 Ke Shi
Ke Shi Di Ke
Di Ke Feng Li2†
Feng Li2† Dan Li
Dan Li
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Oncol., 17 April 2025
Sec. Molecular and Cellular Oncology
Volume 15 - 2025 | https://doi.org/10.3389/fonc.2025.1573378
Autophagy-related genes (ATGs) play a crucial role in tumorigenesis and cancer progression. ATG10, a member of the ATG family, has been implicated in various malignancies, including endometrial cancer, hepatocellular carcinoma, acute leukemia, nasopharyngeal carcinoma, gastric cancer and colorectal cancer. Its overexpression is frequently associated with poor prognosis and increased disease progression. ATG10 promotes cancer growth and metastasis by modulating epithelial-mesenchymal transition and cell cycle regulators such as cyclin B1, CDK1 and CDK2. However, its activity can be inhibited by several factors, including DDX10, PTBP1, sodium orthovanadate, podofilox, SIRT6, FAT1, SOX2 and multiple microRNAs (e.g., miR-369-3p, miR-100-3p, miR-27b-3p, miR-197-3p, let-7i-5p and miR-552). This review explores the functional and clinical significance of ATG10 across various cancers, highlighting its potential as a biomarker and therapeutic target.
Cancer progression involves inevitable alterations in the expression of key regulatory factors (1–5). Huang et al. reported that chondroitin sulfate synthase 3 (CHSY3) is highly expressed in gastric cancer tissues, where its overexpression correlates with poor prognosis and advanced T staging. In vitro and in vivo studies confirm that CHSY3 enhances gastric cancer cell proliferation, migration and invasion (3). Similarly, Xiao et al. identified aberrant expression of T cell receptor-associated transmembrane adaptor 1 (TRAT1) in lung adenocarcinoma, where specific mutations are associated with poor prognosis. TRAT1 overexpression suppresses cell viability, migration and invasion while promoting apoptosis. Moreover, it is significantly linked to immune cell infiltration, including B cells, CD8+ T cells and cytotoxic cells, as well as RNA modification processes (4). These findings underscore the critical roles of key genes in cancer progression.
Autophagy-related genes (ATGs) have also been implicated in tumor development and progression (6–10). In non-small cell lung cancer (NSCLC), ATG5 and circ-FOXM1 are significantly upregulated. Suppressing circ-FOXM1 inhibits NSCLC cell viability, migration and autophagy while inducing apoptosis, an effect mediated by circ-FOXM1’s regulation of miR-149-5p to enhance ATG5 expression (9). Among ATGs, ATG10 has garnered increasing research interest due to its abnormal expression in multiple cancers. Elevated ATG10 levels are strongly associated with tumor progression and poor prognosis (11–25). It is overexpressed in endometrial cancer, hepatocellular carcinoma and acute leukemia, where its upregulation correlates with reduced survival and increased disease progression. This review summarizes the molecular mechanisms and clinical significance of ATG10 across cancers, drawing on findings from clinical tissue samples, in vitro experiments and in vivo models. Our analysis highlights ATG10 as a potential biomarker and therapeutic target in oncology.
ATG10 expression is significantly elevated in endometrial cancer, hepatocellular carcinoma, acute leukemia, nasopharyngeal carcinoma, gastric cancer and colorectal cancer compared to normal tissues (11–17). In hepatocellular carcinoma, ATG10 levels are markedly higher in Hep3B, HepG2 and PLC cell lines compared to normal LO2 cells (2). Similarly, colorectal cancer cell lines (HCT116, HT29, KM12C, WiDr, LoVo, SW480, SW48, HCT15, DLD1, RKO and CaCo2) exhibit significantly higher ATG10 expression than CCD841 cells (16). Moreover, in cancer tissues and cells, there is a significant trend of increased ATG10 expression levels (Table 1).
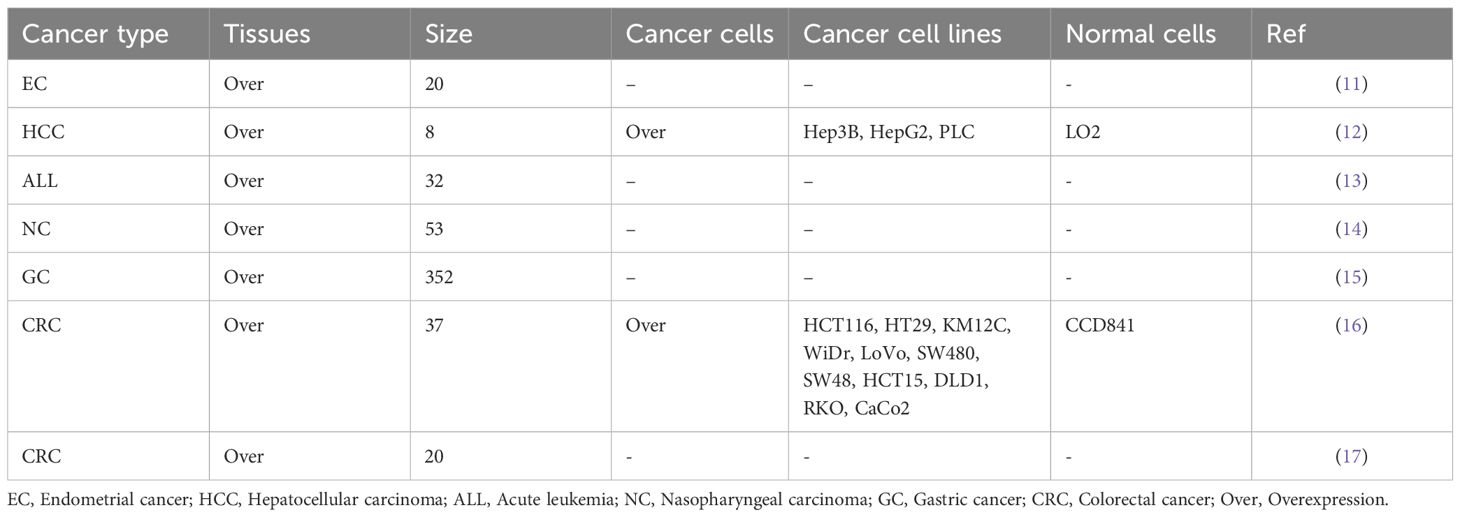
Table 1. ATG10 overexpression in cancer tissues and cells.
Both in vitro and in vivo studies indicate that ATG10 functions as an oncogenic factor in cancer progression (Table 2). ATG10 overexpression enhances the proliferation of endometrial cancer (HEC-1-A), hepatocellular carcinoma (Hep3B and PLC), nasopharyngeal carcinoma (HONE-1, CNE-2 and 5-8F), colorectal cancer (SW480, SW620 and DLD-1), gastric cancer (SGC-7901, MGC-803, GIST-882, GIST-T1, AGS and HGC27) and lung cancer (A549 and H1299) cells. Furthermore, it inhibits apoptosis in colorectal cancer (SW480, SW620 and DLD-1) cells. In vivo experiments confirm that ATG10 overexpression increases tumorigenicity in gastric cancer (SGC-7901) and nasopharyngeal carcinoma (CNE-2) cells in nude mice.
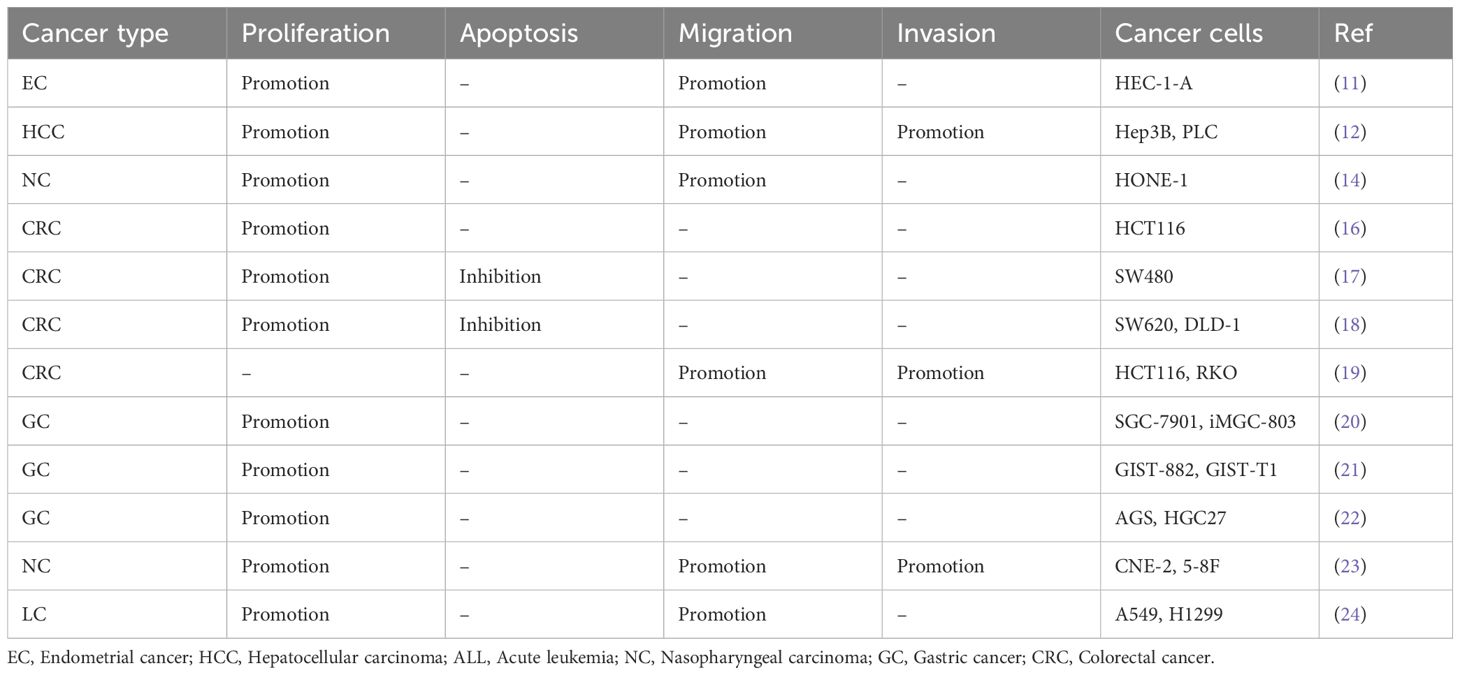
Table 2. In vitro functional characterization of ATG10 in cancer.
ATG10 overexpression has been shown to promote cancer metastasis in vitro (Table 2). Specifically, it enhances the migration and invasion of hepatocellular carcinoma (Hep3B and PLC), colorectal cancer (HCT116 and RKO) and nasopharyngeal carcinoma (CNE-2 and 5-8F) cells (Table 2). Additionally, it promotes the migration of endometrial cancer (HEC-1-A), nasopharyngeal carcinoma (HONE-1) and lung cancer (A549 and H1299) cells (Table 2). Notably, ATG10 overexpression also induces resistance to oxaliplatin in colorectal cancer (SW480) cells (17).
ATG10 promotes cancer growth and metastasis through multiple signaling pathways (Figure 1). Downregulation of ATG10 suppresses hepatocellular carcinoma cell proliferation, migration and invasion by modulating cyclin B1, CDK1 and CDK2 expression (12). In nasopharyngeal carcinoma and ovarian cancer, ATG10 inhibition disrupts the PI3K/AKT signaling pathway and epithelial-mesenchymal transition, respectively (14, 25). ATG10 expression is regulated by various microRNAs, including miR-369-3p, miR-100-3p, miR-27b-3p, miR-197-3p, let-7i-5p and miR-552 (Table 3) and can be suppressed by factors such as DDX10, PTBP1, sodium orthovanadate (SOV), podofilox, SIRT6, FAT1 and SOX2 (18–20, 22, 26–28). For instance, Liu et al. reported that miR-369-3p is downregulated in endometrial cancer, and its overexpression targets ATG10 to inhibit endometrial cancer cell proliferation and migration (11). Similarly, Peng et al. demonstrated that lncRNA ZFAS1 upregulates ATG10 by competitively binding miR-100-3p, thereby promoting nasopharyngeal carcinoma progression via the PI3K/AKT pathway (14).
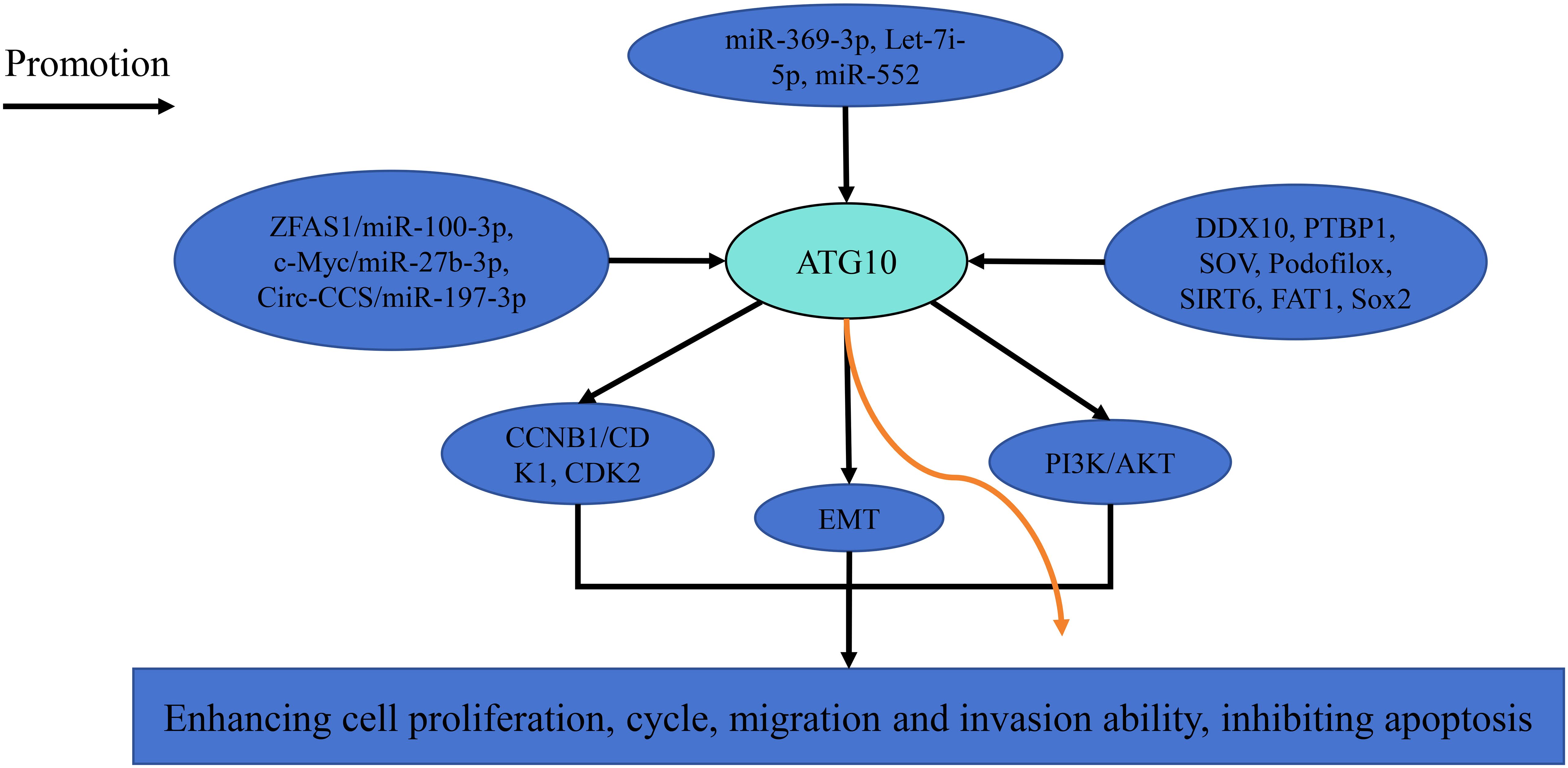
Figure 1. The related pathways of ATG10 in cancer.
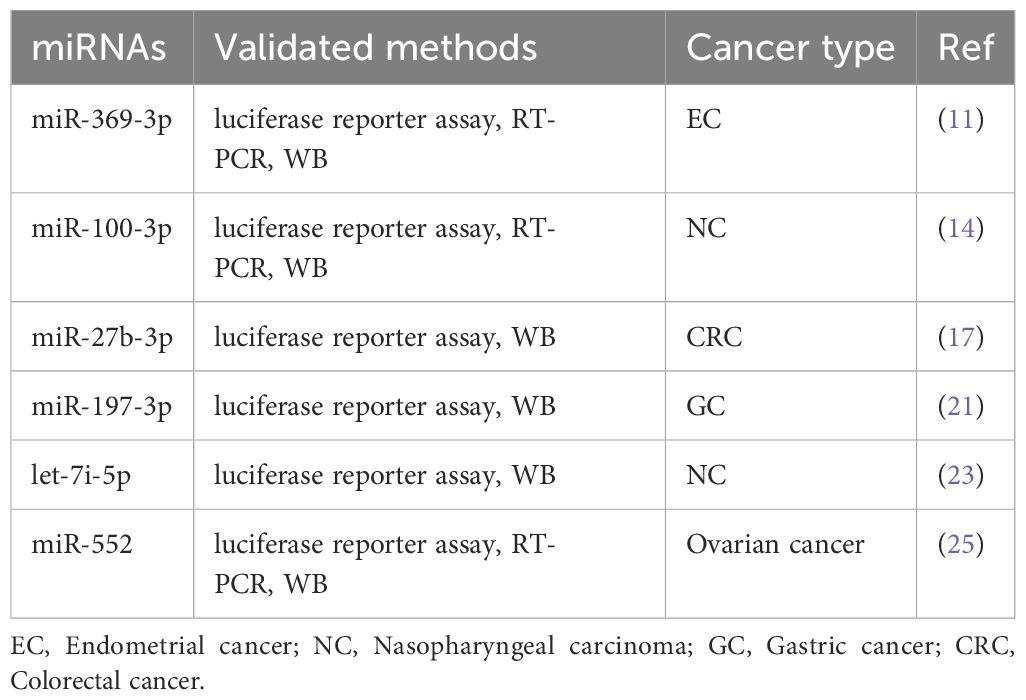
Table 3. ATG10-miRNAs signaling pathways in cancer.
ATG10 overexpression has been associated with poor prognosis and adverse clinicopathological features in gastric cancer, colorectal cancer and acute leukemia, as determined by RT-PCR, immunohistochemistry and western blotting (Table 4). In gastric cancer, elevated ATG10 expression correlates with reduced overall survival, lymph node metastasis and advanced TNM staging (15). Similarly, in colorectal cancer, ATG10 overexpression is linked to poorer overall and disease-specific survival, as well as increased lymphovascular invasion and lymph node metastasis (16). Moreover, in acute leukemia, high ATG10 expression is significantly associated with elevated white blood cell counts (13). These findings underscore ATG10’s potential as a prognostic biomarker and suggest that its inhibition may improve patient survival.

Table 4. Overexpression of ATG10 is associated with poor prognosis and clinical features in patients with cancer.
ATG10 is abnormally overexpressed in multiple cancers, including gastric cancer, colorectal cancer and acute leukemia, and its overexpression is strongly associated with poor prognosis and adverse clinicopathological features. Functionally, ATG10 promotes cancer cell proliferation, migration and invasion, underscoring its potential as a tumor biomarker. Mechanistic studies suggest that ATG10 contributes to tumor development by regulating cell cycle-related proteins, epithelial-mesenchymal transition and interactions with miRNAs. As a potential therapeutic target, ATG10 has garnered increasing attention, with ongoing research exploring the effects of its inhibition on cancer treatment. However, current findings are largely based on in vitro studies, with limited in vivo validation. While preliminary mechanistic insights have been gained, the functional specificity of ATG10 across different cancer types remains unclear. Additionally, clinical studies on ATG10 are scarce, hindering its validation as a reliable biomarker in patient populations. In summary, ATG10 exhibits significant biological functions in cancer progression and holds promise as a novel therapeutic target. However, further systematic in vivo studies and clinical investigations are needed to fully elucidate its role in tumorigenesis and its potential in targeted therapy.
KS: Data curation, Investigation, Methodology, Writing – original draft. DK: Data curation, Investigation, Methodology, Writing – original draft. FL: Data curation, Investigation, Methodology, Writing – original draft. RSS: Methodology, Software, Visualization, Writing – review & editing. TL: Conceptualization, Funding acquisition, Validation, Writing – review & editing. DL: Conceptualization, Funding acquisition, Validation, Writing – review & editing. QXZ: Conceptualization, Funding acquisition, Validation, Visualization, Writing – review & editing.
The author(s) declare that financial support was received for the research and/or publication of this article. This study was funded by the Zunyi City Joint Fund (Zun Shi Ke He HZ Word (2022) No. 244).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Ma H, Kong L, Liu L, Du Y, Zhu X, Wang J, et al. ENO1 contributes to the gemcitabine resistance of pancreatic cancer through the YAP1 signaling pathway. Mol Carcinog. (2024) 63:1221–34. doi: 10.1002/mc.23719
2. Xiang QM, Jiang N, Liu YF, Wang YB, Mu DA, Liu R, et al. Overexpression of SH2D1A promotes cancer progression and is associated with immune cell infiltration in hepatocellular carcinoma via bioinformatics and in vitro study. BMC Cancer. (2023) 23:1005. doi: 10.1186/s12885-023-11315-1
3. Wang H, Zhang J, Wei Z, Chen S, Zheng J, Li Y. The prognostic implications and tumor-promoting functions of CHSY3 in gastric cancer. Front Immunol. (2024) 15:1364979. doi: 10.3389/fimmu.2024.1364979
4. Su Y, Meng L, Ge C, Liu Y, Zhang C, Yang Y, et al. PSMD9 promotes the Malignant progression of hepatocellular carcinoma by interacting with c-Cbl to activate EGFR signaling and recycling. J Exp Clin Cancer Res. (2024) 43:142. doi: 10.1186/s13046-024-03062-3
5. Xiao XY, Guo Q, Tong S, Wu CY, Chen JL, Ding Y, et al. TRAT1 overexpression delays cancer progression and is associated with immune infiltration in lung adenocarcinoma. Front Oncol. (2022) 12:960866. doi: 10.3389/fonc.2022.960866
6. Zhang Y, Yang H, Sun M, He T, Liu Y, Yang X, et al. Alpinumisoflavone suppresses hepatocellular carcinoma cell growth and metastasis via NLRP3 inflammasome-mediated pyroptosis. Pharmacol Rep. (2020) 72:1370–82. doi: 10.1007/s43440-020-00064-8
7. Liu HS, Wang YP, Lin PW, Chu ML, Lan SH, Wu SY, et al. The role of ATG5 gene in tumorigenesis under autophagy deficiency conditions. Kaohsiung J Med Sci. (2024) 40:631–41. doi: 10.1002/kjm2.12853
8. Qin Y, Sun W, Wang Z, Dong W, He L, Zhang T, et al. ATF2-Induced lncRNA GAS8-AS1 Promotes Autophagy of Thyroid Cancer Cells by Targeting the miR-187-3p/ATG5 and miR-1343-3p/ATG7 Axes. Mol Ther Nucleic Acids. (2020) 22:584–600. doi: 10.1016/j.omtn.2020.09.022
9. Wei H, Li L, Zhang H, Xu F, Chen L, Che G, et al. Circ-FOXM1 knockdown suppresses non-small cell lung cancer development by regulating the miR-149-5p/ATG5 axis. Cell Cycle. (2021) 20:166–78. doi: 10.1080/15384101.2020.1867780
10. Wang J, Dong Z, Sheng Z, Cai Y. Hypoxia-induced PVT1 promotes lung cancer chemoresistance to cisplatin by autophagy via PVT1/miR-140-3p/ATG5 axis. Cell Death Discovery. (2022) 8:104. doi: 10.1038/s41420-022-00886-w
11. Liu P, Ma C, Wu Q, Zhang W, Wang C, Yuan L, et al. MiR-369-3p participates in endometrioid adenocarcinoma via the regulation of autophagy. Cancer Cell Int. (2019) 19:178. doi: 10.1186/s12935-019-0897-8
12. Li F, Li K, Li D, Zhang W, Yang KW, Ke D, et al. ATG10 overexpression is related to the dismal prognosis and promotes the growth and migration of hepatocellular carcinoma cells via cyclin B1/CDK1 and CDK2. Am J Cancer Res. (2023) 13:1188–208.
13. Tandel P, Ranjbaran R, Ebrahimi E, Rezvani A, Ramzi M, Tamaddon G. Decreased expression of autophagy-related genes in the complete remission phase of acute myeloid leukemia. Mol Genet Genomic Med. (2022) 10:e1872. doi: 10.1002/mgg3.1872
14. Peng J, Zheng H, Liu F, Wu Q, Liu S. The m6A methyltransferase METTL3 affects autophagy and progression of nasopharyngeal carcinoma by regulating the stability of lncRNA ZFAS1. Infect Agent Cancer. (2022) 17:1. doi: 10.1186/s13027-021-00411-1
15. Cao QH, Liu F, Yang ZL, Fu XH, Yang ZH, Liu Q, et al. Prognostic value of autophagy related proteins ULK1, Beclin 1, ATG3, ATG5, ATG7, ATG9, ATG10, ATG12, LC3B and p62/SQSTM1 in gastric cancer. Am J Transl Res. (2016) 8:3831–47.
16. Jo YK, Kim SC, Park IJ, Park SJ, Jin DH, Hong SW, et al. Increased expression of ATG10 in colorectal cancer is associated with lymphovascular invasion and lymph node metastasis. PLoS One. (2012) 7:e52705. doi: 10.1371/journal.pone.0052705
17. Sun W, Li J, Zhou L, Han J, Liu R, Zhang H, et al. The c-Myc/miR-27b-3p/ATG10 regulatory axis regulates chemoresistance in colorectal cancer. Theranostics. (2020) 10:1981–96. doi: 10.7150/thno.37621
18. Wang K, Zhan H, Fan S, Chu S, Xu H, Jiang H. ATG10-dependent autophagy is required for DDX10 to regulate cell proliferation, apoptosis and stemness in colorectal cancer. J Cancer Res Clin Oncol. (2024) 150:386. doi: 10.1007/s00432-024-05910-3
19. Jo YK, Roh SA, Lee H, Park NY, Choi ES, Oh JH, et al. Polypyrimidine tract-binding protein 1-mediated down-regulation of ATG10 facilitates metastasis of colorectal cancer cells. Cancer Lett. (2017) 385:21–7. doi: 10.1016/j.canlet.2016.11.002
20. Wang W, Jiang XG, Bai YP, Li H, Gao LX, Zhang T, et al. SOV sensitizes gastric cancer cells to radiation by suppressing radiation-induced autophagy in vitro and in vivo. Tissue Cell. (2023) 82:102109. doi: 10.1016/j.tice.2023.102109
21. An J, Liu Y, Duo S, Ma X, An L, Yan Y, et al. Podofilox suppresses gastric cancer cell proliferation by regulating cell cycle arrest and the c-Myc/ATG10 axis. Exp Ther Med. (2021) 22:1203. doi: 10.3892/etm.2021.10637
22. Sui S, Ma F, Mi L, Gao L, Yu W, Li M, et al. Circ-CCS enhances autophagy during imatinib resistance of gastrointestinal stromal tumor by regulating miR-197-3p/ATG10 signaling. J Cancer Res Ther. (2022) 18:1338–45. doi: 10.4103/jcrt.jcrt_625_22
23. You B, Zhang P, Gu M, Yin H, Fan Y, Yao H, et al. Let-7i-5p promotes a Malignant phenotype in nasopharyngeal carcinoma via inhibiting tumor-suppressive autophagy. Cancer Lett. (2022) 531:14–26. doi: 10.1016/j.canlet.2022.01.019
24. Xie K, Liang C, Li Q, Yan C, Wang C, Gu Y, et al. Role of ATG10 expression quantitative trait loci in non-small cell lung cancer survival. Int J Cancer. (2016) 139:1564–73. doi: 10.1002/ijc.30205
25. Wang L, Wang H, Chen J. Linc00261 Inhibited High-Grade Serous Ovarian Cancer Progression through miR-552-ATG10-EMT Axis. Comput Math Methods Med. (2022) 2022:9450353. doi: 10.1155/2022/9450353
26. Garcia-Peterson LM, Ndiaye MA, Singh CK, Chhabra G, Huang W, Ahmad N. SIRT6 histone deacetylase functions as a potential oncogene in human melanoma. Genes Cancer. (2017) 8:701–12. doi: 10.18632/genesandcancer.153
27. Cho YY, Kim DJ, Lee HS, Jeong CH, Cho EJ, Kim MO, et al. Autophagy and cellular senescence mediated by Sox2 suppress Malignancy of cancer cells. PLoS One. (2013) 8:e57172. doi: 10.1371/journal.pone.0057172
Keywords: ATG10, epithelial-mesenchymal transition, cancer, review, therapeutic target
Citation: Shi K, Ke D, Li F, Shi R-S, Liu T, Li D and Zhang Q-X (2025) A review shows that ATG10 has been identified as a potential prognostic marker and therapeutic target for cancer patients based on real-world studies. Front. Oncol. 15:1573378. doi: 10.3389/fonc.2025.1573378
Received: 08 February 2025; Accepted: 27 March 2025;
Published: 17 April 2025.
Edited by:
Ji Zhang, University of South China, ChinaReviewed by:
Yang Zhang, First Affiliated Hospital of Heilongjiang University of Chinese Medicine, ChinaCopyright © 2025 Shi, Ke, Li, Shi, Liu, Li and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dan Li, bHE5NDk2NjEyNTlAMTYzLmNvbQ==; Tao Liu, MzA4ODQyMDEwQHFxLmNvbQ==; Qun-Xian Zhang, cXVueGlhbjA0QDEyNi5jb20=
†These authors share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.