 Ping Cheng
Ping Cheng Tong WangWei Li
Tong WangWei Li Ishfaq MuhammadHe WangXiaoqi SunYuqi YangJiarui LiTianshi Xiao
Ishfaq MuhammadHe WangXiaoqi SunYuqi YangJiarui LiTianshi Xiao Xiuying Zhang*
Xiuying Zhang*- Department of Basic Veterinary Science, College of Veterinary Medicine, Northeast Agricultural University, Harbin, China
As a kind of potent stimulus, lipopolysaccharide (LPS) has the ability to cause cell damage by activating toll-like receptor(TLR)4, then nuclear factor kappa B (NF-κB) translocates into the nucleus and changes the expression of related inflammatory genes. Baicalin is extracted from Radix Scutellariae, which possesses anti-inflammation, antioxidant and antibacterial properties. However, the effects of it on LPS-induced liver inflammation have not been fully elucidated. This study aims to investigate the anti-inflammatory effects of Baicalin on the LPS-induced liver inflammation and its underlying molecular mechanisms in chicken. The results of histopathological changes, serum biochemical analysis, NO levels and myeloperoxidase activity showed that Baicalin pretreatment ameliorated LPS-induced liver inflammation. ELISA and qPCR assays showed that Baicalin dose-dependently suppressed the production of IL-1β, IL-6, and TNF-α. Furthermore, the mRNA expression of inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) were significantly decreased by Baicalin. TLR4 is an important sensor in LPS infection. Molecular studies showed that the expression of TLR4 was inhibited by Baicalin pretreatment. In addition, Baicalin pretreatment inhibited NF-kB signaling pathway activation. All results demonstrated the protective effects of Baicalin pretreatment against LPS-induced liver inflammation in chicken via negative regulation of inflammatory mediators through the down-regulation of TLR4 expression and the inhibition of NF-kB activation.
Introduction
Due to significant morbidity and mortality, people start to pay more attention on liver diseases which have become a serious public health problem worldwide. Many studies showed that an immune-mediated mechanism was regarded as a vital role in the development of liver disease, even in determining its prognosis (Park et al., 2010; Lin et al., 2015). Hepatic inflammation is the characteristic of acute liver disease, which can significantly affect the development of liver pathology and even result in liver failure or cancer (Malhi and Kaufman, 2011). Inflammation is a complicated biological interaction that arises in response to a series of injuries caused by physical, noxious chemical stimuli or microbiological toxins (Kim et al., 2001). LPS is a structural part of endotoxin in the outer membrane of Gram-negative bacteria, which is one of the most potent stimuli to organism (Shin et al., 2004). It has been shown that LPS plays an important role in inducing the inflammatory response and leading to various inflammatory diseases (Brigham and Meyrick, 1986; Kim T. H. et al., 2005). In order to create a ideal model for studying systemic inflammation, LPS administration has been extensively used in chicken to induce inflammatory response (Munyaka et al., 2012). Activation of monocytes, macrophages and various cells is the result of LPS exposure, which promotes the secretion of various pro-inflammatory mediators, containing TNF-α, IL-1β, IL-6, COX-2, and iNOS (Kim et al., 2016). These cytokines will promote TLR4-mediated pathways activation, which involves NF-κB (Zhai et al., 2016).
Toll-like receptors (TLRs) play critical roles in the liver health, which can be found on multiple hepatocytes, containing Kupffer cells, hepatic stellate cells, sinusoidal endothelial cells and biliary epithelial cells (Kim et al., 2010; Xu et al., 2011). It has been reported that almost all liver diseases are associated with TLR medicated signals (Takeuchi and Akira, 2010; Guven-Maiorov et al., 2015), and many therapeutic agents can alleviate liver injury through interfering with the TLR4 signaling pathway (Lian et al., 2010; Qian et al., 2014). As one of the best characterized TLRs, TLR4 has been shown to be the receptor for LPS together with CD14 and MD2 (da Silva Correia et al., 2001; Nagai et al., 2002). LPS signals mainly via TLR4 receptor. Activation of TLR4 by LPS induces the activation of NF-kB and MAPK pathways to regulate the release of pro-inflammatory cytokines (Kagan and Medzhitov, 2006).
NF-κB is regarded as one of the most important regulators of inflammatory process, which is a widely expressed nuclear transcription factor (Pahl, 1999). When the phosphorylation and degradation of IκB-α occur, the downstream NF-κB signaling pathway can be activated through p65 translocation into the nucleus to change the expression of related inflammatory genes induced by LPS (Kwon et al., 2014; Fan et al., 2015). In addition, activation of NF-κB can aggravate the early immune response and inflammatory reaction through promoting overproduction of pro-inflammatory mediators. Therefore, to alleviate inflammatory status, restraining TLR4-mediated NF-κB pathway activation may have potential application (Bocchini et al., 1992).
Nowadays, antibiotics have become the first choice of the treatment for liver inflammation. However, many reports have shown that the prevalence of multi-antibiotic Escherichia coli increased markedly in animals (Zhao et al., 2014). Therefore, we need to put out new prophylactic and therapeutic approaches to treat liver inflammation. In recent years, isolating and developing naturally occurring monomers from medicinal plants have become resurgence of interest (Zheng et al., 2010). Furthermore, numerous Chinese herbal medicines have been widely used in clinical case, which are useful to treat inflammatory diseases, including mastitis, acute lung injury and endometritis (Lv et al., 2015).
Baicalin, 7-glucuronic acid, 5,6-dihydroxy-flavone, a kind of flavonoid compound extracted from Radix Scutellariae, which has potent anti-inflammatory and anti-oxidative activity (Hwang et al., 2005; Lee et al., 2015). It has been applied to the treatment of various inflammatory diseases, including periodontitis, hepatitis and ulcerative colitis (Cheng et al., 2006; Cai et al., 2008; Yu et al., 2014). At present, the excellent pharmacological anti-inflammatory attributions of Baicalin have been proposed by a large number of studies (Cui et al., 2014). Either as a single compound such as its tablets and capsules or as a main active component in more than 40 kinds of preparations, Baicalin has been widely used in clinic recorded in the Chinese National Pharmacopoeia Committee (2015).
In this study, we attempted to investigate the anti-inflammatory effects and liver protection of Baicalin against the LPS-induced liver inflammation in chicken and its underlying molecular mechanisms.
Materials and Methods
Reagents
Baicalin (pure, 98%) was purchased from Shanghai Xin Yu Biological Technology Co., Ltd (Shanghai, China). LPS (E. coli, serotype 055: B5) was purchased from Sigma (St. Louis, MO, USA). ELISA kits of IL-1β, TNF-α, and IL-6 were purchased from BioLegend (Camino Santa Fe, CA, USA), MPO and NO assay kits were obtained from the Nanjing Jiancheng Bioengineering Institute (Nanjing, China). Rabbit anti-phospho-IkBα polyclonal antibody, rabbit anti-phospho-p65 polyclonal antibody and rabbit anti- p65 polyclonal antibody were purchased from Sangon Biotech Co., Ltd (Shanghai, China). Rabbit anti-TLR4 polyclonal antibody was obtained from Santa Cruz Biotechnology (Dallas, TX, USA). Rabbit anti- β-actin polyclonal antibody was obtained from univ Biotechnology (Shanghai, China). Rabbit anti-lamin-B polyclonal antibody was obtained from Santa Cruz Biotechnology. And HRP-conjugated goat anti-rabbit IgG was obtained from ZSGE-BIO (Beijing, China). SYBR Green PCR Master Mix was purchased from Toyobo Life Science Co., Ltd (Shanghai, China). And M-MULV cDNA reverse transcription kit was purchased from Sangon Biotech Co., Ltd (Shanghai, China).
Animals and Treatment
Seventy-five 1-day-old Beijing white chickens were obtained from a local hatchery and housed in cage. Temperature and relative humidity accorded with the requirements of young chicken. The chickens were kept on 12 h light and 12 h dark, fed under standard management conditions. All experimental protocols were approved by the Northeast Agricultural University Animal Care and Use Committee prior to initiation of the study.
At 15 days of age, 75 chickens were randomly divided into five groups, including control group, LPS group, and Baicalin (50, 100, and 200 mg/kg body weight BW) + LPS groups. Baicalin was dissolved in pure dimethyl sulfoxide (DMSO), and diluted in saline to result in final concentrations of 50, 100, and 200 mg/kg BW. The final concentration of DMSO was not more than 0.2%. In Baicalin + LPS groups, Baicalin (50, 100, and 200 mg/kg BW) was administered orally using oral gavages once daily during 7 days and 3 h before LPS injection. At 22 days of age, the chickens of the LPS and Baicalin groups were injected LPS with 2 mg/kg BW in left pectoral muscle, while the chickens of the control group received an equal volume of saline.
We observed the general behavioral and clinical symptoms of these chickens following LPS treatment. In addition, we measured the cloacal temperature of each chicken by thermocouple at 24 h after injection of LPS. Then the blood samples of every chicken were collected through heart puncture. Finally, all chickens were humanely euthanized by cervical dislocation and the liver was collected from the chickens. All the specimen samples for further analysis were either stored at −80°C or prepared for tissue sectioning.
Serum Aspartate Aminotransferase and Alanine Aminotransferase Analysis
Blood extracted from the heart was incubated at 37°C for 30 min, then centrifuged to obtain the serum. Biochemical parameters including serum alanine aminotransferase (ALT), and aspartate aminotransferase (AST) were determined using a Automatic biochemical analyzer (type 7080, Japan Hitachi Co. Ltd.).
Histological Analysis
Liver histopathology was evaluated via hematoxylin and eosin (HE) staining (Wang J. et al., 2016). Liver tissues (1 × 1 cm) were excised and fixed with 10% neutral buffered formalin for 24 h, after processed in a series of graded ethanol and dimethyl benzene, the tissues were embedded in paraffin, sliced (4 μm), and then stained with hematoxylin and eosin (H&E). Finally, we observed pathological changes in the liver tissues by using a light microscope.
MPO Activity Analysis
MPO is regarded as an early marker predicting the risk of inflammatory diseases. The livers from each group were collected and weighted (100 mg), then they were homogenized with 500 μl PBS. After centrifugation, the MPO in the supernatant was detected using an assay kit following the manufacturer's protocol and measured with a spectrophotometry at 460 nm. Results were expressed as units MPO per gram of wet tissue.
NO Levels Detection
The liver concentration of nitric oxide (NO) was determined using an NO assay kit, according to the manufacturer's instructions. As a stable product of nitric oxide, nitrite was used to assess NO level. Griess reaction was used to determine the nitrite concentration in the livers. Each homogenates supernatant (100 μl) was mixed with Griess reagent (50 μl) for 10 min at room temperature. Then the absorbance values were measured at 530 nm. The level of NO production can be determined with reference to standard curve of sodium nitrite.
Cytokines Determination by ELISA Assay
The liver samples from the −80°C freezer were weighed (1g), rinsed in pre-cooled saline to remove excess blood before homogenization, then the tissues were homogenized in a tissue homogenizer (Shanghai, China.) in 1 ml of PBS. The homogenate was centrifuged at 3,000 rpm for 10min at 4°C and the supernatant were isolated and diluted. Then, the liver protein levels of cytokines (IL-1β, TNF-α, and IL-6) were assayed using chicken ELISA kits, according to the manufacturer's instructions. All the samples and standards along with a blank control were run in duplicate and the readings were taken at 450 nm using an iMARK™ microplate reader (Bio-Rad Co., Ltd. Shanghai, China) and the protein concentration was quantified.
RNA Extraction and cDNA Synthesis
The total RNA was isolated from chicken liver homogenates using Trizol reagent according to the manufacturer's instructions and quantified by measuring the absorbance at 260 nm. RNA quality was determined by measuring the 260/280 ratio (>2.0) (Wang X. H. et al., 2016). M-MULV cDNA reverse transcription kit was utilized to synthesize first strand cDNA, according to the manufacturer's protocol. Briefly, 4 μg (0.4 μl) of total RNA from each sample was added to a mixture of 1 μl reverse transcriptase random primers (0.2 μg/μl), 4 μl of 5 × Reaction Buffer, 1 μl of RNase Inhibitor (20U/μl), 2 μl of dNTP Mix (10 mmol/L), 1 μl of M-MULV RT(200U/μl), and 10.6 μl of RNase free ddH2O. The final reaction mixture was kept at 25°C for 10 min, and then heated to 42°C for 50 min, followed by 70°C for 10 min, and finally cooled to 4°C.
Quantification PCR Analysis
Quantitative analysis of specific gene mRNA expression was performed via qPCR by subjecting the cDNA obtained from the above preparation to the LightCycle instrument (Roche). The 20 μl reaction mixture contained o.4 μl of forward primer and 0.4 μl of reverse primer, 10 μl of SYBR Green PCR master mix, 1 μl of cDNA sample, and 8.2 nuclease-free water. The primers used in these assays are listed in Table 1. The amplification conditions were as follows: 95°C for 10 min, 40 cycles of 95°C for 15 s, 60°C for 60 s, and 72°C for 60 s. The mRNA expression levels were determined relative to the blank control after normalization to the β-actin level through the 2−ΔΔCt method (Livak and Schmittgen, 2001). Analysis was carried out in triplicates.
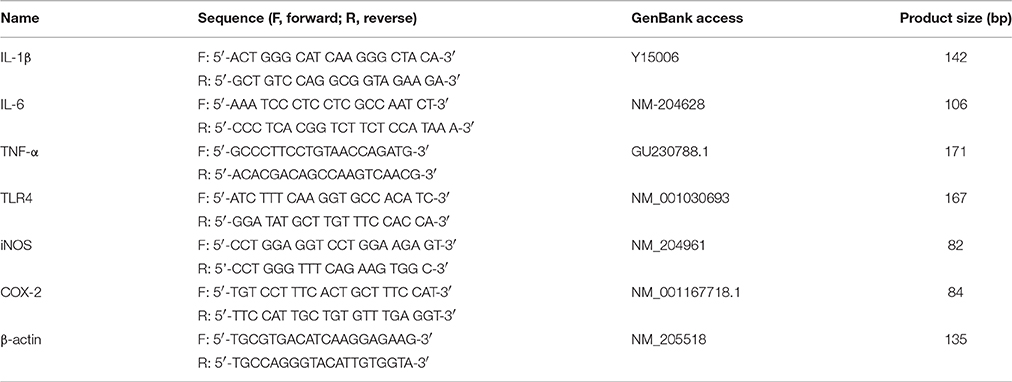
Table 1. Primers used for the quantitative real-time PCR.
Western Blotting Analysis
Liver samples were dissociated by RIPA lysis buffer (P0013B Beyotime Biotechnology, Jiangsu, China) supplemented with protease inhibitor mixture (Roche Applied Science, Indianapolis, USA) and centrifuged at 12,000 g for 15 min at 4°C. Nuclear extracts for p65 were prepared from hepatic homogenates as described previously (Tunon et al., 2011). The protein concentrations were determined by using the BCA protein assay kit (Beyotime, China). Subsequently, samples with the same amount of protein (80 μg) were fractionated using 10% SDS-PAGE (Beyotime, China) and followed by transferring onto polyvinylidene fluoride (PVDF) membranes. These PVDF membranes were blocked with 5% skimmed milk for 2 h and then incubated with special primary antibody (1:1,000 dilution) at 4°C overnight. Afterwards, the membranes were incubated with the corresponding HRP labeled secondary antibodies (1:4,000 dilution) at 37°C for 2 h, and then washed three times using TBST buffer. Finally, protein level was determined using the enhanced chemiluminescent (ECL) reagent (Beyotime, China) and the images were captured with Azure Bio-imaging systems (California, USA). Quantitative analysis was carried out using Amersham Imager 600 (Fairfield, USA). β-actin or Lamin-B were used as the internal control.
Statistical Analysis
SPSS 17.0 software was used to analyze the data. The results are presented as the mean ± SD. The differences between groups were analyzed by one-way ANOVA. A p-value of < 0.05 was considered to be statistically significant.
Results
Effects of Baicalin on LPS-Induced Clinical Symptoms of Chickens
Compared to control group, the chickens in the LPS group showed symptoms of drowsiness, lethargy, ruffled feathers and slight diarrhea at 24 h after injection. In the Baicalin pretreated groups the symptoms were milder than the LPS group. In addition, the cloacal temperature of the chickens in the LPS group was elevated markedly compared with the control group at 24 h after injection, Baicalin pretreatment (50, 100, and 200 mg/kg) appeared to significantly alleviate the increased cloacal temperature induced by LPS (P < 0.05, Figure 1).
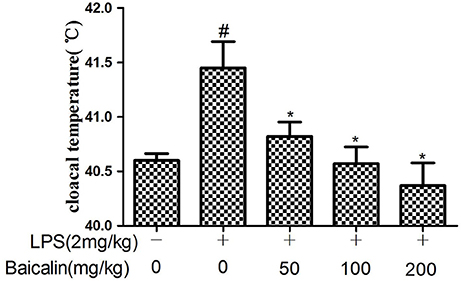
Figure 1. Effects of Baicalin on LPS-induced cloacal temperature. Data are expressed as the mean ± SD. (n = 10). *P < 0.05 vs. LPS group; #P < 0.05 vs. control group.
Baicalin Decreased Serum AST and ALT Levels after LPS Challenge
The severity of LPS-induced liver injury was determined through serum AST and ALT levels as well as histological findings. Serum levels of AST and ALT drastically increased in response to LPS at 24 h after injection, and Baicalin pretreatment (50, 100, and 200 mg/kg) significantly suppressed the release of transaminases ALT and AST into the plasma (P < 0.05, Figures 2A,B).
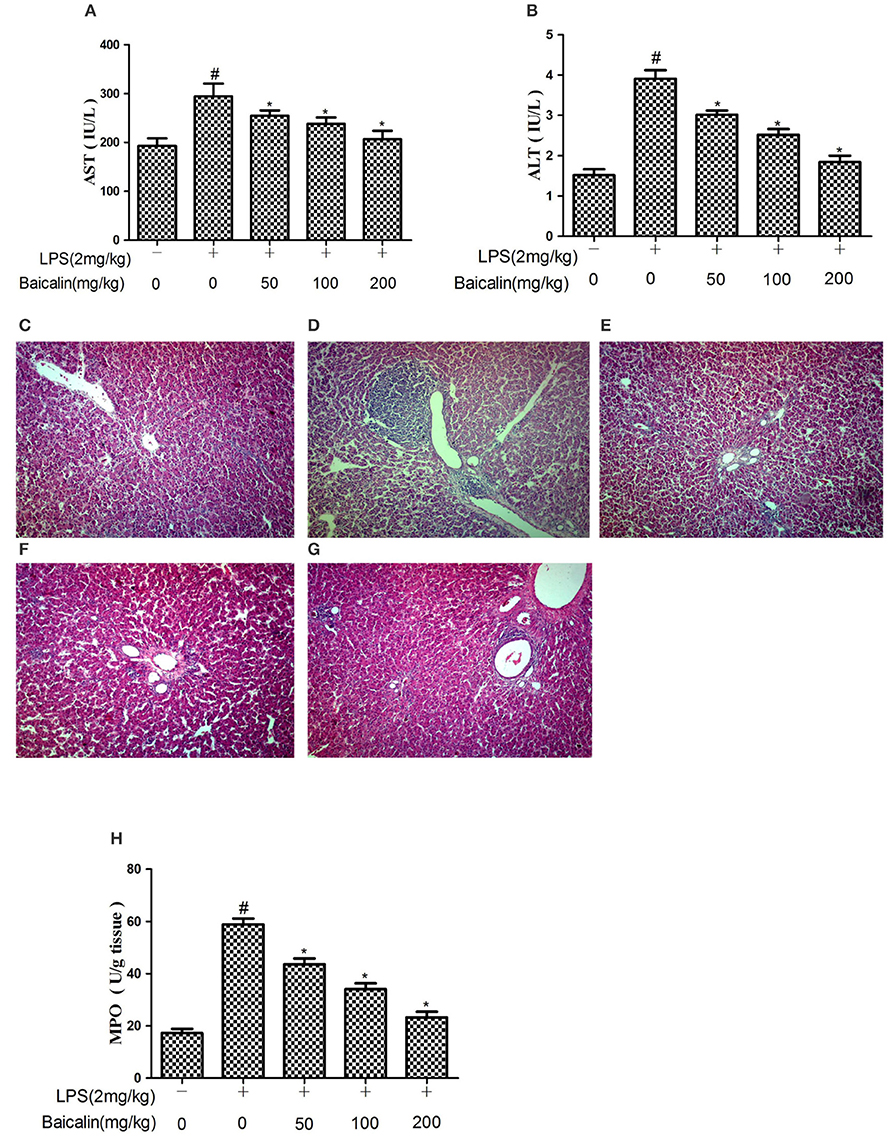
Figure 2. Baicalin pretreatment alleviated LPS-induced liver inflammation. Effects of Baicalin on LPS-induced serum AST (A) and ALT (B) levels. Histopathological changes in liver after LPS stimulation (HE, ×100). (C) control group, (D) LPS group, (E–G) Baicalin (50, 100, 200 mg/kg BW)+LPS groups. And effects of Baicalin on LPS-induced MPO activity in liver (H). Data are expressed as the mean ± SD. (n = 10). *P < 0.05 vs. LPS group; #P < 0.05 vs. control group.
Effects of Baicalin on LPS-Induced Liver Inflammation
The histological analysis and MPO detection were performed to estimate the extent of damage of liver tissues. As shown in Figure 2C, the result of histological analysis displayed a complete morphology and no histopathologic changes in control group. The LPS group had extensive inflammatory cells infiltration around the central veins and even some areas of necrosis within the liver lobules (Figure 2D). The inflammatory cells infiltration was reduced, and the structure of the liver was comparatively complete in the Baicalin pretreated groups (Figures 2E–G). In order to estimate the effectiveness of Baicalin on LPS-stimulated liver inflammation, further analysis of MPO activity was carried out. As shown in Figure 2H, LPS significantly increased the MPO activity. Baicalin pretreatment (50, 100, and 200 mg/kg) dramatically decreased the MPO level in a dose-dependent manner (p < 0.05).
Effects of Baicalin on NO Production Induced by LPS
The results showed that the NO production of liver tissues in the LPS group was approximately two-fold greater than the control group, Baicalin pretreatment had significant inhibitory effects on NO production in a concentration-dependent manner. (P < 0.05, Figure 3A) To further explore the effects of Baicalin on NO production, qPCR was performed to determine the iNOS mRNA expression in liver tissues. As expected, the iNOS mRNA expression was significantly up-regulated in response to LPS, Baicalin pretreatment could significantly reduce the iNOS mRNA expression induced by LPS compared with the LPS group (P < 0.05, Figure 3B). This implied that Baicalin could inhibit NO production stimulated by LPS through inhibiting iNOS gene expression at the transcriptional level.
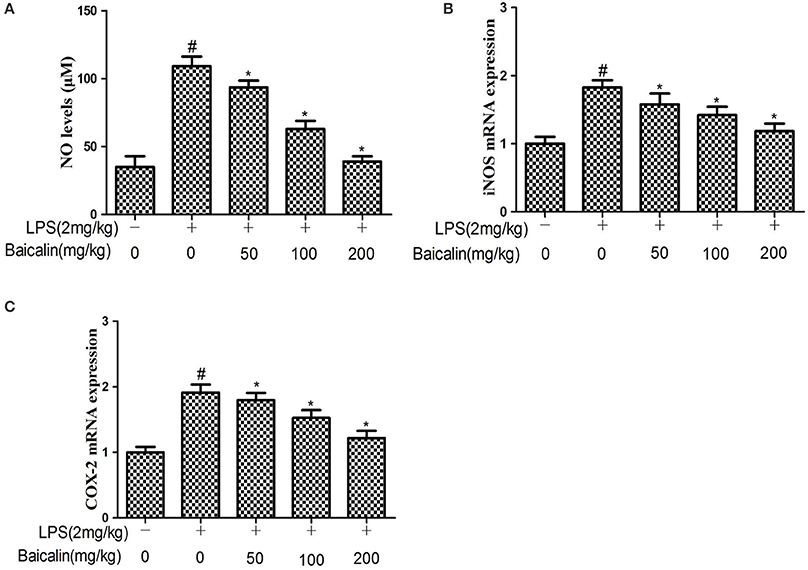
Figure 3. Effects of Baicalin on LPS-induced NO levels (A) and iNOS (B) and COX-2 mRNA expression (C) in liver. Data are expressed as the mean ± SD. (n = 10). *P < 0.05 vs. LPS group; #P < 0.05 vs. control group.
Baicalin Pretreatment Suppressed the Expression of COX-2 Induced by LPS
The effects of Baicalin on the expression COX-2 was determined by qPCR. The results showed that LPS increased COX-2 mRNA expression compared with the control group, while Baicalin pretreatment (50, 100, and 200 mg/kg) could significantly reduce the liver COX-2 mRNA expression in a dose-dependent manner compared with the LPS group (P < 0.05, Figure 3C).
Baicalin Pretreatment Suppressed the Production of Proinflammatory Mediators Induced by LPS
The expressions of cytokines in liver tissues were determined by ELISA and qPCR assays, respectively. ELISA results showed that LPS significantly increased the protein expression of TNF-α, IL-6, and 1L-1β. Baicalin pretreatment significantly attenuated the expression of TNF-α, IL-6, and 1L-1β in a concentration-dependent manner. Similarly, qPCR results suggested that LPS significantly increased the mRNA expression of TNF-α, IL-6, and 1L-1β compared with the control group. Baicalin pretreatment dose-dependently suppressed the mRNA expression of TNF-α, IL-6, and 1L-1β. These results showed that Baicalin inhibited the levels of pro-inflammatory cytokines induced by LPS (P < 0.05, Figure 4).
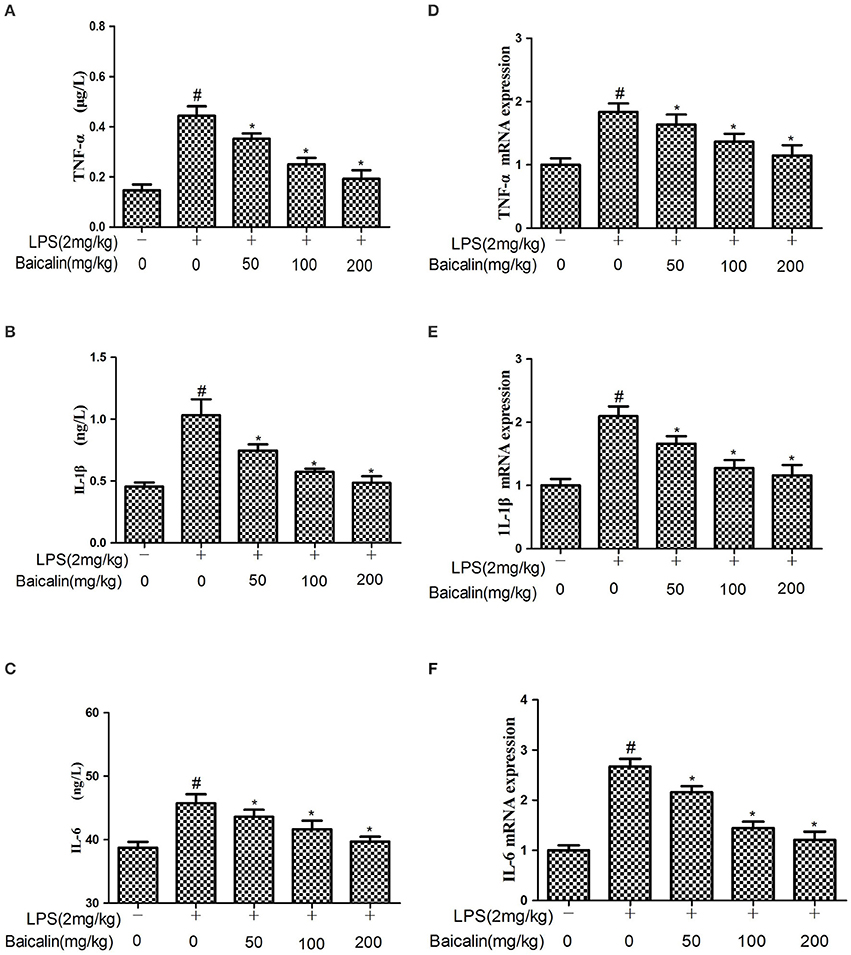
Figure 4. Effects of Baicalin on the production of cytokines. (A–C) The expression of TNF-α, IL-1β and IL-6 proteins induced by LPS. (D–F) The expression of TNF-α, IL-1β and IL-6 mRNA induced by LPS. Data are expressed as the mean ± SD (n = 10). *P < 0.05 vs. LPS group; #P < 0.05 vs. control group.
Baicalin Pretreatment Inhibited TLR4 mRNA and Protein Expression in LPS-Induced Liver Inflammation
The effects of Baicalin on the expression of TLR4 was determined by qPCR and western blot in liver tissues. The results showed that the mRNA and protein expressions of TLR4 were increased in the LPS group. Baicalin pretreatment inhibited the LPS-induced TLR4 mRNA and protein expression in a dose-dependent manner in liver tissues. (P < 0.05, Figure 5).
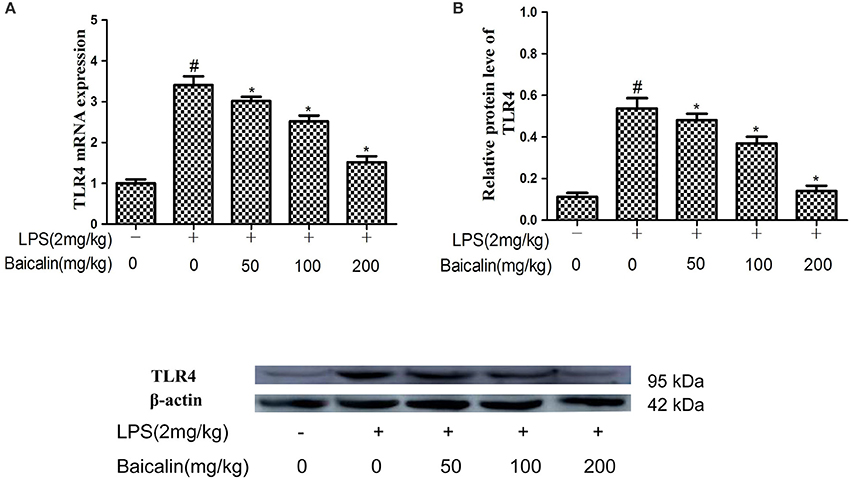
Figure 5. Effects of Baicalin on TLR4 expression. TLR4 mRNA levels (A), TLR4 protein levels (B). Data are expressed as the mean ± SD (n = 10). *P < 0.05 vs. LPS group; #P < 0.05 vs. control group.
Baicalin Pretreatment Inhibited LPS-Induced NF-κB Pathway Activation in LPS-Induced Liver Inflammation
To determine whether the anti-inflammatory mechanism of Baicalin on LPS-induced liver inflammation act through the NF-κB pathway, western blot assays were carried out. Compared to control group, the phosphorylated p65 and IκBα proteins were significantly increased in the LPS group. Moreover, these changes were accompanied by an enhanced nuclear protein concentration of the NF-kB p65 subunit. In contrast, the proteins levels of p-p65, p-IκBα and p65 were dose-dependently reduced in the Baicalin groups (P < 0.05, Figure 6).
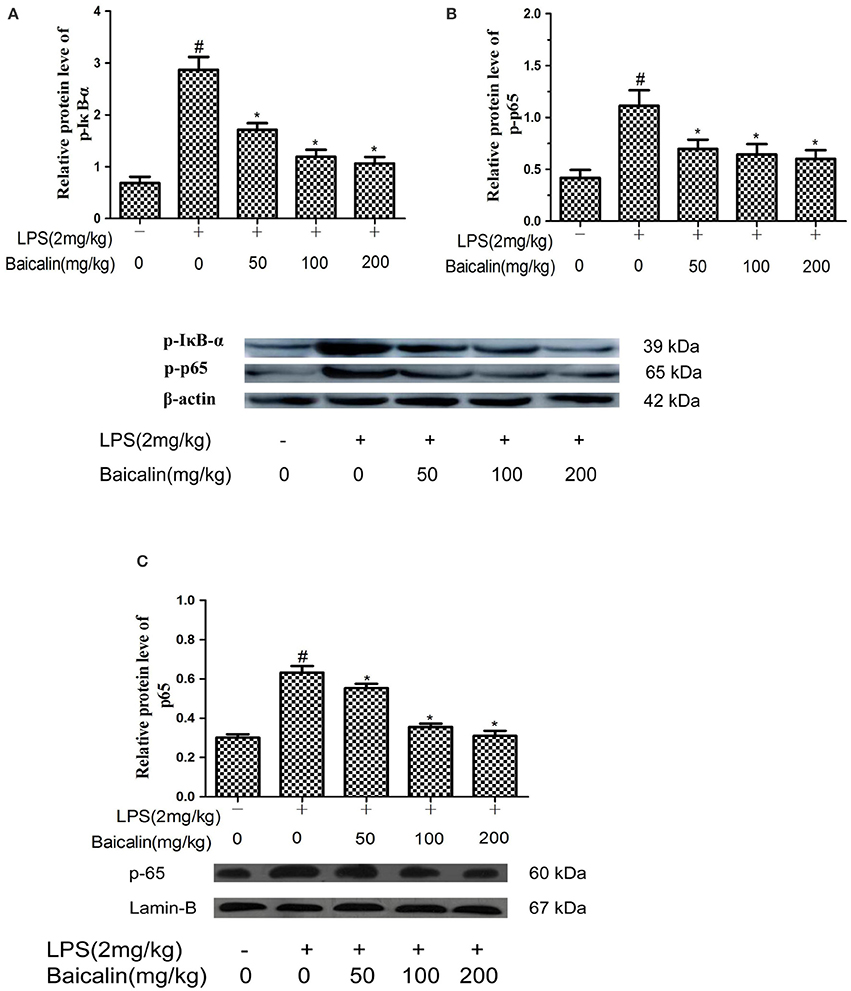
Figure 6. Baicalin inhibited NF-κB pathway activation. The expression of p-IkBα (A), p-p65 (B), and nuclear p65 (C). Data are expressed as the mean ± SD. (n = 6). *P < 0.05 vs. LPS group; #P < 0.05 vs. control group.
Discussion
Flavonoids are often as major pharmacological active constituents in numerous plant medicines, which exist in a wide variety of plants. As a kind of polyphenolic compounds, many studies have demonstrated that flavonoids possess vasodilatory, antibacterial, anti-inflammatory, immune-stimulating, and antiallergic properties (Middleton and Kandaswami, 1992; Duarte et al., 1993). Due to their outstanding antioxidant activity, people start to pay more attention on flavonoids. It has been reported that some flavonoids, including quercetin, luteolin and catechins, can become better antioxidants than the antioxidant nutrients vitamin C, vitamin E and β -carotene on a mole for mole basis (Rice-Evans et al., 1995). In addition, epidemiological studies showed that dietary intake of flavonoids is capable of reducing risk of cancer, inflammation and heart disease (Middleton et al., 2000). Scutellaria baicalensis as one of the most popular and multi-purpose herbs has been used in China for a long time, the growing evidence demonstrated that the molecular basis of the anti-inflammatory effects of Scutellaria is the bioactive phytochemical flavonoids. Indeed, the flavonoids extracted from the roots of Radix Scutellariae, such as Wogonin, Baicalein, and Baicalin possess multiple bioactivities including antioxidant and anti-inflammatory (Gao et al., 1999; Chi et al., 2003).
Baicalin has been reported to possess the anti-inflammatory and antioxidative properties (Li L. et al., 2012). However, the effects of it against LPS-induced liver injury in chicken and inflammatory cytokines involved have been seldom reported so far. In the present study, we observed the effects of Baicalin on LPS-induced liver inflammation. Our results demonstrated that Baicalin exhibited potential anti-inflammatory activity as evidenced by dose-dependent inhibition of the elevation of serum AST and ALT levels as well as histopathological finding. In addition, the MPO activity is often regarded as an early marker for the infiltration of inflammatory cells into tissues, which also predicts the occurrence of inflammatory diseases (Kubota et al., 2012). Thus, weakened MPO activity means decreased anti-inflammatory response. All results suggested that Baicalin had a protective action on LPS-induced liver inflammation in chicken.
Among other events, production of significant quantities of free radicals, nitrogen reactive species and cytokines is the characteristic of inflammatory disorders (Marcus et al., 2003; Korbecki et al., 2013). It has been reported that LPS could stimulate the release of TNF-α, IL-6, and IL-1β, and they are regarded as critical cytokines in the inflammatory cytokine family (Laveti et al., 2013). TNF-α plays a crucial role in exerting variety of biologic effects, which is secreted by activated macrophages and considered to be an efficient pro-inflammatory factor (Wu et al., 2015). IL-6 is necessary to leukocyte recruitment and tissue homeostasis, which is secreted from many cells induced by invasion or injury (Cronin et al., 2016). As a key factor of mediating inflammation, IL-1β is mainly produced by activated macrophages (Smirnova et al., 2002). In addition, TNF-α, IL-6, and IL-1β are able to promote the expression of iNOS (Marcus et al., 2003). Therefore, we can reduce inflammation and its symptoms by inhibiting the overproduction and activity of pro-inflammatory cytokines, which has been applied to clinic in the treatment of certain inflammatory diseases, such as rheumatoid arthritis, obesity, diabetes mellitus, cancer and atherosclerosis (Laveti et al., 2013). Our results showed that Baicalin protected the chicken from LPS-stimulated liver inflammation through decreasing the secretion of pro-inflammatory mediators.
As a kind of highly reactive oxidant, NO can be catalyzed by iNOS, which often participates in diverse biological mechanisms such as regulating mitochondrial functions, apoptosis, neuro transmission and inflammation, and even directly damaging normal cells (Koppula et al., 2012). iNOS is present at low levels under normal physiological conditions, but can get it to high levels rapidly when stimulated by pro-inflammatory mediators, such as LPS (Park et al., 2009). It has been reported that the increasing activity of iNOS could catalyze overproduction of NO which can induce septic shock, cause tissue damage, and contribute to developing pathological complication during ongoing chronic inflammatory response (Yang et al., 2009). Some studies have shown that several inhibitors of NO induction exert anti-inflammatory effects through suppressing iNOS expression (Kim J. Y. et al., 2005). In this research, the results indicated that Baicalin significantly suppressed the production of NO as well as iNOS expression stimulated by LPS in liver tissues, suggesting that inhibition of iNOS enzyme is one of the anti-inflammatory mechanisms of Baicalin.
It's well known that COX-2 can participate in synthesizing eicosanoids as an inducible enzyme during the inflammatory process (Vane et al., 1998). It has been demonstrated that iNOS is highly expressed in cytokines-stimulated macrophages or phorbol ester-stimulated mouse skin through activation of NF-κB and MAPKs (Gust et al., 1999; Kim S. O. et al., 2005). And some studies have provided convincing evidence that overproduction of COX-2 can promote the pathological development of inflammatory-related chronic diseases including rheumatoid arthritis, cancer, and cardiovascular disease (Cerella et al., 2010). In this study, our results showed that the Baicalin pretreatment markedly inhibited the LPS-induced COX-2 mRNA expression in liver tissues. Thus, it indicated that the protective effects of Baicalin against LPS-induced chicken liver inflammation is related with the selective down-regulation of COX-2 in some extent.
TLRs play a great role in the process of liver fibrosis which is caused by many common liver pathogens, including viral, and toxin-induced hepatitis (Cengiz et al., 2015). As an important receptor for LPS, it is well known that the activation of TLR4 by LPS results in the secretion of crucial pro-inflammatory cytokines which are crucial to stimulate potent immune responses (Gorina et al., 2011; Mateu et al., 2015). Many studies have provided convincing evidence that the NF-κB signaling pathway can be activated when LPS triggers the immune response via TLR4 (Byun et al., 2013, 2015). To further understand the mechanism by which Baicalin exerts its anti-inflammatory property, we investigated the effects of Baicalin on the local activities of TLR4 and NF-κB signaling pathway in LPS-induced liver inflammation. The results showed that Baicalin could inhibit the expression of TLR4 in a dose-dependent manner.
As a nuclear transcription factor, NF-κB plays an important role in inflammatory reaction, and the activation of NF-κB can aggravate the expression of pro-inflammatory mediators (Li and Verma, 2002; Morris et al., 2003). NF-kB is sequestered in the cytoplasm through its linkage to the inhibitor of κB (IκB) protein under normal physical condition. Upon activation by LPS, phosphorylation of IκBα activates the NF-κB pathway, which results in NF-kB p65 dissociates from IκBα, then the released NF-κB p65 translocates into the nucleus, and triggers the transcription of related inflammatory genes (Sun and Andersson, 2002; Li et al., 2014). Some studies have reported that Con A could upregulate NF-kB expression in liver (Tiegs et al., 1998), and there was increasing evidence shown that Con A-induced liver injury was significantly attenuated via inhibiting NF-kB activation (Li Y. et al., 2012; Song et al., 2013). In the present study, the results showed that Baicalin obviously inhibited the protein levels of p-p65, p- IκBα and p65 induced by LPS in liver tissues.
Conclusion
In conclusion, our results demonstrate that Baicalin can attenuate LPS-induced inflammatory response in chicken liver. The protective effects of it are due to the inhibition of oxidative stress and the release of inflammatory mediators. In addition, the anti-inflammatory effects of Baicalin are associated with inhibiting the activation of the TLR4-mediated NF-κB signaling pathway.
Ethics Statement
All of the experiments were conducted under the supervision of the Harbin Veterinary Research Institute of the Chinese Academy of Agricultural Sciences in accordance with animal ethics guidelines and approved protocols. The Harbin Veterinary Research Institute Animal Ethics Committee approval number was SYXK (Hei) 2012-2067.
Author Contributions
XZ supervised the whole experiments. PC performed the practical work and completed the experiments, TW, WL, HW, XS, YY, JL, and TX provided help during the experiments. IM helped in improving language expression.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This study was supported by the Harbin Science and Technology Innovation Fund (2013RFXXJ035).
References
Bocchini, V., Mazzolla, R., Barluzzi, R., Blasi, E., Sick, P., and Kettenmann, H. (1992). An immortalized cell line expresses properties of activated microglial cells. J. Neurosci. Res. 31, 616–621. doi: 10.1002/jnr.490310405
Brigham, K. L., and Meyrick, B. (1986). Endotoxin and lung injury. Am. Rev. Respir Dis. 133, 913–927.
Byun, E. B., Sung, N. Y., Byun, E. H., Song, D. S., Kim, J. K., Park, J. H., et al. (2013). The procyanidin trimer C1 inhibits LPS-induced MAPK and NF-kappaB signaling through TLR4 in macrophages. Int. Immunopharmacol. 15, 450–456. doi: 10.1016/j.intimp.2012.11.021
Byun, E. B., Sung, N. Y., Park, J. N., Yang, M. S., Park, S. H., and Byun, E. H. (2015). Gamma-irradiated resveratrol negatively regulates LPS-induced MAPK and NF-kappaB signaling through TLR4 in macrophages. Int. Immunopharmacol. 25, 249–259. doi: 10.1016/j.intimp.2015.02.015
Cai, X., Li, C., Du, G., and Cao, Z. (2008). Protective effects of baicalin on ligature-induced periodontitis in rats. J. Periodontal Res. 43, 14–21. doi: 10.1111/j.1600-0765.2007.00989.x
Cengiz, M., Ozenirler, S., and Elbeg, S. (2015). Role of serum toll-like receptors 2 and 4 in non-alcoholic steatohepatitis and liver fibrosis. J. Gastroenterol. Hepatol. 30, 1190–1196. doi: 10.1111/jgh.12924
Cerella, C., Sobolewski, C., Dicato, M., and Diederich, M. (2010). Targeting COX-2 expression by natural compounds: a promising alternative strategy to synthetic COX-2 inhibitors for cancer chemoprevention and therapy. Biochem. Pharmacol. 80, 1801–1815. doi: 10.1016/j.bcp.2010.06.050
Cheng, Y., Ping, J., Xu, H. D., Fu, H. J., and Zhou, Z. H. (2006). Synergistic effect of a novel oxymatrine-baicalin combination against hepatitis B virus replication, alpha smooth muscle actin expression and type I collagen synthesis in vitro. World J. Gastroenterol. 12, 5153–5159. doi: 10.3748/wjg.v12.i32.5153
Chi, Y. S., Lim, H., Park, H., and Kim, H. P. (2003). Effects of wogonin, a plant flavone from Scutellaria radix, on skin inflammation: in vivo regulation of inflammation-associated gene expression. Biochem. Pharmacol. 66, 1271–1278. doi: 10.1016/S0006-2952(03)00463-5
Cronin, J. G., Kanamarlapudi, V., Thornton, C. A., and Sheldon, I. M. (2016). Signal transducer and activator of transcription-3 licenses toll-like receptor 4-dependent interleukin (IL)-6 and IL-8 production via IL-6 receptor-positive feedback in endometrial cells. Mucosal Immunol. 9, 1125–1136. doi: 10.1038/mi.2015.131
Cui, L., Feng, L., Zhang, Z. H., and Jia, X. B. (2014). The anti-inflammation effect of baicalin on experimental colitis through inhibiting TLR4/NF-kappaB pathway activation. Int. Immunopharmacol. 23, 294–303. doi: 10.1016/j.intimp.2014.09.005
da Silva Correia, J., Soldau, K., Christen, U., Tobias, P. S., and Ulevitch, R. J. (2001). Lipopolysaccharide is in close proximity to each of the proteins in its membrane receptor complex. transfer from CD14 to TLR4 and MD-2. J. Biol. Chem. 276, 21129–21135. doi: 10.1074/jbc.M009164200
Duarte, J., Perez-Vizcaino, F., Zarzuelo, A., Jimenez, J., and Tamargo, J. (1993). Vasodilator effects of quercetin in isolated rat vascular smooth muscle. Eur. J. Pharmacol. 239, 1–7. doi: 10.1016/0014-2999(93)90968-N
Fan, X., Zhang, Y., Dong, H., Wang, B., Ji, H., and Liu, X. (2015). Trilobatin attenuates the LPS-mediated inflammatory response by suppressing the NF-kappaB signaling pathway. Food Chem. 166, 609–615. doi: 10.1016/j.foodchem.2014.06.022
Gao, Z., Huang, K., Yang, X., and Xu, H. (1999). Free radical scavenging and antioxidant activities of flavonoids extracted from the radix of Scutellaria baicalensis Georgi. Biochim. Biophys. Acta 1472, 643–650. doi: 10.1016/S0304-4165(99)00152-X
Gorina, R., Font-Nieves, M., Marquez-Kisinousky, L., Santalucia, T., and Planas, A. M. (2011). Astrocyte TLR4 activation induces a proinflammatory environment through the interplay between MyD88-dependent NFkappaB signaling, MAPK, and Jak1/Stat1 pathways. Glia 59, 242–255. doi: 10.1002/glia.21094
Gust, R., Kozlowski, J. K., Stephenson, A. H., and Schuster, D. P. (1999). Role of cyclooxygenase-2 in oleic acid-induced acute lung injury. Am. J. Respir Crit. Care Med. 160, 1165–1170. doi: 10.1164/ajrccm.160.4.9811073
Guven-Maiorov, E., Keskin, O., Gursoy, A., and Nussinov, R. (2015). A structural view of negative regulation of the toll-like receptor-mediated inflammatory pathway. Biophys. J. 109, 1214–1226. doi: 10.1016/j.bpj.2015.06.048
Hwang, J. M., Wang, C. J., Chou, F. P., Tseng, T. H., Hsieh, Y. S., Hsu, J. D., et al. (2005). Protective effect of baicalin on tert-butyl hydroperoxide-induced rat hepatotoxicity. Arch. Toxicol. 79, 102–109. doi: 10.1007/s00204-004-0588-6
Kagan, J. C., and Medzhitov, R. (2006). Phosphoinositide-mediated adaptor recruitment controls toll-like receptor signaling. Cell 125, 943–955. doi: 10.1016/j.cell.2006.03.047
Kim, H. Y., Kim, J. K., Choi, J. H., Jung, J. Y., Oh, W. Y., Kim, D. C., et al. (2010). Hepatoprotective effect of pinoresinol on carbon tetrachloride-induced hepatic damage in mice. J. Pharmacol. Sci. 112, 105–112. doi: 10.1254/jphs.09234FP
Kim, J. Y., Lee, H. J., Kim, J. S., and Ryu, J. H. (2005). Induction of nitric oxide synthase by saponins of heat-processed ginseng. Biosci. Biotechnol. Biochem. 69, 891–895. doi: 10.1271/bbb.69.891
Kim, S. H., Jeong, H., Kim, Y. K., Cho, S. H., Min, K. U., and Kim, Y. Y. (2001). IgE-mediated occupational asthma induced by herbal medicine, Banha (Pinellia ternata). Clin. Exp. Allergy 31, 779–781. doi: 10.1046/j.1365-2222.2001.01067.x
Kim, S. O., Kundu, J. K., Shin, Y. K., Park, J. H., Cho, M. H., Kim, T. Y., et al. (2005). [6]-Gingerol inhibits COX-2 expression by blocking the activation of p38 MAP kinase and NF-kappaB in phorbol ester-stimulated mouse skin. Oncogene 24, 2558–2567. doi: 10.1038/sj.onc.1208446
Kim, T. H., Yoon, H. J., Lim, C. M., Kim, E. K., Kim, M. J., and Koh, Y. (2005). The role of endogenous histamine on the pathogenesis of the lipopolysaccharide (LPS)-induced, acute lung injury: a pilot study. Inflammation 29, 72–80. doi: 10.1007/s10753-006-9001-3
Kim, Y. S., Hwang, J. W., Jang, J. H., Son, S., Seo, I. B., Jeong, J. H., et al. (2016). Trapa japonica pericarp extract reduces LPS-Induced inflammation in macrophages and acute lung injury in mice. Molecules 21:392. doi: 10.3390/molecules21030392
Koppula, S., Kumar, H., Kim, I. S., and Choi, D. K. (2012). Reactive oxygen species and inhibitors of inflammatory enzymes, NADPH oxidase, and iNOS in experimental models of Parkinson's disease. Mediators Inflamm. 2012:823902. doi: 10.1155/2012/823902
Korbecki, J., Baranowska-Bosiacka, I., Gutowska, I., and Chlubek, D. (2013). The effect of reactive oxygen species on the synthesis of prostanoids from arachidonic acid. J. Physiol. Pharmacol. 64, 409–421.
Kubota, K., Saiwai, H., Kumamaru, H., Maeda, T., Ohkawa, Y., Aratani, Y., et al. (2012). Myeloperoxidase exacerbates secondary injury by generating highly reactive oxygen species and mediating neutrophil recruitment in experimental spinal cord injury. Spine 37, 1363–1369. doi: 10.1097/BRS.0b013e31824b9e77
Kwon, D. J., Bae, Y. S., Ju, S. M., Youn, G. S., Choi, S. Y., and Park, J. (2014). Salicortin suppresses lipopolysaccharide-stimulated inflammatory responses via blockade of NF-kappaB and JNK activation in RAW 264.7 macrophages. BMB Rep. 47, 318–323. doi: 10.5483/BMBRep.2014.47.6.200
Laveti, D., Kumar, M., Hemalatha, R., Sistla, R., Naidu, V. G., Talla, V., et al. (2013). Anti-inflammatory treatments for chronic diseases: a review. Inflamm. Allergy Drug Targets 12, 349–361. doi: 10.2174/18715281113129990053
Lee, W., Ku, S. K., and Bae, J. S. (2015). Anti-inflammatory effects of Baicalin, Baicalein, and Wogonin in vitro and in vivo. Inflammation 38, 110–125. doi: 10.1007/s10753-014-0013-0
Li, J., Zhong, W., Wang, W., Hu, S., Yuan, J., Zhang, B., et al. (2014). Ginsenoside metabolite compound K promotes recovery of dextran sulfate sodium-induced colitis and inhibits inflammatory responses by suppressing NF-kappaB activation. PLoS ONE 9:e87810. doi: 10.1371/journal.pone.0087810
Li, L., Bao, H., Wu, J., Duan, X., Liu, B., Sun, J., et al. (2012). Baicalin is anti-inflammatory in cigarette smoke-induced inflammatory models in vivo and in vitro: a possible role for HDAC2 activity. Int. Immunopharmacol. 13, 15–22. doi: 10.1016/j.intimp.2012.03.001
Li, Q., and Verma, I. M. (2002). NF-kappaB regulation in the immune system. Nat. Rev. Immunol. 2, 725–734. doi: 10.1038/nri910
Li, Y., Wang, Z. L., He, F., Wu, Y., Huang, W., He, Y., et al. (2012). TP-58, a novel thienopyridine derivative, protects mice from concanavalinA-induced hepatitis by suppressing inflammation. Cell. Physiol. Biochem. 29, 31–40. doi: 10.1159/000337584
Lian, L. H., Jin, X., Wu, Y. L., Cai, X. F., Lee, J. J., and Nan, J. X. (2010). Hepatoprotective effects of Sedum sarmentosum on D-galactosamine/lipopolysaccharide-induced murine fulminant hepatic failure. J. Pharmacol. Sci. 114, 147–157. doi: 10.1254/jphs.10045FP
Lin, M., Zhai, X., Wang, G., Tian, X., Gao, D., Shi, L., et al. (2015). Salvianolic acid B protects against acetaminophen hepatotoxicity by inducing Nrf2 and phase II detoxification gene expression via activation of the PI3K and PKC signaling pathways. J. Pharmacol. Sci. 127, 203–210. doi: 10.1016/j.jphs.2014.12.010
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Lv, X., Fu, K., Li, W., Wang, Y., Wang, J., Li, H., et al. (2015). TIIA attenuates LPS-induced mouse endometritis by suppressing the NF-kappaB signaling pathway. Can. J. Physiol. Pharmacol. 93, 967–971. doi: 10.1139/cjpp-2015-0003
Malhi, H., and Kaufman, R. J. (2011). Endoplasmic reticulum stress in liver disease. J. Hepatol. 54, 795–809. doi: 10.1016/j.jhep.2010.11.005
Marcus, J. S., Karackattu, S. L., Fleegal, M. A., and Sumners, C. (2003). Cytokine-stimulated inducible nitric oxide synthase expression in astroglia: role of Erk mitogen-activated protein kinase and NF-kappaB. Glia 41, 152–160. doi: 10.1002/glia.10168
Mateu, A., Ramudo, L., Manso, M. A., and De Dios, I. (2015). Cross-talk between TLR4 and PPARgamma pathways in the arachidonic acid-induced inflammatory response in pancreatic acini. Int. J. Biochem. Cell Biol. 69, 132–141. doi: 10.1016/j.biocel.2015.10.022
Middleton, E. Jr., and Kandaswami, C. (1992). Effects of flavonoids on immune and inflammatory cell functions. Biochem. Pharmacol. 43, 1167–1179. doi: 10.1016/0006-2952(92)90489-6
Middleton, E. Jr., Kandaswami, C., and Theoharides, T. C. (2000). The effects of plant flavonoids on mammalian cells: implications for inflammation, heart disease, and cancer. Pharmacol. Rev. 52, 673–751.
Morris, K. R., Lutz, R. D., Choi, H. S., Kamitani, T., Chmura, K., and Chan, E. D. (2003). Role of the NF-kappaB signaling pathway and kappaB cis-regulatory elements on the IRF-1 and iNOS promoter regions in mycobacterial lipoarabinomannan induction of nitric oxide. Infect. Immun. 71, 1442–1452. doi: 10.1128/IAI.71.3.1442-1452.2003
Munyaka, P. M., Tactacan, G., Jing, M. O. K., House, J. D., and Rodriguez-Lecompte, J. C. (2012). Immunomodulation in young laying hens by dietary folic acid and acute immune responses after challenge with Escherichia coli lipopolysaccharide. Poult. Sci. 91, 2454–2463. doi: 10.3382/ps.2012-02381
Nagai, Y., Akashi, S., Nagafuku, M., Ogata, M., Iwakura, Y., Akira, S., et al. (2002). Essential role of MD-2 in LPS responsiveness and TLR4 distribution. Nat. Immunol. 3, 667–672. doi: 10.1038/ni809
National Pharmacopoeia Committee (2015). Pharmacopoeia of People's Republic of China. Part 2. Beijing: China Medical Science and Technology Press.
Pahl, H. L. (1999). Activators and target genes of Rel/NF-kappaB transcription factors. Oncogene 18, 6853–6866. doi: 10.1038/sj.onc.1203239
Park, E. J., Lee, J. H., Yu, G. Y., He, G., Ali, S. R., Holzer, R. G., et al. (2010). Dietary and genetic obesity promote liver inflammation and tumorigenesis by enhancing IL-6 and TNF expression. Cell 140, 197–208. doi: 10.1016/j.cell.2009.12.052
Park, J. S., Park, E. M., Kim, D. H., Jung, K., Jung, J. S., Lee, E. J., et al. (2009). Anti-inflammatory mechanism of ginseng saponins in activated microglia. J. Neuroimmunol. 209, 40–49. doi: 10.1016/j.jneuroim.2009.01.020
Qian, H., Shi, J., Fan, T. T., Lv, J., Chen, S. W., Song, C. Y., et al. (2014). Sophocarpine attenuates liver fibrosis by inhibiting the TLR4 signaling pathway in rats. World J. Gastroenterol. 20, 1822–1832. doi: 10.3748/wjg.v20.i7.1822
Rice-Evans, C. A., Miller, N. J., Bolwell, P. G., Bramley, P. M., and Pridham, J. B. (1995). The relative antioxidant activities of plant-derived polyphenolic flavonoids. Free Radic. Res. 22, 375–383. doi: 10.3109/10715769509145649
Shin, K. M., Kim, Y. H., Park, W. S., Kang, I., Ha, J., Choi, J. W., et al. (2004). Inhibition of methanol extract from the fruits of Kochia scoparia on lipopolysaccharide-induced nitric oxide, prostaglandin [correction of prostagladin] E2, and tumor necrosis factor-alpha production from murine macrophage RAW 264.7 cells. Biol. Pharm. Bull. 27, 538–543. doi: 10.1248/bpb.27.538
Smirnova, M. G., Kiselev, S. L., Gnuchev, N. V., Birchall, J. P., and Pearson, J. P. (2002). Role of the pro-inflammatory cytokines tumor necrosis factor-alpha, interleukin-1 beta, interleukin-6 and interleukin-8 in the pathogenesis of the otitis media with effusion. Eur. Cytokine Netw. 13, 161–172.
Song, B., Huang, G., Tong, C., Li, G., Wang, Z., Xiong, Y., et al. (2013). Gossypol suppresses mouse T lymphocytes via inhibition of NFkB, NFAT and AP-1 pathways. Immunopharmacol. Immunotoxicol. 35, 615–621. doi: 10.3109/08923973.2013.830126
Sun, Z., and Andersson, R. (2002). NF-kappaB activation and inhibition: a review. Shock 18, 99–106. doi: 10.1097/00024382-200208000-00001
Takeuchi, O., and Akira, S. (2010). Pattern recognition receptors and inflammation. Cell 140, 805–820. doi: 10.1016/j.cell.2010.01.022
Tiegs, G., Kusters, S., Kunstle, G., Hentze, H., Kiemer, A. K., and Wendel, A. (1998). Ebselen protects mice against T cell-dependent, TNF-mediated apoptotic liver injury. J. Pharmacol. Exp. Ther. 287, 1098–1104.
Tunon, M. J., San Miguel, B., Crespo, I., Jorquera, F., Santamaria, E., Alvarez, M., et al. (2011). Melatonin attenuates apoptotic liver damage in fulminant hepatic failure induced by the rabbit hemorrhagic disease virus. J. Pineal Res. 50, 38–45. doi: 10.1111/j.1600-079X.2010.00807.x
Vane, J. R., Bakhle, Y. S., and Botting, R. M. (1998). Cyclooxygenases 1 and 2. Annu. Rev. Pharmacol. Toxicol. 38, 97–120. doi: 10.1146/annurev.pharmtox.38.1.97
Wang, J., Yi, M., Chen, X., Muhammad, I., Liu, F., Li, R., et al. (2016). Effects of colistin on amino acid neurotransmitters and blood-brain barrier in the mouse brain. Neurotoxicol. Teratol. 55, 32–37. doi: 10.1016/j.ntt.2016.03.004
Wang, X. H., Li, C. Y., Muhammad, I., and Zhang, X. Y. (2016). Fatty acid composition in serum correlates with that in the liver and non-alcoholic fatty liver disease activity scores in mice fed a high-fat diet. Environ. Toxicol. Pharmacol. 44, 140–150. doi: 10.1016/j.etap.2016.04.009
Wu, X., Xu, W., Feng, X., He, Y., Liu, X., Gao, Y., et al. (2015). TNF-a mediated inflammatory macrophage polarization contributes to the pathogenesis of steroid-induced osteonecrosis in mice. Int. J. Immunopathol. Pharmacol. 28, 351–361. doi: 10.1177/0394632015593228
Xu, J., Zhang, X., Monestier, M., Esmon, N. L., and Esmon, C. T. (2011). Extracellular histones are mediators of death through TLR2 and TLR4 in mouse fatal liver injury. J. Immunol. 187, 2626–2631. doi: 10.4049/jimmunol.1003930
Yang, G. Y., Taboada, S., and Liao, J. (2009). Inflammatory bowel disease: a model of chronic inflammation-induced cancer. Methods Mol. Biol. 511, 193–233. doi: 10.1007/978-1-59745-447-6_9
Yu, F. Y., Huang, S. G., Zhang, H. Y., Ye, H., Chi, H. G., Zou, Y., et al. (2014). Effects of baicalin in CD4 + CD29 + T cell subsets of ulcerative colitis patients. World J. Gastroenterol. 20, 15299–15309. doi: 10.3748/wjg.v20.i41.15299
Zhai, X. T., Zhang, Z. Y., Jiang, C. H., Chen, J. Q., Ye, J. Q., Jia, X. B., et al. (2016). Nauclea officinalis inhibits inflammation in LPS-mediated RAW 264.7 macrophages by suppressing the NF-kappaB signaling pathway. J. Ethnopharmacol. 183, 159–165. doi: 10.1016/j.jep.2016.01.018
Zhao, H. X., Zhao, J. L., Shen, J. Z., Fan, H. L., Guan, H., An, X. P., et al. (2014). Prevalence and molecular characterization of fluoroquinolone resistance in Escherichia coli isolates from dairy cattle with endometritis in China. Microb. Drug Resist. 20, 162–169. doi: 10.1089/mdr.2013.0073
Zheng, X., Yang, D., Liu, X., Wang, N., Li, B., Cao, H., et al. (2010). Identification of a new anti-LPS agent, geniposide, from Gardenia jasminoides Ellis, and its ability of direct binding and neutralization of lipopolysaccharide in vitro and in vivo. Int. Immunopharmacol. 10, 1209–1219. doi: 10.1016/j.intimp.2010.07.001
Keywords: Baicalin, lipopolysaccharide, anti-inflammation, liver inflammation, TLR4, NF-κB
Citation: Cheng P, Wang T, Li W, Muhammad I, Wang H, Sun X, Yang Y, Li J, Xiao T and Zhang X (2017) Baicalin Alleviates Lipopolysaccharide-Induced Liver Inflammation in Chicken by Suppressing TLR4-Mediated NF-κB Pathway. Front. Pharmacol. 8:547. doi: 10.3389/fphar.2017.00547
Received: 10 March 2017; Accepted: 04 August 2017;
Published: 18 August 2017.
Edited by:
Annalisa Bruno, Università degli Studi “G. d'Annunzio” Chieti - Pescara, ItalyReviewed by:
Amanda Jane Gibson, Royal Veterinary College, United KingdomCyril Seillet, Walter and Eliza Hall Institute of Medical Research, Australia
Copyright © 2017 Cheng, Wang, Li, Muhammad, Wang, Sun, Yang, Li, Xiao and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xiuying Zhang, emhhbmd4aXV5aW5nQG5lYXUuZWR1LmNu