
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Pharmacol. , 16 October 2018
Sec. Experimental Pharmacology and Drug Discovery
Volume 9 - 2018 | https://doi.org/10.3389/fphar.2018.01146
 Karolina Pytka1*
Karolina Pytka1* Monika Głuch-Lutwin2
Monika Głuch-Lutwin2 Elżbieta Żmudzka1
Elżbieta Żmudzka1 Kinga Sałaciak1Agata Siwek2
Kinga Sałaciak1Agata Siwek2 Katarzyna Niemczyk1Maria Walczak3Magdalena Smolik3
Katarzyna Niemczyk1Maria Walczak3Magdalena Smolik3 Adrian Olczyk4
Adrian Olczyk4 Adam Gałuszka4
Adam Gałuszka4 Jarosław Śmieja5Barbara Filipek1Jacek Sapa1Marcin Kołaczkowski6
Jarosław Śmieja5Barbara Filipek1Jacek Sapa1Marcin Kołaczkowski6 Katarzyna Pańczyk7
Katarzyna Pańczyk7 Anna Waszkielewicz7Henryk Marona7
Anna Waszkielewicz7Henryk Marona7Numerous studies have proven that both stimulation and blockade of 5-HT1A and the blockade of 5-HT7 receptors might cause the anxiolytic-like effects. Biased agonists selectively activate specific signaling pathways. Therefore, they might offer novel treatment strategies. In this study, we investigated the anxiolytic-like activity, as well as the possible mechanism of action of 1-[(2,5-dimethylphenoxy)propyl]-4-(2-methoxyphenyl)piperazine hydrochloride (HBK-17). In our previous experiments, HBK-17 showed high affinity for 5-HT1A and 5-HT7 receptors and antidepressant-like properties. We performed the four plate test and the elevated plus maze test to determine anxiolytic-like activity. Toward a better understanding of the pharmacological properties of HBK-17 we used various functional assays to determine its intrinsic activity at 5-HT1A, 5-HT2A, 5-HT7, and D2 receptors and UHPLC-MS/MS method to evaluate its pharmacokinetic profile. We observed the anxiolytic-like activity of HBK-17 in both behavioral tests and the effect was reversed by the pretreatment with WAY-100635, which proves that 5-HT1A receptor activation was essential for the anxiolytic-like effect. Moreover, the compound moderately antagonized D2, weakly 5-HT7 and very weakly 5-HT2A receptors. We demonstrated that HBK-17 preferentially activated ß-arrestin signaling after binding to the 5-HT1A receptor. HBK-17 was rapidly absorbed after intraperitoneal administration and had a half-life of about 150 min. HBK-17 slightly penetrated the peripheral compartment and showed bioavailability of approximately 45%. The unique pharmacological profile of HBK-17 encourages further experiments to understand its mechanism of action fully.
Brain serotonin participates in numerous physiological and pathological processes. It regulates mood, sleep, or cognitive function and modulates fear and anxiety processes. Serotonin exerts its effects interacting with at least 14 serotonin receptor subtypes. Among all of them, the 5-HT1A receptor plays the most important role in the etiology of anxiety (reviewed by Akimova et al., 2009). The 5-HT1A receptors are expressed both presynaptically and postsynaptically. The activation of presynaptic 5-HT1A autoreceptors, present on serotonergic neurons in the raphe nuclei, reduces serotonergic neurons firing and decreases serotonin levels. On the other hand, the stimulation of postsynaptic 5-HT1A receptors, located mainly on glutamatergic and GABAergic pyramidal neurons, modulates serotonergic sensitivity and participates in emotional and cognitive processes (for review see Garcia-Garcia et al., 2014).
Garcia-Garcia et al. (2014) in their review suggested that inhibiting serotonergic neuron firing and decreasing serotonin release, as well as reducing serotonergic signaling at postsynaptic target receptors might result in anxiolytic-like effects. Studies confirmed this theory, as stimulating 5-HT1A receptors in the dorsal raphe nucleus caused an anxiolytic-like effect (Cervo et al., 2000), whereas activating the heteroreceptors in the medial septum and dorsal hippocampus was anxiogenic (File et al., 1996; De Almeida et al., 1998). Collinson and Dawson (1997) presented the anxiolytic-like effect of 5-HT1A receptor agonist, 8-OH-DPAT, which was reversed by 5-HT1A receptor antagonist (WAY-100635). Moreover, 5-HT1A knockout mice, lacking 5-HT1A auto- and heteroreceptors, displayed a heightened anxiety-like phenotype in several tests in rodents (Heisler et al., 1998; Parks et al., 1998; Ramboz et al., 1998) and resistance to benzodiazepines (Sibille et al., 2000). Overall, the above findings demonstrate the involvement of 5-HT1A receptors in anxiogenic processes.
The blockade of 5-HT7 receptors might also be beneficial in anxiety. Yau et al. (2001) showed an increase in 5-HT7 receptor mRNA expression after acute, but not chronic, stress in the rat hippocampus, which suggested the role of the 5-HT7 receptor in stress regulation. Wesołowska et al. (2006b) demonstrated the anxiolytic-like activity of a selective 5-HT7 receptor antagonist, SB-269970, in rodent models of anxiety. Similarly, intrahippocampal administration of the compound induced an antianxiety-like effect in the Vogel conflict test in rats (Wesołowska et al., 2006a). Thus, some studies suggest that 5-HT7 receptor antagonists might decrease anxiety symptoms.
Functional selectivity (also called biased signaling) is an ability of a ligand to direct a G protein-coupled receptor toward a conformation that activates specific signal transduction pathway. Upon stimulation, the 5-HT1A receptor can activate many signaling pathways, including adenylate cyclase inhibition, extracellular signal-regulated protein kinase (ERK) phosphorylation, G protein-coupled inwardly rectifying potassium channel activation, voltage-gated calcium channel inhibition, or the recruitment of ß-arrestin (Chilmonczyk et al., 2015; Stroth et al., 2015). Newman-Tancredi et al. (2009) presented compounds highly selective for a 5-HT1A receptor with biased agonist profiles, i.e., F15599 or F13714. F15599 showed a marked potency for ERK1/2 phosphorylation, which might underlie its potent antidepressant-like activity in rats. On the other hand, F13714, which presented lower potency for ERK1/2 phosphorylation (Newman-Tancredi, 2011), showed an exceptionally potent antidyskinetic activity in rats (Iderberg et al., 2015). This demonstrates that biased agonists at the 5-HT1A receptor might present different pharmacological activity. Although it is not yet clear to what extent functional selectivity can be exploited for therapeutic advantage, some clinically used drugs, e.g., carvedilol (ß-blocker) show this effect (Patel et al., 2010; Kenakin, 2011). Moreover, scientists suggest that functional selectivity can explain the unique features of the antipsychotic drug – aripiprazole (Mailman and Murthy, 2010). Since biased agonists preferentially activate one pathway over others, they might offer novel treatment strategies, i.e., show pharmacological activity without inducing unwanted effects. Thus, compounds with functional selectivity might show interesting pharmacological properties in vivo and therefore are worth investigating.
We previously demonstrated antidepressant-like activity of 1-[(2,5-dimethylphenoxy)propyl]-4-(2-methoxyphenyl)piperazine hydrochloride (HBK-17, Figure 1) in rodents (Waszkielewicz et al., 2015; Kubacka et al., 2016; Pytka et al., 2016a). The compound moderately antagonized 5-HT1A receptors expressed in CHO-K1 cells in Ca2+ mobilization assay and showed the affinity for 5-HT7 receptors (Waszkielewicz et al., 2015; Kubacka et al., 2016; Pytka et al., 2016a). Since 5-HT1A and 5-HT7 receptors play a role in anxiety, we hypothesized that HBK-17 might influence anxiety-like behaviors in rodents. Therefore, our study aimed to investigate anxiolytic-like properties in mice, as well as the possible mechanism of action of HBK-17 using in vitro and in vivo experiments. As knowledge of pharmacokinetics and brain distribution of novel central-acting compounds is essential for the proper analysis of their in vivo effects, we also evaluated the compound’s pharmacokinetic profile.
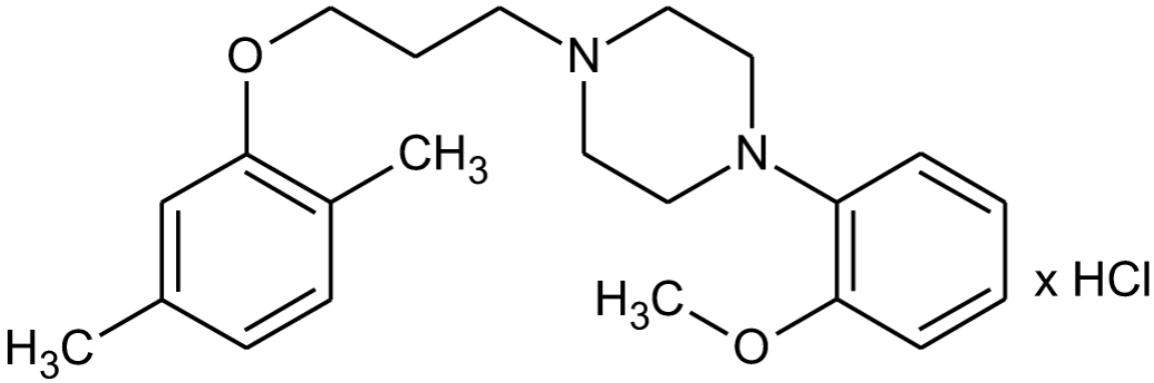
FIGURE 1. Chemical structure of HBK-17.
We used adult male Albino-Swiss mice (CD-1) weighing 18–21 g in all experiments. The animals were kept in groups of 15 mice in cages at room temperature of 22 ± 2°C under light/dark (12:12) cycle and they had free access to food (standard laboratory pellets) and water before experiments. Humidity and ambient temperature of the room were kept constant throughout all tests, which were conducted between 9 a.m. and 4 p.m. The animals were used only once in each test. All injections were given in a volume of 10 ml/kg. Behavioral experiments were carried out by a trained observer blind to the treatments. All experimental procedures were approved by the I Local Ethics Committee for Experiments on Animals of the Jagiellonian University in Krakow (approval number 51/2015), Poland and performed under the guidelines provided by the European Union Directive of 22 September 2010 (2010/63/EU) and Polish legislation concerning animal experimentation.
The tested compound 1-[(2,5-dimethylphenoxy)propyl]-4-(2-methoxyphenyl)piperazine hydrochloride (HBK-17) was synthesized in the Department of Bioorganic Chemistry, Chair of Organic Chemistry, Jagiellonian University Medical College (Figure 1; Waszkielewicz et al., 2015). HBK-17 was dissolved in saline and administered intraperitoneally (i.p.) 30 min before behavioral experiments. p-Chlorophenylalanine (pCPA, Sigma, Germany) and α-methyl-p-tyrosine (AMPT, Sigma, Germany) were suspended in 1% Tween and administered i.p. WAY-100635 (Sigma, Germany) was injected subcutaneously (s.c.) 15 min before the studied compound.
Binding experiments were performed using membranes from CHO-K1 cells stably transfected with the human D2 receptor and rat cerebral cortex (α2-adrenergic and GABAA receptors). [3H]-Methylspiperon (final concentration 0.4 nM; D2 receptor), [3H]-clonidine (final concentration 0.2 nM; α2-adrenergic receptor) and [3H]-muscimol (final concentration 1 nM; GABAA receptor) were used. The final incubation mixture (total volume 250 μl) consisted of a 50 μl solution of the test compound, 50 μl of radioligand and 150 μl of diluted membranes or the tissue suspension. The incubation was terminated by rapid filtration over glass fiber filters GF/B (PerkinElmer, United States) using 96-well FilterMate Harvester (PerkinElmer, United States). Five rapid washes were performed with ice-cold 50 mM Tris–HCl buffer (pH 7.4 or 7.6). The radioactivity was measured in MicroBeta TriLux 1450 – liquid scintillation counter (PerkinElmer, United States). Radioligand binding data were analyzed using iterative curve fitting routines GraphPad Prism 5.0 (GraphPad Software). Ki values were calculated from the Cheng and Prusoff (1973) equation. The concentrations of analyzed compounds ranged from 10−10 to 10−5 M. For measuring unspecific binding, clonidine – 10 μM (in case of [3H]-clonidine) and haloperidol – 1 μM (in case of [3H]-methylspiperon) and diazepam – 100 μM (in case of [3H]-muscimol) were applied. At least two independent experiments were performed for each assay.
Serial dilutions were prepared in 96-well microplate in appropriate dilution buffer (HBSS or medium) with 0.1% BSA added, and 8–10 concentrations were tested in duplicates. At least two independent experiments were performed for each type of method. Assays were done according to manufacturer’s protocols.
The level of cAMP (TRF0263, PerkinElmer assay kit) was monitored using CHO-K1 cells with expression of the human serotonin 5-HT1A receptor. The cells were resuspended in stimulation buffer at 2 × 105 cells/ml. The same volume (10 μl) of cells was added to tested compounds with 10 μM forskolin. The antagonist response was performed using as a reference agonist, serotonin, in EC80 (120 nM). Samples were loaded onto a white opaque half area 96-well microplate, incubated for 40 min at room temperature. The 10 μl of reagents were added, mixed, and incubated for 1 h. The homogeneous TR-FRET signal was read on an EnVision Microplate Reader (PerkinElmer, United States).
The CHO-5HT1A receptor cells were tested for phosphorylated-ERK (p-ERK) using the SureFire ERK-Phosphorylation AlphaLISA Assay Kit according to the manufacturer’s instruction (ALSU-PERK-A10K, PerkinElmer). At the experiment, cells were plated at 50,000 cells/well of 96-well plate and grown 7 h in an incubator (5% CO2, 37°C). The serial dilutions of compounds were added and incubated for 15 min at 37°C. The antagonist response was performed using serotonin as a reference agonist at the concentration equal to EC80 (45 nM). The “lysis buffer” (70 μl) was added and the plate gently agitated on a plate shaker (10 min). The 10 μl of samples were transferred to assay plates (384-OptiPlate, PerkinElmer) in duplicates and 10 μl of the reaction mix was added. The plates were incubated for 2 h at 22°C and measured with an EnVision a multifunction plate reader (PerkinElmer Life Science).
The HTR1A-bla U2OS receptor cells were tested using the Tango LiveBLAzer β-arrestin assay kit according to the manufacturer’s instruction (K1095, Life Technologies). At the experiment, cells were plated at 10,000 cells/well of 384-well black, clear bottom plate and grown 12 h in an incubator (5% CO2, 37°C). The serial dilutions of compounds were added and incubated for 5 h (5% CO2, 37°). The antagonist response was performed using serotonin as a reference agonist at the concentration equal to EC80 (300 nM). After this time, 8 μl of the reaction mix was added. The plates were incubated for 2 h at 22°C and measured with a FLUOstar OPTIMA a multifunction plate reader (PerkinElmer Life Science).
The four plate test was performed on mice according to the method previously described (Aron et al., 1971; Bourin et al., 2005; Pytka et al., 2016b). Mice were placed individually in the four plate apparatus connected to the power source. After a 15 s of habituation period, each mouse crossing from one plate to another (two limbs on one plate, two on another) was punished by an electric shock (0.8 mA, 0.5 s). If the mouse continued running, it received no new shock for the following 3 s. The number of punished crossings was calculated during the 60 s of the test. HBK-17 was dissolved in saline and administered i.p. 30 min before the test.
The elevated plus maze was performed according to the method previously described (Lister, 1987; Pytka et al., 2017). The elevated plus maze for mice consisted of two opposing open (30 cm × 5 cm), and two enclosed arms (30 cm × 5 cm × 25 cm) connected by a central platform forming the shape of a plus sign. The open and closed arms were connected with a central field (5 cm × 5 cm). Each mouse was individually placed at the central field of the apparatus with the head turned toward one of the closed arms. Animal behavior was observed for 5 min. The device was disinfected with 70% ethanol after each mouse. The number of entries to open and closed arms and time spent in the open and closed arms were measured. The experiments were recorded and scored using aLab.io software by a trained observer blind to the treatments. HBK-17 was dissolved in saline and administered i.p. 30 min before the test.
Spontaneous locomotor activity was performed as previously described (Pytka et al., 2015a). Photoresistor actometers (Ugo Basile, Italy) connected to a counter for the recording of light-beam interruptions were used to investigate the effect of the compound on locomotor activity in mice. Locomotor activity was counted as the number of the light-beam crossing. It was recorded individually for each animal. After administration of the tested compound, each mouse was placed separately in a cage for 30 min habituation period. After that time, the parameter was measured for 1 min and 5 min (i.e., time equal to the observation period in four plate test and elevated plus maze test, respectively). The cages were sanitized with 70% ethanol after each mouse. HBK-17 was dissolved in saline and administered i.p. 30 min before the test.
The hot plate test was performed as described by Sałat et al. (2015). The hot plate apparatus (Hot/Cold Plate, Bioseb, France) consisted of an electrically heated surface and was equipped with a temperature controller that keeps the temperature constant at 55–56°C. One day before the experiment, the animals were tested for their pain sensitivity threshold (baseline latency). We chose only mice that showed baseline latencies ≤30 s. The latency time to pain reaction (licking hind paws or jumping) was measured as the indicative of nociception (cutoff time – 60 s). Animals that did not respond within 60 s were removed from the hot plate apparatus and assigned a score of 60 s. HBK-17 was dissolved in saline and administered i.p. 30 min before the test.
To assess the involvement of the serotonergic system in the anxiolytic-like effect of HBK-17, we used pCPA (tryptophan hydroxylase inhibitor) as previously described (Pytka et al., 2015b). We injected mice with pCPA at a dose of 200 mg/kg for three consecutive days. 24 h after the last pCPA administration, we injected mice with saline (i.p.) or HBK-17 (5 mg/kg, i.p.). We performed the four plate test 30 min after the administration.
To investigate the involvement of the noradrenergic and dopaminergic systems in the anxiolytic-like effect of HBK-17, we pretreated mice i.p. with a catecholamine synthesis inhibitor, AMPT (tyrosine hydroxylase inhibitor) at the dose of 100 mg/kg. Control group received vehicle (1% Tween). Four hours after AMPT or 1% Tween injection, mice were pretreated with saline (i.p.) or HBK-17 (5 mg/kg, i.p.). We performed the four plate test 30 min after the compound administration.
The LC/ESI-MS/MS experiments were performed on TSQ Quantum Triple Quadrupole Mass Spectrometer (Thermo Scientific, United States) equipped with an electrospray ionization interface. This instrument was coupled to Dionex 3000 (Dionex, United States) ULPC system. Data acquisition and processing were accomplished using Xcalibur data collection and integration software. The mobile phase consisted of a mixture of acetonitrile with an addition of 0.1% formic acid (Solvent A) and water with an addition of 0.1% formic acid (Solvent B) was set at a flow rate of 0.3 ml/min in gradient elution. Sample preparations were carried out by precipitation procedure using acetonitrile after the addition of the internal standard [IS, 2-(4-methyl-1-piperazinyl)-4-phenylquinazoline]. The dried residue was reconstituted in the mobile phase and injected onto an Acclaim Polar Advantage Column (1.8 μm, 100 mm × 2.1 mm, Dionex).
The method was validated according to validation procedures, parameters and acceptance criteria based on USP XXIII guidelines and FDA criterion 20/15.
All data in pharmacokinetic experiments were processed with the pharmacokinetics software Phoenix WinNonlin (Certara Company, United States). The non-compartmental pharmacokinetic parameters of t1/2, MRT, AUC, Cl, and Vd were calculated based on moment methods. First order elimination rate constant (λz) was calculated by linear regression of time vs. log concentration according to Eq. 1.
where a is a slope of a line.
The terminal half-life (t1/2) was calculated as:
The area under curve the mean plasma and tissue concentration vs. time curve extrapolated to infinity (AUC0→∞) was estimated using the log-linear trapezoidal rule (Eq. 3), here Cn is the concentration of last sampling.
Area under the first-moment curve (AUMC0→∞) was estimated by calculation of total area under the first-moment curve and extrapolated area using the Eq. 4, where tn is the time of last sampling.
Mean residence time (MRT) was calculated as:
Systemic clearance (Cl) was calculated as:
Volume of distribution (Vd) was calculated as:
were Div is an intravenous dose and C0 is an initial concentration.
The bioavailability (F) of the studied compound after an intraperitoneal administration compared to the intravenous route was calculated as:
Results are presented as means ± SEM. They were estimated using one or two-way analysis of variance (ANOVA), followed by Newman-Keuls or Bonferroni post hoc, respectively. Differences between groups were considered as significant if p < 0.05.
The radioligand binding experiments revealed that compared with the reference compound, haloperidol, HBK-17 moderately bound to D2 receptors. HBK-17 did not show the affinity for α2 adrenoceptors or GABAA receptors, while the reference drugs clonidine and diazepam strongly bound to these receptors, respectively (Table 1).
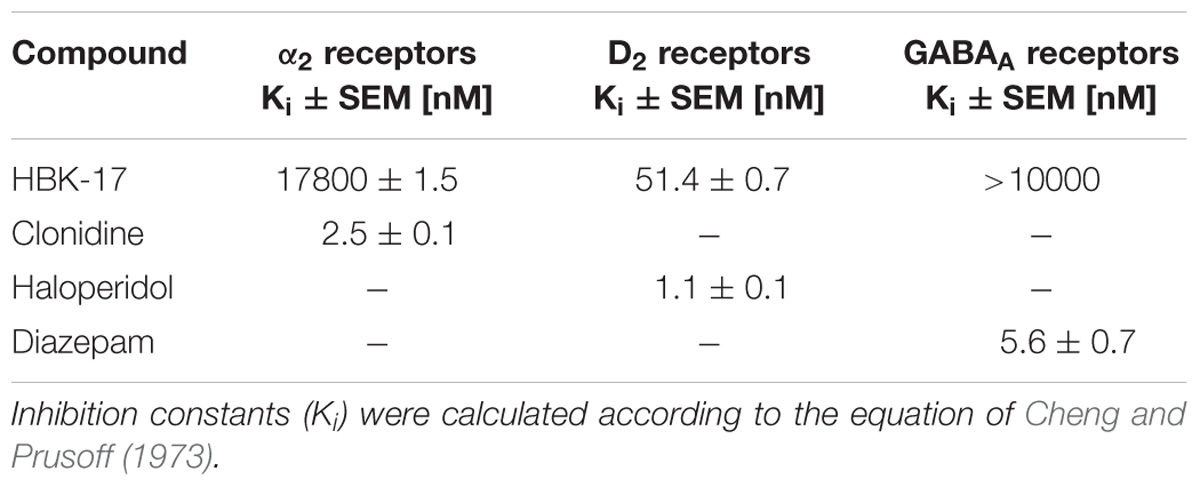
TABLE 1. The affinity of HBK-17 for adrenergic α2, dopaminergic D2, and GABAergic GABAA receptors.
Since HBK-17 showed a moderate affinity for D2, 5-HT2A, and 5-HT7 receptors, we investigated its intrinsic activity toward these receptors. The results of the functional assays showed that HBK-17 had no agonistic properties, but it blocked dopamine D2 receptor. Its value of the equilibrium dissociation constant for a competitive antagonist (Kb) was 7.4-fold lower than the Kb value of the reference antagonist – chlorpromazine (Table 2). Similarly, HBK-17 did not show any agonistic properties at serotonin 5-HT7 receptor but blocked the receptor with Kb value 78-fold lower than the reference antagonist, SB269970 (Table 2). As demonstrated in Table 2 HBK-17 showed no agonistic properties and negligible antagonistic effect at serotonin 5-HT2A receptor. Compared with the reference compound, α-methylserotonin, HBK-17 blocked 5-HT2A receptor 1228.1-fold weaker (Table 2).
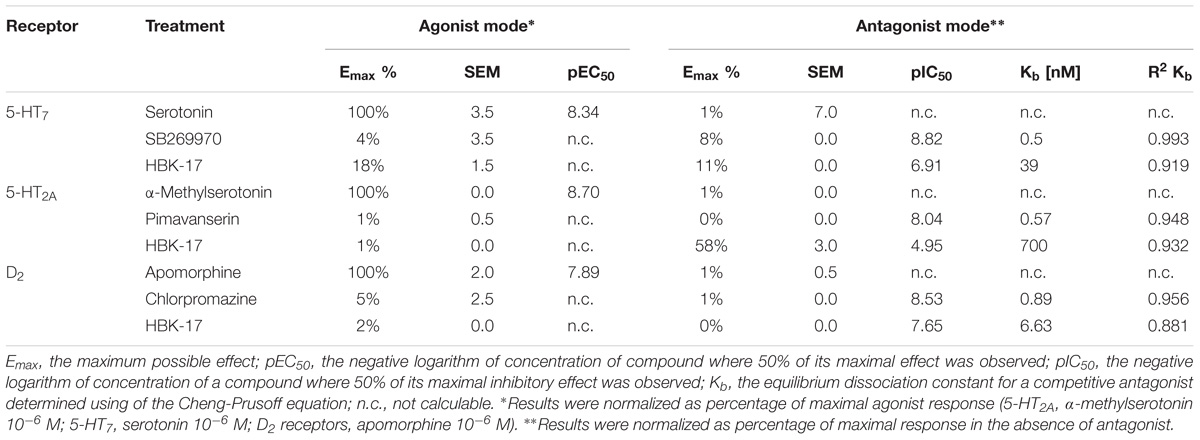
TABLE 2. Intrinsic activity of HBK-17 toward 5-HT2A, 5-HT7, and D2 receptors.
A single receptor can activate numerous signaling pathways and influence cell function oppositely. Biased agonists stabilize the receptor’s conformation preferentially, and consequently activate a selected signaling pathway. In our study we used cell cultures as they provide a valuable complement to in vivo experiments, allowing more controlled manipulation of the cellular functions and processes.
Although 5-HT1A receptor couples to the broad panel of second messengers, the primary coupling linkage is to the inhibition of adenylate cyclase (Chilmonczyk et al., 2015). Therefore, first, we evaluated the influence of HBK-17 on cAMP production using CHO-K1 cells with expression of the human serotonin 5-HT1A receptor. Contrary to the reference compound, serotonin, HBK-17 did not inhibit cAMP formation. However, the studied compound showed antagonistic properties in this assay. HBK-17 demonstrated 6.1-fold lower Kb value compared with the reference compound, WAY-100135 (Table 3).
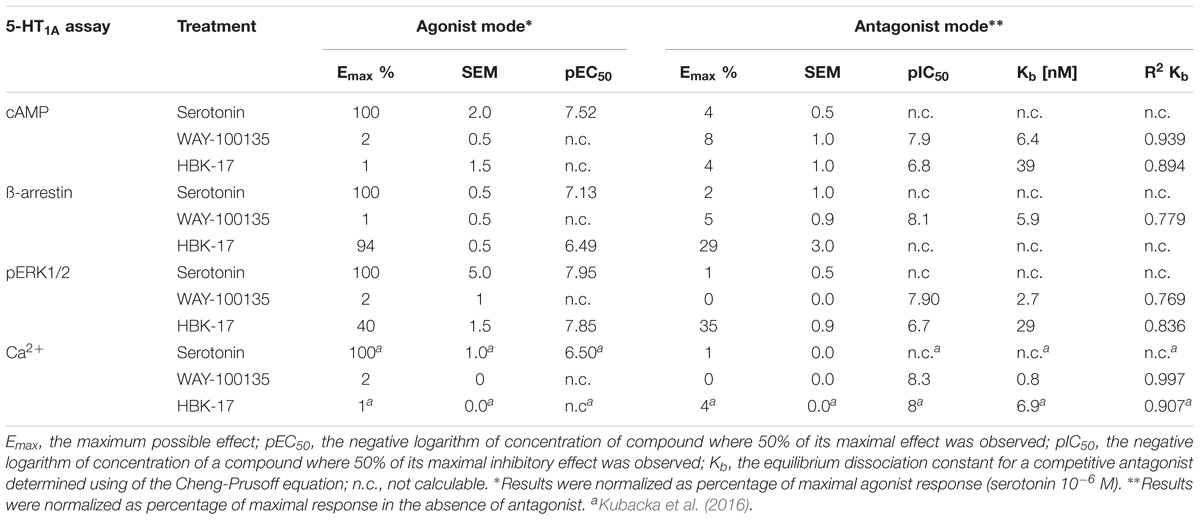
TABLE 3. Intrinsic activity of HBK-17 at 5-HT1A receptor in various functional assays.
To prevent over-stimulation G protein-coupled receptors recruit cytosolic proteins – arrestins – which promote signaling termination. Thus, we determined the influence of HBK-17 on β-arrestin recruitment using HTR1A-bla U2OS receptor cells. The functional studies revealed that HBK-17 relative to serotonin moderately activated ß-arrestin recruitment in 5-HT1A receptor (Table 3). As presented in Table 3 the efficacy of HBK-17 (Emax = 94%) was similar relative to that induced by 10 μmol L−1 serotonin (Emax = 100%), but the potency was 4.4-fold lower. In the antagonist mode, HBK-17 did not show any effect in this assay.
Since stimulation of 5-HT1A receptor may lead to the activation of the ERK/MAPK pathway, we determined the effect of HBK-17 on the level p-ERK1/2 using the CHO-5HT1A receptor cells. The compound showed partial agonistic properties in this assay. In the p-ERK1/2 assay, HBK-17 showed 6.6-fold higher potency than serotonin, but 2.5-fold lower efficacy (Table 3). In the antagonist mode HBK-17, compared with the reference antagonist, WAY-100135, showed 10.7-fold lower Kb value.
Our previous studies demonstrated that in the Ca2+ mobilization assay the reference compound, WAY-100135, showed strong antagonistic properties (Table 3; Kubacka et al., 2016).
In order to evaluate the anxiolytic-like properties of the compound, we performed four plate test in mice. The tested compound HBK-17 at a dose 5 mg/kg (but not 1.25, 2.5, 10, or 20 mg/kg) significantly increased the number of punished crossings by 70.9% compared with non-treated control [F(5,54) = 3.791, p < 0.01] (Figure 2A). A reference 5-HT1A receptor agonist, 8-OD-DPAT, at a dose 0.5 mg/kg (but not 0.25, 1 or 2 mg/kg) significantly increased the number of punished crossings by 38.2% [F(4,37) = 3.791, p < 0.05] (Figure 2C). Another reference compound SB269970, a 5-HT7 receptor antagonist, at a dose 2.5 mg/kg (but not 1.25 or 5 mg/kg) increased the number of punished crossings by 137.5% compared with non-treated control [F(3,29) = 5.903, p < 0.01] (Figure 2D). Compounds with analgesic properties might show false positive results in the four plate test. Therefore, we evaluated the HBK-17 influence on pain responses in the hot plate test in mice. HBK-17 at doses 1.25–20 mg/kg did not increase the latency time to the first hind paw or/and jumping response [F(5,54) = 0.466, ns] (Figure 2B).
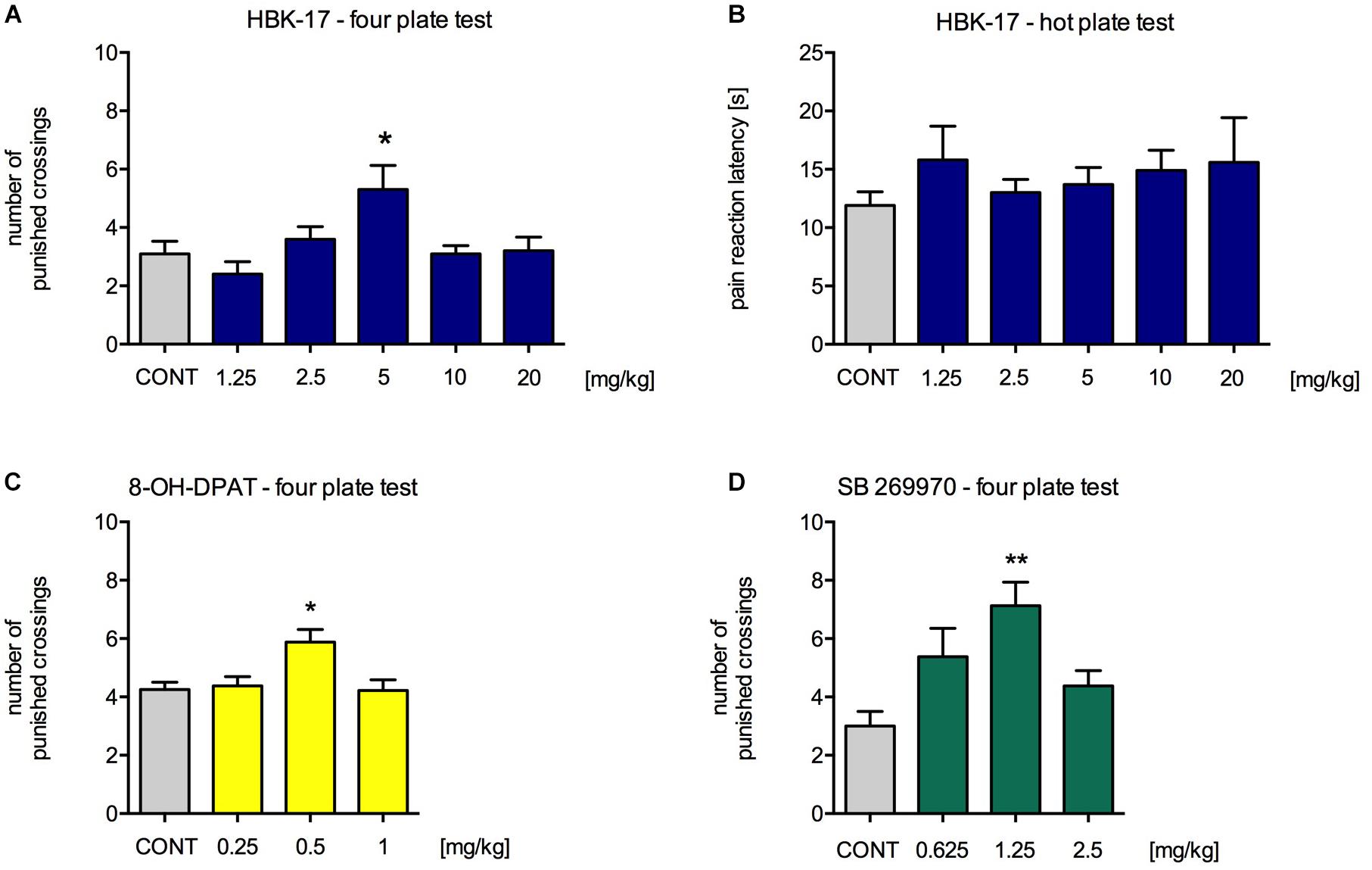
FIGURE 2. The effect of HBK-17 (A), 8-OH-DPAT (C), and SB 269970 (D) on the behavior of mice in the four plate test and the influence of HBK-17 on the pain threshold (B). HBK-17 and SB 269970 were administered intraperitoneally 30 min before the experiment, whereas 8-OH-DPAT was injected 15 min before the four plate test. CONT – control group; Statistical analysis: one-way ANOVA (Newman-Keuls post hoc) ∗p < 0.05, ∗∗p < 0.01 vs. control group; n = 8–10 mice per group.
To confirm our findings, we performed another experiment assessing anxiolytic-like properties of the compound, i.e., elevated plus maze test. HBK-17 at a dose 5 mg/kg (but not 1.25, 2.5, 10, or 20 mg/kg) compared with non-treated control significantly increased the time spent in the open arms by 38.9% [F(5,54) = 4.143, p < 0.01] and the number of open arm entries by 28.6% [F(5,54) = 5.267, p < 0.01] (Figures 3A,B). 8-OH-DPAT at a dose 2 mg/kg (but not 0.25, 0.5 or 1 mg/kg) compared with non-treated control significantly increased the time spent in the open arms by 154.3% [F(3,29) = 3.710, p < 0.05] but had no effect on the number of open arm entries [F(3,29) = 2.224, ns] (Figures 3C,D). SB269970 at a dose 2.5 mg/kg (but not 1.25 or 5 mg/kg) compared with non-treated control significantly increased the time spent in the open arms by 76.4% [F(3,28) = 3.026, p < 0.05] and the number of open arm entries by 48.6% [F(3,30) = 3.396, p < 0.05] (Figures 3E,F). Diazepam at the doses 1.25 and 2.5 mg/kg (but not 0.3125 or 0.625 mg/kg) compared with non-treated control significantly increased the time spent in the open arms by 84.6 and 59.9% [F(4,35) = 12.870, p < 0.0001], respectively. The reference compound significantly increased the number of open arm entries at the doses 0.625, 1.25, and 2.5 mg/kg (but not 0.3125 mg/kg) by 78.5, 73.9, and 124.6% [F(4,35) = 5.908, p < 0.01], respectively (Figures 3G,H).
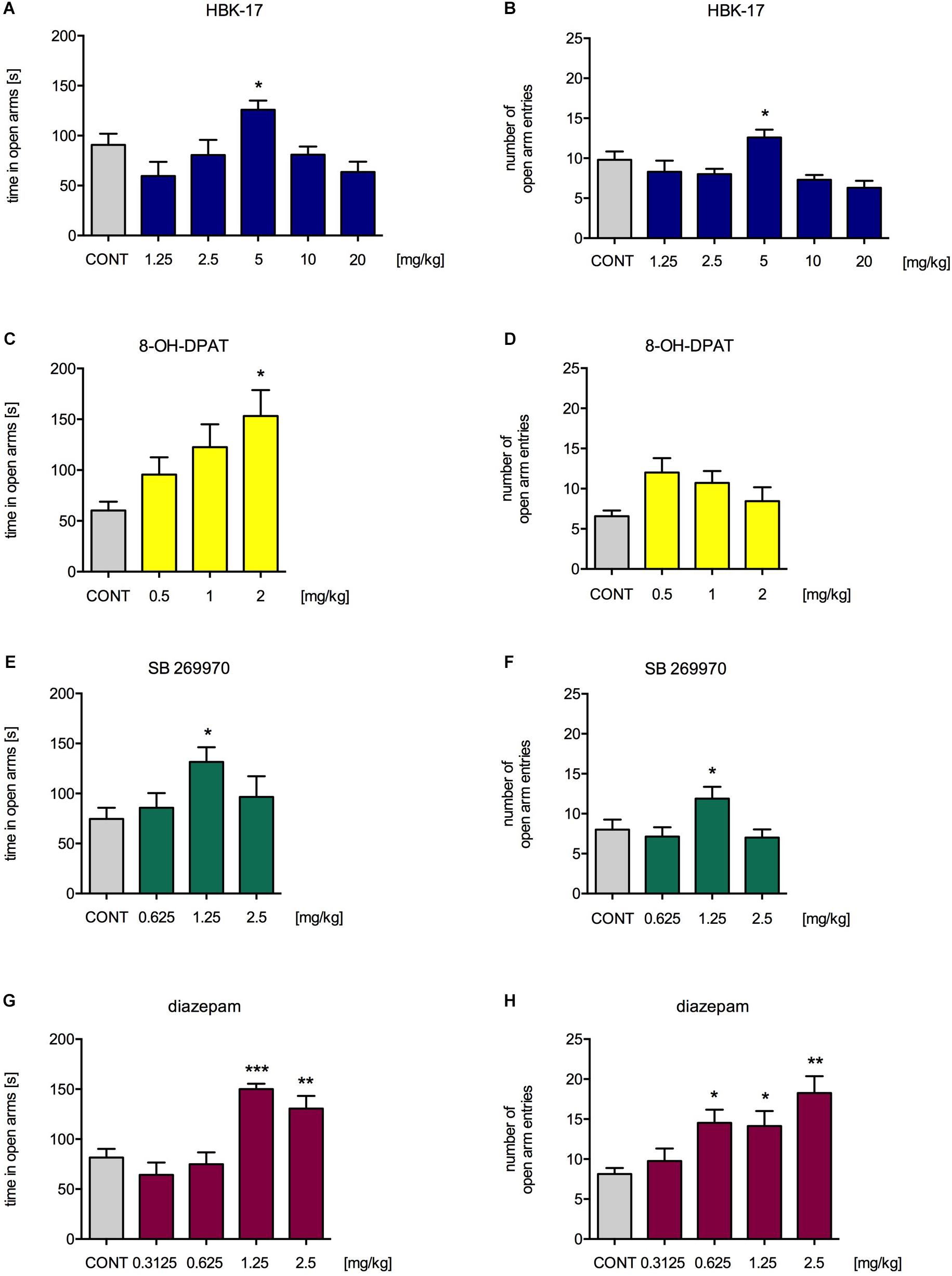
FIGURE 3. The effect of HBK-17 (A,B), 8-OH-DPAT (C,D), SB 269970 (E,F), or diazepam (G,H) on the behavior of mice in the elevated plus maze test. HBK-17, SB 269970, and diazepam were administered intraperitoneally 30 min before the test, whereas 8-OH-DPAT was injected 15 min before the elevated plus maze test. CONT. CONT – control group; Statistical analysis: one-way ANOVA (Newman-Keuls post hoc) ∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001 vs. control group; n = 8–10 mice per group.
To find the possible mechanism of anxiolytic-like activity of HBK-17, we again performed the four plate test, but this time using tryptophan hydroxylase inhibitor (pCPA), tyrosine hydroxylase inhibitor (AMPT), and 5-HT1A receptor antagonist (WAY-100635), which would show the involvement of the serotonergic system, noradrenergic system, and the 5-HT1A receptor, respectively. HBK-17 (5 mg/kg) significantly increased number of punished crossings of mice by 84.2% compared with non-treated control group. Three days treatment with pCPA (200 mg/kg) did not influence the measured parameter but abolished the activity of HBK-17 (Figure 4A). The two-way ANOVA demonstrated no effect of pCPA [F(1,28) = 0.887; ns], significant effect of HBK-17 [F(1,28) = 7.986; p < 0.01], and significant interaction [F(1,28) = 4.831; p < 0.05]. Similarly, the administration of HBK-17 (5 mg/kg) increased the number of punished crossings by 60.0% compared with non-treated control group. Pretreatment with WAY-100635 (0.3 mg/kg) did not influence the number of punished crossings, but it antagonized the effect of HBK-17 (Figure 4B). The two-way ANOVA demonstrated no effect of WAY-100635 [F(1,28) = 3.092; ns], significant effect of HBK-17 [F(1,28) = 4.209; p < 0.05], and significant interaction [F(1,28) = 8.589; p < 0.01]. The injection of HBK-17 (5 mg/kg) caused a significant increase by 76% in the number punished crossings, while AMPT (100 mg/kg) had no effect on this parameter. Pretreatment with AMPT did not influence the effect of HBK-17 (Figure 4C) in this test. The two-way ANOVA showed no effect of AMPT [F(1,28) = 0.209; ns], significant effect of HBK-17 [F(1,28) = 28.490; p < 0.05], and no significant interaction [F(1,28) = 0.209; ns].
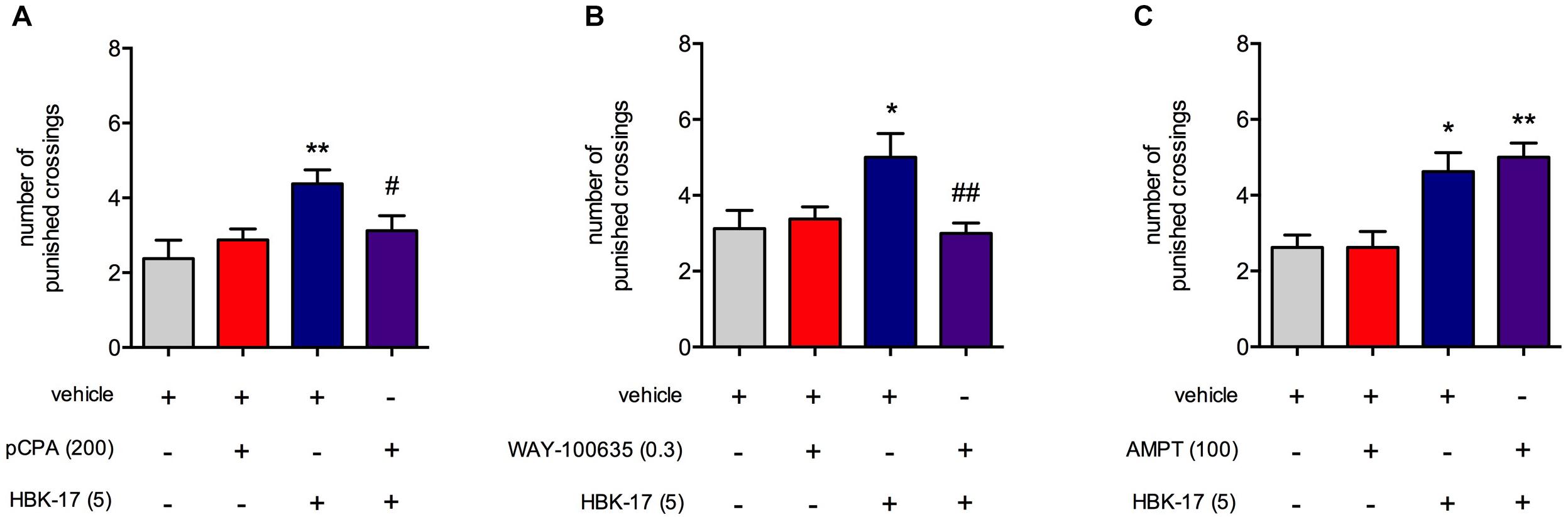
FIGURE 4. The effect of p-chlorophenylalanine [pCPA, (A)], WAY-100635 (B), or α-methylphenylalanine [AMPT, (C)] on anxiolytic-like activity of HBK-17 in the four plate test in mice. pCPA was injected intraperitoneally (i.p.) once daily for three consecutive days. Twenty-four hour after the last injection and 30 min before the test mice received (i.p.) either HBK-17 or 0.9% NaCl. WAY-100635 was administered subcutaneously 15 min before HBK-17. AMPT was injected (i.p.) 4 h before HBK-17. Vehicle-treated groups received 1% Tween (i.p.). Statistical analysis: two-way ANOVA (Bonferroni post hoc); ∗p < 0.05, ∗∗p < 0.01 vs. respective control group receiving vehicle or 1% Tween; #p < 0.05, ##p < 0.01 vs. respective group receiving HBK-17 at the dose 5 mg/kg; n = 8 mice per group.
To investigate whether the observed increased number of punished crossings was not due to psychostimulant properties, we evaluated the effect of HBK-17 on locomotor activity of mice in the 1-min session. The compound did not influence the locomotor activity of mice at the doses 1.25–20 mg/kg during the 1-min session [F(5,42) = 2.616; ns] (Table 4). To exclude false positive results in the elevated plus maze test, we also evaluated the effect of HBK-17 on locomotor activity of mice in 5-min session. The compound did not influence the locomotor activity of mice at the doses 1.25–20 mg/kg during the 5-min session [F(5,42) = 2.485; ns] (Table 4). Neither HBK-17 alone nor in combinations with pCPA or AMPT influenced the locomotor activity of mice (Table 4). The two-way ANOVA demonstrated no effect of pCPA [F(1,28) = 0.405; ns], no effect of HBK-17 [F(1,28) = 0.192; ns], and no significant interaction [F(1,28) = 1.001; ns]. In case of AMPT the two-way ANOVA showed no effect of AMPT [F(1,28) = 0; ns], no effect of HBK-17 [F(1,28) = 0.026; ns], and no significant interaction [F(1,28) = 1.224; ns]. The administration of HBK-17, WAY-100635 or the combination of both, did not affect the locomotor activity of mice (Table 4). The two-way ANOVA demonstrated no effect of WAY-100635 [F(1,28) = 1.452; ns], no effect of HBK-17 [F(1,28) = 0.088; ns], and no significant interaction [F(1,28) = 1.316; ns].
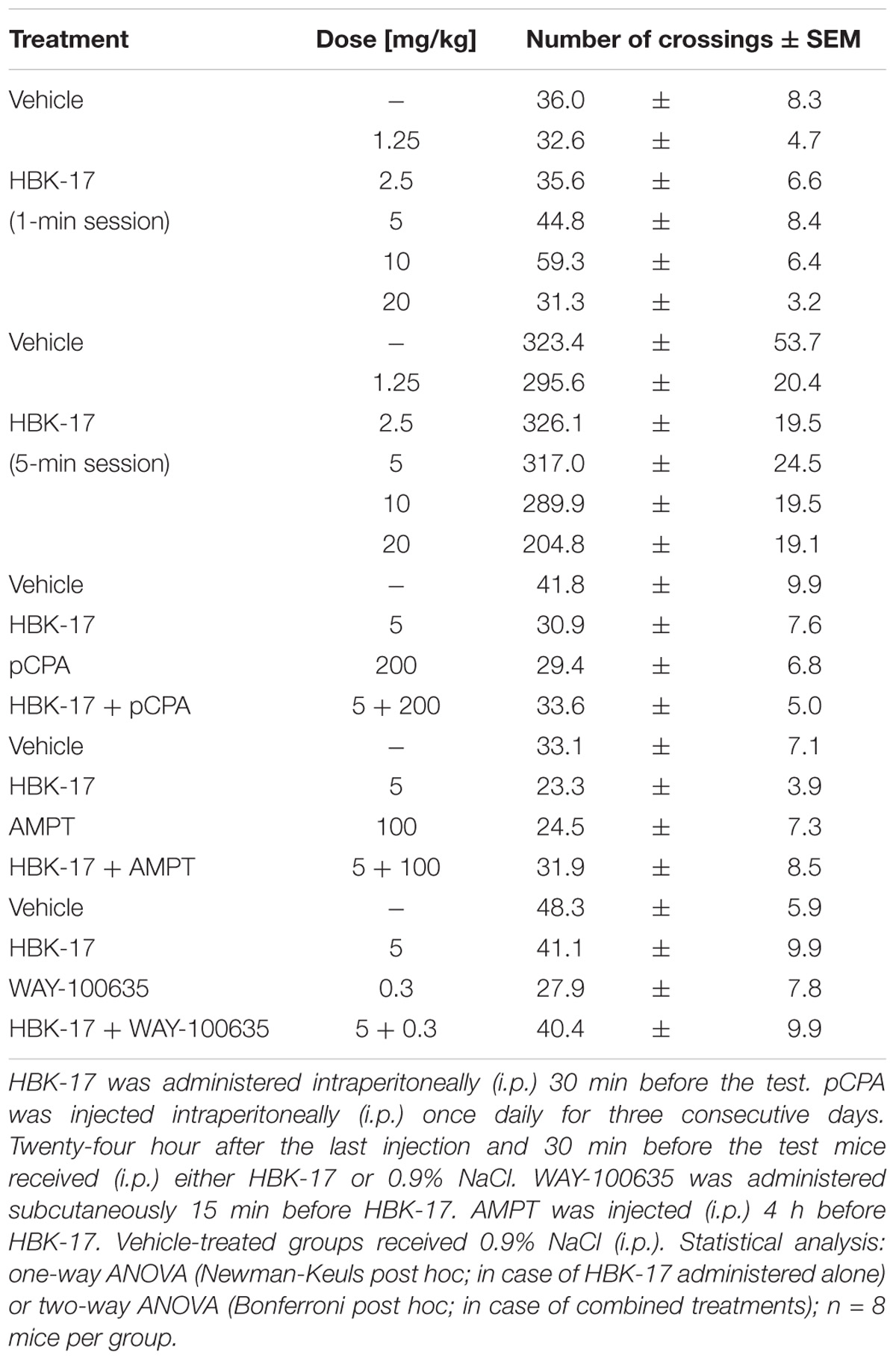
TABLE 4. The influence of HBK-17 and its combinations with pCPA, AMPT, or WAY-100635 on locomotor activity of mice.
We used a well-validated UPLC-MS/MS method to determine pharmacokinetic parameters after i.v. and i.p. administration of HBK-17 at a dose of 5 mg/kg in mice. The parameters for the compound are presented in Table 5. HBK-17 was eliminated rather fast (i.v.: t1/2 = 105 min; i.p.: t1/2 = 150 min). The absorption of the compound after i.p. administration was also fast (tmax = 5 min) with the maximum plasma concentration at 806.7 ng/mL. The volume of distribution was 9.7 L/kg, suggesting the distribution of the compound throughout the total body water. The absolute bioavailability after i.p. administration, which was estimated based on the AUC0→∞ calculated from zero time to infinity, was significant (F = 44.8%).
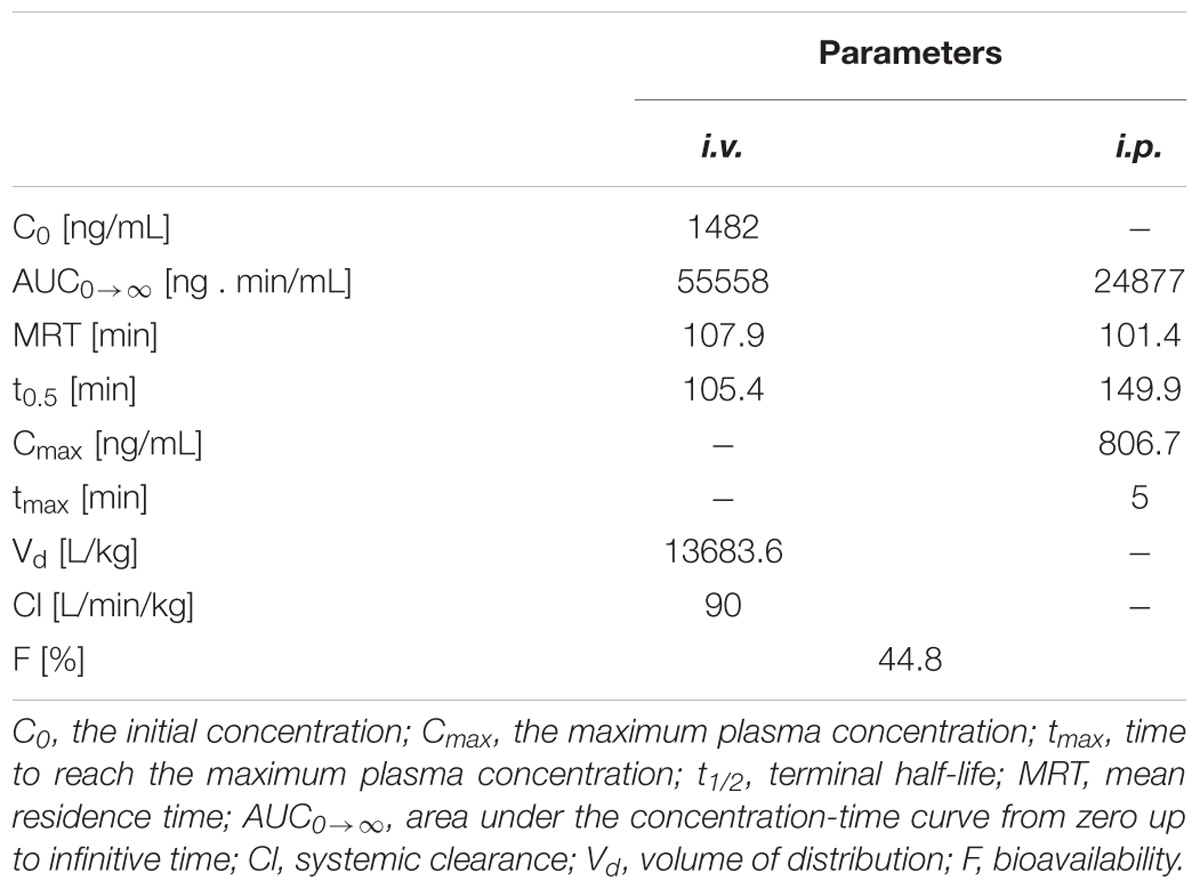
TABLE 5. Pharmacokinetic parameters for HBK-17 after i.v. and i.p administration at a dose of 5 mg/kg to mice.
HBK-17 distributed widely and rapidly in the brain within the time course examined. Thus, we can conclude that the compound penetrated the blood-brain barrier. We detected the highest concentrations of HBK-17 5 min after i.p. administration. The compound’s concentration in the brain decreased significantly within 8 h, which suggests that HBK-17 not tend to accumulate. The AUC ratio between brain and plasma was 1.13 and 0.96 after i.v. or i.p. administration revealed a satisfactory binding affinity to mice brain. The other parameters were as follows: AUC0→∞ = 23860 ng . min/g, MRT = 147.1 min, Cmax = 523.8 ng/g, tmax = 5 min.
We found that HBK-17 possessed anxiolytic-like activity in mice, which depended on the 5-HT1A receptor activation. After binding to the 5-HT1A receptor, the compound preferentially activated β-arrestin recruitment over Ca2+ mobilization, phosphorylation of ERK1/2 or cAMP formation inhibition. We also demonstrated that following a single i.v. or i.p. administration, HBK-17 showed relatively high bioavailability, rapid absorption to the blood but rather low distribution to the brain.
Serotonin 5-HT1A and 5-HT7 receptors play a role in anxiety (Żmudzka et al., 2018). Studies proved that both stimulation and blockade of 5-HT1A and the blockade of 5-HT7 receptors might cause anxiolytic-like effects (Garcia-Garcia et al., 2014; Nikiforuk, 2015). Since HBK-17 showed affinity for both 5-HT1A and 5-HT7 receptors, it might also influence anxiety-like behaviors. Thus, as the first step of our studies, we investigated the compound’s anxiolytic-like properties using a preliminary assay based on the unconditioned fear model of anxiety, i.e., the four plate test in mice. Our results show that the compound possessed significant anxiolytic-like activity at one dose tested (5 mg/kg). Since HBK-17 did not influence pain responses and locomotor activity of mice, we can conclude that the obtained results were specific to anxiety.
Next, we decided to confirm our findings in another experiment, i.e., the elevated plus maze, which uses a natural aversion of rodents for open and elevated areas. Compounds with anxiolytic-like properties increase the time spent and the number of entries to the open arms of the elevated plus maze. Similarly, the administration of HBK-17 at only one dose (5 mg/kg) increased the time spent and the number of entries in the open arms of the elevated plus maze. The compound did not affect the locomotor activity of mice; thus, the observed effect could not be attributed to psychostimulant properties. In our previous experiments, HBK-17 showed strong antiarrhythmic properties in adrenaline-induced arrhythmia in rats (Pytka et al., 2016a). Anxiety disorder patients often report autonomic manifestations of anxiety, such as increased heart rate or blood pressure (Gałuszka, 2013). These factors also determine anxiety levels in animals (Pattij, 2002). The fact that HBK-17 normalized heart rhythm in adrenaline-induced arrhythmia might complement its anxiolytic-like effect.
Interestingly, in both behavioral tests, HBK-17 showed an inverted U-shaped dose-effect (only one dose was active). A similar dose-effect function was demonstrated for a reference compound, a selective 5-HT7 receptor antagonist, SB 269970 and a 5-HT1A receptor agonist, 8-OH-DPAT in the four plate test. In the elevated plus maze test the only active dose of 8-OH-DPAT was the highest dose tested (i.e., 2 mg/kg). Conversely, a well-known anxiolytic, diazepam, showed a dose dependent-effect in both tests (Pytka et al., 2015a). Although scientists very often report such a non-linear relationship in pharmacological studies (Baldi and Bucherelli, 2005), an inverted U-shaped dose-effect it is very poorly understood. Some scientists suggest that this effect might have multifactorial nature, which is difficult to explain. In the case of HBK-17, there might be an explanation of this phenomenon. The anxiolytic-like effect of HBK-17 at higher doses might disappear due to the sedative properties of the compound, which we reported in our previous studies (ED50 = 15.0 mg/kg, observation time: 30 min) (Waszkielewicz et al., 2015). It is also possible that as HBK-17, 8-OH-DPAT, and SB 269970 showed an inverted U-shaped dose-effect, this might be a common feature of serotonin agents.
Toward a better understanding of the pharmacological properties of HBK-17, we evaluated the compound’s affinity for GABAA, adrenergic α2 and dopamine D2 receptors. The studied compound showed a moderate affinity for D2 receptors, whereas no affinity for GABAA and α2 receptors. Our previous studies demonstrated that HBK-17 showed high affinity for α1 (Pytka et al., 2016a) and 5-HT7 (Waszkielewicz et al., 2015), moderate for 5-HT2A (Kubacka et al., 2016), and very low for 5-HT6 and 5-HT3 receptors (Waszkielewicz et al., 2015; Kubacka et al., 2016). Bearing that in mind, we evaluated the compound’s intrinsic activity at 5-HT2A, 5-HT7, and D2 receptors. Our functional studies indicated that HBK-17 was a moderate D2 and very weak 5-HT7 receptors antagonist.
In our previous experiments, HBK-17 showed antagonistic properties at 5-HT1A receptor (Ca2+ mobilization assay), in this study we demonstrated that the compound showed functional selectivity at the 5-HT1A receptor. It preferentially activated ß-arrestin recruitment vs. p-ERK1/2, cAMP production inhibition or Ca2+ mobilization. Interestingly, HBK-17 showed partial agonistic activity in the p-ERK1/2 assay. Studies demonstrated decreased ERK expression and phosphorylation in post-mortem brains of depressed patients (suicide victims) (Dwivedi et al., 2001). Similar changes were observed in animal models of depression (Tiraboschi et al., 2004). Therefore, the fact that HBK-17 increased p-ERK1/2 might at least in part underlie its previously described antidepressant-like properties (Waszkielewicz et al., 2015; Kubacka et al., 2016). At this point, it is also worth mentioning that ß-arrestins desensitize not only G-protein-dependent signal pathways but also promote novel pathways of signal transduction, e.g., ERK, JNK, p38, or Akt (for review see DeWire et al., 2007). Thus, it might be possible that the observed increase in p-ERK1/2 might also be a result of the activation of ß-arrestin recruitment. Nevertheless, we need to perform more experiments to explain this issue.
To find the possible mechanism of anxiolytic-like activity of HBK-17, we performed another set of experiments, i.e., pretreated mice with pCPA (tryptophan hydroxylase inhibitor), WAY-100635 (non-selective 5-HT1A receptor antagonist), or AMPT (tyrosine hydroxylase inhibitor). Imaizumi et al. (1996) demonstrated that a 3-day treatment with pCPA (200 mg/kg) significantly reduced serotonin levels in the cortex (30%), diencephalon (35%), midbrain (24%), and pons with medulla (34%). The depletion of serotonin levels by pCPA abolished the anxiolytic-like activity of HBK-17 in the four plate test. We observed a similar effect after pretreatment with WAY-100635, which implies that the anxiolytic-like activity of HBK-17 depended on the activation of the serotonergic system, and particularly 5-HT1A receptors. In contrast, AMPT did not affect the compound’s activity in this test. This suggests that noradrenergic and dopaminergic systems are not involved in HBK-17 anxiolytic-like activity. We speculate that the anxiolytic-like activity of HBK-17 might have been a result of the postsynaptic 5-HT1A receptors internalization induced by the recruitment of ß-arrestins and possibly the stimulation of presynaptic receptors. Nonetheless, our hypothesis requires confirmation.
Pharmacokinetic parameters of the compound influence its pharmacological activity. Thus, we evaluated the pharmacokinetic profile of HBK-17. We demonstrated that HBK-17 was rapidly absorbed with the peak concentration occurring after 5 min and had a rather long half-life of about 105 and 150 min when administered i.v. or i.p., respectively. The volume of distribution (9.7 L/kg) suggests that the compound was moving throughout the body water. We should emphasize the relatively high bioavailability (44.8%) of the studied compound. HBK-17 could penetrate the brain tissue with the brain/plasma ratio of 1.13 and 0.96 after i.v. or i.p. administration, respectively. The results show that we could administer the compound by both i.v. and i.p. routes. Therefore, we conclude that HBK-17 has a desirable pharmacokinetic profile for pharmacological studies. Moreover, to our knowledge, this is the first report to quantify HBK-17 in biomatrices and assess its pharmacokinetics in mice.
We demonstrated that HBK-17 possessed anxiolytic-like activity in mice, which depended on the 5-HT1A receptor activation. The compound after binding to the 5-HT1A receptor preferentially activated β-arrestin recruitment over Ca2+ mobilization, phosphorylation of ERK1/2 or cAMP formation inhibition. We also demonstrated that following a single i.v. or i.p. administration, HBK-17 showed relatively high bioavailability, rapid absorption to the blood but rather low distribution to brain. The interesting pharmacological profile of HBK-17 encourages further experiments to understand its mechanism of action fully.
KPy, JS, BF, and MK conceived and designed the experiments. KPy, MG-L, KN, AS, MS, KPa, EŻ, and KS performed the experiments. KPy, MG-L, MW, AS, AO, AG, and JŚ analyzed the data. MK, KPy, AW, and HM contributed to reagents, materials, and analysis tools. KPy, MG-L, KS, EŻ, MW, AS, and AW wrote the paper.
This study was supported by Jagiellonian University grant numbers: K/DSC/001955, K/DSC/004285, and K/ZDS/006223, and partially by Students Scientific Association grant number: GS 15/14 UJ, The National Science Centre (NCN) grant numbers: DEC- 2013/11/B/ST7/01713, 2017/01/X/NZ7/00818, and 2015/19/B/NZ7/03543, and Silesian University of Technology BK grant number: 02/010/BK18/0102.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We wish to thank Agnieszka Niedbał and Teresa Dobrut for their technical assistance.
Akimova, E., Lanzenberger, R., and Kasper, S. (2009). The serotonin-1A receptor in anxiety disorders. Biol. Psychiatry 66, 627–635. doi: 10.1016/j.biopsych.2009.03.012
Aron, C., Simon, P., Larousse, C., and Boissier, J. R. (1971). Evaluation of a rapid technique for detecting minor tranquilizers. Neuropharmacology 10, 459–469. doi: 10.1016/0028-3908(71)90074-8
Baldi, E., and Bucherelli, C. (2005). The inverted “u-shaped” dose-effect relationships in learning and memory: modulation of arousal and consolidation. Nonlinearity Biol. Toxicol. Med. 3, 9–21. doi: 10.2201/nonlin.003.01.002
Bourin, M., Masse, F., Dailly, E., and Hascoët, M. (2005). Anxiolytic-like effect of milnacipran in the four-plate test in mice: mechanism of action. Pharmacol. Biochem. Behav. 81, 645–656. doi: 10.1016/j.pbb.2005.04.015
Cervo, L., Mocaër, E., Bertaglia, A., and Samanin, R. (2000). Roles of 5-HT1A receptors in the dorsal raphe and dorsal hippocampus in anxiety assessed by the behavioral effects of 8-OH-DPAT and S 15535 in a modified Geller–Seifter conflict model. Neuropharmacology 39, 1037–1043. doi: 10.1016/s0028-3908(99)00189-6
Cheng, Y., and Prusoff, W. H. (1973). Relationship between the inhibition constant (K1) and the concentration of inhibitor which causes 50 per cent inhibition (I50) of an enzymatic reaction. Biochem. Pharmacol. 22, 3099–3108. doi: 10.1016/0006-2952(73)90196-2
Chilmonczyk, Z., Bojarski, A. J., Pilc, A., and Sylte, I. (2015). Functional selectivity and antidepressant activity of serotonin 1A receptor ligands. Int. J. Mol. Sci. 16, 18474–18506. doi: 10.3390/ijms160818474
Collinson, N., and Dawson, G. R. (1997). On the elevated plus-maze the anxiolytic-like effects of the 5-HT 1A agonist, 8-OH-DPAT, but not the anxiogenic-like effects of the 5-HT 1A partial agonist, buspirone, are blocked by the 5-HT 1A antagonist, WAY 100635. Psychopharmacology 132, 35–43. doi: 10.1007/s002130050317
De Almeida, R. M., Giovenardi, M., Charchat, H., and Lucion, A. B. (1998). 8-OH-DPAT in the median raphe nucleus decreases while in the medial septal area it may increase anxiety in female rats. Neurosci. Biobehav. Rev. 23, 259–264. doi: 10.1016/S0149-7634(98)00026-8
DeWire, S. M., Ahn, S., Lefkowitz, R. J., and Shenoy, S. K. (2007). Beta-arrestins and cell signaling. Annu. Rev. Physiol. 69, 483–510. doi: 10.1146/annurev.ph.69.013107.100021
Dwivedi, Y., Rizavi, H. S., Roberts, R. C., Conley, R. C., Tamminga, C. A., and Pandey, G. N. (2001). Reduced activation and expression of ERK1/2 MAP kinase in the post-mortem brain of depressed suicide subjects. J. Neurochem. 77, 916–928. doi: 10.1046/j.1471-4159.2001.00300.x
File, S. E., Gonzalez, L. E., and Andrews, N. (1996). Comparative study of pre- and postsynaptic 5-HT1A receptor modulation of anxiety in two ethological animal tests. J. Neurosci. 16, 4810–4815. doi: 10.1523/JNEUROSCI.16-15-04810.1996
Gałuszka, A. (2013). Personality and social resources in case of serious somatic disease. Psychol. Health 28, 217–217.
Garcia-Garcia, A. L., Newman-Tancredi, A., and Leonardo, E. D. (2014). 5-HT(1A) [corrected] receptors in mood and anxiety: recent insights into autoreceptor versus heteroreceptor function. Psychopharmacology 231, 623–636. doi: 10.1007/s00213-013-3389-x
Heisler, L. K., Chu, H. M., Brennan, T. J., Danao, J. A., Bajwa, P., Parsons, L. H., et al. (1998). Elevated anxiety and antidepressant-like responses in serotonin 5-HT1A receptor mutant mice. Proc. Natl. Acad. Sci. U.S.A. 95, 15049–15054. doi: 10.1073/pnas.95.25.15049
Iderberg, H., McCreary, A. C., Varney, M. A., Cenci, M. A., and Newman-Tancredi, A. (2015). Activity of serotonin 5-HT1A receptor “biased agonists” in rat models of Parkinson’s disease and l-DOPA-induced dyskinesia. Neuropharmacology 93, 52–67. doi: 10.1016/j.neuropharm.2015.01.012
Imaizumi, M., Miyazaki, S., and Onodera, K. (1996). Effects of theophylline in p-chlorophenylalanine-treated mice in a light/dark test. Methods Find. Exp. Clin. Pharmacol. 18, 513–520.
Kenakin, T. (2011). Functional selectivity and biased receptor signaling. J. Pharmacol. Exp. Ther. 336, 296–302. doi: 10.1124/jpet.110.173948
Kubacka, M., Mogilski, S., Bednarski, M., Nowiński, L., Dudek, M., Żmudzka, E., et al. (2016). Antidepressant-like activity of aroxyalkyl derivatives of 2-methoxyphenylpiperazine and evidence for the involvement of serotonin receptor subtypes in their mechanism of action. Pharmacol. Biochem. Behav. 141, 28–41. doi: 10.1016/j.pbb.2015.11.013
Lister, R. G. (1987). The use of a plus-maze to measure anxiety in the mouse. Psychopharmacology 92, 180–185. doi: 10.1007/BF00177912
Mailman, R. B., and Murthy, V. (2010). Third generation antipsychotic drugs: partial agonism or receptor functional selectivity? Curr. Pharm. Des. 16,488–501.
Newman-Tancredi, A. (2011). Biased agonism at serotonin 5-HT1A receptors: preferential postsynaptic activity for improved therapy of CNS disorders. Neuropsychiatry 1, 149–164. doi: 10.2217/npy.11.12
Newman-Tancredi, A., Martel, J. -C., Assié, M. B., Buritova, J., Lauressergues, E., Cosi, C., et al. (2009). Signal transduction and functional selectivity of F15599, a preferential post-synaptic 5-HT1A receptor agonist. Br. J. Pharmacol. 156, 338–353. doi: 10.1111/j.1476-5381.2008.00001.x
Nikiforuk, A. (2015). Targeting the serotonin 5-HT7 receptor in the search for treatments for CNS disorders: rationale and progress to date. CNS Drugs 29, 265–275. doi: 10.1007/s40263-015-0236-0
Parks, C. L., Robinson, P. S., Sibille, E., Shenk, T., and Toth, M. (1998). Increased anxiety of mice lacking the serotonin1A receptor. Proc. Natl. Acad. Sci. U.S.A. 95, 10734–10739. doi: 10.1073/pnas.95.18.10734
Patel, C. B., Noor, N., and Rockman, H. A. (2010). Functional selectivity in adrenergic and angiotensin signaling systems. Mol. Pharmacol. 78, 983–992. doi: 10.1124/mol.110.067066
Pattij, T. (2002). Autonomic changes associated with enhanced anxiety in 5-HT1A receptor knockout mice. Neuropsychopharmacology 27, 380–390. doi: 10.1016/S0893-133X(02)00317-2
Pytka, K., Głuch-Lutwin, M., Kotańska, M., Waszkielewicz, A., Kij, A., and Walczak, M. (2017). Single administration of HBK-15-a triple 5-HT1A, 5-HT7, and 5-HT3 receptor antagonist-reverses depressive-like behaviors in mouse model of depression induced by corticosterone. Mol. Neurobiol. 68:263. doi: 10.1007/s12035-017-0605-4
Pytka, K., Lustyk, K., Żmudzka, E., Kotańska, M., Siwek, A., Zygmunt, M., et al. (2016a). Chemically homogenous compounds with antagonistic properties at all α1-adrenoceptor Subtypes but not β1-adrenoceptor attenuate adrenaline-induced arrhythmia in rats. Front. Pharmacol. 7:229. doi: 10.3389/fphar.2016.00229
Pytka, K., Żmudzka, E., Lustyk, K., Rapacz, A., Olczyk, A., Gałuszka, A., et al. (2016b). The antidepressant- and anxiolytic-like activities of new xanthone derivative with piperazine moiety in behavioral tests in mice. Indian J. Pharmacol. 48, 286–291. doi: 10.4103/0253-7613.182872
Pytka, K., Partyka, A., Jastrzêbska-Wiêsek, M., Siwek, A., Głuch-Lutwin, M., Mordyl, B., et al. (2015a). Antidepressant- and anxiolytic-like effects of new dual 5-HT1A and 5-HT7 antagonists in animal models. PLoS One 10:e0142499. doi: 10.1371/journal.pone.0142499
Pytka, K., Rapacz, A., Zygmunt, M., Olczyk, A., Waszkielewicz, A. M., Sapa, J., et al. (2015b). Antidepressant-like activity of a new piperazine derivative of xanthone in the forced swim test in mice: the involvement of serotonergic system. Pharmacol. Rep. 67, 160–165. doi: 10.1016/j.pharep.2014.08.016
Ramboz, S., Oosting, R., Amara, D. A., Kung, H. F., Blier, P., Mendelsohn, M., et al. (1998). Serotonin receptor 1A knockout: an animal model of anxiety-related disorder. Proc. Natl. Acad. Sci. U.S.A. 95, 14476–14481. doi: 10.1073/pnas.95.24.14476
Sałat, K., Podkowa, A., Kowalczyk, P., Kulig, K., Dziubina, A., Filipek, B., et al. (2015). Anticonvulsant active inhibitor of GABA transporter subtype 1, tiagabine, with activity in mouse models of anxiety, pain and depression. Pharmacol. Rep. 67, 465–472. doi: 10.1016/j.pharep.2014.11.003
Sibille, E., Pavlides, C., Benke, D., and Toth, M. (2000). Genetic inactivation of the serotonin(1A) receptor in mice results in downregulation of major GABA(A) receptor alpha subunits, reduction of GABA(A) receptor binding, and benzodiazepine-resistant anxiety. J. Neurosci. 20, 2758–2765. doi: 10.1523/JNEUROSCI.20-08-02758.2000
Stroth, N., Niso, M., Colabufo, N. A., Perrone, R., Svenningsson, P., Lacivita, E., et al. (2015). Arylpiperazine agonists of the serotonin 5-HT1A receptor preferentially activate cAMP signaling versus recruitment of Î2-arrestin-2. Bioorg. Med. Chem. 23, 4824–4830. doi: 10.1016/j.bmc.2015.05.042
Tiraboschi, E., Tardito, D., Kasahara, J., Moraschi, S., Pruneri, P., Gennarelli, M., et al. (2004). Selective phosphorylation of nuclear CREB by fluoxetine is linked to activation of CaM kinase IV and MAP kinase cascades. Neuropsychopharmacology 29, 1831–1840. doi: 10.1038/sj.npp.1300488
Waszkielewicz, A. M., Pytka, K., Rapacz, A., Wełna, E., Jarzyna, M., Satała, G., et al. (2015). Synthesis and evaluation of antidepressant-like activity of some 4-substituted 1-(2-methoxyphenyl)piperazine derivatives. Chem. Biol. Drug Des. 85, 326–335. doi: 10.1111/cbdd.12394
Wesołowska, A., Nikiforuk, A., and Stachowicz, K. (2006a). Potential anxiolytic and antidepressant effects of the selective 5-HT7 receptor antagonist SB 269970 after intrahippocampal administration to rats. Eur. J. Pharmacol. 553, 185–190. doi: 10.1016/j.ejphar.2006.09.064
Wesołowska, A., Nikiforuk, A., Stachowicz, K., and Tatarczyńska, E. (2006b). Effect of the selective 5-HT7 receptor antagonist SB 269970 in animal models of anxiety and depression. Neuropharmacology 51, 578–586. doi: 10.1016/j.neuropharm.2006.04.017
Yau, J. L., Noble, J., and Seckl, J. R. (2001). Acute restraint stress increases 5-HT7 receptor mRNA expression in the rat hippocampus. Neurosci. Lett. 309, 141–144. doi: 10.1016/S0304-3940(01)02054-7
Keywords: 5-HT1A receptor, anxiolytic-like, mouse models, pharmacokinetics, ß-arrestin signaling
Citation: Pytka K, Głuch-Lutwin M, Żmudzka E, Sałaciak K, Siwek A, Niemczyk K, Walczak M, Smolik M, Olczyk A, Gałuszka A, Śmieja J, Filipek B, Sapa J, Kołaczkowski M, Pańczyk K, Waszkielewicz A and Marona H (2018) HBK-17, a 5-HT1A Receptor Ligand With Anxiolytic-Like Activity, Preferentially Activates ß-Arrestin Signaling. Front. Pharmacol. 9:1146. doi: 10.3389/fphar.2018.01146
Received: 18 May 2018; Accepted: 21 September 2018;
Published: 16 October 2018.
Edited by:
Herve Boutin, University of Manchester, United KingdomReviewed by:
Gian Marco Leggio, Università degli Studi di Catania, ItalyCopyright © 2018 Pytka, Głuch-Lutwin, Żmudzka, Sałaciak, Siwek, Niemczyk, Walczak, Smolik, Olczyk, Gałuszka, Śmieja, Filipek, Sapa, Kołaczkowski, Pańczyk, Waszkielewicz and Marona. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Karolina Pytka, a2Fyb2xpbmEucHl0a2FAdWouZWR1LnBs
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.