 Lee Han Jie1
Lee Han Jie1 Ibrahim Jantan
Ibrahim Jantan Syaratul Dalina Yusoff
Syaratul Dalina Yusoff Juriyati Jalil
Juriyati Jalil Khairana Husain
Khairana Husain- 1Drug and Herbal Research Centre, Faculty of Pharmacy, Universiti Kebangsaan Malaysia, Kuala Lumpur, Malaysia
- 2Institute of Systems Biology, Universiti Kebangsaan Malaysia, Bangi, Malaysia
Sinensetin, a plant-derived polymethoxylated flavonoid found in Orthosiphon aristatus var. aristatus and several citrus fruits, has been found to possess strong anticancer activities and a variety of other pharmacological benefits and promising potency in intended activities with minimal toxicity. This review aims to compile an up-to-date reports of published scientific information on sinensetin pharmacological activities, mechanisms of action and toxicity. The present findings about the compound are critically analyzed and its prospect as a lead molecule for drug discovery is highlighted. The databases employed for data collection are mainly through Google Scholar, PubMed, Scopus and Science Direct. In-vitro and in-vivo studies showed that sinensetin possessed strong anticancer activities and a wide range of pharmacological activities such as anti-inflammatory, antioxidant, antimicrobial, anti-obesity, anti-dementia and vasorelaxant activities. The studies provided some insights on its several mechanisms of action in cancer and other disease states. However, more detail mechanistic studies are needed to understand its pharmacological effects. More in vivo studies in various animal models including toxicity, pharmacokinetic, pharmacodynamic and bioavailability studies are required to assess its efficacy and safety before submission to clinical studies. In this review, an insight on sinensetin pharmacological activities and mechanisms of action serves as a useful resource for a more thorough and comprehensive understanding of sinensetin as a potential lead candidate for drug discovery.
Introduction
Sinensetin is a polymethoxylated flavonoid found in Orthosiphon aristatus var. aristatus [syn: Orthosiphon stamineus Benth., Orthosiphon spicatus (Thunb.) Backer, Bakh.f. & Steenis] (Lambiaceae) and in many citrus fruits. In citrus fruits, it is particularly more abundant in the peel than other parts of the fruit (Manthey and Grohmann, 2001; Youn et al., 2017). A polymethoxylated flavonoid has a double bond between positions 2 and 3 and a ketone in position 4 of the C ring (Panche et al., 2016). Sinensetin is a pentamethoxyflavone, chemically known as 2-(3,4-dimethoxyphenyl)-5,6,7-trimethoxy-4H-1-benzopyran-4-one. The flavone skeleton is substituted by methoxy groups at positions 5, 6, 7, 3′ and 4’, respectively, with a molecular formula of C20H20O7 and molecular weight of 372.4 g/mol. Figure 1 shows the chemical structure of sinensetin.
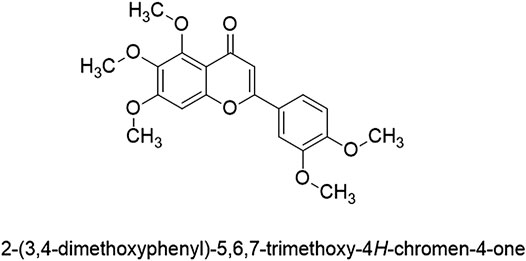
FIGURE 1. Structure of sinensetin.
Orthosiphon aristatus is widely employed in traditional medicine as a diuretic (Beaux et al., 1999; Adam et al., 2009) and to treat hypertension (Mohamed et al., 2012; Manshor et al., 2013), rheumatism, tonsillitis, gout, menstrual disorder and diabetes (Adam et al., 2009; Mohamed et al., 2012). The leaves of O. aristatus have been shown to exhibit excellent pharmacological activities such as antioxidant (Awale et al., 2003), antibacterial (Ho et al., 2014), hepatoprotective (Yam et al., 2007), anti-inflammatory (Hsu et al., 2010), cytotoxic (Yam et al., 2007), antihypertensive (Azizan et al., 2012) and gastroprotective (Yam et al., 2009). Many citrus fruits have been used traditionally for many medicinal purposes. For instance, fruit peels of Citrus reticulata Blanco [syn: Citrus sunki Hort. ex Tanaka, Citrus unshiu (Yu.Tanaka ex Swingle), Citrus depressa Hayata, Citrus deliciosa Ten.] have been used in Asian ethnomedicine to treat indigestion, bronchial, fever, snakebite, stomachache, edema, cardiac diseases, bronchitis and asthmatic conditions (Kang et al., 2013). Moreover, many Citrus species among the different variety of C. reticulate are recognized as food supplements and nutrition while their leaf extracts and essential oils have shown strong pharmacological activities such as antimicrobial (Tao et al., 2014), anti-inflammatory (Nasri et al., 2017), antioxidant (Nasri et al., 2017), anticancer (Benavente-Garcia O and Castillo, 2008), antiproliferative (Du, and Chen, 2010), hypoglycemic (Aruoma et al., 2012) and analgesic effects (Nasri et al., 2017).
There were several reports on the isolation of sinensetin from O. aristatus.Tezuka et al. (2000) reported the isolation of 75 mg of sinensetin from 4 kg of plant material by subjecting its methanol extract to repeated column chromatography followed by preparative thin layer chromatography (TLC). However, lower yields were obtained by other workers. Yuliana et al. (2009) reported 3.03 mg of sinensetin was isolated from 500 g of O. aristatus using silica column, sephadex LH-20 column and preparative TLC. Another study reported that from 1 kg of O. aristatus, only 2.6 mg of the compound was isolated by repeated column chromatography followed by preparative TLC (Amzad Hossain and Mizanur Rahman, 2015). The variable yields of sinensetin isolated by these workers might probably be due to the different separation techniques employed and other factors such as climate variability, soil conditions and geographical factors. Various yields of sinensetin were obtained from different Citrus species. Out of 100 mg ethyl acetate extract of C. reticulata, 23.4 mg sinensetin was isolated by using sephadex LH-20 column chromatography and preparative TLC (Nakanishi et al., 2019). The peels of C. reticulata (500 g of dry weight) yielded 38.8 mg of sinensetin by using silica gel column chromatography (Nagase et al., 2005). Citrus reticulata subjected to high-speed counter current chromatography (HSCCC) separation and preparative high performance liquid chromatography (HLPC) yielded 27.7 mg of sinensetin (Du and Chen, 2010). Polymethoxyflavone-enriched extract of Sweet Portuguese oranges (Newhall variety), obtained by supercritical fluid technology contained sinensetin at 17.36 mg/g as determined by HPLC with diode array detection (HPLC-UV/DAD) (Pereira et al., 2019). Sinensetin was the second most abundant methoxylated flavonoids isolated from 1 kg peels of C. sinensis (L.) Osbeck (Malterud and Rydland, 2000). Iwase et al. (2000), reported that from dried peels (955 g) of C. sinensis 13.4 mg sinensetin was isolated. Sinensetin is also available as a synthetic compound as it can be commercially prepared. Amzad Hossain and Ismail (2004) reported synthesis of sinensetin in high yield by alkaline condensation of 2,4,5,6-tetrahydroxyacetophenone and 3, 4-di (methoxymethoxy) benzaldehyde to give 2′, 4′, 5′, 6′, 3, 4-hexa (methoxymethoxy) chalcone, followed by dimethoxymethylation in acid medium and treatment with 2,3-dichloro-5,6-dicyano-1,4-benzoquinone to yield 5,6,7,3′,4′-pentahydroxyflavone. Sinensetin was obtained by methylation of 5,6,7,3′,4′-pentahydroxyflavone using dimethyl sulphate and potassium carbonate.
Pharmacological Activities and Mechanisms of Action
In this review, an updated overview of the pharmacological activities of sinensetin such as anticancer, anti-inflammatory, antioxidant, antimicrobial, anti-obesity, anti-dementia, vasorelaxant and antitrypanosomal activities and their underlying mechanisms of action are presented. The databases employed for data collection are mainly from Google Scholar, PubMed, Scopus and Science Direct. The keywords used during searching include ‘sinensetin’ AND ‘activity’ OR ‘antiproliferative’ OR ‘antiangiogenesis’ OR ‘antitrypanosomal’ OR ‘antimicrobial’ OR ‘antioxidant’ OR ‘antidementia’ OR ‘vasorelaxant’ OR ‘anti-inflammatory’ OR ‘anticancer’. The papers to be included need to fulfil the inclusion criteria as follows: original research papers related to any activity exhibited by sinensetin; sinensetin used can either be synthetic or an isolated natural compound but not in crude extract or mixture; published in English; experiments conducted are in vivo and/or in vitro and/or ex vivo assays. Most researchers used commercially available synthetic sinensetin to evaluate its pharmacological activities due to its limited supply from natural sources. The scientific information on sinensetin, sourced from natural sources and synthesis, are critically analysed to provide perspectives for future studies on the compound as a lead molecule for the development of new therapeutic agents. In addition, toxicological information have also been included to assess its safety before submission to clinical trials. Table 1 summarizes the various pharmacological activities of sinensetin such as anticancer (Choi et al., 2002, Choi et al., 2004; Mertens-Talcott et al., 2007; Ohtani et al., 2007; Du and Chen, 2010; Du and Chen, 2010; Tan et al., 2013; Zhang et al., 2014; Bai et al., 2019; Pereira et al., 2019); antiobesity (Sergeev et al., 2009; Kang et al., 2013, Kang et al., 2015), antioxidant (Lyckander and Malterud, 1996; Malterud and Rydland, 2000; Jeong et al., 2007; Yao et al., 2009); antidementia (Nagase et al., 2005; Kawahata et al., 2013; Kimura et al., 2013; Swaminathan et al., 2014; Youn et al., 2017), anti-inflammatory (Laavola et al., 2012; Shin et al., 2012; Chae et al., 2017), antiangiogenesis (Lam et al., 2012) and diuretic (Yuliana et al., 2009).
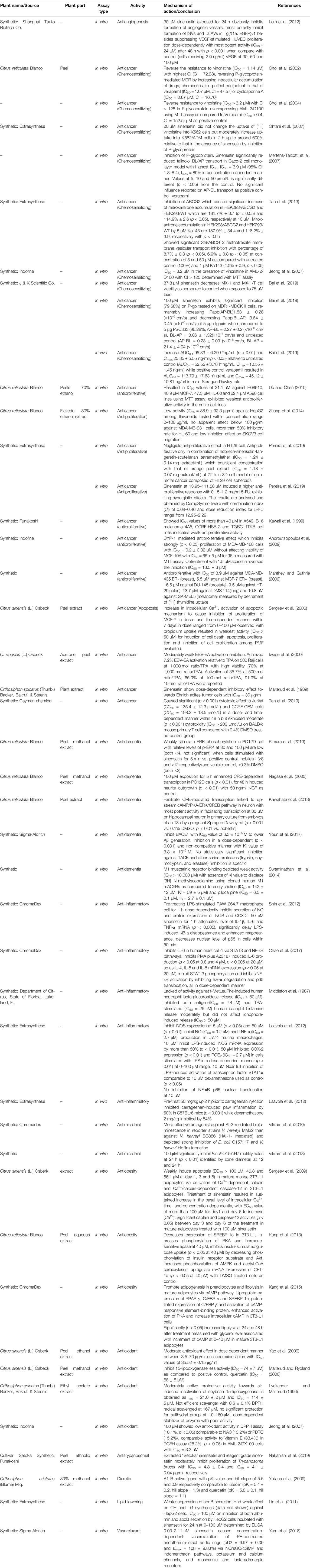
TABLE 1. Pharmacological activities of sinensetin.
Anticancer Activity
The search for new anticancer drugs with high therapeutic index has led to great extent of anticancer research on plant-derived natural products such as flavonoids. Sinensetin is one of the polymethoxylated flavonoids that has been intensively studied for its anticancer activity. Chemotherapy remains as a standard in the treatment and management of cancer (Qiu et al., 2019). However, the major obstacle to successful chemotherapy is the development of multidrug resistance (MDR) by tumor cells after long term administration of chemotherapy drugs (Choi et al., 2002; Qiu et al., 2019). Prominent group of ATP-binding cassette (ABC) transporters such as P-glycoprotein (P-gp), Multiresistance Protein (MRP) and ATP Binding Cassette Subfamily G Member or also known as breast cancer resistance protein (ABCG2 or BCRP) have been characterized in relation to MDR, limiting the exposure to anticancer drugs (Choi and Yu, 2014; Wilkens, 2015; Waghray and Zhang, 2018). Sinensetin has been intensively investigated for its anticancer activities and its mechanisms of anticancer activities involving interaction with P-gp, breast cancer resistance protein (ABCG2/BCRP), antiproliferation, and antiangiogenesis are critically discussed.
Interaction of Sinensetin With P-Glycoprotein
Sinensetin has been reported to inhibit P-glycoprotein (P-gp) in several studies. Elevated expression of P-glycoprotein (P-gp) due to regulation of human gene expression of MDR1 is a significant cause for tumor multidrug resistance (MDR) (Qiu et al., 2019). Overexpression of P-gp has been linked to reduced chemotherapeutic responses and poor clinical outcomes in various cancer types (Waghray and Zhang, 2018). Therefore, P-gp inhibition is nominated as an effective strategy to reverse MDR (Qiu et al., 2019). Sinensetin, was found to significantly inhibit P-gp in a few cell types such as MDR1-MDCK II cells, Caco 2 cells and K562/ADM cells (Mertens-Talcott et al., 2007; Ohtani et al., 2007; Bai et al., 2019). Subsequently, sinensetin was reported to induce antiproliferation response synergistically with 5-fluorouracil in HT29 cell (Pereira et al., 2019). These findings revealed the potential of sinensetin as a new flavonoid chemosensitizer. Chemosensitizers are in need so that MDR cells can be sensitized to anticancer drugs when treated simultaneously (Choi et al., 2002). Calcium channel blocker, verapamil and various drugs such as phenothiazine, quinidine, cyclosporine A, valspodar (PSC 833) and much more had been reported to be potential drugs to overcome MDR (Wang et al., 1995; Fromm et al., 1999; Tidefelt et al., 2000; Summers et al., 2004; Wesolowska et al., 2009). While many compounds able to inhibit P-gp function have been identified, no broadly applicable inhibitor is in use yet. This is due to poor potency, significant side effects, unsatisfactory toxicity and low selectivity of the compounds (Wilkens, 2015). Potent and nontoxic P-gp inhibitors are needed to increase drug accumulation and reversal of drug resistance (Leonard et al., 2003). Choi et al. (2002) reported that sinensetin exhibited the most potent chemosensitizing effect among all tested flavonoids with an IC50 value (1.14 µM) comparable to that of verapamil (IC50 = 1.07 µM) and cyclosporine (IC50 = 0.87 µM), with the presence of vincristine in P-gp overexpressing AML-2/D100 cells. Sinensetin inhibitory activity was observed in the presence of vincristine which is also a good substrate of P-gp. The inhibitory effect of sinensetin on P-gp was well-supported by other studies (Mertens-Talcott et al., 2007; Bai et al., 2019; Danisman Kalindemirtas et al., 2019). However, there was no chemosensitizing activity of sinensetin against MRP-overexpressing cancer cells AML-2/DX100 (Choi et al., 2002). In a further screening of flavonoid sensitizer conducted by Choi et al. (2004), sinensetin was reported to exhibit IC50 value of 3.2 µM (chemosensitizing index, CI = 125).
Aside from chemosensitizing, sinensetin aided in the reversal of MDR phenotype in aspects such as decreased accumulation of the drug and enhanced efflux of anticancer drugs. As previously reported by Choi et al. (2002), sinensetin significantly (p < 0.05) increased the accumulation of rhodamine-123, a drug transported by P-gp in AML-2/D100 at a concentration of 20 µM. However, the dose accumulation determined by flow cytometry of sinensetin was 4- and 16-folds lower than those of verapamil and cyclosporine A, when concentrations were compared to induce the same effect. The difference thought may be due to different incubation times or other cellular effects of sinensetin than P-gp inhibition (Choi et al., 2002). Sinensetin increased digoxin uptake in rats with significant increment relative to control in AUC0-t and Cmax although weaker than verapamil as a positive control. The effects of several flavonoids including sinensetin on the pharmacokinetics of digoxin in rats after administration of 0.25 mg/kg digoxin were evaluated. Results disclosed significant increment in AUC0-t 95.33 (p < 0.01) and Cmax 25.85 (p < 0.05) caused by sinensetin. In addition, Vd/F and Cl/F of digoxin were also decreased after pre-administration with sinensetin with 11.6 (p < 0.05) and 2.48 (p < 0.01), respectively (Bai et al., 2019). These findings on sinensetin were consistent with increment of rhodamine-123 accumulation in AML-2/D100 (Choi et al., 2002), reduced talinolol transport by P-gp from basolateral to apical (BL/AP) side in Caco-2 colon carcinoma cell monolayer model (Mertens-Talcott et al., 2007) and increased uptake of [3H] vincristine into adriamycin-resistant human myelogenous leukemia (K562/ADM) cells (Ohtani et al., 2007).
Sinensetin also decreased P-gp levels even at the maximum non-cytotoxic concentration unlike verapamil, cyclosporine A and nefedipine which increased P-gp expression with an increase in MDR1 mRNA level (Choi et al., 2002). Maximum non cytotoxic concentration (more than 90% of control survival) of sinensetin, 25 µM was administered to AML-2/D100 for 72 h and appeared to decrease P-gp levels in Western blot analysis. Meanwhile, verapamil and cyclosporine A showed an increase in P-gp levels. This may suggest long term use of sinensetin as a better chemosensitizing agent than verapamil and cyclosporine A without concern about P-gp activation. Modulators blocked the substrate transportation by binding competitively and noncompetitively with P-gp. Apart from that, cyclosporin A and verapamil partially inhibited photolabeling of P-gp by [3H] azidopine but not by sinensetin at the same excess molar concentrations of 100 µM. This result may suggest that sinensetin did not interact directly with P-gp at the azidopine-binding site. Collectively, sinensetin was believed to have significant advantages of having a high therapeutic index or chemosensitizing index (CI), of being a non-transportable inhibitor and having no induction of P-gp in long term use.
Interaction of Sinensetin With Breast Cancer Resistance Protein
Sinensetin has been shown to inhibit the efflux transporter, ATP Binding Cassette Subfamily G Member or also known as breast cancer resistance protein (ABCG2 or BCRP), which is one of the players taking part in multidrug resistance same as P-gp (Mo and Zhang, 2012). Overexpression of ABCG2 has been studied and correlated to MDR phenotypes of numerous cancer cell lines derived from tumor types such as SN-38-selected human small cell lung cancer cells PC-6/SN2-5, mitoxantrone-selected human gastric carcinoma cell line EPG85-257RNOV, gefitinib-resistant non-small cell lung cancer (NSCLC) cells, epirubicin-resistant human hepatocyte carcinoma cells HLE-EPI, topotecan and doxorubicin-selected human multiple myeloma cells (Mo and Zhang, 2012). ABCG2 contributed to cancer progression due to its active efflux of chemotherapeutic agents out of neoplastic cells (Natarajan et al., 2012). The inhibiting properties of sinensetin against the efflux transporter ABCG2 was determined by in vitro mitoxantrone accumulation assay using wild type HEK293 (human embryonic kidney), HEK293/WT and ABCG2-overexpressing HEK293 cell line (HEK293/ABCG2) and sf9 (Spodoptera frugiperda) insect cell membrane vesicles overexpressing ABCG2 for the cell-based and the membrane-based methods, respectively. The results showed that 10 µM sinensetin significantly (p < 0.05) enhanced mitoxantrone accumulation in both HEK293/ABCG2 (181.7%) and HEK 293/WT cells (114.9%). However, mitroxantrone accumulation increment in HEK 293/WT cells has no significant difference from 5 µM Ko143 treatment effect (118.2%, p < 0.05). Sinensetin restricted transport of methotrexate into Sf9/ABCG2 to 8.7% (p < 0.05) at 5 μM and 6.9% (p < 0.05) at 50 μM, which were comparable to 1 µM Ko143 (4.0%) (Tan et al., 2013). The identified ABCG2 modulating ability of sinensetin may provide a framework for further investigation to identify potential MDR agents from phytochemicals.
Antiproliferation Effect of Sinensetin
Cancer stem cell (CSC), considered as one of the main causes of resistance to chemotherapeutic drug, is responsible for tumor initiation, relapse, invasion and metastases (Ishimoto et al., 2015; Otoukesh et al., 2018). MicroRNAs can participate in modulating multiple genes and CSCs functions (Otoukesh et al., 2018). Several miRNAs were found to target mRNA of epithelial-mesenchymal transition (EMT) such as ZEB1, ZEB2, or SNAI1 (Vu and Datta, 2017). Thus, it can also be one of the ways to target CSC. Synergistic interaction of sinensetin with 5-fluorouracil besides modulation of EMT genes has been reported (Pereira et al., 2019). EMT genes were modulated by sinensetin such that tumor suppressor gene, CDH1 was increased while ZEB1 and SNAI1 were reduced. Sinensetin alone did not present antiproliferative effect. However, nobiletin-sinensetin-tangeretin-scutellarian tetramethylether (N-S-T-SC), mixture of four polymethoxylated flavones (PMF) with concentration equivalent to that of orange peel extract (OPE) presented similar response as OPE. In terms of antiproliferation, expression of cell cycle marker, CDKN1A (coding for p21) and CCNA2 (coding for cyclin A2) genes and the apoptotic marker BICR5 (encoding baculoviral IAP repeat-containing protein 5) were also evaluated with sinensetin. No significant activity was observed on lowering expression of BICR5, presented lower numbers of caspase-3 active cells to induce apoptosis as BICR5 prevented apoptosis by caspase-3 inhibition. Besides, overexpression of CDKN1A might lead to G1/S cell cycle arrest while reduced CCNA2 was proposed to arrest G2/M cell cycle. However, the effect of sinensetin was negligible in the aspect of antiproliferation in HT29 cell and this was supported by expression data of BICR5, CDKN1A and CCNA2. Expression of cancer stemness markers (PROM1 and LGR5), EMT genes (SNAI1, ZEB1 gene and CDH1) and percentage of ALDH + cells were evaluated to assess the effect of sinensetin in modulating cancer stemness and self-renewal. Sinensetin at 46.62 µM, reduced LGR5 (p ≤ 0.001), decreased expression of mesenchymal markers, SNAI1 (p ≤ 0.001) and ZEB1 (p ≤ 0.001) and increased CDH1 (p ≤ 0.0001), the results were expressed relative to untreated control (Pereira et al., 2019).
According to Androutsopoulos et al. (2009), sinensetin possessed CYP-1 mediated antiproliferative activity in MDA-MB-468 breast adenocarcinoma cell line. Sinensetin strongly inhibited (p < 0.05) in vitro proliferation of MDA-MB-468 cells with IC50 = 0.2 µM without affecting the viability of normal breast cell line MCF-10A with IC50 = 65 µM for 96 h measured with MTT assay. Co-treatment of sinensetin with 1.5 µM acacetin reversed the inhibition (IC50 = 13.5 µM), confirming the involvement of CYP1 as acacetin is a CYP1 family enzyme inhibitor. Sinensetin was metabolised by CYP1 enzymes and produced various metabolites which were unidentified by HPLC analysis. The metabolites were suggested to be involved in its antiproliferative activity (Androutsopoulos et al., 2009). In the study conducted by Iwase et al. (2000), sinensetin exhibited 7.2% relative to control (100%) on Epstein-Barr virus (EBV-EA) activation without causing cytotoxic effect on Raji cells with 70% viability. Evaluation of EBV-EA inhibition is now applied to identify possible anti-tumor agents (Kapadia et al., 2000). The inhibition effect was moderately weak as the reported range of all the compounds tested was 0–12.7% of activation. Manthey and Guthrie (2002) studied the effects of several classes of phenols, 40 natural and synthetic flavonoids on six cancer cell lines including MDA-MB-435 human breast cancer cell, MCF-7 ER + human breast cancer cells, DU-145 androgen receptor-negative human prostate cancer cells, HT-29 human colon cancer cells, DMS-114 human lung cancer cells and SK-MEL5 human melanoma cells. Sinensetin showed a strong antiproliferative activity against MDA-MB-435 ER-and MCF-7 ER+, moderate against HT-29, DMS 114 and SK-MEL5 and weakest against DU-145, measured by decrement of [3H] thymidine uptake in treated cells. In addition, it is interesting to find that the activity of 5-desmethyl sinensetin was significantly higher than sinensetin with lower IC50 values against all cell lines (Manthey and Guthrie, 2002). In another study, sinensetin isolated from the flavedo of C. reticulata exhibited more than 50% inhibitory rate on promyelocytic leukemia HL-60 cells. Activity against Hep G2 liver cells although reported low s, it was the first time reported for antiproliferative effect on the growth of liver cancer (Zhang et al., 2014).
Sinensetin was found to significantly impede Jurkat cell proliferation and additionally triggered apoptosis and autophagy (Tan et al., 2019). Human T-cell acute lymphoblastic leukemia, CCRF-CEM, non-Hodgkin’s lymphoma cell line, Jurkat cell and mouse primary T-cells isolated and purified from six week-old male BALB/c mice were used in this study. Sinensetin inhibited cell viability in different strength at concentration ranged 1–100 μmol/L for 24 or 48 h compared with 0.4% DMSO-treated control group dose- and time-dependently. It caused significant (p < 0.001) cytotoxic effects to Jurkat and CCRF- CEM cells at concentrations of 25–100 µmol but moderate (p < 0.01) toxicity to mouse primary T-cells at a dose range of 100–200 μM at 24 and 48 h. Based on cell viability results evaluated by CCK-8 assays, IC50 values at 48 h were 135.4, 198.3 and >200 μmol/L for Jurkat, CCRF- CEM cells and mouse primary T-cells, respectively. Sinensetin induced sub-G1 population, an indicator of apoptotic death significantly (p < 0.001) when Jurkat cell treated with 50 and 100 µM for 24 and 48 h, suggesting cell-cycle arrest was involved in the growth inhibition. Considerable increase (p < 0.001) in total Annexin V+ apoptotic cells in Jurkat cells treated with sinensetin was observed. Membrane potential and protein expression of cleaved poly (ADP-ribose) polymerase (cleaved-PARP), caspase-3, caspase-8 and caspase-9 were significantly increased in Jurkat cell treated with sinensetin. The results suggested that sinensetin provoked caspase-dependent apoptosis. Acid vesicular organelles (AVOs) were used to detect autophagy. Cells treated also revealed an exhibition of more AVOs (p < 0.001) which increased with time and concentration. In the same study, sinensetin downregulated phosphorylation of Akt and mTOR dose-dependently. Inhibition of those two tremendously led Jurkat cells to be more sensitive to cytotoxicity effect of sinensetin in Jurkat cells, suggest that co-treatment of sinensetin with Akt and mTOR inhibitor synergistically blocked Akt/mTOR signaling pathway (Tan et al., 2019).
Antiangiogenesis Effect of Sinensetin
Growing evidences of the correlation between angiogenesis and chronic inflammation urge the search for effective natural occurring antiangiogenesis agents. In the study by Lam et al. (2012), sinensetin was reported as having the most potent antiangiogenesis activity with the lowest toxicity among seven polymethoxylated flavonoids. Sinensetin inhibited angiogenesis by inducing cell cycle arrest in the G0/G1 phase in human umbilical vein endothelial cell (HUVEC) culture and downregulating the mRNA expressions of angiogenesis genes flt 1, kdrl, and hras in zebrafish (Danio rerio). Sinensetin at 30 µM demonstrated the most potent inhibition on formation of intersegmental vessels (ISVs) and dorsal longitudinal anastomotic vessels (DLAVs) when transgenic zebrafish embryos, Tg (fli1a: EGFP) γ1 incubated for 24 h compared to hesperidin, nobiletin, scutellarein and scutellarein tetramethylether (STE). At 3 μM sinensetin affected the formation of ISVs with mild antiangiogenesis activity while 30 μM almost completely arrested the growth of ISVs. Sinensetin has the most potent inhibitory activity (IC50 = 24 µM) and significant dose-dependent suppression of VEGF-induced HUVEC proliferation after 48 h at a concentration range of 0–100 µM with cells receiving VEGF (20 ng/ml) served as a control. Inhibition of HUVEC proliferation might be due to cell cycle arrest in the G0/G1 phase. The expression of flt1, hras, and kdrl mRNA were downregulated by 0.56, 0.44, and 0.52-fold, respectively, after treatment with 30 μM sinensetin. Meanwhile, after treatment with 10 and 30 μM sinensetin the expression of vegfaa mRNA was significantly upregulated by about 1.70 fold as compared to control.
Anti-inflammatory Activity
Previously, many flavonoids had been found to be affecting the function of cells such as mast cell, basophils, neutrophils and platelets to modulate allergy and inflammatory responses (Middleton et al., 1987). Activated macrophage induces pro-inflammatory mediators such as PGE2, NO, IL-1β, IL-6 and TNF-α produced by inflammatory enzymes iNOS and COX-2 to cause damage to pathogen and host cells (Laavola et al., 2012; Shin et al., 2012). Middleton et al. (1987) reported sinensetin showed moderate activity on the release of basophil histamine but not neutrophil beta-glucuronidase. Basophil histamine release stimulated by antigen, tetradecanoyl phorbol acetate (TPA) and calcium ionophore, A 23,187 were by distinct immunologic, receptor-directed and non-receptor directed pathways, respectively. Sinensetin inhibited both antigen- and TPA-stimulated histamine release with IC50 values of 44 and 26 μM, respectively (Middleton et al., 1987).
Similar activities were displayed in different mechanisms and cells by other workers (Laavola et al., 2012; Shin et al., 2012; Chae et al., 2017). LPS and TNF-α are stimuli that activate macrophages during an inflammatory response. The anti-inflammatory activity of sinensetin was studied by using LPS-stimulated RAW 264.7 cells. LPS produces inflammatory response by activating MAPK and NF-ĸB pathways which involve degradation of inhibitor ĸB (iĸB). Sinensetin inhibited NF-ĸB activation by inhibiting degradation or stabilizing of IĸB-α, suppressing excessive inflammation with no effect on MAPK phosphorylation (Shin et al., 2012; Chae et al., 2017). Activated NF-ĸB can be terminated by the export of nuclear NF-ĸB and degradation of p65 in the nucleus. Pre-treating LPS-stimulated RAW 264.7 macrophage cell for 1 h inhibited secretion of NO and protein expression of iNOS and COX-2 dose-dependently. Sinensetin at 50 μM attenuated the level of pro-inflammatory cytokines, IL-1β, IL-6 and TNF-α mRNA (p < 0.005). Significantly delayed LPS-induced IĸB-α disappearance and enhanced reappearance were also reported, together with the finding of decreased nuclear level of p65 in cells within 50 min (Shin et al., 2012). Similar activity was displayed by dose-dependent inhibition of p65 translocation in human mast cell-1 (HMC-1) (Chae et al., 2017).
Laavola et al. (2012) further revealed sinensetin inhibited the production of prostaglandin E2 (PGE2) with IC50 value of 2.7 µM in a dose-dependent manner besides providing evidence to support the findings of Shin et al. (2012) on iNOS and COX-2 expression and NO production inhibition. In the carrageenan-induced paw inflammation test, sinensetin (50 mg/kg) treatment via intraperitoneal route was reported to inhibit carrageenan-induced paw inflammation in C57BL/6 mice (Laavola et al., 2012). The effect was comparable to 2 mg/kg dexamethasone which inhibited paw edema response by 84%. Chae et al. (2017) investigated human mast cell-1 (HMC1) mediated inflammatory mechanism. It was found that sinensetin inhibited PMA plus A23187-induced IL-6 production in a dose-dependent manner as well as IL-4, IL-5 and IL-8 mRNA expressions. Elevated level of IL-6 in stimulated mast cell was lowered by treatment with sinensetin, at a concentration range of 0.8–0.20 µM. Treatment of PMA plus A23187 induced IL-6 production and suppression of increased mRNA expression with the treatment of sinensetin was assayed by RT-PCR. Increment of IL-4, IL-5 and IL-8 mRNA expression in 6 h stimulated HMC-1 were significantly (p < 0.05) decreased with pretreatment of 20 µM sinensetin for 0.5 h (Chae et al., 2017). Sinensetin also inhibited signal transducer and activation of transcription 3 (STAT3) phosphorylation in PMA plus A23187-stimulated HMC-1. Activated STAT3 up-regulated the expression of inflammation mediators and induced inflammation-relevant gene expression. The inhibition of sinensetin was in a dose-dependent manner with HMC-1 pretreated with sinensetin for 0.5 h and stimulated for 0.5 h to detect phosphorylated STAT3 (Chae et al., 2017). Overall, these findings proposed sinensetin involvement in mast cell-mediated inflammatory responses, providing a clue for further findings on anti-inflammatory mechanisms in the future.
Antioxidant Activity
Inhibition of 15-lipooxygenase is of interest for researchers as it was proposed to have a role in oxidation of low density lipoprotein (LDL) which believed to be a significant step leading to atherosclerotic lesions. Air may induce inactivation of 15-lipooxygenase by loss of sulfhydryl groups through oxidation. Sinensetin was a moderate inhibitor of air-induced inactivation of soybean 15-lipooxygenase with an IC50 value of 114 µM besides acting as a poor stabilizer of enzyme which did not possess significant protection for sulfhydryl group at concentrations up to 160 μM, as reported by Lyckander and Malterud (1996). In a study by Malterud and Ryland (2000), sinensetin inhibited 15-lipooxyygenase with an IC50 value of 74.0 µM which was higher than that of the positive control, quercetin (IC50 = 68 ± 5 µM). Free radical scavenging activity of sinensetin was assayed by using diphenylpicryhydrayzyl (DPPH) and dichlorofluorescin (DCFH). Sinensetin showed low antioxidant activity in DPPH assay (Lyckander and Malterud, 1996), employing AML2/DX100 cells which possessed supersensitivity towards hydrogen peroxide treatment due to down-regulated catalase expression comparable to positive control antioxidants, N-acetyl-l-cysteine (NAC) or pyrrolidine dithiocarbamate (PDTC). However, comparable antioxidant activity to vitamin E (33.4%) was observed in DCFH fluorescence-scavenging assay (26.2%, p < 0.05). Besides, Yao et al. (2009) revealed antioxidant effect of sinensetin in a dose dependent manner between 3.5 and 70 μg/ml on superoxide anion with an IC50 value of 35.52 μg/ml.
Effects on Lipolysis Metabolism
Previous study by Sergeev et al. (2006) indicated that sinensetin induced apoptosis via increasing basal intracellular Ca2+ level and activation of Ca2+ mediated, caplain-dependent caspase-12. There are different in vitro experiments conducted on 3T3-L1 preadipocyte cells to study the effects and mechanisms of sinensetin on lipid metabolism. The adipocytes generated exhibited most of the key features of adipocytes in vivo for the 3T3-L1 cells. Induction of adipocyte apoptosis to reduce body fat may result in long-lasting weight loss, as compared with after caloric restriction as well as bariatric surgery. Sergeev et al. (2006) undertaken a study to determine whether polymethoxyflavones (PMFs) including 5-hydroxy-3,6,7,8,3′,4′-hexamethoxyflavone, 3,5,6,7,8,3′,4′-heptamethoxyflavone, 5,6,7,3′,4′-pentamethoxyflavone and 3′-hydroxy-5,6,7,4′-tetramethoxyflavone could induce Ca2+-mediated apoptosis in mature adipocytes. All the PMFs increased basal levels of intracellular Ca2+ and induced apoptosis in dose and time-dependent manner in mature 3T3-L1 adipocytes treated with PMFs of concentration range between 3.125 and 100 µM for 1, 3, or 6 days. Sinensetin increased intracellular Ca2+ with EC50 of >100 μM, 45.9 µM, >100 µM for 1, 3 and 6 days, respectively. As for the induction of apoptosis, EC50 values of all the PMFs including sinensetin were generally the same range as those for inducing intracellular Ca2+. However, the effective concentration for inducing apoptosis was lower than those for inducing intracellular Ca2+ at day 6 of treatment with an EC50 value of 56.1 µM. Cellular apoptotic changes were also shown when treated at 50 µM for 3 and 6 days labelled with Annexin V-Alexa Fluor 488. Subsequently, it was reported that the sustained increase of intracellular Ca2+ level was accompanied by activation of caplain and caspase-12 activities measured with specific fluorogenic peptide substrates when treated with sinensetin (p < 0.05 for day 3). From this experiment, it was found that sinensetin induced apoptosis via increasing basal intracellular Ca2+ level and activation of Ca2+ mediated, caplain-dependent caspase-12.
Besides, adipose tissue lipolysis actually controls the quantity of triglycerides stored in fat depots and in determining plasma free fatty acid levels (Kang et al., 2013). Effects and lipid metabolism of sinensetin isolated from PMF-rich C. reticulata peel was studied and the mechanism or pathway was elucidated in mature 3T3-L1 adipocytes. A decrease in the level of sterol regulatory element-binding protein (SREBP) 1c expression was revealed when treated with 40 µM of sinensetin. Sinensetin increased glycerol released into culture supernatants and stimulated phosphorylation of protein kinase A (PKA) and hormone-sensitive lipase (HSL), suggesting that sinensetin enhanced lipolysis by cAMP-dependent PKA. Glucose uptake and phosphorylation of Akt and IRS both were inhibited when treated by sinensetin. The results proposed that by reduction of phospho-insulin receptor substrate (IRS) and phosphor-Akt, sinensetin modulated insulin action includes uptake of glucose and insulin antilipolytic action. AMP-activated protein kinase (AMPK) was vital in glucose and lipid metabolism, to increase CPT-1a mRNA expression. 6, 12 or 24 h of 40 µM sinensetin exposure markedly induced the phosphorylation of AMPK and acetyl-CoA carboxylase (ACC), increased phosphorylation of LKB1 and the level of CPT-1a mRNA. These results suggested sinensetin took part in the AMPK signaling pathway (Kang et al., 2013).
An imbalance between adipogenesis and lipolysis leads to obesity. Results reported by Kang et al. (2015) revealed enhancement of adipogenesis in preadipocytes and lipolysis in mature adipocytes. Treatment with 2, 10 and 40 µM sinensetin resulted in increment of cellular lipid accumulation and triglyceride content in a dose-dependent manner in 3T3-L1 cells. Somehow the effect was less than positive control which was treated with 0.5 mM 3-isobutyl-1-methylxanthine (IBMX). Sinensetin also upregulated expression of peroxisome proliferator-activated receptor (PPAR)γ1, PPARγ2, CCAAT/enhancer-binding protein (C/EBP) α and fatty acid-binding protein aP2. These results suggested sinensetin enhanced lipogenesis in preadipocytes 3T3-L1. Adiponectin, as a direct target of PPARγ increased significantly (p < 0.05) in a dose-dependent manner. Sinensetin possessed ability the same as IBMX to increase expression of differentiation signals, SREBP1c and C/EBPβ relative to DMSO-treated cell in 3T3-L1 preadipocytes. It also increased phosphorylation of cAMP-responsive element-binding protein, CREB and extracellular signal-regulated kinase ERK, significantly (p < 0.05) increase phosphorylated forms of PKA at 0.25 and 0.5 h compared with DMSO-treated cells. cAMP levels increased dose-dependently with 30 min treatment of sinensetin. These results suggested sinensetin enhanced lipogenesis in preadipocytes 3T3-L1 via cAMP/PKA pathway. In search of the effect of sinensetin on lipolysis, glycerol levels in culture supernatant were measured. It was reported that sinensetin significantly increased lipolysis at 24 and 48 h after treatment measured with glycerol level associated with an increment of cAMP at 0–40 µM in mature 3T3-L1 adipocytes (Kang et al., 2015).
Antidiabetic Effect
Mohamed et al. (2013) demonstrated that fraction consisted mainly of sinensetin and eupatorin possessed the highest glucose-lowering effect in O. aristatus. It reduced glucose absorption of jejunum without changing the insulin level in streptozocin-induced rat. Hydrolysis of starch by pancreatic α-amylase and uptake of glucose by intestinal α-glucosidase caused hyperglycaemia in type 2 diabetes mellitus when there was a sudden increase in blood glucose level. Potentiation of α-glucosidase and α-amylase inhibition may be used as an effective strategy for post-prandial hyperglycemia linked to type2 diabetes mellitus. Mohamed et al. (2012) reported strong inhibition percentages (89.0–32.0%) for α-glucosidase at concentration range of 2.5–0.31 mg/ml. The activity was comparably strong with IC50 value of 0.66 mg/ml relative to acarbose (IC50 = 1.93 mg/ml, p < 0.01) and 50% ethanolic extract of O. aristatus (IC50 = 4.63 mg/ml, p < 0.01). However, Damsud et al. (2014) had different findings which showed no inhibition towards yeast α-glucosidase and rat intestinal α-glucosidases (maltase and sucrase) when 10 µL sinensetin was mixed with 0.1 U/mL α-glucosidase. In addition, sinensetin showed maximum inhibition of 85.8% at 2.5 mg/ml against porcine α-amylase (Mohamed et al., 2012). IC50 value of sinesetin (1.13 mg/ml) was significantly lower than both acarbose (IC50 = 4.89 mg/ml, p < 0.05) and 50% ethanolic extract of O. aristatus (IC50 = 36.70 mg/ml, p < 0.001). The provided in vitro evidence demonstrated appreciable inhibitory effect of sinensetin on α-amylase and α-glucosidase and generated a stronger rationale for further studies on sinensetin.
Diuretic Effect
Since O. aristatus is used for several kidney disorders, gout and as diuretics traditionally, it was examined for its diuretic and hypoglycemic effects. One of the reports on the diuretic activity of O. aristatus described significant enhancement of ion excretion, especially potassium ion urinary excretion, although activity was lower than that of furosemide treated in Sprague–Dawley rats model (Arafat et al., 2008; Adam et al., 2009). Diuretic action is vital in kidney stone treatment, dissolving the stone when fluid volume flowing through kidney increases. Adenosine A1 receptor antagonist function as diuresis and sodium excretion inducer affords renal protection (Modlinger and Welch, 2003; Welch, 2015; Tofovic et al., 2016). Adenosine A1-receptor binding assay was also carried out on seven active methoxy flavonoids from O. aristatus (Yuliana et al., 2009). Sinensetin exhibited pKi value of 5.5 µM which was comparable to thise of luteolin (pKi = 5.4) and quercetin (pKi = 5.8). All flavonoids including sinensetin, luteolin and quercetin demonstrated a one-site curve with a Hill slope value within 0.9–1.4, not significantly different with unity, indicating the antagonist effect.
Vasorelaxant Effect
Vasorelaxant effect of sinensetin was reported by Yam et al. (2018) recently, employing in vitro pre-contraction aortic ring assay. It was suggested that the mechanism of action might be via NO/sGC/cGMP and indomethacin pathways, potassium and calcium channels, and muscarinic and beta-adrenergic receptors. Sinensetin at a range of 0.03–2.11 µM caused concentration-dependent vasorelaxation of 1 µM phenylephrine (PE) pre-contracted endothelium–intact aortic rings (pD2 = 6.97, Emax = 108%). There were still sinensetin-induced vasorelaxation in various conditions or pretreated with different antagonist and the results obtained from endothelium-denude (pD2 < 5.67, Emax = 31.38%), L-NAME (pD2 = 6.42, Emax = 75.46%), ODQ (pD2 = 6.18, Emax = 76.72%) and methylene blue (pD2 = 5.52, Emax = 52.69%) were significantly reduced (p < 0.001). Potassium channels involvement determined with the application of potassium blocker on endothelium-intact aortic 20 min before pre-contraction with PE. Propanolol as a β-blocker significantly (p < 0.05) decreased the vasorelaxation effect of sinensetin (pD2 = 6.71, Emax = 95.4%). Besides, sinensetin (0.04–0.58 µM) and calcium channel blocker, nifedipine (0.1–1.0 µM), significantly reduced the increase in contraction caused by the addition of 3 mM calcium ions, diminishing the effect of calcium ions induced contraction (p < 0.05) by 0.65–0.31 g and 0.1–0.03 g, respectively. Sinensetin depicted concentration-dependent vasorelaxant effect on endothelium-intact aortic rings. The evidence supported the suggestion that vasorelaxant effect of sinensetin did not solely dependent on endothelium but endothelium-independent relaxing factors might partially contribute.
Antidementia
Nagase et al. (2005) isolated sinensetin from the peel of C. reticulata and reported on its antidementia activity using transient transfection and reporter gene assay. cAMP/protein kinase A (PKA)/Extracellular related kinase (ERK)/cAMP response element-binding protein (CREB) linked to CRE-mediated transcription is vital in the aspect of learning and memory (Kawahata et al., 2013). cAMP response element (CRE) transcription is dysregulated in neurodegenerative disorders in the central nervous system. Protective action against neurodegeneration in CNS includes stimulating CRE-dependent transcription and induce neurite outgrowth. Sinensetin from C. reticulata peel enhanced CRE-dependent transcription (p < 0.01) after 5 h treatment at 100 µM in PC12D cells. In conjunction, 100 µM sinensetin exposition for 48 h induced neurite outgrowth (p < 0.01). Among the compounds tested nobiletin and 6-demethylnobiletin were more potent than sinensetin (Nagase et al., 2005). However, Kawahata et al. (2013) found that sinensetin isolated from C. reticulata peel enhanced transcription to a greater extent than nobiletin at concentration up to 30 µM (p < 0.001) tested on CRE-mediated transcription in cultured hippocampal neuron from embryos of an 18-days pregnant Sprague-Dawley rat. Sinensetin exhibited the most potent activity among the tested compounds (tangeretin, 6-demethoxynobiletin, 6-demethoxytangeretin and nobiletin) with p value <0.01 as compared to nobiletin, facilitating the transcription. The discrepancy between the results of both studies might be due to the cell-type-dependent difference in the response of these compounds. Meanwhile, Kimura et al. (2013) studied the effect on ERK phosphorylation in PC12D cells. No significant result was observed when cells were stimulated with sinensetin. Previous studies have suggested the generation of intraneuronal amyloid-beta (Aβ) oligomers is an early event in the pathogenesis of Alzheimer’s disease (Kimura et al., 2013). Beta-site amyloid precursor protein (APP) cleaving enzyme1 (BACE1) catalyzes the rate-limiting step of Aβ generation, considered as a prime target for Alzheimer’s disease (Youn et al., 2017). Sinensetin inhibited BACE1 with an IC50 value of 6.3 × 10–5 M, to lower Aβ generation in a dose-dependent manner (p < 0.001) with Ki value of 3.8 × 10–5 M. Tangeretin (4.9 × 10–5 M) has the highest potency followed by nobiletin (5.9 × 10–5 M) and sinensetin. Inhibition of sinensetin was specific, exhibiting no statistically significant inhibition against TACE and other serine proteases (trypsin, chymotrypsin, elastase) (Youn et al., 2017).
Antimicrobial Effect
When 19 citrus cultivar were investigated for their antitrypanosomal activities to tackle African trypanosomiasis such as Trypanosoma brucei, T. brucei gambiense and T. brucei rhodesiense, “Setoka” extract exhibited the most potent inhibitory effect (36%) on T. brucei proliferation with highest activity in its ethyl acetate extract (Nakanishi et al., 2019). “Setoka” is one of the second-generation hybrids between “Kiyomi” (C. reticulata) and other Citrus species. All of the compounds subjected to the parasite inhibition assay showed potent activities against T. brucei in a dose-dependent manner. Pentamidine (IC50 = 0.001 μg/ml) used as a standard was the most potent, followed by nobiletin (IC50 = 2.4 μg/ml), reagent grade nobiletin (IC50 = 3.1 μg/ml), reagent grade sinensetin (IC50 = 4.1 μg/ml) and sinensetin (IC50 = 4.8 μg/ml). 293T human embryonic kidney cell was employed to assess toxicity against mammalian cells by resazurin assay. Selectivity was calculated for all these compounds based on IC50 values against T. brucei and 293T human embryonic kidney cells. Descending sequence of selectivity was as follows; pentamidine (SI = 3,400), nobiletin (SI = 5.4), reagent grade nobiletin (SI = 4.5), reagent grade sinensetin (SI = 2.9), sinensetin (SI = 2.7), HMF (SI = 1.3). Taken together all the data reported, sinensetin and nobiletin were responsible for antitrypanosomal activity in “Setoka” peel. Unfortunately, the IC50 values and selective indexes were not comparable with those of modern antitrypanosomal candidates. Vikram et al. (2010) evaluated quorum sensing, biofilm and type three secretion system (TTSS) inhibitory properties of citrus flavonoids including sinensetin using in vitro model. Sinensetin dose-dependently inhibited AI-2- mediated bioluminescence, demonstrating maximal response at 100 μg/ml in Vibrio harveyi mutant strain MM32 (AI-2). Meanwhile, sinensetin strongly inhibited biofilm formation in E. coli O157: H7 and V. harveyi without inhibiting the growth of V. harveyi and E. coli O157: H7. The results were proposing inhibition of A1-mediated cell-cell signalling by sinensetin lead to antagonistic activity against biofilm formation. Continuation of work further discovered 100 µM of sinensetin significantly (p < 0.05) inhibited motility halos of E. coli O157: H7 by measuring zone diameter after culture of EHEC was stabbed in 0.3% LB-agar plate for 12 and 24 h (Vikram et al., 2013).
Toxicology
The medicinal benefits of sinensetin have raised the interest of many researchers to further study its pharmacological and toxicological properties. The results of toxicological studies are listed in Table 2. For instance, Lam et al. (2012) reported the lowest lethal toxicity of sinensetin as compared to nobiletin and hesperetin by using zebrafish embryo. Beating heart rate of zebrafish embryo was the parameter used to indicate the living status of embryo. The toxicity of the compounds at concentration of 30, 100 and 300 µM for 48 h was determined. At 48 h post-treatment with 100 µM sinensetin, about 40% of embryos were reported alive, achieving a 94.6% survival rate. However, Nakanishi et al. (2019) revealed non-negligible cytotoxic effect of sinensetin and reagent grade sinensetin against 293T human embryonic kidney cells with IC50 values of 11.2 and 13.8 μg/ml, respectively, by a resazurin assay in vitro. Most of the cytotoxic effects conducted reported minimal cytotoxicity, as compared to other tested compounds and untreated control. Sinensetin at 20 µM demonstrated 84.5% growth of K562/ADM cells when 10 µL WST-1 was used to evaluate cell viability treated for 48 h (Ohtani et al., 2007). The result was a ratio relative to that in the absence of vincristine, indicating not much different from the untreated control (85.3%). Sinensetin exerted toxicity against AML-2/D100 cells with an IC50 value of more than 400 µM while the effective dose for its synergistic chemosensitizing effect with vincristine has an IC50 value of 3.2 µM (Choi et al., 2004). No activity was observed too as in the study conducted by Jeong at al. (2007) up to concentration of 400 µM. These studies indicate that sinensetin exerted minimal toxicity to normal cells and possessed high selectivity.
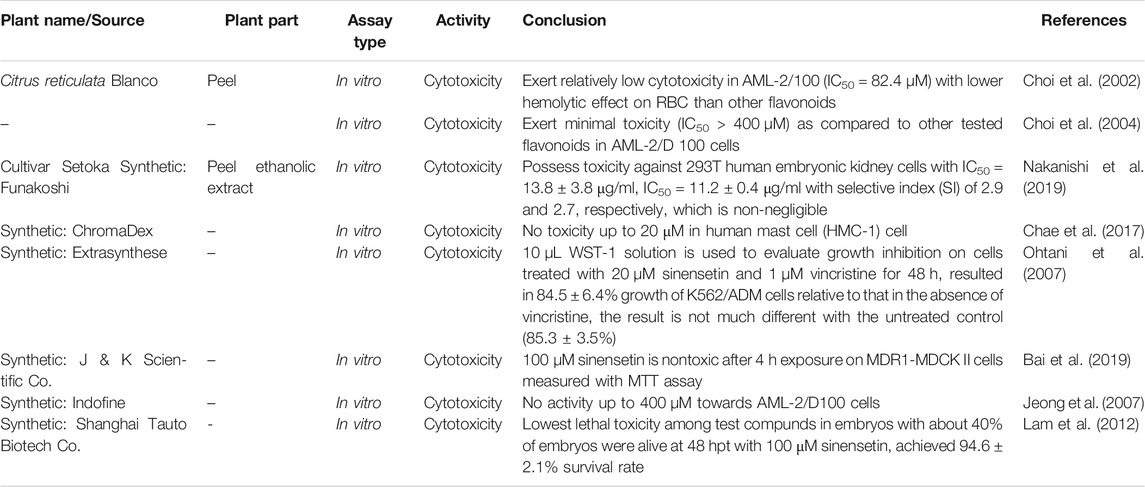
TABLE 2. Toxicity of sinensetin.
Conclusion
Undeniably, in recent years, there is an elevating global demand in natural products for healthcare and also finding leading compounds for several diseases in drug discovery and development. The growing demand and significance of natural products and compounds should not be underestimated. Sinensetin was studied and found to be possessing a diversity of beneficial pharmacological effects such as anticancer, antidiabetic, antidementia, vasorelaxant, antimicrobial and anti-inflammatory activities. Since 1989, people had paid much attention to its anticancer activity. Many in vivo and in vitro studies had indicated that sinensetin not only showed good activity towards tumor cells, but also exert minimal toxicity to normal cells according to the ratio of IC50 values, and possessed high selectivity. Thus, we conclude sinensetin is a promising anticancer drug candidate for future development to delay cancer progression and prolong survival. Since we are still lacking effective anticancer drug with good safety profile, potential of sinensetin will be worth studying. However, most of the mechanisms are poorly understood for all activities aforementioned. Therefore, more investigation and further studies of mechanisms of action need to be performed before it can be used as a therapeutic drug in the future. There is a necessity for performing toxicology profile of sinensetin in vivo in future research to further evaluate its safety profile because as shown in Table 2, all the toxicity studies were performed in vitro. The ability of sinensetin to aid in delaying cancer progression is definitely worth further studying with the evidence of IC50 values and selectivity index in previous studies as aforementioned. Focusing on the anticancer ability of sinensetin, more supporting studies should be carried out such as the exact mechanisms and all the genes and extend of genes expression to further develop sinensetin as a lead compound.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Author Contributions
LHJ obtained the literatures and wrote the draf while KH, IJ, JJ, and SDY edited the manuscript.
Funding
The authors would like to thank Universiti Kebangsaan Malaysia (UKM) for the financial support under Grant No. DIP-2018-029.
References
Adam, Y., Somchit, M. N., Sulaiman, M. R., Nasaruddin, A. A., Zuraini, A., Bustamam, A. A., et al. (2009). Diuretic properties of Orthosiphon stamineus Benth. J. Ethnopharmacol. 124, 154–158. doi:10.1016/j.jep.2009.04.014
Amzad Hossain, M., and Ismail, Z. (2004). Synthsis of sinensetin, a naturally occuring polymethoxyflavone. Pak. J. Sci. Ind. Res. 47, 268–271. doi:1016/j.foodres.2020.109114
Amzad Hossain, M., and Mizanur Rahman, S. M. (2015). Isolation and characterisation of flavonoids from the leaves of medicinal plant Orthosiphon stamineus. Arab. J. Chem. 8, 218–221. doi:10.1016/j.arabjc.2011.06.016
Androutsopoulos, V. P., Ruparelia, K., Arroo, R. R., Tsatsakis, A. M., and Spandidos, D. A. (2009). Cyp1-mediated antiproliferative activity of dietary flavonoids in Mda-Mb-468 breast cancer cells. Toxicology. 264, 162–170. doi:10.1016/j.tox.2009.07.023
Arafat, O. M., Tham, S. Y., Sadikun, A., Zhari, I., Haughton, P. J., and Asmawi, M. Z. (2008). Studies on diuretic and Hypouricemic effects of Orthosiphon stamineus methanol extracts in rats. J. Ethnopharmacol. 118, 354–360. doi:10.1016/j.jep.2008.04.015
Aruoma, O. I., Landes, B., Ramful-Baboolall, D., Bourdon, E., Neergheen-Bhujun, V., Wagner, K. H., et al. (2012). Functional benefits of citrus fruits in the management of diabetes. Prev. Med. 54 (Suppl), S12–S16. doi:10.1016/j.ypmed.2012.02.012
Awale, S., Tezuka, Y., Banskota, A. H., and Kadota, S. (2003). Inhibition of NO production by highly-oxygenated diterpenes of Orthosiphon stamineus and their structure-activity relationship. Biol. Pharm. Bull. 26 (4), 468–473. doi:10.1248/bpb.26.468
Azizan, N. A., Ahmad Mohamed, K., Ahmad, M. Z., and Asmawi, Z. (2012). The in vivo antihypertensive effects of standardized methanol extracts of Orthosiphon stamineus on spontaneous hypertensive rats: a preliminary study. Afr. J. Pharm. Pharmacol. 6, 376–379. doi:10.4103/jpbs.JPBS_253_17
Bai, J., Zhao, S., Fan, X., Chen, Y., Zou, X., Hu, M., et al. (2019). Inhibitory effects of flavonoids on P-glycoprotein in vitro and in vivo: food/Herb-drug interactions and structure-activity relationships. Toxicol. Appl. Pharmacol. 369, 49–59. doi:10.1016/j.taap.2019.02.010
Beaux, D., Fleurentin, J., and Mortier, F. (1999). Effect of extracts of Orthosiphon stamineus Benth, Hieracium pilosella L., Sambucus nigra L. and Arctostaphylos Uva-Ursi (L.) Spreng. In rats. Phytother Res. 13, 222–225. doi:10.1002/(SICI)1099-1573(199905)13:3<222::AID-PTR447>3.0.CO;2-P
Benavente-García, O., and Castillo, J. (2008). Update on uses and properties of Citrus flavonoids: new findings in anticancer, cardiovascular, and anti-inflammatory activity. J. Agric. Food Chem. 56, 6185–6205. doi:10.1021/jf8006568
Chae, H.-S., Kim, Y.-M., and Chin, Y.-W. (2017). Sinensetin inhibits interleukin-6 in human mast cell - 1 via signal transducers and Activators of the transcription 3 (Stat3) and nuclear factor Kappa B (Nf-Κb) pathways. Natural Product Sci. 23, 1–4. doi:10.20307/nps.2017.23.1.1
Choi, C. H., Kim, J. H., and Kim, S. H. (2004). Reversal of P-glycoprotein-mediated MDR by 5,7,3',4',5'-pentamethoxyflavone and SAR. Biochem. Biophys. Res. Commun. 320, 672–679. doi:10.1016/j.bbrc.2004.06.020
Choi, C. H., Sun, K. H., An, C. S., Yoo, J. C., Hahm, K. S., Lee, I. H., et al. (2002). Reversal of P-Glycoprotein-Mediated multidrug resistance by 5,6,7,3',4'-pentamethoxyflavone (sinensetin). Biochem. Biophys. Res. Commun. 295, 832–840. doi:10.1016/s0006-291x(02)00755-6
Choi, Y. H., and Yu, A. M. (2014). Abc transporters in multidrug resistance and pharmacokinetics, and Strategies for drug development. Curr. Pharmaceut. Des. 20, 793–807. doi:10.2174/138161282005140214165212
Damsud, T., Grace, M. H., Adisakwattana, S., and Phuwapraisirisan, P. (2014). Orthosiphol A from the aerial parts of Orthosiphon aristatus is putatively responsible for hypoglycemic effect via alpha-glucosidase inhibition. Nat Prod Commun. 9, 639–641. doi:10.1186/1472-6882-12-176
Danisman Kalindemirtas, F., Birman, H., Candoken, E., Bilgis Gazioglu, S., Melikoglu, G., and Kuruca, S. (2019). Cytotoxic effects of some flavonoids and imatinib on the K562 chronic myeloid leukemia cell line: data analysis using the Combination index method. Balkan Med. J. 36, 96–105. doi:10.4274/balkanmedj.galenos.2018.2017.1244
Du, Q., and Chen, H. (2010). The Methoxyflavones in Citrus reticulata Blanco Cv. Ponkan and their antiproliferative activity against cancer cells. Food Chem. 119, 567–572. doi:10.1016/j.foodchem.2009.06.059
Fromm, M. F., Kim, R. B., Stein, C. M., Wilkinson, G. R., and Roden, D. M. (1999). Inhibition of P-glycoprotein-mediated drug transport: a unifying mechanism to explain the interaction between digoxin and quinidine [seecomments]. Circulation. 99, 552–557. doi:10.1161/01.cir.99.4.552
Ho, S. K., Tan, C. P., Thoo, Y. Y., Abas, F., and Ho, C. W. (2014). Ultrasound-assisted extraction of antioxidants in misai kucing (Orthosiphon stamineus). Molecules. 19 (8), 12640–12659. doi:10.3390/molecules190812640
Hsu, C. L., Hong, B. H., Yu, Y. S., and Yen, G. C. (2010). Antioxidant and anti-inflammatory effects of Orthosiphon aristatus and its bioactive compounds. J. Agric. Food Chem. 58, 2150–2156. doi:10.1021/jf903557c
Ishimoto, T., Izumi, D., Miyamoto, Y., Sakamoto, Y., and Bab, H. (2015). Molecular insights into Colorectal cancer stem cell regulation by environmental factors. J. Cancer Metast. Treatm. 1, 156–162. doi:10.18632/oncotarget.23607
Iwase, Y., Takemura, Y., Ju-Ichi, M., Ito, C., Furukawa, H., Kawaii, S., et al. (2000). Inhibitory effect of flavonoids from citrus plants on epstein-barr virus activation and two-Stage Carcinogenesis of Skin tumors. Canc. Lett. 154, 101–105.
Jeong, J. M., Choi, C. H., Kang, S. K., Lee, I. H., Lee, J. Y., and Jung, H. (2007). Antioxidant and chemosensitizing effects of flavonoids with hydroxy and/or methoxy groups and structure-activity relationship. J. Pharm. Pharmaceut. Sci. 10, 537–546. doi:10.18433/j3kw2z
Kang, S. I., Shin, H. S., and Kim, S. J. (2015). Sinensetin enhances adipogenesis and lipolysis by increasing cyclic Adenosine Monophosphate levels in 3t3-L1 adipocytes. Biol. Pharm. Bull. 38, 552–558. doi:10.1248/bpb.b14-00700
Kang, S. I., Shin, H. S., Ko, H. C., and Kim, S. J. (2013). Effects of sinensetin on lipid metabolism in mature 3t3-L1 adipocytes. Phytother Res. 27, 131–134. doi:10.1002/ptr.4683
Kapadia, G. J., Azuine, M. A., Takayasu, J., Konoshima, T., Takasaki, M., Nishino, H., et al. (2000). Inhibition of epstein-barr virus early antigen activation promoted by 12-O-tetradecanoylphorbol-13-acetate by the non-steroidal anti-inflammatory drugs. Cancer Lett. 161, 221–229.
Kawahata, I., Yoshida, M., Sun, W., Nakajima, A., Lai, Y., Osaka, N., et al. (2013). Potent activity of nobiletin-rich Citrus reticulata peel extract to facilitate Camp/pka/erk/Creb signaling associated with learning and memory in cultured hippocampal neurons: identification of the Substances responsible for the pharmacological action. J. Neural. Transm. 120, 1397–1409. doi:10.1007/s00702-013-1025-x
Kawaii, S., Tomono, Y., Katase, E., Ogawa, K., and Yano, M. (1999). Antiproliferative activity of flavonoids on several cancer cell lines. Biosci. Biotechnol. Biochem. 63, 896–899. doi:10.1271/bbb.63.896
Kimura, J., Nemoto, K., Yokosuka, A., Mimaki, Y., Degawa, M., and Ohizumi, Y. (2013). 6-Demethoxynobiletin, a nobiletin-Analog citrus flavonoid, enhances extracellular signal-regulated kinase phosphorylation in Pc12d cells. Biol. Pharm. Bull. 36, 1646–1649. doi:10.1021/bi050643x
Laavola, M., Nieminen, R., Yam, M. F., Sadikun, A., Asmawi, M. Z., Basir, R., et al. (2012). Flavonoids eupatorin and sinensetin present in Orthosiphon stamineus leaves inhibit inflammatory gene expression and Stat1 activation. Planta Med. 78, 779–786. doi:10.1055/s-0031-1298458
Lam, I. K., Alex, D., Wang, Y. H., Liu, P., Liu, A. L., Du, G. H., et al. (2012). In Vitro and in Vivo structure and activity relationship analysis of polymethoxylated flavonoids: identifying sinensetin as a novel antiangiogenesis agent. Mol. Nutr. Food Res. 56, 945–956. doi:10.1002/mnfr.201100680
Leonard, G. D., Fojo, T., and Bates, S. E. (2003). The role of Abc transporters in clinical practice. Oncology. 8, 411–424. doi:10.1634/theoncologist.8-5-411
Lin, Y., Vermeer, M. A., Bos, W., Van Buren, L., Schuurbiers, E., Miret-Catalan, S., et al. (2011). Molecular structures of citrus flavonoids determine their effects on lipid metabolism in Hepg2 cells by primarily suppressing Apob secretion. J. Agric. Food Chem. 59, 4496–4503. doi:10.1021/jf1044475
Lyckander, I. M., and Malterud, K. E. (1996). Lipophilic flavonoids from Orthosiphon spicatus prevent oxidative inactivation of 15-Lipoxygenase. Prostaglandins Leukot. Essent. Fatty Acids. 54, 239–246. doi:10.1016/s0952-3278(96)90054-x
Malterud, K. E., Hanche-Olsen, I. M., and Smith-Kielland, I. (1989). Flavonoids from Orthosiphon spicatus. Planta Med. 55, 569–570. doi:10.1055/s-2006-962099
Malterud, K. E., and Rydland, K. M. (2000). Inhibitors of 15-Lipoxygenase from orange peel. J. Agric. Food Chem. 48, 5576–5580.
Manshor, N. M., Dewa, A., Asmawi, M. Z., Ismail, Z., Razali, N., and Hassan, Z. (2013). Vascular reactivity concerning Orthosiphon stamineus Benth-mediated antihypertensive in aortic rings of Spontaneously hypertensive rats. Int. J. Vasc. Med. 2013, 456852–456858. doi:10.1155/2013/456852
Manthey, J. A., and Grohmann, K. (2001). Phenols in citrus peel Byproducts. Concentrations of Hydroxycinnamates and polymethoxylated flavones in citrus peel Molasses. J. Agric. Food Chem. 49, 3268–3273. doi:10.1021/jf010011r
Manthey, J. A., and Guthrie, N. (2002). Antiproliferative activities of citrus flavonoids against six human cancer cell lines. J. Agric. Food Chem. 50, 5837–5843. doi:10.1021/jf020121d
Mertens-Talcott, S. U., De Castro, W. V., Manthey, J. A., Derendorf, H., and Butterweck, V. (2007). Polymethoxylated flavones and other phenolic derivates from citrus in their inhibitory effects on P-Glycoprotein-Mediated transport of talinolol in Caco-2 cells. J. Agric. Food Chem. 55, 2563–2568. doi:10.1021/jf063138v
Middleton, E., Drzewiecki, G., and Tatum, J. (1987). The effects of citrus flavonoids on human basophil and neutrophil function. Planta Med. 53, 325–328. doi:10.1055/s-2006-962728
Mo, W., and Zhang, J. T. (2012). Human Abcg2: structure, function, and its role in multidrug resistance. Int J Biochem Mol Biol. 3, 1–27. doi:10.20517/cdr.2018.02
Modlinger, P. S., and Welch, W. J. (2003). Adenosine A1 receptor antagonists and the kidney. Curr. Opin. Nephrol. Hypert. 12, 497–502. doi:10.1097/01.mnh.0000088726.87142.e3
Mohamed, E. A., Siddiqui, M. J., Ang, L. F., Sadikun, A., Chan, S. H., Tan, S. C., et al. (2012). Potent α-glucosidase and α-amylase inhibitory activities of standardized 50% ethanolic extracts and sinensetin from Orthosiphon stamineus Benth as anti-diabetic mechanism. BMC Compl. Alternat. Med. 12, 176–177. doi:10.1186/1472-6882-12-176
Mohamed, E. A., Yam, M. F., Ang, L. F., Mohamed, A. J., and Asmawi, M. Z. (2013). Antidiabetic properties and mechanism of action of Orthosiphon stamineus Benth bioactive sub-fraction in Streptozotocin-induced diabetic rats. J. Acupunct. Meridian Stud. 6, 31–40. doi:10.1016/j.jams.2013.01.005
Nagase, H., Omae, N., Omori, A., Nakagawasai, O., Tadano, T., Yokosuka, A., et al. (2005). Nobiletin and its related flavonoids with Cre-dependent transcription-stimulating and neuritegenic activities. Biochem. Biophys. Res. Commun. 337, 1330–1336. doi:10.1016/j.bbrc.2005.10.001
Nakanishi, M., Hino, M., Yoshimura, M., Amakura, Y., and Nomoto, H. (2019). Identification of sinensetin and nobiletin as major antitrypanosomal factors in a Citrus cultivar. Exp. Parasitol. 200, 24–29. doi:10.1016/j.exppara.2019.03.008
Nasri, M., Bedjou, F., Porras, D., and Martínez-Flórez, S. (2017). Antioxidant, anti-inflammatory, and analgesic activities of Citrus reticulata Blanco leaves extracts: an in vivo and in vitro study. Phytothérapie. 46, 1–13.
Natarajan, K., Xie, Y., Baer, M. R., and Ross, D. D. (2012). Role of breast cancer resistance protein (Bcrp/Abcg2) in cancer drug resistance. Biochem. Pharmacol. 83, 1084–1103. doi:10.1016/j.bcp.2012.01.002
Ohtani, H., Ikegawa, T., Honda, Y., Kohyama, N., Morimoto, S., Shoyama, Y., et al. (2007). Effects of various Methoxyflavones on vincristine uptake and multidrug resistance to vincristine in P-Gp-Overexpressing K562/Adm cells. Pharm. Res. (N. Y.). 24, 1936–1943. doi:10.1007/s11095-007-9320-6
Otoukesh, B., Boddouhi, B., Moghtadaei, M., Kaghazian, P., and Kaghazian, M. (2018). Novel molecular insights and new therapeutic Strategies in Osteosarcoma. Cancer Cell Int. 18, 158–160. doi:10.1186/s12935-018-0654-4
Panche, A. N., Diwan, A. D., and Chandra, S. R. (2016). Flavonoids: an overview. J. Nutr. Sci. 5, e47. doi:10.1017/jns.2016.41
Pereira, C. V., Duarte, M., Silva, P., Bento Da Silva, A., Duarte, C. M. M., Cifuentes, A., et al. (2019). Polymethoxylated flavones target cancer stemness and improve the antiproliferative effect of 5-fluorouracil in a 3d cell model of Colorectal cancer. Nutrients. 11. doi:10.3390/nu11020326
Qiu, Q., Shi, W., Zhao, S., Zhu, Y., Ding, Z., Zhou, S., et al. (2019). Discovery to Solve multidrug resistance: design, synthesis, and Biological evaluation of novel agents. Archiv der Pharmazie- Chem. Life Sci. 37, 28.
Sergeev, I. N., Li, S., Colby, J., Ho, C. T., and Dushenkov, S. (2006). Polymethoxylated flavones induce Ca(2+)-mediated apoptosis in breast cancer cells. Life Sci. 80, 245–253. doi:10.1016/j.lfs.2006.09.006
Sergeev, I. N., Li, S., Ho, C. T., Rawson, N. E., and Dushenkov, S. (2009). Polymethoxyflavones activate Ca2+-dependent apoptotic targets in adipocytes. J. Agric. Food Chem. 57, 5771–5776. doi:10.1021/jf901071k
Shin, H. S., Kang, S. I., Yoon, S. A., Ko, H. C., and Kim, S. J. (2012). Sinensetin attenuates LPS-induced inflammation by regulating the protein level of IκB-α. Biosci. Biotechnol. Biochem. 76, 847–849. doi:10.1271/bbb.110908
Summers, M. A., Moore, J. L., and Mcauley, J. W. (2004). Use of verapamil as a potential P-glycoprotein inhibitor in a patient with refractory epilepsy. Ann. Pharmacother. 38, 1631–1634. doi:10.1345/aph.1E068
Swaminathan, M., Chee, C. F., Chin, S. P., Buckle, M. J., Rahman, N. A., Doughty, S. W., et al. (2014). Flavonoids with M1 muscarinic Acetylcholine receptor binding activity. Molecules. 19, 8933–8948. doi:10.3390/molecules19078933
Tan, K. T., Lin, M. X., Lin, S. C., Tung, Y. T., Lin, S. H., and Lin, C. C. (2019). Sinensetin induces apoptosis and autophagy in the treatment of human T-cell lymphoma. Anti Canc. Drugs. 30, 485–494. doi:10.1097/CAD.0000000000000756
Tan, K. W., Li, Y., Paxton, J. W., Birch, N. P., and Scheepens, A. (2013). Identification of novel dietary phytochemicals inhibiting the efflux transporter breast cancer resistance protein (Bcrp/Abcg2). Food Chem. 138, 2267–2274. doi:10.1016/j.foodchem.2012.12.021
Tao, N., Jia, L., and Zhou, H. (2014). Anti-fungal activity of Citrus reticulata Blanco essential oil against Penicillium italicum and Penicillium digitatum. Food Chem. 153, 265–271. doi:10.1016/j.foodchem.2013.12.070
Tezuka, Y., Stampoulis, P., Banskota, A. H., Awale, S., Tran, K. Q., Saiki, I., et al. (2000). Constituents of the Vietnamese medicinal plant Orthosiphon stamineus. Chem. Pharm. Bull. 48, 1711–1719. doi:10.1248/cpb.48.1711
Tidefelt, U., Liliemark, J., Gruber, A., Liliemark, E., Sundman-Engberg, B., Juliusson, G., et al. (2000). P-glycoprotein inhibitor valspodar (psc 833) increases the intracellular concentrations of daunorubicin in Vivo in patients with P-Glycoprotein-Positive acute myeloid leukemia. J. Clin. Oncol. 18 (9), 1837–1844. doi:10.1200/JCO.2000.18.9.1837
Tofovic, S. P., Salah, E. M., Smits, G. J., Whalley, E. T., Ticho, B., Deykin, A., et al. (2016). Dual A1/A2b receptor blockade improves cardiac and renal outcomes in a rat model of heart Failure with preserved ejection fraction. J. Pharmacol. Exp. Therapeut. 356, 333–340. doi:10.1124/jpet.115.228841
Vikram, A., Jayaprakasha, G. K., Jesudhasan, P. R., Pillai, S. D., and Patil, B. S. (2010). Suppression of Bacterial cell-cell signalling, biofilm formation and type iii secretion system by citrus flavonoids. J. Appl. Microbiol. 109, 515–527. doi:10.1111/j.1365-2672.2010.04677.x
Vikram, A., Jayaprakasha, G. K., Uckoo, R. M., and Patil, B. S. (2013). Inhibition of Escherichia coli O157:H7 motility and biofilm by β-sitosterol glucoside. Biochim. Biophys. Acta. 1830, 5219–5228. doi:10.1016/j.bbagen.2013.07.022
Vu, T., and Datta, P. K. (2017). Regulation of emt in Colorectal cancer: a Culprit in Metastasis. Cancers. 9, 171. doi:10.3390/cancers9120171
Waghray, D., and Zhang, Q. (2018). Inhibit or evade multidrug resistance P-glycoprotein in cancer treatment. J. Med. Chem. 61, 5108–5121. doi:10.1021/acs.jmedchem.7b01457
Wang, Q., Yang, H., Miller, D. W., and Elmquist, W. F. (1995). Effect of the p-glycoprotein inhibitor, cyclosporin A, on the distribution of rhodamine-123 to the brain: an in vivo microdialysis study in freely moving rats. Biochem. Biophys. Res. Commun. 211, 719–726. doi:10.1006/bbrc.1995.1872
Welch, W. J. (2015). Adenosine, type 1 receptors: role in proximal Tubule Na+ reabsorption. Acta Physiol. 213, 242–248. doi:10.1111/apha.12413
Wesołowska, O., Molnar, J., Ocsovszki, I., and Michalak, K. (2009). Differential effect of phenothiazines on Mrp1 and P-glycoprotein activity. In Vivo. 23, 943–947. doi:10.3389/fpsyt.2012.00054
Wilkens, S. (2015). Structure and mechanism of Abc transporters. F1000Prime Rep. 7, 14. doi:10.12703/P7-14
Yam, M. F., Ang, L. F., Salman, I. M., Ameer, O. Z., Lim, V., Ong, L. M., et al. (2009). Orthosiphon stamineus leaf extract protects against ethanol-induced gastropathy in rats. J. Med. Food. 12, 1089–1097. doi:10.1089/jmf.2008.0005
Yam, M. F., Basir, R., Asmawi, M. Z., and Ismail, Z. (2007). Antioxidant and hepatoprotective effects of Orthosiphon stamineus Benth. standardized extract. Am. J. Chin. Med. 35, 115–126. doi:10.1142/S0192415X07004679
Yam, M. F., Tan, C. S., and Shibao, R. (2018). Vasorelaxant effect of sinensetin via the No/Sgc/Cgmp pathway and potassium and calcium channels. Hypertens. Res. 41, 787–797. doi:10.1038/s41440-018-0083-8
Yao, X., Xu, X., Fan, G., Qiao, Y., Cao, S., and Pan, S. (2009). Determination of synergistic effects of polymethoxylated flavone extracts of Jinchen orange peels (Citrus sinensis Osberk) with Amino acids and organic acids using Chemiluminescence. Eur. Food Res. Technol. 229, 743–750. doi:10.1007/s00217-009-1100-6
Youn, K., Yu, Y., Lee, J., Jeong, W. S., Ho, C. T., and Jun, M. (2017). Polymethoxyflavones: novel β-Secretase (BACE1) inhibitors from citrus peels. Nutrients. 9. doi:10.3390/nu9090973
Yuliana, N. D., Khatib, A., Link-Struensee, A. M., Ijzerman, A. P., Rungkat-Zakaria, F., Choi, Y. H., et al. (2009). Adenosine A1 receptor binding activity of methoxy flavonoids from Orthosiphon stamineus. Planta Med. 75, 132–136. doi:10.1055/s-0028-1088379
Keywords: sinensetin, pharmacological activities, mechanistic studies, anticancer, anti-inflammatory, lipolysis
Citation: Han Jie L, Jantan I, Yusoff SD, Jalil J and Husain K (2021) Sinensetin: An Insight on Its Pharmacological Activities, Mechanisms of Action and Toxicity. Front. Pharmacol. 11:553404. doi: 10.3389/fphar.2020.553404
Received: 18 April 2020; Accepted: 11 December 2020;
Published: 20 January 2021.
Edited by:
Francesca Borrelli, University of Naples Federico II, ItalyReviewed by:
Santa Cirmi, University of Messina, ItalyMariateresa Russo, Mediterranea University of Reggio Calabria, Italy
Copyright © 2021 Han Jie, Jantan, Yusoff, Jalil and Husain. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Khairana Husain, a2hhaXJhbmFAdWttLmVkdS5teQ==