 Sebastian Wirsching1
Sebastian Wirsching1 Marina Machtakova2Frauke Borgans1,3Leah Pretsch1Michael Fichter1,2,4
Marina Machtakova2Frauke Borgans1,3Leah Pretsch1Michael Fichter1,2,4 Maximiliano L. Cacicedo1
Maximiliano L. Cacicedo1 Héloïse Thérien-Aubin2,5Katharina Landfester2Stephan Gehring1*
Héloïse Thérien-Aubin2,5Katharina Landfester2Stephan Gehring1*- 1Children's Hospital, University Medical Center of the Johannes Gutenberg University, Mainz, Germany
- 2Max Planck Institute for Polymer Research, Mainz, Germany
- 3Department of Infectious Diseases, University Hospital Frankfurt, Frankfurt, Germany
- 4Department of Dermatology, University Medical Center of the Johannes Gutenberg University, Mainz, Germany
- 5Department of Chemistry, Memorial University of Newfoundland, St. John's, NL, Canada
Childhood mortality represents a major issue with 5. 3 million worldwide deaths of children under 5 years of age in 2019. Approximately half of those deaths can be attributed to easily preventable, infectious diseases. Currently approved neonatal vaccines are typically effective only after multiple doses leaving infants especially vulnerable during the first 6 months of life. Survival rates could be improved significantly by developing new and more potent vaccines that are capable of overcoming inherently tolerogenic neonatal immune systems. TLR agonists have garnered a great deal of attention in recent years due to their extensive capacities to activate innate immunity. Herein, the superior capacity of the TLR7/8 agonist, resiquimod (R848), to activate adult and neonatal primary peripheral blood dendritic cells is demonstrated. Moreover, R848 can be conjugated to polyethylene glycol and encapsulated in ovalbumin nanocapsules to efficiently co-deliver antigen and adjuvant in vitro. This study is among the first to demonstrate the capacity of encapsulated R848 to activate neonatal dendritic cells. These findings support the potential incorporation of R848 as adjuvant in neonatal vaccines, making them more effective in eliciting a robust immune response.
Introduction
Easily preventable, infectious diseases accounted for ~50% of the 5.3 million worldwide deaths of children under 5 years of age in 2019. Although childhood mortality has been on a steady decline in recent years, it is projected that, based on current trends, as many as 48.1 million children who are 5 years or younger will die between 2020 and 2030 (1, 2). The development of novel and more potent vaccines that are able to overcome the limitations of the neonatal immune system represents a primary approach to improving the survival rate.
In utero, T cells are mainly polarized toward a TH2 phenotype in order to prevent fetal allograft rejection (3, 4). Immediately after birth, the immune system is skewed toward a tolerogenic phenotype. This polarization persists through the first few months of life, and is mainly due to a dampened TH1-type response leaving infants extremely susceptible to infectious diseases (5). Regulatory T cells, found in cord blood and infant tissues in greater abundance, further contribute to the compromised state of the neonatal immune system (6, 7). The reduced production of pro-inflammatory cytokines, e.g., IL-12p70 and TNF-α, by antigen presenting cells upon Toll-like receptor (TLR) stimulation represents a further contributing factor (8, 9). Additionally, neonatal dendritic cells (DCs) are functionally immature, expressing fewer cell surface MHC II molecules, as well as co-stimulatory receptors: CD40, CD80 and CD86 (9, 10). The administration of Bacille Calmette-Guérin (BCG) vaccine, however, shows that neonatal DCs are fully capable of secreting pro-inflammatory cytokines and mounting a robust TH1-type immune response (11, 12).
Conventional and plasmacytoid DCs fulfill different functions but are equally important in presenting antigens and activating T cells. Conventional DCs, in particular the more abundant CD1c-expressing subset, exhibit a broad range of TLRs and are capable of secreting IL-12p70, TNF-α and IL-6 after sensing pathogens (13, 14). CD123-expressing plasmacytoid DCs, on the other hand, are uniquely tailored to respond to viral infections by expressing TLR7 and TLR9, which recognize single-stranded RNA and DNA, respectively, and by producing type I interferons (15).
Apart from BCG, there are only two other vaccines, Hepatitis B (HBV) and polio, that are currently approved for administration to neonates. Of the three, only the HBV vaccine uses aluminum-based (alum) agents as an adjuvant to boost immune responses. Today, the CDC recommends three doses of the HBV vaccine administered during the first 6 months after birth to achieve robust protection (16). A big challenge is the mother-to-infant transmission, which can be prevented effectively by administrating HBV immunoglobulin together with HBV vaccine (17, 18). This, however, can be challenging in low-income countries since infants only infrequently come in contact with the health care system or certain medications, in this case HBV immunoglobulin, are not available at all (19, 20). Therefore, novel and more effective adjuvants, which activate the neonatal immune system early and decrease childhood mortality, are needed to improve vaccines.
TLR agonists have garnered a great deal of attention in recent years due to their extensive capabilities to activate the innate immune system (21). Today, two TLR agonists are approved for use as vaccine adjuvants. Monophosphoryl lipid A (MPLA), a component of the Cervarix vaccine, is a TLR4 agonist; and CpG oligodeoxynucleotides, a component of the Heplisav-B vaccine, is a TLR9 agonist (22, 23).
The study presented herein demonstrates the superior capacity of the TLR7/8 agonist, resiquimod (R848), relative to alum and MPLA, to activate both adult and neonatal primary peripheral blood DCs. Stimulation with R848 increased both activation marker expression and pro-inflammatory cytokine secretion by conventional and plasmacytoid DCs. Adult and neonatal DCs stimulated with R848 coupled to polyethylene glycol (PEG) and encapsulated in ovalbumin nanocapsules (OVA-PEG-R848), herein used as model antigen, exhibited an activated phenotype. This supports the potential use of encapsulated PEG-R848 as a versatile platform to co-deliver a variety of antigens and R848 to DCs.
Materials and methods
Synthesis of PEG-R848 and labeling with Rhodamine B
PEG-R848 was prepared as previously described (24). Hydroxyl-PEG-NHS ester (160 mg; Sigma, St. Louis, MO) was dissolved in 4 mL of dry dichloromethane before adding 10 mg R848 (Sigma) and 6 μL trimethylamine. The reaction was stirred overnight. Afterwards, the solvent was evaporated, and water was added to the solid product. The resulting solution was dialyzed against water for 3 days, then freeze-dried. PEG-R848 was labeled with a fluorescent dye by dissolving 50 mg in 4 mL DMSO and adding 2 mg of Rhodamine B isothiocyanate (Sigma). Then, the reaction was stirred overnight, dialyzed against water for 5 days, and lyophilized.
Synthesis of protein nanocapsules and encapsulation of PEG-R848
Ovalbumin (OVA) and Ovalbumin-polyethylene glycol-R848 (OVA-PEG-R848) nanocapsules (NCs) were synthesized as previously described (24). Briefly, 20 mg of OVA (Sigma) were dissolved in 0.2 mL PBS-buffer (Sigma) and 1 mg of PEG-R848 was added to the aqueous solution. The organic phase was prepared by dissolving 75 mg of polyglycerol-polyricinoleat (PGPR) (Danisco, Copenhagen, Denmark) in 3 mL toluene. Both phases were emulsified with an Ultra-Turrax® (IKA, Staufen im Breisgau, Germany) and homogenized by one cycle through a LV1 microfluidizer (MFIC, Newton, MA). Afterwards, a solution of 25 mg PGPR and 0.1 μL of Toluol-2,4-diisocyanat (TDI) (TCI Chemicals, Tokyo, Japan) in 0.5 mL toluene was added dropwise to the mini-emulsion and then stirred (20 h, RT). The nanocapsules were purified by centrifugation three times (30 min, 1,200 g) followed by redispersion in fresh toluene to remove excess surfactant and unreacted crosslinker. The nanocapsules were then transferred to PBS for subsequent use.
Nanocapsule characterization
NCs were characterized as described before (24). Shortly, NC size distribution was measured via dynamic light scattering (DLS) at 20°C using a Malvern NanoS90 (Malvern Instruments, UK) device at 90° angle. Zeta potential was measured through dilution of NCs in potassium chloride solution with a Malvern ZetaSizer (Malvern Instruments, UK).
Encapsulation efficiency and release experiments
Encapsulation efficiency of nanocapsule formulations was measured as described before (24). Shortly, encapsulation efficiency was calculated by measuring the total fluorescence intensity of PEG-Rhodamine (λex = 553 nm and λem = 576 nm) in the aqueous suspension. Afterwards, the samples were centrifuged and the remaining, unencapsulated PEG-Rhodamine in the recovered suspension as well as the PEG-Rhodamine in the redispersed NCs was measured.
Release of payload from NCs was measured as previously described (24). Shortly, the release of the PEG-Rhodamine was analyzed by centrifugation of NCs at 1,770 g for 30 min using Vivaspin® Ultrafiltration tubes (500 μl 1,000 K; Sartorius AG, Germany) and measuring the fluorescence of the filtrate (λex = 553 nm and λem = 576 nm).
Isolation and culture of primary peripheral blood DCs
Leukocytes were obtained from leukapheresis products collected from healthy, voluntary donors (blood bank of the University Medical Center Mainz) after informed consent. Umbilical cord blood was collected after cesarean section from healthy neonates according to guidelines of the University Medical Center Mainz and after ethical approval of the local ethics committee. Peripheral blood mononuclear cells (PBMCs) were isolated by Biocoll density centrifugation. Afterwards, the purified PBMC fraction was washed twice with Hank's Balanced Salt Solution (HBSS; Sigma). cDC2 and pDCs were isolated in one step using the Blood Dendritic Cell Isolation Kit II (Miltenyi Biotec, Bergisch-Gladbach, Germany) according to the manufacturer's instructions.
DC stimulation
Primary DCs were cultured and stimulated as described before (24). Shortly, 2 x 105 bulk DCs in serum-free X-VIVO 15 medium were seeded in triplicates into 96-well U-bottom plates. For stimulation with soluble adjuvants, DCs were stimulated with 1 μg/ml MPLA (Sigma), 5 μg/ml Alhydrogel® (2%; Invivogen, San Diego, CA) or 1 μg/ml R848 (Sigma) for 20 h at 37°C and 5% CO2.
Cells stimulated with nanocapsules were incubated with OVA-PEG-R848 NCs at an effective R848 concentration of 500 ng/ml. Empty OVA NCs were used in equimolar concentration as control. Cells were stimulated with 500 ng/ml soluble PEG-R848 as a positive control.
Flow cytometry
The cells were transferred to FACS round-bottom tubes, washed with buffer, then incubated with 10% Privigen® Immunoglobulin solution (CSL Behring, Marburg, Germany) to block Fc receptors. Primary DCs were stained with fluorochrome-conjugated mouse monoclonal antibodies specific for CD83 (FITC), CD80 (PE), CD123 (PE-Cy7), HLA-DR (APC) (all purchased from BD Pharmingen, Franklin Lakes, NJ), CD86 (BV420), CD80 (BV510) (BD Horizon) and CD1c (PerCP-eFluor 710) (eBioscience, San Diego, CA) for 30 min at 4°C. The gating strategy used for the gating of DC populations can be found in Supplementary Figure S1.
Cytometric bead array
Cell culture supernates, collected after 20 h of incubation, were analyzed using the Human Enhanced Sensitivity Flex system (BD) and the MACSPlex Cytokine assay system (Miltenyi Biotec). The assay was performed with IL-6 and TNF-α beads according to the manufacturer's instructions.
Statistical analysis
Statistical analyses were performed using GraphPad Prism version 7 (GraphPad Software, San Diego, CA). The data were analyzed for Gaussian distribution using a Shapiro-Wilk normality test. The data were further analyzed using a one-way ANOVA and Tukey's post hoc analysis if normality tests were passed; if not, a Kruskal-Wallis test and Dunn's post hoc analysis was used. A two-way ANOVA was used to compare the effects of different concentrations of soluble R848 and PEG-R848.
Results
R848 activates adult and neonatal primary DCs
Three adjuvants: Alum, MPLA, and R848 were analyzed for their immune-stimulatory activities by incubating cDC2 and pDCs together with or without adjuvant. The number of CD1c+CD80+ double-positive cells increased from 9% to 71% after stimulation with R848, whereas alum and MPLA only exerted marginal effects, i.e., 19% and 15% positive cells, respectively (Figure 1A). Additionally, stimulation with R848 increased the expression of CD83 from 16 to 59% on adult cDC2 (Figure 1B). Similar trends were observed after the incubation of neonatal DCs with R848 (Figures 1C,D). R848 stimulation also led to a substantial upregulation of CD80 expression on adult and neonatal pDCs, i.e., the number positive cells increased by 40–50%, whereas stimulation with alum or MPLA had only minor effects (Figures 2A,C). The upregulation of CD83 after R848 stimulation was not as pronounced but the amount of CD123+CD83+ double-positive cells still increased two-fold on adult and 3.3-fold on neonatal cells (Figures 2B,D). In addition to increased activation marker expression, R848 stimulated a substantiate secretion of IL-6 and TNF-α in both adult and neonatal DCs. Alum and MPLA, however, failed to induce any cytokine production (Figure 3).
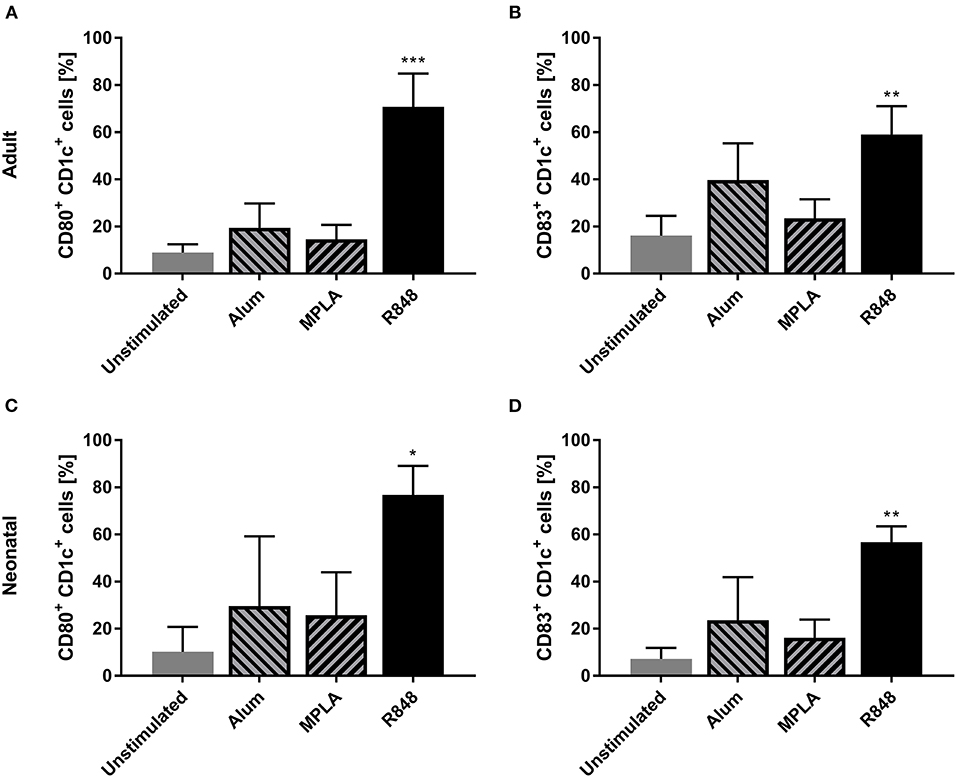
Figure 1. Effect of different adjuvants on CD1c+ cDC2 activation. Adult (A,B) and neonatal (C,D) DCs were incubated with or without alum, MPLA or R848 for 20 h. Flow cytometric analysis of CD80 (A,C) and CD83 (B,D) expression by CD1c+ DCs after stimulation. Data represent the means ± SD (n = 3). Significantly greater than unstimulated control: *P < 0.05; **P < 0.001; ***P < 0.005 (one-way ANOVA).
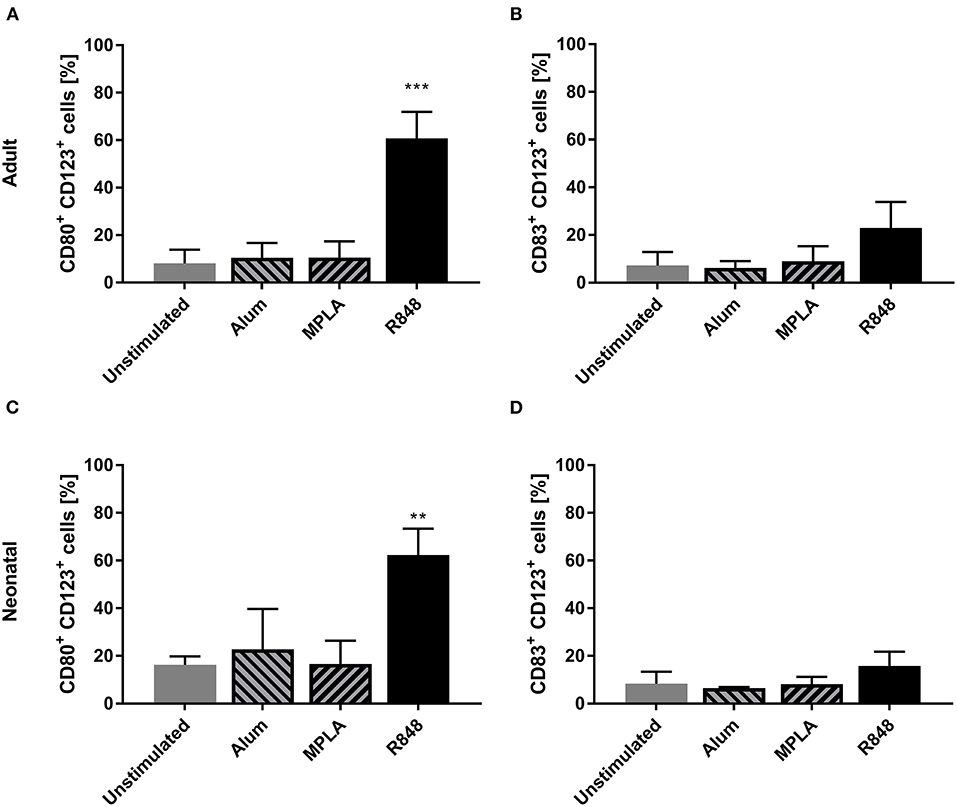
Figure 2. Effect of different adjuvants on CD123+ pDC activation. Adult (A,B) and neonatal (C,D) DCs were incubated with or without alum, MPLA or R848 for 20 h. Flow cytometric analysis of CD80 (A,C) and CD83 (B,D) expression by CD1c+ DCs after stimulation. Data represent the means ± SD (n = 3). Significantly greater than unstimulated control: **P < 0.001; ***P < 0.005 (one-way ANOVA).
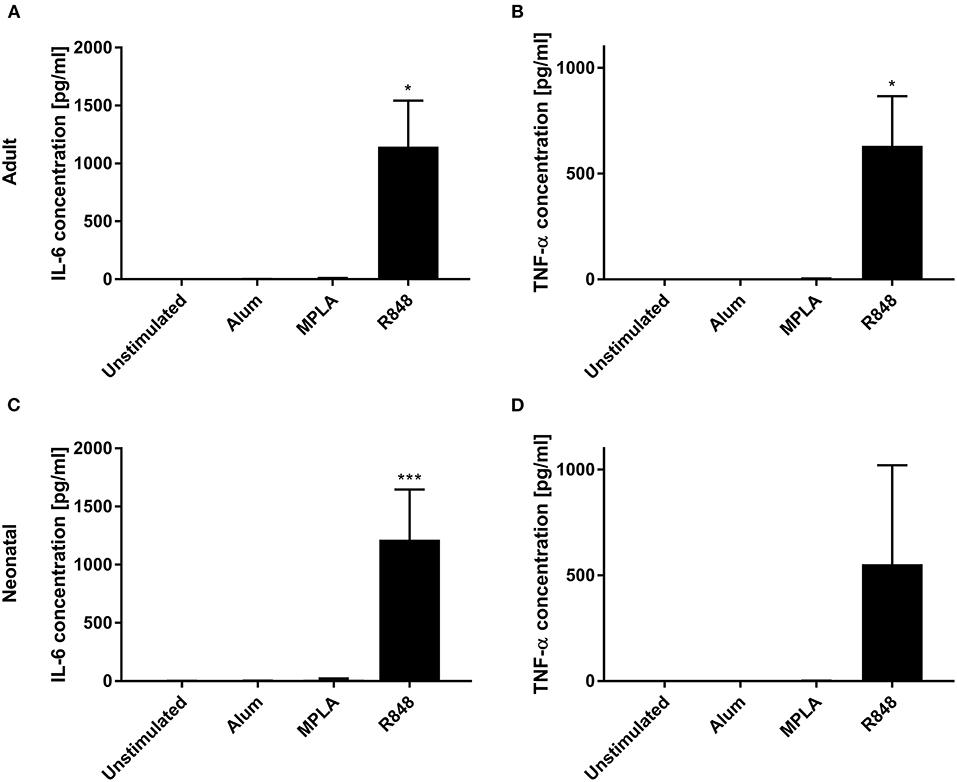
Figure 3. Effect of different adjuvants on DC cytokine secretion. Analysis of IL-6 (A,C) and TNF-α (B,D) secretion by adult (A,B) and neonatal (C,D) DCs after incubation with or without alum, MPLA or R848 for 20 h. Data represent the means ± SD (n = 3). Significantly more than unstimulated: *P < 0.05; ***P < 0.005 [(A,B) one-way ANOVA; (C,D) Kruskal-Wallis test].
PEGylation of R848 does not abrogate its stimulatory activity
To enable the quantification of R848 in OVA NCs and prevent diffusion after encapsulation, R848 was conjugated to a polyethylene glycol (PEG) chain (24). The impact of PEGylation on PEG-R848 was evaluated at multiple concentrations (Figure 4). DCs incubated with lower concentrations of PEG-R848 exhibited an approximately ten-fold decrease in activation relative to cells incubated with the same concentration of R848. Especially at the lowest concentration (10 ng/ml), neither CD1c+ nor CD123+ cells showed any increase in activation relative to the negative control. At the highest concentration tested (1 μg/ml), however, there was no detectable difference between PEG-R848 and R848. Coupling PEG-R848 to Rhodamine B (RhB) made it possible to directly correlate activation marker expression and the internalization of PEG-R848 by cells (Supplementary Figure S2). At 10 ng/ml, neither CD1c+ nor CD123+ RhB+ cells were measurable. At 100 ng/ml the number increased to 13.3% and 54.8%, respectively. At 1 μg/ml, nearly all cells were RhB+.
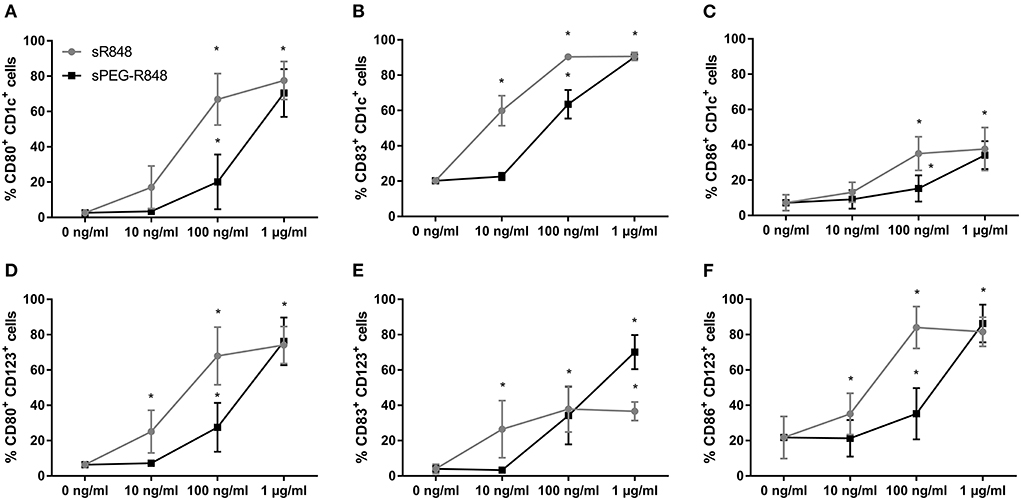
Figure 4. R848 retains stimulatory activity after PEGylation. Flow cytometric analysis of adult CD1c+ (A–C) and CD123+ (D–F) DCs after stimulation with different concentrations of R848 and PEG-R848 for 20h. CD80- (A,D), CD83- (B,E) and CD86-expressing (C,F) cells were evaluated. Data represent the means ± SD (n = 3). Significantly greater than cells incubated with 0 ng/ml: *P < 0.05 (two-way ANOVA).
OVA-PEG-R848 NCs serve as an adjuvant-antigen co-delivery system for DC activation
Co-delivery of soluble adjuvants and antigens is plagued by their rapid dissociation (25). To circumvent this issue, PEG-R848 was encapsulated in OVA NCs (OVA-PEG-R848 NCs), and the capacities of PEG-R848 and OVA-PEG-R848 NCs to activate DCs were assessed and compared. The average size of OVA NCs was 240 nm in toluene and 160 nm in water with a zeta potential of −26.4 (Supplementary Table S1). The observed encapsulation efficiency of OVA-PEG-R848 NCs was approximately 70% (± 5%) of payload retained in NCs, similar to our previous reported data (Supplementary Figure S3A) (24). Moreover, OVA NCs did not show signs of payload leakage after transferring them to water and washing away any non-encapsulated molecules (Supplementary Figure S3B).
DCs were incubated with OVA-PEG-R848 NCs (effective 500 ng/ml R848 concentration), empty OVA NCs or soluble PEG-R848 in equimolar concentrations. Both adult and neonatal CD1c+ cells were significantly activated by incubation with OVA-PEG-R848 NCs (Figure 5). After stimulation, 59% of adult and 61% of neonatal DCs expressed cell surface CD80, compared to 7–8% for the negative control (Figures 5A,E). CD83 expression by both adult and neonatal DCs also increased more than two-fold (Figures 5B,F). Cells incubated with empty NCs, in contrast, showed only a negligible increase in marker expression, clearly demonstrating adjuvant-specific DC activation.
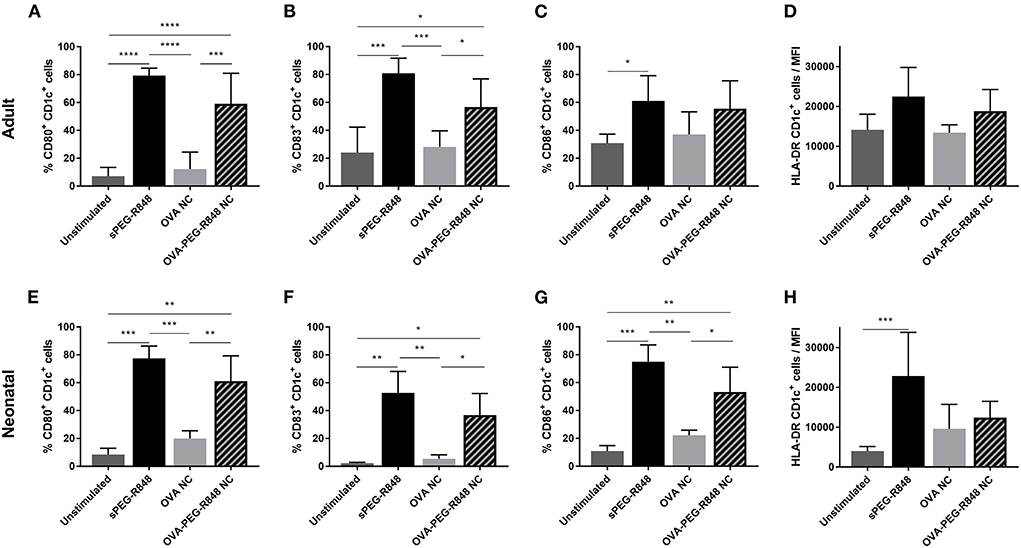
Figure 5. Stimulation of CD1c+ cDC2 by OVA-PEG-R848 NCs. Flow cytometric analysis of adult (A–D) and neonatal (E–H) CD1c+ DCs after incubation with soluble PEG-R848, empty OVA NCs or OVA-PEG-R848 NCs. Frequency of CD80+ (A,E), CD83+ (B,F) and CD86+ (C,G) cells as well as HLA-DR mean fluorescent intensity (D,H) was evaluated. Data represent the means ± SD [(A–D), n = 5; (E–H), n = 3). Significantly different: *P < 0.05; **P < 0.01; ***P < 0.005; ****P < 0.0001 (one-way ANOVA).
Activation of CD123+ cells was not as pronounced as for CD1c+ DCs, but a significant increase in activated cells could still be measured (Figure 6). The expression of CD80 increased from 5 to 37% in adult and 1% to 19% in neonatal DCs (Figures 6A,E). Similarly, the number of CD83+ cells increased from 5–8% to 22% for both adult and neonatal cells (Figures 6B,F). Strikingly, the HLA-DR mean-fluorescent intensity (MFI) increased substantially more in the CD123+ (Figures 6D,H), than the CD1c+, population (Figures 5D,H).
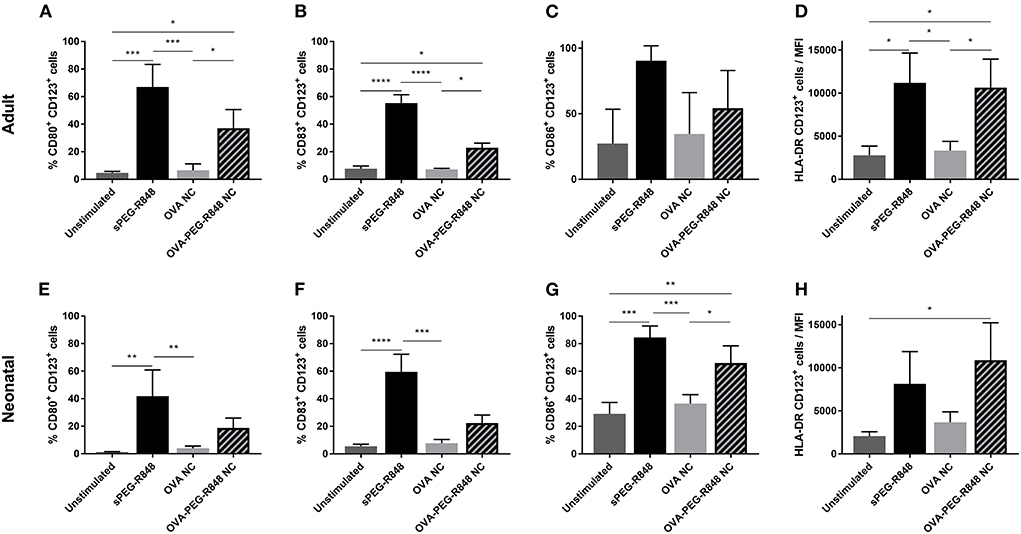
Figure 6. Stimulation of CD123+ pDCs by OVA-PEG-R848 NCs. Flow cytometric analysis of adult (A–D) and neonatal (E–H) CD123+ DCs after incubation with soluble PEG-R848, empty OVA NCs or OVA-PEG-R848 NCs. CD80+ (A,E), CD83+ (B,F) and CD86+ (C,G) cells, and HLA-DR mean fluorescent intensity (D,H) were evaluated. Data represent the means ± SD (n = 3). Significantly different: *P < 0.05; **P < 0.01; ***P < 0.005; ****P < 0.0001 (one-way ANOVA).
For all DC populations and markers tested, soluble PEG-R848 treatment resulted in a higher activation than treatment with OVA-PEG-R848 NCs. This was principally due to significantly greater uptake of soluble PEG-R848 (Supplementary Figure S4). Only 54–64% of CD1c+ DCs were RhB+ after incubation with OVA-PEG-R848 NCs compared to 98% of cells incubated with soluble PEG-R848 (Supplementary Figures S4A,C). The difference was even more pronounced when assessing uptake by CD123+ cells exhibiting a >70% reduction in RhB+ cells (Supplementary Figures S4B,D). After normalizing the expression of CD80 and CD83 in terms of RhB+ levels of OVA-PEG-R848 stimulated cells, and therefore to the amount of ingested PEG-R848, it was evident that stimulation with OVA-PEG-R848 led to slightly higher up-regulation compared to equal amounts of ingested, soluble adjuvant for both the CD1c+ and CD123+ DC populations (Supplementary Figure S5).
Analysis of cell culture supernates showed that OVA-PEG-R848 NCs induced an increase in IL-6 and, especially, TNF-α production (Figure 7). Adult and neonatal cells incubated with OVA-PEG-R848 NCs secreted even more TNF-α than cells incubated with soluble PEG-R848 (Figures 7A,C). Additionally, IL-6 secretion by adult cells was slightly increased compared to the positive control (Figure 7B).
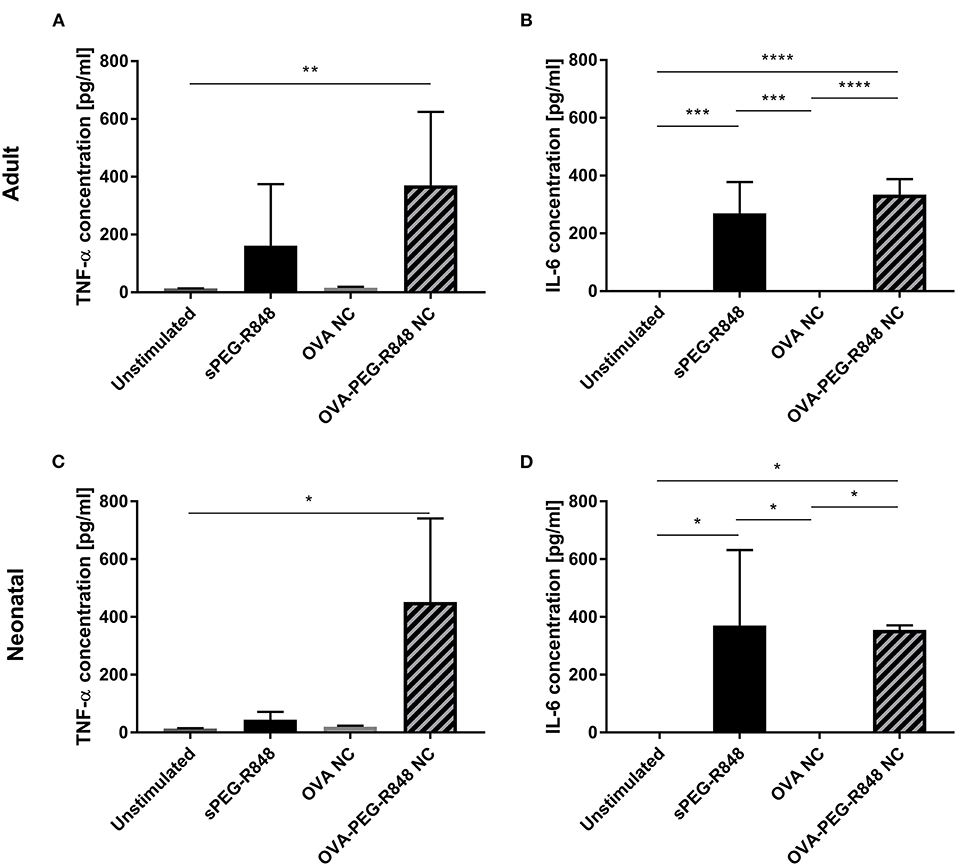
Figure 7. PEG-R848 encapsulation increases cytokine secretion. Analysis of TNF-α (A,C) and IL-6 (B,D) secretion by adult (A,B) and neonatal (C,D) DCs after incubation with soluble PEG-R848, empty OVA NCs or OVA-PEG-R848 NCs. Data represent the means ± SD (n = 3). Significantly different: *P < 0.05; **P < 0.01; ***P < 0.005; ****P < 0.0001 (one-way ANOVA).
Discussion
In 2019, infectious diseases accounted for roughly half of the deaths in children under the age of five; pneumonia and diarrhea alone accounted for 23% of total deaths (1). Many of these cases could be preventable if vaccines, administered during the first weeks of life, elicited a more robust immune response. Most routine vaccines are administered in a three-dose regimen at 2, 4 and 6 months after birth to achieve maximum protection. This, however, leaves children vulnerable to infectious diseases in the first 6 months of life (26). The development and administration of more potent adjuvants could increase vaccine immunogenicity and infant protection. R848, the focus of this study, is a TLR7/8 agonist, which is currently approved for topical use to treat skin lesions and cutaneous T cell lymphoma (27, 28).
Initial experiments showed that R848 was a much more potent adjuvant than either alum or MPLA. Noteworthy, so far most of the studies reporting stimulatory capacities of MPLA and alum were performed in artificially generated DCs (moDCs) (29–31). In this context, primary DCs represent more realistic functional characteristics for the evaluation and screening of potential adjuvants for vaccine development, especially in pediatrics. In this regard, adult primary DCs stimulated with R848 exhibited significantly greater expression of co-stimulatory receptors, CD80 and CD83, and increased secretion of pro-inflammatory cytokines, IL-6 and TNF-α. R848 also activated neonatal DCs isolated from cord blood, confirming the reports of other groups (32, 33).
The rapid dissociation of small molecules co-delivered with antigen upon injection is a major issue (25). Moreover, systemic administration of R848 alone is linked to side effects (34, 35). Encapsulation of R848 in full-protein NCs solves both issues simultaneously. Previously, OVA NCs with three varying degrees of crosslinking were compared for their abilities to release their cargo and subsequently activate adult CD1c+ DCs. It was shown that NC crosslinking density directly influences the release of cargo and that only low-crosslinked NCs successfully activate primary DCs. PEGylation of R848, however, was crucial to avoid diffusion of the small molecule out of low-crosslinked NCs (24), In addition, conjugating RhB to PEG-R848 made it possible to precisely define the amount of encapsulated R848 inside OVA NCs and to measure the number of adjuvant-ingesting cells. PEGylation reduced the effect of R848 by approximately ten-fold when used at lower concentrations to stimulate both CD1c+ and CD123+ DC populations. This difference was no longer observed at the highest concentration tested, a finding that correlated directly with the percentage of RhB+ DCs. OVA-PEG-R848 NCs activated both adult and neonatal primary DCs. CD80, CD83 and CD86 expression levels were elevated significantly following NC stimulation. Notably, incubation with empty OVA NCs did not affect marker expression, confirming the adjuvant-specific effect. Furthermore, OVA-PEG-R848 NCs treatment stimulated the secretion of pro-inflammatory cytokines, i.e., IL-6 and TNF-α, markedly.
It is worth to mention that the study presented here, together with our previous studies regarding this nanosystem (24), provide data supporting the consequent evaluation and characterization of this nanoformulation in vivo. While this study yields answers to relevant immunological questions, such as the possibility to elicit a robust immune stimulation of both primary adult and neonatal DCs, consecutive in vivo studies would need to be performed to provide a complete pre-clinical overview of this nanovaccine platform.
Previous in vitro and in vivo studies showed similar results involving R848 encapsulated in different carrier systems (36–38). In a neonatal context Dowling et al. were able to show the potential of TLR7/8 agonists by encapsulating the agonist CL075. Stimulation of monocyte-derived dendritic cells (moDCs) with CL075 nanoparticles upregulated costimulatory markers and led to an increased secretion of pro-inflammatory cytokines (39). The present study, however, is one of the first to evaluate the stimulatory effects of encapsulated R848 on neonatal primary DCs, providing further proof that R848 is a more potent adjuvant, relative to alum or MPLA, to incorporate into neonatal vaccines. While moDCs and cDCs share a lot of similarities, e.g., the upregulation of costimulatory markers and secretion of IL12p70 upon activation, cDCs overall possess higher migratory as well as T cell activating capabilities (40–42). Further evidence regarding the potential of R848 was also provided in recent studies investigating the effect of conjugating R848 to inactivated influenza virus. Non-human primates vaccinated with conjugated virus showed an increase in activated B cells, DCs and macrophages, as well as elevated antibody titers still measurable 6 months after vaccination (43–45).
HBV vaccine, requiring the administration of three doses over a 6-month period to achieve full protection, could benefit immensely from the increased stimulatory activity of R848 (16). Vaccinating infants three times can be challenging especially in low-income countries where HBV tends to be most prevalent (19, 46). Licensed HBV vaccines are currently adjuvanted with aluminum-based agents (47, 48). Unfortunately, aluminum salts only elicit a TH2-type immune response, however neonatal immunity is already naturally skewed toward this direction (5, 49, 50). Therefore, alum-based vaccines do not sufficiently activate the neonatal immune system nor do they generate specific CD4+ and CD8+ T cells needed to prevent chronic infections (51, 52). This is a significant issue insofar as 90% of HBV infections that occur following mother-to-infant transmission develop into chronic disease (53). It is relevant to note, therefore, that nanocapsules composed of HBV surface antigen (HBsAg) and functionalized with MPLA bear the capacity to generate a specific T cell response in vitro (54).
In summary, R848 is a potent adjuvant capable of activating adult and neonatal primary DCs in vitro. PEGylation of R848 only exerted a minor impact on its effect; labeling with RhB made it possible to track the number of adjuvant-ingesting cells and quantify the amount of R848 inside NCs. Stimulation with functionalized OVA NCs led to a marked increase in the expression of activation markers and secretion of pro-inflammatory cytokines by conventional and plasmacytoid DCs. This study is among the first to demonstrate the capacity of encapsulated R848 to activate neonatal DCs. This finding supports the potential incorporation of R848 as adjuvant in neonatal vaccines, making them more effective in eliciting a robust immune response.
Data availability statement
The datasets generated and/or analysed during the current study are available from the corresponding author on reasonable request.
Ethics statement
The studies involving human participants were reviewed and approved by Landesärztekammer Rheinland-Pfalz. Written informed consent to participate in this study was provided by the participants' legal guardian/next of kin.
Author contributions
SW, MM, MF, MC, HT-A, KL, and SG contributed to the study conception and design. Material preparation and data collection were performed by SW, MM, FB, LP, and MF. Data analysis was performed by SW and MF. The first draft of the manuscript was written by SW and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Funding
This project was funded by the Else Kröner-Fresenius Stiftung (EKFS) grant 2016_A206 and by the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation)—Project-ID 213555243—SFB1066.
Acknowledgments
We are grateful to Dr. Stephen H. Gregory (Providence, RI, USA) for his help writing and editing this manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fped.2022.966113/full#supplementary-material
References
1. Perin J, Mulick A, Yeung D, Villavicencio F, Lopez G, Strong KL, et al. Global, regional, and national causes of under-5 mortality in 2000-19: an updated systematic analysis with implications for the Sustainable Development Goals. Lancet Child Adolesc Health. (2022) 6:106–15. doi: 10.1016/S2352-4642(21)00311-4
2. Sharrow D, Hug L, You D, Alkema L, Black R, Cousens S, et al. Global, regional, and national trends in under-5 mortality between 1990 and 2019 with scenario-based projections until 2030: a systematic analysis by the UN Inter-agency Group for Child Mortality Estimation. Lancet Glob Health. (2022) 10:e195–206. doi: 10.1016/S2214-109X(21)00515-5
3. Makhseed M, Raghupathy R, Azizieh F, Omu A, Al-Shamali E, Ashkanani L. Th1 and Th2 cytokine profiles in recurrent aborters with successful pregnancy and with subsequent abortions. Hum Reprod. (2001) 16:2219–26. doi: 10.1093/humrep/16.10.2219
4. Morein B, Blomqvist G, Hu K. Immune responsiveness in the neonatal period. J Comp Pathol. (2007) 137 Suppl 1:S27–31. doi: 10.1016/j.jcpa.2007.04.008
5. Adkins B. Heterogeneity in the CD4 T cell compartment and the variability of neonatal immune responsiveness. Curr Immunol Rev. (2007) 3:151–9. doi: 10.2174/157339507781483496
6. Godfrey WR, Spoden DJ, Ge YG, Baker SR, Liu B, Levine BL, et al. Cord blood CD4(+)CD25(+)-derived T regulatory cell lines express FoxP3 protein and manifest potent suppressor function. Blood. (2005) 105:750–8. doi: 10.1182/blood-2004-06-2467
7. Thome JJC, Bickham KL, Ohmura Y, Kubota M, Matsuoka N, Gordon C, et al. Early-life compartmentalization of human T cell differentiation and regulatory function in mucosal and lymphoid tissues. Nat Med. (2016) 22:72–7. doi: 10.1038/nm.4008
8. Burl S, Townend J, Njie-Jobe J, Cox M, Adetifa UJ, Touray E, et al. Age-dependent maturation of Toll-like receptor-mediated cytokine responses in Gambian infants. PLoS ONE. (2011) 6:e18185. doi: 10.1371/journal.pone.0018185
9. Nguyen M, Leuridan E, Zhang T, De Wit D, Willems F, Van Damme P, et al. Acquisition of adult-like TLR4 and TLR9 responses during the first year of life. PLoS ONE. (2010) 5:e10407. doi: 10.1371/journal.pone.0010407
10. Hunt DW, Huppertz HI, Jiang HJ, Petty RE. Studies of human cord blood dendritic cells: evidence for functional immaturity. Blood. (1994) 84:4333–43. doi: 10.1182/blood.V84.12.4333.bloodjournal84124333
11. Marchant A, Goetghebuer T, Ota MO, Wolfe I, Ceesay SJ, De Groote D, et al. Newborns develop a Th1-type immune response to Mycobacterium bovis bacillus Calmette-Guérin vaccination. J Immunol. (1999) 163:2249–55.
12. Ravn P, Boesen H, Pedersen BK, Andersen P. Human T cell responses induced by vaccination with Mycobacterium bovis bacillus Calmette-Guérin. J Immunol. (1997) 158:1949–55.
13. Napolitani G, Rinaldi A, Bertoni F, Sallusto F, Lanzavecchia A. Selected TLR agonist combinations synergistically trigger a TH1 polarizing program in dendritic cells. Nat Immunol. (2005) 6:769–76. doi: 10.1038/ni1223
14. Hémont C, Neel A, Heslan M, Braudeau C, Josien R. Human blood mDC subsets exhibit distinct TLR repertoire and responsiveness. J Leukoc Biol. (2013) 93:599–609. doi: 10.1189/jlb.0912452
15. Bao M, Liu YJ. Regulation of TLR7/9 signaling in plasmacytoid dendritic cells. Protein Cell. (2013) 4:40–52. doi: 10.1007/s13238-012-2104-8
16. Birth-18 Years Immunization Schedule. CDC. (2021). Available online at: https://www.cdc.gov/vaccines/schedules/hcp/imz/child-adolescent.html (accessed January 20, 2022).
17. Wong VC, Ip HM, Reesink HW, Lelie PN, Reerink-Brongers EE, Yeung CY, et al. Prevention of the HBsAg carrier state in newborn infants of mothers who are chronic carriers of HBsAg and HBeAg by administration of hepatitis-B vaccine and hepatitis-B immunoglobulin. Double-blind randomised placebo-controlled study. Lancet. (1984) 1:921–6. doi: 10.1016/S0140-6736(84)92388-2
18. Xu ZY, Liu CB, Francis DP, Purcell RH, Gun ZL, Duan SC, et al. Prevention of perinatal acquisition of hepatitis B virus carriage using vaccine: preliminary report of a randomized, double-blind placebo-controlled and comparative trial. Pediatrics. (1985) 76:713–8. doi: 10.1542/peds.76.5.713
19. Morris MC, Surendran N. Neonatal vaccination: challenges and intervention strategies. Neonatology. (2016) 109:161–9. doi: 10.1159/000442460
20. Franco E, Bagnato B, Marino MG, Meleleo C, Serino L, Zaratti L. Hepatitis B: Epidemiology and prevention in developing countries. World J Hepatol. (2012) 4:74–80. doi: 10.4254/wjh.v4.i3.74
21. Luchner M, Reinke S, Milicic ATLR. Agonists as vaccine adjuvants targeting cancer and infectious diseases. Pharmaceutics. (2021) 13:142. doi: 10.3390/pharmaceutics13020142
22. Szarewski AHPV. vaccine: Cervarix. Expert Opin Biol Ther. (2010) 10:477–87. doi: 10.1517/14712591003601944
23. Campbell JD. Development of the CpG adjuvant 1018: a case study. Methods Mol Biol. (2017) 1494:15–27. doi: 10.1007/978-1-4939-6445-1_2
24. Machtakova M, Wirsching S, Gehring S, Landfester K, Thérien-Aubin H. Controlling the semi-permeability of protein nanocapsules influences the cellular response to macromolecular payloads. J Mater Chem B. (2021) 9:8389–98. doi: 10.1039/D1TB01368H
25. Sands H, Gorey-Feret LJ, Cocuzza AJ, Hobbs FW, Chidester D, Trainor GL. Biodistribution and metabolism of internally 3H-labeled oligonucleotides. I Comparison of a phosphodiester and a phosphorothioate. Mol Pharmacol. (1994) 45:932–43.
26. Whittaker E, Goldblatt D, McIntyre P, Levy O. Neonatal immunization: rationale, current state, and future prospects. Front Immunol. (2018) 9. doi: 10.3389/fimmu.2018.00532
27. Szeimies RM, Bichel J, Ortonne JP, Stockfleth E, Lee J, Meng TC, et al. phase II dose-ranging study of topical resiquimod to treat actinic keratosis. Br J Dermatol. (2008) 159:205–10. doi: 10.1111/j.1365-2133.2008.08615.x
28. Rook AH, Gelfand JM, Gelfand JC, Wysocka M, Troxel AB, Benoit B, et al. Topical resiquimod can induce disease regression and enhance T-cell effector functions in cutaneous T-cell lymphoma. Blood. (2015) 126:1452–61. doi: 10.1182/blood-2015-02-630335
29. Kolanowski STHM, Lissenberg-Thunnissen SN, Emal D, van Ham SM, Ten Brinke A. Monophosphoryl lipid A-induced pro-inflammatory cytokine expression does not require CD14 in primary human dendritic cells. Inflamm Res. (2016) 65:449–58. doi: 10.1007/s00011-016-0927-0
30. van Haren SD, Dowling DJ, Foppen W, Christensen D, Andersen P, Reed SG, et al. Age-specific adjuvant synergy: dual TLR7/8 and mincle activation of human newborn dendritic cells enables Th1-polarization. J Immunol. (2016) 197:4413–24. doi: 10.4049/jimmunol.1600282
31. Surendran N, Simmons A, Pichichero METLR. agonist combinations that stimulate Th type I polarizing responses from human neonates. Innate Immun. (2018) 24:240–51. doi: 10.1177/1753425918771178
32. Ahonen CL, Gibson SJ, Smith RM, Pederson LK, Lindh JM, Tomai MA, et al. Dendritic cell maturation and subsequent enhanced T-cell stimulation induced with the novel synthetic immune response modifier R-848. Cell Immunol. (1999) 197:62–72. doi: 10.1006/cimm.1999.1555
33. Schüller S, Wisgrill L, Sadeghi K, Gindl E, Helmer H, Husslein P, et al. The TLR-specific adjuvants R-848 and CpG-B endorse the immunological reaction of neonatal antigen-presenting cells. Pediatr Res. (2016) 80:311–8. doi: 10.1038/pr.2016.71
34. Hasham MG, Baxan N, Stuckey DJ, Branca J, Perkins B, Dent O, et al. Systemic autoimmunity induced by the TLR7/8 agonist Resiquimod causes myocarditis and dilated cardiomyopathy in a new mouse model of autoimmune heart disease. Dis Model Mech. (2017) 10:259–70. doi: 10.1242/dmm.027409
35. Pockros PJ, Guyader D, Patton H, Tong MJ, Wright T, McHutchison JG, et al. Oral resiquimod in chronic HCV infection: safety and efficacy in 2 placebo-controlled, double-blind phase IIa studies. J Hepatol. (2007) 47:174–82. doi: 10.1016/j.jhep.2007.02.025
36. Da Silva CG, Camps MGM Li TMWY, Chan AB, Ossendorp F, Cruz LJ. Co-delivery of immunomodulators in biodegradable nanoparticles improves therapeutic efficacy of cancer vaccines. Biomaterials. (2019) 220:119417. doi: 10.1016/j.biomaterials.2019.119417
37. Paßlick D, Piradashvili K, Bamberger D, Li M, Jiang S, Strand D, et al. Delivering all in one: Antigen-nanocapsule loaded with dual adjuvant yields superadditive effects by DC-directed T cell stimulation. J Control Release. (2018) 289:23–34. doi: 10.1016/j.jconrel.2018.09.008
38. Wagner J, Gößl D, Ustyanovska N, Xiong M, Hauser D, Zhuzhgova O, et al. Mesoporous silica nanoparticles as pH-responsive carrier for the immune-activating drug resiquimod enhance the local immune response in mice. ACS Nano. (2021) 15:4450–66. doi: 10.1021/acsnano.0c08384
39. Dowling DJ, Scott EA, Scheid A, Bergelson I, Joshi S, Pietrasanta C, et al. Toll-like receptor 8 agonist nanoparticles mimic immunomodulating effects of the live BCG vaccine and enhance neonatal innate and adaptive immune responses. J Allergy Clin Immunol. (2017) 140:1339–50. doi: 10.1016/j.jaci.2016.12.985
40. Ten Brinke A, Karsten ML, Dieker MC, Zwaginga JJ, van Ham SM. The clinical grade maturation cocktail monophosphoryl lipid A plus IFNgamma generates monocyte-derived dendritic cells with the capacity to migrate and induce Th1 polarization. Vaccine. (2007) 25:7145–52. doi: 10.1016/j.vaccine.2007.07.031
41. Osugi Y, Vuckovic S, Hart DNJ. Myeloid blood CD11c(+) dendritic cells and monocyte-derived dendritic cells differ in their ability to stimulate T lymphocytes. Blood. (2002) 100:2858–66. doi: 10.1182/blood.V100.8.2858
42. Guilliams M, van de Laar LA. Hitchhiker's guide to myeloid cell subsets: practical implementation of a novel mononuclear phagocyte classification system. Front Immunol. (2015) 6:406. doi: 10.3389/fimmu.2015.00406
43. Holbrook BC, Aycock ST, Machiele E, Clemens E, Gries D, Jorgensen MJ, et al. An R848 adjuvanted influenza vaccine promotes early activation of B cells in the draining lymph nodes of non-human primate neonates. Immunology. (2018) 153:357–67. doi: 10.1111/imm.12845
44. Clemens EA, Holbrook BC, Kanekiyo M, Yewdell JW, Graham BS, Alexander-Miller MA. An R848-conjugated influenza virus vaccine elicits robust immunoglobulin G to hemagglutinin stem in a newborn nonhuman primate model. J Infect Dis. (2021) 224:351–9. doi: 10.1093/infdis/jiaa728
45. Holbrook BC, D'Agostino RB, Tyler Aycock S, Jorgensen MJ, Hadimani MB, Bruce King S, et al. Adjuvanting an inactivated influenza vaccine with conjugated R848 improves the level of antibody present at 6months in a nonhuman primate neonate model. Vaccine. (2017) 35:6137–42. doi: 10.1016/j.vaccine.2017.09.054
46. Schweitzer A, Horn J, Mikolajczyk RT, Krause G, Ott JJ. Estimations of worldwide prevalence of chronic hepatitis B virus infection: a systematic review of data published between 1965 and 2013. Lancet. (2015) 386:1546–55. doi: 10.1016/S0140-6736(15)61412-X
47. Gołoś A, Lutyńska A. Aluminium-adjuvanted vaccines–a review of the current state of knowledge. Przegl Epidemiol. (2015) 69:731–4.
48. He P, Zou Y, Hu Z. Advances in aluminum hydroxide-based adjuvant research and its mechanism. Hum Vaccin Immunother. (2015) 11:477–88. doi: 10.1080/21645515.2014.1004026
49. Marrack P, McKee AS, Munks MW. Towards an understanding of the adjuvant action of aluminium. Nat Rev Immunol. (2009) 9:287–93. doi: 10.1038/nri2510
50. Kool M, Fierens K, Lambrecht BN. Alum adjuvant: some of the tricks of the oldest adjuvant. J Med Microbiol. (2012) 61:927–34. doi: 10.1099/jmm.0.038943-0
51. Das A, Maini MK. Innate and adaptive immune responses in hepatitis B virus infection. Dig Dis. (2010) 28:126–32. doi: 10.1159/000282075
52. Jafarzadeh A, Shokri F. TH1 and TH2 responses are influenced by HLA antigens in healthy neonates vaccinated with recombinant hepatitis B vaccine. Iran J Allergy Asthma Immunol. (2012) 11:308–15.
53. Orlando R, Foggia M, Maraolo AE, Mascolo S, Palmiero G, Tambaro O, et al. Prevention of hepatitis B virus infection: from the past to the future. Eur J Clin Microbiol Infect Dis. (2015) 34:1059–70. doi: 10.1007/s10096-015-2341-x
Keywords: neonatal dendritic cells, conventional dendritic cells, plasmacytoid dendritic cells, vaccine, R848, nanocapsules
Citation: Wirsching S, Machtakova M, Borgans F, Pretsch L, Fichter M, Cacicedo ML, Thérien-Aubin H, Landfester K and Gehring S (2022) OVA-PEG-R848 nanocapsules stimulate neonatal conventional and plasmacytoid dendritic cells. Front. Pediatr. 10:966113. doi: 10.3389/fped.2022.966113
Received: 10 June 2022; Accepted: 17 August 2022;
Published: 13 September 2022.
Edited by:
Anna B. Nilsson, Karolinska Institutet (KI), SwedenReviewed by:
Evan Alexander Scott, Northwestern University, United StatesEun-Ju Ko, Jeju National University, South Korea
Copyright © 2022 Wirsching, Machtakova, Borgans, Pretsch, Fichter, Cacicedo, Thérien-Aubin, Landfester and Gehring. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Stephan Gehring, c3RlcGhhbi5nZWhyaW5nQHVuaS1tYWluei5kZQ==