
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Pharmacol. , 23 June 2017
Sec. Ethnopharmacology
Volume 8 - 2017 | https://doi.org/10.3389/fphar.2017.00409
 Kemin Yan1†Huijuan Zhu1†Jian Xu1,2†Hui Pan1Naishi Li1Linjie Wang1Hongbo Yang1Meijuan Liu1
Kemin Yan1†Huijuan Zhu1†Jian Xu1,2†Hui Pan1Naishi Li1Linjie Wang1Hongbo Yang1Meijuan Liu1 FengYing Gong1*
FengYing Gong1*Objectives: Lotus leaf is a kind of traditional Chinese medicine. We aimed to explore the effects of lotus leaf aqueous extract (LLAE) on peroxisome proliferative activated receptor γ2 (PPARγ2) expression in preadipocytes and adipocytes and further investigate its effects on high fat diet (HFD)-induced obese rats.
Methods: pGL3-Enhancer-PPARγ2 (625 bp)-Luc plasmid, a luciferase reporter gene expression plasmid containing PPARγ2 promoter, was stably transfected into 3T3-L1 preadipocytes. PPARγ2 promoter activities were determined by assaying the luciferase activities. Then PPARγ2 promoter activities in preadipocytes and PPARγ2 mRNA levels in human subcutaneous adipocytes were measured after the administration with LLAE. Additionally, the effects of LLAE on body weight, fat mass, glucose and lipid metabolism and the expression of PPARγ2, insulin receptor substrate 1 and glucose transporter 4 (GLUT4) in visceral adipose tissue (VAT) were measured in HFD-induced obese rats treated with low or high dose [0.5 or 3.0 g crude drug/(kg.d)] LLAE for 6 weeks.
Results: Ten μg/ml LLAE significantly increased the luciferase activities in 3T3-L1 cells and the stimulatory action reached 2.51 folds of controls when LLAE was 1000 μg/ml (P < 0.01). After treating 3T3-L1 cells with 100 μg/ml LLAE, the stimulatory role peaked at 32 h where it was 2.58 folds of controls (P < 0.01). Besides, 100 μg/ml LLAE significantly increased PPARγ2 mRNA levels in human adipocytes to 1.91 folds of controls (P < 0.01). In HFD-induced obese rats, administration with both low and high dose LLAE notably reduced visceral fat mass by 45.5 and 58.4%, respectively, and significantly decreased fasting serum insulin levels (P < 0.05). The high dose LLAE also significantly decreased homeostasis model assessment of insulin resistance in obese rats (P < 0.05). Furthermore, the mRNA levels of PPARγ2 and GLUT4 in VAT of obese rats were significantly increased when compared with control rats, and were notably suppressed by LLAE intervention for 6 weeks (P < 0.05).
Conclusion: LLAE significantly reduces visceral fat mass and ameliorates insulin resistance in HFD-induced obese rats. These beneficial effects of LLAE may associate with its role in stimulating PPARγ2 expression in preadipocytes and subcutaneous adipocytes and suppressing PPARγ2 and GLUT4 expression in VAT.
Obesity is a chronic metabolic disease caused by many factors, which derives from an imbalance between energy intake and expenditure and finally leads to excessive and ectopic fat accumulation in our body. It is becoming a major risk factor for many diseases such as type 2 diabetes, cardiovascular diseases, and several kinds of tumors (Djalalinia et al., 2015). The study of obesity has attracted the attention of scholars all over the world. However, drugs for obesity treatment are limited and have many adverse reactions (Jones and Bloom, 2015).
Some TCMs have been reported to be useful and safe in the treatment of obesity by acting on multiple targets (Yin et al., 2008; Hasani-Ranjbar, 2009; Gu and Huang, 2013). Lotus leaf, also known as Nelumbo nucifera leaf and is rich in a number of alkaloids such as nuciferine, pronuciferine, roemerine and so on, is such kind of TCMs with many beneficial roles in organism (Mukherjee et al., 2009; Sharma et al., 2017), including anti-obesity (Ono et al., 2006; Ahn et al., 2013), antioxidant (Lin et al., 2009; Zhu et al., 2015), hypoglycemic and hypolipidemic effects (Kim et al., 2013; Zeng et al., 2016), and lipolytic activity (Siegner et al., 2010). The anti-obesity effect of lotus leaf was reported to be associated with inhibiting the activities of pancreatic alpha-amylase and lipase in vitro (Ono et al., 2006; Ahn et al., 2013), accelerating lipid metabolism and up-regulating energy expenditure in HFD-induced obese mice and rats (Ono et al., 2006) and suppressing adipocyte differentiation in 3T3-L1 preadipocytes (Ahn et al., 2013). However, the detailed mechanism by which lotus leaf loses weight is largely still unknown.
Peroxisome proliferative activated receptors, which belong to the thyroid/retinoid nuclear receptor family, have been reported to control the expression of genes involved in adipogenesis in preadipocytes differentiation process (Mangelsdorf et al., 1995; Wang et al., 2014). PPARγ, especially the PPARγ2 isoform, is predominately expressed in adipose tissue and plays a significant role in adipogenesis, insulin sensitization and homeostasis of lipid and glucose (Willson et al., 2001; Feng et al., 2016). Adipose-specific activation of PPARγ is sufficient to reverse whole body insulin resistance to a similar degree as systemic TZD treatment (Sugii et al., 2009). TZD, a PPARγ agonist, has been reported to decrease the blood glucose levels and ameliorate insulin resistance in diabetic animal model and patients and has been widely used in clinical practice (Saraf et al., 2012; Janani and Ranjitha Kumari, 2015). Interestingly, Guo et al. (2013) reported that nuciferine, a chemical composition of lotus leaf, could also prevent hepatic steatosis and injury induced by a HFD in hamsters through suppressing the expression of PPARγ in liver. However, whether lotus leaf could affect the expression of PPARγ in adipocytes and adipose tissue has not been elucidated yet.
Therefore, in the present study, we aimed to explore the effects of LLAE on PPARγ2 expression in 3T3-L1 preadipocytes and human subcutaneous adipocytes differentiated from preadipocytes, and further investigate its effects on body weight, fat mass, insulin resistance and PPARγ2 expression in VAT of HFD-induced obese rats.
Lotus leaf (100 g, purchased from Beijing Tongrentang pharmaceuticals company, Beijing, China) was soaked in 300 ml cold water for 2 h, then was boiled for 30 min and filtered with eight layers of gauzes. Then 200 ml water was added to the residue and the mixture was boiled and filtered again as above. The two filtrates were mixed thoroughly and centrifuged at 4000 rpm for 20 min. The supernatant was taken and freeze-dried. The powder obtained was LLAE. 4 g LLAE was dissolved in 4 ml deionized water to make 1000 mg/ml LLAE stock solution.
pGL3-Enhancer-PPARγ2 (625 bp)-Luc plasmid was constructed as described previously (Zhu et al., 2013). In brief, pGL3-PPARγ2 (625 bp)-Luc, which contained the human PPARγ2 gene 5′-promoter fragment spanning -615 to +10 bp (+1 indicates the transcription start site) and was constructed previously in our laboratory, was digested by the restriction enzymes KpnI and BglII (Takara, Japan) to get the 625 bp PPARγ2 gene promoter inserting fragment. The fragment was then inserted into pGL3-Enhancer-Luc vector to yield pGL3-Enhancer-PPARγ2 (625 bp)-Luc plasmid. An electrophoresis was conducted to verify the constructed plasmid, and the plasmid was further sequenced by Bioasia Biotechnology Company (Shanghai, China).
Stably transfected 3T3-L1 preadipocytes were created as described previously (Gong et al., 2006). In brief, 3T3-L1 preadipocytes (obtained as a generous gift from Department of genetics, Institute of life sciences, Peking University, Beijing, China) were cultured with Dulbecco’s modified Eagle’s-F12 (DMEM/F12, Hyclone, Logan, UT, United States) medium supplemented with 10% FBS, then were plated in 12-well plates at a density of 4.5 × 105 cells/well. The cells were co-transfected with 1.2 μg pGL3-Enhancer-PPARγ2 (625 bp)-Luc and 0.4 μg pcDNA3.1(+), which carried the neomycin resistance gene, and 4 μl lipofectamine2000 (Invitrogen, Carlsbad, CA, United States) at a ratio of weight/volume 1/2.5 in 0.4 ml OPTI-MEM medium for 5 h. Then cells were incubated with medium containing 300 μg/ml Geneticin G418 (Invitrogen, Carlsbad, CA, United States) to screen the monoclonal cells. The luciferase activity was measured using commercially available Luciferase Assay System kit (Promega, Madison, WI, United States) in microplate luminescence analyzer (Beijing Hamamatsu Photon Techniques, inc., Beijing, China) as described previously (Zhu et al., 2014). The relative luciferase activity of stably transfected 3T3-L1 preadipocytes, which were transfected with pGL3-Enhancer-PPARγ2 (625 bp)-Luc plasmid, was obtained in comparison with that of cells transfected with blank plasmid. The three appropriate colonies which expressed luciferase in a middle degree were chosen for the following studies.
Stably transfected 3T3-L1 preadipocytes were seeded in 12-well plates at a density of 2.5 × 105 cells/well. Then cells were treated with different concentrations (1 ∼ 1000 μg/ml) of LLAE for 18 h or were treated with 100 μg/ml LLAE for different time (2 ∼ 36 h). Then cells were lysed and the luciferase activities were measured. The relative luciferase activity of LLAE treated cells was obtained in comparison with that of controls. The experiments were performed in three different colonies. Samples in each experiment were in triplicate.
Human primary preadipocytes [obtained from the abdominal subcutaneous adipose tissues (SATs) of three patients with benign diseases (appendicitis, cholelithiasis, and colon benign neoplasm) and preserved in our laboratory] were differentiated as previously described (Zhu et al., 2006). Briefly, cells seeded in six-well plates at a density of 5 × 104 cells/well were grown to 90% confluence, and then were induced to differentiate in serum-free DMEM/F12 medium supplemented with 0.5 μM insulin (Sigma–Aldrich, United States), 0.25 μM dexamethasone (Sigma–Aldrich, United States), 0.2 nM thyroxine (Sigma–Aldrich, United States) and 0.5 nM 3-isobutyl-1-methylxanthine (Sigma–Aldrich, United States) for 6 days. The fully differentiated cells were then administrated with 100 μg/ml LLAE for 36 h. The total RNA was extracted using an EZNA total RNA kit (Omega Bio-Tek, Doraville, GA, United States), and 0.5 μg RNA was reverse transcribed with a SuperScript First-Strand Synthesis System Kit (Invitrogen, Carlsbad, CA, United States). 2 μl RT products (cDNA) were amplified with PPARγ2 primers or β-actin primers as shown in Supplementary Table S1. Amplification was carried out as follows: 10 min at 95°C, 35×(1 min at 94°C, 1 min at 59°C, and 1 min at 70°C), and 10 min at 72°C. The predicted products of PPARγ2 and β-actin were 223 and 540 bp, respectively. 10 μl RT-PCR products were electrophoresed in 2% agarose gel in Tris-acetate-EDTA buffer. The gel was then stained with ethidium bromide and photographed using Alphalmager M2200 (AlphaInnotech, San Leandro, CA, United States). The density ratio of PPARγ2 to β-actin PCR products was calculated to evaluate the PPARγ2 expression changes. The experiments were repeated for three times. Samples in each experiment were in triplicate. The study was approved by the ethics committee of Peking Union Medical College Hospital (No. JS-1093). All patients signed informed consent before being taken the adipose tissue during the surgery.
Four-week-old male Sprague Dawley (SD) rats (weight 45 ± 1.5 g) were purchased from Beijing Vital River Laboratory Animal Technology Company (Beijing, China). All rats were housed in standard cages as in previous animal experiments (Zhu et al., 2016). Rats were randomly divided into SF group (n = 8) and HFD group (n = 22) which was further divided into simple HFD group (n = 7), low dose LLAE intervention group (LLLAE, n = 8) and high dose LLAE intervention group (HLLAE, n = 7). The composition of the experimental diet (Beijing HFK Bioscience, Co. Ltd, Beijing, China) was shown in Supplementary Table S2. Rats in LLLAE and HLLAE groups were administrated with 0.5 g crude drug/(kg.d) and 3.0 g crude drug/(kg.d) LLAE, respectively, by gavages. Rats in SF and HFD groups were given gavages with the same volume of saline. All rats were weighted every 5 ∼ 6 days and the dose of LLAE was changed according to body weight change. Food intake was calculated by subtracting the amount of residue food from the amount of supply food. After 6 weeks of LLAE intervention, rats were anesthetized with 10% urethane (1.0 g/kg) after a 12-h fasting. Blood samples were drawn from abdominal aorta and centrifuged immediately at 3000 rpm for 10 min at 4°C. VAT including epididymal white adipose tissue (eWAT) and perirenal white adipose tissue (pWAT) was dissected and weighted, then was immediately frozen in liquid nitrogen and stored at -80°C. Serum fasting blood glucose (FBG), total cholesterol (TC), triglycerides (TG), low density lipoprotein-cholesterol (LDL-c), and high density lipoprotein-cholesterol (HDL-c) were measured by routine automated laboratory methods. Serum insulin levels were measured by insulin radioimmunoassay kit (Linco, United States). HOMA-IR was calculated according to the following formula: fasting serum insulin (pmol/L) × FBG (mmol/L)/135 (Nordstrand et al., 2013). RT-qPCR analysis was performed in VAT using SYBR Premix Ex Taq (Takara, Japan) as previously described (Zhu et al., 2016). In brief, total RNA was extracted from adipose tissue using RNeasy Lipid Tissue Mini Kit (Qiagen, Germany) according to the supplier’s instructions. Then 1 μg of total RNA was used to reverse transcribe to cDNA using the PrimeScriptTM RT reagent Kit with gDNA Eraser (Takara, Japan). RT-qPCR was conducted to assess the expression of PPARγ2, IRS1, and GLUT4 using an ABI7500 PCR system (Applied Biosystems, San Francisco, CA, United States). β-actin was used for normalization and the relative expression for each target gene was calculated using the formula 2-ΔΔCt. The primers used to amplify target genes and β actin were listed in Supplementary Table S1. All animal experimental protocols were carried out according to the standards of the Guide for the Care and Use of Laboratory Animals and approved by the ethics committee of Peking Union Medical College Hospital (No. XHDW-2016-001).
All values were expressed as mean ± standard deviation (SD) or median (interquartile range). Statistical analysis was performed by t-test for two groups or one-way ANOVA followed by Dunnett t (two-sided) or Dunnett t3 post-test for three or more groups. Skewed data were ln-transformed and Kruskal–Wallis test was used if data were still not normally distributed. Differences between groups were considered as statistically significant at P < 0.05. All statistical computations were run on SPSS 22.0 for Windows (SPSS, Inc., Chicago, IL, United States).
The constructed pGL3-Enhancer-PPARγ2 (625 bp)-Luc plasmid were digested by KpnI and BglII. The fragments produced were electrophoresed. As shown in Figure 1, the digested products of the predicted lengths appeared (5064- and 625-bp). The plasmid was also verified by DNA sequencing, finally demonstrating that construction of pGL3-Enhancer-PPARγ2 (625 bp)-Luc plasmid which contained the human PPARγ2 gene 5′-promoter fragment spanning -615 to +10 bp was successful.
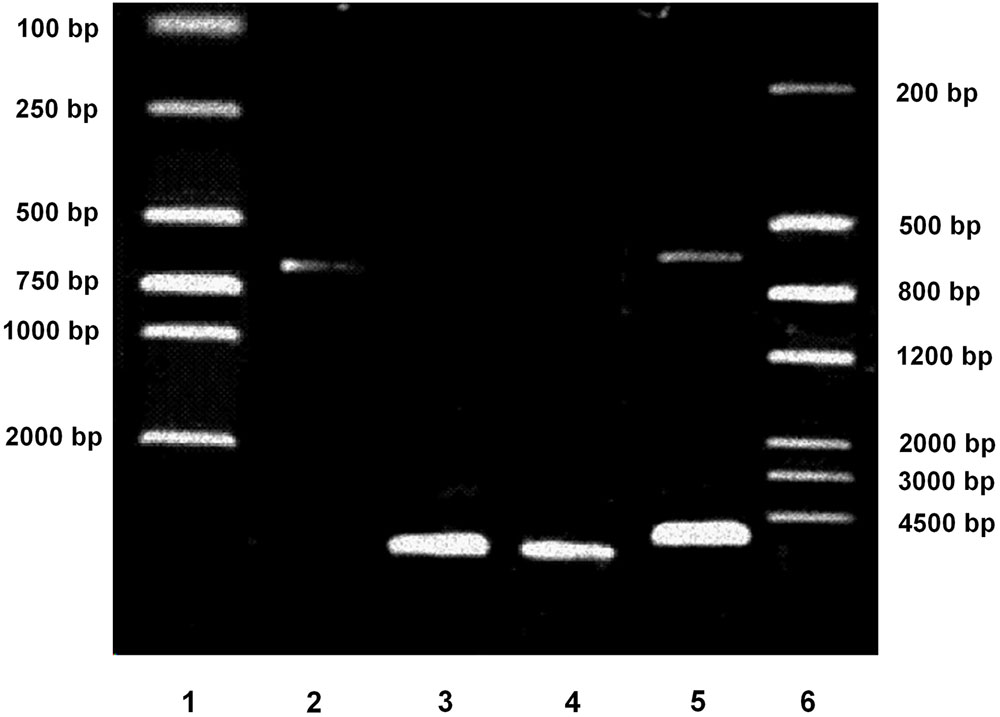
FIGURE 1. Verification of pGL3-Enhancer-PPARγ2 (625 bp)-Luc plasmid by electrophoresis. Lane 1: DNA marker DL2000. Lane 2: Inserting fragment from pGL3-PPARγ2 (625 bp)-Luc digested by KpnI and BglII (625 bp). Lane 3: Vector fragment from pGL3-Enhancer-Luc digested by KpnI and BglII (5064 bp). Lane 4: The constructed pGL3-Enhancer-PPARγ2 (625 bp)-Luc plasmid (5689 bp). Lane 5: The constructed pGL3-Enhancer-PPARγ2 (625 bp)-Luc digested by KpnI and BglII (5064/625 bp). Lane 6: DNA marker III.
The stably transfected 3T3-L1 preadipocytes were treated with 1 ∼ 1000 μg/ml LLAE for 18 h to explore the effects of LLAE on luciferase activities. As shown in Figure 2, 10 μg/ml LLAE significantly increased the luciferase activities in 3T3-L1 preadipocytes. The increase was further augmented with an increase of LLAE concentration. The maximal stimulatory action was noted to be 2.51 folds above controls after treating 3T3-L1 preadipocytes with 1000 μg/ml LLAE (P < 0.01). These findings suggest that LLAE could stimulate PPARγ2 promoter activities in a dose-dependent manner.
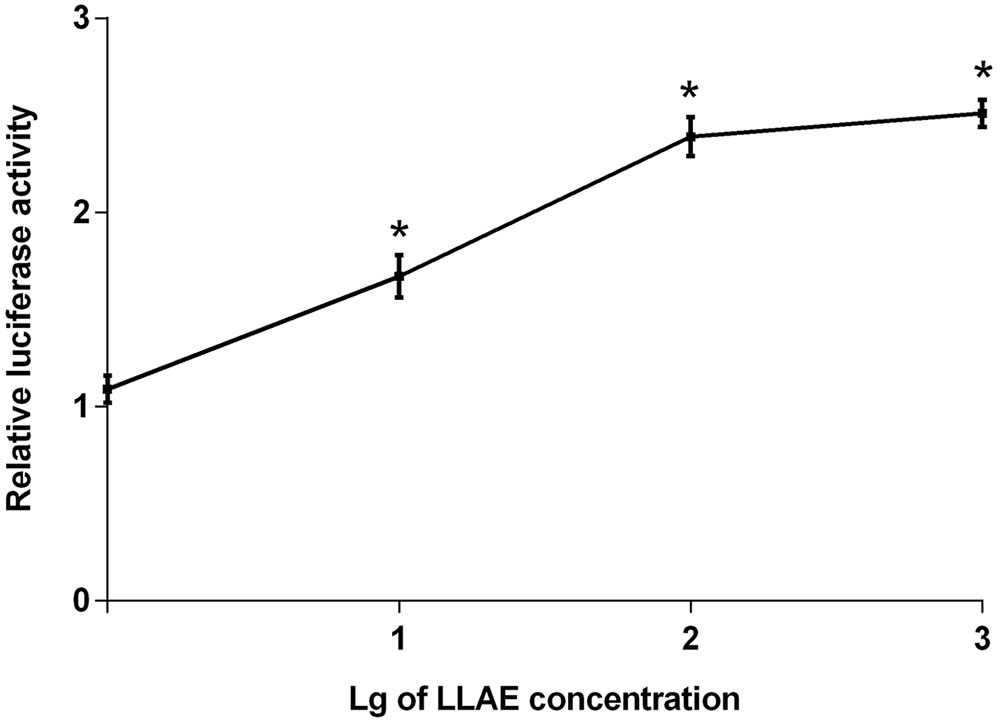
FIGURE 2. Effects of different concentrations of LLAE on PPARγ2 promoter activities in 3T3-L1 preadipocytes. Stably transfected 3T3-L1 preadipocytes were plated in 12-well plates at a density of 2.5 × 105 cells/well and cultured in medium with different concentrations of LLAE (1 ∼ 1000 μg/ml) for 18 h. Then cells were lysed and the luciferase activities were measured. The relative luciferase activity was obtained in comparison with that of controls. The data represent the mean ± SD of three separate wells in three different colonies. ∗P < 0.01 vs. controls (without LLAE treatment).
In order to investigate the effects of different action time of LLAE on luciferase activities, the stably transfected 3T3-L1 preadipocytes were treated with 100 μg/ml LLAE and luciferase activities were detected every 2 h from 0 to 36 h. As presented in Figure 3, the relative luciferase activities in 3T3-L1 preadipocytes started to increase at 6 h and increased gradually with the prolongation of action time. The maximum effect was observed at 32 h after LLAE stimulation where it was 2.58 folds of control cells. It remained up to 36 h where it was still 2.35 folds of controls (P < 0.01). These data indicate that LLAE could stimulate PPARγ2 promoter activities in a time-dependent manner.
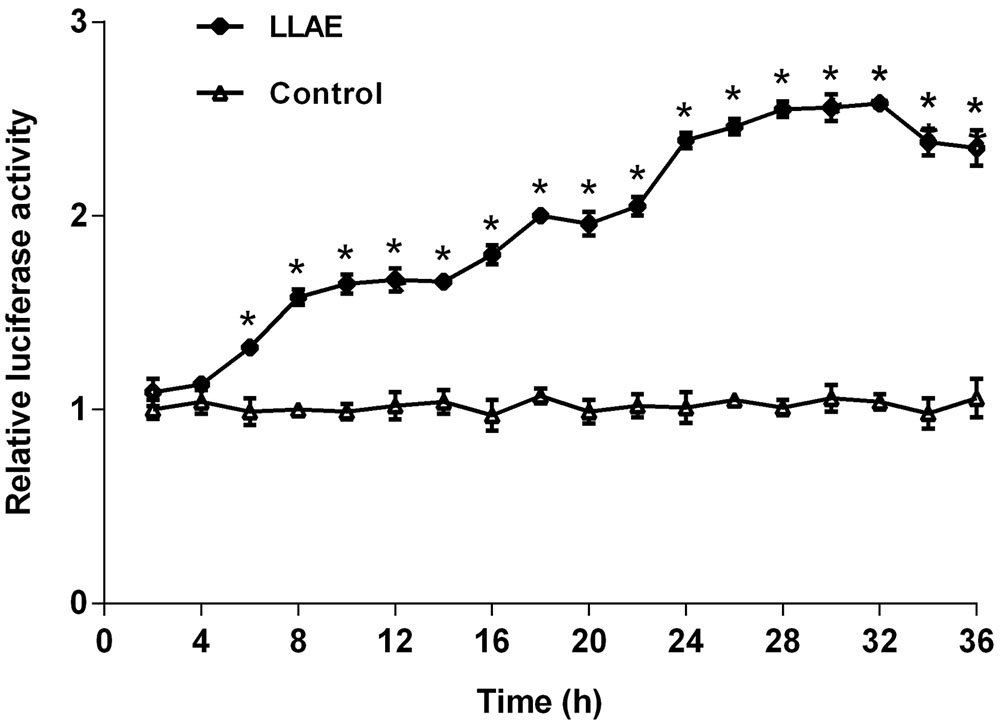
FIGURE 3. Effects of different action time of LLAE on PPARγ2 promoter activities in 3T3-L1 preadipocytes. Stably transfected 3T3-L1 preadipocytes were plated in 12-well plates at a density of 2.5 × 105 cells/well and cultured in medium with 100 μg/ml LLAE for different time (2 ∼ 36 h). Then cells were lysed and the luciferase activities were measured. The relative luciferase activity was obtained in comparison with that of controls. The data represent the mean ± SD of three separate wells in three different colonies. ∗P < 0.01 vs. controls (without LLAE treatment).
Since LLAE could stimulate PPARγ2 promoter activities in 3T3-L1 preadipocytes as evidenced in the above experiments, whether or not it could promote the PPARγ2 expression in adipocytes? In order to answer this question, human preadipocytes preserved in our laboratory were differentiated to adipocytes according to the standard differentiation protocols as shown in Section “Materials and Methods.” On the sixth day after differentiation, cells were treated with 100 μg/ml LLAE for 36 h and then PPARγ2 mRNA levels were observed by RT-PCR. The changes of PPARγ2 mRNA levels were calculated by the density ratio of PPARγ2 to β-actin RT-PCR products in electrophoresis image. As presented in Figure 4A, the band density of PPARγ2 RT-PCR products was higher in LLAE treated cells than that of control cells, and it was 1.91 folds of control cells by the semi-quantitative analysis as shown in Figure 4B (P < 0.01). This result suggests that LLAE also significantly increase PPARγ2 mRNA levels in addition to its stimulatory role in PPARγ2 promoter activity.
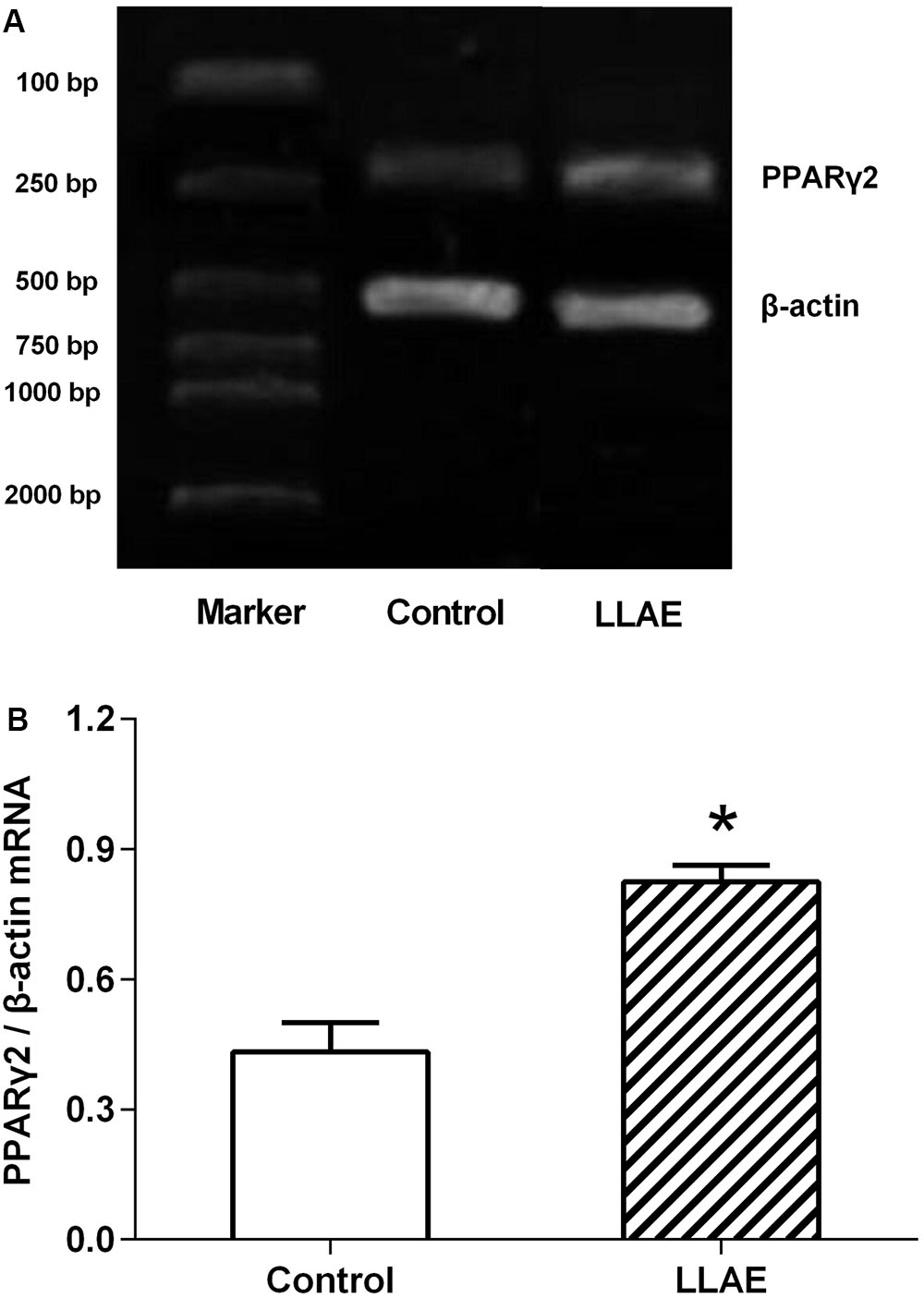
FIGURE 4. Effects of LLAE on PPARγ2 expression in human adipocytes. Human primary preadipocytes were seeded in six-well plates at a density of 5 × 104 cells/well. The cells were then differentiated as described in Section “Materials and Methods.” On the sixth day of differentiation, the cells were treated with 100 μg/ml LLAE for 36 h. The total RNA was extracted and RT-PCR was performed as described in Section “Materials and Methods.” (A) The RT-PCR products were electrophoresed in 2% agarose gel with ethidium bromide and photographed under ultraviolet light. (B) PPARγ2 expression in human adipocytes treated with LLAE was calculated by the density ratio of PPARγ2 to β-actin PCR products. The data represent the mean ± SD of three separate wells in three independent experiments. ∗P < 0.01 vs. controls (without LLAE treatment).
In order to further explore the effects of LLAE in vivo, an animal experiment on HFD-induced obese rats was conducted subsequently. As presented in Figure 5, the visceral fat mass including epididymal and perirenal fat, and the visceral fat percentage (the percentage of body weight occupied by the visceral fat mass) of rats fed with HFD for six weeks were significantly increased to 1.98 and 2.07 folds of rats fed with SF (10.1 ± 2.4 g vs. 5.1 ± 1.3 g; 3.1 ± 0.9% vs. 1.5 ± 0.4%, all P < 0.05) although there were no notable changes of body weight observed in these two group rats. Interestingly, after administration these HFD-induced obese rats with low dose (LLLAE group) or high dose (HLLAE group) LLAE for 6 weeks, the visceral fat mass were significantly decreased by 45.5% (5.5 ± 1.0 g vs. 10.1 ± 2.4 g, P < 0.05) and 58.4% (4.2 ± 1.0 g vs. 10.1 ± 2.4 g, P < 0.05), respectively, as shown in Figure 5B Moreover, visceral fat percentage of rats in HLLAE group was also significantly decreased by 51.6% in comparison with that of rats in HFD group (1.5 ± 0.2% vs. 3.1 ± 0.9%, P < 0.05). In addition, LLAE intervention did not affect food intake of HFD-induced obese rats although there was a decrease in food intake of HFD fed rats when compared with SF fed rats as presented in Figure 5A (P < 0.05). These data show that LLAE significantly reduced visceral fat mass and visceral fat percentage in HFD-induced obese rats.
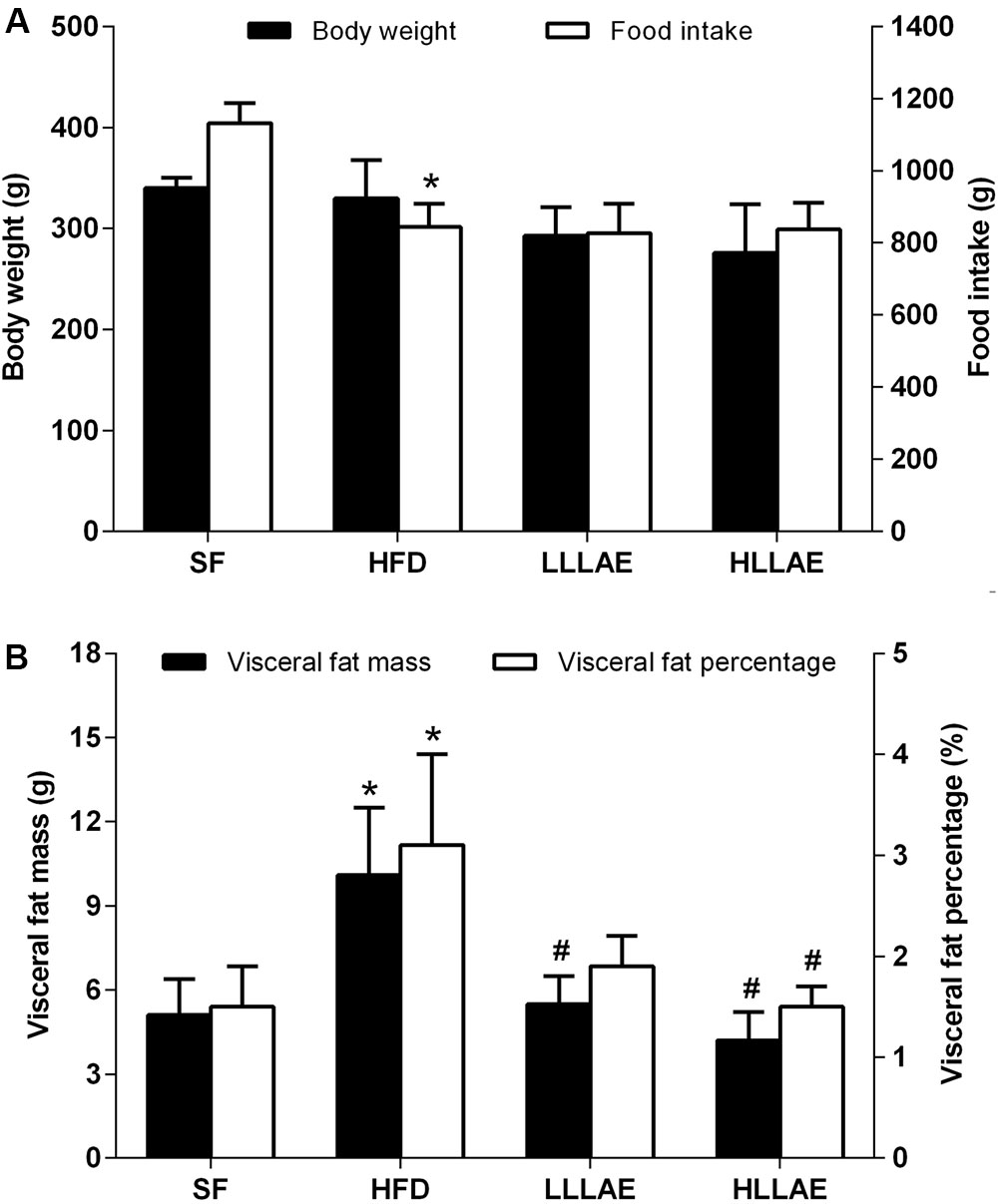
FIGURE 5. Effects of LLAE on body weight, food intake (A) and visceral fat mass (B) in HFD-induced obese rats. HFD-induced obese rats were administrated with low dose LLAE [0.5 g crude drug/(kg.d), LLLAE] or high dose LLAE [3.0 g crude drug/(kg.d), HLLAE] for 6 weeks by gavages. Body weight and food intake were recorded, and visceral fat (epididymal and perirenal fat) mass was weighted as described in Section “Materials and Methods.” The visceral fat percentage was calculated by the percentage of body weight occupied by the visceral fat mass. The data represent the mean ± SD. ∗P < 0.05 vs. SF group; #P < 0.05 vs. HFD group.
As displayed in Table 1, serum TC and LDL-c levels in HFD group rats were higher than that in SF group (P < 0.05), which were 1.95 and 4.47 folds of that in SF group, respectively. Compared with HFD group, rats in both LLLAE and HLLAE groups manifested lower fasting insulin levels (P < 0.05). Meanwhile, rats in HLLAE group also displayed significant decrease in HOMA-IR [3.8 (1.6, 13.8) vs. 21.1 (6.4, 35.8), P < 0.05] when compared with that of HFD group. However, there was no difference in FBG and serum lipid levels in LLAE intervention rats. These data indicate that LLAE significantly ameliorated insulin resistance in HFD-induced obese rats.
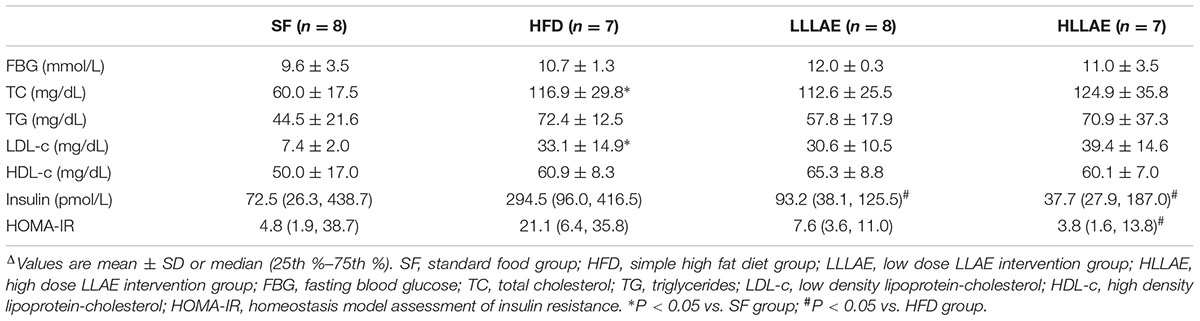
TABLE 1. Effects of LLAE on biochemical parameters, insulin and HOMA-IR in HFD-induced obese ratsΔ.
As presented in Figure 6A, the mRNA levels of PPARγ2 and GLUT4 in pWAT of HFD-induced obese rats were notably increased to 1.64 and 2.93 folds of that in SF group (P < 0.05). After the administration of these obese rats with low and high dose LLAE, GLUT4 mRNA levels were significantly decreased by 59.0 and 75.4%, respectively (P < 0.05). PPARγ2 mRNA levels were significantly decreased by 38.4% after administration with HLLAE (P < 0.05). However, there was no significant difference in IRS1 mRNA levels of pWAT between these groups. As referring to eWAT, the mRNA levels of PPARγ2, IRS1, and GLUT4 in HFD group were significantly increased to 1.53, 1.75, and 2.24 folds of that in SF group (P < 0.05), and GLUT4 mRNA levels were also significantly decreased by 41.1 and 78.1% after LLLAE and HLLAE intervention, respectively (P < 0.05) as presented in Figure 6B. But the mRNA levels of PPARγ2 and IRS1 were not significantly changed in eWAT after LLAE intervention.
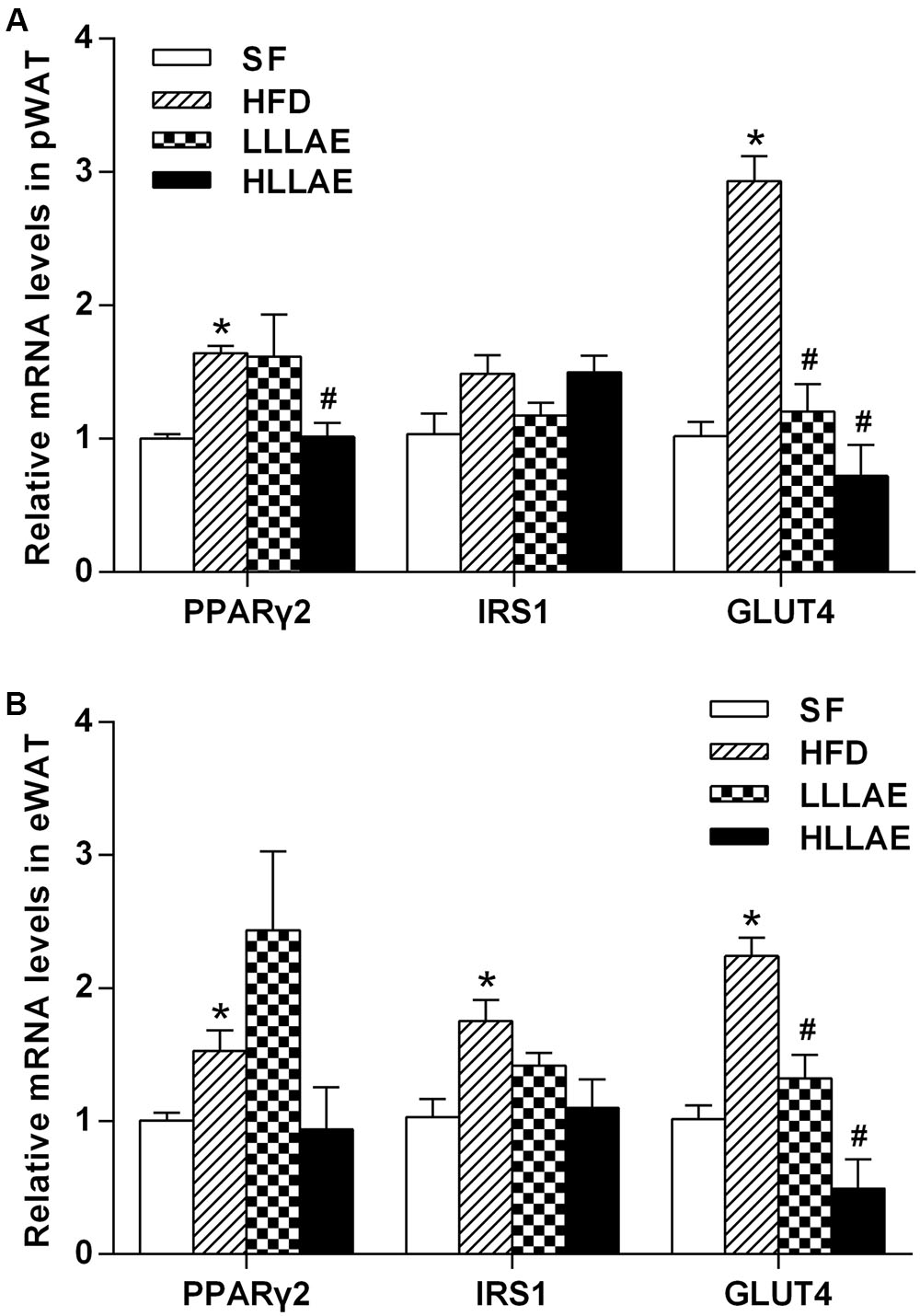
FIGURE 6. Effects of LLAE on the expression of PPARγ2, IRS1, and GLUT4 in pWAT (A) and eWAT (B) of HFD-induced obese rats. HFD-induced obese rats were administrated with low dose LLAE [0.5 g crude drug/(kg.d), LLLAE] or high dose LLAE [3.0 g crude drug/(kg.d), HLLAE] for 6 weeks by gavages. Then perirenal white adipose tissue (pWAT) and epididymal white adipose tissue (eWAT) were taken. The total RNA of adipose tissue was extracted and then RT-qPCR was performed as described in Section “Materials and Methods.” The data represent the mean ± SD. ∗P < 0.05 vs. SF group; #P < 0.05 vs. HFD group.
Lotus leaf has been reported to have some biological roles such as antioxidant, promoting lipolytic activity and up-regulating energy expenditure. However, the detailed mechanism by which lotus leaf functions is largely still unknown. Our current study for the first time demonstrated that LLAE could stimulate PPARγ2 promoter activities in 3T3-L1 preadipocytes and increase PPARγ2 gene expression in human subcutaneous adipocytes differentiated from preadipocytes. LLAE also significantly reduced visceral fat mass, decreased fasting serum insulin levels and ameliorated insulin resistance in HFD-induced obese rats, indicating the anti-obesity and anti-diabetic activity of LLAE. In accordance with our results, Ono et al. (2006) and Du et al. (2010) found that lotus leaf hot water or alcoholic extract both significantly lowered the body weight and the mass of epididymal, retroperitoneal, and parametrial adipose tissues in HFD-induced obese SD rats and mice. Studies performed by Guo et al. (2013) also demonstrated that nuciferine, a major active aporphine alkaloid from lotus leaf, significantly reduced body weight and the mass of epididymal and perirenal adipose tissues in hamsters fed with HFD. The similar anti-diabetic activity of lotus leaf was also reported by other researchers. Zeng et al. (2016) found that the treatment of rats with gestational diabetes mellitus with 50 and 100 mg/kg lotus leaf selenium-polysaccharide, a water extract of lotus leaf, for 2 weeks induced a significant decrease in FBG and insulin levels. Huang et al. (2011) reported that lotus leaf methanolic extract (100 mg/kg) also significantly decreased blood glucose levels of normal mice and HFD-induced diabetic mice. Studies conducted by Kim et al. (2013) demonstrated that oral administration of lotus leaf ethanol extract (500 mg/kg) significantly decreased the area under the glucose response curve in streptozotocin (STZ)-induced diabetic rats and 1% lotus leaf ethanol extract intervention for 7 weeks significantly reduced the plasma glucose level, HOMA-IR and HbA1C level in db/db mice. All these findings, together with our results, suggest that lotus leaf plays the anti-obesity and anti-diabetic roles by decreasing visceral fat and ameliorating insulin resistance.
It is well-known that PPARγ, a member of the nuclear-receptor superfamily, is considered to be the master regulator in promoting adipocyte differentiation and increasing insulin sensitization. Activation of PPARγ has been shown to promote differentiation of preadipocytes, thereby increasing the number of smaller and more insulin sensitive adipocytes (Okuno et al., 1998). Activation of PPARγ by TZD leads to increase the storage capacity of fatty acids in the adipocytes and thereby to decrease the amount of circulating fatty acids and ameliorate the insulin resistance (Siersbaek et al., 2010), which finally leads to the reduced plasma insulin and glucose levels (Olefsky, 2000; Leonardini et al., 2009; Sugii et al., 2009). Studies performed at the cellular level showed that PPARγ activation directly modulated the expression or phosphorylation of specific molecules of insulin signaling cascade to ameliorate insulin resistance (Leonardini et al., 2009). In the present study, LLAE was firstly found to significantly stimulate PPARγ2 promoter activities of 3T3-L1 preadipocytes in the dose and time-dependent manner, and LLAE also directly increased PPARγ2 mRNA levels in human subcutaneous adipocytes differentiated from the preadipocytes. These results indicate that LLAE might play an important role in ameliorating insulin resistance by stimulating PPARγ2 expression in preadipocytes and subcutaneous adipocytes.
The adipocytes from VAT are considered to be metabolically active, more insulin-resistant and sensitive to lipolysis than that from SAT (Ibrahim, 2010). The visceral fat was strongly related to many unhealthy conditions, including cardiovascular disease, insulin resistance, and type 2 diabetes (Carr et al., 2004; Finelli et al., 2013). It has been reported that PPARγ activation could greatly decrease the VAT mass (Mori et al., 1999; Miyazaki et al., 2002). After treated with troglitazone (a kind of TZD) for 6 months, type 2 diabetic patients showed a significant decrease in visceral fat area and a notable improvement in glucose metabolism (Mori et al., 1999). The similar results were also obtained by Miyazaki et al. (2002) who demonstrated that after intervention type 2 diabetic patients with pioglitazone (another kind of TZD) for 4 months, they had decreased visceral fat area and improved hepatic and peripheral insulin sensitivity. Studies performed by Gabriely et al. (2002) and Hirashita et al. (2012) showed that surgical removal of epididymal fat or together with the perinephric fat pad of obese rats led to the decrease of plasma insulin levels and the improvement of insulin sensitivity. All these results indicate that VAT in relative to SAT was bad adipose tissue and the decrease of VAT mass by internal medicine or surgical method could significantly ameliorate insulin resistance. In our present study, visceral fat mass, fasting serum insulin level and HOMA-IR of HFD-induced obese rats were also significantly reduced after LLAE intervention for 6 weeks, suggesting that LLAE could ameliorate insulin resistance by reducing VAT mass of HFD-induced obese rats.
Glucose transporter 4 is one of the sugar transporter proteins that catalyze hexose transport across cell membranes (Huang and Czech, 2007). It is predominantly expressed in muscle cells and adipocytes. The GLUT4 glucose transporter is a major mediator of glucose removal from the circulation to adipocytes. About 50% of the glucose taken up by the adipocytes is normally used to synthesize the glycerol and fatty acid components of triacylglycerol (Festuccia et al., 2009). Furthermore, PPARγ is capable of activating GLUT4 gene expression during adipogenesis (Wu et al., 1998). It was reported that higher PPARγ2 mRNA levels of VAT were accompanied by higher GLUT4 mRNA levels as well as higher capacity to store triacylglycerol and larger adipocyte size in Wistar rats (Palou et al., 2009). These results indicated that the activation of PPARγ2 in VAT could increase GLUT4 expression to transport more glucose into visceral adipocytes thus increasing their fat accumulation. In the current study, HFD-induced obese rats showed increased mRNA levels of PPARγ2 and GLUT4 in VAT as well as an increase of visceral fat mass compared with rats fed with SF, indicating that there was more glucose uptake and lipogenesis in visceral adipocytes in obese rats. In accordance with our results, MacLaren et al. (2007) found that GLUT4 also had a higher expression in omental adipose tissue from insulin-resistant obese subjects when compared with insulin-sensitive subjects, suggesting the increased expression levels of GLUT4 might also indicate a compensatory mechanism under the situation of insulin resistance. Interestingly, the increased expression of PPARγ2 and GLUT4 in VAT of these obese rats was significantly suppressed by LLAE intervention in our present study. These results demonstrate that LLAE could suppress PPARγ2 expression, then decrease GLUT4 expression and inhibit glucose uptake, further reduce substrate supply for triacylglycerol synthesis in VAT adipocytes and finally decrease the visceral fat mass in obese rats. Similarly, studies performed by Lee et al. (2015) also founded that kaempferol, a compound isolated from the methanolic extract of Nelumbo nucifera stamens, significantly suppressed GLUT4 expression and inhibited glucose uptake, further decreased triacylglycerol accumulation in adipocytes. Besides, nuciferine and pronuciferine, the main active components of lotus leaf, were also reported to notably decrease lipid droplets and intracellular triglyceride contents by activating the AMPK signaling pathway in mature 3T3-L1 adipocytes (Ma et al., 2015).
Additionally, we found serum TC and LDL-c levels of rats fed with HFD were higher than those fed with SF. However, the reduction of TC and LDL-c levels was not observed after LLAE administration. On the contrary, studies performed by Du et al. (2010) and Guo et al. (2013) found that lotus leaf hot water extract or its major active aporphine alkaloid nuciferine significantly reduced the serum TG, TC and LDL-c levels in HFD fed SD rats or hamsters. 1% lotus leaf ethanol extract or 50 or 100 mg/kg lotus leaf selenium-polysaccharide also significantly elevated HDL-c levels in db/db mice and gestational diabetes mellitus rats in addition to its role in reducing TG, TC, and LDL-c levels (Kim et al., 2013; Zeng et al., 2016). The different results between ours and literatures may be explained by the different concentrations and action time of lotus leaf or different origins and extraction methods of lotus leaf. Different animal models used in the experiment may also be one reason for these differences.
This is the preliminary study to firstly demonstrate that LLAE could directly stimulate PPARγ2 promoter activities in preadipocytes and increase PPARγ2 gene expression in subcutaneous adipocytes. Meanwhile, LLAE significantly ameliorated insulin resistance of HFD-induced obese rats, and its beneficial role was obtained possibly by reducing visceral fat mass through suppressing the expression of PPARγ2 and GLUT4 in VAT. Further studies need to be done for elucidating the effects of LLAE on the proliferation and differentiation of preadipocytes, especially preadipocytes from VAT, and the effects of LLAE on SAT mass of HFD-induced obese rats. Overall, this is a promising beginning which demonstrates that PPARγ2 is an important factor involved in the mechanism by which LLAE reduces visceral fat mass and ameliorates insulin resistance in obese rats.
KY did the cell experiments, analyzed data and wrote the primary manuscript. HZ designed the experiments and revised the primary manuscript. JX did the cell experiments and animal experiments. HP supervised the experiments. NL, LW, and HY helped to treat the animals and supervised the biochemical parameters measurements. ML analyzed data. FG designed the experiment, supervised the whole experiments and revised the primary manuscript.
The study was supported by grants from the National Natural Science Foundation of China (No. 30600836, 81471024 for HZ, No. 30540036, 30771026, 81370898 for FG), the Beijing Natural Science Foundation (No. 7082079 for FG), the National Key Program of Clinical Science (WBYZ2011-873 for FG and HZ) and PUMCH Foundation (2013-020 for FG).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer MH and handling Editor declared their shared affiliation, and the handling Editor states that the process nevertheless met the standards of a fair and objective review.
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fphar.2017.00409/full#supplementary-material
FBS, fetal bovine serum; GLUT4, glucose transporter 4; HFD, high fat diet; HLLAE, high dose LLAE; HOMA-IR, homeostasis model assessment of insulin resistance; IRS1, insulin receptor substrate 1; LLAE, lotus leaf aqueous extract; LLLAE, low dose LLAE; PPARs, peroxisome proliferative activated receptors; SF, standard food; TCMs, traditional Chinese medicines; TZD, thiazolidinedione; VAT, visceral adipose tissue.
Ahn, J. H., Kim, E. S., Lee, C., Kim, S., Cho, S.-H., Hwang, B. Y., et al. (2013). Chemical constituents from Nelumbo nucifera leaves and their anti-obesity effects. Bioorg. Med. Chem. Lett. 23, 3604–3608. doi: 10.1016/j.bmcl.2013.04.013
Carr, D. B., Utzschneider, K. M., Hull, R. L., Kodama, K., Retzlaff, B. M., Brunzell, J. D., et al. (2004). Intra-abdominal fat is a major determinant of the National Cholesterol Education Program Adult Treatment Panel III criteria for the metabolic syndrome. Diabetes 53, 2087–2094.
Djalalinia, S., Qorbani, M., Peykari, N., and Kelishadi, R. (2015). Health impacts of Obesity. Pak. J. Med. Sci. 31, 239–242. doi: 10.12669/pjms.311.7033
Du, H., You, J. S., Zhao, X., Park, J. Y., Kim, S. H., and Chang, K. J. (2010). Antiobesity and hypolipidemic effects of lotus leaf hot water extract with taurine supplementation in rats fed a high fat diet. J. Biomed. Sci. 17(Suppl. 1):S42. doi: 10.1186/1423-0127-17-S1-S42
Feng, S., Reuss, L., and Wang, Y. (2016). Potential of natural products in the inhibition of adipogenesis through regulation of PPARgamma expression and/or its transcriptional activity. Molecules 21:1278. doi: 10.3390/molecules21101278
Festuccia, W. T., Blanchard, P. G., Turcotte, V., Laplante, M., Sariahmetoglu, M., Brindley, D. N., et al. (2009). Depot-specific effects of the PPARgamma agonist rosiglitazone on adipose tissue glucose uptake and metabolism. J. Lipid Res. 50, 1185–1194. doi: 10.1194/jlr.M800620-JLR200
Finelli, C., Sommella, L., Gioia, S., La Sala, N., and Tarantino, G. (2013). Should visceral fat be reduced to increase longevity? Ageing Res. Rev. 12, 996–1004. doi: 10.1016/j.arr.2013.05.007
Gabriely, I., Ma, X. H., Yang, X. M., Atzmon, G., Rajala, M. W., Berg, A. H., et al. (2002). Removal of visceral fat prevents insulin resistance and glucose intolerance of aging an adipokine-mediated process. Diabetes 51, 2951–2958.
Gong, F. Y., Shi, Y. F., and Deng, J. Y. (2006). The regulatory mechanism by which interleukin-6 stimulates GH-gene expression in rat GH3 cells. J. Endocrinol. 190, 397–406. doi: 10.1677/joe.1.06736
Gu, M., and Huang, C. (2013). Research and development of anti-obesity drugs: the hope behind the dilemma. Chin. J. New Drugs 22, 535–541.
Guo, F., Yang, X., Li, X., Feng, R., Guan, C., Wang, Y., et al. (2013). Nuciferine prevents hepatic steatosis and injury induced by a high-fat diet in hamsters. PLoS ONE 8:e63770. doi: 10.1371/journal.pone.0063770
Hasani-Ranjbar, S. (2009). A systematic review of the efficacy and safety of herbal medicines used in the treatment of obesity. World J. Gastroenterol. 15:3073. doi: 10.3748/wjg.15.3073
Hirashita, T., Ohta, M., Endo, Y., Masuda, T., Iwashita, Y., and Kitano, S. (2012). Effects of visceral fat resection and gastric banding in an obese diabetic rat model. Surgery 151, 6–12. doi: 10.1016/j.surg.2011.06.025
Huang, C. F., Chen, Y. W., Yang, C. Y., Lin, H. Y., Way, T. D., Chiang, W., et al. (2011). Extract of lotus leaf ( Nelumbo nucifera ) and its active constituent catechin with insulin secretagogue activity. J. Agric. Food Chem. 59, 1087–1094. doi: 10.1021/jf103382h
Huang, S., and Czech, M. P. (2007). The GLUT4 glucose transporter. Cell Metab. 5, 237–252. doi: 10.1016/j.cmet.2007.03.006
Ibrahim, M. M. (2010). Subcutaneous and visceral adipose tissue: structural and functional differences. Obes. Rev. 11, 11–18. doi: 10.1111/j.1467-789X.2009.00623.x
Janani, C., and Ranjitha Kumari, B. D. (2015). PPAR gamma gene–a review. Diabetes Metab. Syndr. 9, 46–50. doi: 10.1016/j.dsx.2014.09.015
Jones, B. J., and Bloom, S. R. (2015). The new era of drug therapy for obesity: the evidence and the expectations. Drugs 75, 935–945. doi: 10.1007/s40265-015-0410-1
Kim, A. R., Jeong, S. M., Kang, M. J., Jang, Y. H., Choi, H. N., and Kim, J. I. (2013). Lotus leaf alleviates hyperglycemia and dyslipidemia in animal model of diabetes mellitus. Nutr. Res. Pract. 7, 166–171. doi: 10.4162/nrp.2013.7.3.166
Lee, B., Kwon, M., Choi, J. S., Jeong, H. O., Chung, H. Y., and Kim, H. R. (2015). Kaempferol isolated from Nelumbo nucifera inhibits lipid accumulation and increases fatty acid oxidation signaling in adipocytes. J. Med. Food 18, 1363–1370. doi: 10.1089/jmf.2015.3457
Leonardini, A., Laviola, L., Perrini, S., Natalicchio, A., and Giorgino, F. (2009). Cross-Talk between PPARgamma and insulin signaling and modulation of insulin sensitivity. PPAR Res. 2009:818945. doi: 10.1155/2009/818945
Lin, H. Y., Kuo, Y. H., Lin, Y. L., and Chiang, W. (2009). Antioxidative effect and active components from leaves of Lotus ( Nelumbo nucifera ). J. Agric. Food Chem. 57, 6623–6629. doi: 10.1021/jf900950z
Ma, C., Li, G., He, Y., Xu, B., Mi, X., Wang, H., et al. (2015). Pronuciferine and nuciferine inhibit lipogenesis in 3T3-L1 adipocytes by activating the AMPK signaling pathway. Life Sci. 136, 120–125. doi: 10.1016/j.lfs.2015.07.001
MacLaren, R., Cui, W., Simard, S., and Cianflone, K. (2007). Influence of obesity and insulin sensitivity on insulin signaling genes in human omental and subcutaneous adipose tissue. J. Lipid Res. 49, 308–323. doi: 10.1194/jlr.M700199-JLR200
Mangelsdorf, D. J., Thummel, C., Beato, M., Herrlich, P., Schütz, G., Umesono, K., et al. (1995). The nuclear receptor superfamily: the second decade. Cell 83, 835–839.
Miyazaki, Y., Mahankali, A., Matsuda, M., Mahankali, S., Hardies, J., Cusi, K., et al. (2002). Effect of pioglitazone on abdominal fat distribution and insulin sensitivity in type 2 diabetic patients. J. Clin. Endocrinol. Metab. 87, 2784–2791. doi: 10.1210/jcem.87.6.8567
Mori, Y., Murakawa, Y., Okada, K., Horikoshi, H., Yokoyama, J., Tajima, N., et al. (1999). Effect of troglitazone on body fat distribution in type 2 diabetic patients. Diabetes Care 22, 908–912.
Mukherjee, P. K., Mukherjee, D., Maji, A. K., Rai, S., and Heinrich, M. (2009). The sacred lotus (Nelumbo nucifera) - phytochemical and therapeutic profile. J. Pharm. Pharmacol. 61, 407–422. doi: 10.1211/jpp/61.04.0001
Nordstrand, N., Gjevestad, E., Hertel, J. K., Johnson, L. K., Saltvedt, E., Roislien, J., et al. (2013). Arterial stiffness, lifestyle intervention and a low-calorie diet in morbidly obese patients-a nonrandomized clinical trial. Obesity 21, 690–697. doi: 10.1002/oby.20099
Okuno, A., Tamemoto, H., Tobe, K., Ueki, K., Mori, Y., Iwamoto, K., et al. (1998). Troglitazone increases the number of small adipocytes without the change of white adipose tissue mass in obese Zucker rats. J. Clin. Invest. 101, 1354–1361. doi: 10.1172/JCI1235
Olefsky, J. M. (2000). Treatment of insulin resistance with peroxisome proliferator-activated receptor gamma agonists. J. Clin. Invest. 106, 467–472. doi: 10.1172/JCI10843
Ono, Y., Hattori, E., Fukaya, Y., Imai, S., and Ohizumi, Y. (2006). Anti-obesity effect of Nelumbo nucifera leaves extract in mice and rats. J. Ethnopharmacol. 106, 238–244. doi: 10.1016/j.jep.2005.12.036
Palou, M., Priego, T., Sánchez, J., Rodríguez, A. M., and Palou, A. P. C. (2009). Gene expression patterns in visceral and subcutaneous adipose depots in rats are linked to their morphologic features. Cell Physiol. Biochem. 24, 547–556. doi: 10.1159/000257511
Saraf, N., Sharma, P. K., Mondal, S. C., Garg, V. K., and Singh, A. K. (2012). Role of PPARg2 transcription factor in thiazolidinedione-induced insulin sensitization. J. Pharm. Pharmacol. 64, 161–171. doi: 10.1111/j.2042-7158.2011.01366.x
Sharma, B. R., Gautam, L. N., Adhikari, D., and Karki, R. (2017). A comprehensive review on chemical profiling of Nelumbo nucifera: potential for drug development. Phytother. Res. 31, 3–26. doi: 10.1002/ptr.5732
Siegner, R., Heuser, S., Holtzmann, U., Sohle, J., Schepky, A., Raschke, T., et al. (2010). Lotus leaf extract and L-carnitine influence different processes during the adipocyte life cycle. Nutr. Metab. 7:66. doi: 10.1186/1743-7075-7-66
Siersbaek, R., Nielsen, R., and Mandrup, S. (2010). PPARgamma in adipocyte differentiation and metabolism–novel insights from genome-wide studies. FEBS Lett. 584, 3242–3249. doi: 10.1016/j.febslet.2010.06.010
Sugii, S., Olson, P., Sears, D. D., Saberi, M., Atkins, A. R., Barish, G. D., et al. (2009). PPARgamma activation in adipocytes is sufficient for systemic insulin sensitization. Proc. Natl. Acad. Sci. U.S.A. 106, 22504–22509. doi: 10.1073/pnas.0912487106
Wang, L., Waltenberger, B., Pferschy-Wenzig, E. M., Blunder, M., Liu, X., Malainer, C., et al. (2014). Natural product agonists of peroxisome proliferator-activated receptor gamma (PPARgamma): a review. Biochem. Pharmacol. 92, 73–89. doi: 10.1016/j.bcp.2014.07.018
Willson, T. M., Lambert, M. H., and Kliewer, S. A. (2001). Peroxisome proliferator-activated receptor gamma and metabolic disease. Annu. Rev. Biochem. 70, 341–367. doi: 10.1146/annurev.biochem.70.1.341
Wu, Z., Xie, Y., Morrison, R. F., Bucher, N. L., and Farmer, S. R. (1998). PPARgamma induces the insulin-dependent glucose transporter GLUT4 in the absence of CEBPalpha during the conversion of 3T3 fibroblasts into adipocytes. J. Clin. Invest. 101, 22–32. doi: 10.1172/JCI1244
Yin, J., Zhang, H., and Ye, J. (2008). Traditional chinese medicine in treatment of metabolic syndrome. Endocr. Metab. Immune Disord. Drug Targets 8, 99–111.
Zeng, Z. H., Xu, Y., and Zhang, B. (2016). Antidiabetic activity of a lotus leaf selenium (Se)-polysaccharide in rats with gestational diabetes mellitus. Biol. Trace Elem. Res. 176, 321–327. doi: 10.1007/s12011-016-0829-6
Zhu, H. J., Deng, J. Y., and Gong, F. Y. (2006). Primary culture of human preadipocyte in a serum-free medium. Basic Clin. Med. 7, 770–773.
Zhu, H. J., Ding, H. H., Deng, J. Y., Pan, H., Wang, L. J., Li, N. S., et al. (2013). Inhibition of preadipocyte differentiation and adipogenesis by zinc-alpha2-glycoprotein treatment in 3T3-L1 cells. J. Diabetes Investig. 4, 252–260. doi: 10.1111/jdi.12046
Zhu, H. J., Wang, L. J., Wang, X. Q., Pan, H., Li, N. S., Yang, H. B., et al. (2014). Hormone-sensitive lipase is involved in the action of hydroxysafflor yellow A (HYSA) inhibiting adipogenesis of 3T3-L1cells. Fitoterapia 93, 182–188. doi: 10.1016/j.fitote.2014.01.001
Zhu, H. J., Wang, X. Q., Pan, H., Dai, Y. F., Li, N. S., Wang, L. J., et al. (2016). The mechanism by which safflower yellow decreases body fat mass and improves insulin sensitivity in HFD-induced obese mice. Front. Pharmacol. 7:127. doi: 10.3389/fphar.2016.00127
Zhu, M. Z., Wu, W., Jiao, L. L., Yang, P. F., and Guo, M. Q. (2015). Analysis of flavonoids in lotus (Nelumbo nucifera) leaves and their antioxidant activity using macroporous resin chromatography coupled with LC-MS/MS and antioxidant biochemical assays. Molecules 20, 10553–10565. doi: 10.3390/molecules200610553
Keywords: lotus leaf aqueous extract (LLAE), PPARγ2, visceral adipose tissue, insulin resistance, obese rats
Citation: Yan K, Zhu H, Xu J, Pan H, Li N, Wang L, Yang H, Liu M and Gong F (2017) Lotus Leaf Aqueous Extract Reduces Visceral Fat Mass and Ameliorates Insulin Resistance in HFD-Induced Obese Rats by Regulating PPARγ2 Expression. Front. Pharmacol. 8:409. doi: 10.3389/fphar.2017.00409
Received: 01 March 2017; Accepted: 09 June 2017;
Published: 23 June 2017.
Edited by:
Adolfo Andrade-Cetto, National Autonomous University of Mexico, MexicoReviewed by:
Marcia Hiriart, National Autonomous University of Mexico, MexicoCopyright © 2017 Yan, Zhu, Xu, Pan, Li, Wang, Yang, Liu and Gong. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Feng Ying Gong, Znlnb25nQGFsaXl1bi5jb20=
†These authors have contributed equally to this work.
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.