 Hélène Grégoire1
Hélène Grégoire1 Loris Roncali1Audrey Rousseau1,2
Loris Roncali1Audrey Rousseau1,2 Michel Chérel3Yves Delneste1,4Pascale Jeannin1,4
Michel Chérel3Yves Delneste1,4Pascale Jeannin1,4 François Hindré1,5
François Hindré1,5 Emmanuel Garcion1,6*
Emmanuel Garcion1,6*- 1CRCINA, INSERM, Université de Nantes, Université d’Angers, Angers, France
- 2Département de Pathologie Cellulaire et Tissulaire, CHU Angers, Angers, France
- 3CRCINA, INSERM, Université d’Angers, Université de Nantes, Nantes, France
- 4Laboratoire d’Immunologie et Allergologie, CHU d’Angers, Angers, France
- 5PRIMEX, Plateforme de radiobiologie et d’imagerie expérimentale, SFR ICAT, Université d’Angers, Angers, France
- 6PACeM, Plateforme d’analyses cellulaires et moléculaires, SFR ICAT, Université d’Angers, Angers, France
Glioblastoma (GB) is the most common and devastating form of brain cancer. Despite conventional treatments, progression or recurrences are systematic. In recent years, immunotherapies have emerged as an effective treatment in a number of cancers, leaving the question of their usefulness also faced with the particular case of brain tumors. The challenge here is major not only because the brain is the seat of our consciousness but also because of its isolation by the blood-brain barrier and the presence of a unique microenvironment that constitutes the central nervous system (CNS) with very specific constituent or patrolling cells. Much of the microenvironment is made up of immune cells or inflammation. Among these, tumor-associated macrophages (TAMs) are of significant interest as they are often involved in facilitating tumor progression as well as the development of resistance to standard therapies. In this review, the ubiquity of TAMs in GB will be discussed while the specific case of microglia resident in the brain will be also emphasized. In addition, the roles of TAMs as accomplices in the progression of GB and resistance to treatment will be presented. Finally, clinical trials targeting TAMs as a means of treating cancer will be discussed.
Introduction
Glioblastoma (GB) is the most frequent and malignant form of brain tumors. It is associated with a poor prognosis and the median overall survival of GB patients is about 15 months after standard of care (Stupp et al., 2009). Conventional treatments consist of maximal safe resection followed by external radiotherapy and concomitant chemotherapy based on the use of the alkylating agent temozolomide (TMZ) (Stupp et al., 2005). However, recurrence inevitably occurs. Currently, no therapy can completely cure GB; current treatments can only marginally improve the overall survival of patients. The current strategy focuses mostly on targeting the tumor cells, failing to account for other cellular constituents present in the tumor. Hence, to cure and achieve a complete resection of GB tumors, new therapeutic strategies are in great demand.
GB is a highly heterogeneous tumor, with diverse co-existing cell types that include tumor cells, endothelial cells, fibroblasts and different cell types from the immune system (Charles et al., 2011; Quail and Joyce, 2017). A particular emphasis has been placed on the immune system and especially on tumor-associated macrophages (TAMs) as they are the dominant infiltrating immune cell population in GB. These cells interact with tumor cells to promote tumor growth and progression (Feng et al., 2015). The host defense is composed of both innate and adaptative immune cells and they are both involved in cancer immune surveillance in early stages of the disease. However, the tumor is able to escape this immune surveillance during its development. At that point, the tumor can recruit immune cells and change their original function to be one of its accomplices (Brown et al., 2018; Finn, 2018). Tumor cells can inhibit the cytotoxic function of the immune system by secreting immunosuppressive factors or recruiting immunosuppressive inflammatory cells. In relation to this, macrophages appear to be a promising target to improve the effectiveness of actual therapy as more and more information on their physiological and pathological roles in the brain is being uncovered.
Macrophages are the most abundant infiltrating immune cells in GB. Their function is different from their homolog in healthy tissues (Nishie et al., 1999; Hussain et al., 2006). They are able to discriminate the components of the self from the non-self (microbes) but also the altered components of the self. When recognizing the non-self or altered self-components, they can begin their process of elimination. Macrophages located in the tumor microenvironment are called tumor-associated macrophages. Under normal physiological conditions, macrophages are implicated in different processes such as organ development, tissue homeostasis, host defense against infections. These cells can also participate in metabolic disorders, immune diseases and cancer development (Sica et al., 2015). Normally, the myeloid population is the major player of the innate immune system and represents up to 30% of the tumor mass (Rossi et al., 1987; Graeber et al., 2002). Both the activation status and the number of TAMs present in the tumor microenvironment seem to influence GB prognosis (Komohara et al., 2008; Lu-Emerson et al., 2013; Pyonteck et al., 2013).
Macrophages are characterized by their plasticity and heterogeneity. They can be activated by different types of stimuli (growth factors, cytokines, microbial products, nucleotides) which in turn will affect macrophages differently (Poh and Ernst, 2018). In vitro, the stimulation of macrophages by interferon-γ (IFN–γ) and/or lipopolysaccharides (LPS) induces the classical (M1) macrophage polarization (Nielsen and Schmid, 2017). M1 macrophages favor the generation of T helper Type 1 (Th1) lymphocytes. Classically activated macrophages are good effectors to fight malignant tumors and are associated with chronic inflammation (Atri et al., 2018). Those macrophages are characterized by a high expression of IL-12, IL-23, and a low expression of IL-10. They can also produce high levels of pro-inflammatory cytokines IL-1β, tumor necrosis factor α (TNF-α), and IL-6, and increase the expression of inducible nitric oxide synthase (iNOS, NOSII) and reactive oxygen species (ROS). Another known stimulus for M1 macrophages is GM-CSF (Granulocyte Macrophage Colony-Stimulating Factor). It activates STAT5, which leads to the activation of the PI3K-AKT pathway (Jeannin et al., 2018).
On the contrary, macrophages stimulated in vitro by IL-4 and/or IL-13 are called alternatively activated (M2) macrophages (Murray et al., 2014). They are known effectors for promoting Th2 lymphocytes. They are involved in angiogenesis and tumor progression (Martinez and Gordon, 2014). This phenotype is associated with a low expression of IL-12, IL-23, and a high expression of IL-10 and TGF-β. Furthermore, M2 macrophages also have high levels of arginase 1 (Arg1), mannose receptors and scavenger receptors. M-CSF (Macrophage Colony-Stimulating Factor) and IL-34 also induce a M2 phenotype. M-CSF and IL-34 express the same receptor named CD115 and activate the MAP kinases signaling pathway (Jeannin et al., 2018).
Although the traditional M1/M2 dichotomy is useful for understanding the functionality of TAMs, recent analyzes, in particular of single-cell, revealed a spectrum of activation states much more complex than these traditional polarizations (Locati et al., 2020). Hence, macrophages in cancer are double-edged swords exerting pro- and antitumor functions. More than a real opposition, the M1/M2 signature crystallize a continuum of two extremes capable of specific adaptations (eg., chromatin remodeling, epigenetic marks, trained immunity, metabolic reprogramming,…) to various loco-regional cues (eg., cytokines, chemokines, miRNA, or immune checkpoints). In addition, proliferating monocytes could persist in a state of self-renewal within tumor tissues, rather than immediately differentiate into macrophages indicating a much higher complexity (Lin et al., 2019). It should again be emphasized that the M1 and M2 markers are distinct across species and in particular between humans and mice (eg., in human NOSII and Arg1 do not account for M1 and M2 macrophages, respectively) (Thomas and Mattila, 2014). In this regard, there are no specific surface markers in humans except a privileged panel of produced cytokines.
TAMs that are described in the tumor have in most cases pro-tumorigenic functions that promote tumor growth, invasion, angiogenesis, and tumor metastasis. In the GB microenvironment, both TAMs derive from blood monocytes; some originate from resident macrophages called microglia. Hence, macrophages appear to be an attractive target for new therapeutic strategies (Noy and Pollard, 2014).
The goal of this review is to discuss whether macrophages are worth considering as therapeutic targets in GB and to summarize the existing drugs targeting macrophages. In the second part of this review, the presence of microglia in brain tumor will be discussed. Then, the roles of TAMs in regulating the tumor development, progression, and the response to conventional therapy will be reviewed. Finally, a survey of clinical trials testing drugs against macrophages in cancer will be presented.
The Presence of TAMs in GB: Reality or Not?
The World Health Organization (WHO) classification of Central Nervous System (CNS) tumors was restructured in 2016. Diagnoses are based on both molecular alterations and histopathologic features (integrated diagnosis) in contrast to the 2007 WHO classification that only included histopathologic features (Louis et al., 2007; Louis et al., 2016). The tumor is essentially defined by the characteristics of the tumor cells that compose it, independently of the ecosystem in which they evolve and which they could themselves modify. GB also consists of many different noncancerous cells. The following cells are known to define the tumor microenvironment: endothelial cells, pericytes, fibroblasts, and immune cells in addition to cancer cells (Quail and Joyce, 2013).
The tumor microenvironment is now emerging as an important regulator of cancer progression (Quail and Joyce, 2017). Data from the literature seem to suggest that distinct molecular profiles in GB are correlated with differences in their microenvironment (Zhernakova et al., 2018). Even if the WHO classification now includes molecular data, no information on the tumor microenvironment has been integrated so far. Despite the fact that a solid tumor has never been seen without infiltrating immune cells, current diagnostic guidelines often forget voluntarily to take this into account. Although this does not necessarily modify the diagnosis as it is perceived today, it could be useful as regards the consideration of patient management and escape or not to new well identified therapies. The presence of TAMs has already been well described in GB (Saha et al., 2017; Séhédic et al., 2017; Roesch et al., 2018). In a mouse model, TAMs were observed in perivascular areas in the tumor and seem to be implicated in gliomagenesis Feng et al., 2015. Interestingly, their localization in the tumor appears to depend on their phenotypes Schiffer et al., 2018. In 2012, a meta-analysis showed that a high density of TAMs appeared to be associated with a poor prognosis in head and neck, ovarian and breast cancer and with a better prognosis in colorectal cancer (Zhang et al., 2012; Yuan et al., 2017; Zhao et al., 2017). Further evidence revealed that human GB display a mixed population of M1/M2 macrophages, and the ratio M1:M2 correlated with survival in IDH1 R132H wild type GB (Zeiner et al., 2018). In high-grade gliomas, M2 macrophages were correlated with an unfavorable prognostic (Sørensen et al., 2018). Caponegro et al. also described a correlation between the presence of TAMs and a poorest prognosis in GB (Caponegro et al., 2018). Furthermore, a study based on magnetic resonance imaging in GB showed that highly aggressive tumors were also correlated with the presence of TAMs (Zhou et al., 2018). Taking into account these findings, the presence of TAMs in GB has been well proven. Macrophages are important for the progression of GB and assessing them may give more information on the prognosis.
Microglia: The Resident Macrophages of the CNS
Microglia are the resident macrophages of the CNS and a healthy CNS macrophage population consists only of resident microglia. The blood brain barrier is impaired in neuropathological diseases, thus allowing an infiltration of monocytes form peripheral blood. In GB, both resident microglia and peripheral macrophages can be detected (Lisi et al., 2017). It is crucial to understand their molecular differences and their specific roles in the tumor. Resident microglia and newly recruited macrophages, hereafter referred to as peripheral macrophages have a distinct origin, as microglia arise from the yolk sac primitive macrophages (Ginhoux et al., 2013; Ginhoux and Guilliams, 2016). Although their origin differs, they share common histologic characteristics. Differentiating between microglia and peripheral macrophages is a difficult task, since they share common surface markers. The name TAM may very well include both resident microglia and monocyte-derived macrophages (Szulzewsky et al., 2015; Kloepper et al., 2016). In order to separate macrophages of hematopoietic origin from resident microglia, CD45 was used in flow cytometry analysis (Badie et al., 2000). However, resident microglia can upregulate their CD45 expression, making them indistinguishable from peripheral macrophages (Müller et al., 2015). Using a genetically engineered mouse, it was demonstrated that peripheral macrophages represent the majority of TAMs in the tumor, and resident microglia form a minor TAM population (Chen et al., 2017). Moreover, resident microglia and peripheral macrophages have different preferential localizations. Peripheral macrophages mostly appear in perivascular areas while resident macrophages are usually located in the peritumoral zone. A recent study showed that only a small batch of common genes toward species (rat, mice, human) differentiates GB-induced polarization of resident microglia (Walentynowicz et al., 2018). Although many studies tried to decipher the origin of TAMs in the tumor, no clear answer has yet been obtained.
Resident microglia are described to be involved in many processes including tumor growth and progression (Bryukhovetskiy et al., 2016; Matias et al., 2018). Microglia were shown to contribute to the invasiveness of GB by upregulating serpin family A member 3 (SERPINA3) expression in GB stem cells (GSCs), that is implicated in the remodeling of the extracellular matrix (Li et al., 2018). Resident microglia were also shown to mediate GB progression and stemness through the activation of interferon regulatory factor 7 (IRF7) that generates an inflammatory environment (Li Z. et al., 2017). Resident microglia are also involved in antitumor immunity processes through the expression of toll-like receptor 2 (TLR2) that down regulates their major histocompatibility complex class II (MHCII) expression (Qian et al., 2018). In a murine model, enhancer of zeste homolog 2 (EZH2) expression in GB was shown to be involved in the polarization of TAMs toward the M2 phenotype, creating an immune deficient environment (Yin et al., 2017). A 6 cytokine-related gene signature in resident microglia was shown to be sufficient to predict survival and identify M2 cells in GB (Cai et al., 2015). Both resident and peripheral macrophages are uniquely involved in supporting GB growth and progression. Hence, if we wish to target TAMs as a mean to treat GB, we must first characterize this population as peripheral macrophages and/or resident microglia and counter their exact roles in GB initiation and maintenance.
Tumor-Associated Macrophages: A Partner in Crime for Tumor Cells
A tumor can influence its microenvironment, and inversely. Thus, the interactions between the tumor cells and the nearby non-tumor cells are crucial to promote tumor angiogenesis, peripheral immune tolerance, and tumor growth. As previously said, TAMs are highly represented inside the tumor microenvironment. They are known for their heterogeneous phenotype, which by simplification can be with either anti-tumor (M1-like) or pro-tumor functions (M2-like). As TAMs are highly plastic cells, they can program themselves into both subpopulations. This gives them the ability to have different functions in different tumor areas and at different times during the tumor development.
Biology of the Tumor
Tumor Cells
The effect of TAMs on tumor cells is dependent on their type of activation. The reprogrammed M1 TAMs suppress the growth of GB cells (Li T. et al., 2017) meanwhile the M2 macrophages are described to favor tumor growth and resistance to therapy (Xue et al., 2017).
A macrophage with pro-tumor function in the tumor microenvironment is a macrophage that enhances tumor initiation and growth. TAMs and tumor cells actively communicate with each other leading to tumor progression. Their communication is mediated by interleukins IL-6 and IL-10 and transforming growth factor-β1 (TGF-β1) (Wagner et al., 1999; Ye et al., 2012). These cytokines activate signaling pathways in the tumor cells that boost processes such as proliferation, invasion and vascularization (Figure 1). TGF-β1 secretion by TAMs is responsible for the recruitment of cancer stem-like cells (CSCs) expressing CD133. Another consequence of TGF-β1 secretion is the production of metalloproteinase 9 (MMP-9) by CSCs rendering them highly invasive (Ye et al., 2012). TAMs are able to secrete pleiotrophin (PTN); CSCs express the PTN receptor PTPRZ1 on their cell surface. Once PTN is recognized by its receptor, it stimulates CSCs maintenance and tumorigenic potential, and therefore promotes GB growth (Shi et al., 2017). PTN- expressing TAMs also express CD163 which is an M2 lineage marker. Wang et al. showed that macrophages support GB invasiveness through the CCL4-CCR5 axis that enhances MMP-9 expression (Wang et al., 2016). Hypoxia was also shown to positively contribute to this mechanism by enhancing CCL4 and CCR5 expression. An increase of TAMs in a mouse model was shown to decrease the survival of the mice associated with a reduction of CD8+ T cells (Chae et al., 2015). On top of that, EGFR activation level correlates with TAM infiltration. Consequently, EGF can induce an upregulation of vascular cell adhesion molecule-1 (VCAM-1) that favors the interaction between TAMs and tumor cells, which in turn promoted tumor cell invasion (Zheng et al., 2013). MerTK (Myeloid-Epithelial-Reproductive Tyrosine Kinase) is a tyrosine kinase expressed by macrophages that suppresses the innate immune response. Its expression was shown to be higher in tumor recurrences. TAMs that express MerTK are also associated with tumor growth and resistance to treatment, making MerTK a potential therapeutic target (Wu et al., 2018). The molecular crosstalk between tumor cells and macrophages appears to be important for tumor growth and malignant progression. Therefore, modulating the exchange between those two cell populations may be therapeutically relevant.
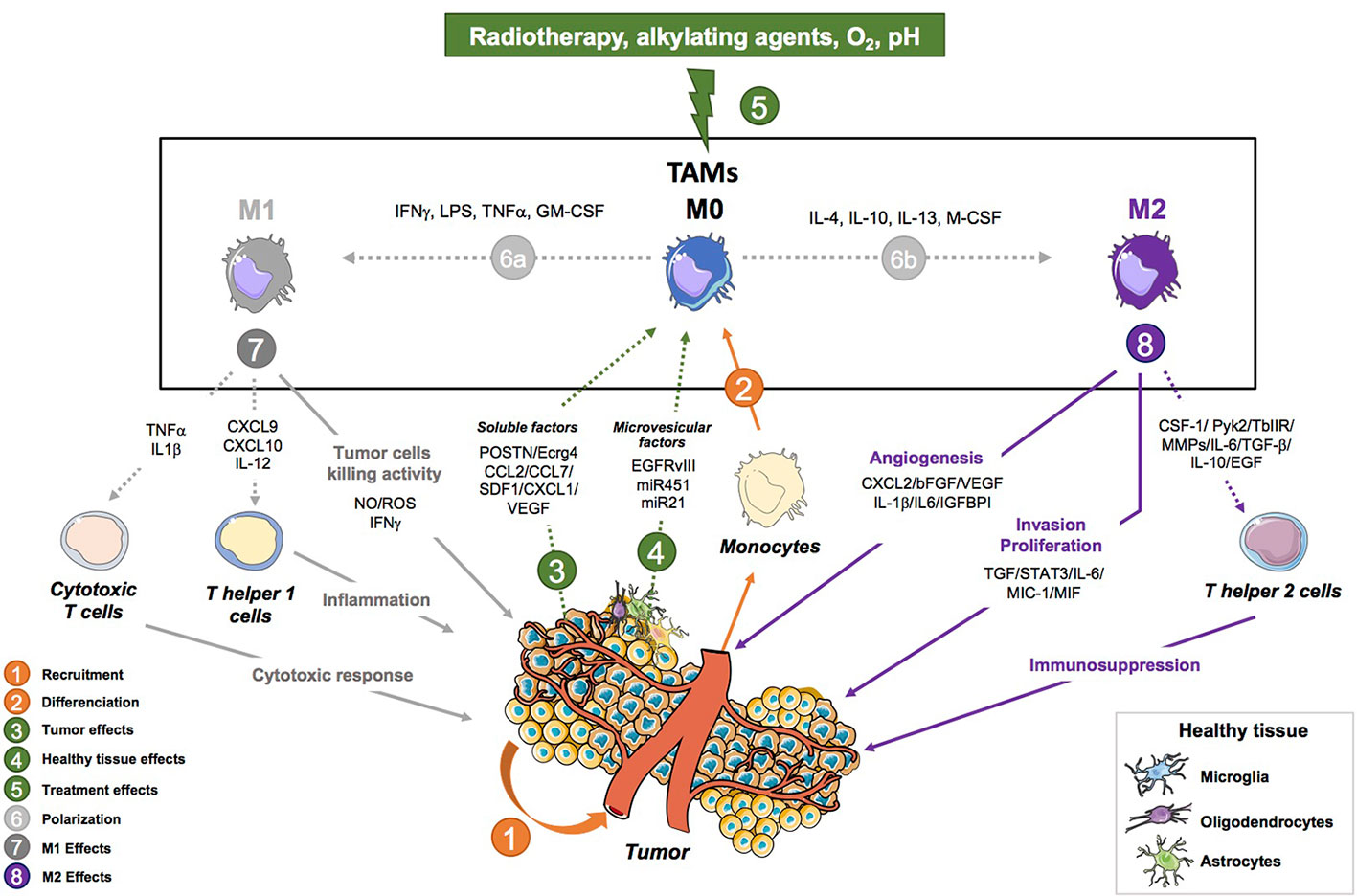
Figure 1 Tumor-associated macrophage activities in glioblastoma progression. This figure shows the pro-tumoral (angiogenesis, invasion, proliferation and immunosuppressive properties) and anti-tumor (Tumor cell killing, Th1 response and anti-tumor activity) activities of tumor-associated macrophages (TAMs) in brain tumors. (1) Monocytes are recruited to the tumor where they differentiate into macrophages. The tumor is involved in their programming as it sends different signals to induce a specific phenotype in favor of the tumor. (2) TAMs that are recruited can either polarize into a continuum of macrophage states that are described with two extremes: an M1 (2a) or an M2 (2b) phenotype depending on the signal they receive (IFNγ/LPS/GM-CSF for M1 and IL-4/IL-13/M-CSF for M2) Pyonteck et al., 2013; Kast et al., 2017; Roesch et al., 2018. (3) M1-like TAMs are macrophages with anti-tumor properties such as tumor cell kill abilities mediated by the production of NO, ROS, IFNγ Kennedy et al., 2013; Leblond et al., 2017. They also mediate the Th1 response in the tumor through the activation of Th helper cells by secreting CXCL9, CXCL10, IL-12 Poon et al., 2017. Finally, they also display an anti-tumor activity by activating cytotoxic T cells via TNFα and IL1β. (4) M2-like TAMs have pro tumoral properties such as enhancing the invasive and proliferative ability of GB cells by secreting CSF-1, MMPs, Pyk2, TGFβIIR, TGFβ, IL-6, IL-10, and EGF. They can also mediate the immunosuppressive environment through the expression of IL-6, MIC-1, MIF, STAT3, and TGFβ. Finally, TAMs also regulate angiogenesis through the following factors: IL-6, MIC-1, MIF, STAT3, and TGFβ. (5) The tumor controls the polarization of TAMs through the production of soluble factors (CCL2/CCL7/SDF-1/CX3CL1/VEGF/POSTN/Ecrg4) Feng et al., 2015; Hambardzumyan et al., 2015; Lee et al., 2015; Zhou et al., 2015; Chang et al., 2016; Chen and Hambardzumyan, 2018; Turkowski et al., 2018 and microvesicle factors (EGFRvIII, miR451, miR21) Van Der Vos et al., 2016; Manda et al., 2018. (6) The tumor is also able to send signals to recruit new peripherical macrophages. (7) Environmental cues including radiotherapy, chemotherapy, O2 level, pH are involved in the programing and functions of macrophages Hardee et al., 2012. (8) Healthy brain cells and TAMs probably interact and are involved in the programming of TAMs. Their interaction has yet to be studied. CCL2, C-C motif chemokine ligand 2; CCL7, C-C motif chemokine ligand 7; CSF-1, colony stimulating factor 1; CXCL2, C-X3-C motif chemokine ligand 2; CX3CL1, C-X3- C motif chemokine ligand 1; Ecrg4, esophageal cancer-related gene 4; EGF, endothelial growth fact; IGFBP1, insulin-like growth factor-binding protein 1; IL-1β, interleukin-1 beta; IL-10, interleukin-10; IL-6, interleukin-6; MIC-1, macrophage inhibitory cytokine 1; MIF, macrophage migration inhibitory factor; MMPs, matrix metalloproteinases; POSTN, periostin; Pyk2, proline rich tyrosine kinase 2; SDF-1, stromal cell-derived factor 1; STAT3, signal transducer and activator of transcription3; TGF-β, transforming growth factor-beta; TGFβIIR, TGF-beta type II receptor; VEGF, vascular endothelial growth factor; βFGF, basic fibroblast growth factor.
Angiogenesis
GB is a highly hypoxic tumor with prominent necrotic regions due to the rapid proliferation of GB cells. The cell composition of the tumor core is quite different from that of the peritumoral area. The tumor core is more hypoxic, contains more CD163+ TAMs and has a higher expression of VEGF-A (Tamura et al., 2018) (a major factor for vascularization). A downstream effect of hypoxia and necrosis is an increase in vascular proliferation. In the tumor microenvironment, TAMs are located near blood vessels. In mice, endothelial cells produce IL-6 that induces the expression of Arg1 and thus the alternative phenotype in TAMs (Wang et al., 2018). This alternative activation is mediated by the hypoxia-inducible factor-2α (HIF-2α). Wang et al. targeted IL-6 expression in a mouse model and improved the survival of GB-bearing mice. VEGF was shown to be implicated in promoting pro-angiogenic functions of TAMs in a GB rodent model (Turkowski et al., 2018). Gliomas overexpressing VEGF were correlated with an increase in the expression of MHCI and MHCII on macrophages. Endothelial cells and TAMs interaction leads to angiogenesis through the expression of TGF-β1 and integrin αvβ3, which induces the activation of the SRC-PI3K-YAP signaling (Cui et al., 2018) (Figure 1). The pro-angiogenic properties of TAMs are mediated by the protein CRCR1. This protein activates the PDGFB–PDGFRβ pathways and promotes pericytes recruitment, migration, and tumor angiogenesis (Zhu C. et al., 2017). In sum, TAMs have a proangiogenic function in GB. Thus, targeting macrophages may improve the response to anti-angiogenic therapies (Deng et al., 2017; Gagner et al., 2017). Indeed, blocking the macrophages recruitment by combining the chemokine SDF-1 and VEGF inhibitors was more effective and decreased tumor invasiveness and vascular density.
Immune Environment
Each tumor is characterized by an immune suppressive environment that forms one hallmark of cancer (Hanahan et al., 2011). This is in part due to the presence of TAMs in tumors but also to a complex regulation of the expression of immune and inflammatory genes by the global tumor ecosystem. It was found that IKKβ levels were reduced in GB; consequently, the NF-κB expression was decreased leading to defective immune and inflammatory gene expression in macrophages (Mieczkowski et al., 2015). NF-κB signaling is required for macrophage polarization and immune suppression in GB, making NF-κB a suitable target to improve overall survival in GB (Achyut et al., 2017). TAMs strongly inhibit the proliferation of antitumor T cells in the tumor microenvironment (Kumar et al., 2017). It was shown that an inhibition of transcription factors such as NF-κB, a mediator of M2 macrophages polarization, led to slower tumor growth and prolonged survival in a mouse model. It also decreased T cell induction which made the tumor less immunosuppressive (Barberi et al., 2018). Targeting NF-κB may improve the effectiveness of the current standard therapies.
TAMs express IL-4Rα that promotes immunosuppression. In mice, they also express Arg1 that is critical for T cell inhibition (Kohanbash et al., 2013). Chemokine ligand 22 (CCL22) is produced by TAMs and its expression is associated with a low survival rate and CD4+ T cell activation (Zhou et al., 2015). One of the key regulators of the immunosuppressive environment in GB is fibrinogen-like protein 2 (FGL2). Its expression was correlated with a higher number of CD4+ T cells and M2 macrophages (Latha et al., 2018). The colony stimulating factor receptor (CSF1R) is required for the recruitment of TAMs in the tumor microenvironment. It is also involved in promoting the polarization of macrophages toward the M2 phenotype. Inhibition of CSF1R attenuates the recruitment of TAMs and also increases the CD8+ T cell infiltration (Strachan et al., 2013) (Figure 1). Another regulator of the immune microenvironment is the receptor tyrosine kinase AXL that is expressed in TAMs (Sadahiro et al., 2018). Its inhibition in a GB mouse model was associated with prolonged survival. Furthermore, myeloid derived suppressor cells (MDSC) such as TAMs have been described to be activated by GB CSCs through MIF expression, having then an immunosuppressive activity on CD8+ T cells, notably through the Arg1 expression in mice models (Flavahan et al., 2016). Overall, targeting TAMs may disturb the immunosuppressive environment of the tumor, allowing the immune cells to function more effectively.
Loco-Regional Cues for Metabolic Reprogramming
A peculiarity of GB is that it affects the seat of our consciousness, the CNS, whose immune status remains privileged due notably to the presence of the blood-brain barrier (BBB) and of unique resident cells (microglia, astrocytes, endothelial cells) (cf. Box 1). Although a precise control of the inflammatory or immune infiltrate is realized, the physiological and anatomical characteristics of the CNS is fed by the field of new recent knowledge, such as the identification of direct vascular channels connecting skull bone marrow to the brain surface enabling myeloid cell migration (Herisson et al., 2018), and make evolve our representation of its immune status. It should be stressed, however, that depending on the therapeutic strategy envisaged, the drug used can have a distinct impact when used according to a peripheral or loco-regional mode of administration (cf. Tables 1–3). Hence, if TAMs influence immune and adaptive signaling, reciprocally, loco-regional metabolic signals produced in tumor environments (glucose, glutamine, cystéine, lactate, IDO, adenosine, itaconic acid, acidic pH) impacted the polarization fate and immunosuppressive functions of TAMs, thus possibly resulting in immune tolerance and treatment resistance in GB (for review, see Won et al., 2019). Hence, tolerance can be reversed at both the promoters and enhancers of tolerized genes involved in metabolism and lipid biosynthesis, leading to transcriptional programs that rewired the intracellular signaling of innate immune cells thus increasing the capability of macrophages to respond to stimulation (for review see, Locati et al., 2020). In line with this, it has been observed that inhibition of fatty acid synthase (FAS), which catalyzes the synthesis of long-chain fatty acids, prevents the pro-inflammatory response in macrophages (Carroll et al., 2018). Interestingly, using metabolic profiling, it was found that exposure to β-amyloid triggers acute reactive microglial inflammation accompanied by metabolic reprogramming from oxidative phosphorylation to glycolysis while metabolic boosting with recombinant interferon-γ treatment reversed the defective glycolytic metabolism and inflammatory functions of microglia (Baik et al., 2019). Such microglial metabolic switch may also have a strong impact on GB development.
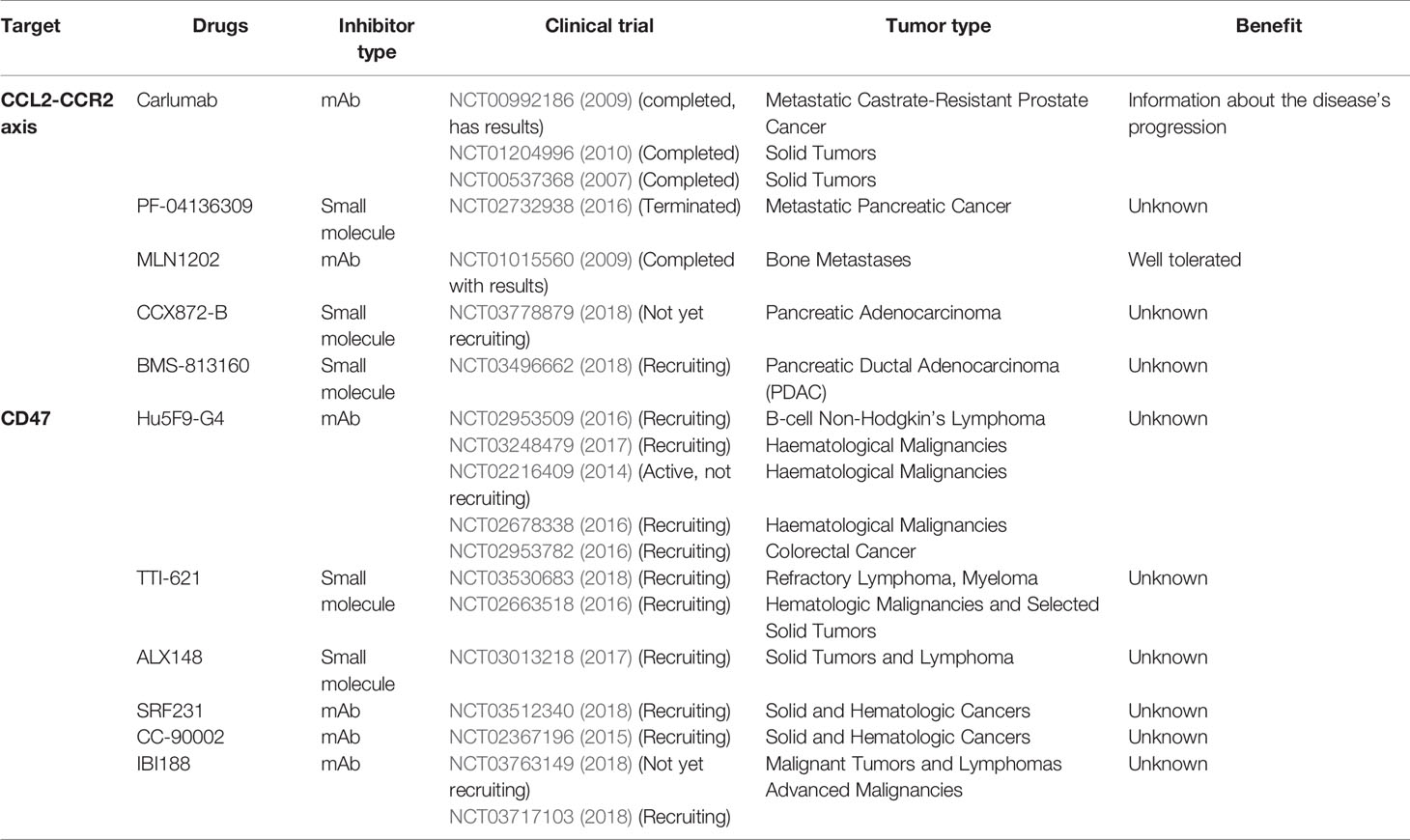
Table 1 Clinical trials targeting the recruitment of macrophages.

Table 2 Clinical trials with toll-like receptor (TLR) agonists for macrophages reprogramming.
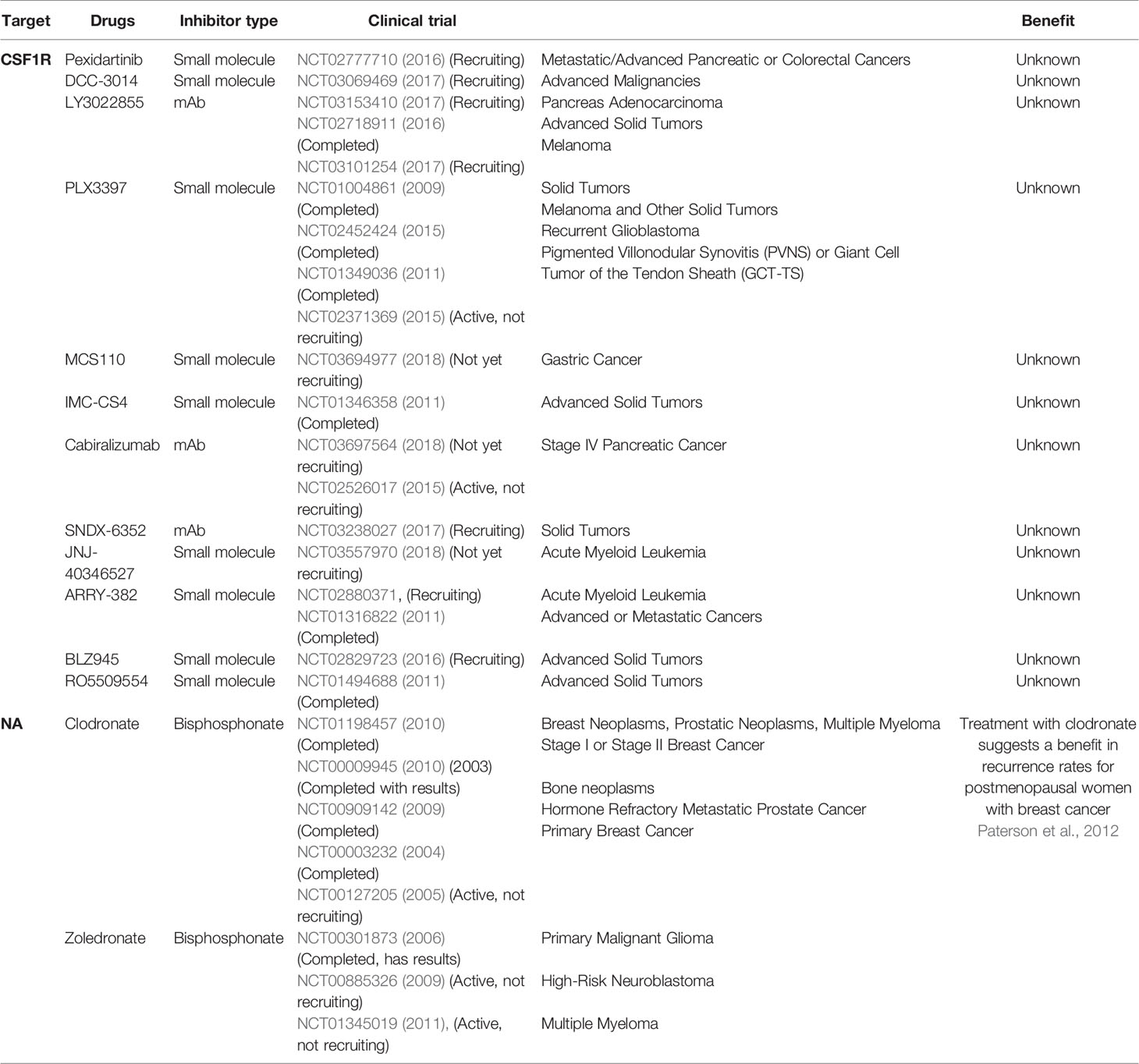
Table 3 Clinical trials using drugs to deplete macrophages from the tumor’s microenvironment.
Box 1. Non-cancerous brain cells alter macrophages polarization and functions.
Tumor cells cooperate with its surroundings such as the tumor microenvironment. The brain is also the home of specific cell types with their own characteristics and functions; although those cells are not part of the tumor, they can also interact with it. The interaction between cells residing in the brain and TAMs are very poorly understood in cancer but has been studied in depth in other pathologies, which will be quickly reviewed in this box. Both neurons and astrocytes can produce CX3CL1R, the receptor for CX3CL1 found on microglia Matias et al., 2018. CX3CL1 promotes TAM recruitment and increases the expression of MMPs and thus invasive properties. When an ischaemic stroke happens, ischaemic neurons are able to prime microglia toward an M1 phenotype during an injury Hu et al., 2012. Another cell type is oligodendrocyte which accounts for the formation of the myelin sheath in the CNS. It was found that macrophages and oligodendrocyte progenitor cells colocalized near the tumor border. At this site of colocalization, those cells induced stemness and resistance to therapy in GB cells Hide et al., 2018. In the peripheral nervous system, Schwann cells are the cells responsible for myelin sheath formation. Schwann cells were shown to promote cancer invasion by direct contact with tumor cells Deborde et al., 2016. The mechanism involved in this process remains unclear. In neurofibromas (peripheral nerve sheath tumors due to NF1 loss in Schwann cells), macrophages were shown to be abundant Stratton et al., 2018. In this case, Schwann cells and macrophages communicate with each other and are involved in the regulation of inflammatory gene expression. As Schwann cells and oligodendrocytes share a common function in normal tissue, it may be interesting to further study the involvement of oligodendrocytes in GB. Non-cancerous cells of the CNS and peripheral nervous system interact with macrophages and lead them to polarize toward a specific phenotype.
TAMs and Therapeutics
TAMs and Surgical Resection
Surgical resection is the current standard treatment for GB. However, limited data on the biological consequences of surgical resection have been published so far. It was reported that surgical resection increases proliferation and angiogenesis (Kong et al., 2010). After surgical resection, TAMs were shown to express higher levels of CD163, a M2 macrophage marker, and their localization was close to the site of recurrence (Zhu H. et al., 2017). Both TAMs and oligodendrocyte progenitor cells are localized near the tumor periphery. They enhance the stemness and chemo-radioresistance in GB cells (Hide et al., 2018). It was shown that tumor phenotypes associated with telomerase overexpression and TAMs infiltration were more complicated to resect, probably due to improvement of GB cell migratory capabilities (Hung et al., 2016). The inability to surgically remove the whole tumor contributes to the poor prognosis and recurrence of GB.
TAMs and Radiotherapy
Macrophages inside the tumor mass are involved in multiple phenomena that include radiation resistance. Radiation therapy itself induces changes in the tumor microenvironment and renders the tumor more aggressive. In fact, recurrence mostly appears near the irradiated area (Gupta and Burns, 2018). Radiotherapy induces a rapid inflammatory response leading to TAMs recruitment. This inflammatory response is correlated with a short survival time (Tabatabaei et al., 2017). TAMs participate in the induction of GB cell differentiation to a mesenchymal state through NF-κB production, an event that correlated with radiation resistance (Bhat et al., 2013). Recently, Leblond et al. showed that M1 macrophages are more sensitive to radiation than M2 macrophages (Leblond et al., 2017). The proportion of M2 macrophages in irradiated tissues is thus increased. Moreover, M2 macrophages were described to contribute to relapses in oral cancer by promoting vascularization after radiation treatment (Okubo et al., 2016). In a radioresistant GB model, the total RNA was sequenced and it was found that there was a positive regulation of macrophage chemotaxis following radiation (Doan et al., 2018). Also, in a murine glioma model, an increase in SDF-1α at the tumor invasion front after radiotherapy was correlated with the recruitment of TAMs and radioresistance (Wang et al., 2013). Irradiation of the tumor leads to the alteration of multiple pathways. In particular, it modifies the macrophage activation type, rendering them more supportive of tumor growth.
TAMs and Chemotherapy
The standard treatment of GB affects the molecular profiles of the tumor. Temozolomide (TMZ) is commonly used to treat GB. TAMs that express CD74 were described to be involved in TMZ resistance by inducing AKT and Erk1/2 activation in tumor cells (Kitange et al., 2010). Gene expression profiling showed that the tumor that recurred after treatment did not match the primary treatment-naïve tumor. After treatment, the polarization toward the M2 phenotype was upregulated (Hudson et al., 2018). Tumor protein 53 (p53) is involved in promoting the development of the tumor. GB with the p53 isoform Δ133p53β had increased CD163+ macrophages (Kazantseva et al., 2018). Moreover, Δ133p53β supports cancer stemness (Arsic et al., 2015). In addition, it is correlated with resistance to TMZ (Kazantseva et al., 2018). GB is able to evade the toxic effects of chemotherapy, but it can equally evade the action of the immune system. Hence, a cocktail of multiple drugs targeting different pathways may provide the most effective therapy for GB and improve overall survival.
Current Therapies Targeting Tumor-Associated Macrophages in Cancer
Targeting the Recruitment of TAMs
One strategy to target TAMs is to block their recruitment to the tumor site. It can be achieved by targeting the chemokine ligand 2 (CCL2) - chemokine receptor 2 (CCR2) axis. CCL2 is an inflammatory chemokine that can recruit macrophages and Treg lymphocytes leading to an immunosuppressive environment (Chang et al., 2016). To achieve this, a human IgG1k mAb called Carlumab was developed. A survey of clinical trials involving the CCL2-CCR2 axis is provided in Table 1.
A phase 2 study showed that this antibody was well-tolerated. However, it did not block the CCL2-CCR2 axis or have any antitumor activity as a single agent in metastatic prostate cancer (Pienta et al., 2013) (NCT00992186). When Carlumab was combined with four other chemotherapies, the treatment was still well tolerated but the suppression of CCL2-CCR2 axis remained elusive (Brana et al., 2015) (NCT01204996). In other studies, Carlumab was shown to transiently suppress CCL2 and had a preliminary antitumor activity (Sandhu et al., 2013) (NCT00537368, 2007). PF-04136309 combined with chemotherapy was also shown to be well-tolerated and led to a tumor response (Nywening et al., 2016).
Reprogramming of TAMs Toward an Antitumoral Phenotype
As mentioned previously, TAMs can exist in different functional states between the M1 and M2 phenotypes, making them highly heterogeneous and plastic cells (Biswas and Mantovani, 2010). Thus, they can be either pro- or anti-tumoral (Wynn et al., 2013). Reprogramming the TAMs toward a tumoricidal or a tumor-inhibition state may be a plausible therapeutic strategy. Different strategies are being studied in the clinic. These are reported in Table 2 (please refer also to Box 2).
Inhibition of CD47
Inhibition of CD47 is a strategy that can facilitate phagocytosis of tumor cells by macrophages. Indeed, CD47 expressed by cancer cells inhibits phagocytosis through its interaction with signal regulatory protein-α (SIRPα) expressed by macrophages thus sending out a “do not eat me” signal. Alternatively, CD47 can serve as a receptor for thrombospondin 1 (TSP1) to trigger specific signaling. Many tumors are described to overexpress CD47 (Zhang et al., 2015; Zhao et al., 2016). Inhibition of CD47 in a preclinical model showed a modification of microglia phenotypes in GB that was correlated with better survival (Hutter et al., 2019). Furthermore, in vivo, the anti-CD47 treatment is able to shift the macrophage phenotype toward an M1 type (Zhang et al., 2016) and induces anti-tumor effects (Li F. et al., 2017). The preclinical study of Hu5F9-G4 in pediatric malignant primary brain model demonstrated that this CD47 inhibitor is a safe and effective therapeutic agent (Gholamin et al., 2017). Hu5F9-G4 was also shown to be well tolerated in a clinical trial (Sikic et al., 2018) (NCT02216409, Table 2). TTI-621, a small molecule inhibiting CD47, is being investigated in an ongoing clinical trial. Interestingly, however, it has recently been observed that CD47 inhibition may result in cancer cell resistance to chemotherapy through escape to senescence (Guillon et al., 2019).
Activation of CD40
CD40 is expressed on monocytes, macrophages, dendritic cells, and B cells. It is a receptor that belongs to the TNF receptor superfamily. Many clinical trials targeting CD40 notably through agonistic or activating antibodies are ongoing (Table 3). In a mouse model, targeting CD40 was useful in producing antitumor effects that greatly improved the overall survival (Shoji et al., 2016). Targeting CD40 modulated the immune cell number and led to an antitumor response (Vonderheide et al., 2013; Nowak et al., 2015). In a mouse model, the combination of CSF1R inhibition and CD40 activation induced the reprogramming of TAMs (Hoves et al., 2018), thus allowing the protective response of T cells (Perry et al., 2018).
TLR Agonist
Toll-like receptors (TLRs) are normally activated by microbial moieties (including nucleic acids) allowing macrophages to acquire a M1 phenotype. Using a TLR agonist to reprogram macrophages was thus of interest in cancer treatment (Feng et al., 2019). Numerous TLR7 ligands, TLR9 ligands, and one TLR8 ligand have been tested for their antitumoral properties in clinical trials (Table 2). For example, the TLR7 agonist Imiquimod has been tested. It was well tolerated and associated to tumor regression and increased lymphocytic infiltrate (Adams et al., 2013) (NCT00899574). The TLR7 agonist 852A was also well tolerated with reversible side effects (Dudek et al., 2007). IMO-2055, a TLR9 agonist, demonstrated a possible antitumor activity when combined with erlotinib and bevacizumab (Smith et al., 2014) (NCT00633529).
Box 2. The content of exosomes as a therapeutic target to control TAMs phenotype.
Exosomes are microvesicles (30-120µm) that are secreted through exocytosis by various cells. They exert a variety of biological effects. GB cells can secrete exosomes that carry proteins such as EGFR variant III (EGFRvIII) Manda et al., 2018. The content of exosomes was shown to be different depending on partial pressure in O2 as cancer cells can adapt to their surroundings Zhang et al., 2017. Exosomes can mediate immunosuppressive properties in GB through their internalization in monocytes. Once they are internalized, they cause a rearrangement of the monocyte cytoskeleton and induce an M2 phenotype Gabrusiewicz et al., 2018. Vos et al. visualized the effect of GB-derived exosomes on TAMs and observed a shift of their cytokine profile to an immune-suppressive profile Van Der Vos et al., 2016. They also observed an elevation of miR-21 expression in TAMs associated with a decrease in c-Myc mRNA levels. GB-derived exosomes were shown to modify the expression of cell surface proteins and cytokines (IL-6 and VEGF), and to increase phagocytic activity in macrophages De Vrij et al., 2015. Also, blood samples from patients with GB were analyzed and shown to harbor GB-derived exosomes containing immunoglobulin (Ig) G2 and IgG4 antibody isotypes Harshyne et al., 2016. Those exosomes were able to induce the expression of CD163, associated with the M2 phenotype. Exosomes appear to be important for the communication between tumor cells and TAMs in GB. As key players from the tumor ecosystem, targeting them may impair the regulatory effects of GB cells on TAM immunosuppressive properties.
Depletion of TAMs
The activation of TAMs is dependent on the CSF1R signaling pathway. Therefore, CSF1R may be a way to target macrophages specifically. Many small molecules and antibodies were developed against CSF1R, and numerous clinical trials have been completed or are ongoing (Table 3). PLX3397 is a small molecule targeting CSF1R, it reduced the number of TAMs in a preclinical GB model and showed an antitumor activity (Coniglio and Segall, 2013; Yan et al., 2017). In clinical studies, PLX3397 was also well tolerated and showed anti-tumor responses after treatment (Tap et al., 2015) (NCT01004861). PLX3397 was also well tolerated but showed no efficacy in GB (Butowski et al., 2016) (NCT01349036). BLZ945, another small molecule inhibitor of CSF1R, can alter the polarization of TAMs in glioma (Pyonteck et al., 2013). It is currently being assessed in a clinical trial.
Another way to deplete the number of TAMs in the tumor is to use bisphosphonates. They are described for both direct and indirect anti-tumor effects such as induction of tumor apoptosis and inhibition of cell adhesion. More importantly, they alter the behavior of TAMs (Van Acker et al., 2016). Bisphosphonates are divided in two classes depending on their structure and mechanism of action. Clodronate belongs to the first group while zoledronate belongs to the second group. Both zoledronate and clodronate are still being assessed in clinical trials (Table 3).
Conclusion
In GB microenvironment, both resident and peripheral macrophages are present and there is an urgent need to understand their specific roles in tumor progression and resistance to treatment. It is obvious that macrophages may be a useful target to improve the outcome of cancer. Currently, many drugs targeting macrophages are being tested in the clinic. However, only a few are tested specifically in GB. The immune landscape in GB, and in cancer in general, has to be investigated further as there is a lack of efficacy in the clinic when only TAMs are targeted. The targeting of TAMs must be implemented hand in hand with the standard treatment to potentially improve the overall effect. In summary, TAMs seem to be a promising target to overcome resistance that arises in GB.
Author Contributions
HG, LR and EG wrote the manuscript. FH and EG contributed to the conception and design of the work. HG, LR, AR, MC, YD, PJ, FH, and EG contributed to manuscript amendments and revisions. All authors read and approved the submitted version.
Funding
This work was supported by the French national research agency (ANR) through the LabEx IRON << Innovative Radiopharmaceuticals in Oncology and Neurology>> as part of the French government “Investissements d’Avenir” program (ANR-11-LABX-0018). It was also supported by the ANR under the frame of EuroNanoMed III (project GLIOSILK). The work was additionally funded by the “Institut National de la Santé et de la Recherche Médicale” (INSERM) and by the University of Angers (Angers, France). It was also related to: (i) the PL-BIO 2014-2020 INCa (Institut National du Cancer) consortium MARENGO <<; MicroRNA agonist and antagonist Nanomedicines for GliOblastoma treatment: from molecular programmation to preclinical validation>>, (ii) to the MuMoFRaT project << Multi-scale Modeling & simulation of the response to hypo-Fractionated Radiotherapy or repeated molecular radiation Therapies>> supported by “La Région Pays-de-la-Loire” and by the Cancéropôle Grand-Ouest (Vectorization, imaging and radiotherapies network), (iii) the LabEX IGO and the ANR through the investment of the future program ANR-11-LABX-0016-01, (iv) the SIRIC ILIAD program supported by INCa, and (v) the Ministry of Health and the Institute for Health and Medical Research (Inserm) (contract INCa-DGOS-Inserm_12558). HG and LR were PhD fellows funded by the LabEx IRON and by the LabEx IRON-2 and the University of Angers, respectively.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Achyut, B. R., Angara, K., Jain, M., Borin, T. F., Rashid, M. H., Iskander, A. S. M., et al. (2017). Canonical NFκB signaling in myeloid cells is required for the glioblastoma growth. Sci. Rep. 7, 1–12. doi: 10.1038/s41598-017-14079-4
Adams, S., Kozhaya, L., Martiniuk, F., Meng, T., Chiriboga, L., Liebes, L., et al. (2013). Rejection of Skin Metastases in Patients With Breast Cancer. Clin. Cancer Res. 18, 6748–6757. doi: 10.1158/1078-0432.CCR-12-1149
Arsic, N., Gadea, G., Lagerqvist, E. L., Busson, M., Cahuzac, N., Brock, C., et al. (2015). The p53 isoform Δ133p53β promotes cancer stem cell potential. Stem Cell Rep. 4, 531–540. doi: 10.1016/j.stemcr.2015.02.001
Atri, C., Guerfali, F. Z., Laouini, D. (2018). Role of Human Macrophage Polarization in Inflammation during Infectious Diseases. Int. J. Mol. Sci. 19, 1801. doi: 10.3390/ijms19061801
Badie, B., Schartner, J., Vorpahl, J., Preston, K. (2000). Interferon-γ Induces Apoptosis and Augments the Expression of Fas and Fas Ligand by Microglia in Vitro. Exp. Neurol. 162, 290–296. doi: 10.1006/exnr.1999.7345
Baik, S. H., Kang, S., Lee, W., Choi, H., Chung, S., Kim, J., et al. (2019). A Breakdown in Metabolic Reprogramming Causes Microglia Dysfunction in Alzheimer’s Disease. Cell Metab. doi: 10.1016/j.cmet.2019.06.005
Barberi, T., Martin, A., Suresh, R., Barakat, D. J., Harris-Bookman, S., Drake, C. G., et al. (2018). Absence of host NF-κB p50 induces murine glioblastoma tumor regression, increases survival, and decreases T-cell induction of tumor-associated macrophage M2 polarization. Cancer Immunol. Immunother. 67, 1491–1503. doi: 10.1007/s00262-018-2184-2
Bhat, K. P. L., Balasubramaniyan, V., Vaillant, B., Ezhilarasan, R., Hummelink, K., Hollingsworth, F., et al. (2013). Mesenchymal Differentiation Mediated by NF-κB Promotes Radiation Resistance in Glioblastoma. Cancer Cell 24, 331–346. doi: 10.1016/j.ccr.2013.08.001
Biswas, S. K., Mantovani, A. (2010). Macrophage plasticity and interaction with lymphocyte subsets: Cancer as a paradigm. Nat. Immunol. 11, 889–896. doi: 10.1038/ni.1937
Brana, I., Calles, A., LoRusso, P. M., Yee, L. K., Puchalski, T. A., Seetharam, S., et al. (2015). Carlumab, an anti-C-C chemokine ligand 2 monoclonal antibody, in combination with four chemotherapy regimens for the treatment of patients with solid tumors: an open-label, multicenter phase 1b study. Targeting Oncol. 10, 111–123. doi: 10.1007/s11523-014-0320-2
Brown, N. F., Carter, T. J., Ottaviani, D., Mulholland, P. (2018). Harnessing the immune system in glioblastoma. Br. J. Cancer 119, 1171–1181. doi: 10.1038/s41416-018-0258-8
Bryukhovetskiy, I. S., Dyuizen, I. V., Shevchenko, V. E., Bryukhovetskiy, A. S., Mischenko, P. V., Milkina, E. V., et al. (2016). Hematopoietic stem cells as a tool for the treatment of glioblastoma multiforme. Mol. Med. Rep. 14, 4511–4520. doi: 10.3892/mmr.2016.5852
Butowski, N., Colman, H., De Groot, J. F., Omuro, A. M., Nayak, L., Wen, P. Y., et al. (2016). Orally administered colony stimulating factor 1 receptor inhibitor PLX3397 in recurrent glioblastoma: An Ivy Foundation Early Phase Clinical Trials Consortium phase II study. Neuro. Oncol. 18, 557–564. doi: 10.1093/neuonc/nov245
Cai, J., Zhang, W., Yang, P., Wang, Y., Li, M., Zhang, C., et al. (2015). Identification of a 6-cytokine prognostic signature in patients with primary glioblastoma harboring M2 microglia/macrophage phenotype relevance. PloS One 10, e0126022–e0126022. doi: 10.1371/journal.pone.0126022
Caponegro, M. D., Moffitt, R. A., Tsirka, S. E. (2018). Expression of neuropilin-1 is linked to glioma associated microglia and macrophages and correlates with unfavorable prognosis in high grade gliomas. Oncotarget 9, 35655–35665. doi: 10.18632/oncotarget.26273
Carroll, R. G., Zasłona, Z., Galván-Peña, S., Koppe, E. L., Sévin, D. C., Angiari, S., et al. (2018). An unexpected link between fatty acid synthase and cholesterol synthesis in proinflammatory macrophage activation. J. Biol. Chem. doi: 10.1074/jbc.RA118.001921
Chae, M., Peterson, T. E., Balgeman, A., Chen, S., Zhang, L., Renner, D. N., et al. (2015). Increasing glioma-associated monocytes leads to increased intratumoral and systemic myeloid-derived suppressor cells in a murine model. Neuro. Oncol. 17, 978–991. doi: 10.1093/neuonc/nou343
Chang, A. L., Miska, J., Wainwright, D. A., Dey, M., Rivetta, C. V., Yu, D., et al. (2016). CCL2 Produced by the Glioma Microenvironment Is Essential for the Recruitment of Regulatory T Cells and Myeloid-Derived Suppressor Cells. Cancer Res. 76, 5671–5682. doi: 10.1158/0008-5472.CAN-16-0144
Charles, N. A., Holland, E. C., Gilbertson, R., Glass, R., Kettenmann, H. (2011). The brain tumor microenvironment. Glia 59, 1169–1180. doi: 10.1002/glia.21136
Chen, Z., Hambardzumyan, D. (2018). Immune Microenvironment in Glioblastoma Subtypes. Front. Immunol. 9, 1004. doi: 10.3389/fimmu.2018.01004
Chen, Z., Feng, X., Herting, C. J., Garcia, V. A., Nie, K., Pong, W. W., et al. (2017). Cellular and Molecular Identity of Tumor-Associated Macrophages in Glioblastoma. Cancer Res. 77, 2266–2278. doi: 10.1158/0008-5472.CAN-16-2310
Coniglio, S. J., Segall, J. E. (2013). Review: Molecular mechanism of microglia stimulated glioblastoma invasion. Matrix Biol. 32, 372–380. doi: 10.1016/j.matbio.2013.07.008
Cui, X., Morales, R. T. T., Qian, W., Wang, H., Gagner, J. P., Dolgalev, I., et al. (2018). Hacking macrophage-associated immunosuppression for regulating glioblastoma angiogenesis. Biomaterials 161, 164–178. doi: 10.1016/j.biomaterials.2018.01.053
De Vrij, J., Niek Maas, S. L., Kwappenberg, K. M. C., Schnoor, R., Kleijn, A., Dekker, L., et al. (2015). Glioblastoma-derived extracellular vesicles modify the phenotype of monocytic cells. Int. J. Cancer 137, 1630–1642. doi: 10.1002/ijc.29521
Deborde, S., Hall, A., Wong, R.J., Deborde, S., Omelchenko, T., Lyubchik, A., et al. (2016). Schwann cells induce cancer cell dispersion and invasion Find the latest version : Schwann cells induce cancer cell dispersion and invasion. J. Clin. Invest. 126, 1538–1554. doi: 10.1172/JCI82658
Deng, L., Stafford, J. H., Liu, S. C., Chernikova, S. B., Merchant, M., Recht, L., et al. (2017). SDF-1 Blockade Enhances Anti-VEGF Therapy of Glioblastoma and Can Be Monitored by MRI. Neoplasia (United States) 19, 1–7. doi: 10.1016/j.neo.2016.11.010
Doan, N. B., Nguyen, H. S., Alhajala, H. S., Jaber, B., Al-Gizawiy, M. M., Ahn, E.-Y. E., et al. (2018). Identification of radiation responsive genes and transcriptome profiling via complete RNA sequencing in a stable radioresistant U87 glioblastoma model. Oncotarget 9, 23532–23542. doi: 10.18632/oncotarget.25247
Dudek, A. Z., Yunis, C., Harrison, L. I., Kumar, S., Hawkinson, R., Cooley, S., et al. (2007). First in human phase I trial of 852A, a novel systemic toll-like receptor 7 agonist, to activate innate immune responses in patients with advanced cancer. Clin. Cancer Res. 13, 7119–7125. doi: 10.1158/1078-0432.CCR-07-1443
Feng, X., Szulzewsky, F., Yerevanian, A., Chen, Z., Heinzmann, D., Rasmussen, R. D., et al. (2015). Loss of CX3CR1 increases accumulation of inflammatory monocytes and promotes gliomagenesis. Oncotarget 6, 15077–15094. doi: 10.18632/oncotarget.3730
Feng, Y., Mu, R., Wang, Z., Xing, P., Zhang, J., Dong, L., et al. (2019). A toll-like receptor agonist mimicking microbial signal to generate tumor-suppressive macrophages. Nat. Commun. 10, 2272. doi: 10.1038/s41467-019-10354-2
Finn, O. J. A. (2018). Believer’s Overview of Cancer Immunosurveillance and Immunotherapy. J. Immunol. 200, 385–391. doi: 10.4049/jimmunol.1701302
Flavahan, W. A., Nakano, I., Rich, J. N., Otvos, B., Silver, D. J., Sinyuk, M., et al. (2016). Cancer Stem Cell-Secreted Macrophage Migration Inhibitory Factor Stimulates Myeloid Derived Suppressor Cell Function and Facilitates Glioblastoma Immune Evasion. Stem Cells 34, 2026–2039. doi: 10.1002/stem.2393
Gabrusiewicz, K., Li, X., Wei, J., Hashimoto, Y., Marisetty, A. L., Ott, M., et al. (2018). Glioblastoma stem cell-derived exosomes induce M2 macrophages and PD-L1 expression on human monocytes. Oncoimmunology 7 (e1412909), 1–10. doi: 10.1080/2162402X.2017.1412909
Gagner, J. P., Sarfraz, Y., Ortenzi, V., Alotaibi, F. M., Chiriboga, L. A., Tayyib, A. T., et al. (2017). Multifaceted C-X-C Chemokine Receptor 4 (CXCR4) Inhibition Interferes with Anti–Vascular Endothelial Growth Factor Therapy–Induced Glioma Dissemination. Am. J. Pathol. 187, 2080–2094. doi: 10.1016/j.ajpath.2017.04.020
Gholamin, S., Mitra, S. S., Feroze, A. H., Liu, J., Kahn, S. A., Zhang, M., et al. (2017). Disrupting the CD47-SIRPα anti-phagocytic axis by a humanized anti-CD47 antibody is an efficacious treatment for malignant pediatric brain tumors. Sci. Transl. Med. 9, 1–14. doi: 10.1126/scitranslmed.aaf2968
Ginhoux, F., Guilliams, M. (2016). Tissue-Resident Macrophage Ontogeny and Homeostasis. Immunity 44, 439–449. doi: 10.1016/j.immuni.2016.02.024
Ginhoux, F., Lim, S., Hoeffel, G., Low, D., Huber, T. (2013). Origin and differentiation of microglia. Front. Cell. Neurosci. 7, 45. doi: 10.3389/fncel.2013.00045
Graeber, M. B., Scheithauer, B. W., Kreutzberg, G. W. (2002). Microglia in brain tumors. Glia 40, 252–259. doi: 10.1002/glia.10147
Guillon, J., Petit, C., Moreau, M., Toutain, B., Henry, C., Roché, H., et al. (2019). Regulation of senescence escape by TSP1 and CD47 following chemotherapy treatment. Cell Death Dis. doi: 10.1038/s41419-019-1406-7
Gupta, K., Burns, T. C. (2018). Radiation-Induced Alterations in the Recurrent Glioblastoma Microenvironment: Therapeutic Implications. Front. Oncol. 8, 503. doi: 10.3389/fonc.2018.00503
Hambardzumyan, D., Gutmann, D. H., Kettenmann, H. (2015). The role of microglia and macrophages in glioma maintenance and progression. Nat. Neurosci. 19, 20. doi: 10.1038/nn.4185
Hanahan, D., Weinberg, R. A., Adams, J. M., Cory, S., Aguirre-Ghiso, J. A., Ahmed, Z., et al. (2011). Hallmarks of cancer: the next generation. Cell 144, 646–674. doi: 10.1016/j.cell.2011.02.013
Hardee, M. E., Marciscano, A. E., Medina-Ramirez, C. M., Zagzag, D., Narayana, A., Lonning, S. M., et al. (2012). Resistance of glioblastoma-initiating cells to radiation mediated by the tumor microenvironment can be abolished by inhibiting transforming growth factor-β. Cancer Res. 72, 4119–4129. doi: 10.1158/0008-5472.CAN-12-0546
Harshyne, L. A., Nasca, B. J., Kenyon, L. C., Andrews, D. W., Hooper, D. C. (2016). Serum exosomes and cytokines promote a T-helper cell type 2 environment in the peripheral blood of glioblastoma patients. Neuro. Oncol. 18, 206–215. doi: 10.1093/neuonc/nov107
Herisson, F., Frodermann, V., Courties, G., Rohde, D., Sun, Y., Vandoorne, K., et al. (2018). Direct vascular channels connect skull bone marrow and the brain surface enabling myeloid cell migration. Nat. Neurosci. doi: 10.1038/s41593-018-0213-2
Hide, T., Komohara, Y., Miyasato, Y., Nakamura, H., Makino, K., Takeya, M., et al. (2018). Oligodendrocyte Progenitor Cells and Macrophages/Microglia Produce Glioma Stem Cell Niches at the Tumor Border. EBioMedicine 30, 94–104. doi: 10.1016/j.ebiom.2018.02.024
Hoves, S., Ooi, C.-H., Wolter, C., Sade, H., Bissinger, S., Schmittnaegel, M., et al. (2018). Rapid activation of tumor-associated macrophages boosts preexisting tumor immunity. J. Exp. Med. doi: 10.1084/jem.20171440
Hu, X., Li, P., Guo, Y., Wang, H., Leak, R. K., Chen, S., et al. (2012). Microglia/macrophage polarization dynamics reveal novel mechanism of injury expansion after focal cerebral ischemia. Stroke 43, 3063–3070. doi: 10.1161/STROKEAHA.112.659656
Hudson, A. L., Parker, N. R., Khong, P., Parkinson, J. F., Dwight, T., Ikin, R. J., et al. (2018). Glioblastoma Recurrence Correlates With Increased APE1 and Polarization Toward an Immuno-Suppressive Microenvironment. Front. Oncol. 8 (314), 1–10. doi: 10.3389/fonc.2018.00314
Hung, N. A., Eiholzer, R. A., Kirs, S., Zhou, J., Ward-Hartstonge, K., Wiles, A. K., et al. (2016). Telomere profiles and tumor-associated macrophages with different immune signatures affect prognosis in glioblastoma. Mod. Pathol. 29, 212–226. doi: 10.1038/modpathol.2015.156
Hussain, S. F., Yang, D., Suki, D., Aldape, K., Grimm, E., Heimberger, A. B. (2006). The role of human glioma-infiltrating microglia/macrophages in mediating antitumor immune responses. Neuro. Oncol. 8, 261–279. doi: 10.1215/15228517-2006-008
Hutter, G., Theruvath, J., Graef, C. M., Zhang, M., Schoen, M. K., Manz, E. M., et al. (2019). Microglia are effector cells of CD47-SIRPα antiphagocytic axis disruption against glioblastoma. Proc. Natl. Acad. Sci. U. S. A. 116, 997–1006. doi: 10.1073/pnas.1721434116
Jeannin, P., Paolini, L., Adam, C., Delneste, Y. (2018). The roles of CSFs on the functional polarization of tumor-associated macrophages. FEBS J. 285, 680–699. doi: 10.1111/febs.14343
Kast, R. E., Hill, Q. A., Wion, D., Mellstedt, H., Focosi, D., Karpel-Massler, G., et al. (2017). Glioblastoma-synthesized G-CSF and GM-CSF contribute to growth and immunosuppression: Potential therapeutic benefit from dapsone, fenofibrate, and ribavirin. Tumor Biol. 39, 1–10. doi: 10.1177/1010428317699797
Kazantseva, M., Eiholzer, R. A., Mehta, S., Taha, A., Bowie, S., Roth, I., et al. (2018). Elevation of the TP53 isoform Δ133p53β in glioblastomas: an alternative to mutant p53 in promoting tumor development. J. Pathol. 246, 77–88. doi: 10.1002/path.5111
Kennedy, B. C., Showers, C. R., Anderson, D. E., Anderson, L., Canoll, P., Bruce, J. N., et al. (2013). Tumor-associated macrophages in glioma: friend or foe? J. Oncol. 2013, 486912. doi: 10.1155/2013/486912
Kitange, G. J., Carlson, B. L., Schroeder, M. A., Decker, P. A., Morlan, B. W., Wu, W., et al. (2010). Expression of CD74 in high grade gliomas: a potential role in temozolomide resistance. J. Neurooncol. 100, 177–186. doi: 10.1007/s11060-010-0186-9
Kloepper, J., Riedemann, L., Amoozgar, Z., Seano, G., Susek, K., Yu, V., et al. (2016). Ang-2/VEGF bispecific antibody reprograms macrophages and resident microglia to anti-tumor phenotype and prolongs glioblastoma survival. Proc. Natl. Acad. Sci. 113, 4476 LP– 4481. doi: 10.1073/pnas.1525360113
Kohanbash, G., McKaveney, K., Sakaki, M., Ueda, R., Mintz, A. H., Amankulor, N., et al. (2013). GM-CSF promotes the immunosuppressive activity of glioma-infiltrating myeloid cells through interleukin-4 receptor-α. Cancer Res. 73, 6413–6423. doi: 10.1158/0008-5472.CAN-12-4124
Komohara, Y., Ohnishi, K., Kuratsu, J., Takeya, M. (2008). Possible involvement of the M2 anti-inflammatory macrophage phenotype in growth of human gliomas. J. Pathol. 216, 15–24. doi: 10.1002/path.2370
Kong, B., Michalski, C. W., Friess, H., Kleeff, J. (2010). Surgical procedure as an inducer of tumor angiogenesis. Exp. Oncol. 32, 186–189.
Kumar, R., De Mooij, T., Peterson, T. E., Kaptzan, T., Johnson, A. J., Daniels, D. J., et al. (2017). Modulating glioma-mediated myeloid-derived suppressor cell development with sulforaphane. PloS One 12, 1–26. doi: 10.1371/journal.pone.0179012
Latha, K., Yan, J., Yang, Y., Gressot, L. V., Kong, L.-Y., Manyam, G., et al. (2018). The Role of Fibrinogen-Like Protein 2 on Immunosuppression and Malignant Progression in Glioma. JN7CI J. Natl. Cancer Inst. 111, 1–9. doi: 10.1093/jnci/djy107
Leblond, M. M., Pérès, E. A., Helaine, C., Gérault, A. N., Moulin, D., Anfray, C., et al. (2017). M2 macrophages are more resistant than M1 macrophages following radiation therapy in the context of glioblastoma. Oncotarget 8, 72597–72612. doi: 10.18632/oncotarget.19994
Lee, J., Dang, X., Borboa, A., Coimbra, R., Baird, A., Eliceiri, B. P. (2015). Thrombin-processed Ecrg4 recruits myeloid cells and induces antitumorigenic inflammation. Neuro. Oncol. 17, 685–696. doi: 10.1093/neuonc/nou302
Li, Z., Huang, Q., Chen, H., Lin, Z., Zhao, M., Jiang, Z. (2017). Interferon Regulatory Factor 7 Promoted Glioblastoma Progression and Stemness by Modulating IL-6 Expression in Microglia. J. Cancer 8, 207–219. doi: 10.7150/jca.16415
Li, T.-F., Li, K., Wang, C., Liu, X., Wen, Y., Xu, Y.-H., et al. (2017). Harnessing the cross-talk between tumor cells and tumor-associated macrophages with a nano-drug for modulation of glioblastoma immune microenvironment. J. Control. Release 268, 128–146. doi: 10.1016/j.jconrel.2017.10.024
Li, F., Lv, B., Liu, Y., Hua, T., Han, J., Sun, C., et al. (2017). Blocking the CD47-SIRPα axis by delivery of anti-CD47 antibody induces antitumor effects in glioma and glioma stem cells. Oncoimmunology 7, e1391973–e1391973. doi: 10.1080/2162402X.2017.1391973
Li, Y., Dong, X., Cai, J., Yin, S., Sun, Y., Yang, D., et al. (2018). SERPINA3 induced by astroglia/microglia co-culture facilitates glioblastoma stem-like cell invasion. Oncol. Lett. 15, 285–291. doi: 10.3892/ol.2017.7275
Lin, J. D., Nishi, H., Poles, J., Niu, X., McCauley, C., Rahman, K., et al. (2019). Single-cell analysis of fate-mapped macrophages reveals heterogeneity, including stem-like properties, during atherosclerosis progression and regression. JCI Insight 4 (e124574), 1–15. doi: 10.1172/jci.insight.124574
Lisi, L., Ciotti, G. M. P., Braun, D., Kalinin, S., Currò, D., Dello Russo, C., et al. (2017). Expression of iNOS, CD163 and ARG-1 taken as M1 and M2 markers of microglial polarization in human glioblastoma and the surrounding normal parenchyma. Neurosci. Lett. 645, 106–112. doi: 10.1016/j.neulet.2017.02.076
Locati, M., Curtale, G., Mantovani, A. (2020). Diversity, Mechanisms, and Significance of Macrophage Plasticity. Annu. Rev. Pathol. Mech. Dis 15, 123–147. doi: 10.1146/annurev-pathmechdis-012418-012718
Louis, D. N., Ohgaki, H., Wiestler, O. D., Cavenee, W. K., Burger, P. C., Jouvet, A., et al. (2007). The 2007 WHO classification of tumours of the central nervous system. Acta Neuropathol. 114, 97–109. doi: 10.1007/s00401-007-0243-4
Louis, D. N., Perry, A., Reifenberger, G., von Deimling, A., Figarella-Branger, D., Cavenee, W. K., et al. (2016). The 2016 World Health Organization Classification of Tumors of the Central Nervous System: a summary. Acta Neuropathol. 131, 803–820. doi: 10.1007/s00401-016-1545-1
Lu-Emerson, C., Snuderl, M., Kirkpatrick, N. D., Goveia, J., Davidson, C., Huang, Y., et al. (2013). Increase in tumor-associated macrophages after antiangiogenic therapy is associated with poor survival among patients with recurrent glioblastoma. Neuro. Oncol. 15, 1079–1087. doi: 10.1093/neuonc/not082
Müller, A., Brandenburg, S., Turkowski, K., Müller, S., Vajkoczy, P. (2015). Resident microglia, and not peripheral macrophages, are the main source of brain tumor mononuclear cells. Int. J. Cancer 137, 278–288. doi: 10.1002/ijc.29379
Martinez, F. O., Gordon, S. (2014). The M1 and M2 paradigm of macrophage activation: time for reassessment. F1000Prime Rep. 6, 13. doi: 10.12703/P6-13
Matias, D., Balça-Silva, J., da Graça, G. C., Wanjiru, C. M., Macharia, L. W., Nascimento, C. P., et al. (2018). Microglia/Astrocytes–Glioblastoma Crosstalk: Crucial Molecular Mechanisms and Microenvironmental Factors. Front. Cell. Neurosci. 12, 235. doi: 10.3389/fncel.2018.00235
Mieczkowski, J., Kocyk, M., Nauman, P., Gabrusiewicz, K., Sielska, M., Przanowski, P., et al. (2015). Down-regulation of IKKβ expression in glioma-infiltrating microglia/macrophages is associated with defective inflammatory/immune gene responses in glioblastoma. Oncotarget 6, 33077–33090. doi: 10.18632/oncotarget.5310
Murray, P. J., Allen, J. E., Biswas, S. K., Fisher, E. A., Gilroy, D. W., Goerdt, S., et al. (2014). Macrophage activation and polarization: nomenclature and experimental guidelines. Immunity 41, 14–20. doi: 10.1016/j.immuni.2014.06.008
NCT00003232 (2004). Combination Chemotherapy in Treating Pain in Patients With Hormone Refractory Metastatic Prostate Cancer. ClinicalTrials.gov. Available at https://clinicaltrials.gov/ct2/show/NCT00003232
NCT00009945 (2003). Clodronate With or Without Chemotherapy and/or Hormonal Therapy in Treating Women With Stage I or Stage II Breast Cancer. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT00009945
NCT00127205 (2005). S0307 Phase III Trial of Bisphosphonates as Adjuvant Therapy for Primary Breast Cancer. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT00127205
NCT00301873 (2006). Zoledronate in Preventing Osteoporosis in Patients With Primary Malignant Glioma. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT00301873
NCT00537368 (2007). First Study of the Safety of CNTO 888 in Patients With Solid Tumors. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT00537368
NCT00633529 (2008). Safety of Adding IMO-2055 to Erlotinib + Bevacizumab in 2nd Line Treatment for Patients With NSCLC. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT00633529
NCT00719199 (2008). Study of FOLFIRI Plus Cetuximab Plus IMO-2055 in Patients With Colorectal Cancer. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT00719199
NCT00821652 (2009). Randomized, Double Blind, Placebo-controlled Topical Resiquimod Adjuvant for NY-ESO-1 Protein Vaccination. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT00821652
NCT00885326 (2009). N2007-02:Bevacizumab,Cyclophosphamide,& Zoledronic Acid in Patients W/ Recurrent or Refractory High-Risk Neuroblastoma. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT00885326
NCT00899574 (2009). Imiquimod for Breast Cancer Patients With Chest Wall Recurrence or Skin Metastases. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT00899574
NCT00909142 (2009). Bonefos and the Consumption of Analgesics (BICAM). ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT00909142
NCT00992186 (2009). A Study of the Safety and Efficacy of Single-agent Carlumab (an Anti-Chemokine Ligand 2 [CCL2]) in Participants With Metastatic Castrate-Resistant Prostate Cancer. ClinicalTrials.gov.Available at: https://clinicaltrials.gov/ct2/show/NCT00992186
NCT01004861 (2009). Safety Study of PLX108-01 in Patients With Solid Tumors. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT01004861
NCT01015560 (2009). S0916, MLN1202 in Treating Patients With Bone Metastases. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT01015560
NCT01040832 (2009). EMD 1201081 in Combination With Cetuximab in Second-Line Cetuximab-Naïve Subjects With Recurrent or Metastatic Squamous Cell Carcinoma of the Head and Neck. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT01040832
NCT01198457 (2010). Study to Investigate Adherence of Patients to Clodronate (Bonefos) Treatment (BONA). ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT01198457
NCT01204996 (2010). A Study of the Safety and Efficacy of CNTO 888 in Combination With SoC (Standard of Care) Chemotherapy in Patients With Solid Tumors. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT01204996
NCT01294293 and NCT01334177 (2011). TLR8 Agonist VTX-2337 and Pegylated Liposomal Doxorubicin Hydrochloride or Paclitaxel in Treating Patients With Recurrent or Persistent Ovarian Epithelial, Fallopian Tube, or Peritoneal Cavity Cancer. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT01294293
NCT01316822 (2011). A Study of ARRY-382 in Patients With Selected Advanced or Metastatic Cancers. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT01316822
NCT01345019 (2011). Denosumab Compared to Zoledronic Acid in the Treatment of Bone Disease in Patients With Multiple Myeloma. ClinicalTrials.gov.
NCT01346358 (2011). A Study of IMC-CS4 in Subjects With Advanced Solid Tumors. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT01346358
NCT01349036 (2011). A Phase 2 Study of PLX3397 in Patients With Recurrent Glioblastoma. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT01349036
NCT01421017 (2011). Toll-like Receptor (TLR) 7 Agonist, Cyclophosphamide, and Radiotherapy for Breast Cancer With Skin Metastases. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT01421017
NCT01494688 (2011). A Study of RO5509554 as Monotherapy and in Combination With Paclitaxel in Participants With Advanced Solid Tumors. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT01494688
NCT01561911 and NCT00607048. (2012) A Phase I Study of the Chimeric Anti-CD40 Monoclonal Antibody ChiLob 7/4 to Treat Advanced Malignancies Refractory to Conventional Anti-cancer Treatment. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT01561911
NCT02216409 (2014). Phase 1 Trial of Hu5F9-G4, a CD47-targeting Antibody. ClinicalTrials.gov. Available at https://clinicaltrials.gov/ct2/show/NCT02216409
NCT02252146. Dose Escalation Study in Patients With Relapsed or Refractory DLBCL and MyD88 L265P Mutation. ClinicalTrials.gov.
NCT02254772 (2014). A Phase I/II Study of Intratumoral Injection of SD-101. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT02254772
NCT02304393 (2014). A Study of Selicrelumab (RO7009789) in Combination With Atezolizumab in Participants With Locally Advanced and/or Metastatic Solid Tumors. ClinicalTrials.gov.Available at: https://clinicaltrials.gov/ct2/show/NCT02304393
NCT02367196 (2015). A Phase 1, Dose Finding Study of CC-90002 in Subjects With Advanced Solid and Hematologic Cancers. ClinicalTrials.gov. Available at https://clinicaltrials.gov/ct2/show/NCT02367196
NCT02371369 (2015). Phase 3 Study of Pexidartinib for Pigmented Villonodular Synovitis (PVNS) or Giant Cell Tumor of the Tendon Sheath (GCT-TS) (ENLIVEN). ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT02371369
NCT02431559 (2015). A Phase 1/2 Study of Motolimod (VTX-2337) and MEDI4736 in Subjects With Recurrent, Platinum-Resistant Ovarian Cancer for Whom Pegylated Liposomal Doxorubicin (PLD) is Indicated. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT02431559
NCT02452424 (2015). A Combination Clinical Study of PLX3397 and Pembrolizumab To Treat Advanced Melanoma and Other Solid Tumors. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT02452424
NCT02452697 (2015). Ph2 NK Cell Enriched DCIs w/wo RLR9 Agonist, DUK-CPG-001 From Donors Following Allogeneic SCT (NK-DCI). ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT02452697
NCT02482168 (2015). Study of the CD40 Agonistic Monoclonal Antibody APX005M. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT02482168
NCT02526017 (2015). Study of Cabiralizumab in Combination With Nivolumab in Patients With Selected Advanced Cancers (FPA008-003). ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT02526017
NCT02663518 (2016). A Trial of TTI-621 for Patients With Hematologic Malignancies and Selected Solid Tumors. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT02663518
NCT02678338 (2016). CAMELLIA: Anti-CD47 Antibody Therapy in Haematological Malignancies. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT02678338
NCT02718911 (2016). A Study of LY3022855 in Combination With Durvalumab or Tremelimumab in Participants With Advanced Solid Tumors. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT02718911
NCT02732938 (2016). Ph1b/2 Study of PF-04136309 in Combination With Gem/Nab-P in First-line Metastatic Pancreatic Patients (CCR2i). ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT02732938
NCT02777710 (2016). Evaluation of Safety and Activity of an Anti-PDL1 Antibody (DURVALUMAB) Combined With CSF-1R TKI (PEXIDARTINIB) in Patients With Metastatic/Advanced Pancreatic or Colorectal Cancers (MEDIPLEX). ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT02777710
NCT02829723 (2016). Phase I/II Study of BLZ945 Single Agent or BLZ945 in Combination With PDR001 in Advanced Solid Tumors. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT02829723
NCT02927964 (2016). TLR9 Agonist SD-101, Ibrutinib, and Radiation Therapy in Treating Patients With Relapsed or Refractory Grade 1-3A Follicular Lymphoma. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT02927964
NCT02953509 (2016). Trial of Hu5F9-G4 in Combination With Rituximab in Relapsed/Refractory B-cell Non-Hodgkin’s Lymphoma. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT02953509
NCT02953782 (2016). Trial of Hu5F9-G4 in Combination With Cetuximab in Patients With Solid Tumors and Advanced Colorectal Cancer. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT02953782
NCT03007732 (2017). Pembrolizumab in Combination With Intratumoral SD-101 Therapy. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT03007732
NCT03013218 (2017). A Study of ALX148 in Patients With Advanced Solid Tumors and Lymphoma. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT03013218
NCT03069469 (2017). Study of DCC-3014 in Patients With Advanced Malignancies. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT03069469
NCT03101254 (2017). LY3022855 With BRAF/MEK Inhibition in Patients With Melanoma. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT03101254
NCT03123783 (2017). CD40 Agonistic Antibody APX005M in Combination With Nivolumab. ClinicalTrials.gov.
NCT03153410 (2017). Pilot Study With CY, Pembrolizumab, GVAX, and IMC-CS4 (LY3022855) in Patients With Borderline Resectable Adenocarcinoma of the Pancreas. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT03153410
NCT03165994 (2017). APX005M With Concurrent Chemoradiation for Resectable Esophageal and Gastroesophageal Junction Cancers. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT03165994
NCT03238027 (2017). A Phase 1 Study to Investigate SNDX-6352 Alone or in Combination With Durvalumab in Patients With Solid Tumors. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT03238027
NCT03248479 (2017). Hu5F9-G4 Monotherapy or Hu5F9-G4 in Combination With Azacitidine in Patients With Hematological Malignancies. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT03248479
NCT03301896 (2017). Study of the Safety and Efficacy of LHC165 Single Agent and in Combination With PDR001 in Patients With Advanced Malignancies. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT03301896
NCT03329950 (2017). A Study of CDX-1140 as Monotherapy or in Combination in Patients With Advanced Malignancies. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT03329950
NCT03389802 (2018). Phase I Study of APX005M in Pediatric CNS Tumors. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT03389802
NCT03410901 (2018). TLR9 Agonist SD-101, Anti-OX40 Antibody BMS 986178, and Radiation Therapy in Treating Patients With Low-Grade B-Cell Non-Hodgkin Lymphomas. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT03410901
NCT03416335 (2018). A Study of DSP-0509 in Patients With Advanced Solid Tumors to Determine the Safety and the Pharmacokinetic Profile. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT03416335
NCT03435640 (2018). A Study of NKTR-262 in Combination With NKTR-214 and With NKTR-214 Plus Nivolumab in Patients With Locally Advanced or Metastatic Solid Tumor Malignancies (REVEAL). ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT03435640
NCT03496662 (2018). BMS-813160 With Nivolumab and Gemcitabine and Nab-paclitaxel in Borderline Resectable and Locally Advanced Pancreatic Ductal Adenocarcinoma (PDAC). ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT03496662
NCT03502330 (2018). APX005M With Nivolumab and Cabiralizumab in Advanced Melanoma, Non-small Cell Lung Cancer or Renal Cell Carcinoma. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT03502330
NCT03507699 (2018). Combined Immunotherapy and Radiosurgery for Metastatic Colorectal Cancer. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT03507699
NCT03512340 (2018). Study of SRF231 in Patients With Advanced Solid and Hematologic Cancers. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT03512340
NCT03530683 (2018). A Trial of TTI-622 in Patients With Advanced Relapsed or Refractory Lymphoma or Myeloma (TTI-622-01). ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT03530683
NCT03557970 and NCT02880371 (2018). CSF1R Inhibitor JNJ-40346527 in Treating Participants With Relapsed or Refractory Acute Myeloid Leukemia. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT03557970
NCT03618641 (2018). CMP-001 in Combo With Nivolumab in Stage IIIB/C/D Melanoma Patients With Clinically Apparent Lymph Node Disease. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT03618641
NCT03694977 (2018). Biomarker Study of PDR001 in Combination With MCS110 in Gastric Cancer. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT03694977
NCT03697564 (2018). Nivolumab + Cabiralizumab + Gemcitabine Versus Gemcitabine in Patients With Stage IV Pancreatic Cancer Achieving Disease Control in Response to First-line Chemotherapy (GemCaN Trial). ClinicalTrials.gov.Available at: https://clinicaltrials.gov/ct2/show/NCT03697564
NCT03717103 (2018). A Phase 1 Study Evaluating the Safety, Tolerability, and Initial Efficacy of Recombinant Human Anti-cluster Differentiation Antigen 47 (CD47) Monoclonal Antibody Injection (IBI188) in Patients With Advanced Malignancies. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT03717103
NCT03763149 (2018). A Study Evaluating the Safety, Tolerability, and Initial Efficacy of Recombinant Human Anti-cluster Differentiation Antigen 47 (CD47) Monoclonal Antibody Injection (IBI188) in Patients With Advanced Malignant Tumors and Lymphomas. ClinicalTrials.gov.
NCT03778879 (2018). Pre-operative Stereotactic Body Radiation Therapy for Pancreatic Adenocarcinoma With or Without CCX872-B. ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT03778879
Nielsen, S. R., Schmid, M. C. (2017). Macrophages as Key Drivers of Cancer Progression and Metastasis. Mediators Inflamm. 2017. 9624760, 1–11. doi: 10.1155/2017/9624760
Nishie, A., Ono, M., Shono, T., Fukushi, J., Otsubo, M., Onoue, H., et al. (1999). Macrophage Infiltration and Heme Oxygenase-1 Expression Correlate with Angiogenesis in Human Gliomas. Clin. Cancer Res. 5, 1107 LP–1113.
Nowak, A. K., Millward, M. J., Cook, A. M., McDonnell, A. M., Creaney, J., Lake, R. A., et al. (2015). A phase 1b clinical trial of the CD40-activating antibody CP-870,893 in combination with cisplatin and pemetrexed in malignant pleural mesothelioma. Ann. Oncol. 26, 2483–2490. doi: 10.1093/annonc/mdv387
Noy, R., Pollard, J. W. (2014). Tumor-Associated Macrophages: From Mechanisms to Therapy. Immunity 41, 49–61. doi: 10.1016/j.immuni.2014.06.010
Nywening, T. M., Wang-Gillam, A., Sanford, D. E., Belt, B. A., Panni, R. Z., Cusworth, B. M., et al. (2016). Targeting tumour-associated macrophages with CCR2 inhibition in combination with FOLFIRINOX in patients with borderline resectable and locally advanced pancreatic cancer: a single-centre, open-label, dose-finding, non-randomised, phase 1b trial. Lancet Oncol. 17, 651–662. doi: 10.1016/S1470-2045(16)00078-4
Okubo, M., Kioi, M., Nakashima, H., Sugiura, K., Mitsudo, K., Aoki, I., et al. (2016). M2-polarized macrophages contribute to neovasculogenesis, leading to relapse of oral cancer following radiation. Sci. Rep. 6, 1–12. doi: 10.1038/srep27548
Paterson, A. H. G., Anderson, S. J., Lembersky, B. C., Fehrenbacher, L., Falkson, C. I., King, K. M., et al. (2012). Oral clodronate for adjuvant treatment of operable breast cancer (National Surgical Adjuvant Breast and Bowel Project protocol B-34): a multicentre, placebo-controlled, randomised trial. Lancet Oncol. 13, 734–742. doi: 10.1016/S1470-2045(12)70226-7
Perry, C. J., et al. (2018). Myeloid-targeted immunotherapies act in synergy to induce inflammation and antitumor immunity. J. Exp. Med. 215, 877–893. doi: 10.1084/jem.20171435
Perry, C. J., Muñoz-Rojas, A. R., Meeth, K. M., Kellman, L. N., Amezquita, R. A., Thakral, D., et al. (2013). Phase 2 study of carlumab (CNTO 888), a human monoclonal antibody against CC-chemokine ligand 2 (CCL2), in metastatic castration-resistant prostate cancer. Invest. New Drugs 31, 760–768. doi: 10.1007/s10637-012-9869-8
Poh, A. R., Ernst, M. (2018). Targeting Macrophages in Cancer: From Bench to Bedside. Front. Oncol. 8, 49. doi: 10.3389/fonc.2018.00049
Poon, C. C., Sarkar, S., Yong, V. W., Kelly, J. J. P. (2017). Glioblastoma-associated microglia and macrophages: Targets for therapies to improve prognosis. Brain 140, 1548–1560. doi: 10.1093/brain/aww355
Pyonteck, S. M., Akkari, L., Schuhmacher, A. J., Bowman, R. L., Sevenich, L., Quail, D. F., et al. (2013). CSF-1R inhibition alters macrophage polarization and blocks glioma progression. Nat. Med. 19, 1264. doi: 10.1038/nm.3337
Qian, J., Luo, F., Yang, J., Liu, J., Liu, R., Wang, L., et al. (2018). TLR2 Promotes Glioma Immune Evasion by Downregulating MHC Class II Molecules in Microglia. Cancer Immunol. Res. 6, 1220 LP–1233. doi: 10.1158/2326-6066.CIR-18-0020
Quail, D. F., Joyce, J. A. (2013). Microenvironmental regulation of tumor progression and metastasis. Nat. Med. 19, 1423–1437. doi: 10.1038/nm.3394
Quail, D. F., Joyce, J. A. (2017). The Microenvironmental Landscape of Brain Tumors. Cancer Cell 31, 326–341. doi: 10.1016/j.ccell.2017.02.009
Manda, S. V., Kataria, Y., Tatireddy, B. R., Ramakrishnan, B., Ratnam, B. G., Lath, R., et al. (2018). Exosomes as a biomarker platform for detecting epidermal growth factor receptor–positive high-grade gliomas. J. Neurosurg. 128, 1091–1101. doi: 10.3171/2016.11.jns161187
Roesch, S., Rapp, C., Dettling, S., Herold-Mende, C. (2018). When Immune Cells Turn Bad-Tumor-Associated Microglia/Macrophages in Glioma. Int. J. Mol. Sci. 19, 436. doi: 10.3390/ijms19020436
Rossi, M. L., Hughes, J. T., Esiri, M. M., Coakham, H. B., Brownell, D. B. (1987). Immunohistological study of mononuclear cell infiltrate in malignant gliomas. Acta Neuropathol. 74, 269–277. doi: 10.1007/BF00688191
Ruzsa, A., Sen, M., Evans, M., Lee, L. W., Hideghety, K., Rottey, S., et al. (2014). Phase 2, open-label, 1:1 randomized controlled trial exploring the efficacy of EMD 1201081 in combination with cetuximab in second-line cetuximab-naïve patients with recurrent or metastatic squamous cell carcinoma of the head and neck (R/M SCCHN). Invest. New Drugs 32, 1278–1284. doi: 10.1007/s10637-014-0117-2
Séhédic, D., Chourpa, I., Tétaud, C., Griveau, A., Loussouarn, C., Avril, S., et al. (2017). Locoregional confinement and major clinical benefit of188re-loaded CXCR4-targeted nanocarriers in an orthotopic human to mouse model of glioblastoma. Theranostics 7, 4517–4536. doi: 10.7150/thno.19403
Sørensen, M. D., Dahlrot, R. H., Boldt, H. B., Hansen, S., Kristensen, B. W. (2018). Tumour-associated microglia/macrophages predict poor prognosis in high-grade gliomas and correlate with an aggressive tumour subtype. Neuropathol. Appl. Neurobiol. 44, 185–206. doi: 10.1111/nan.12428
Sadahiro, H., Kang, K.-D., Gibson, J. T., Minata, M., Yu, H., Shi, J., et al. (2018). Activation of the Receptor Tyrosine Kinase AXL Regulates the Immune Microenvironment in Glioblastoma. Cancer Res. 78, 3002 LP–3013. doi: 10.1158/0008-5472.CAN-17-2433
Saha, D., Martuza, R. L., Rabkin, S. D. (2017). Macrophage Polarization Contributes to Glioblastoma Eradication by Combination Immunovirotherapy and Immune Checkpoint Blockade. Cancer Cell 32, 253–267.e5. doi: 10.1016/j.ccell.2017.07.006
Sandhu, S. K., Papadopoulos, K., Fong, P. C., Patnaik, A., Messiou, C., Olmos, D., et al. (2013). A first-in-human, first-in-class, phase i study of carlumab (CNTO 888), a human monoclonal antibody against CC-chemokine ligand 2 in patients with solid tumors. Cancer Chemother. Pharmacol. 71, 1041–1050. doi: 10.1007/s00280-013-2099-8
Schiffer, D., Annovazzi, L., Casalone, C., Corona, C., Mellai, M. (2018). Glioblastoma: Microenvironment and Niche Concept. Cancers 11 (5), 1–17. doi: 10.3390/cancers11010005
Shi, Y., Ping, Y.-F., Zhou, W., He, Z.-C., Chen, C., Bian, B.-S.-J., et al. (2017). Tumour-associated macrophages secrete pleiotrophin to promote PTPRZ1 signalling in glioblastoma stem cells for tumour growth. Nat. Commun. 8, 15080. doi: 10.1038/ncomms15080
Shoji, T., Saito, R., Chonan, M., Shibahara, I., Sato, A., Kanamori, M., et al. (2016). Local convection-enhanced delivery of an anti-CD40 agonistic monoclonal antibody induces antitumor effects in mouse glioma models. Neuro. Oncol. 18, 1120–1128. doi: 10.1093/neuonc/now023
Sica, A., Erreni, M., Allavena, P., Porta, C. (2015). Macrophage polarization in pathology. Cell. Mol. Life Sci. 72, 4111–4126. doi: 10.1007/s00018-015-1995-y
Sikic, B. I., Lakhani, N. J., Patnaik, A., Shah, S., Chandana, S. R., Rasco, D. W., et al. (2018). A first-in-class, first-in-human phase 1 pharmacokinetic (PK) and pharmacodynamic (PD) study of Hu5F9-G4, an anti-CD47 monoclonal antibody (mAb), in patients with advanced solid tumors. J. Clin. Oncol. 36, 3002. doi: 10.1200/JCO.2018.36.15_suppl.3002
Smith, D. A., Conkling, P., Richards, D. A., Nemunaitis, J. J., Boyd, T. E., Mita, A. C., et al. (2014). Antitumor activity and safety of combination therapy with the Toll-like receptor 9 agonist IMO-2055, erlotinib, and bevacizumab in advanced or metastatic non-small cell lung cancer patients who have progressed following chemotherapy. Cancer Immunol. Immunother. 63, 787–796. doi: 10.1007/s00262-014-1547-6
Strachan, D. C., Ruffell, B., Oei, Y., Bissell, M. J., Coussens, L. M., Pryer, N., et al. (2013). CSF1R inhibition delays cervical and mammary tumor growth in murine models by attenuating the turnover of tumor-associated macrophages and enhancing infiltration by CD8+T cells. Oncoimmunology 2, 1–12. doi: 10.4161/onci.26968
Stratton, J. A., Holmes, A., Rosin, N. L., Sinha, S., Vohra, M., Burma, N. E., et al. (2018). Macrophages Regulate Schwann Cell Maturation after Nerve Injury. Cell Rep. 24, 2561–2572.e6. doi: 10.1016/j.celrep.2018.08.004
Stupp, R., Mason, W. P., van den Bent, M. J., Weller, M., Fisher, B., Taphoorn, M. J. B., et al. (2005). Radiotherapy plus Concomitant and Adjuvant Temozolomide for Glioblastoma. N. Engl. J. Med. 352, 987–996. doi: 10.1056/NEJMoa043330
Stupp, R., Hegi, M. E., Mason, W. P., van den Bent, M. J., Taphoorn, M. J. B., Janzer, R. C., et al. (2009). Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomised phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet Oncol. 10, 459–466. doi: 10.1016/S1470-2045(09)70025-7
Szulzewsky, F., Pelz, A., Feng, X., Synowitz, M., Markovic, D., Langmann, T., et al. (2015). Glioma-associated microglia/macrophages display an expression profile different from M1 and M2 polarization and highly express Gpnmb and Spp1. PloS One 10, e0116644–e0116644. doi: 10.1371/journal.pone.0116644
Tabatabaei, P., Visse, E., Bergström, P., Brännström, T., Siesjö, P., Bergenheim, A. T. (2017). Radiotherapy induces an immediate inflammatory reaction in malignant glioma: a clinical microdialysis study. J. Neurooncol. 131, 83–92. doi: 10.1007/s11060-016-2271-1
Tamura, R., Ohara, K., Sasaki, H., Morimoto, Y., Kosugi, K., Yoshida, K., et al. (2018). Difference in Immunosuppressive Cells Between Peritumoral Area and Tumor Core in Glioblastoma. World Neurosurg. 120, e601–e610. doi: 10.1016/j.wneu.2018.08.133
Tap, W. D., Wainberg, Z. A., Anthony, S. P., Ibrahim, P. N., Zhang, C., Healey, J. H., et al. (2015). Structure-Guided Blockade of CSF1R Kinase in Tenosynovial Giant-Cell Tumor. N. Engl. J. Med. 373, 428–437. doi: 10.1056/NEJMoa1411366
Thomas, A. C., Mattila, J. T. (2014). ‘Of mice and men’: arginine metabolism in macrophages. Front. Immunol. 5, 479. doi: 10.3389/fimmu.2014.00479
Turkowski, K., Brandenburg, S., Mueller, A., Kremenetskaia, I., Bungert, A. D., Blank, A., et al. (2018). VEGF as a modulator of the innate immune response in glioblastoma. Glia 66, 161–174. doi: 10.1002/glia.23234
Van Acker, H. H., Anguille, S., Willemen, Y., Smits, E. L., Van Tendeloo, V. F. (2016). Bisphosphonates for cancer treatment: Mechanisms of action and lessons from clinical trials. Pharmacol. Ther. 158, 24–40. doi: 10.1016/j.pharmthera.2015.11.008
Van Der Vos, K. E., Abels, E. R., Zhang, X., Lai, C., Carrizosa, E., Oakley, D., et al. (2016). Directly visualized glioblastoma-derived extracellular vesicles transfer RNA to microglia/macrophages in the brain. Neuro. Oncol. 18, 58–69. doi: 10.1093/neuonc/nov244
Vonderheide, R. H., Burg, J. M., Mick, R., Trosko, J. A., Li, D., Shaik, M. N., et al. (2013). Phase i study of the CD40 agonist antibody CP-870,893 combined with carboplatin and paclitaxel in patients with advanced solid tumors. Oncoimmunology 2, 1–10. doi: 10.4161/onci.23033
Wagner, S., Czub, S., Greif, M., Vince, G. H., Suss, N., Kerkau, S., et al. (1999). Microglial/macrophage expression of interleukin 10 in human glioblastomas. Int. J. Cancer 82, 12–16. doi: 10.1002/(SICI)1097-0215(19990702)82:1<12::AID-IJC3>3.0.CO;2-O
Walentynowicz, K. A., Ochocka, N., Pasierbinska, M., Wojnicki, K., Stepniak, K., Mieczkowski, J., et al. (2018). In Search for Reliable Markers of Glioma-Induced Polarization of Microglia. Front. Immunol. 9, 1329. doi: 10.3389/fimmu.2018.01329
Wang, S. C., Yu, C. F., Hong, J. H., Tsai, C. S., Chiang, C. S. (2013). Radiation Therapy-Induced Tumor Invasiveness Is Associated with SDF-1-Regulated Macrophage Mobilization and Vasculogenesis. PloS One 8 (e69182), 1–10. doi: 10.1371/journal.pone.0069182
Wang, Y., Liu, T., Yang, N., Xu, S., Li, X., Wang, D. (2016). Hypoxia and macrophages promote glioblastoma invasion by the CCL4-CCR5 axis. Oncol. Rep. 36, 3522–3528. doi: 10.3892/or.2016.5171
Wang, Q., He, Z., Huang, M., Liu, T., Wang, Y., Xu, H., et al. (2018). Vascular niche IL-6 induces alternative macrophage activation in glioblastoma through HIF-2α. Nat. Commun. 9, 559. doi: 10.1038/s41467-018-03050-0
Won, W.-J., Deshane, J. S., Leavenworth, J. W., Oliva, C. R., Griguer, C. E. (2019). Metabolic and functional reprogramming of myeloid-derived suppressor cells and their therapeutic control in glioblastoma. Cell Stress. 3, 47–65 doi: 10.15698/cst2019.02.176
Wu, J., Frady, L. N., Bash, R. E., Cohen, S. M., Schorzman, A. N., Su, Y.-T., et al. (2018). MerTK as a therapeutic target in glioblastoma. Neuro. Oncol. 20, 92–102. doi: 10.1093/neuonc/nox111
Wynn, T. A., Chawla, A., Pollard, J. W. (2013). Origins and Hallmarks of Macrophages: Development, Homeostasis, and Disease. Nature 496, 445–455. doi: 10.1038/nature12034
Xue, N., Zhou, Q., Ji, M., Jin, J., Lai, F., Chen, J., et al. (2017). Chlorogenic acid inhibits glioblastoma growth through repolarizating macrophage from M2 to M1 phenotype. Sci. Rep. 7, 39011. doi: 10.1038/srep39011
Yan, D., Kowal, J., Akkari, L., Schuhmacher, A. J., Huse, J. T., West, B. L., et al. (2017). Inhibition of colony stimulating factor-1 receptor abrogates microenvironment-mediated therapeutic resistance in gliomas. Oncogene 36, 6049–6058. doi: 10.1038/onc.2017.261
Ye, X., Xu, S., Xin, Y., Yu, S., Ping, Y., Chen, L., et al. (2012). Tumor-Associated Microglia/Macrophages Enhance the Invasion of Glioma Stem-like Cells via TGF-β1 Signaling Pathway. J. Immunol. 189, 444 LP–44 453. doi: 10.4049/jimmunol.1103248
Yin, Y., Qiu, S., Li, X., Huang, B., Xu, Y., Peng, Y. (2017). EZH2 suppression in glioblastoma shifts microglia toward M1 phenotype in tumor microenvironment. J. Neuroinflammation 14, 220. doi: 10.1186/s12974-017-0993-4
Yuan, D., Zhao, Y., Banks, W. A., Bullock, K. M., Haney, M., Batrakova, E., et al. (2017). Macrophage exosomes as natural nanocarriers for protein delivery to inflamed brain. Biomaterials 142, 1–12. doi: 10.1016/j.biomaterials.2017.07.011
Zeiner, P. S., Preusse, C., Golebiewska, A., Zinke, J., Iriondo, A., Muller, A., et al. (2018). Distribution and prognostic impact of microglia/macrophage subpopulations in gliomas. Brain Pathol. 29, 513–529. doi: 10.1111/bpa.12690
Zhang, Q., et al. (2012). Prognostic significance of tumor-associated macrophages in solid tumor: a meta-analysis of the literature. PloS One 7, e50946–e50946. doi: 10.1371/journal.pone.0050946
Zhang, H., Lu, H., Xiang, L., Bullen, J. W., Zhang, C., Samanta, D., et al. (2015). HIF-1 regulates CD47 expression in breast cancer cells to promote evasion of phagocytosis and maintenance of cancer stem cells. Proc. Natl. Acad. Sci. 112, E6215–E6223. doi: 10.1073/pnas.1520032112
Zhang, M., Hutter, G., Kahn, S. A., Azad, T. D., Gholamin, S., Xu, C. Y., et al. (2016). Anti-CD47 Treatment Stimulates Phagocytosis of Glioblastoma by M1 and M2 Polarized Macrophages and Promotes M1 Polarized Macrophages In Vivo. PloS One 11, e0153550–e0153550. doi: 10.1371/journal.pone.0153550
Zhang, G., Zhang, Y., Cheng, S., Wu, Z., Liu, F., Zhang, J. (2017). CD133 positive U87 glioblastoma cells-derived exosomal microRNAs in hypoxia- versus normoxia-microenviroment. J. Neurooncol. 135, 37–46. doi: 10.1007/s11060-017-2566-x
Zhao, H., Wang, J., Kong, X., Li, E., Liu, Y., Du, X., et al. (2016). CD47 promotes tumor invasion and metastasis in non-small cell lung cancer. Sci. Rep. 6 (29719), 1–11. doi: 10.1038/srep29719
Zhao, X., Qu, J., Sun, Y., Wang, J., Liu, X., Wang, F., et al. (2017). Prognostic significance of tumor-associated macrophages in breast cancer: a meta-analysis of the literature. Oncotarget 8, 30576–30586. doi: 10.18632/oncotarget.15736
Zheng, Y., Yang, W., Aldape, K., He, J., Lu, Z. (2013). Epidermal growth factor (EGF)-enhanced vascular cell adhesion molecule-1 (VCAM-1) expression promotes macrophage and glioblastoma cell interaction and tumor cell invasion. J. Biol. Chem. 288, 31488–31495. doi: 10.1074/jbc.M113.499020
Zhernakova, A., Garmaeva, S., Fu, J., Chen, L., Wijmenga, C. (2018). A system biology perspective on environment–host–microbe interactions. Hum. Mol. Genet. 27, R187–R194. doi: 10.1093/hmg/ddy137
Zhou, M., Bracci, P. M., McCoy, L. S., Hsuang, G., Wiemels, J. L., Rice, T., et al. (2015). Serum macrophage-derived chemokine/CCL22 levels are associated with glioma risk, CD4 T cell lymphopenia and survival time. Int. J. Cancer 137, 826–836. doi: 10.1002/ijc.29441
Zhou, W., Ke, S. Q., Huang, Z., Flavahan, W., Fang, X., Paul, J., et al. (2015). Periostin secreted by glioblastoma stem cells recruits M2 tumour-associated macrophages and promotes malignant growth. Nat. Cell Biol. 17, 170–182. doi: 10.1038/ncb3090
Zhou, J., Reddy, M. V., Wilson, B. K. J., Blair, D. A., Taha, A., Frampton, C. M., et al. (2018). MR Imaging Characteristics Associate with Tumor-Associated Macrophages in Glioblastoma and Provide an Improved Signature for Survival Prognostication. Am. J. Neuroradiol. 39, 252 LP–25 259. doi: 10.3174/ajnr.A5441
Zhu, C., Chrifi, I., Mustafa, D., Van Der Weiden, M., Leenen, P. J. M., Duncker, D. J., et al. (2017). CECR1-mediated cross talk between macrophages and vascular mural cells promotes neovascularization in malignant glioma. Oncogene 36, 5356–5368. doi: 10.1038/onc.2017.145
Keywords: glioblastoma, macrophages, microglia, resistance, radiation, crosstalks, tumor-associated macrophage
Citation: Grégoire H, Roncali L, Rousseau A, Chérel M, Delneste Y, Jeannin P, Hindré F and Garcion E (2020) Targeting Tumor Associated Macrophages to Overcome Conventional Treatment Resistance in Glioblastoma. Front. Pharmacol. 11:368. doi: 10.3389/fphar.2020.00368
Received: 23 December 2019; Accepted: 10 March 2020;
Published: 08 April 2020.
Edited by:
Robert Clarke, Georgetown University, United StatesReviewed by:
David Soto-Pantoja, Wake Forest School of Medicine, United StatesBiana Godin, Houston Methodist Research Institute, United States
Copyright © 2020 Grégoire, Roncali, Rousseau, Chérel, Delneste, Jeannin, Hindré and Garcion. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Emmanuel Garcion, ZW1tYW51ZWwuZ2FyY2lvbkB1bml2LWFuZ2Vycy5mcg==