensitive and resistant clones of Plasmodium falciparum. From the species, the Z. zanthoxyloides was widely studied in vitro, having the activity of its extracts, fractions, and isolated substances evaluated (Gansane et al., 2010; Goodman et al., 2019). Extracts obtained from this species were active in a chloroquine-resistant clone of P. falciparum, while the fractions were promising in a sensitive clone (Table 3). Also, extracts from the bark of this species were promising against L. donovani (Table 5).
The species Z. zanthoxyloides stood out due to its activity against African trypanosomiasis (sleeping sickness) caused by T. brucei. The dichloromethane extract from this species was the most promising (Dofuor et al., 2019a), followed by petroleum ether and methanol extracts (Dofuor et al., 2019a). From these, methanol extract showed less cytotoxic effect with a good selectivity index (Freiburghaus et al., 1996). In the study by Dofuor et al. (2019a), the dichloromethane extract presented aromatic hydrocarbons as its main constituents, indicative of metabolites such as terpenes. Thus, it suggested terpenes may be responsible for the antitrypanosomal activity.
All alkaloids isolated from the species were promising as antimalarials (Table 3). The likely mechanism of action of the alkaloids is explained by their ability to form complexes with the heme group and inhibit the formation of b-hematin (O'Neill et al., 2009; Kelly et al., 2009; Bouquet et al., 2012). The fagaronin inhibits topoisomerases I and II and acts as a DNA-intercalating agent (Larsen et al., 1993), and berberine inhibits the Plasmodium telomerase (Sriwilaijareon et al., 2002; Parida et al., 2014). The alkaloid flavopereirin also was active against L. amazonensis promastigotes, and this activity is attributable to oligopeptidase binding (Silva e Silva et al., 2020).
Other alkaloids have already been subjected to molecular docking studies to suggest possible therapeutic targets against parasites, such as cassin and (-)-3-O- acetylspectaline, which bind to the arginase in L. amazonensis, being embedded in the binding site of the enzyme, with hydrophobic bonds forming the ligand–arginase complex, which may explain the leishmanicidal profile of the compounds shown in the studies (Lacerda et al., 2018). This is one of the possible mechanisms of action of this class of substances.
From Z. chiloperone, the alkaloid canthin-6-one was isolated. This alkaloid showed antimicrobial (Thouvenel et al., 2003) and leishmanicidal (Ferreira et al., 2002) activities and was active against T. cruzi (Ferreira et al., 2007). However, the mechanism of action of canthin-6-one is entirely unknown, but its trypanocidal activity may suggest, as a first hypothesis, the inhibition of the sterol 14α-demethylase in T. cruzi intracellular amastigotes (Ferreira et al., 2007), a mechanism similar to that proposed for triazole antifungals (Molina et al., 2000; Lira et al., 2001; Urbina et al., 2003). This alkaloid also showed promising activity against L. amazonenzis (Ferreira et al., 2002), making it urgent to investigate its possible mechanism of action.
The alkaloids from the benzophenanthridine class (chelerythrine, skimmianine, and buesgenin) were also active against chloroquine-sensitive strains (Table 3; Goodman et al., 2019), with anti-inflammatory (Chaturvedi et al., 1997; Dvořák et al., 2006) and antimicrobial (Croaker et al., 2016) activities. An in vivo study using methanol extract of Z. zanthoxyloides administered orally in Wistar albino mice was found. This sample showed chemosuppression and inhibition of the parasite load proportional to dosage (Table 4; Enechi et al., 2019), the antimalarial mechanism of action being explained by the presence of fagaronine and berberine in its extract (Elujoba et al., 2005).
The alkylamide tortozanthoxylamide isolated from Z. zanthoxyloides showed promising results against T. brucei and a good selectivity index (Dofuor et al., 2019b). Some studies suggest the activity of this compound may occur by targeting the functioning of the parasite’s cell cycle through the inhibition of DNA synthesis, which is replicated in the S phase of parasite reproduction, and inhibiting the karyokinesis processes and parasite cytokinesis (Dofuor et al., 2019b). In summary, the antiparasitic activity of this genus is related to the alkaloids; however, preliminary results suggest that different signaling pathways may be involved in the activities (Dofuor et al., 2019b).
The extracts from Z. rhoifolium presented the best in vitro activity against strains resistant to CQ, and fractions obtained from these extracts were active in sensitive clones. From the isolated compounds of the genus, in in vitro studies, the alkaloid chelerythrine, isolated from Z. rhoifolium, showed the strongest activity. The compound showed good in vivo activity in parasitemia indices, and it was the least toxic compound (Castillo et al., 2014). The alkaloid nitidine isolated from this species obtained IC50 < 1 μg/ml in a resistant clone and a satisfactory selectivity index (>10) when compared to a non-tumor cell line (Table 3; Bouquet et al., 2012), being described as an active antimalarial principle (Bai et al., 2006; Nyangulu et al., 2010; BOUQUET et al., 2012).
Also, it showed anti-leishmania activity in vitro, with the hexane fraction (apolar), rich in terpenes, being the most promising (Melo-Neto et al., 2016), and the following compounds were isolated from this fraction: 7-O-2-quinolone ether; (13S)-labdane-8α, 15-diol; (13R)-labdane-8α; 15-diol; and 13-(S)-8α-13-epoxylabd-14-ene (Santiago-Brugnoli et al., 2013). The leishmanicidal activity of terpenes has been described in different studies (Cechinel-Filho and Yunes, 1998; Arruda et al., 2005), as having possible direct mechanisms of action in the inhibition of protease activity, lipid synthesis, cell cycle, or indirectly through modulation of macrophage activation (Soares et al., 2012). Metabolites as the nerolidol, an oxygenated sesquiterpene, effectively inhibit the biosynthesis of isoprenoids such as dolicho, ergosterol, and ubiquinone in promastigotes (Arruda et al., 2005; Rodrigues et al., 2013). Two terpenes isolated from C. cajucara, namely, t-dehydrocrotonin and t-crotonin, inhibited trypanothione reductase in L. amazonensis promastigotes (Lima, 2014).
Among the analyzed species, the Z. chalybeum stood out due to the activity of its ethyl acetate extract and the alkaloid fagaramide against sensitive and resistant strains of Plasmodium (Adia et al., 2016). Also, from this species, two extracts (methanol + aqueous extract and methanol + chloroform + aqueous extract) decreased parasitemia and promoted high chemosuppression in Swiss mice infected with Plasmodium berghei (Musila et al., 2013; Mugunga et al., 2014). Considering that phytochemical analyses of this species revealed several alkaloid substances, such as benzophenanthridine, chelerythrine, nitidine, and 8-O-demethylchelerythrine (Kato et al., 1996; Tian et al., 2017), it is suggested that its activity against Plasmodium results from the blocking or depolymerization of the cytoskeleton in red blood cells to prevent the entry of the protozoan (Fernandes, 2017).
The species Z. heitzii also had good activity against sensitive Plasmodium strains. Their study reports the isolation of some classes of substances; however, only the alkaloids didydronitidine and heitziquinone showed activity against the tested strains. The study suggested that the activity of the alkaloid dihydronitidine is equal to that of “delayed death” drugs (Goodman et al., 2016), common to compounds that target the parasite’s epicoplast (Goodman et al., 2007). However, the alkaloid heitziquinone comes from benzo(c)phenanthridine, and previous studies found that this class is very sensitive to small changes in chemical structure (Nyangulu et al., 2010), and its structure–activity mechanism comes from an open C-ring (Goodman et al., 2016); this can cause difficulty to fully understand its antiplasmodial mechanism.
In the compilation of studies of the genus Zanthoxylum against T. cruzi, some in vitro analyses were highlighted. The hexane extract from Z. naranjillo against the T. cruzi trypomastigote form presented good activity in both tested strains (sensitive and resistant; Bastos et al., 1999); however, this is the only study that affirms the trypanocidal activity of the species. Complementary studies to identify the metabolite responsible for the activity and the mechanism are important.
The dichloromethane and methanol extracts and ethyl acetate, hexane, and butanol fractions from Z. minutiflorum showed strong activity against the trypomastigote form (Mafezoli et al., 2000). The in vivo trypanocidal action has also been reported in the literature through an index of published abstracts (Ferreira et al., 2003); however, this was the only in vivo study for this species reported in the literature.
The trypanocidal activity may be related to different chemical constituents of plants, such as isoquinoline alkaloids, lignans, coumarins, flavonoids, and terpenes (Prieto et al., 2011). In the in vitro study of the Z. tingoassuiba, the isolated compound syringoresinol, a furofuran lignan, showed activity against the epimastigote form of the Y strain, and its isolation from this plant species was reported for the first time in the literature (Costa et al., 2018). Through molecular docking studies, it is suggested that the likely mechanism of action of lignans occurs by interrupting the divisions and other cellular functions of the parasite, with tubulin being a possible biological target (Corrêa, 2015).
5 Conclusion
Isolated fractions and substances from different Zanthoxylum species seem to be promising as sources of active molecules in disease-causing parasites. When considering the cytotoxicity and antimalarial activity in vitro and in vivo, Z. rhoifolium is the most promising species due to metabolites such as alkaloids in its composition. When considering the antileishmanial activity, it is suggested that alkaloids and terpenes are responsible for the activity, mainly on the Z. rhoifolium. For Chagas disease, in the in vitro analysis, the most promising species was Z. minutiflorum due to its activity against trypomastigotes. As for in vivo analysis, the most promising species was Z. chiloperone, based on alkaloids in its composition. Finally, for sleeping sickness, the most promising species in vitro studies was Z. zanthoxyloides, and its activity may be related to metabolites such as terpenes and tortozanthoxylamide. In general, the most promising metabolites for the studied diseases are described in Figure 2. So far, there are no studies on the in vivo infection of the disease. The importance of toxicity studies in animals and the evaluation of the genotoxic, mutagenic, and carcinogenic potential of the species are highlighted.
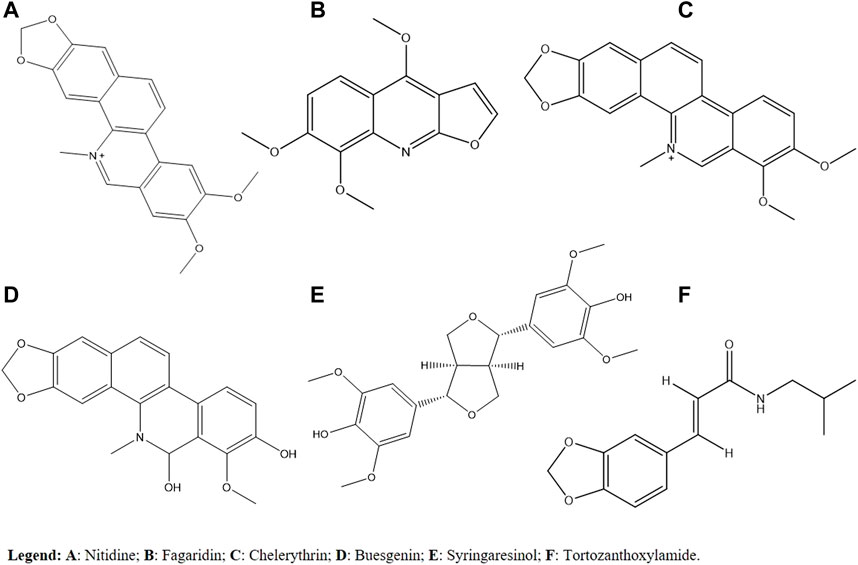
FIGURE 2. Most promising secondary metabolites for the diseases studied.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material; further inquiries can be directed to the corresponding author.
Author contributions
JC-B, DS, and PN searched the articles, created the outline, and drafted the manuscript. DS and PN assisted in searching relative articles and drafting the manuscript. JC-B and MD rechecked the articles and revised the manuscript. JC-B and MD supervised the manuscript writing and revised the manuscript.
Funding
This work was supported by the Dean of research and post-graduate studies—UFPA (Propesp–UFPA).
Acknowledgments
The authors also acknowledge the funding support from the National Council for Scientific and Technological Development.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphar.2022.873208/full#supplementary-material
References
Adia, M. M., Emami, S. N., Byamukama, R., Faye, I., and Borg-Karlson, A. K. (2016). Antiplasmodial activity and phytochemical analysis of extracts from selected Ugandan medicinal plants. J. Ethnopharmacol. 186, 14–19. doi:10.1016/J.JEP.2016.03.047
Alam, F., and Najam us Saqib, Q. N. (2017). Evaluation of Zanthoxylum armatum Roxb for in vitro biological activities. J. Tradit. Complement. Med. 7, 515–518. doi:10.1016/J.JTCME.2017.01.006
Aratikatla, E. K., Valkute, T. R., Puri, S. K., Srivastava, K., and Bhattacharya, A. K. (2017). Norepinephrine alkaloids as antiplasmodial agents: Synthesis of syncarpamide and insight into the structure-activity relationships of its analogues as antiplasmodial agents. Eur. J. Med. Chem. 138, 1089–1105. doi:10.1016/J.EJMECH.2017.07.052
Arruda, D. C., D’Alexandri, F. L., Katzin, A. M., and Uliana, S. R. B. (2005). Antileishmanial activity of the terpene nerolidol. Antimicrob. Agents Chemother. 49, 1679–1687. doi:10.1128/AAC.49.5.1679-1687.2005
Bai, L. P., Zhao, Z. Z., Cai, Z., and Jiang, Z. H. (2006). DNA-binding affinities and sequence selectivity of quaternary benzophenanthridine alkaloids sanguinarine, chelerythrine, and nitidine. Bioorg. Med. Chem. 14, 5439–5445. doi:10.1016/J.BMC.2006.05.012
Bannister, L. H., and Sherman, I. W. (2009). Plasmodium. California, United States: Wiley. doi:10.1002/9780470015902.a0001970.pub2
Bastos, J. K., Albuquerque, S., and Silva, M. L. A. (1999). Evaluation of the trypanocidal activity of lignans isolated from the leaves of Zanthoxylum naranjillo. Planta Med. 65, 541–544. doi:10.1055/s-1999-14012
Bates, P. A. (2007). Transmission of Leishmania metacyclic promastigotes by phlebotomine sand flies. Int. J. Parasitol. 37, 1097–1106. doi:10.1016/J.IJPARA.2007.04.003
Belloze, K. T. (2013). Priorização de alvos para fármacos no combate a doenças tropicais negligenciadas causadas por protozoários. xiv,272-xiv. Available at: http://157.86.8.8/reports/doutorado_bibcb/kele_belloze_ioc_dout_2013.pdf (Accessed February 8, 2022).
Bertani, S., Bourdy, G., Landau, I., Robinson, J. C., Esterre, P., and Deharo, E. (2005). Evaluation of French Guiana traditional antimalarial remedies. J. Ethnopharmacol. 98, 45–54. doi:10.1016/J.JEP.2004.12.020
Bouquet, J., Rivaud, M., Chevalley, S., Deharo, E., Julian, V., and Velentin, A. (2012). Biological activities of nitidine, a potential antimalarial lead compound. Malar. J. 167, 67. doi:10.1186/1475-2875-11-67
BRASIL (2018). Ministério da saúde. Protocolo Clínico e Diretrizes Terapêuticas Doença de Chagas. Brasília. Available at: http://conitec.gov.br/images/Protocolos/Relatorio_PCDT_Doenca_de_Chagas.pdf (Accessed February 3, 2021).
BRASIL (2020). Ministry of Health. Malaria treatment guide in Brazil. Available at: https://bvsms.saude.gov.br/bvs/publicacoes/guia_tratamento_malaria_brasil.pdf (Accessed February 5, 2021).
Castillo, D., Sauvain, M., Rivaud, M., and Jullian, V. (2014). In vitro and in vivo activity of benzo[c]phenanthridines against leishmania amazonensis. Planta Med. 80, 902–906. doi:10.1055/S-0034-1382826
Cebrián-Torrejón, G., Spelman, K., Leblanc, K., Muñoz-Durango, K., Gutiérrez, S. T., Ferreira, M. E., et al. (2011). The antiplasmodium effects of a traditional South American remedy: Zanthoxylum chiloperone var. angustifolium against chloroquine resistant and chloroquine sensitive strains of Plasmodium falciparum. Rev. Bras. Farmacogn. 21, 652–661. doi:10.1590/S0102-695X2011005000104
Cechinel-Filho, V., and Yunes, R. A. (1998). Estratégias para a obtenção de compostos farmacologicamente ativos a partir de plantas medicinais: Conceitos sobre modificação estrutural para otimização da atividade. Quim. Nova 21, 99–105. doi:10.1590/S0100-40421998000100015
Chaturvedi, M. M., Kumar, A., Darnay, B. G., Chainy, G. B. N., Agarwal, S., and Aggarwal, B. B. (1997). Sanguinarine (pseudochelerythrine) is a potent inhibitor of NF-kappaB activation, IkappaBalpha phosphorylation, and degradation. J. Biol. Chem. 272, 30129–30134. doi:10.1074/JBC.272.48.30129
Chavez, E. E., Coy-Barrera, O., Patino, L., and Delgado, G. (2014). Evaluation of the leishmanicidal activity of rutaceae and lauraceae ethanol extracts on golden Syrian hamster (Mesocricetus auratus) peritoneal macrophages. Indian J. Pharm. Sci. 76, 178–197.
Chinchilla-Carmona, M., Valerio, I., Sánchez, R., Mora, V., Bagnarello, V., Martínez, L., et al. (2012). In vitro antimalarial activity of extracts of some plants from a biological reserve in Costa Rica. Rev. Biol. Trop. 60 (2), 881–891. doi:10.15517/rbt.v60i2.4024
Comandolli-Wyrepkowski, C. D., Wyrepkowski, C. D. C., Paz, A. D. C., Jensen, B. B., and Franco, A. M. R. (2020). Aspectos farmacológicos da terapia medicamentosa utilizada para a leishmaniose cutânea: uma revisão de literatura. Acervo Saude 12, e3352. doi:10.25248/REAS.E3352.2020
Corrêa, D. S. (2015). Modelagem por homologia da tubulina do Plasmodium falciparum e o estudo de lignanas ariltetralônicas antimaláricas por docking molecular. Available at: https://repositorio.ufscar.br/handle/ufscar/7309 (Accessed February 8, 2021).
Costa, R. S., Souza Filho, O. P., Dias Júnior, O. C. S., Silva, J. J., Hyaric, L. S., Santos, M. A. V., et al. (2018). In vitro antileishmanial and antitrypanosomal activity of compounds isolated from the roots of Zanthoxylum tingoassuiba. Rev. Bras. Farmacogn. 28, 551–558. doi:10.1016/J.BJP.2018.04.013
Croaker, A., King, G. J., Pyne, J. H., Anoopkumar-Dukie, S., and Liu, L. (2016). Sanguinaria canadensis: Traditional medicine, phytochemical composition, biological activities and current uses. Int. J. Mol. Sci. 17, 1414. doi:10.3390/IJMS17091414
Da Silva, F. B., Trovó, M., Gomes, A. L., Mazotto, A. M., Vermelho, A. B., Martins, R. C. C., et al. (2019). Antiparasitic activity and characterization of lignans from the ethanolic extract of zanthoxylum monogynum A. St-hil. Leaves. Rev. Virtual Quim. 11, 1498–1512. doi:10.21577/1984-6835.20190104
Dofuor, A. K., Djameh, G. I., Ayertey, F., Bolah, P., Amoa-Bosompem, M., Kyeremeh, K., et al. (2019a). Antitrypanosomal effects of zanthoxylum zanthoxyloides (lam.) zepern. & timler extracts on african trypanosomes. Evid. Based. Complement. Altern. Med. 2019, 1730452. doi:10.1155/2019/1730452
Dofuor, A. K., Kwain, S., Osei, E., Tetevi, G. M., Okine, L. K., Ohashi, M., et al. (2019b). N-(Isobutyl)-3, 4-methylenedioxy cinnamoyl amide. Molbank 2019, M1070. doi:10.3390/M1070
Dolabela, M. F., Povoa, M. M., Brandão, G. C., Rocha, F. D., Soares, L. F., Paula, R. C., et al. (2015). Aspidosperma species as sources of anti-malarials: Uleine is the major anti-malarial indole alkaloid from aspidosperma parvifolium (apocynaceae). Malar. J. 266, 498–522. doi:10.1186/s12936-015-0997-4
Don, R., and Ioset, J. R. (2014). Screening strategies to identify new chemical diversity for drug development to treat kinetoplastid infections. Parasitology 141 (01), 140–146. doi:10.1017/S003118201300142X
Dvořák, Z., Vrzal, R., Maurel, P., and Ulrichová, J. (2006). Differential effects of selected natural compounds with anti-inflammatory activity on the glucocorticoid receptor and NF-kappaB in HeLa cells. Chem. Biol. Interact. 159, 117–128. doi:10.1016/J.CBI.2005.10.105
Elujoba, A., Odeleye, O., and Ogunyemi, C. (2005). Traditional medicine development for medical and dental primary health care delivery system in Africa. Afr. J. Tradit. Complement. Altern. Med. 2. doi:10.4314/AJTCAM.V2I1.31103
Enciso, N. C., Coy-Barrera, E. D., Patiño, O. J., Cuca, L. E., and Delgado, G. (2014). Evaluation of the leishmanicidal activity of rutaceae and lauraceae ethanol extracts on golden Syrian hamster (Mesocricetus auratus) peritoneal macrophages. Indian J. Pharm. Sci. 76 (3), 188–197.
Enechi, O. C., Amah, C. C., Okagu, I. U., Ononiwu, C. P., Azidiegwu, V. C., Ugwuoke, E. O., et al. (2019). Methanol extracts of Fagara zanthoxyloides leaves possess antimalarial effects and normalizes haematological and biochemical status of Plasmodium berghei-passaged mice. Pharm. Biol. 57, 577–585. doi:10.1080/13880209.2019.1656753
Faggion, C. M., Listl, S., and Giannakopoulos, N. N. (2012). The methodological quality of systematic reviews of animal studies in dentistry. Vet. J. 48, 140–147. doi:10.1016/j.tvjl.2011.08.006
Fernandes, N. D. S. (2017). Investigação de Alcaloides de lauraceae da amazônia como tratamento para tripanosomíase e leishmaniose: Avaliação fenotípica e busca de alvos moleculares. Available at: https://tede.ufam.edu.br/handle/tede/5965 (Accessed February 8, 2021).
Ferreira, M. E., Cebrián-Torrejónb, G., and Corrales, A. S. (2011). Zanthoxylum chiloperone leaves extract: First sustainable Chagas disease treatment. J. Ethnopharmacol. 133, 986–993. doi:10.1016/j.jep.2010.11.032
Ferreira, M. E., De Arias, A. R., De Ortiz, S. T., Inchausti, A., Nakayama, H., Thouvenel, C., et al. (2002). Leishmanicidal activity of two canthin-6-one alkaloids, two major constituents of Zanthoxylum chiloperone var. angustifolium. J. Ethnopharmacol. 80 (2-3), 199–202. doi:10.1016/S0378-8741(02)00025-9
Ferreira, M. E., Nakayama, H., de Arias, A. R., Schinini, A., de Bilbao, N. V., Serna, E., et al. (2007). Effects of canthin-6-one alkaloids from Zanthoxylum chiloperone on Trypanosoma cruzi-infected mice. J. Ethnopharmacol. 109 (2), 258–263. doi:10.1016/j.jep.2006.07.028
Ferreira, P. C., Souza, G. H. M. F., and Silva, R. M. G. (2003). Atividade tripanomicida do extrato bruto de Zanthoxyllum minutiflorum, rutaceae (mama-de-porca) na infecção experimental de camundongos por trypanosoma cruzi. Available at: https://www.scielo.br/j/rsbmt/a/fVrY4Y8YnKX9vqNwQ5jtfxr/?lang=pt (Accessed February 8, 2021).
Ferreira, R. T. B., Branquinho, M. R., and Cardarelli-Leite, P. (2014). Transmissão oral da doença de Chagas pelo consumo de açaí: Um desafio para a vigilância sanitária. Visa. em Debate 2. doi:10.3395/VD.V2I4.358
Fidalgo, A. S., Da Costa, A. C., Da Silva Filho, J. D., Da Silva Cândido, D., Freitas, E. C., Pereira, L. D. S., et al. (2018). Insect vectors of chagas disease (trypanosoma cruzi) in northeastern Brazil. Rev. Soc. Bras. Med. Trop. 51, 174–182. doi:10.1590/0037-8682-0408-2017
França, T. C. C., Dos Santos, M. G., and Figueroa-Villar, J. D. (2008). Malária: Aspectos históricos e quimioterapia. Quim. Nova 31, 1271–1278. doi:10.1590/S0100-40422008000500060
Freiburghaus, F., Kaminsky, R., Nkunya, M. H. H., and Brun, R. (1996). Evaluation of African medicinal plants for their in vitro trypanocidal activity. J. Ethnopharmacol. 55, 1–11. doi:10.1016/S0378-8741(96)01463-8
Gansane, A., Sanon, S., Ouattara, P. L., Hutter, S., Olhvier, E., Azas, N., et al. (2010). Antiplasmodial activity and cytotoxicity of semi purified fractions from zanthoxylum zanthoxyloïdes lam. Bark of trunk. Int. J. Pharmacol. 6, 921–925. doi:10.3923/IJP.2010.921.925
Gessler, M. C., Nkunya, M. H. H., Mwasumbi, L. B., Heinrich, M., and Tanner, M. (1994). Screening Tanzanian medicinal plants for antimalarial activity. Acta Trop. 56, 65–77. doi:10.1016/0001-706X(94)90041-8
Goodman, C. D., Austarheim, I., Mollard, V., Mikolo, B., Malterud, K. E., McFadden, G. I., et al. (2016). Natural products from Zanthoxylum heitzii with potent activity against the malaria parasite. Malar. J. 15, 481. doi:10.1186/S12936-016-1533-X
Goodman, C. D., Hoang, A. T., Diallo, D., Malterud, K. E., McFadden, G. I., and Wangensteen, H. (2019). Anti-plasmodial effects of zanthoxylum zanthoxyloides. Planta Med. 85, 1073–1079. doi:10.1055/A-0973-0067
Goodman, C. D., Su, V., and McFadden, G. I. (2007). The effects of anti-bacterials on the malaria parasite Plasmodium falciparum. Mol. Biochem. Parasitol. 152, 181–191. doi:10.1016/J.MOLBIOPARA.2007.01.005
Hilou, A., Nacoulmaa, O. G., and Guiguemde, T. R. (2006). In vivo antimalarial activities of extracts from Amaranthus spinosus L. and Boerhaavia erecta L. in mice. J. Ethnopharmacol. 103, 236–240. doi:10.1016/j.jep.2005.08.006
Hooijmans, C. R., Rovers, M. M., Vries, R. B. M., Leenaars, M., Ritskes-Hoitinga, M., and Langendam, M. W. (2014). SYRCLE’s risk of bias tool for animal studies. BMC Med. Res. Methodol. 43, 43–49. doi:10.1186/1471-2288-14-43
Jullian, V., Bourdy, G., Georges, S., Maurel, S., and Sauvain, M. (2006). Validation of use of a traditional antimalarial remedy from French Guiana, Zanthoxylum rhoifolium Lam. J. Ethnopharmacol. 106, 348–352. doi:10.1016/J.JEP.2006.01.011
Kamanzi Atindehou, K., Schmid, C., Brun, R., Koné, M. W., and Traore, D. (2004). Antitrypanosomal and antiplasmodial activity of medicinal plants from Côte d’Ivoire. J. Ethnopharmacol. 90, 221–227. doi:10.1016/J.JEP.2003.09.032
Kato, A., Moriyasu, M., Ichimaru, M., Nishiyama, Y., Juma, F. D., Nganga, J. N., et al. (1996). Isolation of alkaloidal constituents of zanthoxylum usambarense and zanthoxylum chalybeum using ion-pair HPLC. J. Nat. Prod. 59, 316–318. doi:10.1021/NP960183W
Kelly, J. X., Smilkstein, M. J., Brun, R., Wittlin, S., Cooper, R. A., Lane, K. D., et al. (2009). LETTERS Discovery of dual function acridones as a new antimalarial chemotype. Nature 459 (7244), 270–273. doi:10.1038/nature07937
Kirira, P. G., Rukunga, G. M., Wanyonyi, A. W., Muregi, F. M., Gathirwa, J. W., Muthaura, C. N., et al. (2006). Anti-plasmodial activity and toxicity of extracts of plants used in traditional malaria therapy in Meru and Kilifi Districts of Kenya. J. Ethnopharmacol. 106, 403–407. doi:10.1016/J.JEP.2006.01.017
Lacerda, R. B. M., Freitas, T. R., Martins, M. M., Teixeira, T. L., da Silva, C. V., Candido, P. A., et al. (2018). Isolation, leishmanicidal evaluation and molecular docking simulations of piperidine alkaloids from Senna spectabilis. Bioorg. Med. Chem. 26, 5816–5823. doi:10.1016/J.BMC.2018.10.032
Larsen, A. K., Grondard, L., Couprie, J., Desoize, B., Comoe, L., Jardillier, J. C., et al. (1993). The antileukemic alkaloid fagaronine is an inhibitor of DNA topoisomerases I and II. Biochem. Pharmacol. 46, 1403–1412. doi:10.1016/0006-2952(93)90105-6
Ledoux, A., Cao, M., Jansen, O., Mamede, L., Campos, P. E., Payet, B., et al. (2018). Antiplasmodial, anti-chikungunya virus and antioxidant activities of 64 endemic plants from the Mascarene Islands. Int. J. Antimicrob. Agents 52, 622–628. doi:10.1016/J.IJANTIMICAG.2018.07.017
Lima, G. S. (2014). Ufrrj pró-reitoria de pesquisa E pós-graduação programa de pós-graduação em ciência. Rio de Janeiro, Brazil. TECNOLOGIA E INOVAÇÃO AGROPECUÁRIA TESE Estudo da Atividade Tripanossomicida e Leishmanicida de Extrato, Frações e Terpenos de croton cajucara Benth.
Lira, R., Contreras, L. M., Santa Rita, R. M., and Urbina, J. A. (2001). Mechanism of action of anti-proliferative lysophospholipid analogues against the protozoan parasite trypanosoma cruzi: Potentiation of in vitro activity by the sterol biosynthesis inhibitor ketoconazole. J. Antimicrob. Chemother. 47, 537–546. doi:10.1093/JAC/47.5.537
Mafezoli, J., Vieira, P. C., Fernandes, J. B., Da Silva, M. F. G. F., and De Albuquerque, S. (2000). In vitro activity of Rutaceae species against the trypomastigote form of Trypanosoma cruzi. J. Ethnopharmacol. 73, 335–340. doi:10.1016/S0378-8741(00)00315-9
Matthews, K. R. (2005). The developmental cell biology of Trypanosoma brucei. J. Cell Sci. 118, 283–290. doi:10.1242/JCS.01649
Melo-Neto, B., Leitão, J. M. S. R., Oliveira, L. G. C., Santos, S. E. M., Carneiro, S. M. P., Rodrigues, K. A. F., et al. (2016). Inhibitory effects of Zanthoxylum rhoifolium Lam. (Rutaceae) against the infection and infectivity of macrophages by Leishmania amazonensis. An. Acad. Bras. Cienc. 88, 1851–1861. doi:10.1590/0001-3765201620150131
Ministério da Saúde Brasil (2020). Biblioteca Virtual em Saúde do Ministério da Saúde Boletim Epidemiológico. Available at: http://bvsms.saude.gov.br (Accessed February 2, 2021).
Molina, J., Brener, Z., Romanha, A. J., and Urbina, J. A. (2000). In vivo activity of the bis-triazole D0870 against drug-susceptible and drug-resistant strains of the protozoan parasite Trypanosoma cruzi. J. Antimicrob. Chemother. 46, 137–140. doi:10.1093/JAC/46.1.137
Mota, E. F., Rosario, D. M., Silva-Veiga, A. S., Brasil, S., Silveira, F. T., and Dolabela, M. F. (2015). Biological activities of Croton palanostigma Klotzsch. Pharmacogn. Mag. 43, S96–S101. doi:10.4103/0973-1296.176109
Muganga, R., Angenot, L., Tits, M., and Frédérich, M. (2014). In vitro and in vivo antiplasmodial activity of three Rwandan medicinal plants and identification of their active compounds. Planta Med. 80, 482–489. doi:10.1055/S-0034-1368322
Musila, M. F., Dossaji, S. F., Nguta, J. M., Lukhoba, C. W., and Munyao, J. M. (2013). In vivo antimalarial activity, toxicity and phytochemical screening of selected antimalarial plants. J. Ethnopharmacol. 146 (2), 557–561. doi:10.1016/j.jep.2013.01.023
Muganga, R., Angenot, L., Tits, M., and Frederich, M. (2010). Antiplasmodial and cytotoxic activities of Rwandan medicinal plants used in the treatment of malaria. J. Ethnopharmacol. 128, 52–57. doi:10.1016/j.jep.2009.12.023
Muganga, R., Angenot, L., Tits, M., and Frédérich, M. (2014). In vitro and in vivo antiplasmodial activity of three Rwandan medicinal plants and identification of their active compounds. Planta Med. 80, 482–489. doi:10.1055/s-0034-1368322
Muthaura, C. N., Keriko, J. M., Mutai, C., Yenesew, A., Gathirwa, J. W., Irungu, B. N., et al. (2015). Antiplasmodial potential of traditional antimalarial phytotherapy remedies used by the Kwale community of the Kenyan Coast. J. Ethnopharmacol. 170, 148–157. doi:10.1016/J.JEP.2015.05.024
Nibret, E., Ashour, M. L., Rubanza, C. D., and Wink, M. (2010). Screening of some Tanzanian medicinal plants for their trypanocidal and cytotoxic activities. Phytother. Res. 24, 945–947. doi:10.1002/PTR.3066
Nyangulu, J. M., Hargreaves, S. L., Sharples, S. L., Mackay, S. P., Waigh, R. D., Duval, O., et al. (2010). Antimalarial benzo[c]phenanthridines. Bioorg. Med. Chem. Lett. 15, 2007–2010. doi:10.1016/J.BMCL.2005.02.074
Ohashi, M., Amoa-Bosompem, M., Kwofie, K. D., Agyapong, J., Adegle, R., Sakyiamah, M. M., et al. (2018). In vitro antiprotozoan activity and mechanisms of action of selected Ghanaian medicinal plants against Trypanosoma, Leishmania, and Plasmodium parasites. Phytother. Res. 32, 1617–1630. doi:10.1002/PTR.6093
O’Neill, P. M., Park, B. K., Shone, A. E., Maggs, J. L., Roberts, P., Stocks, P. A., et al. (2009). Candidate selection and preclinical evaluation of N-tert-Butyl isoquine (GSK369796), an affordable and effective 4-aminoquinoline antimalarial for the 21st century. J. Med. Chem. 52, 1408–1415. doi:10.1021/jm8012618
Parida, P., Kalita, S., Shankar, B., Yadav, R. N. S., and Das, A. (2014). Homology modeling and docking studies of <i>Plasmodium falciparum</i> telomerase reverse transcriptase with berberine and some of its derivatives. Bangladesh J. Pharmacol. 9, 96–104. doi:10.3329/BJP.V9I1.17535
Penali, L., Mulholand, D. A., Tano, K. D., Cheplogoi, P. K., and Randrianarivelojosia, M. (2007). Low antiplasmodial activity of alkaloids and amides from the stem bark of Zanthoxylum rubescens (Rutaceae). Parasite 14, 161–164. doi:10.1051/PARASITE/2007142161
Prieto, J. A., Patiño, O. J., Delgado, W. A., Moreno, J. P., and Cuca, L. E. (2011). Chemical composition, insecticidal, and antifungal activities of fruit essential oils of three colombian Zanthoxylum species. Chil. J. Agric. Res. 71, 73–82. doi:10.4067/S0718-58392011000100009
Randrianarivelojosia, M., Rasidimanana, V., and Rabarison, H. (2003). Plants traditionally prescribed to treat tazo (malaria) in the eastern region of Madagascar. Malaria J. 2, 1–9. doi:10.1186/1475-2875-2-1
Rodrigues, K. A. D. F., Amorim, L. V., Oliveira, J. M. G. De, Dias, C. N., Moraes, D. F. C., Andrade, E. H. D. A., et al. (2013). Eugenia uniflora L. Essential oil as a potential anti-leishmania agent: Effects on leishmania amazonensis and possible mechanisms of action. Evid. Based. Complement. Altern. Med. 2013, 279726. doi:10.1155/2013/279726
Rukunga, G. M., Gathirwa, J. W., Omar, S. A., Muregi, F. W., Muthaura, C. N., Kirira, P. G., et al. (2009). Anti-plasmodial activity of the extracts of some Kenyan medicinal plants. J. Ethnopharmacol. 121, 282–285. doi:10.1016/J.JEP.2008.10.033
Sandjo, L. P., de Moraes, M. H., Kuete, V., Kamdoum, B. C., Ngadjui, B. T., and Steindel, M. (2016). Individual and combined antiparasitic effect of six plant metabolites against Leishmania amazonensis and Trypanosoma cruzi. Bioorg. Med. Chem. Lett. 26, 1772–1775. doi:10.1016/J.BMCL.2016.02.044
Santiago-Brugnoli, L., Rosquete-Porcar, C., Pouységu, L., and Quideau, S. (2013). Zanthoxyfolina, un nuevo meroterpenoide y derivados del labdano aislados de las hojas de Zanthoxylum rhoifolium LAM (Rutaceae), 85–88. Available at: www.saber.ula.ve/avancesenquimica (Accessed February 8, 2021).
Silva e Silva, J. V., Cordovil Brigido, H. P., Oliveira de Albuquerque, K. C., Carvalho, J. M., Reis, J. F., Faria, L. V., et al. (2020). Flavopereirine—an alkaloid derived from geissospermum vellosii—presents leishmanicidal activity in vitro. Molecules 24, 785. doi:10.3390/MOLECULES24040785
Soares, D. C., Calegari-Silva, T. C., Lopes, U. G., Teixeira, V. L., de Palmer Paixão, I. C. N., Cirne-Santos, C., et al. (2012). Dolabelladienetriol, a compound from dictyota pfaffii algae, inhibits the infection by leishmania amazonensis. PLoS Negl. Trop. Dis. 6, e1787. doi:10.1371/JOURNAL.PNTD.0001787
Sousa, F., das, C. A., Soares, H. V. A., Lemos, L. E. A. S., Reis, D. M., Silva, W. C. D., et al. (2019). Perfil epidemiológico de doenças negligenciadas de notificação compulsória no Brasil com análise dos investimentos governamentais nessa área. Res. Soc. Dev. 9, e62911610. doi:10.33448/RSD-V9I1.1610
Stangeland, T., Wangensteen, H., Katuura, E., Lye1, K. A., and Paulsen, B. S. (2010). Antioxidant and anti-plasmodial activity of extracts from three Ugandan medicinal plants. J. Med. Plants Res. 18, 1916–1923. doi:10.5897/JMPR10.44
Sriwilaijareon, N., Petmitr, S., Mutirangura, A., Ponglikitmongkol, M., and Wilairat, P. (2002). Stage specificity of Plasmodium falciparum telomerase and its inhibition by berberine. Parasitol. Int. 51, 99–103. doi:10.1016/S1383-5769(01)00092-7
Tanoh, E., Boué, G., and Nea, F. (2020). Seasonal effect on the chemical composition, insecticidal properties and other biological activities of zanthoxylum leprieurii guill. and perr. essential oils. Foods 4, 66–87. doi:10.3390/foods9050550
Tchinda, A. T., Fuendjiep, V., Sajjad, A., Matchawe, C., Wafo, P., Khan, S., et al. (2009). Bioactive compounds from the fruits of zanthoxylum leprieurii. Pharmacologyonline 1, 406–415. Available at: https://www.researchgate.net/publication/257487101_BIOACTIVE_COMPOUNDS_FROM_THE_FRUITS_OF_ZANTHOXYLUM_LEPRIEURII (Accessed February 8, 2021).
Thouvenel, C., Gantier, J. C., Duret, P., Fourneau, C., Hocquemiller, R., Ferreira, M. E., et al. (2003). Antifungal compounds from Zanthoxylum chiloperone var. angustifolium. Phytother. Res. 17, 678–680. doi:10.1002/PTR.1137
Tian, Y., Zhang, C., and Guo, M. (2017). Comparative study on alkaloids and their anti-proliferative activities from three Zanthoxylum species. BMC Complement. Altern. Med. 17, 460–516. doi:10.1186/s12906-017-1966-y
Urbina, J. A., Payares, G., Sanoja, C., Molina, J., Lira, R., Brener, Z., et al. (2003). Parasitological cure of acute and chronic experimental Chagas disease using the long-acting experimental triazole TAK-187. Activity against drug-resistant Trypanosoma cruzi strains. Int. J. Antimicrob. Agents 21, 39–48. doi:10.1016/S0924-8579(02)00274-1
Were, P. S., Kinyanjui, P., Gicheru, M. M., Mwangi, E., and Ozwara, H. S. (2010). Prophylactic and curative activities of extracts from warburgia ugandensis sprague (canellaceae) and zanthoxylum usambarense (engl.) kokwaro (rutaceae) against Plasmodium knowlesi and Plasmodium berghei. J. Ethnopharmacol. 130, 158–162. doi:10.1016/J.JEP.2010.04.034
Glossary
AC activity classification
Bolivia sensitive strain of T. cruzi
DeCs descriptors in health sciences
NDs neglected diseases
NTDs neglected tropical diseases
BVS Virtual Health Library–biblioteca virtual da saúde
Mesh medical subjects
N not relevant to the item
S showed relevance to the item
M presented medium relevance to the item
IC50 50% inhibitory concentration
CC50 50% cytotoxic concentration
SI selectivity index
Ext extract
EtOAc ethyl acetate
H2O water
CH2CL2 dichloromethane
EtOH ethanol
MeOH methanol
PE petroleum ether
Hex hexane
CHCL3 chloroform
MCW dichloromethane, methanol, and water
Act acetone
lr root
s shell
n-hex n-hexane
F fraction
FA fraction of alkaloids
b bark
sb stem bark
rb root bark
l leaves
fr fruit
ND not determined
SND strain not determined
I inactive
MA moderately active
A active
VA very active
GA good active
LA low active
FCR3 Plasmodium falciparum-resistant strain
W2 Plasmodium falciparum-resistant strain
K1 Plasmodium falciparum-resistant strain
ENT 30 Plasmodium falciparum-resistant strain
FcB1 Plasmodium falciparum-resistant strain
PFB Plasmodium falciparum-resistant strain
FcM29 Plasmodium falciparum-resistant strain
NF54 Plasmodium falciparum-sensitive strain
3D7 Plasmodium falciparum-sensitive strain
F32 Plasmodium falciparum-sensitive strain
D6 Plasmodium falciparum-sensitive strain
MRA-285 Plasmodium falciparum-sensitive strain
MRC5 fibroblasts
K562S chronic myeloid leukemia
A.s. artemia saline
L-6 rat skeletal myoblasts
J774 murine macrophages
Jurkat leukemia/lymphoma T lymphocytes
VERO monkey kidney fibroblasts
MCF-7 human breast cancer
HPM hamster peritoneal macrophage
BALB/c Albino mice c genetic line
PCSM peritoneal cavity of Swiss mice
M macrophages
THP-1 human acute monocytic leukemia
But butanol
Dm28c resistant strain of T. cruzi
Y sensitive strain of T. cruzi
RAW 264.7 murine macrophages
WI-38 diploid human cell line composed of fibroblasts from lung tissue
HL-60 human leukemia cells
RAW rat macrophages
Keywords: Malaria, Leishmaniasis, Chagas disease, sleeping sickness, Zanthoxylum
Citation: Correa-Barbosa J, Sodré DF, Nascimento PHC and Dolabela MF (2023) Activity of the genus Zanthoxylum against diseases caused by protozoa: A systematic review. Front. Pharmacol. 13:873208. doi: 10.3389/fphar.2022.873208
Received: 10 February 2022; Accepted: 15 November 2022;
Published: 09 January 2023.
Edited by:
Karl Hassan, The University of Newcastle, AustraliaReviewed by:
Wanderley De Souza, Federal University of Rio de Janeiro, BrazilDebora Botura Scariot, Northwestern University, United States
Florenci Vicent González, University of Jaume I, Spain
Copyright © 2023 Correa-Barbosa, Sodré, Nascimento and Dolabela. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Maria Fâni Dolabela, ZmFuaUB1ZnBhLmJy