 Jie Zhu
Jie Zhu Mingxin He
Mingxin He Simin Li1
Simin Li1 Xiaochen Xiang
Xiaochen Xiang Zhi Guo
Zhi Guo Qiang Wang
Qiang Wang- 1Institute of Infection, Immunology and Tumor Microenvironment, Wuchang Hospital Affiliated to Wuhan University of Science and Technology, Medical College, Wuhan University of Science and Technology, Wuhan, China
- 2Department of Hematology, Huazhong University of Science and Technology Union Shenzhen Hospital, Shenzhen, China
The first 1, 000 days of life, from the fetal stage of a woman's pregnancy to 2 years of age after the baby is born, is a critical period for microbial colonization of the body and development of the immune system. The immune system and microbiota exhibit great plasticity at this stage and play a crucial role in subsequent development and future health. Two-way communication and interaction between immune system and microbiota is helpful to maintain human microecological balance and immune homeostasis. Currently, there is a growing interest in the important role of the microbiota in the newborn, and it is believed that the absence or dysbiosis of human commensal microbiota early in life can have lasting health consequences. Thus, this paper summarizes research advances in the establishment of the oral and intestinal microbiome and immune system in early life, emphasizing the substantial impact of microbiota diversity in the prenatal and early postnatal periods, and summarizes that maternal microbes, mode of delivery, feeding practices, antibiotics, probiotics, and the environment shape the oral and intestinal microbiota of infants in the first 1, 000 days of life and their association with the immune system.
1 Introduction
The immune system in early life does not develop in isolation, but is strongly influenced by maternal and autoantigens, commensal bacteria, and pathogens (1). The microbiota establishes or maintains homeostasis when the balance of immune system tolerance to commensal, conditionally pathogenic and harmful bacteria is established (2); microbiota regulate the function and activity of immune cells, stimulate mucosal specificity and activate the immune system and induce the development and maturation of immune cells and organs, with health-promoting and beneficial immunomodulatory properties (3). Therefore, the two-way communication between immune system and microbiota is helpful to maintain the microecological balance and immune homeostasis of human body. Indeed, microbe-dependent modulation of the host immune system has a limited window of opportunity, and there is no way to make up for the missed window early in life in adulthood, which may even promote inflammation or increase the risk of disease progression, making it particularly important to focus on the establishment of the microbiota and the immune system early in life.
Streptococcus spp. are the first bacteria to colonize the infant's mouth and are known as “early colonizers” (4). Early colonization affects subsequent microbial colonization due to the occupation of binding sites, efficient use of nutrients, production of antimicrobial agents and end products, and changes in the environment (5). Therefore, early colonization is very important for the development of microbiota, and may have long-term consequences (6). Facultative anaerobic bacteria are early colonizers in the human intestinal and are distinctly dominant in the first few weeks of life, such as Enterobacterales, Enterococci and Staphylococci. Subsequently, the dominant microbiota gradually changed to anaerobic bacteria, such as C. leptum subgroup, Bifidobacterium and B. fragilis (7). With the introduction of solid food, intestinal microbiota are dominated by Firmicutes and Bacteroidetes (8). Acquisition of early colonizers may be altered by a variety of maternal and infant factors (Figure 1), which may lead to differences in oral and intestinal microbiota development (9, 10).
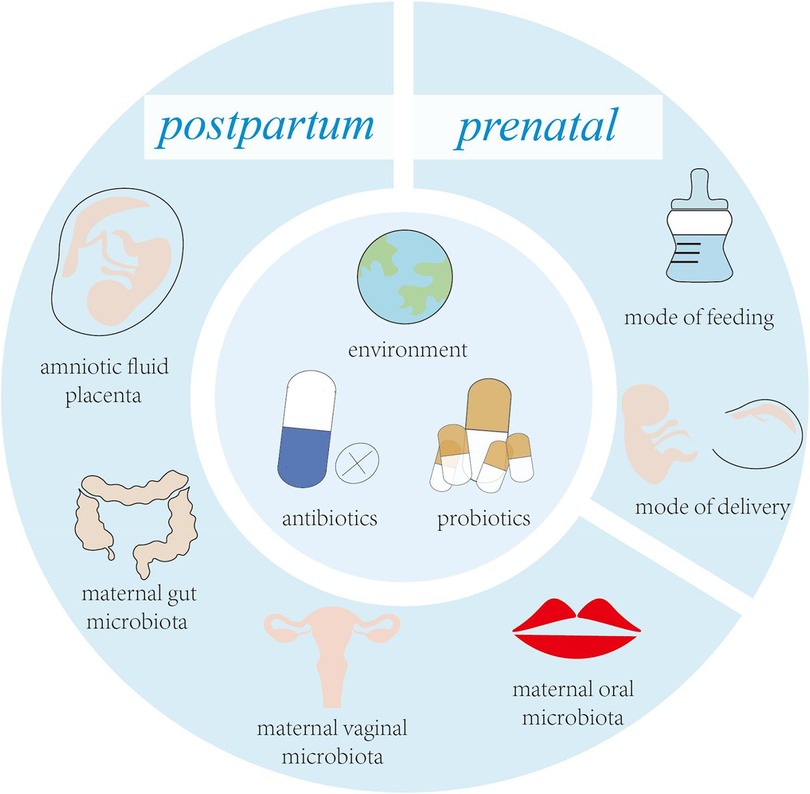
Figure 1. Key factors influencing early colonizers before and after birth.
Disruption of early oral colonization may affect the progression of oral and systemic disease in children. The oral microbiota has shown significant correlations with systemic disorders such as weight gain, rheumatoid arthritis (RA), and autism (11–13). In addition, the oral cavity is the window to the gut, and the oral microbiota affects the colonization of the gut microbiota to a certain extent, and it has been shown that the oral microbiota shows its relevance and possibilities as a substitute for the gut microbiota (14). Intestinal microbiota have the ability to generate resistance to pathogens and enhance the resistance of the immune system through colonization of mucosa surface and production of different antimicrobial substances, which play an important role in maintaining normal gut physiology and health. Thus, the oral and intestinal microbiota has a unique role in promoting immune system development and modulating host defence (15, 16). The aim of this review is to provide a summary of the latest research advances in the establishment of the oral and gut microbiome and the immune system during the first 1,000 days of life. At the same time, it is emphasized that maternal factors, mode of delivery, breastfeeding, antibiotics, host factors, and living environment reciprocally shape the oral and intestinal microbiota of infants during the first 1,000 days of life and their association with the immune system.
2 Prenatal factors affecting the development of the fetal microbiota and immune system
2.1 Effects of maternal microbiota on the fetus
For many years, scientists and doctors have believed that the fetus grows and develops in a sterile environment. Recent research data questioned this common understanding. New research found that there are microbiota in meconium, placenta and amniotic fluid, and put forward the view that uterus is not a sterile organ (17). The most common bacteria in meconium samples are Staphylococcus, followed by Enterobacteriaceae, Enterococcus, Lactobacillus and Bifidobacterium (18–20). However, Rehbinder et al. suggested that the presence of foetal faecal microbiota was the result of DNA contamination from laboratory reagents or acquired during labour (21). Stinson et al. combined PacBio single-molecule real-time (SMRT) sequencing technology with a workflow designed to reduce contamination and showed that the foetal faecal and amniotic fluid microbiomes were present beyond background contamination levels (22). Meanwhile, Aagaard et al. performed the first macrogenomic characterisation of the placental microbiome using whole-genome birdshot sequencing and 454 pyrophosphate sequencing, and found that the placenta contains a unique microbiome composed of a non-pathogenic gut microbiota dominated by Proteobacteria, and composed of non-pathogenic intestinal microbiota of Firmicutes, Bacteroidetes and Fusobacteria phyla. At the same time, it is proposed that placental microorganisms are most similar to maternal oral microorganisms and that their composition may also influence pregnancy outcome (23). The microbiota in amniotic fluid is similar to that in placenta, and an imbalance in its microbial composition may lead to chorioamnionitis. Previous studies have shown that the microbial status in amniotic fluid can be used to predict the occurrence of premature birth (24). Preterm birth and delivery are associated with bacterial colonization of the amniotic cavity and fetal membranes, and the amount of microbial DNA in amniotic fluid correlates with increased levels of leukocytes and inflammatory mediators, suggesting that a higher microbial load can lead to inflammation, which may be an endocrine mechanism that triggers preterm labor (25). These results suggest that colonization of the fetal oral cavity and intestine by the microbiota may have occurred during pregnancy.
Vertical transfer of bacterial species between mother and baby occurs before the baby is born, suggesting that the physiological state of the mother can influence the microecology of the fetus. Maternal microbiota is a key source of microbiota during initial colonization of the fetal oral and gut microbiota. Maternal factors that shape the fetal microbiota are mainly the maternal oral, intestinal and vaginal microbiota (26). Interestingly, the composition of the maternal oral, intestinal and vaginal microbiota changes dramatically during pregnancy (Figure 2), which may be the source of the close connection between the maternal microbiota and the fetal and infant microbiota. The total number of oral microbiota was higher than that of non-pregnant bacteria at different stages of pregnancy. In the early and second trimester of pregnancy, the abundance of Porphyromonas gingivalis and Aggregatibacter actinomycetemcomitans increased significantly, Candida levels were significantly higher in mid and late pregnancy, and the abundance of Actinomyces was even more significantly higher throughout pregnancy, changes that may be due to changes in the overall immune status during pregnancy (27–30). From the first trimester to the third trimester, changes in the maternal gut microbiota are mainly manifested by increased abundance of Actinomycetes and Proteobacteria, a significant decrease in the levels of butyrate-producing Faecalibacterium with anti-inflammatory activity in the third trimester, and a decrease in α-diversity at the individual level. In addition, in the third trimester of pregnancy, the β-diversity of the maternal microbiota increases among individuals, reflecting the immune manifestations of chronic inflammation (29, 31, 32). Alterations in the maternal gut microbiota during pregnancy play a positive role in the regulation of metabolic responses in the fetus (33). In view of the influence of intestinal microbiota on fetal immune response, the adaptation and regulation of maternal immune response during pregnancy is a necessary condition for fetal tissue cells to be accepted by the mother and avoid maternal immune rejection, while maintaining their own immune defense mechanism to ensure the survival of the mother and fetus (34). The vaginal microbiota also changes significantly during pregnancy, with a decrease in overall diversity and number of organisms in the first trimester, but an increase in the stability of the microbiota composition, as well as an enrichment of Lactobacillus, which leads to a decrease in vaginal pH and an increase in secretions (35). Studies have shown that the increase of Lactobacillus has a direct or indirect protective effect on maintaining the stability of the vaginal microbiota during pregnancy and preventing ascending infections (36). In addition, a decrease in the proportion of Lactobacillus is associated with adverse pregnancy outcomes, such as preterm birth, miscarriage, etc (37).
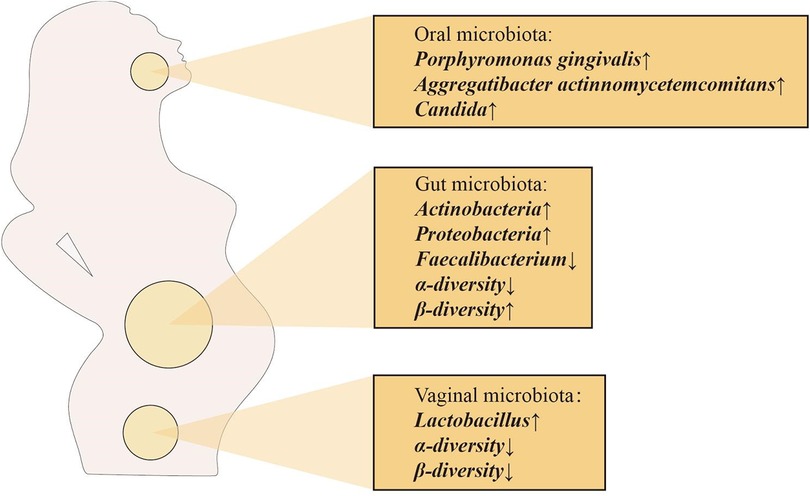
Figure 2. Major changes in the maternal oral, intestinal and vaginal microbiota during pregnancy.
The above changes in the mother's own microbiota during pregnancy can reach the placenta and affect the fetal microbiota through different routes of transmission, with blood transmission being the most direct and effective route. Scientists hypothesize that the pathway of transmission of the microbiota from mother to fetus is similar to the potential pathway for intrauterine infections (38). Some of the maternal immune cells are thought to be transporter cells for placenta-acquired microbiota (39). For example, dendritic cells (DCs) can cross the gap between intestinal epithelial cells and capture bacteria directly from the intestinal lumen. However, DCs do not kill the captured microorganisms, which can be transmitted through the dendritic cells through the bloodstream to other sites, such as the placenta and amniotic fluid, and through the umbilical cord to the fetus (40). Intrauterine microbiota colonization may occur via the ascending route through the genitourinary tracts such as the urinary tract, cervix, and vagina, as well as the hematogenous route through the placenta after translocation from the digestive tracts such as the oral cavity and intestines (41). Microbiota associated with the gastrointestinal tract such as Bacteroides and Proteus as well as microbiota associated with the vaginal microbiota such as Lactobacillus iners, L. crispatus, and Prevotella amnii were found in endometrial samples, so Verstraelen et al. concluded that the endometrium is colonized by both vaginal and intestinal microbiota (42). In addition, transplacental passage of these colonized microorganisms or their metabolites may result in increased expression of microbe-associated molecular pattern (MAMP) recognizing receptors, thymic regulatory T (Treg) cell promotion, and the establishment of dendritic cell (DC) networks in the gut, lung, and other tissues, as well as accelerated transitions from TH2 to TH1, interferon-regulated transcription factor 7 (RF7), and TH-17 (43).
2.2 Effects of antenatal antibiotic use on the fetus
Current clinical evidence suggests that antibiotics used by pregnant women may cross the placental barrier and reach the fetus, directly or indirectly disrupting the oral and intestinal microbiota of the fetus (Table 1) (48). By comparing fecal samples from PAT (prenatal antibiotic treatment) and PAF (prenatal antibiotic-free) preterm infants on days 7 and 14 after birth, Zou et al. found that prenatal antibiotic exposure resulted in a decrease in the intestinal microbiota of preterm infants in Bacteroides spp. (PAF 9.11%, PAT 2.93%) and an increase in Escherichia-Shigella (d7, PAF 27.35%, PAT 43.35%; d14, PAF 29.47%, PAT 40.36%) increases (49). A meta-analysis showed a significant decrease in the relative abundance of Actinobacteria and an increase in the abundance of Firmicutes and Proteobacteria in infants whose mothers were treated with antibiotics before or during labor and delivery, compared to infants who were not exposed to antibiotics (50). In the current study, Marilen et al. used a mouse pregnancy model to explore the effects of antibiotic treatment on maternal immunity, and the microbial intervention strategy chosen was shown to affect offspring immunity (51). Using a mouse model, Xu et al. showed that prenatal antibiotic exposure led to a reduction in microbiota-derived butyrate production, which in turn enhanced neonatal ILC2 activation by downregulating IFN1 signaling (52). At the same time, antibiotic use in pregnant and lactating females reduces antiviral-specific immune responses in pups, suggesting that antibiotics cause extensive immune damage in the offspring (53). A large number of studies have shown that maternal antibiotic use increases the risk of obesity, otitis media, asthma and other diseases after birth (54–56). The above results suggest that prenatal antibiotic use can affect the baby's oral and intestinal microbiota, which in turn affects the development of the baby's immune system.

Table 1. Effects of maternal prenatal or perinatal antibiotic use on the infant microbiota.
2.3 Effects of prenatal environmental exposure on the fetus
Exposure to environmental pollutants and bacteria early in life can affect the establishment of the fetal oral and intestinal microbiota and the development of the immune system. Some recent studies have shown that maternal inhalation of PM2.5 may induce oxidative stress, inflammatory responses, endocrine disruption and epigenetic changes, thus indirectly affecting normal fetal development (52). Inhaled PM2.5 may also penetrate the alveolar epithelial barrier and subsequently enter the circulation and be deposited in the placenta, where these particles may directly damage the structure and function of the placenta and further affect fetal growth (57). Tao et al. exposed pregnant mice to filtered air (FA) or concentrated ambient PM2.5 (CAP) and found that CAP exposure altered the metabolome and disrupted metabolic pathways (e.g., amino acid, lipid pathways) in maternal serum and placenta (58). Prenatal maternal exposure to air pollutants, especially in the first and third trimesters, affects the distribution of white blood cells in the fetus, and may also lead to an imbalance in fetal T helper(Th) cell subsets, increasing the risk of allergic reactions in children (59, 60). Loss et al. assessed mRNA expression of Toll-like receptors (TLR) 1 through TLR9 and CD14 in cord blood and found that gene expression of innate immunity receptors was higher overall in the cord blood of newborns from a rural setting (P for multifactorial multivariate ANOVA = .041), particularly so for TLR7 [adjusted geometric means ratio (aGMR), 1.15; 95% CI, 1.02–1.30] and TLR8 (aGMR, 1.15; 95% CI, 1.04–1.26) (61). In addition, prenatal exposure to farms can modulate the immune system of offspring, and from umbilical cord blood cells obtained from mothers born to mothers who were exposed to farms before birth, a decreased TH2 immune response, a decrease in the number of white blood cells, an increase in the number of Treg cells, and an increase in immunosuppressive ability, while pro-inflammatory cytokines such as TNF-α and IL-6 in umbilical cord blood are increased (62, 63).
3 Postnatal factors affecting the development of the infant microbiota and immune system
3.1 Effects of mode of delivery on infants
Although the fetus has begun to colonize and establish microbiota in the mother's body, the main maternal origin of microbiota may begin to build up in large quantities during production. Vaginal delivery(VD) or caesarean section(CS) largely determines the commensal microbiota that begins to settle in the newborn, and there are significant differences in the degree of oral and intestinal microbial development between different modes of delivery (64, 65).
VD infants have oral and intestinal microbiota enriched with the mother's vaginal microbiota, such as Lactobacillus, Prevotella and Bifidobacterium, during the first week of life (66). At the same time, VD infants are consistently enriched with maternal gut microbiota such as Bacteroidetes, Bifidobacterium and Escherichia coli, which may be due to the better adaptation of maternally transmitted high-abundance bacteria to the intestinal environment (29). It was found that neonates VD had high levels of gram-negative bacteria, which led to a significant enrichment in lipopolysaccharide (LPS) biosynthesis, and that LPS stimulation of primary human immune cells led to higher levels of TNF-α and IL-18, suggesting a link between the immunostimulatory potential of gut microbial LPS and the overall immune status of neonates (67). Another study also showed that LPS was involved in building tolerance to the colonized gut microbiota and initiating the neonatal immune system, and that stimulatory LPS helped reduce the risk of immune-mediated disease (68). Cord blood from vaginally delivered infants has higher immune cell counts and activity (e.g., neutrophils, monocytes, NK cells), higher concentrations of cytokines and expression levels of TLR 2 and TLR 4, and expression levels of CD 16 and CD 56 surface receptors (68).
Newborns delivered by CS have microbiota similar to the maternal skin microbiota, such as Staphylococcus, Corynebacterium, Propionibacterium and environmental microbiota, whereas Bifidobacterium spp. which are common in VD are not found (69). Bacteroides are associated with a lower proportion of the oral and intestinal microbiota of neonates born by CS compared with VD, which may further contribute to reduced bacterial diversity in caesarean section infants (70). Compared with VD, the potential pathogens of CS infants are more abundant, such as Clostridium perfringens or Escherichia coli (71, 72). It may be related to the reduction of leukocytes such as lymphocytes and dendritic cells(DC) in the umbilical cord blood of infants born by CS, as well as the expression of lower levels of the surface innate antigen receptors TLR2 and TLR4 by mononuclear cells in the umbilical cord blood (73, 74). In addition, CS infants had reduced responses to TLR1/2 stimulation of TNF-α and IL-6 (74). CS may affect the critical window period of immune system startup and destroy the mother-to-child transmission and immune stimulation potential of specific microbiota (67). Based on mouse models, it has been found that differences in the earliest initiation of the immune system may continue to affect the development and healthy development of the human immune system (75). Another study also found that newborns born by CS were at higher risk of developing chronic diseases in the future due to changes in early immune system stimulation (76). Compared with VD, CS is associated with an increased risk of childhood immune diseases such as asthma and allergies, leukemia and IBD. The above results suggest that the mode of delivery affects the colonization of neonatal microbiota and the development of the immune system, thereby providing different defenses against infection in the later stages. CS will increase the risk of antibiotic exposure and change the composition of breast milk. At the same time, it leads to an increase in the relative abundance of opportunistic pathogens in neonates, affecting the immune and metabolic development of infants. In addition, by analysing the early microbiota composition and dynamics of 34 mother-infant pairs, Selma-Royo et al. found a significant effect of home vs. hospital births, which was also evident at 6 months of age, but by which time the differences associated with mode of delivery had disappeared (77). Therefore, while focusing on the mode of delivery, relevant factors (e.g., place of delivery, etc.) should be added to the mix to better understand infant bacterial colonisation and its potential long-term impact on infant development.
3.2 Effects of feeding methods on infants
Traditionally, breast milk has been considered sterile, but several studies have confirmed that breast milk provides an important source of bacteria for infants, with Staphylococcus, Streptococcus, Bifidobacterium, Propionibacterium and Lactobacillus present in breast milk samples (78, 79). According to WHO's recommendation, breast-feeding begins within one hour after birth, and exclusive breast-feeding can last until the baby is 6 months old (5). The oral microbiota of breastfed infants is highly similar to that of their mother's mouth, breast milk and areolas. Streptococcus dominates the oral microbiota of exclusively breastfed infants, while Actinomyces and Prevotella are higher in formula-fed infants (80). Bifidobacterium were less represented in the intestinal microbiota of infants fed formula that was not supplemented with probiotics or human milk oligosaccharides (HMO) compared to breastfed children, and supplementation of formula with Bifidobacterium did not significantly increase Bifidobacterium in the infant gut (81). Differences in the initial microbiota due to different feeding patterns can have long-term effects on the oral and intestinal microbiota of infants (82).
The development and maturation of the innate and adaptive immune systems of neonates is time-dependent. In the first few days, immune-related substances such as secretory immunoglobulins in breast milk are the main source of antibodies and immune cells in newborns (82, 83). Breast milk contains high levels of sIgA, which has a crucial role in pathogen clearance, microbiota colonization and in microbiota homeostasis by influencing microbiota gene expression. Studies have shown that maternal sIgA may regulate the development of the oral microbiota by limiting the colonization of potentially pathogenic species, protecting and regulating the homeostasis of mucosal epithelial cells, and has been found to inhibit the local adhesion of specific pathogenic bacteria (84, 85). SIgA in the feces of breastfed infants around 6 months of age was significantly higher than that of formula-fed infants (86). Breast milk also provides immunoglobulins, complement proteins, lysozyme and lactoferrin, antimicrobial substances that protect infants from pathogens and influence their immune maturation. Depending on the Fc receptor (FcRn) of newborn mice, IgG derived from breast milk can enter the blood stream of mice, which is very important to protect against mucosal diseases induced by Escherichia coli (87). The main cytokine present in human milk is IL-10, which suppresses the immune response and participates in tolerance to dietary and microbial antigens (88). Breast milk contains macrophages, neutrophils and lymphocytes, which can be transferred directly to the infant through breast milk and trigger an immune response in the infant and influence the phenotype of the infant's immune cells, particularly the B and T cell phenotype (89, 90).
There are other bioactive components present in human milk, mainly composed of species involved in HMO metabolism in human milk (91, 92). Charbonneau et al. dissected the presence of microbiota-dependent effects of HMO (93). Because HMO can't be digested by infants, they play a role as prebiotics, supporting the growth of some beneficial bacterial strains such as Bifidobacterium, Streptococcus, Staphylococcus and so on in infants' gastrointestinal tract (94), regulates intestinal epithelial cell response and improves intestinal barrier function, provides the primary substrate that shapes the intestinal microbiota of infants and influences the maturation of the intestinal mucosal immune systemand as immunomodulators that compete with potentially pathogenic microbiota to prevent infection and the adhesion and invasion of certain pathogens (95, 96). In addition, HMOs can be fermented by certain gut bacteria to produce short-chain fatty acids(SCFAs), SCFAs are known to be immunomodulators and can even shape the adult immune system by activating G-protein coupled receptor 41/43(GPR 41/43). This pathway was shown to reduce the severity of allergic asthma and colitis in mouse models (97–99). The above studies suggest that increasing HMO content and thus short-chain fatty acid levels by breastfeeding newborns may be a new option for preventing allergic diseases in children.
The above results indicate that breast milk contributes to the establishment of oral and intestinal microbiota, and that immune-related substances in breast milk directly or indirectly promote the development of the infant's immune system. In the long run, breastfeeding is associated with enhanced cognitive development and reduced risk of diseases such as nephrotic syndrome, obesity and type 2 diabetes in children (100–103). However, Gámez-Valdez et al. found that in colostrum samples from women with gestational diabetes mellitus, the GDM group showed higher microbial diversity (104). Also in Piñeiro-Salvador et al. peripheral blood and colostrum paired samples from mothers with BMI > 30 and BMI < 25 were analysed and found that compared to the lean group, the colostrum B lymphocyte compartment was significantly reduced, and CD16 blood monocytes had an increased CD16 expression compared to the lean group in obesity (105). In conclusion, gestational diabetes mellitus (GDM) and obesity affect colostrum composition and lead to abnormal oral and intestinal microbiota colonisation and impact on the establishment of the immune system via the breast milk microbiota and maternal-derived cytokines and leukocytes. Therefore, more long-term follow-up clinical studies are needed to elucidate the influence of maternal factors on breast milk composition and the control mechanisms linking breast milk composition to a diverse infant microbiome and health or disease status later in life.
3.3 Effects of antibiotic exposure on infants
Exposure to antibiotics in early life will destroy most oral and intestinal microbiota, reduce the diversity of microbiota and change the bacterial community structure (Table 2). Dzidic et al's research shows that there are a lot of unique bacteria in infants treated with amoxicillin and penicillin in early life (Effect of caries removal status and time, p = 0.05).For example, the genus Granulicatella was higher in abundance at 24 months of age (p = 0.003) in children not taking antibiotics while Prevotella (p = 0.020) was more prevalent at 7 years of age in children treated with antibiotics in early life (82). Kennedy et al. showed that antibiotic treatment was positively correlated with two OTUs in Pasteurellaceae and Neisseriaceae families, and negatively correlated with OTUs within the Prevotellaceae family (112). It is reported that antibiotic exposure reduces α diversity and increases the abundance of Enterobacteriaceae, and antibiotic-specific enrichment of antibiotic resistance genes (ARGs) and multidrug resistant organisms (MDROs) (113, 114).
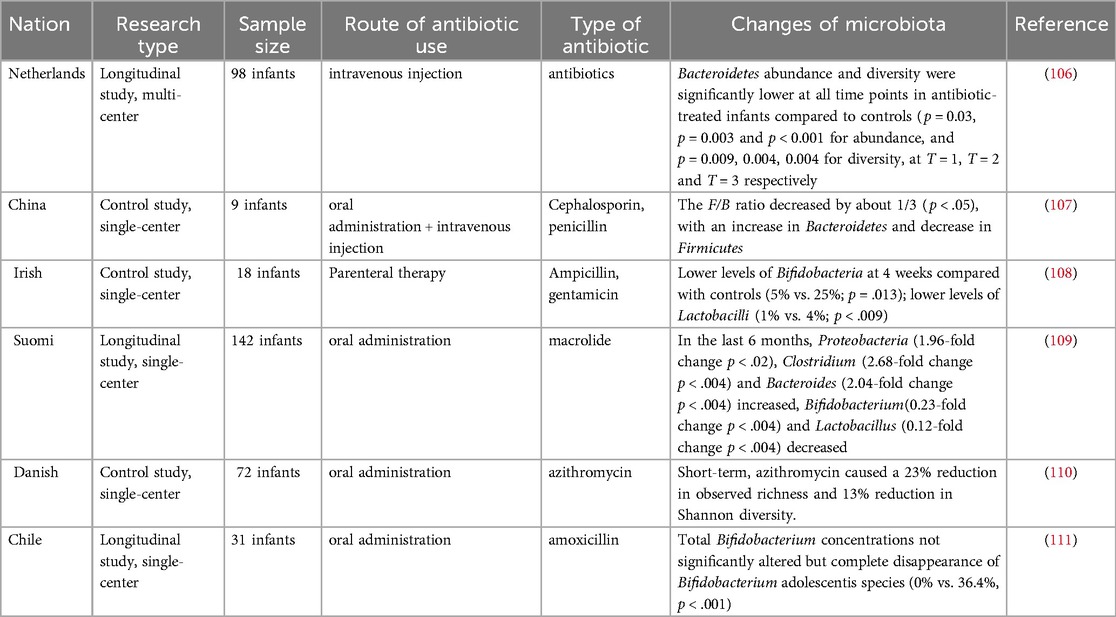
Table 2. Effects of antibiotic exposure on infant microbiota.
Antibiotic use may have potential long-term effects on microbiota development. The results of the study showed that the proportion of Neisseria and Streptococcus mitis/dentisani was increased in the oral cavity of 7-year-old children who had not been exposed to antibiotics in the first two years of life, while the levels of Prevotella and Actinomyces were higher in children exposed to antibiotics (82). In a study by Yassour et al, antibiotic treatment in the first year of life was found to be associated with a reduction in microbial diversity at 3 years of age (115). At the same time, mouse pups were treated with antibiotics, and it was found that the levels of interleukin IL-4 and IgE increased, the number of Treg cells decreased, and the Treg/Th balance was disrupted (116, 117). Early antibiotic exposure has been reported to predispose to microecological dysregulation and immune system dysfunction, increasing susceptibility to asthma, allergic diseases, IBD, Crohn's disease, type 1 diabetes, and other diseases (118, 119).
3.4 Effects of environmental exposure on infants
The results of many studies suggest that it is environmental factors, rather than host genetic factors, that shape the human gut microbiota (120). The biodiversity of the environment, the human commensal microbiota and the human immune system are a complex system of interactions. Current findings show that air pollutants can be ingested directly into the body through the oral cavity with food and liquids, and also by being inhaled into the lungs, where smaller particles reach the alveolar space and are transported by alveolar macrophages to the oropharynx and gastrointestinal tract.The intake of particulate matter will change the intestinal microbiota (for example, the relative numbers of Bacteroides, Firmicutes and Verrucomicrons have changed significantly), or increase the production of ROS and the release of inflammatory factors to promote the disorder of intestinal microbiota (121, 122). Lehtimäki et al. showed that urbanization-related changes in the infant microbiota increase the risk of asthma and atopic features, possibly through crosstalk with the developing immune system (123). In contrast, exposure to the farm environmental microbiota directs the infant gut microbiota toward appropriate tolerances, thereby reducing their risk of developing asthma. Thus, postnatal environmental exposure is an important factor in the infant oral and gut microbiota (124).
4 Discussion
Dysbiosis of the oral microbiota increases the risk of developing oral and systemic diseases, and many related diseases overlap with systemic diseases caused by dysbiosis of the gut microbiota (125). Research has shown that the oral microbiota is an endogenous reservoir of the gut microbiota (126). Interactions between multiple factors such as bacterial translocation, circulating bacteria, bacterial metabolites, immune cells, and inflammatory factors influence the homeostasis of the oral and intestinal microbiota in a bidirectional manner (127). Thus, researchers propose that oral microbes are major contributors to the overall health of the host and that the oral-gut axis may serve as a potential causal mechanism linking systemic diseases (128). It is particularly important to establish healthy oral and gut microbiota during the critical window of 1,000 days of life.
The abundance of bifidobacteria in infants decreases with generations and there is a trend toward increased oral and intestinal pathogens due to antibiotic use, the presence of latent pathogens in the mother and the environment, and genetic disorders (129). The depletion and exhaustion of the collective microbial reservoir by the misuse of antibiotics leads to the emergence and spread of drug-resistant microorganisms, making the re-establishment of a healthy microbiota difficult, with increased susceptibility to infections, greater symptomatology and higher mortality rates (130). The World Health Organization has identified antibiotic resistance as one of the top ten global health threats (131). Probiotics have great potential as a “new age” immunotherapy and an alternative to antibiotics. Probiotics are microorganisms that contain sufficient quantities of defined microorganisms to interfere with the growth of pathogenic bacteria through the secretion of bacteriostatic substances, competition with pathogenic bacteria for nutrients and adhesion sites. Enomoto et al. reported that prenatal supplementation of pregnant women with Bifidobacterium breve M-16 V and Bifidobacterium longum BB536 and subsequent postnatal supplementation of newborns may activate the anti-allergic mechanisms of the immature immune system and significantly reduced the risk of eczema/atopic dermatitis found in infants in the probiotic group during the first 18 months of life [OR: 0.231 [95% CI: 0.084–0.628 ] and 0.304 [0.105–0.892] at 10 and 18 months of age, respectively]. In conclusion, probiotics increase the diversity of the oral and intestinal microbiota, enhance immunity in infants and children, and combat viral respiratory infections through strain-specific immunomodulatory effects and stimulation of the interferon (IFN) pathway (132, 133). Numerous basic and clinical studies have demonstrated the effectiveness of probiotics in the treatment of oral and intestinal diseases in infants (Table 3). In addition, probiotics have specific effects on reducing the expression of resistance genes in addition to their general effects on pathogenic bacteria. Studies have shown that probiotic strains such as C. butyricum MIYAIRI 588 reduce the expression of resistance genes (143). Probiotic strains may be a potential solution for mitigating the problem of antibiotic resistance, and more research is expected to advance the understanding of probiotic strains for mitigating antibiotic resistance.

Table 3. Effects of various probiotics on the microbiology of the infant's mouth and intestines.
It has been found that during the critical window of the first 1,000 days of life, it is possible to regulate the health of newborns in a non-invasive and inexpensive way, such as supplementing with probiotics and prebiotics to regulate and restore the oral and intestinal microbiota in order to reduce children's risk of diseases such as asthma, allergies, and obesity, as well as global morbidity and mortality rates associated with childhood malnutrition (144–147). In the future, personalized infant diets should be further explored and developed to restore microbiota disorders caused by factors such as cesarean section, prenatal or postnatal antibiotics. The study by Gámez-Valdez and Piñeiro-Salvador found that differences in bacterial communities and leukocytes in samples from colostrum from obese women may have an impact on the establishment of an infant's oral and intestinal microbiomes and immune system, and may even pose a threat to the infant's future health (104, 105). Korpela et al. transferred fecal microbiota from mothers to infants to correct gut microbiota imbalances common in CS-born infants and showed that FMT restored microbiota development in infants born vaginally, while FMT attenuated increased levels of potentially opportunistic pathogens in infants born as a result of CS (148). Microbial Ecosystem Transplantation (MET), on the other hand, is a more advanced and precise method than Fecal Microbiota Transplantation (FMT), which involves the purification and cultivation of selected beneficial bacteria from a sample to produce a well-defined and stable microbial ecosystem that can be transplanted into a recipient (149). Therefore, an innovative application of MET may be to address the specific needs of the infant gut microbiota by developing age-specific formulas for the first 1,000 days of life to promote healthy development and prevent or treat dysbiosis. These age-specific MET formulations may have long-term benefits for infant health by supporting immune and metabolic development. Further research is needed to evaluate the safety, efficacy, and long-term consequences of probiotics, FMT, and MET in infants as promising avenues for future research in the field of infant microbiota therapy.Currently, many questions remain about microbiota-immune system interactions, the oro-intestinal axis, or the mechanisms of their interactions with systemic diseases. There is a need for more in-depth exploration of the impact of maternal and child factors (e.g., obesity, diabetes) on the establishment of the microbiota and the immune system, and the mechanism of action of how alterations in the microbiota can increase susceptibility to disease, in order to provide a scientific basis for targeted interventions and therapeutic approaches.
Author contributions
JZ: Formal Analysis, Investigation, Software, Writing – original draft. MH: Formal Analysis, Investigation, Software, Writing – original draft. SL: Validation, Writing – review & editing. YL: Validation, Writing – review & editing. XX: Validation, Writing – review & editing. ZG: Validation, Writing – original draft, Writing – review & editing. QW: Validation, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. The authors are cordially thankful to the Scientific Research and Innovation Fund of Wuhan Wuchang Hospital (Project No.WCYY-2022K02) for their financial support to conduct this project successfully.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Westaway JAF, Huerlimann R, Kandasamy Y, Miller CM, Norton R, Watson D, et al. To probiotic or not to probiotic: a metagenomic comparison of the discharge gut microbiome of infants supplemented with probiotics in NICU and those who are not. Front Pediatr. (2022) 10:838559. doi: 10.3389/fped.2022.838559
2. La Fata G, Weber P, Mohajeri MH. Probiotics and the gut immune system: indirect regulation. Probiotics Antimicrob Proteins. (2018) 10(1):11–21. doi: 10.1007/s12602-017-9322-6
3. Mazziotta C, Tognon M, Martini F, Torreggiani E, Rotondo JC. Probiotics mechanism of action on immune cells and beneficial effects on human health. Cells. (2023) 12(1):184. doi: 10.3390/cells12010184
4. Choo SW, Mohammed WK, Mutha NVR, Rostami N, Ahmed H, Krasnogor N, et al. Transcriptomic responses to coaggregation between streptococcus gordonii and streptococcus oralis. Appl Environ Microbiol. (2021) 87(22):e0155821. doi: 10.1128/AEM.01558-21
5. Blum J, Silva M, Byrne SJ, Butler CA, Adams GG, Reynolds EC, et al. Temporal development of the infant oral microbiome. Crit Rev Microbiol. (2022) 48(6):730–42. doi: 10.1080/1040841X.2021.2025042
6. Korpela K, de Vos WM. Infant gut microbiota restoration: state of the art. Gut Microbes. (2022) 14(1):2118811. doi: 10.1080/19490976.2022.2118811
7. Nagpal R, Tsuji H, Takahashi T, Nomoto K, Kawashima K, Nagata S, et al. Ontogenesis of the gut microbiota composition in healthy, full-term, vaginally born and breast-fed infants over the first 3 years of life: a quantitative bird’s-eye view. Front Microbiol. (2017) 8:1388. doi: 10.3389/fmicb.2017.01388
8. Mohajeri MH, Brummer RJM, Rastall RA, Weersma RK, Harmsen HJM, Faas M, et al. The role of the microbiome for human health: from basic science to clinical applications. Eur J Nutr. (2018) 57(Suppl 1):1–14. doi: 10.1007/s00394-018-1703-4
9. Vandenplas Y, Carnielli VP, Ksiazyk J, Luna MS, Migacheva N, Mosselmans JM, et al. Factors affecting early-life intestinal microbiota development. Nutrition. (2020) 78:110812. doi: 10.1016/j.nut.2020.110812
10. Raspini B, Vacca M, Porri D, De Giuseppe R, Calabrese FM, Chieppa M, et al. Early life microbiota colonization at six months of age: a transitional time point. Front Cell Infect Microbiol. (2021) 11:590202. doi: 10.3389/fcimb.2021.590202
11. Craig SJC, Blankenberg D, Parodi ACL, Paul IM, Birch LL, Savage JS, et al. Child weight gain trajectories linked to oral microbiota composition. Sci Rep. (2018) 8(1):14030. doi: 10.1038/s41598-018-31866-9
12. Juárez-Chairez MF, Cid-Gallegos MS, Jiménez-Martínez C, Prieto-Contreras LF, Bollain Y-l-R. The role of microbiota on rheumatoid arthritis onset. Int J Rheum Dis. (2024) 27(3):e15122. doi: 10.1111/1756-185X.15122
13. Hicks SD, Uhlig R, Afshari P, Williams J, Chroneos M, Tierney-Aves C, et al. Oral microbiome activity in children with autism spectrum disorder. Autism Res. (2018) 11(9):1286–99. doi: 10.1002/aur.1972
14. Kaan AMM, Kahharova D, Zaura E. Acquisition and establishment of the oral microbiota. Periodontol 2000. (2021) 86(1):123–41. doi: 10.1111/prd.12366
15. Mills S, Stanton C, Lane JA, Smith GJ, Ross RP. Precision nutrition and the microbiome, part I: current state of the science. Nutrients. (2019) 11(4):923. doi: 10.3390/nu11040923
16. Jiao Y, Wu L, Huntington ND, Zhang X. Crosstalk between gut microbiota and innate immunity and its implication in autoimmune diseases. Front Immunol. (2020) 11:282. doi: 10.3389/fimmu.2020.00282
17. He Q, Kwok LY, Xi X, Zhong Z, Ma T, Xu H, et al. The meconium microbiota shares more features with the amniotic fluid microbiota than the maternal fecal and vaginal microbiota. Gut Microbes. (2020) 12(1):1794266. doi: 10.1080/19490976.2020.1794266
18. Hansen R, Scott KP, Khan S, Martin JC, Berry SH, Stevenson M, et al. First-pass meconium samples from healthy term vaginally-delivered neonates: an analysis of the microbiota. PLoS One. (2015) 10(7):e0133320. doi: 10.1371/journal.pone.0133320
19. Martin R, Makino H, Cetinyurek Yavuz A, Ben-Amor K, Roelofs M, Ishikawa E, et al. Early-life events, including mode of delivery and type of feeding, siblings and gender, shape the developing gut microbiota. PLoS One. (2016) 11(6):e0158498. doi: 10.1371/journal.pone.0158498
20. Collado MC, Rautava S, Aakko J, Isolauri E, Salminen S. Human gut colonisation may be initiated in utero by distinct microbial communities in the placenta and amniotic fluid. Sci Rep. (2016) 6(1):23129. doi: 10.1038/srep23129
21. Rehbinder EM, Lødrup Carlsen KC, Staff AC, Angell IL, Landrø L, Hilde K, et al. Is amniotic fluid of women with uncomplicated term pregnancies free of bacteria? Am J Obstet Gynecol. (2018) 219(3):289.e1–.e12. doi: 10.1016/j.ajog.2018.05.028
22. Stinson LF, Boyce MC, Payne MS, Keelan JA. The not-so-sterile womb: evidence that the human fetus is exposed to bacteria prior to birth. Front Microbiol. (2019) 10:1124. doi: 10.3389/fmicb.2019.01124
23. Aagaard K, Ma J, Antony KM, Ganu R, Petrosino J, Versalovic J. The placenta harbors a unique microbiome. Sci Transl Med. (2014) 6(237):237ra65. doi: 10.1126/scitranslmed.3008599
24. Theis KR, Romero R, Winters AD, Greenberg JM, Gomez-Lopez N, Alhousseini A, et al. Does the human placenta delivered at term have a microbiota? Results of cultivation, quantitative real-time PCR, 16S rRNA gene sequencing, and metagenomics. Am J Obstet Gynecol. (2019) 220(3):267.e1–.e39. doi: 10.1016/j.ajog.2018.10.018
25. Neu J. The microbiome during pregnancy and early postnatal life. Semin Fetal Neonatal Med. (2016) 21(6):373–9. doi: 10.1016/j.siny.2016.05.001
26. Ansari A, Bose S, You Y, Park S, Kim Y. Molecular mechanism of microbiota metabolites in preterm birth: pathological and therapeutic insights. Int J Mol Sci. (2021) 22(15):8145. doi: 10.3390/ijms22158145
27. Borgo PV, Rodrigues VA, Feitosa AC, Xavier KC, Avila-Campos MJ. Association between periodontal condition and subgingival microbiota in women during pregnancy: a longitudinal study. J Appl Oral Sci. (2014) 22(6):528–33. doi: 10.1590/1678-775720140164
28. Fujiwara N, Tsuruda K, Iwamoto Y, Kato F, Odaki T, Yamane N, et al. Significant increase of oral bacteria in the early pregnancy period in Japanese women. J Investig Clin Dent. (2017) 8(1):e12189. doi: 10.1111/jicd.12189
29. Xiao L, Zhao F. Microbial transmission, colonisation and succession: from pregnancy to infancy. Gut. (2023) 72(4):772–86. doi: 10.1136/gutjnl-2022-328970
30. Jang H, Patoine A, Wu TT, Castillo DA, Xiao J. Oral microflora and pregnancy: a systematic review and meta-analysis. Sci Rep. (2021) 11(1):16870. doi: 10.1038/s41598-021-96495-1
31. Koren O, Goodrich JK, Cullender TC, Spor A, Laitinen K, Bäckhed HK, et al. Host remodeling of the gut microbiome and metabolic changes during pregnancy. Cell. (2012) 150(3):470–80. doi: 10.1016/j.cell.2012.07.008
32. Nuriel-Ohayon M, Neuman H, Ziv O, Belogolovski A, Barsheshet Y, Bloch N, et al. Progesterone increases bifidobacterium relative abundance during late pregnancy. Cell Rep. (2019) 27(3):730–6.e3. doi: 10.1016/j.celrep.2019.03.075
33. Steegers-Theunissen RP, Twigt J, Pestinger V, Sinclair KD. The periconceptional period, reproduction and long-term health of offspring: the importance of one-carbon metabolism. Hum Reprod Update. (2013) 19(6):640–55. doi: 10.1093/humupd/dmt041
34. Mor G. Introduction to the immunology of pregnancy. Immunol Rev. (2022) 308(1):5–8. doi: 10.1111/imr.13102
35. Aagaard K, Riehle K, Ma J, Segata N, Mistretta TA, Coarfa C, et al. A metagenomic approach to characterization of the vaginal microbiome signature in pregnancy. PLoS One. (2012) 7(6):e36466. doi: 10.1371/journal.pone.0036466
36. Gilbert NM, Foster LR, Cao B, Yin Y, Mysorekar IU, Lewis AL. Gardnerella vaginalis promotes group B streptococcus vaginal colonization, enabling ascending uteroplacental infection in pregnant mice. Am J Obstet Gynecol. (2021) 224(5):530.e1–.e17. doi: 10.1016/j.ajog.2020.11.032
37. Fettweis JM, Serrano MG, Brooks JP, Edwards DJ, Girerd PH, Parikh HI, et al. The vaginal microbiome and preterm birth. Nat Med. (2019) 25(6):1012–21. doi: 10.1038/s41591-019-0450-2
38. Hoo R, Nakimuli A, Vento-Tormo R. Innate immune mechanisms to protect against infection at the human decidual-placental interface. Front Immunol. (2020) 11:2070. doi: 10.3389/fimmu.2020.02070
39. Jennewein MF, Goldfarb I, Dolatshahi S, Cosgrove C, Noelette FJ, Krykbaeva M, et al. Fc glycan-mediated regulation of placental antibody transfer. Cell. (2019) 178(1):202–15.e14. doi: 10.1016/j.cell.2019.05.044
40. Schoenmakers S, Steegers-Theunissen R, Faas M. The matter of the reproductive microbiome. Obstet Med. (2019) 12(3):107–15. doi: 10.1177/1753495X18775899
41. Ramírez-Lozada T, Loranca-García MC, Fuentes-Venado CE, Rodríguez-Cerdeira C, Ocharan-Hernández E, Soriano-Ursúa MA, et al. Does the fetus limit antibiotic treatment in pregnant patients with COVID-19? Antibiotics. (2022) 11(2):252. doi: 10.3390/antibiotics11020252
42. Verstraelen H, Vilchez-Vargas R, Desimpel F, Jauregui R, Vankeirsbilck N, Weyers S, et al. Characterisation of the human uterine microbiome in non-pregnant women through deep sequencing of the V1–2 region of the 16S rRNA gene. PeerJ. (2016) 4:e1602. doi: 10.7717/peerj.1602
43. Gao Y, Nanan R, Macia L, Tan J, Sominsky L, Quinn TP, et al. The maternal gut microbiome during pregnancy and offspring allergy and asthma. J Allergy Clin Immunol. (2021) 148(3):669–78. doi: 10.1016/j.jaci.2021.07.011
44. Azad MB, Konya T, Persaud RR, Guttman DS, Chari RS, Field CJ, et al. Impact of maternal intrapartum antibiotics, method of birth and breastfeeding on gut microbiota during the first year of life: a prospective cohort study. BJOG. (2016) 123(6):983–93. doi: 10.1111/1471-0528.13601
45. Tapiainen T, Koivusaari P, Brinkac L, Lorenzi HA, Salo J, Renko M, et al. Impact of intrapartum and postnatal antibiotics on the gut microbiome and emergence of antimicrobial resistance in infants. Sci Rep. (2019) 9(1):10635. doi: 10.1038/s41598-019-46964-5
46. Coker MO, Hoen AG, Dade E, Lundgren S, Li Z, Wong AD, et al. Specific class of intrapartum antibiotics relates to maturation of the infant gut microbiota: a prospective cohort study. BJOG. (2020) 127(2):217–27. doi: 10.1111/1471-0528.15799
47. Nogacka A, Salazar N, Suárez M, Milani C, Arboleya S, Solís G, et al. Impact of intrapartum antimicrobial prophylaxis upon the intestinal microbiota and the prevalence of antibiotic resistance genes in vaginally delivered full-term neonates. Microbiome. (2017) 5(1):93. doi: 10.1186/s40168-017-0313-3
48. Samarra A, Flores E, Bernabeu M, Cabrera-Rubio R, Bäuerl C, Selma-Royo M, et al. Shaping microbiota during the first 1000 days of life. Adv Exp Med Biol. (2024) 1449:1–28. doi: 10.1007/978-3-031-58572-2_1
49. Zou ZH, Liu D, Li HD, Zhu DP, He Y, Hou T, et al. Prenatal and postnatal antibiotic exposure influences the gut microbiota of preterm infants in neonatal intensive care units. Ann Clin Microbiol Antimicrob. (2018) 17(1):9. doi: 10.1186/s12941-018-0264-y
50. Grech A, Collins CE, Holmes A, Lal R, Duncanson K, Taylor R, et al. Maternal exposures and the infant gut microbiome: a systematic review with meta-analysis. Gut Microbes. (2021) 13(1):1–30. doi: 10.1080/19490976.2021.1897210
51. Benner M, Lopez-Rincon A, Thijssen S, Garssen J, Ferwerda G, Joosten I, et al. Antibiotic intervention affects maternal immunity during gestation in mice. Front Immunol. (2021) 12:685742. doi: 10.3389/fimmu.2021.685742
52. Xu H, Yi X, Cui Z, Li H, Zhu L, Zhang L, et al. Maternal antibiotic exposure enhances ILC2 activation in neonates via downregulation of IFN1 signaling. Nat Commun. (2023) 14(1):8332. doi: 10.1038/s41467-023-43903-x
53. Nuriel-Ohayon M, Neuman H, Koren O. Microbial changes during pregnancy, birth, and infancy. Front Microbiol. (2016) 7:1031. doi: 10.3389/fmicb.2016.01031
54. Pedersen TM, Stokholm J, Thorsen J, Mora-Jensen AC, Bisgaard H. Antibiotics in pregnancy increase children’s risk of otitis media and ventilation tubes. J Pediatr. (2017) 183:153–8.e1. doi: 10.1016/j.jpeds.2016.12.046
55. Li DK, Chen H, Ferber J, Odouli R. Maternal infection and antibiotic use in pregnancy and the risk of childhood obesity in offspring: a birth cohort study. Int J Obes. (2020) 44(4):771–80. doi: 10.1038/s41366-019-0501-2
56. Alhasan MM, Cait AM, Heimesaat MM, Blaut M, Klopfleisch R, Wedel A, et al. Antibiotic use during pregnancy increases offspring asthma severity in a dose-dependent manner. Allergy. (2020) 75(8):1979–90. doi: 10.1111/all.14234
57. Zhu N, Ji X, Geng X, Yue H, Li G, Sang N. Maternal PM(2.5) exposure and abnormal placental nutrient transport. Ecotoxicol Environ Saf. (2021) 207:111281. doi: 10.1016/j.ecoenv.2020.111281
58. Tao S, Yang M, Pan B, Wang Y, Tian F, Han D, et al. Maternal exposure to ambient PM(2.5) perturbs the metabolic homeostasis of maternal serum and placenta in mice. Environ Res. (2023) 216(Pt 3):114648. doi: 10.1016/j.envres.2022.114648
59. García-Serna AM, Hernández-Caselles T, Jiménez-Guerrero P, Martín-Orozco E, Pérez-Fernández V, Cantero-Cano E, et al. Air pollution from traffic during pregnancy impairs newborn’s cord blood immune cells: the NELA cohort. Environ Res. (2021) 198:110468. doi: 10.1016/j.envres.2020.110468
60. Fussell JC, Jauniaux E, Smith RB, Burton GJ. Ambient air pollution and adverse birth outcomes: a review of underlying mechanisms. BJOG. (2024) 131(5):538–50. doi: 10.1111/1471-0528.17727
61. Loss G, Bitter S, Wohlgensinger J, Frei R, Roduit C, Genuneit J, et al. Prenatal and early-life exposures alter expression of innate immunity genes: the PASTURE cohort study. J Allergy Clin Immunol. (2012) 130(2):523–30.e9. doi: 10.1016/j.jaci.2012.05.049
62. Schaub B, Liu J, Höppler S, Schleich I, Huehn J, Olek S, et al. Maternal farm exposure modulates neonatal immune mechanisms through regulatory T cells. J Allergy Clin Immunol. (2009) 123(4):774–82.e5. doi: 10.1016/j.jaci.2009.01.056
63. Pfefferle PI, Büchele G, Blümer N, Roponen M, Ege MJ, Krauss-Etschmann S, et al. Cord blood cytokines are modulated by maternal farming activities and consumption of farm dairy products during pregnancy: the PASTURE study. J Allergy Clin Immunol. (2010) 125(1):108–15.e1-3. doi: 10.1016/j.jaci.2009.09.019
64. Dominguez-Bello MG, Costello EK, Contreras M, Magris M, Hidalgo G, Fierer N, et al. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc Natl Acad Sci. (2010) 107(26):11971–5. doi: 10.1073/pnas.1002601107
65. Bäckhed F, Roswall J, Peng Y, Feng Q, Jia H, Kovatcheva-Datchary P, et al. Dynamics and stabilization of the human gut microbiome during the first year of life. Cell Host Microbe. (2015) 17(5):690–703. doi: 10.1016/j.chom.2015.04.004
66. Ma X, Ding J, Ren H, Xin Q, Li Z, Han L, et al. Distinguishable influence of the delivery mode, feeding pattern, and infant sex on dynamic alterations in the intestinal microbiota in the first year of life. Microb Ecol. (2023) 86(3):1799–813. doi: 10.1007/s00248-023-02188-9
67. Wampach L, Heintz-Buschart A, Fritz JV, Ramiro-Garcia J, Habier J, Herold M, et al. Birth mode is associated with earliest strain-conferred gut microbiome functions and immunostimulatory potential. Nat Commun. (2018) 9(1):5091. doi: 10.1038/s41467-018-07631-x
68. Gensollen T, Iyer SS, Kasper DL, Blumberg RS. How colonization by microbiota in early life shapes the immune system. Science. (2016) 352(6285):539–44. doi: 10.1126/science.aad9378
69. Dierikx TH, Visser DH, Benninga MA, van Kaam A, de Boer NKH, de Vries R, et al. The influence of prenatal and intrapartum antibiotics on intestinal microbiota colonisation in infants: a systematic review. J Infect. (2020) 81(2):190–204. doi: 10.1016/j.jinf.2020.05.002
70. Pivrncova E, Kotaskova I, Thon V. Neonatal diet and gut microbiome development after C-section during the first three months after birth: a systematic review. Front Nutr. (2022) 9:941549. doi: 10.3389/fnut.2022.941549
71. Mueller NT, Differding MK, Østbye T, Hoyo C, Benjamin-Neelon SE. Association of birth mode of delivery with infant faecal microbiota, potential pathobionts, and short chain fatty acids: a longitudinal study over the first year of life. BJOG. (2021) 128(8):1293–303. doi: 10.1111/1471-0528.16633
72. García-Serna AM, Martín-Orozco E, Hernández-Caselles T, Morales E. Prenatal and perinatal environmental influences shaping the neonatal immune system: a focus on asthma and allergy origins. Int J Environ Res Public Health. (2021) 18(8):3962. doi: 10.3390/ijerph18083962
73. Shen CM, Lin SC, Niu DM, Kou YR. Labour increases the surface expression of two toll-like receptors in the cord blood monocytes of healthy term newborns. Acta Paediatr. (2009) 98(6):959–62. doi: 10.1111/j.1651-2227.2009.01280.x
74. Lurà MP, Gorlanova O, Müller L, Proietti E, Vienneau D, Reppucci D, et al. Response of cord blood cells to environmental, hereditary and perinatal factors: a prospective birth cohort study. PLoS One. (2018) 13(7):e0200236. doi: 10.1371/journal.pone.0200236
75. Vatanen T, Kostic AD, d'Hennezel E, Siljander H, Franzosa EA, Yassour M, et al. Variation in microbiome LPS immunogenicity contributes to autoimmunity in humans. Cell. (2016) 165(6):1551. doi: 10.1016/j.cell.2016.05.056
76. Keag OE, Norman JE, Stock SJ. Long-term risks and benefits associated with cesarean delivery for mother, baby, and subsequent pregnancies: systematic review and meta-analysis. PLoS Med. (2018) 15(1):e1002494. doi: 10.1371/journal.pmed.1002494
77. Selma-Royo M, Dubois L, Manara S, Armanini F, Cabrera-Rubio R, Valles-Colomer M, et al. Birthmode and environment-dependent microbiota transmission dynamics are complemented by breastfeeding during the first year. Cell Host Microbe. (2024) 32(6):996–1010.e4. doi: 10.1016/j.chom.2024.05.005
78. Damaceno QS, Gallotti B, Reis IMM, Totte YCP, Assis GB, Figueiredo HC, et al. Isolation and identification of potential probiotic bacteria from human milk. Probiotics Antimicrob Proteins. (2023) 15(3):491–501. doi: 10.1007/s12602-021-09866-5
79. Cabrera-Rubio R, Collado MC, Laitinen K, Salminen S, Isolauri E, Mira A. The human milk microbiome changes over lactation and is shaped by maternal weight and mode of delivery1234. Am J Clin Nutr. (2012) 96(3):544–51. doi: 10.3945/ajcn.112.037382
80. Oba PM, Holscher HD, Mathai RA, Kim J, Swanson KS. Diet influences the oral microbiota of infants during the first six months of life. Nutrients. (2020) 12(11):3400. doi: 10.3390/nu12113400
81. Bazanella M, Maier TV, Clavel T, Lagkouvardos I, Lucio M, Maldonado-Gòmez MX, et al. Randomized controlled trial on the impact of early-life intervention with bifidobacteria on the healthy infant fecal microbiota and metabolome. Am J Clin Nutr. (2017) 106(5):1274–86. doi: 10.3945/ajcn.117.157529
82. Dzidic M, Collado MC, Abrahamsson T, Artacho A, Stensson M, Jenmalm MC, et al. Oral microbiome development during childhood: an ecological succession influenced by postnatal factors and associated with tooth decay. ISME J. (2018) 12(9):2292–306. doi: 10.1038/s41396-018-0204-z
83. Camacho-Morales A, Caba M, García-Juárez M, Caba-Flores MD, Viveros-Contreras R, Martínez-Valenzuela C. Breastfeeding contributes to physiological immune programming in the newborn. Front Pediatr. (2021) 9:744104. doi: 10.3389/fped.2021.744104
84. Cravioto A, Tello A, Villafán H, Ruiz J, del Vedovo S, Neeser JR. Inhibition of localized adhesion of enteropathogenic escherichia coli to HEp-2 cells by immunoglobulin and oligosaccharide fractions of human colostrum and breast milk. J Infect Dis. (1991) 163(6):1247–55. doi: 10.1093/infdis/163.6.1247
85. Qi C, Tu H, Zhou J, Tu R, Chang H, Chen J, et al. Widespread vertical transmission of secretory immunoglobulin A coated trace bacterial variants from the mother to infant gut through breastfeeding. Food Funct. (2022) 13(22):11543–54. doi: 10.1039/D2FO01244H
86. Donald K, Petersen C, Turvey SE, Finlay BB, Azad MB. Secretory IgA: linking microbes, maternal health, and infant health through human milk. Cell Host Microbe. (2022) 30(5):650–9. doi: 10.1016/j.chom.2022.02.005
87. Zheng W, Zhao W, Wu M, Song X, Caro F, Sun X, et al. Microbiota-targeted maternal antibodies protect neonates from enteric infection. Nature. (2020) 577(7791):543–8. doi: 10.1038/s41586-019-1898-4
88. Pabst O, Mowat AM. Oral tolerance to food protein. Mucosal Immunol. (2012) 5(3):232–9. doi: 10.1038/mi.2012.4
89. Field CJ. The immunological components of human milk and their effect on immune development in infants. J Nutr. (2005) 135(1):1–4. doi: 10.1093/jn/135.1.1
90. Ninkina N, Kukharsky MS, Hewitt MV, Lysikova EA, Skuratovska LN, Deykin AV, et al. Stem cells in human breast milk. Hum Cell. (2019) 32(3):223–30. doi: 10.1007/s13577-019-00251-7
91. Delvaux A, Rathahao-Paris E, Guillon B, Cholet S, Adel-Patient K, Fenaille F, et al. Trapped ion mobility spectrometry time-of-flight mass spectrometry for high throughput and high resolution characterization of human milk oligosaccharide isomers. Anal Chim Acta. (2021) 1180:338878. doi: 10.1016/j.aca.2021.338878
92. Pallarés N, Ferrer E, Berrada H, Barba FJ, Salgado-Ramos M, Collado MC. Mind the gap regarding the knowledge of infant exposure to mycotoxins, acrylamide, bisphenols and heavy metals through human milk. Food Control. (2024) 166:110731. doi: 10.1016/j.foodcont.2024.110731
93. Charbonneau MR, O'Donnell D, Blanton LV, Totten SM, Davis JC, Barratt MJ, et al. Sialylated milk oligosaccharides promote microbiota-dependent growth in models of infant undernutrition. Cell. (2016) 164(5):859–71. doi: 10.1016/j.cell.2016.01.024
94. Ward RE, Niñonuevo M, Mills DA, Lebrilla CB, German JB. In vitro fermentation of breast milk oligosaccharides by bifidobacterium infantis and Lactobacillus gasseri. Appl Environ Microbiol. (2006) 72(6):4497–9. doi: 10.1128/AEM.02515-05
95. Masi AC, Stewart CJ. Untangling human milk oligosaccharides and infant gut microbiome. iScience. (2022) 25(1):103542. doi: 10.1016/j.isci.2021.103542
96. Zhang S, Li T, Xie J, Zhang D, Pi C, Zhou L, et al. Gold standard for nutrition: a review of human milk oligosaccharide and its effects on infant gut microbiota. Microb Cell Fact. (2021) 20(1):108. doi: 10.1186/s12934-021-01599-y
97. Gollwitzer ES, Marsland BJ. Impact of early-life exposures on immune maturation and susceptibility to disease. Trends Immunol. (2015) 36(11):684–96. doi: 10.1016/j.it.2015.09.009
98. Trompette A, Gollwitzer ES, Yadava K, Sichelstiel AK, Sprenger N, Ngom-Bru C, et al. Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nat Med. (2014) 20(2):159–66. doi: 10.1038/nm.3444
99. Maslowski KM, Vieira AT, Ng A, Kranich J, Sierro F, Yu D, et al. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature. (2009) 461(7268):1282–6. doi: 10.1038/nature08530
100. Keim SA, Sullivan JA, Sheppard K, Smith K, Ingol T, Boone KM, et al. Feeding infants at the breast or feeding expressed human milk: long-term cognitive, executive function, and eating behavior outcomes at age 6 years. J Pediatr. (2021) 233:66–73.e1. doi: 10.1016/j.jpeds.2021.02.025
101. Yang Z, Li N, Cui H, Liu B, Wang X, Zhang L, et al. Influence of the interaction between genetic factors and breastfeeding on children’s weight status: a systematic review. Adv Nutr. (2024) 15(11):100312. doi: 10.1016/j.advnut.2024.100312
102. Park PG, Heo JS, Ahn YH, Kang HG. Association between exclusive breastfeeding and the incidence of childhood nephrotic syndrome. J Pediatr. (2025) 276:114266. doi: 10.1016/j.jpeds.2024.114266
103. Horta BL, de Lima NP. Breastfeeding and type 2 diabetes: systematic review and meta-analysis. Curr Diab Rep. (2019) 19(1):1. doi: 10.1007/s11892-019-1121-x
104. Gámez-Valdez JS, García-Mazcorro JF, Montoya-Rincón AH, Rodríguez-Reyes DL, Jiménez-Blanco G, Rodríguez MTA, et al. Differential analysis of the bacterial community in colostrum samples from women with gestational diabetes mellitus and obesity. Sci Rep. (2021) 11(1):24373. doi: 10.1038/s41598-021-03779-7
105. Piñeiro-Salvador R, Vazquez-Garza E, Cruz-Cardenas JA, Licona-Cassani C, García-Rivas G, Moreno-Vásquez J, et al. A cross-sectional study evidences regulations of leukocytes in the colostrum of mothers with obesity. BMC Med. (2022) 20(1):388. doi: 10.1186/s12916-022-02575-y
106. Eck A, Rutten N, Singendonk MMJ, Rijkers GT, Savelkoul PHM, Meijssen CB, et al. Neonatal microbiota development and the effect of early life antibiotics are determined by two distinct settler types. PLoS One. (2020) 15(2):e0228133. doi: 10.1371/journal.pone.0228133
107. Bai L, Zhou P, Li D, Ju X. Changes in the gastrointestinal microbiota of children with acute lymphoblastic leukaemia and its association with antibiotics in the short term. J Med Microbiol. (2017) 66(9):1297–307. doi: 10.1099/jmm.0.000568
108. Fouhy F, Guinane CM, Hussey S, Wall R, Ryan CA, Dempsey EM, et al. High-throughput sequencing reveals the incomplete, short-term recovery of infant gut microbiota following parenteral antibiotic treatment with ampicillin and gentamicin. Antimicrob Agents Chemother. (2012) 56(11):5811–20. doi: 10.1128/AAC.00789-12
109. Korpela K, Salonen A, Virta LJ, Kekkonen RA, Forslund K, Bork P, et al. Intestinal microbiome is related to lifetime antibiotic use in finnish pre-school children. Nat Commun. (2016) 7:10410. doi: 10.1038/ncomms10410
110. Wei S, Mortensen MS, Stokholm J, Brejnrod AD, Thorsen J, Rasmussen MA, et al. Short- and long-term impacts of azithromycin treatment on the gut microbiota in children: a double-blind, randomized, placebo-controlled trial. EBioMedicine. (2018) 38:265–72. doi: 10.1016/j.ebiom.2018.11.035
111. Mangin I, Suau A, Gotteland M, Brunser O, Pochart P. Amoxicillin treatment modifies the composition of bifidobacterium species in infant intestinal microbiota. Anaerobe. (2010) 16(4):433–8. doi: 10.1016/j.anaerobe.2010.06.005
112. Kennedy B, Peura S, Hammar U, Vicenzi S, Hedman A, Almqvist C, et al. Oral microbiota development in early childhood. Sci Rep. (2019) 9(1):19025. doi: 10.1038/s41598-019-54702-0
113. Tanaka S, Kobayashi T, Songjinda P, Tateyama A, Tsubouchi M, Kiyohara C, et al. Influence of antibiotic exposure in the early postnatal period on the development of intestinal microbiota. FEMS Immunol Med Microbiol. (2009) 56(1):80–7. doi: 10.1111/j.1574-695X.2009.00553.x
114. Timens W, Rozeboom T, Poppema S. Fetal and neonatal development of human spleen: an immunohistological study. Immunology. (1987) 60(4):603–9.3294575
115. Bokulich NA, Chung J, Battaglia T, Henderson N, Jay M, Li H, et al. Antibiotics, birth mode, and diet shape microbiome maturation during early life. Sci Transl Med. (2016) 8(343):343ra82. doi: 10.1126/scitranslmed.aad7121
116. Oyama N, Sudo N, Sogawa H, Kubo C. Antibiotic use during infancy promotes a shift in the T(H)1/T(H)2 balance toward T(H)2-dominant immunity in mice. J Allergy Clin Immunol. (2001) 107(1):153–9. doi: 10.1067/mai.2001.111142
117. Noverr MC, Falkowski NR, McDonald RA, McKenzie AN, Huffnagle GB. Development of allergic airway disease in mice following antibiotic therapy and fungal microbiota increase: role of host genetics, antigen, and interleukin-13. Infect Immun. (2005) 73(1):30–8. doi: 10.1128/IAI.73.1.30-38.2005
118. Nguyen LH, Örtqvist AK, Cao Y, Simon TG, Roelstraete B, Song M, et al. Antibiotic use and the development of inflammatory bowel disease: a national case-control study in Sweden. Lancet Gastroenterol Hepatol. (2020) 5(11):986–95. doi: 10.1016/S2468-1253(20)30267-3
119. Kere M, Melén E. Linking antibiotic treatment in early life with childhood allergic disease. Acta Paediatr. (2023) 112(1):14–6. doi: 10.1111/apa.16581
120. Seyedsayamdost MR. High-throughput platform for the discovery of elicitors of silent bacterial gene clusters. Proc Natl Acad Sci. (2014) 111(20):7266–71. doi: 10.1073/pnas.1400019111
121. Salim SY, Kaplan GG, Madsen KL. Air pollution effects on the gut microbiota: a link between exposure and inflammatory disease. Gut Microbes. (2014) 5(2):215–9. doi: 10.4161/gmic.27251
122. Cruells A, Cabrera-Rubio R, Bustamante M, Pelegrí D, Cirach M, Jimenez-Arenas P, et al. The influence of pre- and postnatal exposure to air pollution and green spaces on infant’s gut microbiota: results from the MAMI birth cohort study. Environ Res. (2024) 257:119283. doi: 10.1016/j.envres.2024.119283
123. Lehtimäki J, Thorsen J, Rasmussen MA, Hjelmsø M, Shah S, Mortensen MS, et al. Urbanized microbiota in infants, immune constitution, and later risk of atopic diseases. J Allergy Clin Immunol. (2021) 148(1):234–43. doi: 10.1016/j.jaci.2020.12.621
124. Depner M, Taft DH, Kirjavainen PV, Kalanetra KM, Karvonen AM, Peschel S, et al. Maturation of the gut microbiome during the first year of life contributes to the protective farm effect on childhood asthma. Nat Med. (2020) 26(11):1766–75. doi: 10.1038/s41591-020-1095-x
125. Martínez-García M, Hernández-Lemus E. Periodontal inflammation and systemic diseases: an overview. Front Physiol. (2021) 12:709438. doi: 10.3389/fphys.2021.709438
126. Zhou T, Xu W, Wang Q, Jiang C, Li H, Chao Y, et al. The effect of the “oral-gut” axis on periodontitis in inflammatory bowel disease: a review of microbe and immune mechanism associations. Front Cell Infect Microbiol. (2023) 13:1132420. doi: 10.3389/fcimb.2023.1132420
127. Tortora SC, Agurto MG, Martello LA. The oral-gut-circulatory axis: from homeostasis to colon cancer. Front Cell Infect Microbiol. (2023) 13:1289452. doi: 10.3389/fcimb.2023.1289452
128. Kerstens R, Ng YZ, Pettersson S, Jayaraman A. Balancing the oral-gut-brain axis with diet. Nutrients. (2024) 16(18):3206. doi: 10.3390/nu16183206
129. Finlay BB, Amato KR, Azad M, Blaser MJ, Bosch TCG, Chu H, et al. The hygiene hypothesis, the COVID pandemic, and consequences for the human microbiome. Proc Natl Acad Sci. (2021) 118(6):e2010217118. doi: 10.1073/pnas.2010217118
130. Morehead MS, Scarbrough C. Emergence of global antibiotic resistance. Prim Care. (2018) 45(3):467–84. doi: 10.1016/j.pop.2018.05.006
131. World Health Organization. World Health Organization Ten Threats to Global Health in 2019. (2019). Available online: https://www.who.int/news-room/spotlight/ten-threats-to-global-health-in-2019 (Accessed September 02, 2024).
132. Spacova I, De Boeck I, Cauwenberghs E, Delanghe L, Bron PA, Henkens T, et al. Development of a live biotherapeutic throat spray with lactobacilli targeting respiratory viral infections. Microb Biotechnol. (2023) 16(1):99–115. doi: 10.1111/1751-7915.14189
133. Lehtoranta L, Latvala S, Lehtinen MJ. Role of probiotics in stimulating the immune system in viral respiratory tract infections: a narrative review. Nutrients. (2020) 12(10):3163. doi: 10.3390/nu12103163
134. Guo H, Xiang X, Lin X, Wang Q, Qin S, Lu X, et al. Oropharyngeal probiotic ENT-K12 as an effective dietary intervention for children with recurrent respiratory tract infections during cold season. Front Nutr. (2022) 9:900448. doi: 10.3389/fnut.2022.900448
135. Mageswary MU, Ang XY, Lee BK, Chung YF, Azhar SNA, Hamid IJA, et al. Probiotic bifidobacterium lactis Probio-M8 treated and prevented acute RTI, reduced antibiotic use and hospital stay in hospitalized young children: a randomized, double-blind, placebo-controlled study. Eur J Nutr. (2022) 61(3):1679–91. doi: 10.1007/s00394-021-02689-8
136. Luoto R, Pärtty A, Vogt JK, Rautava S, Isolauri E. Reversible aberrancies in gut microbiome of moderate and late preterm infants: results from a randomized, controlled trial. Gut Microbes. (2023) 15(2):2283913. doi: 10.1080/19490976.2023.2283913
137. Li P, Ren Z, Zhou J, Zhao A, Wang S, Xun Y, et al. Effect of lacticaseibacillus paracasei N1115 on immunomodulatory and gut microbial composition in young children: a randomized, placebo-controlled study. Nutrients. (2023) 15(8):1970. doi: 10.3390/nu15081970
138. Kubota M, Ito K, Tomimoto K, Kanazaki M, Tsukiyama K, Kubota A, et al. Lactobacillus reuteri DSM 17938 and magnesium oxide in children with functional chronic constipation: a double-blind and randomized clinical trial. Nutrients. (2020) 12(1):225. doi: 10.3390/nu12010225
139. Berni Canani R, De Filippis F, Nocerino R, Laiola M, Paparo L, Calignano A, et al. Specific signatures of the gut microbiota and increased levels of butyrate in children treated with fermented cow’s milk containing heat-killed lactobacillus paracasei CBA L74. Appl Environ Microbiol. (2017) 83(19):e01206–17. doi: 10.1128/AEM.01206-17
140. Hiraku A, Nakata S, Murata M, Xu C, Mutoh N, Arai S, et al. Early probiotic supplementation of healthy term infants with bifidobacterium longum subsp. infantis M-63 is safe and leads to the development of bifidobacterium-predominant gut microbiota: a double-blind, placebo-controlled trial. Nutrients. (2023) 15(6):1402. doi: 10.3390/nu15061402
141. Sowden M, van Weissenbruch MM, Bulabula ANH, van Wyk L, Twisk J, van Niekerk E. Effect of a multi-strain probiotic on the incidence and severity of necrotizing enterocolitis and feeding intolerances in preterm neonates. Nutrients. (2022) 14(16):3305. doi: 10.3390/nu14163305
142. Korpela K, Salonen A, Virta LJ, Kumpu M, Kekkonen RA, de Vos WM. Lactobacillus rhamnosus GG intake modifies preschool children’s intestinal microbiota, alleviates penicillin-associated changes, and reduces antibiotic use. PLoS One. (2016) 11(4):e0154012. doi: 10.1371/journal.pone.0154012
143. Kunishima H, Ishibashi N, Wada K, Oka K, Takahashi M, Yamasaki Y, et al. The effect of gut microbiota and probiotic organisms on the properties of extended spectrum beta-lactamase producing and carbapenem resistant Enterobacteriaceae including growth, beta-lactamase activity and gene transmissibility. J Infect Chemother. (2019) 25(11):894–900. doi: 10.1016/j.jiac.2019.04.021
144. Wawryk-Gawda E, Markut-Miotła E, Emeryk A. Postnatal probiotics administration does not prevent asthma in children, but using prebiotics or synbiotics may be the effective potential strategies to decrease the frequency of asthma in high-risk children—a meta-analysis of clinical trials. Allergol Immunopathol. (2021) 49(4):4–14. doi: 10.15586/aei.v49i4.69
145. Fiocchi A, Cabana MD, Mennini M. Current use of probiotics and prebiotics in allergy. J Allergy Clin Immunol Pract. (2022) 10(9):2219–42. doi: 10.1016/j.jaip.2022.06.038
146. Kuitunen M, Kukkonen K, Juntunen-Backman K, Korpela R, Poussa T, Tuure T, et al. Probiotics prevent IgE-associated allergy until age 5 years in cesarean-delivered children but not in the total cohort. J Allergy Clin Immunol. (2009) 123(2):335–41. doi: 10.1016/j.jaci.2008.11.019
147. Wiedmer EB, Herter-Aeberli I. The potential of prebiotic and probiotic supplementation during obese pregnancy to improve maternal and offspring’s metabolic health and reduce obesity risk-A narrative review. Front Nutr. (2022) 9:819882. doi: 10.3389/fnut.2022.819882
148. Korpela K, Helve O, Kolho KL, Saisto T, Skogberg K, Dikareva E, et al. Maternal fecal microbiota transplantation in cesarean-born infants rapidly restores normal gut microbial development: a proof-of-concept study. Cell. (2020) 183(2):324–34.e5. doi: 10.1016/j.cell.2020.08.047
Keywords: first 1,000 days of life, oral microbiota, intestinal microbiota, microecology, immune system
Citation: Zhu J, He M, Li S, Lei Y, Xiang X, Guo Z and Wang Q (2025) Shaping oral and intestinal microbiota and the immune system during the first 1,000 days of life. Front. Pediatr. 13:1471743. doi: 10.3389/fped.2025.1471743
Received: 28 July 2024; Accepted: 10 January 2025;
Published: 21 January 2025.
Edited by:
Luis Ignacio Gonzalez-Granado, University Hospital October 12, SpainReviewed by:
Cuauhtemoc Licona-Cassani, Monterrey Institute of Technology and Higher Education (ITESM), MexicoZhengrui Li, Shanghai Jiao Tong University, China
Copyright: © 2025 Zhu, He, Li, Lei, Xiang, Guo and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zhi Guo, Z3VvemhpNzdAMTI2LmNvbQ==; Qiang Wang, d2FuZ3FpYW5nQHd1c3QuZWR1LmNu
†These authors share first authorship