 Dezhi Pan
Dezhi Pan Xue Gong
Xue Gong Xiaoqin Wang1*
Xiaoqin Wang1* Minhui Li
Minhui Li
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Pharmacol. , 22 January 2021
Sec. Cardiovascular and Smooth Muscle Pharmacology
Volume 11 - 2020 | https://doi.org/10.3389/fphar.2020.594050
This article is part of the Research Topic Angiogenesis and Nutraceuticals View all 5 articles
Angiogenesis refers to the formation of new blood vessels from the endothelial cells of existing arteries, veins, and capillaries. Angiogenesis is involved in a variety of physiological and pathological processes, such as the formation of malignant and development of atherosclerosis and other diseases. In recent years, many studies have shown that the active components of food have a certain regulatory effect on angiogenesis and negligible clinical limitations. With the increasing attention being paid to medicine and food homology, exploring the effect of active food components on angiogenesis is of great significance. In this review, we discuss the source, composition, pharmacological activity, and mechanism of action of certain active components of medicinal foods in detail. These could help prevent angiogenesis-related complications or provide a basis for healthier dietary habits. This review can provide a theoretical basis for the research and development of highly efficient anti-angiogenic drugs with low toxicity.
Angiogenesis is a sign of several physiological and pathological states and is the basis of many diseases, such as malignant tumors, cancers, atherosclerosis, and cerebrovascular diseases (Yan and Zhao, 2020). The mechanism underlying angiogenesis is complex and involves several factors, such as transforming growth factor-β, platelet-derived endothelial growth factor, cyclooxygenase-2 (COX-2), hypoxia-inducible factor-1, vascular endothelial growth factor (VEGF), and VEGF receptor (VEGFR). Among them, the VEGF, which is a homodimeric glycoprotein encoded by a single gene that can the movement, proliferation, and division of vascular endothelial cells, as well as increase microvascular permeability, is the main angiogenic factor (Yan and Zhao, 2020). The active ingredients in food regulate angiogenesis mainly by affecting VEGF/VEGFR signal transduction (Figure 1A), and can also play an anti-tumor angiogenesis effect by affecting VEGF/VEGFR (Figure 1B).
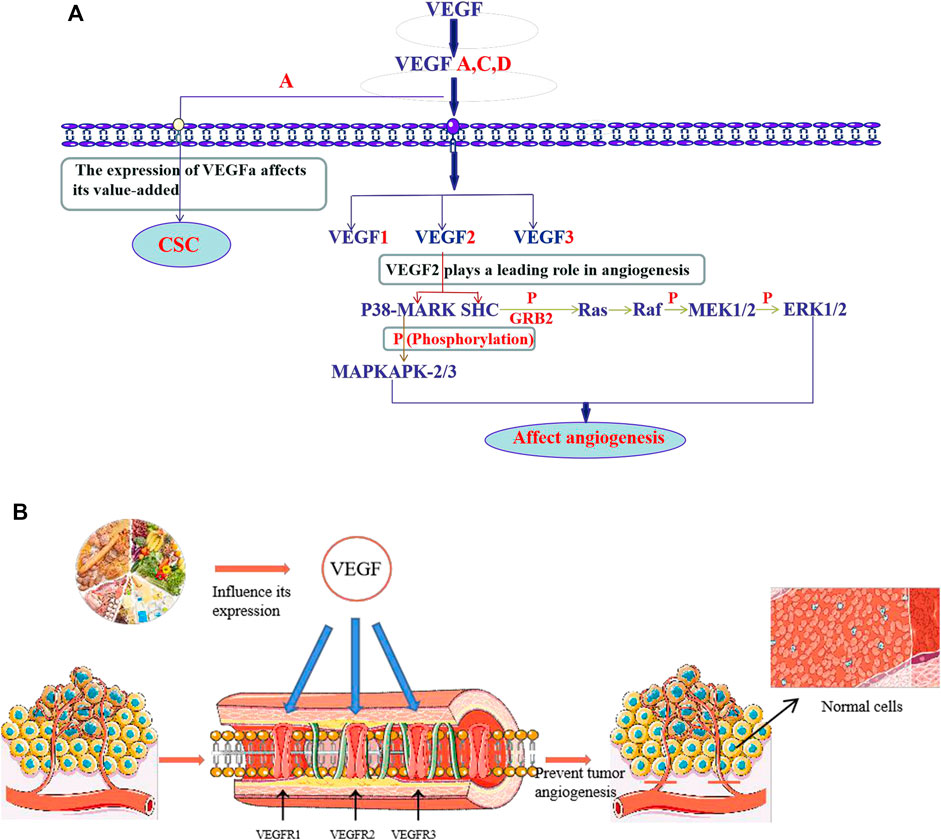
FIGURE 1. Structure of some flavonoids regulating angiogenesis in medicinal food.
In the past 20 years, research has shown that tumour angiogenesis plays an important role in tumour growth and that anti-angiogenesis is fundamental in inhibiting tumour growth, invasion, and metastasis (Qin et al., 2017). Therefore, anti-angiogenesis is a good starting point to treat tumors. With the discovery of angiogenesis inhibitors and their accompanying clinical limitations, modern molecular medicine has developed a new field of study in which the anti-angiogenic effects and anti-tumour effects of traditional Chinese medicine are examined and developed (Bagchi et al., 2004). At the same time, it has also been found that drug and food homologous food promote angiogenesis, such as cerebral functional ischemia, through endogenous repair and regeneration is not enough to help the brain recover from cerebral ischemia or brain injury caused by cerebral ischemia, promoting angiogenesis can better solve this problem, and food tonic can also treat cardiovascular and cerebrovascular diseases. Therefore, the medicine-food homology food promote angiogenesis in the treatment of cardiovascular and cerebrovascular diseases and tumor diseases has broad prospects (Yang et al., 2016; Vaeyens et al., 2020). And with the passage of time, people have found the active ingredients in food that can help regulate angiogenesis; and nutritional management of diseases may be easier, safer and more acceptable to patients. Therefore, the research on the role of nutrition in the regulation of angiogenesis has become the main research focus of modern molecular medicine.
Modern clinical trials have shown that the existing angiogenic regulators have certain clinical limitations, such as short therapeutic effects and adverse reactions, which have led to a gradual decrease in the use of anti-angiogenic drugs (Wang et al., 2017; Nikhil et al., 2020). The medical community is committed to finding a better and healthier way to treat angiogenic diseases, therefore, their research direction has gradually changed and now encompasses studies on traditional Chinese medicine and the medicinal properties of food. There are long-standing historical records on the use of food as medicine. The “medicine and food homology” concept is mentioned in the Yellow Emperor’s Classic of Internal Medicine and the Suwen: “fasting food is food, and medicine replaces medicine,” which embodies the theory of medicine and food homology (Shan et al., 2015; Gong et al., 2019; Adelman, 2018). Food can also be used as medicine; when researchers started believing that medicinal foods exert a regulatory effect on angiogenesis, they studied them intensively.
The aim of this study was to provide a comprehensive review of the regulation of angiogenesis by medicinal foods and their active components. To this end, we collated, analysed, and summarised the relevant recent research. This review can be used as a theoretical basis for future research on the regulation of angiogenesis.
In this section, we analysed six chemical components of medicinal foods that possess angiogenic activity—flavonoids, terpenes, alkaloids, polyphenols, polysaccharides, and saponins.
Flavonoids are natural compounds and secondary plant metabolites with a 2-phenylchromone structure (Li et al., 2016). They have a wide range of pharmacological properties (anti-inflammatory, antibacterial, and anti-tumor), while they are also beneficial for the cardiovascular and central nervous systems. Some studies have shown that flavonoids can regulate angiogenesis, including anti-tumour angiogenesis, as well as promote it. The anti-tumor angiogenesis mechanism entails the down-regulation of matrix metalloproteinases (MMP) (endogenous proteolytic enzyme), VEGF, COX-2, and angiotensin-2 (Huang and Wang, 2019). Figure 1B shows the angiogenic regulatory factors and their anti-tumor mechanisms. The flavonoids from some medicinal plants have been found to regulate angiogenesis. For example, Wang et al. (2019) used zebrafish embryo and rabbit corneal neovascularization models to study the inhibitory effect of isoliquiritigenin on angiogenesis. In particular, they studied whether the triple inhibition of COX-2, microsomal prostaglandin E synthase-1, and cytochrome P450 4A would inhibit tumor angiogenesis through the competing endogenous RNA effect. The results confirmed the anti-angiogenic effect of isoliquiritigenin, and determined that the effect was the most effective at EC50 concentration of 5.9 μmol L −1 (50% concentration for maximum effect) (Wang et al., 2019a).
In another study, the anti angiogenesis effect and mechanism of Puerarin glycosides (PGs) were detected by scratch test, migration test, lumen formation test and cell cycle arrest test. Human umbilical vein endothelial cells (HUVECs) were treated with different concentrations of PGs (100 μg ml−1, 200 μg ml−1 and 400 μg ml−1), and MDA-MB-231 cells were treated with PGs at the same time. Tubulin is an important part of cytoskeleton, and its disordered polymerization hinders the progress of cell cycle. The expression of α - tubulin mRNA was detected by RT-PCR-western blot with α - tubulin, β - actin, cyclin A1 and CDK2 as the main antibodies. The results showed that PGs had anti angiogenic activity and cell cycle blocking ability, and PGs could induce Sphase arrest of MDA-MB-231 cells, inhibit cell proliferation, and the expression level of CDK2 also decreased significantly with the increase of PGs concentration The antiangiogenesis activity of PGA may be related to the ability of cell cycle arrest and the signal pathway of promoting microtubule polymerization. Therefore, PGs have potential antitumor activity (Wu and Jiang, 2010; Li et al., 2018).
The effects of the total flavones of Abelmoschus manihot at different concentrations on angiogenesis were observed using the chick embryo chorioallantoic membrane (CAM) model and gelatin sponges as carriers. The results showed that 5 μg ml−1, 10 μg ml−1, and 20 μg ml−1 of total flavones of A. manihot increased the vascular network and CAM count significantly. This effect was the most obvious at the concentration of 10 μg ml−1. Additionally, the total flavonoids of A. manihot can protect the heart and brain from ischaemic injury and promote angiogenesis (Pan and Jiang, 2010).
In a study on plant-based diets and plant compounds, the inhibitor effect of 6-methoxy-equol (6-ME) extracted from soybean on tumor angiogenesis was assessed. The HUVECs were either left untreated or treated with 6-ME at different concentrations (1–50 μmol L−1) in vitro. Quantitative RT-PCR (qRT-PCR) was performed to study the effect of different 6-ME concentrations (0 μmol L−1, μmol·L−1, and 10 μmol L−1) on VEGF expression. The total DNA concentration after isolation was measured. The results showed that 6-ME had the best inhibitory effect at 5 μmol L−1. The latest research on the effects of mitogen-activated protein kinase 1/2 and extracellular signal-regulated kinase 1/2 has shown that 6-ME can inhibit tumor angiogenesis by targeting the phosphorylation of mitogen-activated protein kinase 1/2 and its downstream substrate extracellular signal-regulated kinase ½. It thus inhibits the proliferation of ECS induced by VEGF and fibroblast growth factor (FGF) 2 (Sofia et al., 2012).
These studies indicate that the flavonoids in medicinal plants, which play an important role in the regulation of angiogenesis, and the vascular regulation mechanism of flavonoids is mainly affecting the formation of blood vessels in HUVECs and. Some flavonoids have natural hormone like activities. Moreover, compared with other anti-angiogenic drugs, flavonoids are more beneficial to the human body and have no harmful effects. The flavonoids that have an angiogenic regulation ability, as well as their sources, are summarised in Table 1, and the chemical structure of some flavonoids is shown in Figure 1.

TABLE 1. Flavonoids with angiogenic regulation.
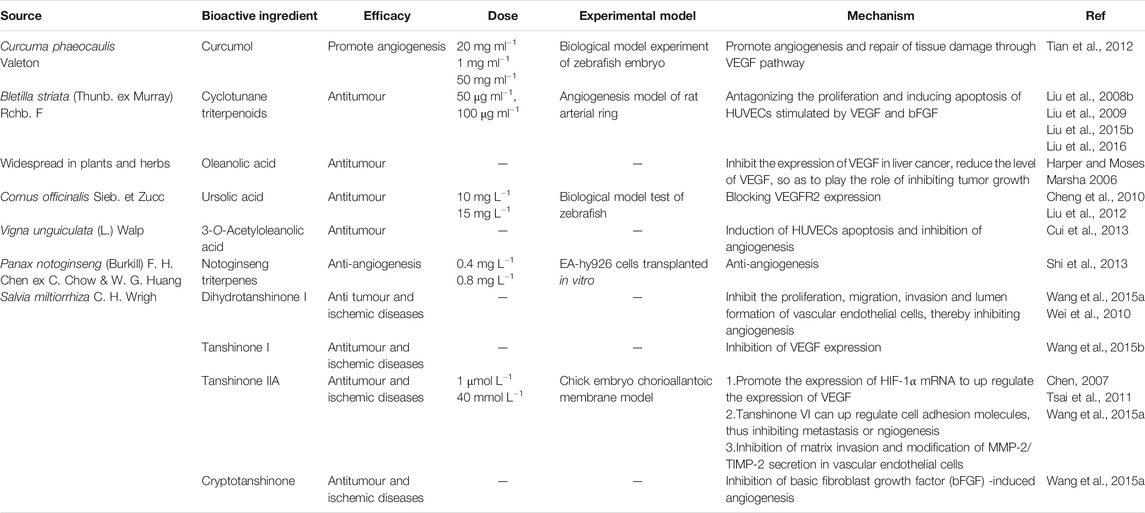
TABLE 2. Terpenoids with angiogenesis regulation.
Terpenoids are natural hydrocarbons, which are abundant in nature and can be linked by isoprene or isopentane units in various ways. All kinds of terpenoids have been proved to be effective chemical raw materials and have significant disease prevention and treatment effects. Especially, they show good antitumor activities. They have potential to be used as lead compounds to develop efficient and safe new antitumor drugs. In addition, studies have shown that some terpenoids can regulate angiogenesis and have a good effect on the treatment of cardiovascular diseases. Therefore, further study on the angiogenesis regulation of terpenoids not only contributes to the development of new anti-tumor drugs, but also has great advantages in the treatment of angiogenesis related diseases. Therefore, terpenoids have great application potential and broad development prospects due to their special structure and function as well as extensive medical uses (Zhang et al., 2018). For this reason, some terpenoids with angiogenic effect (Table 3) are sorted out and the structural diagram of some terpenoids is shown (Figure 2).
FIGURE 2. Structure of some terpenoids regulating angiogenesis in medicinal food.
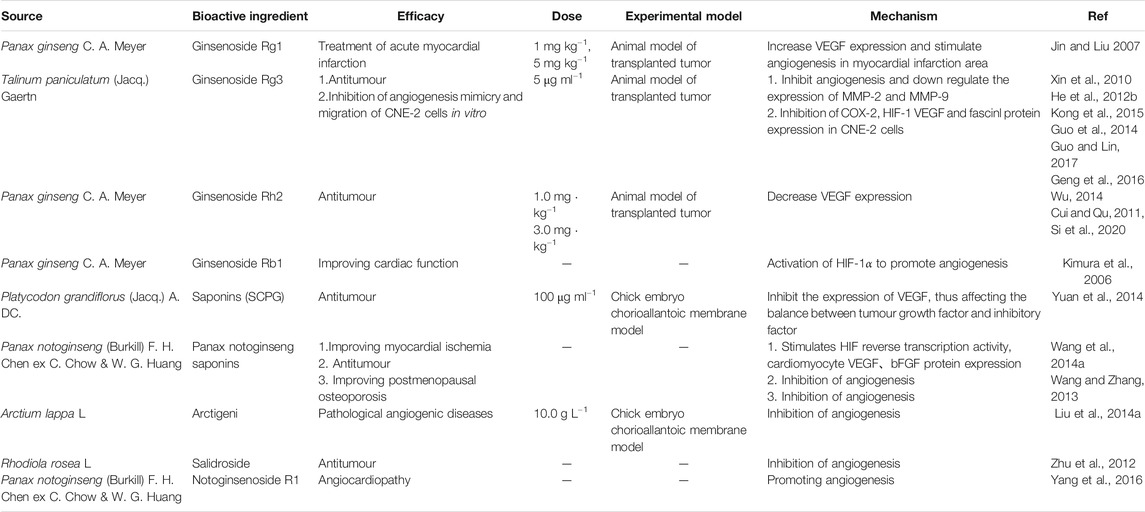
TABLE 3. Saponins with angiogenesis regulation.
The zebrafish biological model has been used to observe the effects of curcumol on the vascular growth of embryonic bodies, the vascular regeneration of adult fish after cutting their tails, and the tissue regeneration of larvae after cutting their tails. VEGF and VEGFR2 gene expressions were detected by relative quantitative fluorescence PCR. For this, zebrafish embryos were treated with different curcumol concentrations (1 mg L−1, 5 mg L−1, 10 mg L−1, 20 mg L−1, and 50 mg L−1). The results showed that 20 mg L−1 promoted the vascular endothelial growth of zebrafish embryo internodes, whereas 50 mg L−1 promoted VEGFR2 growth in zebrafish embryos. Moreover, 1 mg L−1 curcumol had the strongest effect on tissue regeneration. Therefore, curcumol could promote angiogenesis by promoting the expression of VEGF and VEGFR2 (Tian et al., 2012).
Additionally, some studies have examined, by fluorescence microscopy, the effects of different doses of ursolic acid (5 mg L−1, 10 mg L−1, and 20 mg L−1) on the formation of intersegmental vessels on the backs of AB and transgenic fluorescent zebrafish (VEGFR2:GFP) that were used as experimental animals. The results showed that at ursolic acid doses of 5 mg L−1 and 10 mg L−1, intersegmental vessel proliferation induced by transplanted cancer was inhibited significantly, whereas the inhibition of angiogenesis and VEGFR2 was also possible. It has been suggested that ursolic acid significantly inhibits the physiological characteristics of zebrafish and the angiogenesis caused by transplanted tumors, which is related to the inhibition of VEGFR2 (Cheng et al., 2010).
Furthermore, the anti-angiogenic effect of different tanshinone IIA (Tan ⅡA) doses (1 μmol L−1, 5 μmol L−1, and 10 μmol L−1) on chicken embryos and human umbilical vein endothelial cells (HUVECs) was studied using the CAM model. The mRNA expression of MMP-2, MMP-3, MMP-9, and MMP-14, tissue inhibitor of metalloproteinase, and reversion-inducing cysteine-rich protein was detected by RT-PCR. The results showed that Tan IIA inhibits angiogenesis on chicken embryos and HUVECs, and increasing Tan IIA concentration decreases the inhibitory effect. The results of western blotting and ELISA further confirmed that after Tan ⅡA treatment, MMP-2 decreased in a dose-dependent manner, whereas the level of tissue inhibitor of metalloproteinase secretion increased. These results suggest that Tan ⅡA has anti-angiogenic effects both in vivo and in vitro. Additionally, its mechanism of action is related to its inhibitory effect on MMP-2, whereas it has the opposite regulatory effect on the tissue inhibitor of metalloproteinase secretion, thereby decreasing the MMP-2 activity in vascular endothelial cells (Tsai et al., 2011).
Saponins are glycosides composed of triterpenoids or spiral sterane compounds and are widely distributed in nature, including in monocotyledons and dicotyledons (Liu and Henkel, 2002). Saponins have important physiological and pharmacological activities. In recent years, scholars have been actively looking for angiogenesis regulators from natural drug sources. As natural active ingredients in food, saponins have been tested in animal models and have been found to be safe and regulate angiogenesis. The saponins that regulate angiogenesis are shown in Table 3, and the structure of few saponins is shown in Figure 3.
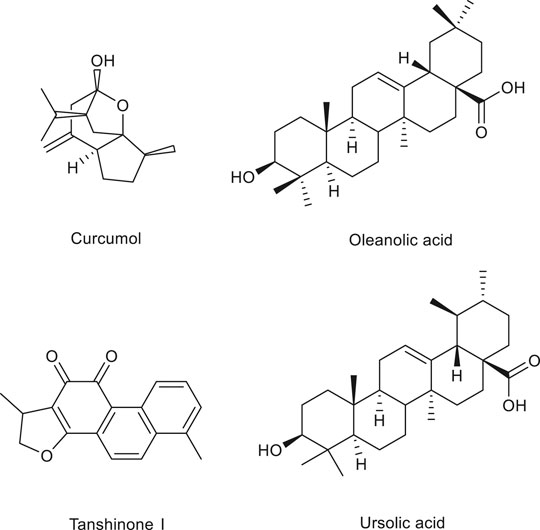
FIGURE 3. Structure of some saponins regulating angiogenesis in medicinal food.
Ginsenoside Rg1 and RB1 have been reported to regulate cardiac function and promote angiogenesis, while ginsenoside Rh2 has antitumor and antiangiogenic effects (Jin and Liu, 2007).
Some studies have divided the rats into sham operation group, acute myocardial infarction control group, ginsenoside Rgl low-dose treatment group (1 mg kg−1) and high-dose treatment group (5 mg kg−1) to study the angiogenesis promoting effect and mechanism of Ginsenoside Rg1. Myocardial enzymes, infarct size and microvessel density were measured at different time points. The expression of VEGF-mRNA was detected by RT-PCR. The results showed that the microvessel density and VEGF-mRNA expression in the sham operation group were lower than those in the operation group at different time points. In addition, myocardial enzyme activity and infarct size in the treatment group were significantly lower than those in the control group, and the degree of angiogenesis in the infarct area continued to increase steadily, with significant difference compared with the control group. In conclusion, severe myocardial ischemia can stimulate the production of a large amount of VEGF, and the difference in each experimental group can show the mechanism of ginsenoside Rgl in promoting angiogenesis, which is related to the increase of VEGF-mRNA expression in myocardial infarction area (Jin and Liu, 2007).
Additionally, the anti-tumor mechanism and effect of ginsenoside Rh2 on Lewis lung carcinoma in mice was studied using a solid tumor model of the carcinoma. The anti-tumor effect of ginsenoside Rh2 and VEGF expression in the tumor were observed by immunohistochemistry. The experimental group was administered different Rh2 doses (0.3 mg kg−1, 1.0 mg kg−1, and 3.0 mg kg−1), and a cyclophosphamide group (20 mg kg−1) was established. The results showed that the tumor weight and the microvessel density in the ginsenoside Rh2 group was significantly lower than that in the control group. VEGF protein was mainly expressed in the new capillaries of tumor cells and some tumor cell membranes and plasma that were stained. The results demonstrate that the positive rate of VEGF protein expression decreases with the increase in ginsenoside Rh2 concentration. Therefore, the inhibitory effect of ginsenoside Rh2 on tumor angiogenesis can be induced by inhibiting the expression of VEGF protein, thereby inhibiting the release of VEGF and tumor angiogenesis (Cui and Qu, 2011).
Polysaccharides, which consist of 10 or more single sugar molecules polymerised by a glucosidic bond, have relatively high molecular weight and generally consist of hundreds or even tens of thousands of single sugar molecules. Polysaccharides are important biological macromolecules, which exist widely in animals, plants, and microorganisms. The biological activities of polysaccharides have recently attracted considerable attention in biochemical and medical research because of their immunomodulatory effects (Lu et al., 2000). Polysaccharides have a wide range of biological activities and low toxicity as well as have a good potential for the development of clinical drugs. Therefore, they can be potentially used for the treatment of angiogenic diseases. In this section, we collated studies that have reported the regulation of angiogenesis by polysaccharides (Table 4).
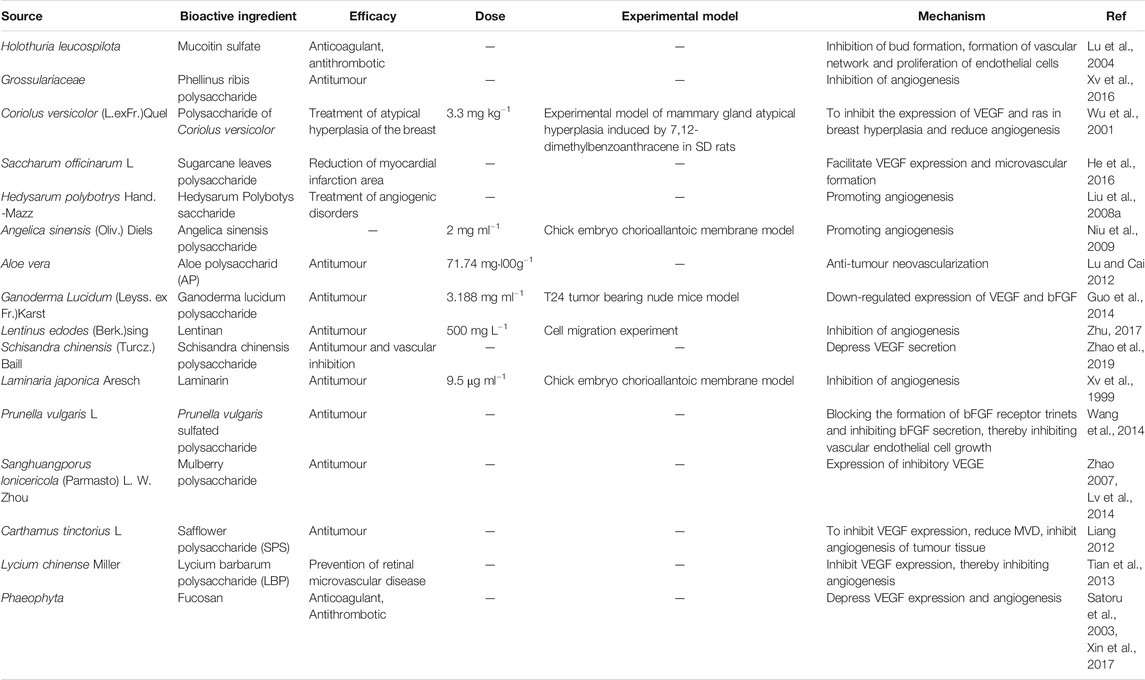
TABLE 4. Polysaccharide compounds with angiogenesis regulation.
In one study, the effect of five different lentinan concentrations (31.25 mg L−1, 62.5 mg L−1, 125 mg L−1, 250 mg L−1, and 500 mg L−1) and thalidomide (200 mg L−1) as the positive control, on HUVEC proliferation was examined using the MTT method. Additionally, the transwell cell migration test, FN adhesion test, and tuber formation assay were used to detect the effect of lentinan on cell migration, cell adhesion, and angiogenesis in vitro, respectively. The results showed that lentinan inhibits cell migration, cell adhesion, and angiogenesis in a dose-dependent manner, with the most significant inhibitory effect produced by the 500 mg L−1 concentration. The anti-tumour effect of lentinan is related to its inhibitory effect on HUVEC proliferation, migration, adhesion, and angiogenesis in vitro (Zhu, 2017).
There are studies on the inhibition of G. lucidum polysaccharide on tumor angiogenesis and its mechanism. The effect of G. lucidum polysaccharide combined with cisplatin on the proliferation of human bladder cancer cell line T24 in vitro was determined by MTS method, and the T24 tumor bearing nude mice model was established to observe the combined treatment effect of G. lucidum polysaccharide and cisplatin. The microvessel density (MVD) and the expression levels of vascular endothelial growth factor (VEGF) and basic fibroblast growth factor (bFGF) were detected by immunohistochemistry Real time PCR and Western blotting were used to detect the expression of VEGF and bFGF. The results showed that G. lucidum polysaccharide could effectively inhibit the proliferation of T24 cells in vitro, and had synergistic effect with cisplatin. Moreover, G. lucidum polysaccharide (3.188 mg L−1) could significantly enhance the inhibitory effect of cisplatin on tumor growth of nude mice, inhibit the angiogenesis of tumor tissue and the expression of VEGF and bFGF. Therefore, G. lucidum polysaccharide can inhibit the growth and angiogenesis of T24 tumor bearing nude mice, and its mechanism may be related to the down-regulation of VEGF and bFGF expression (Guo et al., 2014).
Alkaloids are a class of nitrogen-containing organic compounds found in nature and derived mainly from plants. Most alkaloids have complex ring structures on which the nitrogen atoms are bound. Additionally, most alkaloids are basic, have significant biological activities, and are important pharmaceutical bioactive components in medicinal food (Jin et al., 2004). It has been confirmed that plant alkaloids have a wide range of anti-tumour and anti-angiogenesis effects and can be obtained from a wide range of sources; therefore, they can be potentially used to treat various clinical conditions (Feng and Tang, 2014).
In one study, the authors isolated, purified and identified the active components from the fruit of Evodia rutaecarpa. Through the zebrafish experimental model method, nine of them were detected to have angiogenesis inhibitory effect (Yin et al., 2016).
The antitumor effect of 5,2,4′- trihydroxy-6,7,5' - trimethoxyflavone (TTF1) was studied in this paper. The mechanism of TTF1 action included inhibition of tumor angiogenesis and induction of tumor cell apoptosis. In order to further study the molecular mechanism of its inhibitory effect on tumor angiogenesis, cam method was used to determine the angiogenesis inhibitory effect of TTF1. In order to detect whether the angiogenesis inhibitory effect of TTF1 is related to the expression of VEGF, KDR, bFGF, COX-2 and HIF-1α, the protein levels of these factors in hepg-2-induced mouse tumor were measured The expression of VEGF, KDR, bFGF, COX-2 and HIF-1α was decreased by TTF1 at different concentrations (5 μmol kg−1, 10 μmol kg−1 and 20 μmol kg−1), respectively White matter level to achieve anti angiogenesis effect (Liu et al., 2011a). Table 5 shows the recent research progress on the anti-angiogenic mechanisms of food alkaloids. Figure 4 lists the structural formulas of alkaloids in two common medicinal foods.
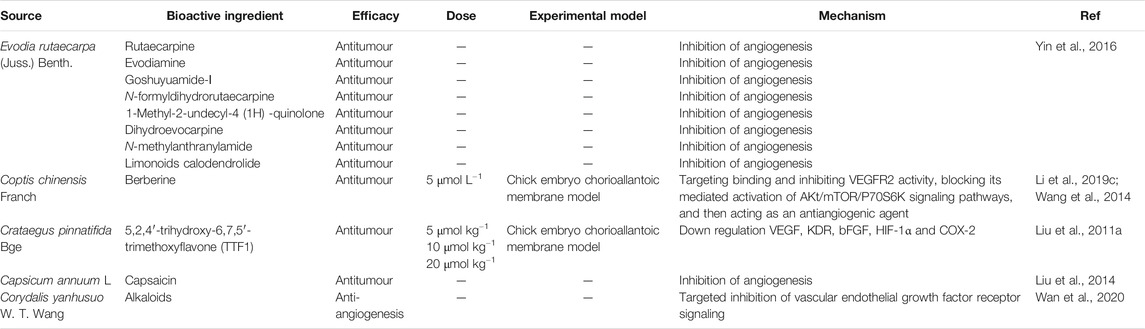
TABLE 5. Alkaloids with angiogenic regulatory action.

FIGURE 4. Structure of some alkaloids regulating angiogenesis in medicinal food.
Polyphenols (tannins) are important natural products that are found widely in plants. Polyphenols have a complex structure and active chemical properties and their structure comprises many homologous compounds; therefore, research on these compounds has progressed relatively slowly (Song et al., 2000). However, in the past 30 years with the development of polyphenol chemistry, the chemical structure and properties of polyphenols have been revealed in detail. Plant polyphenols are recognised as safe anti-tumour agents that act via several mechanisms, including affecting tumour angiogenesis. Moreover, polyphenols have antioxidant, antimutagenic, anticancer, and anti-inflammatory properties, thus ensuring the stability of the human genome and preventing occurrence of diseases. Therefore, polyphenols present in some foods can be potentially used as therapeutic agents (Anupama et al., 2002; Wang et al., 2005).
The effect of emodin on human liver cancer has been studied by examining the angiogenesis of HepG2 cells. To this end, the effects of HIF1A and VEGF on HepG2 cells, as well as the anti-hepatoma effects of rhein, were examined using an in vivo chicken CAM model. The experiment included randomly divided groups: a negative control group (normal saline), emodin low-dose (10 μmol L−1) and high-dose groups (20 μmol L−1), and a positive control group (0.15 mg L−1; dexamethasone). The inhibitory effect of emodin on CAM angiogenesis was observed using in vitro culture methods. For this, HepG2 cells were treated with CoCl2 to simulate chemical hypoxia. Hypoxia untreated (negative control group [normal saline]), hypoxia emodin low-dose (10 μmol L−1), hypoxia emodin high-dose (20 μmol L−1), and hypoxia positive control groups (10 μmol L−1; 5-fluorouracil) were established. After 24 hours of treatment, the expression of HIF1A and VEGF was detected by immunocytochemistry, and the expression of HIF1A and VEGF mRNA was detected by qPCR. The CAM results showed that the number of neovascularization in the experimental group was significantly reduced, especially in the low-dose group (10 μmol L−1), the HIF1A and VEGF positive cells in the low-dose, high-dose and positive control groups were significantly reduced; the HIF1A and VEGF positive cells in the low-dose group (10 μmol L−1) were significantly reduced, and the levels of HIF1A and VEGF mRNA in the experimental group were lower than those in the negative control group. Therefore, it can be concluded that emodin may inhibit the angiogenesis of HepG2 cells by inhibiting the expression of HIF1A mRNA, thereby also reducing VEGF mRNA level (Gao et al., 2016a).
In another study, the regulation of chlorogenic acid on cox2-mmp signaling pathway in transgenic zebrafish was studied. Therefore, 100 fli1a EGFP transgenic zebrafish embryos were randomly divided into blank group, positive control group and chlorogenic acid high, medium and low dose groups. 20 positive controls in each group were treated with PTK787 5 pg ml−1, chlorogenic acid high, medium and low dose groups were treated with chlorogenic acid 200 μg ml−1, 100 μg ml−1 and 50 μg ml−1, respectively. Twenty six hours after treatment, vascular phenotypes were observed and photographed with stereomicroscope (bright field) and stereofluorescence microscope. The expression levels of COX-2 mRNA, MMP-9 mRNA and MMP-2 mRNA in zebrafish embryos were detected by real-time PCR. The results showed that compared with the blank group, the vascular inhibition was obvious in chlorogenic acid group. The levels of COX-2 mRNA, MMP-9 mRNA and MMP-2 mRNA in chlorogenic acid and ptk797 groups were significantly lower than those in the blank control group. The levels of MMP-9 mRNA and MMP-2 mRNA in high-dose chlorogenic acid group were significantly lower than those in medium dose chlorogenic acid group, while those in medium dose chlorogenic acid group were significantly lower than those in low-dose chlorogenic acid group. It is concluded that chlorogenic acid can inhibit the angiogenesis of transgenic zebrafish embryos. The mechanism of action is related to the down regulation of cox-2-mmp signal pathway and the inhibition of MMP-9 and MMP-2 gene expression (Cai et al., 2019).
The results of the aforementioned studies demonstrate that the polyphenols found in some medicinal plants have a certain regulatory effect on angiogenesis. Table 6 shows current progress on the regulation of angiogenesis by polyphenols. Figure 5 lists the structural formulas of common polyphenols present in several medicinal foods.
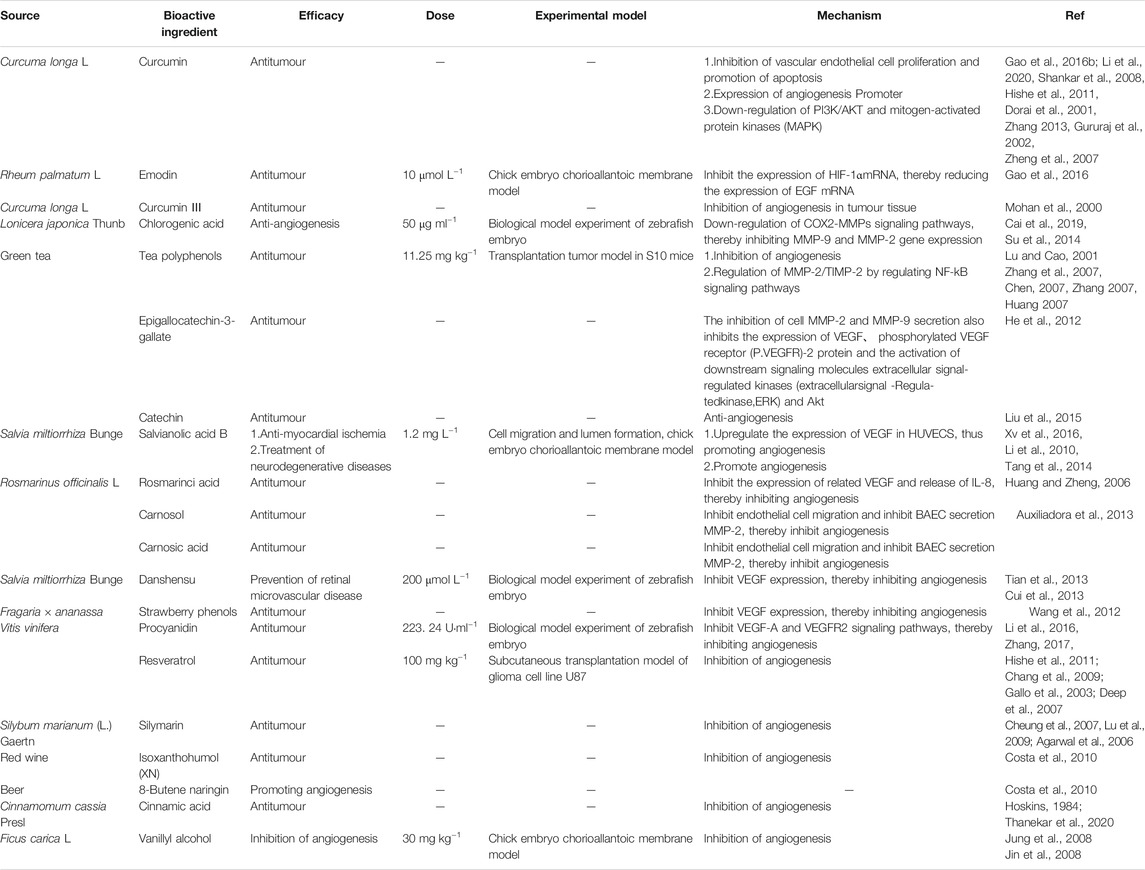
TABLE 6. Polyphenols with angiogenesis regulation.
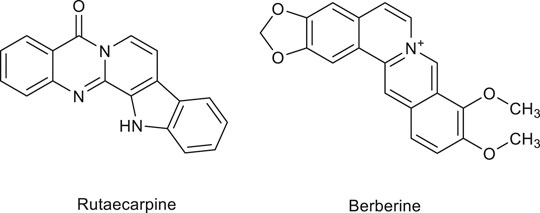
FIGURE 5. Structure of some polyphenols regulating angiogenesis in medicinal food.
FIGURE 6. Other active compounds regulating angiogenesis in medicinal foods.
In addition to the aforementioned compounds, studies have shown that other active components extracted from food can regulate angiogenesis as well, this is consistent with the clearly defined anti-angiogenetic activity of various compounds. Therefore, although these active components have not been clearly classified yet, they can still be used in the research and development of drugs related to angiogenesis regulation (Liu et al., 2011a).
One example is the effect of black garlic on angiogenesis. In one study (Sheng et al., 2018), the authors studied the effect of black garlic on angiogenesis by establishing transgenic zebrafish in vivo and HUVEC in vitro. The transgenic zebrafish Flk-1: GFP labeled with green fluorescent protein was used as an in vivo evaluation model. The experimental group, control group, blank control group and vascular model group (0.2 μg ml−1 PTK787) were established. Positive control group (30 μg ml−1 Danhong injection and 0.2 μg ml−1 PTK787) and black garlic extract treatment group (concentration gradient of 0.2 μg ml−1, 0.8 μg ml−1, 3.2 μg ml−1 and 3.5 μg ml−1 PTK787) were cultured under normal conditions. The results showed that black garlic extract could significantly improve the pathological changes of embryo internodes and lower intestinal veins induced by PTK787, and increase the number of embryonic internodes vessels and small intestinal veins in a dose-dependent manner. When the concentration of black garlic extract increased to 3.2 μg ml−1, PTK787 inhibited the number of embryonic internodes vessels and inferior intestinal veins to normal. In addition, black garlic extract could enhance the activity of HUVEC and promote its growth in a dose-dependent manner. The above results showed that black garlic could promote angiogenesis by promoting HUVEC mechanism (Sheng et al., 2018).
In another study, the effect of Dalbergia odorifera extract on angiogenesis was studied (Fan et al., 2018). Using transgenic zebrafish model, the effects and mechanism of the extract of Dalbergia odorifera B3 were evaluated by observing the subintestinal angiogenesis and the damage of internode vessels in transgenic zebrafish. In this experiment, 50 g of the extract of Dalbergia odorifera B3 was extracted by reflux with 500 ml 75% ethanol, 50 g of Dalbergia odorifera decoction pieces were placed in Soxhlet extractor for 1 hour, and then the drug residue was boiled in boiling water for 1 h, concentrated to 100 ml, and then the extract was dissolved in 900 ml 95% ethanol. Next, the control group and experimental group were established. The extracts were treated with 3 μg ml−1, 10 μg ml−1 and 30 μg ml−1. The results showed that the extract of Dalbergia odorifera B3 had a certain promoting effect on the capillary germination of transgenic zebrafish, and the effect of 30 μg ml−1 was the best. These results indicate that the extract of Dalbergia odorifera B3 can promote angiogenesis and repair vascular injury induced by VEGFR kinase inhibitor II, and its mechanism is related to the promotion of VEGFR mRNA expression (Table 7).
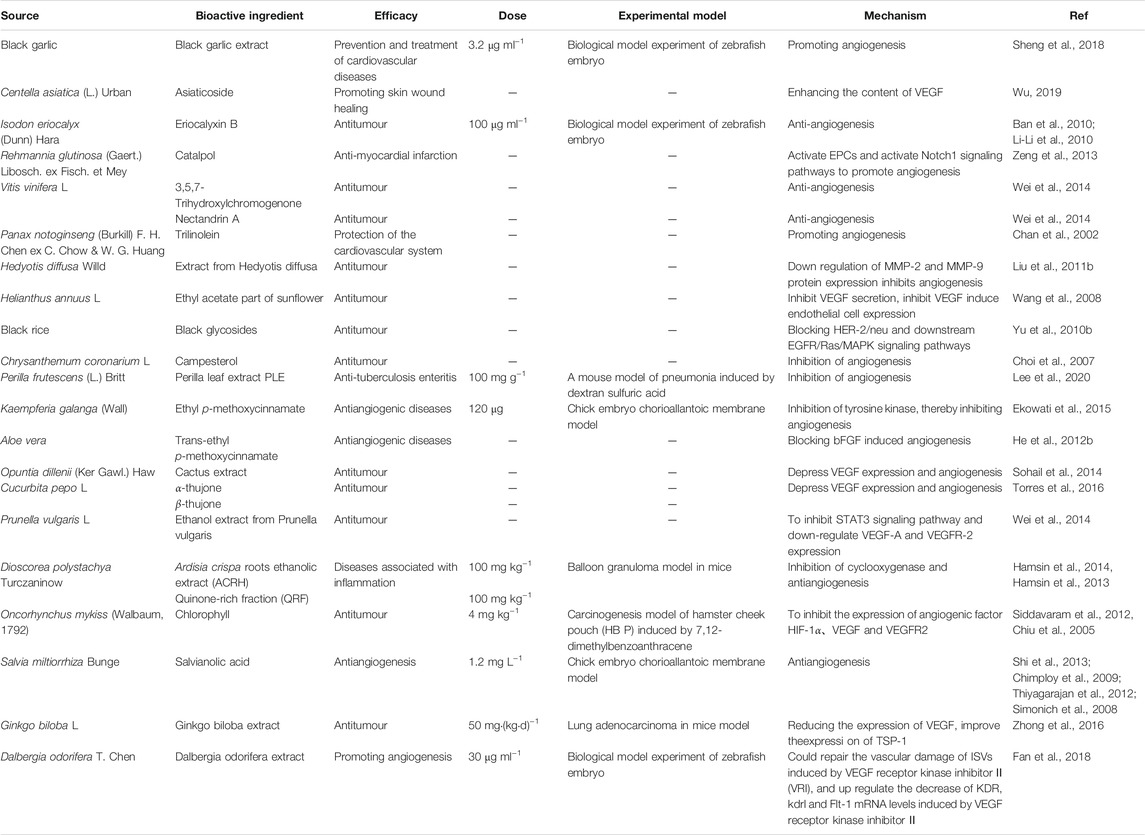
TABLE 7. Other components with vascular regulation.
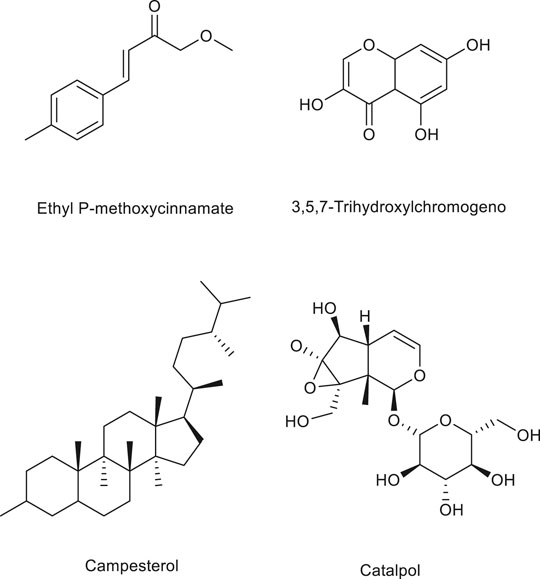
FIGURE 7.
Studies have shown that angiogenesis is associated with many diseases (Qu et al., 2018a). In terms of angiogenesis regulation, the active components of medicinal foods mainly inhibit and promote angiogenesis by altering the corresponding signal transduction pathways. The VEGF and VEGFR are the two main regulatory factors the signal transduction of which is affected, thereby promoting regulatory effects. The VEGF, also known as vascular permeability factor, appears to be one of the key growth factors participating in physiological and pathological angiogenesis. This factor is present in many human tumors and may contribute to vascular hyperpermeability and enhanced angiogenesis (Dvorak et al., 1991; Kim et al., 1993; Teleanu et al., 2019). Blocking the VEGF expression by antibodies or antisense strategies has been shown to inhibit the growth of some tumors; overexpression enhances tumorigenesis. The mechanism of angiogenesis regulation by food and its active components is analysed in the following sections.
The anti-angiogenic effect of the active components of medicinal foods is mainly achieved by inhibiting the transduction pathway of the corresponding angiogenesis signal. And mainly focuses on tumor angiogenesis and can be used to treat a small number of other angiogenesis-induced diseases.
As a study showed that tumor growth is dependent on vasculature (Folkman, 1971), the treatment of tumors has mainly focused on regulating tumor vascular growth and angiogenesis. Inhibiting the growth and migration of vascular endothelial cells, as well as regulating angiogenic factors, is one of the basic anti-angiogenic mechanisms of food active components. The specific mechanisms are directly inhibiting the proliferation and migration of vascular endothelial cells, inhibiting extracellular matrix metalloproteinase (MMP) activity, inhibiting the signal transduction of tumor angiogenic factors, and promoting the expression of tumor angiogenesis inhibitors (Lv et al., 2014).
Many studies have shown that imposing a restriction on total food intake or energy inhibits tumorigenesis (Incio et al., 2016). For example, chlorogenic acid from Lonicera japonica extract can inhibit the expression of MMP-9 and MMP-2 by down-regulating the COX-2-MMP signaling pathway (Cai et al., 2019); curcumin from Curcuma longa extract can inhibit tumor angiogenesis by inhibiting the proliferation of vascular endothelial cells and the expression of angiogenesis promoting factors (Gao et al., 2016b); rosmarinic acid extracted from rosemary can inhibit angiogenesis and achieve an anti-tumour effect by inhibiting VEGF expression and IL-8 release from related cells (Huang and Zheng, 2006).
In recent years, studies have shown that VEGF can promote endothelial cell division and proliferation, increase vascular permeability, and regulate thrombosis. Therefore, VEGF is closely related to the occurrence and development of certain cardiovascular diseases (Xie and Li, 2019; Qu et al., 2018b).
The active components extracted from Kaempferia galanga, ethyl p-methoxycinnamate and ethyl trans-p-methoxycinnamate, are used to treat angiogenic diseases by inhibiting the tyrosine kinase inhibitor of angiogenesis and blocking bFGF-induced angiogenesis (Jung et al., 2010; He et al., 2012a).
Cardiovascular diseases are mainly treated through the promotion of angiogenesis. For example, ginsenoside Rb1 promotes angiogenesis and improves heart function by promoting HIF1A activity (You et al., 2019; Kimura et al., 2006). Catalpol extracted from Rehmannia glutinosa can activate endothelial progenitor cells and the Notch1 signaling pathway, promote angiogenesis, and prevent myocardial infarction (Zeng et al., 2013).
The active food component-induced angiogenic regulation is not only a means to treat tumors and cardiovascular diseases, but also has therapeutic effects on other angiogenesis-related diseases, as it promotes the healing of skin wounds and eye diseases caused by blood vessels.
Beta-sitosterol, an active component of aloe vera, has been found to promote wound healing by promoting angiogenesis (Moon et al., 1999). Asiaticoside promotes skin wound healing by increasing the VEGF content (Wu, 2014). Fucoidan can not only inhibit the mitosis and chemotaxis of VEGF165 in HUVEC by blocking the binding of VEGF to its cell surface receptors but also has an anti-coagulation effect by inhibiting the expression of VEGF (Satoru et al., 2003; Xin et al., 2017).
The aforementioned studies indicate that a medicinal diet often has a good regulatory effect on angiogenesis. They can regulate by inhibiting and promoting the transmission of different angiogenesis-related signals and can be used in the treatment of angiogenic diseases such as cardiovascular diseases and tumors. Cardiovascular and cerebrovascular diseases and tumors are two conditions that have attracted much attention since the 20th century. In the past, studies on these mainly focused on vasodilators and antineoplastic drugs. However, clinical trials have shown that both drug types have clinical limitations, including drug resistance, unclear treatment window, and lack of effective biomarkers. In addition, long-term human use of these drugs can lead to the development of drug resistance and dependence (Fu et al., 2020). Compared with traditional medicine, medicinal food is safer and more easily accepted by people, and the natural chemical products present in it are safer and more reliable. Therefore, since the first discovery that medicinal food and its active ingredients have a good regulatory effect on angiogenesis, the role of food in regulating angiogenesis in disease treatment has attracted extensive attention (He, 2019).
With the advancement of research, the angiogenic regulatory effect of active ingredients in medicinal foods has been gradually revealed. Therefore, it is incredibly important to understand the pathogenesis and regulatory mechanism of angiogenesis to use active ingredients in foods for mediating their angiogenic effect. This will also contribute to a solid foundation for the research and development of therapeutic drugs for vascular diseases based on the characteristics of medicinal foods and their active ingredients.
Some experimental studies have shown that medicinal foods and their active ingredients can regulate angiogenesis and be directly used to treat angiogenic diseases. However, they also have broad application prospects in the field of research, development, and synthesis of angiogenesis-regulating drugs or health products. Although it is safer to treat vascular diseases with medicinal foods and their active ingredients than with traditional medicines, the currently available knowledge has many limitations. First, the current experimental research is only limited to the laboratory construction of animal experimental models or basic pharmacological activity experiments, and there is no regulation on the follow-up clinical trials and other studies. Second, the research on active ingredients of medicinal food is obviously insufficient. Many studies have shown that active ingredients in foods can regulate angiogenesis, but no studies have specified a dose. Most studies only indicate a certain dose range, and the measurement is not uniform, even though the correct dosage is extremely important. Third, the regulatory mechanism of medicinal foods and their active ingredients on angiogenesis is not clear as such foods constitute many ingredients. As previous studies have shown that VEGF is highly concentrated in tumor cells, the inhibition of VEGF-related receptors may be an effective treatment for such diseases (Ge et al., 2019). However, owing to the many regulatory factors, it is not simple to determine the main mechanism of action of medicinal foods and their active ingredients. Finally, an important research problem that needs to be solved is the lack of quality control in these studies owing to the influence of many factors on the active ingredients of medicinal foods. Therefore, the follow-up study of angiogenesis-related factors and their receptors is crucial, as it will provide more reliable targets for the future research and development of new drugs. Moreover, it is particularly important for later clinical trials, which can provide more powerful evidence for later drug development.
Conceptualization, ML, XW, and DP. Writing—originaldraftpreparation, DP and XG. Writing—review and editing, DP. Supervision, ML and XW. All authors have read and agreed to the published version of the manuscript.
This work was supported by National Natural Science Foundation of China (No. 81760686), 2018 Chinese medicine public health service subsidy special “the fourth survey on Chinese materia medica resource”, University level scientific research and innovation team of Inner Mongolia Medical University (YKD2017KJCXTD013).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
DP would like to thank ML for giving me the opportunity to write this article. DP would like to thank ML and XW for their support and encouragement during my writing. At the same time, DP would like to thank XG and other students for their modification suggestions during this period. Thank you very much.
Adelman, J., and Haushofer, L.. (2018). Introduction: food as medicine, medicine as food. Hist Med. Allied Sci. 73 (2), 127–134. doi:10.1093/jhmas/jry010
Agarwal, R., Agarwal, C., Ichikawa, H., Singh, R. P., and Aggarwal, B. B. (2006). Anticancer potential of silymarin: from bench to bed side. Anticancer Res. 26 (6B), 4457–4498. doi:10.1016/j.jembe.2004.06.008
Agnieszka, K., Aleksandra, B. P., Maria, P., Honorata, K., Agnieszka, S., Elżbieta, W., et al. (2020). Microvesicles from human immortalized cell lines of endothelial progenitor cells and mesenchymal stem/stromal cells of adipose tissue origin as carriers of bioactive factors facilitating angiogenesis. Stem Cell. Int. 2020, 1289380. doi:10.1155/2020/1289380
Anupama, E. G., Madesh, B., Deepak, A. V., Dieter, M., and Bharathi, P. S. (2002). Molecular mechanisms of antiangiogenic effect of curcumin. Biochem. Biophys. Res. Comm. 297 (4), 934–942. doi:10.1016/s0006-291x(02)02306-9
Auxiliadora, L. L., Melissa, G. C., Miguel, Á. M., and Ana, R. Q. (2013). Anti-angiogenic properties of carnosol and carnosic acid, two major dietary compounds from rosemary. 52 (1), 85–95. doi:10.1007/s00394-011-0289-x
Bagchi, D., Sen, C. K., Bagchi, M., and Atalay, M. (2004). Anti-angiogenic, antioxidant, and anti-carcinogenic properties of a novel anthocyanin-rich berry extract formula. Biochemistry 69 (1), 75–80. doi:10.1023/B:BIRY.0000016355.19999.93
Ban, L. L., Zhang, H., Luo, M., Yang, B. B., Han, H. S., Pan, M., et al. (2010). Study on the inhibitory effect of calyx B on human normal and hepatoma angiogenesis. Chinese Pharm. 21 (15), 1351–1353.
Bayliss, A. L., Sundararaman, A., Granet, C., and Mellor, H. (2020). Raftlin is recruited by neuropilin-1 to the activated VEGFR2 complex to control proangiogenic signaling. Angiogenesis 23 (3), 371–383. doi:10.1007/s10456-020-09715-z10.1007/s10456-020-09715-z
Bian, W., Chen, F., Bai, L., Zhang, P., and Qin, W. (2008). Dihydrotanshinone I inhibits angiogenesis both in vitro and in vivo. Acta Biochim. Sinica 40 (1), 1–6. doi:10.1111/j.1745-7270.2008.00370.x
Cai, Y., Zheng, Q., He, X. Y., Qian, L., and Yao, D. J. (2019). The regulatory effect of chlorogenic acid on the Cox 2-mmps signaling pathway in flila EGFP transgenic zebrafish. J. Mod. Chinese Western Med. 28 (36), 3995–3998. doi:10.3969/j.issn.1008-8849.2019.36.002
Cao, H. J., Yang, Y., Wei, B., Li, Q. M., He, Y. J., Li, Y. Y., et al. (2020). Decylubiquinone suppresses breast cancer growth and metastasis by inhibiting angiogenesis via the ROS/p53/BAI1 signaling pathway. Angiogenesis 23 (1), 325–338. doi:10.1007/s10456-020-09707-z10.1007/s10456-020-09707-z
Chan, P., Thomas, G. N., and Tomlinson, B. (2002). Protective effects of trilinolein extracted from Panax notoginseng against cardiovascular disease. Acta Pharmacol. Sin. 23 (12), 1157–1162. doi:10.1021/ar010155r
Chang, H. B., Zhang, X., Fei, Z., and Zhen, H. N. (2009). Effect of resveratrol on the growth and angiogenesis of U87 cell transplanted tumour in nude mice. Chinese J. Neuromed. (10), 986–989. doi:10.3760/cma.j.issn.1671-8925.2009.10.004
Chen, X. Y. (2007). “Study on the mechanism of anti angiogenesis of transplanted mouse breast cancer by tea polyphenols [C].,”in International congress of Chinese medicine oncology (China Association of Chinese Medicine: China Society of Chinese Medicine), 21–32.
Chen, X. H., Liang, J. J., Liu, X. S., and Xv, Y. (2017). Effect of Ginsenoside Rg1 on myocardial vascular regeneration after myocardial infarction in rats. J. Cardiovas. Rehabil. Med. 26 (3), 245–250. doi:10.3969/j.issn.1008-0074.2017.03.005
Cheng, C., he, Z. X., and Wang, H. (2010). Ginsenoside RH- 2 research progress of antitumour effect. Traffic Med. 24 (01), 23–25. doi:10.3969/j.issn.1006-2440.2010.01.008
Cheung, C. W., Taylor, P. J., KJrkpatrick, C. M., Vesey, D. A., Gobe, G. C., Winterford, C., et al. (2007). Therapeuticvalue of orallyadministeredsilibi- ninin renal cellcarcinoma:manipulation of insulin- likegrowth fac-tor binding pmtein-3levels. BJUIra 100 (2), 438–444. doi:10.1111/j.1464-410x.2007.07012.x
Chimploy, K., Diaz, G. D., Li, Q., Carter, O., Dashwood, W. M., Mathews, C. K., et al. (2009). E2F4 and ribonucleotide reductase mediate S phase arrest in colon cancer cells treated with chlorophyllin. Int. J. Canc. 125, 2086–2094 doi:10.1002/ijc.24559
Chiu, L. C., Kong, C. K., and Ooi, V. E. (2005). The chlorophyllin-induced cell cycle arrest and apoptosis in human breast cancer MCF-7 cells is associated with ERK deactivation and cyclin D1 depletion. Int. Mol. Med. 16, 735–740. doi:10.1089/hum.2005.16.1227
Choi, J. M., Lee, E. O., Lee, H. J., Kim, K. H., Ahn, K. S., Shim, B. S., et al. (2007). Identification of campesterol from Chrysanthemum coronarium L. and its antiangiogenic activities. Phytother Res. 21 (10), 954–959. doi:10.1002/ptr.2189
Costa, R., Negrao, R., Duarte, D., and Taveira Gomes, T. (2010). Angiogenesis and inflammation signaling are targets of beer polyphenols on vascular cells. Cell Biochem. 111, 1270–1279. doi:10.1002/jcb.22850
Cui, Y. R., and Qu, F. (2011). Antitumour effect of ginsenoside RH 2 on Lewis lung cancer mice. Oncology 1 (5), 422–425. doi:CNKI:SUN:LIYX.0.2011-05-010
Cui, E. J., Jeon, H. B., Park, J. H., Baek, N. I., Kim, J. Y., Hong, S. G., et al. (2013). 3-O-Acetyloleanolic acid exhibits anti-angiogenic effects and induces apoptosis in human umbilical vein endothelial cells. Biotechnol. Lett. 35 (11), 1807–1815. doi:10.1007/s10529-013-1266-7
Deep, G., and Agarwal, R.. (2007). Chemopreventive efficacy of silymarin in skin and prostate cancer. Integr. Canc. Ther. 6 (2), 130–l45. doi:10.1177/1534735407301441
Ding, X. W., Ding, X. L., Zheng, S., and Yang, H. J. (2004). CHL prevent colon neoplasms in mice and its selective inhibition on COX-2. Ai Zheng 23, 1409–1413. doi:10.3969/j.issn.1000-467X.2004.z1.011
Dorai, T., Cao, Y. C., Dorai, B., Buttyan, R., and Katz, A. E. (2001). Therapeutic potential of curcumin in human prostate cancer. III. Curcumin inhibits proliferation, induces apoptosis, and inhibits angiogenesis of LNCaP prostate cancer cells in vivo. Prostate 47 (4), 293–303. doi:10.1002/pros.1074
Du, D. J., Shi, X. F., Ran, C. Q., Qi, J. N., Wen, Z. J., Xie, D. C., et al. (1994). Anti-tumour effect of Pueraria lobata. Pharmacol. Clin. Med. (2), 16–20. doi:10.13412/j.cnki.zyyl.1994.02.006
Dvorak, H. F., Sioussat, T. M., Brown, L. F., Berse, B., Nagy, J. A., Sotrel, A., et al. (1991). Distribution of vascular permeability factor (vascular endothelial growth factor) in tumors: concentration in tumour blood vessels. Exp. Med. 174, 1275–1278. doi:10.1084/jem.174.5.1275
Ekowati, J., Hardjono, S., and Hamid, I. S. (2015). Ethyl p-methoxycinnamate from Kaempferia galanga inhibits angiogenesis through tyrosine kinase. 34, 43. doi:10.18051/UnivMed.2015.v34.43-51
Fan, Z. M., Wang, Y. H., Zhou, X., Yang, A. L., Lin, J. X., Fan, H., et al. (2018). Study on the effect of Jiangxiang extract B3 on promoting angiogenesis of zebrafish model. Pharmacol. Clin. Trad. Chinese Med. 34 (04), 66–69. doi:10.13412/j.cnki.zyyl.2018.04.015
Fang, J., Zhou, Q., Ling-Zhi, L., Xia, C., Hu, X., Shi, X., et al. (2007). Apigenin inhibits tumor angiogenesis through decreasing HIF‐1alpha and VEGF expression. Carcinogenesis 858–864. doi:10.1093/carcin/bgl205
Feng, X. W., and Tang, X. D. (2014). Antitumour mechanism of plant alkaloids. Int. J. Oncol. 41 (4), 254–258. doi:10.3760/cma.j.issn.1673-422X.2014.04.005
Feng, L. L., Zhong, J. Y., and Liu, B. X. (2014). The effect of grape procyanidins on the growth and angiogenesis of H22 hepatoma transplanted tumour. Oncol. Trans. Med. 15 (2), 737–741. doi:10.7314/apjcp.2014.15.2.737
Folkman, J. (1971). Tumor angiogenesis: therapeutic implication. N. Engl. J. Med. 285. doi:10.1056/NEJM197111182852108
Fotsis, T., Pepper, M., Adlercreutz, H., Hase, T., Montesano, R., and Schweigerer, L. (1995). Genistein, a dietary ingested isoflavonoid, inhibits cell proliferation and in vitro angiogenesis. J. Nutr. 125 (Suppl. 3), 790S–797S. doi:10.1093/jn/125.3_Suppl.790S
Fu, W. W., Ouyang, Y. L., Huang, C. Y., Huang, J. W., and Lu, S. (2020). Research progress of autophagy treatment based on mTOR regulation [J/OL]. J. Hainan Med. Coll., 1–7. doi:10.13210/j.cnki.jhmu.20200617.002
Gallo, D., Giacomelli, S., Ferlini, C., Raspaglio, G., Apollonio, P., Prislei, S., et al. (2003). Antitumouractivity of the silybin phosphatidyl cholinecomplex, IdB1016, against human ovarian cancer. EurJ Can. 39 (16), 2403–2410. doi:10.1016/s0959-8049(03)00624-5
Gao, J. X., Liu, S., Ren, F. R., and Qiang, L. V. (2016a). Study on the inhibition of emodin on angiogenesis of HepG2 cells and its mechanism. Chin. J. Cancer Prev. Treat. 23 (3), 164–170.
Gao, W., He, Y. J., and Liang, F. M. (2016b). Research progress on molecular mechanism of curcumin against tumour angiogenesis. Int. J. Ophthalmol. 16 (3), 466–468. doi:10.3980/j.issn.1672-5123.2016.3.1510.3390/s16081230
Gao, W., Xu, H., Liang, Y. L., Li, Y. Y., and Liu, Y. X. (2019). Root extract of Prunella vulgaris inhibits in vitro and in vivo carcinogenesis in MCF-5 human breast carcinoma via suppression of angiogenesis, induction of apoptosis, cell cycle arrest and modulation of PI3K/AKT signalling pathway. J. BUON. 24 (2), 549554.
Ge, J. R., Wang, W., and He, Y. F. (2019). Current situation and challenges of antiangiogenic therapy. Oncology 9 (6), 845–849. 864.
Geng, L., Fan, J., Gao, Q. L., Yu, J., and Hua, B. J. (2016a). Preliminary study for the roles and mechanisms of 20 (R) - ginsenoside Rg3 and peg-plga-rg3 nanoparticles in the Lewis lung cancer mice. J. Peking Univ. (Heal. Sci.) 48 (3), 496–501. doi:10.3969/j.issn.1671-167X.2016.03.021
Geng, L., Fan, J., Gao, Q. L., Yu, J., and Hua, B. J. (2016b). The effect and mechanism of Ginsenoside Rg3 and peg-plga-rg3 nanoparticles on Lewis lung cancer mice. J. Peking Univ. (Heal. Sci.) 48 (3), 496–501.
Gong, X., Ji, M. Y., Xu, J. P., Zhang, C. H., and Li, M. H. (2019). Hypoglycemic effects of bioactive ingredients from medicine food homology and medicinal health food species used in China. Crit. Rev. Food Sci. Nutr. 60, 2303–2326. doi:10.1080/10408398.2019.1634517
Guo, J. Q., and Lin, S. Z. (2014). The effect of Ginsenoside Rg3 on angiogenesis mimicry of pancreatic cancer. J. Hepato-Biliary-Pancreatic Surg. 26 (4), 308–311.+ 322.
Guo, J. Q., and Lin, S. Z. (2017). Inhibitory effect of Ginsenoside Rg3 on angiogenesis mimicry of pancreatic cancer cell line sw-1990 in vitro. Chin. J. Trad. Chin. Med. 32 (3), 1310–1312. doi:CNKI:SUN:BXYY.0.2017-03-119
Guo, L., Ma, Q., Chen, W., Fan, W. X., Zhang, J., and Dai, B.. (2019). Synthesis and biological evaluation of novel N9-heterobivalent β-carbolines as angiogenesis inhibitors. J. Enzym. Inhib. Med. Chem. 34 (1), 375–387. doi:10.1080/14756366.2018.1497619
Guo, P. R., Sheng, Y. W., Liu, B., Wang, C., Liu, Y. D., Fu, D. W., et al. (2014). The effect of Ganoderma lucidum polysaccharide on the inhibition of cisplatin on tumour growth and angiogenesis in nude mice bearing bladder cancer T24 cells. PLA Med. J. 39 (6), 470–474. doi:10.11855/j.issn.0577-7402.2014.06.09
Gururaj, A. E., Belakavadi, M., Venkatesh, D. A., Marmé, D., and Salimath, B. P. (2002). Molecular mechanisms of antiangiogenic effect of curcumin. Biochem. Biophys. Res. Commun. 297 (4), 934–942. doi:10.1016/s0006-291x(02)02306-9
Hamsin, D. E. Z, Abdul Hamid, R., Saiful Yazan, L., Taib, C., N, M., Yeong, L., et al. (2014). Ardisia crispa roots inhibit cyclooxygenase and suppress angiogenesis. BMC Compl. Altern. Med. 14, 102. doi:10.1186/1472-6882-14-102
Hamsin, D. E. Z. A., Hamid, R. A., Yazan, L. S., et al. (2013). The hexane fraction of Ardisia crispaThunb. A. DC. roots inhibits inflammation-induced angiogenesis. BMC Compl. Altern. Med. 13 (1), 5. doi:10.1186/1472-6882-13-5
Han, B., Cheng, D. M., Yu, X. P., Peng, X. L., and Jia, H. (2011). Genistein inhibits vascular endothelial cell activation and protein kinase K activity induced by vascular endothelial growth factor. J. Chengdu Med. Coll. 6 (3), 192–195. doi:CNKI:SUN:CDYU.0.2011-03-005
Harper, J., and Moses, M. A. (2006). Molecular regulation of tumour angiogenesis: mechanisms and therapeutic implications. EXS (96), 223–268. doi:10.1007/3-7643-7378-4_10
He, L. J. (2011). Inhibition of tumour angiogenesis by two Chinese herbal monomers [D]. Shanghai, China: East China Normal University. doi:10.7666/d.y1904077
He, L. (2019). Pharmacological analysis of cardiovascular disease and traditional Chinese medicine treatment. Electron. J. Integr. Chin. Western Med. Ardiovas. Disease 7 (22), 25. doi:10.16282/j.cnki.cn11-9336/r.2019.22.018
He, L., and Tang, X. D. (2012). Antitumour mechanism of epigallocatechin gallate. Int. J. Oncol. 39 (4), 255–259. doi:10.3760/cma.j.issn.1673-422X.2012.04.005
He, F., Sun, X. L., Su, Y. L., Sun, D., and Kang, T. G. (2014). Study on the antitumour effect of arctiin. Pharmacol. Clin. Trad. Chin. Med. 30 (4), 19–23. doi:10.13412/j.cnki.zyyl.2014.04.007
He, F. J., Wang, J., Niu, J. Z., Bai, J. W., Yang, M. J., Zhao, L. Y., et al. (2003). Experimental study on inhibition of tumour growth and angiogenesis in nude mice by soybean isoflavone. Chin. Pharmacol. Bull. (1), 73–76. doi:10.3321/j.issn:1001-1978.2003.01.021
He, J. B., Liao, H. Z., Yang, K., Yi, G. Z., and Xie, M. F. (2012b). Inhibitory effect of Ginsenoside Rg3 on growth and metastasis of Lewis lung cancer in mice and its relationship with tumour related macrophages. Can. Prev. Treat. Res. 39 (12), 1411–1415. doi:10.3971/j.issn.t000-8578.2012.12.002415
He, T., Hu, S., Hou, X. T., and Qin, C. (2016). Effects of sugarcane leaf polysaccharide on electrocardiogram and microvascular formation in myocardial infarction rats. J. Guangxi Med. Univ. 33 (2), 229–231. doi:10.16190/j.cnki.45-1211/r.2016.02.011
He, Y. L., Zhao, C. B. E. M. Y., Yin, H. T., and Gai, K. (2013). Research progress of anti-tumour mechanism of Ginsenoside Rg3. Prog. Mod. Biomed. 13 (17), 3397–3400. doi:10.13241/j.cnki.pmb.2013.17.006
He, Z. H., Yue, G. L., Lau, B. S., Wei, G., Paul, P., and Hay, B. (2012a). Antiangiogenic effects and mechanisms of trans-ethyl p-methoxycinnamate from Kaempferia galanga L. J. Agric. Food Chem. 60 (45), 11309–11317. doi:10.1021/jf304169j
Hishe, H., El-Azab, M., Moustafa, Y., and El-Awady, E. -S. (2011). Anti-angiogenic effect of resveratrol or curcumin in Ehrlich ascites carcinoma-bearing mice. Eur. J. Pharmacol. 652 (1), 7–14. doi:10.1016/j.ejphar.2010.11.008
Hoskins, J. A. (1984). The occurrence, metabolism and toxicity of cinnamic acid and related compounds. J. Appl. Toxicol. 4 (6), 283–292. doi:10.1002/jat.2550040602
Hua, H., Zhang, M., and Tao, Y. H. (2017). Inhibitory effect of galangin on tumour angiogenesis in vitro and in vivo. Internat. J. Pharm. Res. 44 (10), 970–974. doi:10.13220/j.cnki.jipr.2017.10.012
Huang, D. (2007). Inhibitory effect of tea polyphenols and tea pigments on retinal neovascularization induced by alloxan in diabetic mice [D]. Zhejiang University
Huang, H. Z. (2015). Study on the antioxidative and anticancer functions and mechanism of Bayberry phenols [D]. Zhejiang University.
Huang, S. S., and Zheng, R. (2006). Rosmarinic acid inhibits angiogenesis and its mechanism of action in vitro. Cancer Lett. 239 (2), 271–280. doi:10.1016/j.canlet.2005.08.025
Huang, Y. H., Wang, X., and Xin, X. Y. (2007). Inhibitory effect of genistein on human ovarian cancer transplanted tumour. Canc. Clin. China 21, 1244–1247. doi:10.3969/j.issn.1000-8179.2007.21.013
Jin, H. Z., Lee, J. H., Lee, D., Lee, H. S., Hong, Y. S., Kim, Y. S., et al. (2004). Quinolone alkaloids with inhibitory activity against nuclear factor of activated T cells from the fruits of Evodia rutaecarpa. Biol. Pharm. Bull. 27, 926–928.
Incio, J., Tam, J., Rahbari, N. N., Suboj, P., McManus, D. T., Chin, S. M., et al. (2016). PlGF/VEGFR-1 signaling promotes macrophage polarization and accelerated tumour progression in obesity. Clin. Canc. Res. 22 (12), 2993–3004. doi:10.1158/1078-0432.CCR-15-1839
Jin, H. Z., Lee, J. H., Lee, D., Lee, H. S., Hong, Y. S., Kim, Y. S., et al. (2004). Quinolone alkaloids with inhibitory activity against nuclear factor of activated T cells from the fruits of Evodia rutaecarpa. Biol. Pharm. Bull. 27, 926–928.
Jin, Y., and Liu, B. N. N. (2007). Effect of Ginsenoside Rg1 on angiogenesis in rats with acute myocardial infarction. J. China Med. Univ. (5), 517–519. doi:10.3969/j.issn.0258-4646.2007.05.008
Jin, J. F., Hortense, M., Robert, H., Van D, H. H., Albert, J. H., Dick, B. J., et al. (2008). Covalent flavinylation of vanillyl‐alcohol oxidase is an autocatalytic process. FEBS J. 275 (20), 5191–51200. doi:10.1111/j.1742-4658.2008.06649.x10.1111/j.1742-4658.2008.06649.x
Jin, Y., Liu, G. N., and Zhu, X. Y. (2006). Effect of Ginsenoside Rg1 on VEGF and HIF-1 α after acute myocardial infarction. J. PLA Med. 31 (11), 1079–1081. doi:10.3321/j.issn:0577-7402.2006.11.017
Jung, H.-J., Song, Y. S., Kim, K., Lim, C.-J., and Park, E.-H. (2010). Assessment of the anti-angiogenic, anti-inflammatory and antinociceptive properties of ethyl vanillin. Arch. Pharm. Res. doi:10.1007/s12272-010-0217-2
Jung, H.-J., Song, Y. S., Lim, C.-J., and Park, E.-H. (2008). Anti-angiogenic, anti-inflammatory and anti-nociceptive activities of vanillyl alcohol. Arch Pharm. Res. 31 (10), 1275–1279. doi:10.1007/s12272-001-2106-110.1007/s12272-001-2106-1
Kang, X. M., Wang, H., Wang, Li., Tang, D. B., and Zhang, Q. Y. (2015). The effect of Daidzein on the growth and angiogenesis of breast cancer. Carcinog., Distort., Mutat. 27 (1), 16–20. doi:10.3969/j.issn.1004-616x.2015.01.004
Kim, K., Li, B., Winer, J., Armanini, M., Gillett, N., Phillips, H., et al. (1993). Inhibition of vascular endothelial growth factor induced angiogenesis suppresses tumour growth in vivo. Nature 362, 841–844. doi:10.1038/362841a0
Kimura, Y., Sumiyoshi, M., Kawahira, K., and Sakanaka, M. (2006). Effects of ginseng saponins isolated from Red Ginseng roots on burn wound healing in mice. Br. J. Pharmacol. 148 (6), 860–870. doi:10.1038/sj.bjp.0706794
Kondo, K., Muramatsu, M., Okamoto, Y., Jin, D., Takai, S., Tanigawa, N., et al. (2006). Expression of chymase-positive cells in gastric cancer and its correlation with the angiogenesis. J. Surg. Oncol. 93, 36–42. doi:10.1002/jso.20394
Kong, X., Shang, J. L., Zhou, Y. F., Wang, H. G., and Gu, D. S. (2015). The effect of Ginsenoside Rg3 on angiogenesis mimicry and migration of CNE-2 cells in human nasopharyngeal carcinoma. Guangdong Med. J. 36 (9), 1314–1317.
Lee, Y., Lee, J., and Ju, J. (2020). Perilla frutescens Britton var. frutescens leaves attenuate dextran sulfate sodium-induced acute colitis in mice and lipopolysaccharide-stimulated angiogenic processes in human umbilical vein endothelial cells. Food Sci. Biotechnol. 29 (1), 131–140. doi:10.1007/s10068-019-00711-8
Li, L., Wu, M. F, and Yu, H. Z. (2019c). Study on the inhibition of angiogenesis by berberine targeting combined with VEGFR2. J. Anhui Med. Univ. 54 (4), 503–508. doi:10.19405/j.cnki.issn1000-1492.2019.04.001
Li, J. B., Zhang, J., Luo, Y. F., and Li, X. H. (2020). Inhibitory effect of curcumin on high concentration glucose induced angiogenesis in RF/6A cells. Chin. J. Ophthalmol. 30 (6), 396–401. doi:10.13444/j.cnki.zgzyykzz.2020.06.004
Li, W., Zhao, Y., Zhang, Q., Gao, G. Y., Zhang, S. L., Gao, Y., et al. (2019a). Alkaloid extract of Corydalis yanhusuo inhibits angiogenesis via targeting vascular endothelial growth factor receptor signaling. BMC Compl. Alter. Med. 19 (1), 359. doi:10.1186/s12906-019-2739-6
Li, X. H., Li, R., and Zhu, Q. P. (2019b). Effect of quercetin on high glucose induced angiogenesis of choroid and retinal cells and its relationship with autophagy. J. Shanxi Med. Univ. 50 (12), 1732–1737. doi:10.13753/j.issn.1007-6611.2019.12.018
Li, X. P., Tang, R., Zhao, C., Li, X. R., and Liu, Y. (2018). Study on the antiangiogenic activity and cell cycle arrest activity of fructosylpuerarin. Pharmaceut. Biotechnol. 25 (2), 114–118. doi:10.19526/j.cnki.1005-8915.20180204
Li, Y. J., Duan, C. L., Li, H., Hi, , and Liu, J. X. (2010). Effect of salvianolic acid B on angiogenesis of chorioallantoic membrane of chicken embryo. J. Shenyang Pharm. Univ. 27 (11), 905–909. doi:10.1016/j.jep.2010.07.040
Li, Z., Chen, L. Z., and Liu, C. (2016). Study on the inhibitory effect of citrinin on angiogenesis. Shizhen Guoyi Guoyao 27 (2), 277–279.
Li-Li, B., Hui, Z., Min, L., et al. (2010). Inhibition effect of eriocalyxin B against angiogenesis of hepatocelluar carcinoma and human normal endothelial cell. China Pharm.. doi:CNKI:SUN:ZGYA.0.2010-15-007
Liang, Y. (2012). Experimental study on the effect of safflower polysaccharide on the expression of tumour metastasis related genes [D]. Harbin, China: Heilongjiang University Of Chinese Medicine.
Lin, W., Zheng, L. P., Zhuang, Q. C., Zhao, J. Y., Cao, Z. Y., Zeng, J. W., et al. (2013). Spica prunellae promotes cancer cell apoptosis, inhibits cell proliferation and tumour angiogenesis in a mouse model of colorectal cancer via suppression of stat3 pathway. BioMed Central 13 (1), 144. doi:10.1186/1472-6882-13-144
Liu, C., Li, X. W., Cui, L. M., Li, L. C., Chen, L. Y., and Zhang, X. W. (2011a). Inhibition of tumour angiogenesis by TTF1 from extract of herbal medicine. World J. Gastroenterol. 17 (44), 4875–4882. doi:10.3748/wjg.v17.i44.4875
Liu, C., Shen, X. H., Tang, W. J., Gu, Z. X., and Cai, W. (2015a). Study on the inhibitory effect of green tea polyphenols on the angiogenesis of chicken chorioallantoic membrane. J. Shanghai Jiaot. Univ. (Med. Sci.) 35 (1), 13–16. doi:11.3969/j.issn.1674-8115.2015.01.003
Liu, F., Xue, F. X., Guan, Y. X., Yao, J. C., and Yan, Y. (2014a). Effect of arctiin on angiogenesis of chorioallantoic membrane of chicken embryo. Shandong Med. 54 (22), 1–4. doi:10.3969/j.issn.1002-266X.2014.22.001
Liu, J., and Henkel, J. (2002). Traditional Chinese medicine (TCM): are polyphenols and saponins the key ingredients triggering biological activities? Curr. Med. Chem. doi:10.2174/0929867023369709
Liu, K., Zhong, X., and Li, Y. D. (2008a). Experimental study on the effect of Hedysari polysaccharide on the angiogenesis of chorioallantoic membrane of chicken embryo. Chinese Med. Res. 21 (12), 14–16. doi:10.3969/j.issn.1001-6910.2008.12.008
Liu, M. Z. (2016). Inhibitory effect of cycloartetane triterpenoids from Bletilla striata on angiogenesis and tumor cell proliferation. J. Huizhou Univ. 36 (06), 31–34 + 39. doi:10.16778/j.cnki.1671-5934.2016.06.005
Liu, M. Z., Qiang, N., and Lin, F. H. (2015b). Identification and antiangiogenic activity of triterpenes of Bletilla cycloatenane type. Pharmaceut. Biotechnol. 22 (2),121–124. doi:CNKI:SUN:YWSW0.2015-02-007
Liu, M. Z., Tang, J. Z., Zhang, J. S., Qiu, Q., and Huang, Y. Y. (2008b). Terpenoids in Bletilla striata inhibit angiogenesis by inducing apoptosis of vascular endothelial cells. J. Mol. Cell Biol. (5), 383–392. doi:10.3321/j.issn:1673-520X.2008.05.006
Liu, M. Z., Tang, J. Z., Zhang, J. S., and Xu, F. (2009). Effect of Bletilla terpenoids on apoptosis and cytoskeleton of human umbilical vein endothelial cells. Life Sci. Res. 13 (6), 482–486. doi:10.16605/j.cnki.1007-7847.2009.06.007
Liu, Y., Zhang, C. Y., Zhang, D. T., Jiang, H. M., and Zhang, H. (2011b). Experimental study on apoptosis and angiogenesis inhibition of GRC-1 cells induced by Hedyotis diffusa extract. Shandong Med. J. 51 (8), 97–98. doi:10.3969/j.issn.1002-266X.2011.08.051
Liu, Y. M., Zhang, D. L., Xiao, X., Liu, H. H., and Zhao, X. Y. (2012). Research progress on antitumour effect and mechanism of ursolic acid. Chongqing Med. J. 41 (4), 398–400. doi:10.3969/j.issn.1671-8348.2012.04.037
Liu, Z. G., Tao, Y., Wu, H. Y., and Wei, Z.-H. (2014b). Research progress on antitumour effect of capsaicin. Tumour 34 (4), 383–386. doi:10.3781/j.issn.1000-7431.2014.04.016
Lu, B. Y., and Cai, X. L. (2012). Preliminary study on the effect of aloe polysaccharide on tumour angiogenesis. Mod. Med. Health 28 (21), 3201–3203. CNKI:SUN:XYWS0.2012-21-002
Lu, F. B., Zhang, N., and Yu, S. W. (2015). Research progress of anti-tumour mechanism of Sanghuang polysaccharide. Shanghai J. Tradit. Chin. Med. 49 (9), 87–89. CNKI:SUN:SHZZ0.2015-09-035
Lu, X. M., Wang, S., Liu, R. J., Fan, R. L., and Song, Q. H. (2009). Research progress on antitumour effect and mechanism of silymarin. Chin. J. Pharmacol. Toxicol. 23 (04), 320–324.
Lu, Y., Wang, A. T., Chen, W. X., et al. (2004). Application of Polygonatum japonicum polysaccharide in the preparation of anticoagulants and angiogenesis inhibitors. Nanjing, China: Nanjing University of Traditional Chinese Medicine, CN200310112713.9 [P]
Luo, F. J., and Cao, Y. (2001). Molecular mechanism of tea polyphenols inhibiting tumor cell proliferation. J. Clin. Pathol. 21 (004), 251–253. doi:10.3969/j.issn.1673-2588.2001.04.005
Lv, X., Feng, Y. J., Xia, Y., Liu, J., Hou, F. G., Li, Y., et al. (2014). Inhibition of Icariin on angiogenesis of human umbilical vein endothelial cells and its mechanism. World J. Trad. Chin. Med. 9 (1), 71–74. doi:10.3969/j.issn.1673-7202.2014.01.026
Mohan, R. (2000). Curcuminoids inhibit the angiogenic response stimulated by fibroblast growth factor-2, including expression of matrix metalloproteinase gelatinase B. J. Biol. Chem. 275 (14), 10405–10412. doi:10.1074/jbc.275.14.10405
Moon, E. J., Lee, Y. M., Lee, O. H., et al. (1999a).A novel angiogenic factor derived from Aloe vera gel: β-sitosterol, a plant sterol. 3 (2), 117–123. doi:10.1023/a:1009058232389
Moon, E. J., Lee, Y. M., Lee, O. H., Lee, M. J., Lee, S. K., Chung, M. H., et al. (1999b). A novel angiogenic factor derived from Aloe vera gel: beta-sitosterol, a plant sterol. 3 (2), 117–123. doi:10.1023/A:1009058232389
Nikhil, A. S., Melissa, N. Y., and Nasser, H. (2020). .The story of angiogenesis inhibitors in non–small-cell lung cancer: the past, present, and future. Clin. Lung Canc. 21 (4), 308–313. doi:10.1016/j.cllc.2020.02.024
Niu, Y. L., Liu, S. Y., and Ding, H. X. (2009). Experimental study on Angelica polysaccharide promoting chorioallantoic membrane angiogenesis in chicken embryo. Gansu Trad. Chin. Med. 22 (9), 71–72. doi:10.3969/j.issn.1004-6852.2009.09.048
Pan, W., and Jiang, M. (2010). Effect of total flavonoids of Abelmoschus manihot on angiogenesis of chorioallantoic membrane of chicken embryo. Henan Trad. Chin. Med. 30 (6), 555–557. doi:10.16367/j.issn.1003-5028.2010.06.047
Qin, Y., Gao, Y., Gao, Y., and Li, W. M. (2017). Effect of flavonoids extract from cortex mori on angiogenesis related genes of nonalcoholic fatty liver in type 2 diabetic rats. Chin. J. Exp. Pharmacol. 23 (17), 144–148. doi:10.13422/j.cnki.syfjx.2017170144
Qu, Y. N., Fang, S. H., Li, H. L., Wan, C., Wang, X., and Sun, B. (2018a). Research progress of angiogenesis and its correlation with disease. Adv. Mod. Biomed. 18 (23), 4593–4596. doi:10.13241/j.cnki.pmb.2018.23.044
Qu, Y. N., Fang, S. H., Li, H. L., Wan, C., Wang, X., and Sun, B. (2018b). The formation of angiogenesis and its correlation with disease. Prog. Mod. Biomed. 18 (23), 4593–5459.6
Raquel, C., Ilda, R., Luísa, G., Joana, Q. L., Emília, S., Raquel, S., et al. (2017). Modulation of VEGF signaling in a mouse model of diabetes by xanthohumol and 8‐prenylnaringenin: unveiling the angiogenic paradox and metabolism interplay. Mol. Nutr. Food Res. 61 (4), 1600488. doi:10.1002/mnfr.201600488
Rhode, J., Fogoros, S., Zick, S., Wahl, H., Griffith, K., Huang, J., et al. (2007). Ginger inhibits cell growth and modulates angiogenic factors in ovarian cancer cells. BMC Compl. Alter. Med. 7, 44. doi:10.1186/1472-6882-7-44
Torres, A., Vargas, Y., Uribe, D., Carrasco, C., Torres, C., Rocha, R., et al. (2016). Pro-apoptotic and anti-angiogenic properties of the alpha/beta-thujone fraction from Thuja occidentalis on glioblastoma cells. J. Neuro Oncol. 128, 9–19. doi:10.1007/s11060-016-2076-2
Satoru, K., Noboru, T., Hiroo, N., Shinji, S., and Hiroshi, S. (2003). Oversulfation of fucoidan enhances its anti-angiogenic and antitumour activities. Biochem. Pharmol. 65 (2), 173–179. doi:10.1016/s0006-2952(02)01478-8
Shan, F., Huang, L. Q., Guo, J., and Chen, M. (2015). History and development of “one root of medicine and food. Chin. Bull. Life Sci. 27 (8), 1061–1069.
Shankar, S., Ganapathy, S., Chen, Q., and Srivastava, R. (2008). Curcumin sensitizes TRAIL-resistant xenografts: molecular mechanisms of apoptosis, metastasis and angiogenesis. Mol. Canc. 7, 16. doi:10.1186/1476-4598-7-16
Sheng, W. L., Wu, Y., Liu, F., Li, J. J., Jin, M., He, Q. X., et al. (2018). Effect of black garlic on angiogenesis. Food Ind. Technol. 39 (14), 301–305. doi:10.13386/j.issn1002-0306.2018.14.057
Shi, H. B., Chen, J. D., Chen, X. H., et al. (2013). Effects of salvianolic acid and notoginseng triterpenes on angiogenesis in EA-hy926 cells in vitro. Chin. J. Nat. Med. 11 (3), 254–257. doi:10.1016/S1875-5364(13)60024-2
Si, J. G., and Tian, C. G. (2016). Research progress of anti-tumour mechanism of ginsenoside RH 2. Shizhen Guoyi Guoyao 27 (5), 1190–1192. doi:10.3969/j.issn.1008-0805.2016.05.067
Siddavaram, N., Ramamurthi, V. P., and Veeran, V. (2012). Chlorophyllin abrogates canonical Wnt/β-catenin signaling and angiogenesis to inhibit the development of DMBA-induced hamster cheek pouch carcinomas. 35, 385–395. doi:10.1007/s13402-012-0099-z
Simonich, M. T., McQuistan, T., Jubert, C., Pereira, C., Hendricks, J. D, et al. (2008). Low-dose dietary chlorophyll inhibits multi-organ carcinogenesis in the rainbow trout. Food Chem. Toxicol. 46, 1014–1024. doi:10.1016/j.fct.2007.10.034
Sofia, B., Evdoxia, K., Eleni, B., Nawaf, A. M., Lucia, M., Marina, Z., et al. (2012). The isoflavone metabolite 6-methoxyequol inhibits angiogenesis and suppresses tumour growth. BioMed Central 11 (1), 35. doi:10.1186/1476-4598-11-35
Sohail, E., Khaleeq, A., Robina, T., and Muhammad, A. (2014). A novel link between angiogenesis and natural products: anti-angiogenic effects of Opuntia dillenii. Cen. Eur. J. Biol. 9 (3), 298–308. doi:10.2478/s11535-013-0266-x
Song, L., Di, Y., and Shi, B. (2000). The significance and development trend in research of plant polyphenols. Prog. Chem. 12 (02), 161–170. doi:10.3321/j.issn:1005-281X.2000.02.005
Su, Y. L., He, F., Sun, D., Sun, X. L., and Kang, T. G. (2014). Study on the inhibition of chlorogenic acid on angiogenesis and its mechanism. Pharmacol. Clin. Trad. Chin. Med. 30 (2), 37–40. doi:10.1186/1476-4598-11-35
Sun, M. H., Deng, R., Wang, Y., Wu, H., Zhang, Z. G., Bu, Y. H., et al. (2020). Sphingosine kinase 1/sphingosine 1-phosphate/sphingosine 1-phosphate receptor 1 pathway: a novel target of geniposide to inhibit angiogenesis. Life Sci. 256, 117988. doi:10.1016/j.lfs.2020.117988
Tang, M. K., Fan, Y., Liu, C. S., and Zhang, J. T. (2014). Mechanism and research progress of neuroprotection of salvianolic acid B. J. Neuropharmacol. 4 (1), 1–10. doi:10.3969/j.issn.13-1404/R.2014.01.001
Teleanu, R. I., Chircov, C., Grumezescu, A. M., and Teleanu, D. M. (2019). Tumour angiogenesis and anti-angiogenic strategies for cancer treatment.. J. Clin. Med. 9 (1), 84. doi:10.3390/jcm9010084
Thanekar, D., Dhodi, J., Gawali, N., Raju, A., Deshpande, P., Degan, M., et al. (2016). Evaluation of antitumour and anti-angiogenic activity of bioactive compounds from Cinnamomum tamala: in vitro, in vivo and in silico approach. South Afr. J. Bot. 104, 6–14. doi:10.1016/j.sajb.2015.09.014
Thiyagarajan, P., Senthil Murugan, R., Kavitha, K., Anitha, P., Prathiba, D., and Nagini, S. (2012). Dietary chlorophyllin inhibits the canonical NF-κB signaling pathway and induces intrinsic apoptosis in a hamster model of oral oncogenesis. Food Chem. Toxicol. 50, 867–876. doi:10.1016/j.fct.2011.12.019
Tian, L. L., Dong, J. Y., and Huang, C. J. (2012). Study on the angiogenesis activity and mechanism of Curcumol on zebrafish. Chin. J. Trad. Chinese Med. 37 (12), 1822–1825. doi:10.4268/cjcmm20121227
Tian, X. M., Wang, R., Zhang, B. K., Wang, C. L., Guo, H., and Zhang, S. J. (2013). Impact of Lycium Barbarum Polysaccharide and Danshensu on vascular endothelial growth factor in the process of retinal neovascularization of rabbit. Int. J. Ophthalmol. 6 (1), 59–61. doi:10.3980/j.issn.2222-3959.2013.01.12
Torres, A., Vargas, Y., Uribe, D., Carrasco, C., Torres, C., Rocha, R., et al. (2016). Pro-apoptotic and anti-angiogenic properties of the α /β-thujone fraction from Thuja occidentalis on glioblastoma cells. J. Neurooncol. 128 (1), 1–11. doi:10.1007/s11060-016-2076-2
Tsai, M. Y., Yang, R. C., Wu, H. T., Pang, J. H.- S., and Huang, S.- T. (2011). Anti-angiogenic eect of tanshinone IIA involves inhibition of matrix invasion and modification of MMP-2TIMP-2 secretion in vascular endothelial cells. Canc. Lett. 310 (2), 198–206. doi:10.1016/j.canlet.2011.06.031
Vaeyens, M. M., Alvaro, J. P., Jorge, B., Christian, F. S., Tommy, H., Carmeliet, P., et al. (2020). Matrix deformations around angiogenic sprouts correlate to sprout dynamics and suggest pulling activity. Angiogenesis 23 (3), 315–324. doi:10.1007/s10456-020-09708-y10.1007/s10456-020-09708-y
Wang, X. Y. (2013b). “Effect of Panax Notoginsenoside on bidirectional regulation of angiogenesis under the condition of tumour combined with myocardial ischemia injury [C],” in Proceedings of the third symposium of tonic pharmacology committee of Chinese pharmacology society (Tonic pharmacology Committee of Chinese Pharmacology Society: Chinese Pharmacology Society), 47–48.
Wang, X. Y. (2013a). “Study on the effect and mechanism of Notoginsenoside in the pathological environment of myocardial ischemia combined with tumour [C] China Association of traditional Chinese and Western medicine deficiency syndrome and geriatric medicine professional committee,” in Proceedings of the 13th National Symposium on deficiency syndrome and geriatric medicine China Association of traditional Chinese and Western medicine deficiency syndrome and geriatric medicine professional committee (Jinan University: China Association of Traditional Chinese and Western Medicine), 257–258.
Wang, Z. Q. (2015). Study on the antitumour and antioxidant effects of phenolic components in strawberry [D]. Jilin University.
Wang, X. Y., and Zhang, T. (2013). Research progress in the bidirectional regulation of neovascularization by total saponins of notoginseng. Chin J. Trad. Chinese Med. 28 (6), 1818–1822.
Wan, W. F., Zhang, X., Huang, C. G., Chen, L. G., Yang, X. B., Bao, K. Y., et al. (2020). Monensin inhibits glioblastoma angiogenesis via targeting multiple growth factor receptor signaling. Biochem. Biophys. Res. Commun. 530, 479–484. doi:10.1016/j.bbrc.2020.05.057
Wang, A. Y., Tao, L., Lu, Y., Lin, Y. P., Chen, W. X., and Zhou, L. (2015a). Advances in the study of salvia miltiorrhiza intervention in tumour and ischemic Disease angiogenesis. Chin. Herbal Med. 46 (9), 1399–1404. doi:10.7501/j.issn.0253-2670.2015.09.025
Wang, C., Chen, Y., and Wang, Y. (2019a). Inhibition of COX-2, mPGES-1 and CYP4A by isoliquiritigenin blocks the angiogenic Akt signaling in glioma through ceRNA effect of miR-194-5p and lncRNA NEAT1. J. Exp. Clin. Canc. Res. 38 (1), 371. doi:10.1186/s13046-019-1361-2
Wang, C., Gong, L. H., Guo, C. C., He, L. F., Zhao, X. T., and Li, Y. X. (2019b). Application progress of zebrafish as a model organism in screening effective substances of traditional Chinese medicine. Chinese Herbal Med. 50 (24), 6125–6134.
Wang, H., Li, A., Dong, X. P., and Xu, X. Y. (2008). Screening of antitumour active sites and antiangiogenic effect of Taraxacum seeds. Chinese Materia Medica (5), 718–722.
Wang, H. B., Lin, Y. C., Lin, W., et al. (2010). The effect of Ginsenoside Rg3 on the proliferation and angiogenesis of HNE-1 cells in human nasopharyngeal carcinoma. Canc. Preven. Treat. Res. 11. doi:10.3971/j.issn.1000-8578.2010.11.004
Wang, J., Chen, X. Y., Hou, L., Li, L., Lu, H., and Liu, W. (2012). Effect of tea polyphenols on the expression of NF - κ B, COX-2 and Survivin in Lewis lung cancer transplanted tumour. Chin. J. Lung Canc. 15 (5), 271–276. doi:10.3779/j.issn.1009-3419.2012.05.04
Wang, W., Yu, S. Y., Wu, J. W., Huang, S., Wu, Y. Y., Chen, W. X., et al. (2017). Treatment limitations and Strategies of tumour angiogenesis inhibitors. Chin. Pharmacol. Bull. 33 (11), 1489–1492. doi:10.3969/j.issn.1001-1978.2017.11.003
Wang, X. J., Yuan, S. L., and Wei, Y. Q. (2005). Anticancer effect of plant polyphenols. Natural product research and development 17 (4), 508–517. doi:10.3969/j.issn.1001-6880.2005.04.033
Wang, X. Y., Li, G. W., and Chang, S. X. (2014a). Mechanism of Panax Notoginsenoside improving postmenopausal osteoporosis through angiogenesis. Chin. J. Osteoporos. 20 (8), 964–967. doi:10.3969/j.issn.1006-7108.2014.08.0237
Wang, Y., Li, J. X., Wang, Y. Q., and Miao, Z. H. (2015b). Medical frontier forum and the 14th National Academic Conference of tumour pharmacology and chemotherapy abstracts collection. Department of medicine and healthChina Anti Cancer Association, 141.Antiangiogenic effect and mechanism of tanshinone I [C] Department of medicine and health, Chinese Academy of engineering, professional committee of anticancer drugs of China Anti Cancer Association, professional committee of tumour pharmacology of Chinese pharmacology AssociationChinese Academy of engineering, professional committee of anticancer drugs of China Anti Cancer Association, professional committee of tumour pharmacology of Chinese pharmacology Association2015.
Wang, Y. N., Cao, R., Zhu, C., and Wu, X. Z. (2014b). Study on the effect and mechanism of sulfated polysaccharide of Prunella vulgaris on angiogenesis of liver cancer. Chin. J. Oncol. 41 (12), 758–761. doi:10.3969/j.issn.1000-8179.2014067210.4028/www.scientific.net/amr.1030-1032.758
Wang, Z., Li, J. A., Xiao, W. A., Long, J. F., and Zhang, H. M. (2018). The STAT3 inhibitor S3I-201 suppresses fibrogenesis and angiogenesis in liver fibrosis. Lab. Invest.: Advancing the understanding of human and experimental disease 98 (12), 1600–1613. doi:10.1038/s41374-018-0127-310.1038/s41374-018-0127-3
Wang, Z. G., Chen, Z., Yang, S. S., Wang, Y., Huang, Z. Y., Gao, J. F., et al. (2014c). Berberine ameliorates collagen-induced arthritis in rats associated with anti-inflammatory and anti-angiogenic effects. Inflammation 37 (5), 1789–1798. doi:10.1007/s10753-014-9909-y
Wang, Z. Q., et al. (2015c). Antitumour activity and preliminary mechanism of isoliquiritigenin. Chin. Pharmacol. Bull. 31 (8), 159–1165. doi:10.3969/j.issn.1001-1978.2015.08.026
Wei, J. G., Yang, D. S., Chen, W. Y., Wang, X. M., Wang, Y. Y., Yang, Y. P., et al. (2014). Studies on the chemical constituents and antiangiogenic activity of Ampelopsis amurensis. Chinese herbal medicine 45 (7), 900–905. doi:10.7501/j.issn.0253-2670.2014.07.002
Wei, P. B, Fei, C., and Ling, B. (2010). Dihydrotanshinone I inhibits angiogenesis both in vitro and in vivo[J]. Acta Biochim. Sinica. (1), 1–6. doi:10.1111/j.1745-7270.2008.00370.x
Wen, P. N., Lin, M. S., Jin, M., Xu, X. X., and Chen, L. Y. (2020). Effect of Dihydromyricetin on angiogenesis of gastric cancer in vivo and in vitro through ERK/VEGFA/VEGFR2 pathway. Chin. J. Canc. 30 (2), 113–121. doi:10.19401/j.cnki.1007-3639.2020.02.005
Wu, J. (2014). Research progress of anti-tumour mechanism of ginsenoside RH 2. Changzhou Prac. Med. (6), 418–420.
Wu, Y. W. (2019). Mechanism of asiaticoside in the treatment of burn wounds [D]. Nanjing University of traditional Chinese medicine.
Wu, P., and Jiang, M. (2010). Study on the effect of total flavonoids of Abelmoschus manihot on the angiogenesis of chicken embryo allantoic membrane. Chinese Folk Med. 19 (11), 22–23. doi:10.3969/j.issn.1007-8517.2010.11.015
Wu, K. N., Lin, H., Kong, L. Q., Liu, S. C., and Li, H. Y. (2001). Effect of polysaccharide of Coriolus versicolor on atypical hyperplasia and angiogenesis of breast. Chinese J. Exp. Surgery (4), 84. doi:10.3760/j.issn:1001-9030.2001.04.038
Xie, L. J., and Li, L. H. (2019). The role of vascular endothelial growth factor in cardiovascular diseases. Int. J. Cardiol. 46 (4), 219–222. doi:10.3969/j.issn.1673—6583.2019.04.008
Xin, R., Pan, H. F., Shao, S. L., and Xu, X. M. (2017). Anti-tumour and anti-angiogenic effects of Fucoidan on prostate cancer: possible JAK-STAT3 pathway. BioMed Central 17 (1), 378. doi:10.1186/s12906-017-1885-y
Xin, Y., Jiang, X., Cui, J. S., and Ni, J. S. (2010). Inhibitory effect of 20 (s) - ginsenoside Rg3 on proliferation and migration of vascular endothelial cells. Cancer Preven. Treat. Res. 37 (12), 1352–1355. doi:10.3971/j.issn.1000-8578.2()10.12.005
Xing, Y. F., Liu, X. J., Wang, Z. B., and Li, F. Q. (2014). Study on the antiangiogenic effect of genistein on MKN45 gastric cancer cells and their subcutaneous transplanted tumors. J. Chongqing Med. Univ. 39 (12), 1686–1691. doi:10.13406/j.cnki.cyxb.000435
Xu, B. H., Shen, X. C., Zhang, L., Yuan, D. P., Long, J., Yu, B., et al. (2012). The effect of salvianolic acid B on endothelial cell migration and lumen formation. Chin. Pharmacol. Bull. 28 (4), 496–500. doi:10.3969/j.issn.1001-1978.2012.04.013
Xv, J. Z., Liu, W., and Liu, Y. H. (2016). Inhibitory effect of sulfated polysaccharide of Laminaria Scirpus on chicken embryo allantoic membrane angiogenesis. Shandong J. Tradit. Chin. Med. 35 (3), 241–243. doi:10.16295/j.cnki.0257-358x.2016.03.024
Xv, Z. P., Li, F. C., and Wang, H. R. (1999). Inhibition of angiogenesis and antitumour effect of laminarin sulfate. Chinese Herbal Med. (7), 551–553. doi:10.3321/j.issn:0253-2670.1999.07.036
Yan, H. P., and Zhao, Y. (2020). The role of EGF like domain 7 in angiogenesis. J Prac. Med. 37 (2), 179–182. doi:10.14172/j.issn1671-4008.2020.02.027
Yang, B. R., Hong, S.-J., Simon, M.-Y. L., Cong, W. H., Wan, J.-B., et al. (2016). Pro-angiogenic activity of notoginsenoside R1 in human umbilical vein endothelial cells in vitro and in a chemical-induced blood vessel loss model of Zebrafish in vivo. Chin. J. Integr. Med.: English Edition 22 (6), 420–429. doi:10.1007/s11655-014-1954-8
Yang, Y., and Zhou, Z. T. (2005). Research progress of genistein's anti-tumour angiogenesis mechanism. Internat. J. Stomatol. 32 (3), 210–211. doi:10.3969/j.issn.1673-5749.2005.03.015
Yang, M., Chen, G. L., and Chen, C. (2009). Study on the effect of Ginsenoside Rg1 on angiogenesis in the heart of rats with myocardial infarction. Chin. Med. Emerg. 18 (3), 403–405. doi:10.3969/j.issn.1004-745X.2009.03.044
Ye, L. H., Pan, H. J., Huang, Y. Y., and Xu, S. T. (2018). Diagnostic value of combined detection of angiopoietin-2, vascular endothelial growth factor and carbohydrate antigen 72-4 in gastric cancer. Lab. Med. Clin. 15 (24), 3715–3717. doi:10.3969/j.issn.1672-9455.2018.24.01910.20892/j.issn.2095-3941.2017.0182
Yin, Y. Y., Liu, S. S., Han, L. W., He, Q. X., Zhang, Q. W., Liu, K. C., et al. (2016). Analysis of alkaloids and antiangiogenic activity of Evodia rutaecarpa. Chin. J. Exper. Pharmacol. 22 (5), 45–53. doi:10.13422/j.cnki.syfjx.2016050045
Yu, X. P. (2003). Effects of trihydroxyl isoflavones on angiogenesis in HER-2/neu highly expressed breast cancer and its mechanism [D]. Third Military Medical University. doi:10.7666/d.y550975
Yu, X. R. (2002). Definition, scope and research progress of medicinal food. Chin. Herb. Med. (12), 914–916. doi:10.13863/j.issn1001-4454.2002.12.029
Yuan, L., and Pu, H. Z. (2011). Research progress of antitumour effect of capsaicin. Med. J. Yanbian Univ. 34 (1), 68–71. doi:10.16068/j.1000-1824.2011.01.012
Yu, B., Yi, L., Zhu, J. D., Zhang, Q. Y., and Mi, M. T. (2010a). “Study on the mechanism of Heimi anthocyanin inhibiting the expression of angiogenic factor in HER-2/neu high expression human breast cancer cell line MDA-MB-453 [C]. Chongqing nutrition society,” in Proceedings of the academic conference of Chongqing nutrition society (Chongqing Nutrition Society: Chongqing Science and Technology Association), 63.
Yu, B., Yu, X. P., Yi, L., Zhu, J. D., Zhang, Q. Y., and Mi, M. T. (2010b). Mechanism of inhibitory effect of anthocyanin of black rice on the expression of angiogenic factors in human breast cancer cells. J. Nutr. 32 (6), 545–550.
Yu, X. P., Mi, M. T., and Zhu, J. D. (2003). Effect of genistein on angiogenesis of breast cancer xenograft tumour. China Pub. Health (10), 20–22. doi:10.11847/zgggws2003-19-10-13
Yuan, Y., Yu, H. Y., sun, T. T., et al. (2013). Experimental study on inhibition of angiogenesis by saponins in Platycodon grandiflorum. Chinese Pat. Med. (5), 1068–1070. doi:10.3969/j.issn.1001-1528.2013.05.050
Yuan, Y., Zhu, L., Sun, T. T., Zhao, F. L., Liu, M. X., and Li, J. P. (2014). Effect of saponins in Platycodon grandiflorum on angiogenesis of sarcoma S180 chicken embryo chorioallantoic membrane transplant tumour. Chin. J. New drugs Clin. Med. 33 (12), 911–915.
Zeng, J., Huang, F., Tu, Y. Q., Wu, S. C., Li, M. P., and Tong, X. Y. (2013). Protective effect of catalpol on myocardium in rats with isoprenaline-induced myocardial infarcts via angiogenesis through endothelial progenitor cells and Notch1 signaling pathway. Sci. Res. Publ. 4, 8. doi:10.4236/pp.2013.48088
Zhang, F. (2013). Study on the anti fibrosis mechanism of curcumin and Ligustrazine on hepatic stellate cells [D]. Nanjing, China: Nanjing University of Traditional Chinese Medicine.
Zhang, L. J. (2017). Experimental study of resveratrol targeting peripheral cells to inhibit angiogenesis and anti glioma effect [D]. Zhengzhou, China: Henan University of traditional Chinese medicine.
Zhang, J. H., Liu, W. J., and Luo, H. M. (2018). Research progress in the activity of terpenoids in medicinal plants. World Sci. Technol. – Modernization Trad. Chinese Med. 20 (3), 419–430. doi:10.11842/wst.2018.03.015
Zhang, Y. M. (2007). Study on the mechanism of tea polyphenols against angiogenesis of transplanted mouse breast cancer [D]. Beijing University of traditional Chinese medicine.
Zhang, Y. J., Cao, J. H., and Xu, X. D. (2018). The role and mechanism of Sema4D in tumour angiogenesis. Shandong Med. J. 58 (48), 94–96. doi:10.3969/j.issn.1002-266X.2018.48.027
Zhang, Y. M., Chen, X. Y., and Xu, L. (2007). Study on the expression of angiogenesis related factors in breast cancer tissues and important organs by tea polyphenols. Prog. Mod. Biomed. (10), 1441–1444. doi:CNKI:SUN:SWCX0.2007-10-003
Zhao, X. Y., Ding, J., and Liu, Y. H. (2019). Inhibitory effect of Schisandra polysaccharide on chicken embryo allantoic membrane angiogenesis. J. Shandong Med. College 41 (2), 93–95. doi:10.3969/j.issn.1674-0947.2019.02.007
Zheng, F., Cui, X. D., Zhu, J. B., and Yin, X. Z. (2018). Inhibitory effect of soybean isoflavone on angiogenesis of transplanted tumour of liver cancer in mice. Soybean Sci. 37 (1), 101–110. doi:10.11861/j.issn.1000-9841.2018.01.01014
Zheng, Wei., Yang, Xh., and Wu, Lina. (2007). Molecular mechanism of curcumin in antiangiogenesis. Chin. Clin. Oncol. 34 (12), 683–685. doi:10.3969/j.issn.1000-8179.2007.12.008
Zheng, X. Y., Cao, N. N., Pei, S., Zhang, D. D., Lin, Y. X., Xiao, X. F., et al. (2019). Ginsenoside RH-2 research progress in the mechanism of anti hepatoma action. Chinese Journal of experimental formulary 25 (20), 208–213. doi:10.13422/j.cnki.syfjx.20191329
Zhong, J. H., Xu, F. T., Xu, M. J., Wang, X. C., Wang, Y. L., Liu, X., et al. (201). Effect of Ginkgo biloba extract EGb761 on angiogenesis and expression of VEGF and TSP-1 in mouse lung adenocarcinoma. Jiangxi Med. 51 (12), 1344–1346. doi:10.3969/j.issn.1006-2238.2016.12.015
Zhou, P., Cheng, Y. Z., Liu, F. L., Wu, K., Qiu, W. L., and Wang, S. Q. (2019). Tanshindiol-C suppresses in vitro and in vivo hepatocellular cancer cell growth by inducing mitochondrial-mediated apoptosis, cell cycle arrest, inhibition of angiogenesis and modulation of key tumour-suppressive miRNAs. J. BUON 24 (2), 622–627
Zhu, Q. J. (2017). Experimental study on the antiangiogenic effect of Lentinan in vitro. J. Tradit. Chin. Med. 32 (12), 2426–2429. doi:10.16368/j.issn.16748999.2017.12.631
Zhu, J. D., Yu, X. P., and Mi, M. T. (2005). Inhibitory effect of genistein on angiogenesis of breast cancer xenografts in nude mice with HER-2/neu overexpression. China Clin. Rehabil. (6), 228–230. doi:10.7666/d.y550975
Zhu, Y. J., Cheng, P., Su, X. M., Liu, Z., Yang, B., and Zhang, T. (2012). Study on the effect of salidroside on the proliferation of hepatoma cells and the angiogenesis of zebrafish. Mod. Oncol. 20 (11), 2239–2243. doi:CNKI:SUN:SXZL0.2012-11-008
Keywords: angiogenesis, food active components, homology of medicine and food, nutritional food, medicinal food
Citation: Pan D, Gong X, Wang X and Li M (2021) Role of Active Components of Medicinal Food in the Regulation of Angiogenesis. Front. Pharmacol. 11:594050. doi: 10.3389/fphar.2020.594050
Received: 12 August 2020; Accepted: 10 November 2020;
Published: 22 January 2021.
Edited by:
Federica Finetti, University of Siena, ItalyReviewed by:
Simona D’Aguanno, Istituti Fisioterapici Ospitalieri (IRCCS), ItalyCopyright © 2021 Pan, Gong, Wang and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xiaoqin Wang, bnl3YW5neGlhb3FpbkAxNjMuY29t; Minhui Li, bGltaW5odWlAeWVhaC5uZXQ=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.