 Carlota Recio
Carlota Recio Francesco Maione
Francesco Maione Asif J. Iqbal
Asif J. Iqbal Nicola Mascolo
Nicola Mascolo Vincenzo De Feo
Vincenzo De Feo
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Pharmacol. , 06 January 2017
Sec. Cardiovascular and Smooth Muscle Pharmacology
Volume 7 - 2016 | https://doi.org/10.3389/fphar.2016.00526
Cardiovascular disease (CVD) remains a leading cause of mortality and morbidity worldwide. Numerous therapies are currently under investigation to improve pathological cardiovascular complications, but yet, there have been very few new medications approved for intervention/treatment. Therefore, new approaches to treat CVD are urgently required. Attempts to prevent vascular complications usually involve amelioration of contributing risk factors and underlying processes such as inflammation, obesity, hyperglycaemia, or hypercholesterolemia. Historically, the development of peptides as therapeutic agents has been avoided by the Pharmaceutical industry due to their low stability, size, rate of degradation, and poor delivery. However, more recently, resurgence has taken place in developing peptides and their mimetics for therapeutic intervention. As a result, increased attention has been placed upon using peptides that mimic the function of mediators involved in pathologic processes during vascular damage. This review will provide an overview on novel targets and experimental therapeutic approaches based on peptidomimetics for modulation in CVD. We aim to specifically examine apolipoprotein A-I (apoA-I) and apoE mimetic peptides and their role in cholesterol transport during atherosclerosis, suppressors of cytokine signaling (SOCS)1-derived peptides and annexin-A1 as potent inhibitors of inflammation, incretin mimetics and their function in glucose-insulin tolerance, among others. With improvements in technology and synthesis platforms the future looks promising for the development of novel peptides and mimetics for therapeutic use. However, within the area of CVD much more work is required to identify and improve our understanding of peptide structure, interaction, and function in order to select the best targets to take forward for treatment.
Cardiovascular disease remains a leading cause of mortality and morbidity worldwide. In developed countries, risk factors such as hypertension, hyperglycemia, and hypercholesterolemia are accepted as having a key role in driving CVD (Leening et al., 2016). Researchers and clinicians have spent significant time and effort investigating the role of these risk factors in the development and progression of CVD, yet there have been a limited number of new medications approved for CVD-related intervention and/or treatment. Therefore, new approaches to treat CVD are needed. Attempts to prevent vascular complications usually involve amelioration of contributing risk factors and underlying processes such as inflammation, obesity, hyperglycaemia, or hypercholesterolemia (Navickas et al., 2016; Pirlamarla and Bond, 2016).
Targeting lipids has been the major strategy used in treating CVD to date. Hypercholesterolemia plays a key role in peripheral coronary artery disease progression, mainly atherosclerosis. High concentration of low density-lipoprotein (LDL) particles in plasma drives cholesterol accumulation in arteries setting up the initial stage of atheroma plaque formation (Libby et al., 2011; Manduteanu and Simionescu, 2012). Excessive lipid accumulation in the arterial intima induces a significant inflammatory response resulting in increased pro-inflammatory cytokines, adhesion molecules, and chemokine expression, which leads to endothelial dysfunction and leukocyte infiltration (Manduteanu and Simionescu, 2012; Schett et al., 2013; LeBert and Huttenlocher, 2014). Further influx (mainly macrophages, T cells, and vascular smooth muscle cells) of cells into the lesion area triggers plaque hardening and growth. Finally, vessel diameter decreases and, if the plaque is unstable, it can cause significant clinical consequences such MI or stroke (Fuster et al., 2005).
The current first line drugs used in CVD treatment to date are ACEIs, ARBs, anticoagulants, cholesterol-lowering drugs (statins), beta-blockers, and some anti-inflammatory medicines (NSAID, glucocorticoids). The majority of these drugs have shown efficacy but many are also associated with a wide range of side-effects and are therefore inadequate to use in long-term treatment regimens (Nathan, 2002; Lawrence et al., 2002; Costopoulos et al., 2013; Cheng et al., 2014; Pellicori and Costanzo, 2015; Stein and Raal, 2015).
This review will explore novel targets and experimental therapeutic approaches based on peptidomimetics for modulation in CVD including atherosclerosis, vascular diabetic complications and MI, among others.
Therapeutic peptides are described as naturally occurring short amino acid monomer chains, shorter than 100 amino acids, and they act by binding to specific cell surface receptors, where they trigger intracellular pathways (Vlieghe et al., 2010). They have been shown to possess desirable pharmacological profiles and their specificity has been seen to translate into outstanding safety, tolerability, and efficacy profiles in humans, in stark contrast to traditional small molecules (Vlieghe et al., 2010; Goodwin et al., 2012).
The idea of using peptides as therapeutic agents has been historically ignored by pharmaceutical companies due to several limitations including size, which makes them very susceptible to degradation by peptidases, the lack of effective methods for delivery, poor transport properties through biologic membranes, low oral bioavailability, rapid excretion, and poor target specificity resulting from the flexible nature of peptides (Table 1) (Vagner et al., 2008). More recently, however, in light of advances in processing technologies, there has been a renewed interest in peptides and peptidomimetics as potential therapeutic agents. This is partly due to numerous improvements made to stability, transport, affinity profiles, and oral availability (Goodwin et al., 2012; Fosgerau and Hoffmann, 2015). Furthermore, the introduction of alternative delivery methods by new adjuvant and carrier systems have been developed, and the advance of proteomics identifying innumerable PPI targets, has increased the interest in peptides and their mimetics as potential therapeutic drugs (Liskamp et al., 2011; Akram et al., 2014).

TABLE 1. Advantages and disadvantages of peptides as therapeutics.
To date, more than 7000 naturally occurring peptides have been described (Fosgerau and Hoffmann, 2015). The first chemical synthesis of a therapeutic peptide was that of oxytocin in 1953. Recombinant synthesis of proteins was introduced in 1974, and recombinant human insulin, the first approved therapeutic peptide to be manufactured by recombinant fermentation, was introduced in 1982 (Puttagunta and Toth, 1998). Although being used for the last five decades, it still enjoys the fame of being the most generally prescribed peptide worldwide (McGill et al., 2016). At present, there are more than 60 peptide-based drug products that have reached approval and nearly 140 in clinical trials (Lax and Meenan, 2012; Uhlig et al., 2014).
To address these key technical hurdles to use peptides as medicines, a number of modifications strategies (thanks to robust peptide-chemistry approaches developed in recent years) have been widely adopted. Several bioactive peptides have proven to be highly functional with many serving as potent agonists and antagonists against numerous receptors implicated in disease progression (Kaspar and Reichert, 2013; Peptide Therapeutics Market, 2015).
Transformation of peptides to peptidomimetics is one intriguing mechanism to use peptide sequences as potential therapeutic agents. Peptides can be adapted to stable mimics that expose similar effects to their peptide analog but show increased consistency in structure, more target specificity, increased stability to proteolytic digestion and greater cell membrane permeability. Therefore, peptides have been chemically altered to include unnatural amino acid substitutions, backbone amide bond modifications, or rigid scaffolds or the addition of hydrophobic residues (Gentilucci et al., 2006; Vagner et al., 2008; Vlieghe et al., 2010).
Among the major drawbacks faced by the use of peptides as drugs is their delivery. To date, injections remain the most common route of administration. However, oral delivery would be preferable because of its high level of patient compliance which increases the therapeutic value of a drug. So the challenge remains to improve the oral bioavailability from less than 1% to at least 30–50%. Recently, development of orally and nasal active preparations have been proposed as a result of encapsulation of peptides in nanoparticles (e.g., liposomes, synthetic polymers, or fullerenes) which shields the drug from protease digestion until required, therefore increasing stability (Mason, 2010; Bruno et al., 2013). Other strategies under investigation to overcome peptide barriers include use of protease inhibitors, absorption enhancers, or conjugated molecules in combination with the peptide structure, such as antibodies to improve targeting, carbohydrates to increase solubility, or lipids to enhance peptide permeability (Shaji and Patole, 2008; Bruno et al., 2013; Di, 2015)
Equally, synthesis costs are a big issue. The synthesis of peptides relies heavily on expensive coupling reagents, resins and protected amino acids, so cheaper methods for their synthesis and purification are required (Mason, 2010). In line with this, chemical methods such as click chemistry and peptide synthesis reactors that can handle large amounts of reaction material for solid-phase peptide synthesis can substantially lower costs as well as improve the chemistry (Sohma et al., 2004; Fabbrizzi et al., 2014).
Another major breakthrough has been the variety drive in technology platforms to study PPIs. With more information related to 3D structure of protein complexes and PPIs and their importance in human diseases, peptide- and peptidomimetic-based therapeutic agents have become a major area of drug design, competing with natural products, synthetic small molecules, and antibody-based therapies (Gao et al., 2015).
Considering all the upgrades in peptide systems, and the rapid developments in proteomics, bioinformatics, and peptide libraries, it is expected that by 2020, the global Peptide Therapeutics market will reach over $25 billion (Global Peptide Therapeutics Market, 2016). This dramatic market increase is driven by both growing incidences of cardiovascular and metabolic diseases, and also technological enhancements in peptide synthesis that include high-throughput approaches.
Attempts to prevent cardiovascular diseases usually involve control and improvement of causative risk factors such as hypercholesterolemia, inflammation, hyperglycaemia, obesity, insulin resistance, or high blood pressure. Limitations in currently available device therapies and pharmacologic drugs in CVD has prompted wider investigation into new treatment modalities such peptides and their mimetics.
Dyslipidemia is one of most relevant risk factors for coronary artery disease. Therefore, one of the key goals of cardiovascular therapies is to reduce LDL cholesterol accumulation in the subendothelial space lining the artery wall, thereby preventing the progression of atherosclerosis and reducing the risk of heart attack and stroke. A plasma LDL cholesterol reduction of 1 mmol/L has been reported to reduce the risk of cardiovascular events by approximately 20% (Stoekenbroek et al., 2015). HDL is considered to promote the removal of free cholesterol from peripheral tissue and its transport to the liver for eventual clearance. ApoA-I, the major protein component of the HDL particle, is predominantly responsible for the anti-atherogenic properties attributed to HDL (Fisher et al., 2012). ApoA-I is critical for the process of reverse cholesterol transport and cellular cholesterol homeostasis. Several murine pre-clinical models of atherosclerosis have shown potent protective effects of apoA-I following prophylactic and therapeutic intervention (Gordon et al., 2011). Furthermore, genetic ablation of apoA-I in LDL receptor knockout mice, was shown to significantly promote atherosclerosis progression (Moore et al., 2003).
In addition to its role in cholesterol transport, other vascular beneficial effects have been attributed to apoA-I (Mangaraj et al., 2016). Recent studies have reported a potential anti-inflammatory role for apoA-I in the regulation of monocyte/macrophage recruitment to local sites of inflammation, via modulation of lipid rafts in cellular membranes which resulted in suppression of PI3K/Akt signaling (Iqbal et al., 2016). It also displays anti-oxidant properties as shown by its ability to inhibit LDL oxidation, remove lipid hydroperoxides, and also protect endothelial cells from apoptosis (Suc et al., 1997; Podrez, 2010; Rosenbaum et al., 2015). Furthermore, its structural homology with PSF has contributed to its anti-clotting and anti-aggregation effects on platelets which has strengthened its cardioprotective role (Yui et al., 1988). All in all, apoA-I is widely considered as a promising target for CVD treatment, and different therapeutic approaches have been developed to mimic its function.
ApoA-I is a 243 amino acid molecule with a secondary structure of 10 amphipathic αα-helices necessary for its interaction with lipids (Davidson et al., 1996). This secondary structure has been used as a template to design a range of apoA-I mimetic peptides. Although they are functionally similar to the native protein, they have unique structural properties (Table 2) (Stoekenbroek et al., 2015).
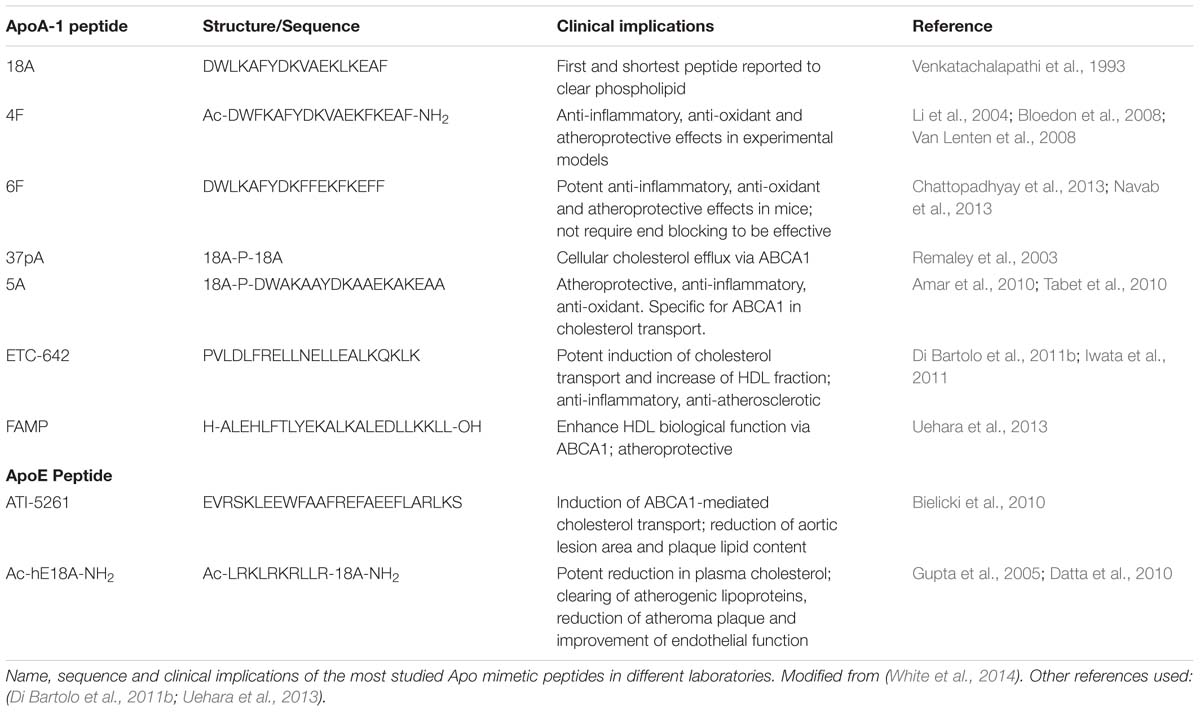
TABLE 2. Apolipoprotein mimetic peptides.
The first apoA-I mimetic peptide, 18A, was synthesized by Anantharamaiah et al. (1985; Venkatachalapathi et al., 1993). Subsequently, this 18 amino acid peptide has undergone numerous modifications to generate variant mimetic peptides with increased homology to apoA-I, higher lipid affinity and enhanced anti-atherogenic properties (Garber et al., 1992). An example of a such a variant is 4F, the most well-studied apoA-I mimetic, that reproduces the helical and amphipathic portion of apoA-I which is key for its function (Navab et al., 2005). Other peptides such as D-4F and L-4F, consist of the D- and L-isomers of the amino acids and show similar functionality as apoA-I, with D-4F being more stable via oral administration (Navab et al., 2002). However, although initially demonstrating potent anti-inflammatory, anti-oxidant, and atheroprotective effects in pre-clinical experimental models in apoE null mice and in human aortic cell cultures, 4F peptides have failed to show any efficacy in human trials (Li et al., 2004; Bloedon et al., 2008; Van Lenten et al., 2008; Watson et al., 2011).
Several other apoA-I mimetics have been developed to overcome some of the weaknesses of previous peptides. For example, the 6F peptide emerged as a promising apoA-I mimetic which did not require end blocking to be effective, and therefore reduced overall costs for synthesis. This peptide was also shown to possess potent anti-inflammatory, anti-oxidant, and atheroprotective effects in pre-clinical experimental models in LDL receptor-null mice (Chattopadhyay et al., 2013; Navab et al., 2013). 5A peptide was synthesized based on an existing 37 pA peptide structure to which five amino acids where replaced in order to decrease its cytotoxicity associated with its elevated lipid affinity (Remaley et al., 2003). In this way, 5A was less toxic and more specific to the ATP-binding cassette transporter A1(ABCA1) in cholesterol transport. Moreover, this apoA-I mimetic reduced pro-inflammatory adhesion molecule expression, neutrophil infiltration, and oxidative stress in animal models of inflammation in rabbits and also in vitro in human coronary artery endothelial cells (Tabet et al., 2010). 5A was also shown to be atheroprotective in pre-clinical mouse models and there are current proposals under consideration to take this mimetic forward into clinical trials (Amar et al., 2010).
ETC-642 is a 22 amino acid apoA-I mimetic peptide that offers numerous beneficial effects on LDL and HDL particles, including reduction of pro-inflammatory oxidized LDLs, potent induction of cholesterol transport, and increase of cholesterol content in the HDL fraction. It has also been attributed with significant anti-inflammatory properties in several studies of acute and chronic inflammation in rabbits, where it was shown to reduce TNFα induced expression of NF-Kb and endothelial adhesion molecule expression (Di Bartolo et al., 2011a,b). Furthermore, ETC-642 was shown to inhibit plaque formation in an experimental model of atherosclerosis in hyperlipidemic rabbits (Iwata et al., 2011).
In 2010, a systematic study of 22 different apoA-I mimetic peptides reported by D’Souza et al. (2010) showed that the structural modifications of each peptide were related with their different capacity and specificity of cholesterol efflux and their inhibitory effects on inflammation and LDL oxidation. In this analysis none of the peptides tested were found to be equally effective in all anti-atherogenic functions (D’Souza et al., 2010).
Many of these apoA-I mimetic peptides are in pre-clinical stages of development (Smith, 2010; White et al., 2014; Uehara et al., 2015). A newly described apoA-I mimetic peptide, called FAMP (Fukuoka University APOA-I mimetic peptide), has been reported to function via ABCA1 in a highly specific manner. This novel mimetic peptide has been shown to effectively enhance HDL biological function and it also has atheroprotective functions in apoE-deficient mice (Uehara et al., 2013).
More recently, apoE mimetic peptides were shown to have a beneficial impact on HDL functionality. ApoE is a 299 amino acid protein that plays an important role in clearing apoB-containing remnant particles mainly chylomicrons (that absorb lipids from the diet in the intestine), very low-density lipoproteins (VLDL, that transport triglycerides to tissues), and other lipoproteins that can be atherogenic (Bocksch et al., 2001). ApoE clears lipoproteins by LDL receptor-independent mechanisms. It also plays a crucial role in the regulation of plasma cholesterol levels, given that it contains an LDL binding domain in its structure (Hatters et al., 2006; Mahley et al., 2006). In addition, other beneficial effects have been attributed to apoE including anti-inflammatory, anti-oxidant, and anti-coagulant properties (Ali et al., 2005; Pham et al., 2005; Gaudreault et al., 2012).
Several mimetic peptides based on apoE structure have been recently designed (Table 2). Among them, ATI-5261 is a 36 amino acid peptide that has been reported to induce ABCA1-mediated cholesterol transport and reduce aortic lesion area and plaque lipid content in several pre-clinical models of atherosclerosis in mice (Bielicki et al., 2010). Anantharamaiah et al. (1985) developed various synthetic dual-domain apolipoprotein peptides which are structurally and functionally similar to apoA-I and apoE but mimic the cholesterol-lowering properties of apoE (Datta et al., 2001; Sharifov et al., 2011). The most characterized is Ac-hE18A-NH2, composed of a region of the LDL binding domain of apoE linked to the apoA-I mimetic 18A (Sharifov et al., 2011). This peptide was shown to dramatically reduce plasma cholesterol in several dyslipidemic animal models and had the extra advantage of clearing atherogenic lipoproteins due to the presence of the LDL binding domain, resulting in the reduction of atheroma plaque formation and the improvement of endothelial function (Gupta et al., 2005; Datta et al., 2010). This novel intravenously administered-peptide has been assigned orphan drug status and, under the name AEM-28, is currently undergoing initial (phases 1 and 2) clinical assessment (White et al., 2014).
Many of these peptides are still in pre-clinical phases of development and to date it has been difficult to identify an efficacy parameter for apo mimetics in human trials collectively. One major reason for the discrepancy observed in humans and mice could be differences in the composition of lipid associated proteins. A study from Gordon et al. (2015) utilized a mass spectrometry approach to demonstrate a high degree of shared homology amongst a range of proteins associated with LDL and HDL. However, a small minority of proteins did exhibit significant differences which could reflect in major metabolic differences between species (Gordon et al., 2015).
Surprisingly, there are no reported studies which have compared the efficacy of statins versus apoA-I mimetics in humans to date. LDL-lowering statin therapy is currently considered the ‘gold standard’ treatment for CVD. Statins are very effective and safe in atherogenic dyslipidemia treatment. However, they have shown to lack benefit for retarding residual adverse cardiovascular events. Even under optimal statin treatment, patients with familial hypercholesterolemia present with high level of LDL cholesterol and there are also patients who are intolerant or unresponsive to statins, highlighting a potential role for the use of apo mimetics in such patients (Boekholdt et al., 2013; Ahn and Choi, 2015; Uehara et al., 2015). Given the current interest in this field we can expect to have novel apo mimetic peptides in the near future to aid in the prevention and treatment of patients with cardiovascular disorders.
It is widely accepted that inflammation participates in all stages of atherosclerosis, from its initiation to its thrombotic complications (Libby et al., 2011). Therefore, targeting inflammatory mediators that dynamically take part in chronic inflammation which underlies disease could be an interesting clinical strategy. In this context, SOCS proteins, which are at the crossroad of multiple inflammatory pathways, have recently emerged as a potential therapeutic target with anti-inflammatory functions (Linossi et al., 2013; Trengove and Ward, 2013). SOCS are negative-feedback regulators of the JAK/STAT pathway, which drive the production of cytokines and inflammatory factors that affect atherosclerotic processes, including leukocyte recruitment, migration, and proliferation of vascular cells, foam cell formation and apoptosis (Marrero, 2005; Miklossy et al., 2013). Among the eight members of this family of proteins (SOCS1-7 and CIS), SOCS1 and SOCS3 are of particular interest because they contain a conserved 12-residue KIR that is involved in direct suppression of JAK activity and they have also been linked to a variety of pro-inflammatory and pro-atherogenic factors including lipoproteins, lipids, high glucose, angiotensin II, and insulin (Alexander, 2002; Yoshimura et al., 2007; Liang et al., 2013). Furthermore, experimental studies in mice and murine aortic cells demonstrate that SOCS overexpression reduces inflammation and cardiovascular disease (Tajiri et al., 2012; Qin et al., 2014). Studies based on peptides mimicking the action of SOCS proteins have been reported in different experimental settings (Table 3). The first SOCS mimetic peptide developed was JAK2 Tkip (Flowers et al., 2004). This short 12-mer peptide was shown to suppress the expression of inflammatory cytokines such as TNFα, inhibit lymphocyte proliferation as well as IFNγ-induced macrophage activation and NO production in mice (Mujtaba et al., 2005; Ahmed et al., 2009). SOCS1-KIR peptidomimetic was reported to inhibit STAT activation by Th1 and Th17 cytokines in leukocytes as well as suppress the expression of pro-inflammatory mediators and activation and migration of vascular cells and macrophages in vitro (Ahmed et al., 2015). SOCS1-KIR was also shown to be atheroprotective in a type I diabetes mouse model, decreasing vascular plaque accumulation of lipids, macrophages, and T cells, and reducing aorta expression of pro-inflammatory cytokines and chemokines (Recio et al., 2014). More recently, this SOCS1-KIR peptide was demonstrated to further improve diabetes associated-renal damage in mice as well as reduce inflammation and fibrosis in diabetic kidneys (Recio et al., 2016).
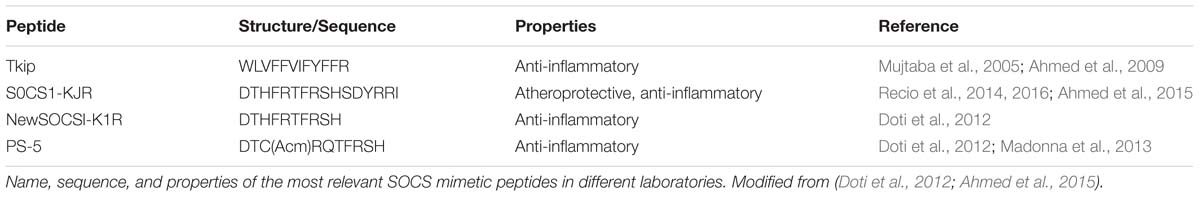
TABLE 3. SOCS mimetic peptides.
Doti et al. (2012) recent studies focused on identifying new improved mimetic peptides of the KIR region of SOCS1 but with enhanced affinity, stability, and potency profiles. Among them, PS-5 was highlighted because it bound to JAK2 more efficiently than KIR and also prevented the IFNγ-induced activation of STAT and its downstream inflammatory effects (Doti et al., 2012; Madonna et al., 2013).
In summary these peptidomimetics emerge not only as potent anti-inflammatory agents but also as promising future drugs in the treatment of cardiovascular complications in diabetic patients.
In diabetic patients, the control of blood glucose levels is a major goal to prevent further tissue damage and cardiovascular events such as stroke, heart attack, or end-stage renal disease (Snell-Bergeon and Wadwa, 2012). Incretin mimetic-based therapies, with particular focus on GLP-1R agonists and DPP4 inhibitors, are currently leading therapeutic agents available for type 2 diabetes treatment (Drucker and Nauck, 2006). As peptidomimetics, GLP-1R agonists mimic the actions of the endogenous hormone GLP-1 in that they stimulate glucose-induced insulin secretion, suppress glucagon secretion and hepatic glucose production and delay gastric emptying. In addition, GLP-1 has been reported to enhance peripheral glucose disposal (very important in diabetes) as well as promote pancreatic beta cell growth and differentiation (Drucker and Nauck, 2006; Meier, 2012). Furthermore, GLP-1R agonists can act both in a short and long-term manner, allowing personalized patient regimes to be offered (Neumiller, 2015).
DPP-4 is the enzyme that inactivates GLP-1, therefore its inhibition emerges as another potential target to increase circulating levels of GLP-1 thereby increase circulating incretin levels (Deacon et al., 1998, Deacon, 2011).
Incretin mimetics present other favorable properties such as a low hypoglycaemia risk, the ability to address postprandial hyperglycemia (DPP-4 inhibitors and short-acting GLP-1R agonists), and potential for weight reduction (GLP-1R agonists; Neumiller, 2015).
Interestingly, besides regulation of glucose homeostasis, GLP-1 mimetic peptides have also been shown to exert cardioprotective effects in cardiovascular-related death, non-fatal MI, and non-fatal stroke (Advani et al., 2013; Wroge and Williams, 2016).
In the last decade, three different GLP-1R agonists have been approved for clinical use; Exenatide, first approved in 2005, Liraglutide and Lixisenatide; Albiglutide, Dulaglutide, and Semaglutide are in last phases of evaluation (Table 4) (Eng et al., 1992; Meier, 2012). One particular feature of Exenatide and Lixisenatide is that, in contrast to the endogenous GLP-1 which is degraded within 1–2 min by DDP-4, they are both DDP-4-resistant. While Exenatide requires a twice daily dosing regime, Lixisenatide can be given once a day because it has a higher affinity for GLP-1R. However, they have a similar half-life (2–4 h; Madsbad et al., 2011; Bhavsar et al., 2013; Kalra et al., 2016). Extended stability and longer half-life of these compounds would be favorable. In contrast, Liraglutide, can be administered once a day and has half-life of 13 h as a result of a modification to the peptide backbone with palmitic acid. This compound has been reported to induce significant weight loss and reduce blood pressure as well as diabetes prevalence in type 2 diabetic patients (Juhl et al., 2002; Russell, 2013).
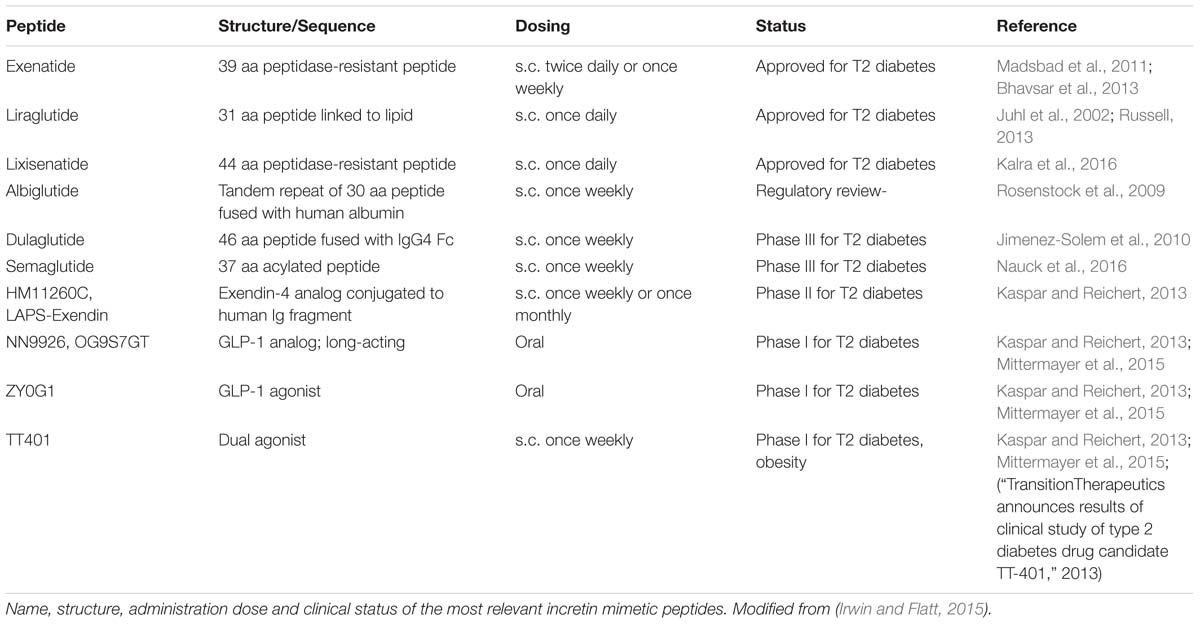
TABLE 4. Incretin mimetic peptides.
Albiglutide and Dulaglutide are incretin mimetics with increased half-life (4–7 days) which allow for weekly administration. This extension of their half-life is feasible owing to their fusion with different molecules that confer them stability (i.e., human albumin; Rosenstock et al., 2009). There are two completed studies with Albiglutide that demonstrate the safety and efficacy of weekly, subcutaneously injected doses compared to other treatments such as Liraglutide or insulin. Dulaglutide is administered by subcutaneous injection once weekly for up to 24 months at seven doses (Jimenez-Solem et al., 2010). Semaglutide is to date the last one in the list of GLP-1 mimetics which are under clinical assessment. Phase III studies of this compound confirm that it can be administered subcutaneously once weekly and it improves glycaemic control in type 2 diabetes patients in a superior way than Exenatide. Semaglutide also reduces the risk of major cardiovascular events and decreases appetite and food intake, therefore becoming an interesting drug to be used in obese patients (Nauck et al., 2016).
Incretin mimetics are the current preferred drug to treat type 2 diabetes owing to their wide range of beneficial effects. However, although many of them are already in clinical use, evolution of this group of peptides is not complete. There are still a high number of studies focused on improving patient convenience and compliance so looking for strategies to reduce dosing frequency or developing oral administrated compounds.
Given that MI remains a major cause of death worldwide and the current therapies based in revascularization of the ischemic tissue (anti-oxidants and calcium channel blockers) have shown insufficient success, novel strategies are needed to treat patients with MI. In this context, the therapeutic potential of glucocorticoid-regulated anti-inflammatory mediator annexin-A1 has been demonstrated in different systemic inflammatory disorders. Annexin-A1 is a glucocorticoid-inducible 37 kDa protein, highly expressed by macrophages, that activates the family of formyl peptide receptors and inhibits different processes related to myocardial reperfusion injury such as polymorphonuclear leukocyte activation, migration, and infiltration (Ambrose et al., 1992; De Caterina et al., 1993; La et al., 2001; Perretti and Gavins, 2003; Qin et al., 2015).
Due to the potent anti-inflammatory and cardioprotective properties of endogenous annexin-A1, several studies utilized experimental models to examine the role of the exogenous protein and its derived peptides (Perretti and Gavins, 2003). The main benefits attributed to annexin-A1 peptide mimetics include cardioprotection based on their anti-inflammatory effect to preserve myocardial viability after MI but also other inflammation-independent properties that directly protect cardiomyocytes viability and contractile function (Qin et al., 2015). The subcutaneous administration of annexin-A1 N-terminal derived peptide Ac2-26 has been shown to confer protection against ischemia-reperfusion injury by reducing myeloperoxidase activity and IL-1β levels in the infarcted heart, as well as down-regulate monocyte accumulation and inhibit phagocytic activity of macrophages in different rodent experimental models (Getting et al., 1997; La et al., 2001). Another annexin-A1 mimetic is CGEN-855A, a 21 amino acid peptide displays anti-inflammatory effects by inhibition of polymorphonuclear neutrophils recruitment and also provides protection against ischemia-reperfusion-mediated injury to the myocardium after being injected intravenously in mice (Hecht et al., 2009).
The view that peptides hold multiple properties as therapeutics, including suitable pharmacokinetic profiles, low toxicity and immunogenicity, and desirable solubility features, is broadly accepted. Since many of the classical limitations they possess to act as drug agents are being overcome by improving techniques and modifications, the use of peptides and peptidomimetics as a therapeutic strategy is growing.
Although the use of these molecules in CVD treatment is gaining traction, more effort is needed to improve therapeutic potential. With further studies of the structures, interactions, and functions of proteins and mediators implicated in CVD, more peptides will be discovered and developed. With this strategy, the use of these molecules could provide good opportunities for cardiovascular prevention and treatment, surpassing some of the limitations of current therapies.
CR and FM drafted the manuscript; AI drafted part of the manuscript and designed it; NM and VDF drafted part of the manuscript and revised it critically for intellectual content.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
ABCA1, ATP-binding cassette transporter A1; ACEIs, Angiotensin-converting enzyme inhibitors; Apo, Apolipoprotein; ARBs, Angiotensin II Receptor Blockers; CVD, Cardiovascular disease; DPP4, Dipeptidyl peptidase 4; GLP-1R, Glucagon-like peptide-1 receptor; KIR, Kinase inhibitory region; MI, Myocardial infarction; NO, Nitric oxide; NSAID, Non-steroidal anti-inflammatory drug; PPI, Protein-protein interactions; PSF, Prostacyclin stabilization factor; SOCS, Suppressors of Cytokine Signaling; STAT, Signal transducer and activator of transcription; Tkip, Tyrosine kinase inhibitor peptide; VLDL, Very low-density lipoproteins.
Advani, A., Bugyei-Twum, A., and Connelly, K. A. (2013). Cardiovascular effects of incretins in diabetes. Can. J. Diabetes 37, 309–314. doi: 10.1016/j.jcjd.2013.06.010
Ahmed, C. M., Dabelic, R., Waiboci, L. W., Jager, L. D., Heron, L. L., and Johnson, H. M. (2009). SOCS-1 mimetics protect mice against lethal poxvirus infection: identification of a novel endogenous antiviral system. J. Virol. 83, 1402–1415. doi: 10.1128/JVI.01138-08
Ahmed, C. M., Larkin, J., and Johnson, H. M. (2015). SOCS1 mimetics and antagonists: a complementary approach to positive and negative regulation of immune function. Front. Immunol. 6:183. doi: 10.3389/fimmu.2015.00183
Ahn, C. H., and Choi, S. H. (2015). New drugs for treating dyslipidemia: beyond statins. Diabetes Metab. J. 39, 87–94. doi: 10.4093/dmj.2015.39.2.87
Akram, O. N., DeGraff, D. J., Sheehan, J. H., Tilley, W. D., Matusik, R. J., Ahn, J. M., et al. (2014). Tailoring peptidomimetics for targeting protein-protein interactions. Mol. Cancer Res. 12, 967–978. doi: 10.1158/1541-7786.MCR-13-0611
Alexander, W. S. (2002). Suppressors of cytokine signalling (SOCS) in the immune system. Nat. Rev. Immunol. 2, 410–416. doi: 10.1038/nri818
Ali, K., Middleton, M., Puré, E., and Rader, D. J. (2005). Apolipoprotein E suppresses the type I inflammatory response in vivo. Circ. Res. 97, 922–927. doi: 10.1161/01.RES.0000187467.67684.43
Amar, M. J., D’Souza, W., Turner, S., Demosky, S., Sviridov, D., Stonik, J., et al. (2010). 5A apolipoprotein mimetic peptide promotes cholesterol efflux and reduces atherosclerosis in mice. J. Pharmacol. Exp. Ther. 334, 634–641. doi: 10.1124/jpet.110.167890
Ambrose, M. P., Bahns, C. L., and Hunninghake, G. W. (1992). Lipocortin I production by human alveolar macrophages. Am. J. Respir. Cell Mol. Biol. 6, 17–21. doi: 10.1165/ajrcmb/6.1.17
Anantharamaiah, G. M., Jones, J. L., Brouillette, C. G., Schmidt, C. F., Chung, B. H., Hughes, T. A., et al. (1985). Studies of synthetic peptide analogs of the amphipathic helix. Structure of complexes with dimyristoyl phosphatidylcholine. J. Biol. Chem. 260, 10248–10255.
Bhavsar, S., Mudaliar, S., and Cherrington, A. (2013). Evolution of exenatide as a diabetes therapeutic. Curr. Diabetes Rev. 9, 161–193. doi: 10.2174/1573399811309020007
Bielicki, J. K., Zhang, H., Cortez, Y., Zheng, Y., Narayanaswami, V., Patel, A., et al. (2010). A new HDL mimetic peptide that stimulates cellular cholesterol efflux with high efficiency greatly reduces atherosclerosis in mice. J. Lipid Res. 51, 1496–1503. doi: 10.1194/jlr.M003665
Bloedon, L. T., Dunbar, R., Duffy, D., Pinell-Salles, P., Norris, R., DeGroot, B. J., et al. (2008). Safety, pharmacokinetics, and pharmacodynamics of oral apoA-I mimetic peptide D-4F in high-risk cardiovascular patients. J. Lipid Res. 49, 1344–1352. doi: 10.1194/jlr.P800003-JLR200
Bocksch, L., Stephens, T., Lucas, A., and Singh, B. (2001). Apolipoprotein E: possible therapeutic target for atherosclerosis. Curr. Drug Targets Cardiovasc. Haematol. Disord. 1, 93–106. doi: 10.2174/1568006013337944
Boekholdt, S. M., Arsenault, B. J., Hovingh, G. K., Mora, S., Pedersen, T. R., Larosa, J. C., et al. (2013). Levels and changes of HDL cholesterol and apolipoprotein A-I in relation to risk of cardiovascular events among statin-treated patients: a meta-analysis. Circulation 128, 1504–1512. doi: 10.1161/CIRCULATIONAHA.113.002670
Bruno, B. J., Miller, G. D., and Lim, C. S. (2013). Basics and recent advances in peptide and protein drug delivery. Ther. Deliv. 4, 1443–1467. doi: 10.4155/tde.13.104
Chattopadhyay, A., Navab, M., Hough, G., Gao, F., Meriwether, D., Grijalva, V., et al. (2013). A novel approach to oral apoA-I mimetic therapy. J. Lipid Res. 54, 995–1010. doi: 10.1194/jlr.M033555
Cheng, J., Zhang, W., Zhang, X., Han, F., Li, X., He, X., et al. (2014). Effect of angiotensin-converting enzyme inhibitors and angiotensin II receptor blockers on all-cause mortality, cardiovascular deaths, and cardiovascular events in patients with diabetes mellitus: a meta-analysis. JAMA Intern. Med. 174, 773–785. doi: 10.1001/jamainternmed.2014.348
Costopoulos, C., Niespialowska-Steuden, M., Kukreja, N., and Gorog, D. A. (2013). Novel oral anticoagulants in acute coronary syndrome. Int. J. Cardiol. 167, 2449–2455. doi: 10.1016/j.ijcard.2012.08.014
Datta, G., Garber, D. W., Chung, B. H., Chaddha, M., Dashti, N., Bradley, W. A., et al. (2001). Cationic domain 141-150 of apoE covalently linked to a class A amphipathic helix enhances atherogenic lipoprotein metabolism in vitro and in vivo. J. Lipid Res. 42, 959–966.
Datta, G., White, C. R., Dashti, N., Chaddha, M., Palgunachari, M. N., Gupta, H., et al. (2010). Anti-inflammatory and recycling properties of an apolipoprotein mimetic peptide, Ac-hE18A-NH(2). Atherosclerosis 208, 134–141. doi: 10.1016/j.atherosclerosis.2009.07.019
Davidson, W. S., Hazlett, T., Mantulin, W. W., and Jonas, A. (1996). The role of apolipoprotein AI domains in lipid binding. Proc. Natl. Acad. Sci. U.S.A. 93, 13605–13610. doi: 10.1073/pnas.93.24.13605
De Caterina, R., Sicari, R., Giannessi, D., Paggiaro, P. L., Paoletti, P., Lazzerini, G., et al. (1993). Macrophage-specific eicosanoid synthesis inhibition and lipocortin-1 induction by glucocorticoids. J. Appl. Physiol. (1985) 75, 2368–2375.
Deacon, C. F. (2011). Dipeptidyl peptidase-4 inhibitors in the treatment of type 2 diabetes: a comparative review. Diabetes Obes. Metab. 13, 7–18. doi: 10.1111/j.1463-1326.2010.01306.x
Deacon, C. F., Hughes, T. E., and Holst, J. J. (1998). Dipeptidyl peptidase IV inhibition potentiates the insulinotropic effect of glucagon-like peptide 1 in the anesthetized pig. Diabetes Metab. Res. Rev. 47, 764–769.
Di, L. (2015). Strategic approaches to optimizing peptide ADME properties. AAPS J. 17, 134–143. doi: 10.1208/s12248-014-9687-3
Di Bartolo, B. A., Nicholls, S. J., Bao, S., Rye, K. A., Heather, A. K., Barter, P. J., et al. (2011a). The apolipoprotein A-I mimetic peptide ETC-642 exhibits anti-inflammatory properties that are comparable to high density lipoproteins. Atherosclerosis 217, 395–400. doi: 10.1016/j.atherosclerosis.2011.04.001
Di Bartolo, B. A., Vanags, L. Z., Tan, J. T., Bao, S., Rye, K. A., Barter, P. J., et al. (2011b). The apolipoprotein A-I mimetic peptide, ETC-642, reduces chronic vascular inflammation in the rabbit. Lipids Health Dis. 10:224. doi: 10.1186/1476-511X-10-224
Doti, N., Scognamiglio, P. L., Madonna, S., Scarponi, C., Ruvo, M., Perretta, G., et al. (2012). New mimetic peptides of the kinase-inhibitory region (KIR) of SOCS1 through focused peptide libraries. Biochem. J. 443, 231–240. doi: 10.1042/BJ20111647
Drucker, D. J., and Nauck, M. A. (2006). The incretin system: glucagon-like peptide-1 receptor agonists and dipeptidyl peptidase-4 inhibitors in type 2 diabetes. Lancet 368, 1696–1705. doi: 10.1016/S0140-6736(06)69705-5
D’Souza, W., Stonik, J. A., Murphy, A., Demosky, S. J., Sethi, A. A., Moore, X. L., et al. (2010). Structure/function relationships of apolipoprotein a-I mimetic peptides: implications for antiatherogenic activities of high-density lipoprotein. Circ. Res. 107, 217–227. doi: 10.1161/CIRCRESAHA.110.216507
Eng, J., Kleinman, W. A., Singh, L., Singh, G., and Raufman, J. P. (1992). Isolation and characterization of exendin-4, an exendin-3 analogue, from Heloderma suspectum venom. Further evidence for an exendin receptor on dispersed acini from guinea pig pancreas. J. Biol. Chem. 267, 7402–7405.
Fabbrizzi, P., Menchi, G., Guarna, A., and Trabocchi, A. (2014). Use of click-chemistry in the development of peptidomimetic enzyme inhibitors. Curr. Med. Chem. 21, 1467–1477. doi: 10.2174/0929867321666131218093611
Fisher, E. A., Feig, J. E., Hewing, B., Hazen, S. L., and Smith, J. D. (2012). High-density lipoprotein function, dysfunction, and reverse cholesterol transport. Arterioscler. Thromb. Vasc. Biol. 32, 2813–2820. doi: 10.1161/ATVBAHA.112.300133
Flowers, L. O., Johnson, H. M., Mujtaba, M. G., Ellis, M. R., Haider, S. M., and Subramaniam, P. S. (2004). Characterization of a peptide inhibitor of Janus kinase 2 that mimics suppressor of cytokine signaling 1 function. J. Immunol. 172, 7510–7518. doi: 10.4049/jimmunol.172.12.7510
Fosgerau, K., and Hoffmann, T. (2015). Peptide therapeutics: current status and future directions. Drug Discov. Today 20, 122–128. doi: 10.1016/j.drudis.2014.10.003
Fuster, V., Moreno, P. R., Fayad, Z. A., Corti, R., and Badimon, J. J. (2005). Atherothrombosis and high-risk plaque: part I: evolving concepts. J. Am. Coll. Cardiol. 46, 937–954. doi: 10.1016/j.jacc.2005.03.074
Gao, M., Cheng, K., and Yin, H. (2015). Targeting protein-protein interfaces using macrocyclic peptides. Biopolymers 104, 310–316. doi: 10.1002/bip.22625
Garber, D. W., Venkatachalapathi, Y. V., Gupta, K. B., Ibdah, J., Phillips, M. C., Hazelrig, J. B., et al. (1992). Turnover of synthetic class A amphipathic peptide analogues of exchangeable apolipoproteins in rats. Correlation with physical properties. Arterioscler. Thromb. 12, 886–894. doi: 10.1161/01.ATV.12.8.886
Gaudreault, N., Kumar, N., Posada, J. M., Stephens, K. B., Reyes, de Mochel, N. S., et al. (2012). ApoE suppresses atherosclerosis by reducing lipid accumulation in circulating monocytes and the expression of inflammatory molecules on monocytes and vascular endothelium. Arterioscler. Thromb. Vasc. Biol. 32, 264–272. doi: 10.1161/ATVBAHA.111.238964
Gentilucci, L., Tolomelli, A., and Squassabia, F. (2006). Peptides and peptidomimetics in medicine, surgery and biotechnology. Curr. Med. Chem. 13, 2449–2466. doi: 10.2174/092986706777935041
Getting, S. J., Flower, R. J., and Perretti, M. (1997). Inhibition of neutrophil and monocyte recruitment by endogenous and exogenous lipocortin 1. Br. J. Pharmacol. 120, 1075–1082. doi: 10.1038/sj.bjp.0701029
Global Peptide Therapeutics Market (2016). Global Peptide Therapeutics Market to Reach US$23.7 bn by 2020, Rising Prevalence of Cancer to Drive Growth. Available at: http://www.transparencymarketresearch.com/pressrelease/global-peptide-therapeutics-market.htm
Goodwin, D., Simerska, P., and Toth, I. (2012). Peptides as therapeutics with enhanced bioactivity. Curr. Med. Chem. 19, 4451–4461. doi: 10.2174/092986712803251548
Gordon, S. M., Hofmann, S., Askew, D. S., and Davidson, W. S. (2011). High density lipoprotein: it’s not just about lipid transport anymore. Trends Endocrinol. Metab. 22, 9–15. doi: 10.1016/j.tem.2010.10.001
Gordon, S. M., Li, H., Zhu, X., Shah, A. S., Lu, L. J., and Davidson, W. S. (2015). A comparison of the mouse and human lipoproteome: suitability of the mouse model for studies of human lipoproteins. J. Proteome Res. 14, 2686–2695. doi: 10.1021/acs.jproteome.5b00213
Gupta, H., White, C. R., Handattu, S., Garber, D. W., Datta, G., Chaddha, M., et al. (2005). Apolipoprotein E mimetic Peptide dramatically lowers plasma cholesterol and restores endothelial function in watanabe heritable hyperlipidemic rabbits. Circulation 111, 3112–3118. doi: 10.1161/CIRCULATIONAHA.104.497107
Hatters, D. M., Peters-Libeu, C. A., and Weisgraber, K. H. (2006). Apolipoprotein E structure: insights into function. Trends Biochem. Sci. 31, 445–454. doi: 10.1016/j.tibs.2006.06.008
Hecht, I., Rong, J., Sampaio, A. L., Hermesh, C., Rutledge, C., Shemesh, R., et al. (2009). A novel peptide agonist of formyl-peptide receptor-like 1 (ALX) displays anti-inflammatory and cardioprotective effects. J. Pharmacol. Exp. Ther. 328, 426–434. doi: 10.1124/jpet.108.145821
Iqbal, A. J., Barrett, T. J., Taylor, L., McNeill, E., Manmadhan, A., Recio, C., et al. (2016). Acute exposure to apolipoprotein A1 inhibits macrophage chemotaxis in vitro and monocyte recruitment in vivo. Elife 5:e15190. doi: 10.7554/eLife.15190
Irwin, N., and Flatt, P. R. (2015). New perspectives on exploitation of incretin peptides for the treatment of diabetes and related disorders. World J. Diabetes 6, 1285–1295. doi: 10.4239/wjd.v6.i15.1285
Iwata, A., Miura, S., Zhang, B., Imaizumi, S., Uehara, Y., Shiomi, M., et al. (2011). Antiatherogenic effects of newly developed apolipoprotein A-I mimetic peptide/phospholipid complexes against aortic plaque burden in Watanabe-heritable hyperlipidemic rabbits. Atherosclerosis 218, 300–307. doi: 10.1016/j.atherosclerosis.2011.05.029
Jimenez-Solem, E., Rasmussen, M. H., Christensen, M., and Knop, F. K. (2010). Dulaglutide, a long-acting GLP-1 analog fused with an Fc antibody fragment for the potential treatment of type 2 diabetes. Curr. Opin. Mol. Ther. 12, 790–797.
Juhl, C. B., Hollingdal, M., Sturis, J., Jakobsen, G., Agersø, H., Veldhuis, J., et al. (2002). Bedtime administration of NN2211, a long-acting GLP-1 derivative, substantially reduces fasting and postprandial glycemia in type 2 diabetes. Diabetes Metab. Res. Rev. 51, 424–429.
Kalra, S., Baruah, M. P., Sahay, R. K., Unnikrishnan, A. G., Uppal, S., and Adetunji, O. (2016). Glucagon-like peptide-1 receptor agonists in the treatment of type 2 diabetes: past, present, and future. Indian J. Endocrinol. Metab. 20, 254–267. doi: 10.4103/2230-8210.176351
Kaspar, A. A., and Reichert, J. M. (2013). Future directions for peptide therapeutics development. Drug Discov. Today 18, 807–817. doi: 10.1016/j.drudis.2013.05.011
La, M., D’Amico, M., Bandiera, S., Di Filippo, C., Oliani, S. M., Gavins, F. N., et al. (2001). Annexin 1 peptides protect against experimental myocardial ischemia-reperfusion: analysis of their mechanism of action. FASEB J. 15, 2247–2256. doi: 10.1096/fj.01-0196com
Lawrence, T., Willoughby, D. A., and Gilroy, D. W. (2002). Anti-inflammatory lipid mediators and insights into the resolution of inflammation. Nat. Rev. Immunol. 2, 787–795. doi: 10.1038/nri915
Lax, R., and Meenan, C. (2012). Challenges for therapeutic peptides part 1: on the inside, looking out. Innov. Pharm. Technol. 42, 54–56.
LeBert, D. C., and Huttenlocher, A. (2014). Inflammation and wound repair. Semin. Immunol. 26, 315–320. doi: 10.1016/j.smim.2014.04.007
Leening, M. J., Cook, N. R., and Ridker, P. M. (2016). Should we reconsider the role of age in treatment allocation for primary prevention of cardiovascular disease? Eur. Heart J. ehw287. doi: 10.1093/eurheartj/ehw287
Li, X., Chyu, K. Y., Faria Neto, J. R., Yano, J., Nathwani, N., Ferreira, C., et al. (2004). Differential effects of apolipoprotein A-I-mimetic peptide on evolving and established atherosclerosis in apolipoprotein E-null mice. Circulation 110, 1701–1705. doi: 10.1161/01.CIR.0000142857.79401.69
Liang, X., He, M., Chen, T., Liu, Y., Tian, Y. L., Wu, Y. L., et al. (2013). Multiple roles of SOCS proteins: differential expression of SOCS1 and SOCS3 in atherosclerosis. Int. J. Mol. Med. 31, 1066–1074. doi: 10.3892/ijmm.2013.1323
Libby, P., Ridker, P. M., and Hansson, G. K. (2011). Progress and challenges in translating the biology of atherosclerosis. Nature 473, 317–325. doi: 10.1038/nature10146
Linossi, E. M., Babon, J. J., Hilton, D. J., and Nicholson, S. E. (2013). Suppression of cytokine signaling: the SOCS perspective. Cytokine Growth Factor Rev. 24, 241–248. doi: 10.1016/j.cytogfr.2013.03.005
Liskamp, R. M., Rijkers, D. T., Kruijtzer, J. A., and Kemmink, J. (2011). Peptides and proteins as a continuing exciting source of inspiration for peptidomimetics. Chembiochem 12, 1626–1653. doi: 10.1002/cbic.201000717
Madonna, S., Scarponi, C., Doti, N., Carbone, T., Cavani, A., Scognamiglio, P. L., et al. (2013). Therapeutical potential of a peptide mimicking the SOCS1 kinase inhibitory region in skin immune responses. Eur. J. Immunol. 43, 1883–1895. doi: 10.1002/eji.201343370
Madsbad, S., Kielgast, U., Asmar, M., Deacon, C. F., Torekov, S. S., and Holst, J. J. (2011). An overview of once-weekly glucagon-like peptide-1 receptor agonists–available efficacy and safety data and perspectives for the future. Diabetes Obes. Metab. 13, 394–407. doi: 10.1111/j.1463-1326.2011.01357.x
Mahley, R. W., Huang, Y., and Weisgraber, K. H. (2006). Putting cholesterol in its place: apoE and reverse cholesterol transport. J. Clin. Invest. 116, 1226–1229. doi: 10.1172/JCI28632
Manduteanu, I., and Simionescu, M. (2012). Inflammation in atherosclerosis: a cause or a result of vascular disorders? J. Cell. Mol. Med. 16, 1978–1990. doi: 10.1111/j.1582-4934.2012.01552.x
Mangaraj, M., Nanda, R., and Panda, S. (2016). Apolipoprotein A-I: a molecule of diverse function. Indian J. Clin. Biochem. 31, 253–259. doi: 10.1007/s12291-015-0513-1
Marrero, M. B. (2005). Introduction to JAK/STAT signaling and the vasculature. Vascul. Pharmacol. 43, 307–309. doi: 10.1016/j.vph.2005.09.002
Mason, J. M. (2010). Design and development of peptides and peptide mimetics as antagonists for therapeutic intervention. Future Med. Chem. 2, 1813–1822. doi: 10.4155/fmc.10.259
McGill, J. B., Ahn, D., Edelman, S. V., Kilpatrick, C. R., and Santos Cavaiola, T. (2016). Making insulin accessible: does inhaled insulin fill an unmet need? Adv. Ther. 33, 1267–1278. doi: 10.1007/s12325-016-0370-1
Meier, J. J. (2012). GLP-1 receptor agonists for individualized treatment of type 2 diabetes mellitus. Nat. Rev. Endocrinol. 8, 728–742. doi: 10.1038/nrendo.2012.140
Miklossy, G., Hilliard, T. S., and Turkson, J. (2013). Therapeutic modulators of STAT signalling for human diseases. Nat. Rev. Drug Discov. 12, 611–629. doi: 10.1038/nrd4088
Mittermayer, F., Caveney, E., De Oliveira, C., Gourgiotis, L., Puri, M., Tai, L.-J., et al. (2015). Addressing unmet medical needs in type 2 diabetes: a narrative review of drugs under development. Curr. Diabetes Rev. 11, 17–31. doi: 10.2174/1573399810666141224121927
Moore, R. E., Kawashiri, M. A., Kitajima, K., Secreto, A., Millar, J. S., Pratico, D., et al. (2003). Apolipoprotein A-I deficiency results in markedly increased atherosclerosis in mice lacking the LDL receptor. Arterioscler. Thromb. Vasc. Biol. 23, 1914–1920. doi: 10.1161/01.ATV.0000092328.66882.F5
Mujtaba, M. G., Flowers, L. O., Patel, C. B., Patel, R. A., Haider, M. I., and Johnson, H. M. (2005). Treatment of mice with the suppressor of cytokine signaling-1 mimetic peptide, tyrosine kinase inhibitor peptide, prevents development of the acute form of experimental allergic encephalomyelitis and induces stable remission in the chronic relapsing/remitting form. J. Immunol. 175, 5077–5086.
Nauck, M. A., Petrie, J. R., Sesti, G., Mannucci, E., Courrèges, J. P., Lindegaard, M. L., et al. (2016). A phase 2, randomized, dose-finding study of the novel once-weekly human GLP-1 analog, semaglutide, compared with placebo and open-label liraglutide in patients with type 2 diabetes. Diabetes Care 39, 231–241. doi: 10.2337/dc15-0165
Navab, M., Anantharamaiah, G. M., Hama, S., Garber, D. W., Chaddha, M., Hough, G., et al. (2002). Oral administration of an Apo A-I mimetic Peptide synthesized from D-amino acids dramatically reduces atherosclerosis in mice independent of plasma cholesterol. Circulation 105, 290–292. doi: 10.1161/hc0302.103711
Navab, M., Anantharamaiah, G. M., Reddy, S. T., Hama, S., Hough, G., Grijalva, V. R., et al. (2005). Apolipoprotein A-I mimetic peptides. Arterioscler. Thromb. Vasc. Biol. 25, 1325–1331. doi: 10.1161/01.ATV.0000165694.39518.95
Navab, M., Hough, G., Buga, G. M., Su, F., Wagner, A. C., Meriwether, D., et al. (2013). Transgenic 6F tomatoes act on the small intestine to prevent systemic inflammation and dyslipidemia caused by Western diet and intestinally derived lysophosphatidic acid. J. Lipid Res. 54, 3403–3418. doi: 10.1194/jlr.M042051
Navickas, R., Gal, D., Laucevičius, A., Taparauskaitė, A., Zdanytė, M., and Holvoet, P. (2016). Identifying circulating microRNAs as biomarkers of cardiovascular disease: a systematic review. Cardiovasc. Res. 111, 322–337. doi: 10.1093/cvr/cvw174
Neumiller, J. J. (2015). Incretin-based therapies. Med. Clin. North Am. 99, 107–129. doi: 10.1016/j.mcna.2014.08.013
Pellicori, P., and Costanzo, P. (2015). [Betablockers in patients with heart failure and atrial fibrillation]. G. Ital. Cardiol. (Rome) 16, 613–616. doi: 10.1714/2066.22428
Peptide Therapeutics Market (2015). Peptide Therapeutics Market (by Applications, by Route of Administration, and by Marketing Status) – Global Industry Analysis, Size, Share, Growth, Trends and Forecast 2014 – 2020. Available at: http://www.transparencymarketresearch.com/peptide-therapeutics-market.html
Perretti, M., and Gavins, F. N. (2003). Annexin 1: an endogenous anti-inflammatory protein. News Physiol. Sci. 18, 60–64.
Pham, T., Kodvawala, A., and Hui, D. Y. (2005). The receptor binding domain of apolipoprotein E is responsible for its antioxidant activity. Biochemistry 44, 7577–7582. doi: 10.1021/bi0472696
Pirlamarla, P., and Bond, R. M. (2016). FDA labeling of NSAIDs: review of nonsteroidal anti-inflammatory drugs in cardiovascular disease. Trends Cardiovasc. Med. 26, 675–680. doi: 10.1016/j.tcm.2016.04.011
Podrez, E. A. (2010). Anti-oxidant properties of high-density lipoprotein and atherosclerosis. Clin. Exp. Pharmacol. Physiol. 37, 719–725. doi: 10.1111/j.1440-1681.2010.05380.x
Puttagunta, A. L., and Toth, E. L. (1998). Insulin lispro (Humalog), the first marketed insulin analogue: indications, contraindications and need for further study. CMAJ 158, 506–511.
Qin, C., Yang, Y. H., May, L., Gao, X., Stewart, A. G., Tu, Y., et al. (2015). Cardioprotective potential of annexin-A1 mimetics in myocardial infarction. Pharmacol. Ther. 148, 47–65. doi: 10.1016/j.pharmthera.2014.11.012
Qin, L., Huang, Q., Zhang, H., Liu, R., Tellides, G., Min, W., et al. (2014). SOCS1 prevents graft arteriosclerosis by preserving endothelial cell function. J. Am. Coll. Cardiol. 63, 21–29. doi: 10.1016/j.jacc.2013.08.694
Recio, C., Lazaro, I., Oguiza, A., Lopez-Sanz, L., Bernal, S., Blanco, J., et al. (2016). Suppressor of cytokine signaling-1 peptidomimetic limits progression of diabetic nephropathy. J. Am. Soc. Nephrol. doi: 10.1681/ASN.2016020237
Recio, C., Oguiza, A., Lazaro, I., Mallavia, B., Egido, J., and Gomez-Guerrero, C. (2014). Suppressor of cytokine signaling 1-derived peptide inhibits Janus kinase/signal transducers and activators of transcription pathway and improves inflammation and atherosclerosis in diabetic mice. Arterioscler. Thromb. Vasc. Biol. 34, 1953–1960. doi: 10.1161/ATVBAHA.114.304144
Remaley, A. T., Thomas, F., Stonik, J. A., Demosky, S. J., Bark, S. E., Neufeld, E. B., et al. (2003). Synthetic amphipathic helical peptides promote lipid efflux from cells by an ABCA1-dependent and an ABCA1-independent pathway. J. Lipid Res. 44, 828–836. doi: 10.1194/jlr.M200475-JLR200
Rosenbaum, M. A., Chaudhuri, P., Abelson, B., Cross, B. N., and Graham, L. M. (2015). Apolipoprotein A-I mimetic peptide reverses impaired arterial healing after injury by reducing oxidative stress. Atherosclerosis 241, 709–715. doi: 10.1016/j.atherosclerosis.2015.06.018
Rosenstock, J., Reusch, J., Bush, M., Yang, F., Stewart, M., and Group, A. S. (2009). Potential of albiglutide, a long-acting GLP-1 receptor agonist, in type 2 diabetes: a randomized controlled trial exploring weekly, biweekly, and monthly dosing. Diabetes Care 32, 1880–1886. doi: 10.2337/dc09-0366
Russell, S. (2013). Incretin-based therapies for type 2 diabetes mellitus: a review of direct comparisons of efficacy, safety and patient satisfaction. Int. J. Clin. Pharm. 35, 159–172. doi: 10.1007/s11096-012-9729-9
Schett, G., Elewaut, D., McInnes, I. B., Dayer, J. M., and Neurath, M. F. (2013). How cytokine networks fuel inflammation: toward a cytokine-based disease taxonomy. Nat. Med. 19, 822–824. doi: 10.1038/nm.3260
Shaji, J., and Patole, V. (2008). Protein and Peptide drug delivery: oral approaches. Indian J. Pharm. Sci. 70, 269–277. doi: 10.4103/0250-474X.42967
Sharifov, O. F., Nayyar, G., Garber, D. W., Handattu, S. P., Mishra, V. K., Goldberg, D., et al. (2011). Apolipoprotein E mimetics and cholesterol-lowering properties. Am. J. Cardiovasc. Drugs 11, 371–381. doi: 10.2165/11594190-000000000-00000
Smith, J. D. (2010). Apolipoprotein A-I and its mimetics for the treatment of atherosclerosis. Curr. Opin. Investig. Drugs 11, 989–996.
Snell-Bergeon, J. K., and Wadwa, R. P. (2012). Hypoglycemia, diabetes, and cardiovascular disease. Diabetes Technol. Ther. 14(Suppl. 1), S51–S58. doi: 10.1089/dia.2012.0031
Sohma, Y., Sasaki, M., Hayashi, Y., Kimura, T., and Kiso, Y. (2004). Novel and efficient synthesis of difficult sequence-containing peptides through O-N intramolecular acyl migration reaction of O-acyl isopeptides. Chem. Commun. (Camb.) 1, 124–125. doi: 10.1039/b312129a
Stein, E. A., and Raal, F. J. (2015). Lipid-lowering drug therapy for CVD prevention: looking into the future. Curr. Cardiol. Rep. 17:104. doi: 10.1007/s11886-015-0659-8
Stoekenbroek, R. M., Stroes, E. S., and Hovingh, G. K. (2015). ApoA-I mimetics. Handb. Exp. Pharmacol. 224, 631–648. doi: 10.1007/978-3-319-09665-0_21
Suc, I., Escargueil-Blanc, I., Troly, M., Salvayre, R., and Nègre-Salvayre, A. (1997). HDL and ApoA prevent cell death of endothelial cells induced by oxidized LDL. Arterioscler. Thromb. Vasc. Biol. 17, 2158–2166. doi: 10.1161/01.ATV.17.10.2158
Tabet, F., Remaley, A. T., Segaliny, A. I., Millet, J., Yan, L., Nakhla, S., et al. (2010). The 5A apolipoprotein A-I mimetic peptide displays antiinflammatory and antioxidant properties in vivo and in vitro. Arterioscler. Thromb. Vasc. Biol. 30, 246–252. doi: 10.1161/ATVBAHA.109.200196
Tajiri, K., Imanaka-Yoshida, K., Matsubara, A., Tsujimura, Y., Hiroe, M., Naka, T., et al. (2012). Suppressor of cytokine signaling 1 DNA administration inhibits inflammatory and pathogenic responses in autoimmune myocarditis. J. Immunol. 189, 2043–2053. doi: 10.4049/jimmunol.1103610
Trengove, M. C., and Ward, A. C. (2013). SOCS proteins in development and disease. Am. J. Clin. Exp. Immunol. 2, 1–29.
Uehara, Y., Ando, S., Yahiro, E., Oniki, K., Ayaori, M., Abe, S., et al. (2013). FAMP, a novel apoA-I mimetic peptide, suppresses aortic plaque formation through promotion of biological HDL function in ApoE-deficient mice. J. Am. Heart Assoc. 2:e000048. doi: 10.1161/JAHA.113.000048
Uehara, Y., Chiesa, G., and Saku, K. (2015). High-density lipoprotein-targeted therapy and apolipoprotein A-I mimetic peptides. Circ. J. 79, 2523–2528. doi: 10.1253/circj.CJ-15-0960
Uhlig, T., Kyprianou, T., Martinelli, F. G., Oppici, C. A., Heiligers, D., Hills, D., et al. (2014). The emergence of peptides in the pharmaceutical business: from exploration to exploitation. EuPA Open Proteomics 4, 58–69. doi: 10.1016/j.euprot.2014.05.003
Vagner, J., Qu, H., and Hruby, V. J. (2008). Peptidomimetics, a synthetic tool of drug discovery. Curr. Opin. Chem. Biol. 12, 292–296. doi: 10.1016/j.cbpa.2008.03.009
Van Lenten, B. J., Wagner, A. C., Jung, C. L., Ruchala, P., Waring, A. J., Lehrer, R. I., et al. (2008). Anti-inflammatory apoA-I-mimetic peptides bind oxidized lipids with much higher affinity than human apoA-I. J. Lipid Res. 49, 2302–2311. doi: 10.1194/jlr.M800075-JLR200
Venkatachalapathi, Y. V., Phillips, M. C., Epand, R. M., Epand, R. F., Tytler, E. M., Segrest, J. P., et al. (1993). Effect of end group blockage on the properties of a class A amphipathic helical peptide. Proteins 15, 349–359. doi: 10.1002/prot.340150403
Vlieghe, P., Lisowski, V., Martinez, J., and Khrestchatisky, M. (2010). Synthetic therapeutic peptides: science and market. Drug Discov. Today 15, 40–56. doi: 10.1016/j.drudis.2009.10.009
Watson, C. E., Weissbach, N., Kjems, L., Ayalasomayajula, S., Zhang, Y., Chang, I., et al. (2011). Treatment of patients with cardiovascular disease with L-4F, an apo-A1 mimetic, did not improve select biomarkers of HDL function. J. Lipid Res. 52, 361–373. doi: 10.1194/jlr.M011098
White, C. R., Garber, D. W., and Anantharamaiah, G. M. (2014). Anti-inflammatory and cholesterol-reducing properties of apolipoprotein mimetics: a review. J. Lipid Res. 55, 2007–2021. doi: 10.1194/jlr.R051367
Wroge, J., and Williams, N. T. (2016). Glucagon-like peptide-1 (GLP-1) receptor agonists in cardiac disorders. Ann. Pharmacother. 50, 1041–1050. doi: 10.1177/1060028016663218
Yoshimura, A., Naka, T., and Kubo, M. (2007). SOCS proteins, cytokine signalling and immune regulation. Nat. Rev. Immunol. 7, 454–465. doi: 10.1038/nri2093
Keywords: cardiovascular disease, cardiovascular system, inflammation, peptides, peptidomimetics
Citation: Recio C, Maione F, Iqbal AJ, Mascolo N and De Feo V (2017) The Potential Therapeutic Application of Peptides and Peptidomimetics in Cardiovascular Disease. Front. Pharmacol. 7:526. doi: 10.3389/fphar.2016.00526
Received: 11 October 2016; Accepted: 19 December 2016;
Published: 06 January 2017.
Edited by:
Concepción Peiró, Universidad Autonoma de Madrid, SpainReviewed by:
Pui Man Maggie Hoi, University of Macau, MacauCopyright © 2017 Recio, Maione, Iqbal, Mascolo and De Feo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Francesco Maione, ZnJhbmNlc2NvLm1haW9uZUB1bmluYS5pdA== Vincenzo De Feo, ZGVmZW9AdW5pc2EuaXQ=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.