
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Pharmacol. , 18 January 2022
Sec. Pharmacogenetics and Pharmacogenomics
Volume 12 - 2021 | https://doi.org/10.3389/fphar.2021.773848
This article is part of the Research Topic Brief Research Reports in Pharmacogenetics and Pharmacogenomics: 2022 View all 5 articles
 Salvador Resino1,2*
Salvador Resino1,2* María Ángeles Jiménez-Sousa1,2
María Ángeles Jiménez-Sousa1,2 Julià Blanco2,3
Julià Blanco2,3 Yolanda M. Pacheco4Jorge del Romero5Joaquim Peraire2,6Ana Virseda-Berdices1,2María José Muñoz-Gómez1,2Carlos Galera-Peñaranda7Lucio Jesus García-Fraile2,8
Yolanda M. Pacheco4Jorge del Romero5Joaquim Peraire2,6Ana Virseda-Berdices1,2María José Muñoz-Gómez1,2Carlos Galera-Peñaranda7Lucio Jesus García-Fraile2,8 José M. Benito9,10†
José M. Benito9,10† Norma Rallón9,10†on behalf of CoRIS and the HIV Biobank integrated in the Spanish AIDS Research Network Project RIS/EPICLIN 10_2015
Norma Rallón9,10†on behalf of CoRIS and the HIV Biobank integrated in the Spanish AIDS Research Network Project RIS/EPICLIN 10_2015Background: The lack of the recovery of CD4+ T-cells (CD4+ recovery) among immunodeficiency virus (HIV)-infected patients on antiretroviral therapy (ART) is not well known. We aimed to analyze the association between single nucleotide polymorphisms (SNPs) underlying vitamin D metabolism and the CD4+ recovery in naïve HIV-infected patients who started ART with low baseline CD4+.
Methods: We conducted a retrospective study in 411 naïve individuals with plasma HIV load >200 copies/mL and CD4+ <200 cells/mm3. During 24 months of follow-up, all patients had plasma HIV load <50 copies/mL. DNA genotyping was performed using the Sequenom MassARRAY platform. The outcome variable was the change in CD4+ during the study.
Results: CD4+ recovery was higher in patients carrying DBP rs7041 AA genotype (AA versus CC/AC) and DHCR7 rs3829251 AA genotype (AA versus GG/AG) (p-value < 0.05). DBP rs7041 AA genotype was linked to increase in CD4+ (adjusted arithmetic mean ratio (aAMR) = 1.22; q-value = 0.011), increase in CD4+ ≥P75th [adjusted odds ratio (aOR) = 2.31; q-value = 0.005], slope of CD4+ recovery (aAMR = 1.25; q-value = 0.008), slope of CD4+ recovery ≥ P75th (aOR = 2.55; q-value = 0.005) and achievement of CD4+ ≥500 cells/mm3 (aOR = 1.89; q-value = 0.023). Besides, DHCR7 rs3829251 AA genotype was related to increase in CD4+ (aAMR = 1.43; q-value = 0.031), increase in CD4+ ≥P75th (aOR = 3.92; q-value = 0.030), slope of CD4+ recovery (aAMR = 1.40; q-value = 0.036), slope of CD4+ recovery ≥ P75th (aOR = 3.42; q-value = 0.031) and achievement of CD4+ ≥500 cells/mm3 (aOR = 5.68; q-value = 0.015).
Conclusion: In summary, DHCR7 rs3829251 and DBP rs7041 polymorphisms were associated with CD4+ recovery in HIV-infected patients who started cART with low CD4+ T-cell counts.
Combination antiretroviral therapy (cART) tends to achieve undetectable plasma viral load levels in the vast majority of the human immunodeficiency virus (HIV)-infected patients treated. This control of viral replication allows the recovery of CD4+ T-cells (CD4+ recovery) in peripheral blood, and many immune functions are restored (Panel de expertos de Gesi, 2011; Thompson et al., 2012). Despite this, there is still a significant percentage (around 30%) of HIV-infected patients who fail to have complete CD4+ recovery (≥500 CD4+ T‐cell count/µl) after long periods of cART (Yang et al., 2020). Those patients who maintain low CD4+ counts remain at risk of acquired immunodeficiency syndrome (AIDS) progression, developing non-AIDS-related morbidity, and dying (Baker et al., 2008; Kelley et al., 2009; Helleberg et al., 2013).
The causes of this lack of CD4+ recovery among cART-treated patients are not well known, but it appears to be a complex and multifactorial phenomenon (Yang et al., 2020). In this regard, many factors involved in CD4+ recovery have been described, among which include age, low CD4+ T-cells nadir, severe immunodeficiency at the time of cART initiation, low baseline CD4/CD8 ratio, immune exhaustion, abnormal immune activation, reduced output in the bone marrow and thymic, increased senescence and apoptosis of T-cells, lymphoid tissue fibrosis, imbalance in Treg and Th17 cells, microbial translocation, persistent HIV replication, and host genetic background, among others (Yang et al., 2020). However, all these factors do not fully explain the great variability of immune reconstitution in cART-treated patients.
Vitamin D (VitD) deficiency is common in HIV-infected patients (around 70–85%) (Mansueto et al., 2015; Gois et al., 2017). VitD regulates different antimicrobial pathways of immunity that can be crucial against HIV infection (Jiménez-Sousa et al., 2018). Besides, VitD deficiency has been related to higher HIV viral load values in plasma, inflammation, immune activation, decreased CD4+ T-cells, rapid AIDS progression in cART-naïve patients, and impaired CD4+ recovery in HIV-infected patients on cART (Jiménez-Sousa et al., 2018).
Most VitD is produced in the body, and only a small percentage is ingested in the diet. In the first step, pro-VitD (7-dehydro-cholesterol) is transported to the skin, where it is isomerized to pre-VitD (cholecalciferol) by ultraviolet irradiation (Herrmann et al., 2017). However, 7-dehydrocholesterol reductase (DHCR7) may oxidate 7-dehydro-cholesterol to cholesterol, decreasing the amount of 7-DHC available for photochemical conversion to VitD in the skin (Prabhu et al., 2016). Next, pre-VitD is hydroxylated to 25-hydroxy-VitD [25(OH)D] in the liver by cytochrome P450 enzymes (CYP27A1 and CYP2R1). Then, 25 (OH)D is transported to the kidneys by the vitamin D-binding protein (DBP), where the 25-hydroxyvitamin D3 1-alpha-hydroxylase (CYP27B1) forms calcitriol [1,25 (OH)2D], which is the metabolically active form (Herrmann et al., 2017; Jiménez-Sousa et al., 2018). Besides, 25 (OH)D is catabolized by CYP3A4, and CYP24A1 catabolizes 1,25 (OH)2D. In the nucleus, VitD binds to vitamin D receptor (VDR) and promote the formation of a heterodimer with the retinoid X receptor alpha (RXRA), which binds to vitamin D response elements (VDRE), initiating the transcription of more than 4,000 genes (around 5% of the human genome) (Herrmann et al., 2017; Jiménez-Sousa et al., 2018) (Figure 1).
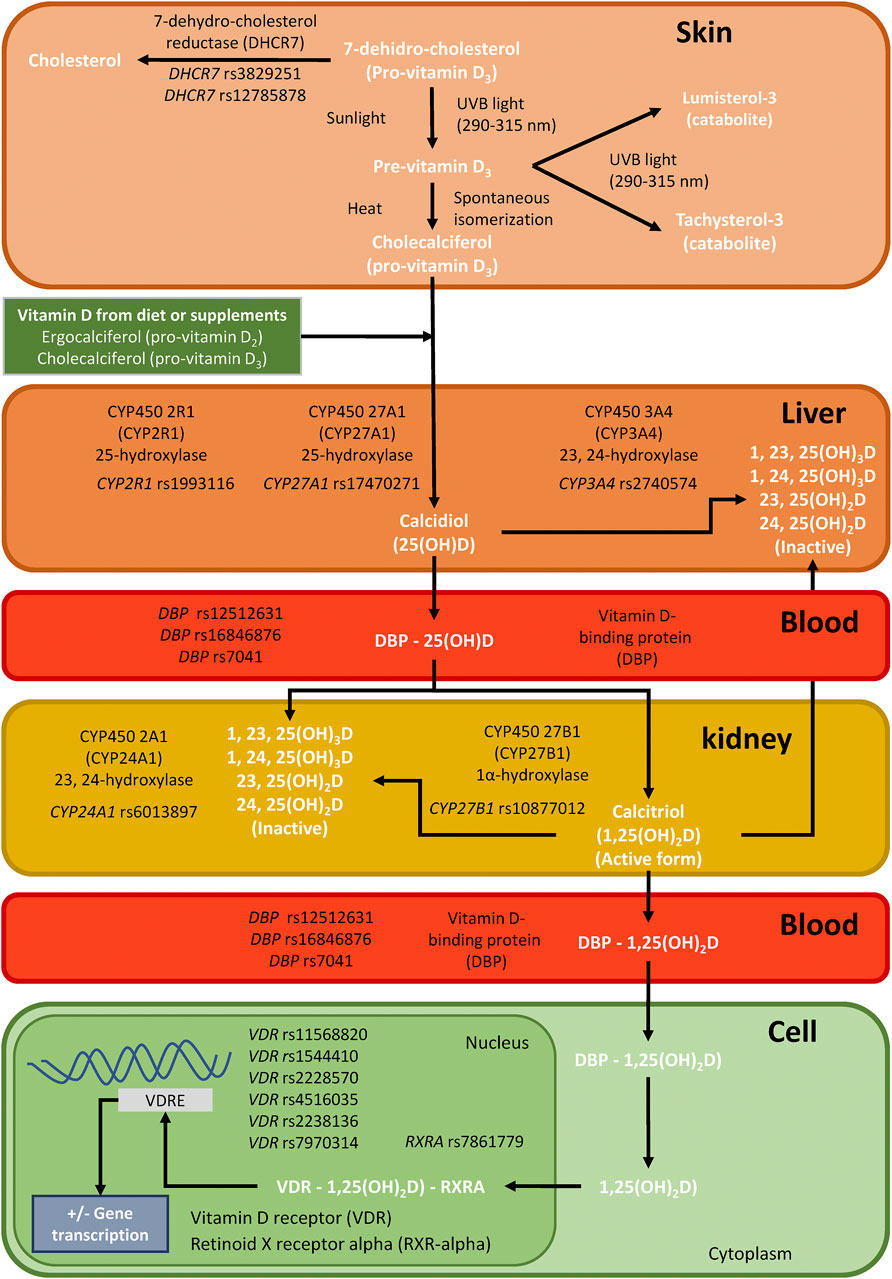
FIGURE 1. Summary of single nucleotide polymorphisms (SNPs) analyzed for this study in genes related to the synthesis, transport, and mechanism of action of vitamin D in the body.; Abbreviations: DHCR7, 7-dehydrocholesterol reductase; CYP2R1, cytochrome P450 enzymes; CYP27A1, cytochrome P450 enzymes; CYP27B1, cytochrome P450 enzymes; CYP3A4, cytochrome P450 enzymes; CYP24A1, cytochrome P450 enzymes DBP, vitamin D-binding protein; VDR, vitamin D receptor; and RXRA, retinoid X receptor alpha.
Several single nucleotide polymorphisms (SNPs) in genes related to VitD metabolism (DHCR7, CYP2R1, CYP27A1, CYP27B1, CYP3A4, CYP24A1, DBP, VDR, and RXRA, among others) have been associated with plasma levels of 25(OH)D and infectious diseases (Jolliffe et al., 2016). In HIV infection, VDR SNPs have been related to protection against HIV infection (de la Torre et al., 2008; Alagarasu et al., 2009; Torres et al., 2010). Also, DHCR7, DBP, VDR, and CYP27B1 SNPs were associated with AIDS progression in ART-naïve HIV-infected patients of European origin (Eales et al., 1987; Nieto et al., 2004; Moodley et al., 2013; Laplana et al., 2014; Jiménez-Sousa et al., 2019a; Jiménez-Sousa et al., 2019b; Jiménez-Sousa et al., 2020). However, there is no data about the relationship between these SNPs in genes related to VitD metabolism and immune recovery in patients on cART.
We aimed to analyze the association between genetic variants underlying VitD metabolism (DHCR7, CYP2R1, CYP27A1, CYP27B1, CYP3A4, CYP24A1, DBP, VDR, and RXRA genes) and the CD4+ recovery in naïve HIV-infected patients who started cART with low baseline CD4+ T cells (<200 cells/mm3).
We carried out a retrospective study in 411 naïve HIV-infected patients who started cART, included in two cohorts, the Spanish AIDS Research Network cohort (CoRIS, see Appendix 1) and the AIDS Research Institute IrsiCaixa-HIVACAT cohort. This cohort study has been previously described (Restrepo et al., 2019). Patients signed informed consent before participating in the study. The Ethics Committee of the “Fundación Jiménez Díaz” approved the research project (Ref.: PIC 52/2015_FJD). This study was performed under the Declaration of Helsinki.
The inclusion criteria were: 1) at baseline: naïve-ART, plasma HIV-RNA >200 copies/mL, CD4+ <200 cells/mm3, and DNA sample available; 2) during the follow-up period (2 years): plasma HIV-RNA <50 copies/mL, periodic monitoring of CD4 and plasma HIV-RNA. The two cohorts totaled 6,160 HIV-infected patients, of which 4,000 started cART, but only 1,259 had CD4+ <200 cells/mm3 before cART initiation. Of them, only 503 patients had a minimum follow-up period of 24 months, but 92 patients had no undetectable viral load (VL, HIV-RNA <50 copies/mL) after cART or DNA sample available. Finally, 411 patients met all the inclusion criteria. Besides, 145 healthy donors from the “Centro de Transfusión de la Comunidad de Madrid” were used as Control-group (HIV, HCV, and HBV seronegative subjects).
Demographic, clinical, virological, and laboratory data were collected from medical records. Gender was by self-identification and there were no transgender people. Time since HIV diagnosis was calculated from the first positive blood test for HIV. The mode of transmission (injecting drug use and sexual behavior) was inferred from the medical record. Hepatitis C and hepatitis B coinfection were determined by a standard laboratory test. The clinic management of patients was carried out following national clinical guidelines (Panel de expertos de Gesi, 2011).
Blood samples were collected by venipuncture in EDTA tubes. The blood samples were then sent to the HIV HGM BioBank (http://hivhgmbiobank.com/?lang=en), where the samples were processed and frozen immediately upon receipt. DNA isolation was performed from peripheral blood mononuclear cells using a QIAamp DNA kit (Qiagen, Spain).
We selected 17 SNPs of VitD metabolism [DHCR7 (rs3829251 and rs12785878), CYP2R1 (rs1993116), CYP27A1 (rs17470271), CYP27B1 (rs10877012), CYP3A4 (rs2740574), CYP24A1 (rs6013897), DBP (rs12512631, rs16846876, and rs7041), VDR (rs11568820, rs1544410, rs2228570, rs4516035, rs2238136, rs7970314), and RXRA (rs7861779)] (Figure 1), which have been related to circulating concentrations of VitD and non-skeletal diseases (Jolliffe et al., 2016). DNA genotyping was performed using the iPLEX® Gold technology and Agena Bioscience’s MassARRAY platform (San Diego, CA, United States) in the Spanish National Genotyping Center (CeGen; http://www.cegen.org/). All SNPs had a DNA genotyping success rate greater than 95%.
The validation and quality control of the genotyping process was performed using: 1) Negative controls, no template controls (NTC). The NTCs were used to confirm that no artifacts associated with design or chemistry were generated during the genotyping assays; 2) Positive controls. As positive controls, a trio of Coriell samples from the Human Genetic Cell Repository (NA10861, NA11994, and NA11995) was included in each genotyping assay. These Coriell samples were included in the set of genotyped samples in 1000GENOMES_phase_3 (EUR), so we could confirm the concordance of our results with those obtained for the 1000 Genomes Project. During the genotyping assays, we have replicated more than 18% of obtained genotypes, and we have observed a total concordance among replicated samples. Additionally, a phenotype-blind genotyping process was followed, since all patients who met the inclusion criteria were genotyped anonymously without information on their phenotype.
Outcome variables were related to changes in CD4+ values during the 24 months of the study. The outcome variables analyzed were: 1) increases in CD4+ (ΔCD4+, continuous), which is the difference between the baseline and end of follow-up (month 24). 2) increases in CD4+ ≥P75th (dichotomous). 3) slope or gradient of CD4+ recovery (continuous), which is the ratio between the change in CD4+ and the time elapsed. 4) slope of CD4+ recovery ≥ P75th (dichotomous). 5) achieving CD4+ at the end of follow-up ≥500 cells/mm3 (dichotomous).
Both SPSS 22.0 (IBM Corp., Chicago, United States) and Stata 15.0 (StataCorp, Texas, United States) were used to carry out the statistical analysis. P-values < 0.05 were considered significant, and all tests were two-tailed.
For the descriptive study, the Chi-squared test or Fisher´s exact tests were used to compare categorical data and evaluate the Hardy-Weinberg equilibrium (HWE). Mann-Whitney U test and Kruskal-Wallis tests were used to compare continuous variables. The genetic association study between SNPs and clinical outcomes was assessed according to dominant, recessive, and additive models by Generalized Linear Models (GLMs). Specifically, we used a GLM with a gamma distribution (log-link) for continuous variables and a GLM with a binomial distribution (logit-link) for dichotomous variables, which reported the arithmetic mean ratio (AMR), and the odds ratio (OR), respectively. Additionally, the raw p-values were corrected using the false discovery rate (FDR) with Benjamini and Hochberg (q-values), a widely used multiple comparison adjustment method. The selected SNPs with q-values < 0.05 were evaluated by multivariate regression using GLM models adjusted by the main clinical characteristics at baseline: age, gender, Caucasian origin, hepatitis C and hepatitis B coinfection, HIV transmission by intravenous drugs use (IDU), cART regimen with protease inhibitors (PI), time since HIV diagnosis, and baseline CD4+ T cell count.
In addition, pairwise linkage disequilibrium (LD) analysis was computed by Haploview 4.2 software (Barrett et al., 2005). Haplotype-based association testing was performed using the PLINK package (Purcell et al., 2007).
Table 1 shows the baseline characteristics of HIV-infected patients, whose median age was 40 years, around 79% were male, and 13% were coinfected with HCV or HBV. All patients were naïve ART and had CD4+ <200 cells/mm3 and detectable plasma HIV-RNA.
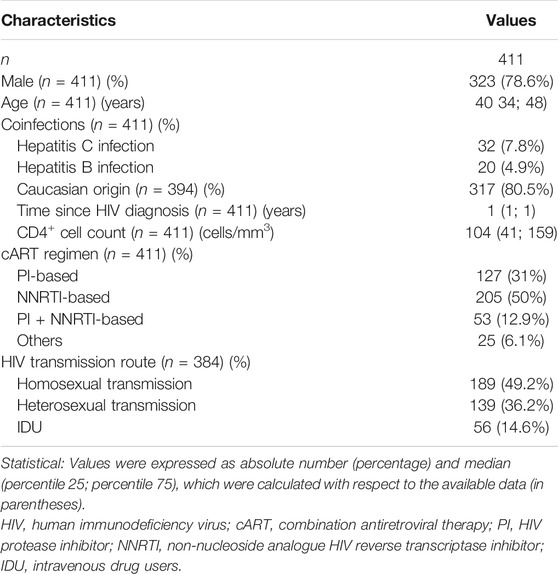
TABLE 1. Clinical and epidemiological characteristics at baseline of HIV infected patients who started cART with very low CD4+T-cells count (<200 cells/mm3).
The distribution of SNPs related to the VitD pathway in healthy controls and HIV-infected patients is shown in Supplementary Table S1. All SNPs had values for minor allelic frequency (MAF) higher than 5%, and they were in HWE (q-value > 0.05). Healthy controls and HIV-infected patients had similar genotypic frequencies. All SNPs’ genotypic and allelic frequencies were in line with the NCBI SNP database for the European population (http://www.ncbi.nlm.nih.gov/projects/SNP/).
We also analyzed the LD between SNPs of the same gene (Supplementary Figure S1), finding very high LD values (D’ = 1.0) for VDR SNPs (rs4516035, rs11568820, and rs7970314) and DHCR7 SNPs (rs12785878 and rs3829251). However, r2 values were low for all SNPs (except rs11568820 vs rs7970314), indicating that each SNP provides different information.
Supplementary Table S2 shows the association between SNPs and CD4+ recovery by unadjusted GLMs. Overall, after correcting the p-values for multiple testing (FDR - Benjamini–Hochberg procedure), CD4+ recovery was higher in patients carrying DBP rs7041 AA genotype (AA versus CC/AC; recessive model) and DHCR7 rs3829251 AA genotype (AA versus GG/AG; recessive model) (q-value <0.05).
Specifically, DBP rs7041 AA and DHCR7 rs3829251 AA carriers had higher values of CD4+ T-cell count increased (p-value = 0.013 (Figure 2A) and p-value = 0.011 (Figure 2B), respectively), CD4+ T-cell count increased ≥ P75th (p-value = 0.001 (Figure 2C) and p-value = 0.011 (Figure 2D), respectively), slope of CD4+ recovery (p-value = 0.017 (Figure 2A) and p-value = 0.008 (Figure 2B), respectively), slope of CD4+ recovery ≥ P75th (p-value = 0.001 (Figure 2C) and p-value = 0.012 (Figure 2D), respectively), and percentage of patients achieving CD4+ ≥500 cells/mm3 (p-value = 0.002 (Figure 2C) and p-value = 0.005 (Figure 2D), respectively) than patients with other genotypes.
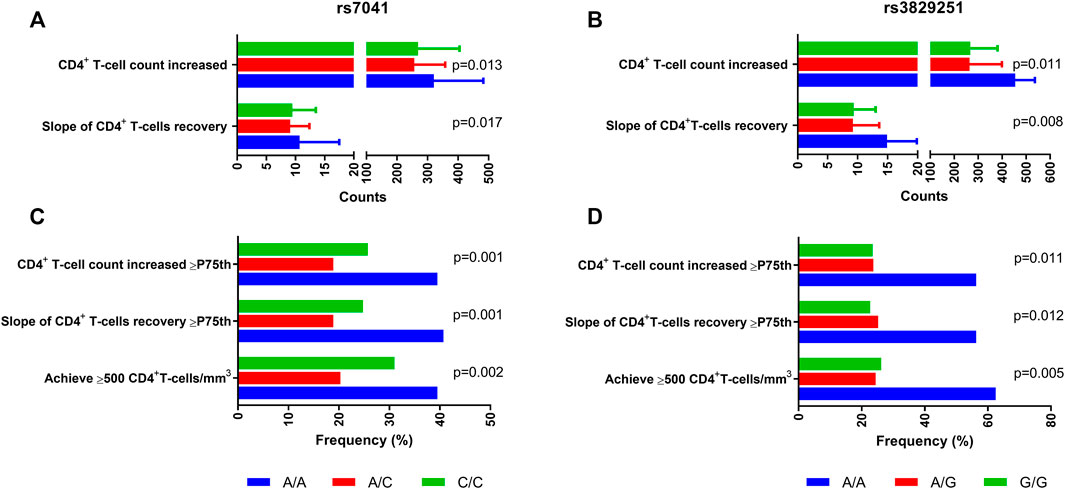
FIGURE 2. Summary of outcome variables of CD4+ recovery in HIV-infected patients who started ART with very low CD4+ T-cells count (<200 cells/mm3) according to DBP rs7041 and DHCR7 rs3829251 polymorphisms. Statistics: Values were expressed as percentages and median (percentile 25; percentile 75). P-values were calculated by Chi-square and Kruskal-Wallis Tests.; Abbreviations: DBP, vitamin D binding protein; DHCR7, 7-Dehidrocolesterol reductase; HIV, human immunodeficiency virus.
Then, the association of DBP rs7041 and DHCR7 rs3829251 polymorphisms with CD4+ recovery was evaluated by adjusted GLMs (Table 2). DBP rs7041 AA genotype was linked to increase in CD4+ (adjusted arithmetic mean ratio (aAMR) = 1.22; q-value = 0.011), increase in CD4+ ≥P75th (adjusted odds ratio (aOR) = 2.31; q-value = 0.005), slope of CD4+ recovery (aAMR = 1.25; q-value = 0.008), slope of CD4+ recovery ≥ P75th (aOR = 2.55; q-value = 0.005) and achievement of CD4+ ≥500 cells/mm3 (aOR = 1.89; q-value = 0.023). Besides, DHCR7 rs3829251 AA genotype was related to increase in CD4+ (aAMR = 1.43; q-value = 0.031), increase in CD4+ ≥P75th (aOR = 3.92; q-value = 0.030), slope of CD4+ recovery (aAMR = 1.40; q-value = 0.036), slope of CD4+ recovery ≥ P75th (aOR = 3.42; q-value = 0.031) and achievement of CD4+ ≥500 cells/mm3 (aOR = 5.68; q-value = 0.015).
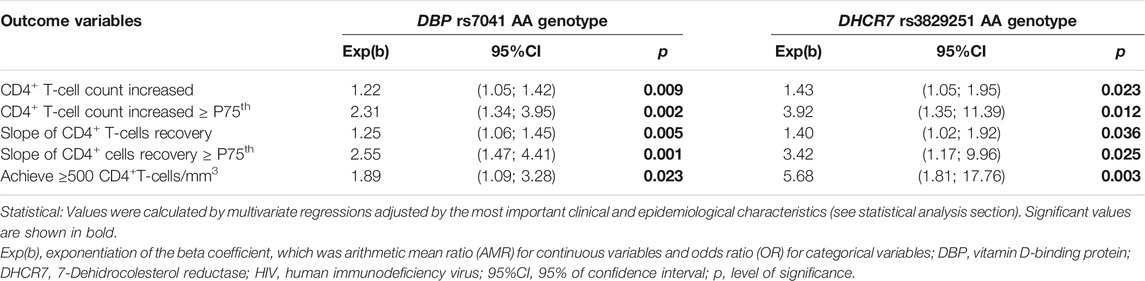
TABLE 2. Association of DBP rs7041 and DHCR7 rs3829251 single nucleotide polymorphisms with CD4+ T-cells recovery under a recessive inheritance model in HIV-infected patients who started ART with very low CD4+T-cells count (<200 cells/mm3).
Supplementary Table S3 shows the association of VDR, DBP, DHCR7 haplotypes with CD4+ recovery. We found some significant associations for the three genes and three dichotomic outcomes analyzed (p-value < 0.05), but these disappeared after FDR adjustment (q-value > 0.05).
Genetic background variability may confer differences in CD4+recovery in cART-treated HIV-infected patients (Guzmán-Fulgencio et al., 2013; Hartling et al., 2014; Yong et al., 2016; Hartling et al., 2017; Masson et al., 2018; Medrano et al., 2018; García et al., 2019). Our study found HIV-infected patients carrying the DBP rs7041 AA and DHCR7 rs3829251 AA genotypes had a better CD4+ recovery after starting cART with low CD4+ T-cells count (<200 cells/mm3). We analyzed five outcome variables related to CD4+ recovery, and we found that rs7041 and rs3829251 were significantly associated with all outcome variables, which seems to indicate a clear impact on immune reconstitution. To our knowledge, this is the first time that these two SNPs (DBP rs7041 AA and DHCR7 rs3829251 AA) have been linked to CD4+ recovery.
Multiple mechanisms have been reported by which VitD could influence the immune system (Jiménez-Sousa et al., 2018). VitD induces antiviral response against HIV by promoting antiviral gene expression, reducing the CCR5 expression on CD4+ T-cells, promoting an HIV-1-restrictive CD4+HLA-DR- phenotype, and decreasing the impact of TNF-α in upregulating HIV replication in latently infected CD4+ T-cells (Aguilar-Jimenez et al., 2016; Nunnari et al., 2016). Interestingly, DBP rs7041 and DHCR7 rs3829251 polymorphisms contribute to variation in plasma VitD level (Ahn et al., 2010; Lu et al., 2012), which has also been related to CD4+recovery (Jiménez-Sousa et al., 2018). Therefore, DBP rs7041 and DHCR7 rs3829251 polymorphisms may be genetic variants to be further explored to increase our current knowledge of the mechanistic pathways involved in the poor immune recovery status and, also, assessing them as a potential target for improving immune reconstitution and prevent disease progression and death.
The DBP is a highly polymorphic gene that contains 13 exons and 12 introns and encodes an α2-glycosylated globulin. DBP SNPs have been associated with plasma VitD levels, affecting the function of VitD (Jolliffe et al., 2016). DBP rs7041 polymorphism is a missense variation of GAT→GAG, which changes aspartic acid at position 416 to glutamic acid. Rs7041 has been implicated in the generation of different DBP isoforms with different affinity for VitD metabolites (Speeckaert et al., 2006; Sinotte et al., 2009), which seem to affect the VitD delivery in the cell (Hibler et al., 2012). Additionally, DBP rs7041 polymorphism is related to the pathogenesis of various infectious diseases, such as coronavirus disease 2019 (COVID-19) (Karcioglu Batur and Hekim, 2020), susceptibility to HCV infection (Xie et al., 2018), chronic hepatitis C progression (Petta et al., 2013; Azevedo et al., 2017), respiratory syncytial virus bronchiolitis (Randolph et al., 2014) and AIDS progression in ART-naïve HIV-infected patients (Eales et al., 1987), as well as the response to antiviral therapy in HCV-infected patients (Falleti et al., 2012). However, the association with the cART response in HIV-infected patients has not yet been described. In our study, we found a positive impact of DBP rs7041 AA genotype on CD4+ recovery in naïve patients who started cART, but due to the high variability of the DBP protein (Jolliffe et al., 2016), we do not rule out that other DBP SNPs may be involved in the observed effect on CD4+ recovery.
The DHCR7 gene encodes 7-dehydrocholesterol reductase that catalyzes the transformation of pro-VitD (7-dehydro-cholesterol) into cholesterol, acting as a switch between cholesterol and vitamin D synthesis (Prabhu et al., 2016). Regarding the genetic factors, several studies have reported that DHCR7 rs3829251 contributes to variation in plasma VitD levels (Ahn et al., 2010; Wang et al., 2010; Lu et al., 2012), although its functional role is still unknown. Rs3829251 SNP is located within an intronic region of the NADSYN1 gene and upstream of the DHCR7 gene. Using the rVarBase database (Guo et al., 2016), we observed that rs3829251 SNP is implicated in changes in the chromatin state in different cell lines and tissues. These chromatin modifications can affect the DNA accessibility to transcription factors and thus, contribute to changes in the expression of both NADSYN1 and DHCR7 genes. In fact, rs3829251 has been associated with the DHCR7 expression in the literature (Strawbridge et al., 2014). It seems to have a regulatory role on NADSYN1 gene as expression and to splice quantitative trait loci (eQTL and sQTL), as described in numerous tissues by the Genotype-Tissue Expression Portal (GTEx Portal, https://gtexportal.org). Additionally, it is also important to note that a large number of SNPs between the nicotinamide adenine dinucleotide (NAD) synthetase‐1 (NADSYN1) gene and DHCR7 gene (NADSYN1/DHCR7 locus) are in high linkage disequilibrium (LD). Thus, we cannot rule out that other SNPs in high LD with rs3829251 SNP could be the causal polymorphism. Further studies would be needed to corroborate its functional role.
All newly diagnosed HIV-infected patients should initiate cART, regardless of CD4+ T cell count, to decrease the risk of HIV transmission and prevent the progression of AIDS and the occurrence of AIDS-related events (Saag et al., 2018). Late presentation to HIV care is a significant and persistent problem worldwide (Kranzer et al., 2012; Suárez-García et al., 2016; Croxford et al., 2018), even in developed countries with good healthcare access (Croxford et al., 2018). Due to the delay in the diagnosis of HIV infection, late presenters are a significant group of patients (Mocroft et al., 2013; Darling et al., 2016), who tend to have CD4+ T cell below 200 cells/mm3 in many cases and start cART late (Antinori et al., 2011), and thus, having worse CD4+ recovery rates (Negredo et al., 2010; Pérez-Molina et al., 2012).
Many reports have shown the inability to have a CD4+ recovery after long periods of cART, evaluating the outcomes in terms of immunological response and disease progression (Yang et al., 2020). There is no consensus on the definition of incomplete immune reconstitution. Our study analyzed the threshold for CD4+ T‐cell count >500/µL, which is one of the most accepted as an adequate immune response to cART since HIV‐1‐infected patients with >500 CD4+ T‐cells/µl have morbidity and mortality rates similar to those of HIV-negative people (Yang et al., 2020). Moreover, we also analyzed other immunological outcomes that indicate efficient CD4+ recovery but have a difficult clinical interpretation. However, we have not evaluated the relationship between poor immune recovery and clinical outcomes.
Strengths: 1) We studied a very representative sample of the Spanish population infected with HIV because our cohort comes from a large number of hospitals spread throughout Spain. 2) We analyzed patients who had a baseline CD4+ T-cells <200 cells/mm3 and undetectable viral load during the whole follow-up period, strict criteria that help better define profiles of patients who recover and do not recover CD4+ T-cells. 3) The study period was the same in all patients (24 months after starting cART). 4) We evaluated different threshold values of CD4+ recovery that allow greater certainty when confirming the statistical association with CD4+ recovery.
Limitations: 1) Selection bias due to retrospective design and restrictive inclusion criteria. 2) Low statistical power due to the relatively small sample size, which may have affected the detection of less strong associations. 3) Our study was mostly performed on Caucasian individuals and more studies should be done in other populations. 4) Genotyping of more polymorphisms within genes involved in the VitD metabolim could provide additional insight into CD4+ T-cells recovery. 5) We have not performed functional assays to confirm the effect of DHCR7 rs3829251 and DBP rs7041 polymorphisms on CD4+ T-cells recovery.
In summary, DHCR7 rs3829251 and DBP rs7041 polymorphisms were associated with CD4+ recovery in HIV-infected patients who started cART with low CD4+ T-cell counts. These SNPs in the VitD pathway could help detect HIV-infected patients with lower likelihood of CD4+ recovery after cART. However, further studies with more polymorphisms, in different ethnicities, and with larger samples are needed about the role of VitD genetic variants on CD4+ recovery in late presenters initiating cART.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: (https://data.mendeley.com/datasets/6pf269zztf/1).
This study was approved by the “Fundación Jiménez Díaz” Ethics Committee (Ref.: PIC 52/2015_FJD). All patients/participants provided their written informed consent.
Funding body: SR, JMB, and NR. Study concept and design: SR, MAJ-S, JMB, and NR. Patients’ selection and clinical data acquisition: JB, YP, JR, JP, CG-P, LJ-GF, JMB, and NR. Sample preparation, DNA isolation, and genotyping: MAJ-S, AV-B, and MJ-MG. Statistical analysis and interpretation of data: MAJ-S and SR. Writing of the manuscript: SR and MAJ-S. Critical revision of the manuscript for important intellectual content: JMB, and NR. Study supervision: SR, JMB, and NR. All authors read and approved the final manuscript.
This study has been (partially) funded by grants from Instituto de Salud Carlos III (RD16/0025/0013 to JMB and RD16CIII/0002/0002 to SR) as part of the Health Research and Development Strategy, State Plan for Scientific and Technical Research and Innovation (2008–2011; 2013–2016) and co-financed by Institute of Health Carlos III, ISCIII – Sub-Directorate General for Research Assessment and Promotion and European Regional Development Fund (ERDF). The study was also funded by the Centro de Investigación Biomédica en Red (CIBER) de Enfermedades Infecciosas (CB21/13/00044, CB21/13/00020, and CB21/13/00063). MAJ-S is supported and funded by ISCIII (grant number CP17CIII/00007). NR is a Miguel Servet researcher from the ISCIII (grant number CPII19/00025).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We also acknowledge the patients in this study for their participation and the Centro de Transfusión of Comunidad de Madrid for the healthy donor blood samples provided. The authors also acknowledge the Spanish National Genotyping Center (CeGen) for providing SNP genotyping service (http://www.cegen.org). We acknowledge the Spanish HIV BioBank integrated in the Spanish AIDS Research Network (RIS) and collaborating centers for the generous gifts of clinical samples provided. The Spanish HIV BioBank is supported by Instituto de Salud Carlos III, Spanish Healt Ministry (Grant no RD06/0006/0035, RD12/0017/0037 and RD16/0025/0019) as part of the Plan Nacional R + D + I and cofinanced by ISCIII- Subdirección General de Evaluación y el Fondo Europeo de Desarrollo Regional (FEDER). The RIS Cohort (CoRIS) is funded by the Instituto de Salud Carlos III through the Red Temática de Investigación Cooperativa en SIDA (RIS C03/173, RD12/0017/0018 and RD16/0002/0006) as part of the Plan Nacional R + D + I and cofinanced by ISCIII-Subdirección General de Evaluacion y el Fondo Europeo de Desarrollo Regional (FEDER). The clinical centers and research groups that contribute to CoRIS are shown in Appendix 1.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphar.2021.773848/full#supplementary-material
Aguilar-Jimenez, W., Villegas-Ospina, S., Gonzalez, S., Zapata, W., Saulle, I., Garziano, M., et al. (2016). Precursor Forms of Vitamin D Reduce HIV-1 Infection In Vitro. J. Acquir Immune Defic Syndr. 73 (5), 497–506. doi:10.1097/QAI.0000000000001150
Ahn, J., Yu, K., Stolzenberg-Solomon, R., Simon, K. C., McCullough, M. L., Gallicchio, L., et al. (2010). Genome-wide Association Study of Circulating Vitamin D Levels. Hum. Mol. Genet. 19 (13), 2739–2745. doi:10.1093/hmg/ddq155
Alagarasu, K., Selvaraj, P., Swaminathan, S., Narendran, G., and Narayanan, P. R. (2009). 5' Regulatory and 3' Untranslated Region Polymorphisms of Vitamin D Receptor Gene in South Indian HIV and HIV-TB Patients. J. Clin. Immunol. 29 (2), 196–204. doi:10.1007/s10875-008-9234-z
Antinori, A., Coenen, T., Costagiola, D., Dedes, N., Ellefson, M., Gatell, J., et al. (2011). Late Presentation of HIV Infection: a Consensus Definition. HIV Med. 12 (1), 61–64. doi:10.1111/j.1468-1293.2010.00857.x
Azevedo, L. A., Matte, U., Silveira, T. R., Bonfanti, J. W., Bruch, J. P., and Álvares-da-Silva, M. R. (2017). Effect of Vitamin D Serum Levels and GC Gene Polymorphisms in Liver Fibrosis Due to Chronic Hepatitis C. Ann. Hepatol. 16 (5), 742–748. doi:10.5604/01.3001.0010.2748
Baker, J. V., Peng, G., Rapkin, J., Krason, D., Reilly, C., Cavert, W. P., et al. (2008). Poor Initial CD4+ Recovery with Antiretroviral Therapy Prolongs Immune Depletion and Increases Risk for AIDS and Non-AIDS Diseases. J. Acquir Immune Defic Syndr. 48 (5), 541–546. doi:10.1097/QAI.0b013e31817bebb3
Barrett, J. C., Fry, B., Maller, J., and Daly, M. J. (2005). Haploview: Analysis and Visualization of LD and Haplotype Maps. Bioinformatics 21 (2), 263–265. doi:10.1093/bioinformatics/bth457
Croxford, S., Yin, Z., Burns, F., Copas, A., Town, K., Desai, S., et al. (2018). Linkage to HIV Care Following Diagnosis in the WHO European Region: A Systematic Review and Meta-Analysis, 2006-2017. PLoS One 13 (2), e0192403. doi:10.1371/journal.pone.0192403
Darling, K. E., Hachfeld, A., Cavassini, M., Kirk, O., Furrer, H., and Wandeler, G. (2016). Late Presentation to HIV Care Despite Good Access to Health Services: Current Epidemiological Trends and How to Do Better. Swiss Med. Wkly 146, w14348. doi:10.4414/smw.2016.14348
de la Torre, M. S., Torres, C., Nieto, G., Vergara, S., Carrero, A. J., Macías, J., et al. (2008). Vitamin D Receptor Gene Haplotypes and Susceptibility to HIV-1 Infection in Injection Drug Users. J. Infect. Dis. 197 (3), 405–410. doi:10.1086/525043
Eales, L. J., Nye, K. E., Parkin, J. M., Weber, J. N., Forster, S. M., Harris, J. R., et al. (1987). Association of Different Allelic Forms of Group Specific Component with Susceptibility to and Clinical Manifestation of Human Immunodeficiency Virus Infection. Lancet 1 (8540), 999–1002. doi:10.1016/s0140-6736(87)92269-0
Falleti, E., Bitetto, D., Fabris, C., Fattovich, G., Cussigh, A., Cmet, S., et al. (2012). Vitamin D Binding Protein Gene Polymorphisms and Baseline Vitamin D Levels as Predictors of Antiviral Response in Chronic Hepatitis C. Hepatology 56 (5), 1641–1650. doi:10.1002/hep.25848
García, M., Jiménez-Sousa, M. A., Blanco, J., Restrepo, C., Pacheco, Y. M., Brochado-Kith, Ó., et al. (2019). CD4 Recovery Is Associated with Genetic Variation in IFNγ and IL19 Genes. Antivir. Res 170, 104577. doi:10.1016/j.antiviral.2019.104577
Gois, P. H. F., Ferreira, D., Olenski, S., and Seguro, A. C. (2017). Vitamin D and Infectious Diseases: Simple Bystander or Contributing Factor? Nutrients 9 (7). doi:10.3390/nu9070651
Guo, L., Du, Y., Qu, S., and Wang, J. (2016). rVarBase: an Updated Database for Regulatory Features of Human Variants. Nucleic Acids Res. 44 (D1), D888–D893. doi:10.1093/nar/gkv1107
Guzmán-Fulgencio, M., Berenguer, J., Micheloud, D., Fernández-Rodríguez, A., García-Álvarez, M., Jiménez-Sousa, M. A., et al. (2013). European Mitochondrial Haplogroups Are Associated with CD4+ T Cell Recovery in HIV-Infected Patients on Combination Antiretroviral Therapy. J. Antimicrob. Chemother. 68 (10), 2349–2357. doi:10.1093/jac/dkt206
Hartling, H. J., Ryder, L. P., Ullum, H., Ødum, N. S. D., and Nielsen, S. D. (2017). Gene Variation in IL-7 Receptor (IL-7R)α Affects IL-7R Response in CD4+ T Cells in HIV-Infected Individuals. Sci. Rep. 7, 42036. doi:10.1038/srep42036
Hartling, H. J., Thørner, L. W., Erikstrup, C., Harritshøj, L. H., Kronborg, G., Pedersen, C., et al. (2014). Polymorphism in Interleukin-7 Receptor α Gene Is Associated with Faster CD4⁺ T-Cell Recovery after Initiation of Combination Antiretroviral Therapy. AIDS 28 (12), 1739–1748. doi:10.1097/QAD.0000000000000354
Helleberg, M., Kronborg, G., Larsen, C. S., Pedersen, G., Pedersen, C., Obel, N., et al. (2013). Poor CD4 Response Despite Viral Suppression Is Associated with Increased Non-AIDS-related Mortality Among HIV Patients and Their Parents. AIDS 27 (6), 1021–1026. doi:10.1097/QAD.0b013e32835cba4c
Herrmann, M., Farrell, C. L., Pusceddu, I., Fabregat-Cabello, N., and Cavalier, E. (2017). Assessment of Vitamin D Status - a Changing Landscape. Clin. Chem. Lab. Med. 55 (1), 3–26. doi:10.1515/cclm-2016-0264
Hibler, E. A., Hu, C., Jurutka, P. W., Martinez, M. E., and Jacobs, E. T. (2012). Polymorphic Variation in the GC and CASR Genes and Associations with Vitamin D Metabolite Concentration and Metachronous Colorectal Neoplasia. Cancer Epidemiol. Biomarkers Prev. 21 (2), 368–375. doi:10.1158/1055-9965.EPI-11-0916
Jiménez-Sousa, M. Á., Jiménez, J. L., Fernández-Rodríguez, A., Bellón, J. M., Rodríguez, C., Riera, M., et al. (2019). DBP Rs16846876 and Rs12512631 Polymorphisms Are Associated with Progression to AIDS Naïve HIV-Infected Patients: a Retrospective Study. J. Biomed. Sci. 26 (1), 83. doi:10.1186/s12929-019-0577-y
Jiménez-Sousa, M. A., Jiménez, J. L., Fernández-Rodríguez, A., Brochado-Kith, O., Bellón, J. M., Gutierrez, F., et al. (2019). VDR Rs2228570 Polymorphism Is Related to Non-progression to AIDS in Antiretroviral Therapy Naïve HIV-Infected Patients. J. Clin. Med. 8 (3). doi:10.3390/jcm8030311
Jiménez-Sousa, M. Á., Martínez, I., Medrano, L. M., Fernández-Rodríguez, A., and Resino, S. (2018). Vitamin D in Human Immunodeficiency Virus Infection: Influence on Immunity and Disease. Front. Immunol. 9, 458. doi:10.3389/fimmu.2018.00458
Jiménez-Sousa, M. A., Jiménez, J. L., Bellón, J. M., Fernández-Rodríguez, A., Iribarren, J. A., López-Cortés, L. F., et al. (2020). Brief Report: CYP27B1 Rs10877012 T Allele Was Linked to Non-AIDS Progression in ART-Naïve HIV-Infected Patients: A Retrospective Study. J. Acquir Immune Defic Syndr. 85, 659–664. doi:10.1097/QAI.0000000000002485
Jolliffe, D. A., Walton, R. T., Griffiths, C. J., and Martineau, A. R. (2016). Single Nucleotide Polymorphisms in the Vitamin D Pathway Associating with Circulating Concentrations of Vitamin D Metabolites and Non-skeletal Health Outcomes: Review of Genetic Association Studies. J. Steroid Biochem. Mol. Biol. 164, 18–29. doi:10.1016/j.jsbmb.2015.12.007
Karcioglu Batur, L., and Hekim, N. (2020). The Role of DBP Gene Polymorphisms in the Prevalence of New Coronavirus Disease 2019 Infection and Mortality Rate. J. Med. Virol. 93, 1409–1413. doi:10.1002/jmv.26409
Kelley, C. F., Kitchen, C. M., Hunt, P. W., Rodriguez, B., Hecht, F. M., Kitahata, M., et al. (2009). Incomplete Peripheral CD4+ Cell Count Restoration in HIV-Infected Patients Receiving Long-Term Antiretroviral Treatment. Clin. Infect. Dis. 48 (6), 787–794. doi:10.1086/597093
Kranzer, K., Govindasamy, D., Ford, N., Johnston, V., and Lawn, S. D. (2012). Quantifying and Addressing Losses along the Continuum of Care for People Living with HIV Infection in Sub-saharan Africa: a Systematic Review. J. Int. AIDS Soc. 15 (2), 17383. doi:10.7448/IAS.15.2.17383
Laplana, M., Sánchez-de-la-Torre, M., Puig, T., Caruz, A., and Fibla, J. (2014). Vitamin-D Pathway Genes and HIV-1 Disease Progression in Injection Drug Users. Gene 545 (1), 163–169. doi:10.1016/j.gene.2014.04.035
Lu, L., Sheng, H., Li, H., Gan, W., Liu, C., Zhu, J., et al. (2012). Associations between Common Variants in GC and DHCR7/NADSYN1 and Vitamin D Concentration in Chinese Hans. Hum. Genet. 131 (3), 505–512. doi:10.1007/s00439-011-1099-1
Mansueto, P., Seidita, A., Vitale, G., Gangemi, S., Iaria, C., and Cascio, A. (2015). Vitamin D Deficiency in HIV Infection: Not Only a Bone Disorder. Biomed. Res. Int. 2015, 735615. doi:10.1155/2015/735615
Masson, J. J. R., Cherry, C. L., Murphy, N. M., Sada-Ovalle, I., Hussain, T., Palchaudhuri, R., et al. (2018). Polymorphism Rs1385129 within Glut1 Gene SLC2A1 Is Linked to Poor CD4+ T Cell Recovery in Antiretroviral-Treated HIV+ Individuals. Front. Immunol. 9, 900. doi:10.3389/fimmu.2018.00900
Medrano, L. M., Gutiérrez-Rivas, M., Blanco, J., García, M., Jiménez-Sousa, M. A., Pacheco, Y. M., et al. (2018). Mitochondrial Haplogroup H Is Related to CD4+ T Cell Recovery in HIV Infected Patients Starting Combination Antiretroviral Therapy. J. Transl Med. 16 (1), 343. doi:10.1186/s12967-018-1717-y
Mocroft, A., Lundgren, J. D., Sabin, M. L., Monforte, Ad., Brockmeyer, N., Casabona, J., et al. (2013). Risk Factors and Outcomes for Late Presentation for HIV-Positive Persons in Europe: Results from the Collaboration of Observational HIV Epidemiological Research Europe Study (COHERE). Plos Med. 10 (9), e1001510. doi:10.1371/journal.pmed.1001510
Moodley, A., Qin, M., Singh, K. K., and Spector, S. A. (2013). Vitamin D-Related Host Genetic Variants Alter HIV Disease Progression in Children. Pediatr. Infect. Dis. J. 32 (11), 1230–1236. doi:10.1097/INF.0b013e31829e4d06
Negredo, E., Massanella, M., Puig, J., Pérez-Alvarez, N., Gallego-Escuredo, J. M., Villarroya, J., et al. (2010). Nadir CD4 T Cell Count as Predictor and High CD4 T Cell Intrinsic Apoptosis as Final Mechanism of Poor CD4 T Cell Recovery in Virologically Suppressed HIV-Infected Patients: Clinical Implications. Clin. Infect. Dis. 50 (9), 1300–1308. doi:10.1086/651689
Nieto, G., Barber, Y., Rubio, M. C., Rubio, M., and Fibla, J. (2004). Association between AIDS Disease Progression Rates and the Fok-I Polymorphism of the VDR Gene in a Cohort of HIV-1 Seropositive Patients. J. Steroid Biochem. Mol. Biol. 89-90 (1-5), 199–207. doi:10.1016/j.jsbmb.2004.03.086
Nunnari, G., Fagone, P., Lazzara, F., Longo, A., Cambria, D., Di Stefano, G., et al. (2016). Vitamin D3 Inhibits TNFα-Induced Latent HIV Reactivation in J-LAT Cells. Mol. Cell Biochem 418 (1-2), 49–57. doi:10.1007/s11010-016-2732-z
Panel de expertos de Gesida y Plan Nacional sobre el Sida (2011). National Consensus Document by GESIDA/National Aids Plan on Antiretroviral Treatment in Adults Infected by the Human Immunodeficiency Virus (January 2011 Update). Enferm Infecc Microbiol. Clin. 29 (3), 209–103. doi:10.1016/j.eimc.2010.12.004
Pérez-Molina, J. A., Díaz-Menéndez, M., Plana, M. N., Zamora, J., López-Vélez, R., and Moreno, S. (2012). Very Late Initiation of HAART Impairs Treatment Response at 48 and 96 Weeks: Results from a Meta-Analysis of Randomized Clinical Trials. J. Antimicrob. Chemother. 67 (2), 312–321. doi:10.1093/jac/dkr478
Petta, S., Grimaudo, S., Marco, V. D., Scazzone, C., Macaluso, F. S., Cammà, C., et al. (2013). Association of Vitamin D Serum Levels and its Common Genetic Determinants, with Severity of Liver Fibrosis in Genotype 1 Chronic Hepatitis C Patients. J. Viral Hepat. 20 (7), 486–493. doi:10.1111/jvh.12072
Prabhu, A. V., Luu, W., Li, D., Sharpe, L. J., and Brown, A. J. (2016). DHCR7: A Vital Enzyme Switch between Cholesterol and Vitamin D Production. Prog. Lipid Res. 64, 138–151. doi:10.1016/j.plipres.2016.09.003
Purcell, S., Neale, B., Todd-Brown, K., Thomas, L., Ferreira, M. A., Bender, D., et al. (2007). PLINK: a Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 81 (3), 559–575. doi:10.1086/519795
Randolph, A. G., Yip, W. K., Falkenstein-Hagander, K., Weiss, S. T., Janssen, R., Keisling, S., et al. (2014). Vitamin D-Binding Protein Haplotype Is Associated with Hospitalization for RSV Bronchiolitis. Clin. Exp. Allergy 44 (2), 231–237. doi:10.1111/cea.12247
Restrepo, C., Gutierrez-Rivas, M., Pacheco, Y. M., García, M., Blanco, J., Medrano, L. M., et al. (2019). Genetic Variation in CCR2 and CXCL12 Genes Impacts on CD4 Restoration in Patients Initiating cART with Advanced Immunesupression. PLoS One 14 (3), e0214421. doi:10.1371/journal.pone.0214421
Saag, M. S., Benson, C. A., Gandhi, R. T., Hoy, J. F., Landovitz, R. J., Mugavero, M. J., et al. (2018). Antiretroviral Drugs for Treatment and Prevention of HIV Infection in Adults: 2018 Recommendations of the International Antiviral Society-USA Panel. JAMA 320 (4), 379–396. doi:10.1001/jama.2018.8431
Sinotte, M., Diorio, C., Bérubé, S., Pollak, M., and Brisson, J. (2009). Genetic Polymorphisms of the Vitamin D Binding Protein and Plasma Concentrations of 25-hydroxyvitamin D in Premenopausal Women. Am. J. Clin. Nutr. 89 (2), 634–640. doi:10.3945/ajcn.2008.26445
Speeckaert, M., Huang, G., Delanghe, J. R., and Taes, Y. E. (2006). Biological and Clinical Aspects of the Vitamin D Binding Protein (Gc-Globulin) and its Polymorphism. Clin. Chim. Acta 372 (1-2), 33–42. doi:10.1016/j.cca.2006.03.011
Strawbridge, R. J., Deleskog, A., McLeod, O., Folkersen, L., Kavousi, M., Gertow, K., et al. (2014). A Serum 25-hydroxyvitamin D Concentration-Associated Genetic Variant in DHCR7 Interacts with Type 2 Diabetes Status to Influence Subclinical Atherosclerosis (Measured by Carotid Intima-media Thickness). Diabetologia 57 (6), 1159–1172. doi:10.1007/s00125-014-3215-y
Suárez-García, I., Sobrino-Vegas, P., Dalmau, D., Rubio, R., Iribarren, J. A., Blanco, J. R., et al. .(2016). Clinical Outcomes of Patients Infected with HIV through Use of Injected Drugs Compared to Patients Infected through Sexual Transmission: Late Presentation, Delayed Anti-retroviral Treatment and Higher Mortality. Addiction 111 (7), 1235–1245. doi:10.1111/add.13348
Thompson, M. A., Aberg, J. A., Hoy, J. F., Telenti, A., Benson, C., Cahn, P., et al. (2012). Antiretroviral Treatment of Adult HIV Infection: 2012 Recommendations of the International Antiviral Society-USA Panel. JAMA 308 (4), 387–402. doi:10.1001/jama.2012.7961
Torres, C., Sánchez de la Torre, M., García-Moruja, C., Carrero, A. J., Trujillo, Mdel. M., Fibla, J., et al. (2010). Immunophenotype of Vitamin D Receptor Polymorphism Associated to Risk of HIV-1 Infection and Rate of Disease Progression. Curr. HIV Res. 8 (6), 487–492. doi:10.2174/157016210793499330
Wang, T. J., Zhang, F., Richards, J. B., Kestenbaum, B., van Meurs, J. B., Berry, D., et al. (2010). Common Genetic Determinants of Vitamin D Insufficiency: a Genome-wide Association Study. Lancet 376 (9736), 180–188. doi:10.1016/S0140-6736(10)60588-0
Xie, C. N., Yue, M., Huang, P., Tian, T., Fan, H. Z., Wu, M. P., et al. (2018). Vitamin D Binding Protein Polymorphisms Influence Susceptibility to Hepatitis C Virus Infection in a High-Risk Chinese Population. Gene 679, 405–411. doi:10.1016/j.gene.2018.09.021
Yang, X., Su, B., Zhang, X., Liu, Y., Wu, H., and Zhang, T. (2020). Incomplete Immune Reconstitution in HIV/AIDS Patients on Antiretroviral Therapy: Challenges of Immunological Non-responders. J. Leukoc. Biol. 107 (4), 597–612. doi:10.1002/JLB.4MR1019-189R
Keywords: HIV, antiretroviral therapy, DHCR7, DBP, SNP, immune reconstitution, CD4+ T cells
Citation: Resino S, Jiménez-Sousa MÁ, Blanco J, Pacheco YM, del Romero J, Peraire J, Virseda-Berdices A, Muñoz-Gómez MJ, Galera-Peñaranda C, García-Fraile LJ, Benito JM and Rallón N (2022) DBP rs7041 and DHCR7 rs3829251 are Linked to CD4+ Recovery in HIV Patients on Antiretroviral Therapy. Front. Pharmacol. 12:773848. doi: 10.3389/fphar.2021.773848
Received: 13 September 2021; Accepted: 09 December 2021;
Published: 18 January 2022.
Edited by:
Chonlaphat Sukasem, Mahidol University, ThailandReviewed by:
Umamaheswaran Gurusamy, University of California San Francisco, United StatesCopyright © 2022 Resino, Jiménez-Sousa, Blanco, Pacheco, del Romero, Peraire, Virseda-Berdices, Muñoz-Gómez, Galera-Peñaranda, García-Fraile, Benito and Rallón. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Salvador Resino, c3Jlc2lub0Bpc2NpaWkuZXM=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.