 Chenwen Wang1†
Chenwen Wang1† Zixiong Chen
Zixiong Chen
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Oncol. , 04 March 2025
Sec. Cancer Immunity and Immunotherapy
Volume 15 - 2025 | https://doi.org/10.3389/fonc.2025.1555270
This article is part of the Research Topic Cancer Metastases: Mechanisms of Tumor Dissemination, Formation of Metastatic Niche and Anti-metastatic Therapy View all 10 articles
Background: MiR-136 is abnormally expressed in many types of metastatic tumors and is closely associated with tumor cell proliferation, apoptosis, invasion, and metastasis, indicating its important role in tumor development and progression. This review summarizes current knowledge regarding miR-136’s molecular mechanisms, functional roles, and impact on chemotherapy in different human cancers.
Methods: A literature search was conducted in PubMed and Web of Science using “miR-136” and “metastatic tumors” as English keywords, and in CNKI and Wanfang databases using the same terms in Chinese. Studies related to miR-136 research in metastatic tumors and high-quality evidence from similar studies were included. Meta-analyses, dissertations, conference papers, low-quality articles, unavailable full-text articles, and republished articles were excluded.
Results: This review synthesizes the current understanding of miR-136’s role in various cancers, including osteosarcoma, gastric cancer, gallbladder cancer, esophageal cancer, prostate cancer, colorectal cancer, breast cancer, glioma, and thyroid cancer. miR-136 acts as a tumor suppressor by targeting various genes, including MTDH, PTEN, MAP2K4, MUC1, LRH-1, MIEN1, RASAL2, CYR61, and KLF7. It influences multiple signaling pathways, including the ERK/mitogen-activated protein kinase, Wnt/β-catenin, Ha-Ras, PI3K/Akt, Aurora-A kinase, nuclear factor-κB, and JNK pathways. Furthermore, miR-136 is involved in chemoresistance by modulating ROCK1, PPP2R2A, and the miR-136-Notch3 signaling axis.
Conclusions: MiR-136 demonstrates promising potential as a novel biomarker and therapeutic target in various human cancers. Further research is needed to fully elucidate its complex roles in cancer development, progression, and drug resistance, particularly regarding its potential in immunotherapy.
MicroRNAs (miRNAs) are a class of non-coding, single-stranded RNA molecules, approximately 22 nucleotides long, encoded by endogenous genes. They participate in the transcriptional regulation of gene expression in animals and plants (1). By recognizing and binding to the 3′ untranslated region of target gene mRNA, miRNAs inhibit mRNA translation or promote degradation (2, 3). miRNAs can also affect histone modifications and promoter site methylation to regulate target gene expression (4, 5). A single miRNA can regulate the expression of multiple target genes (6, 7), by modulating various mRNAs, miRNAs functionally participate in a range of physiological and pathological processes, including cell differentiation, proliferation, migration, and apoptosis (8, 9). Abnormal miRNA expression has been reported in various types of metastatic tumors (10, 11), suggesting their potential oncogenic or tumor-suppressive functions. MiR-136 is abnormally expressed in many types of metastatic tumors and is closely associated with tumor cell proliferation, apoptosis, invasion, and metastasis, indicating its significant role in tumor development and progression (12–21). Moreover, miR-136 contributes to cancer resistance to various chemotherapeutic agents (21–24). Given its critical involvement in tumor biology, miR-136 represents a promising biomarker and therapeutic target for early cancer detection and treatment. This review summarizes the signaling pathways and mechanisms of action of miR-136 in various metastatic tumors, providing a comprehensive understanding of its role in metastatic tumors.
This review was indexed in PubMed and Web of Science using “miR-136” and “metastatic tumors” as English keywords. “miR-136” and “metastatic tumors” were used as Chinese keywords in CNKI and Wanfang databases. Studies were included based on the following criteria: (1) relevance to miR-136 research in metastatic tumors; and (2) provision of high-quality evidence. The following exclusion criteria were applied: (1) studies not published in English or Chinese; (2) meta-analysis, dissertations, conference papers, and (3) low-quality, unavailable full-text and republished articles. Studies were selected according to these criteria (Figure 1). miR-136 and the target genes in tumors are listed in Table 1.
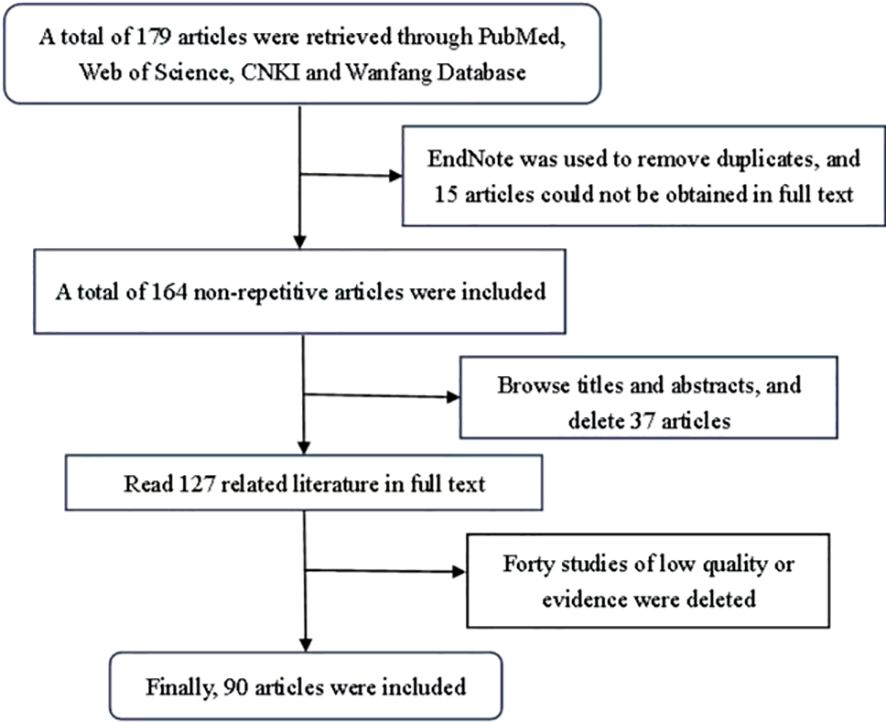
Figure 1. Literature search strategy.
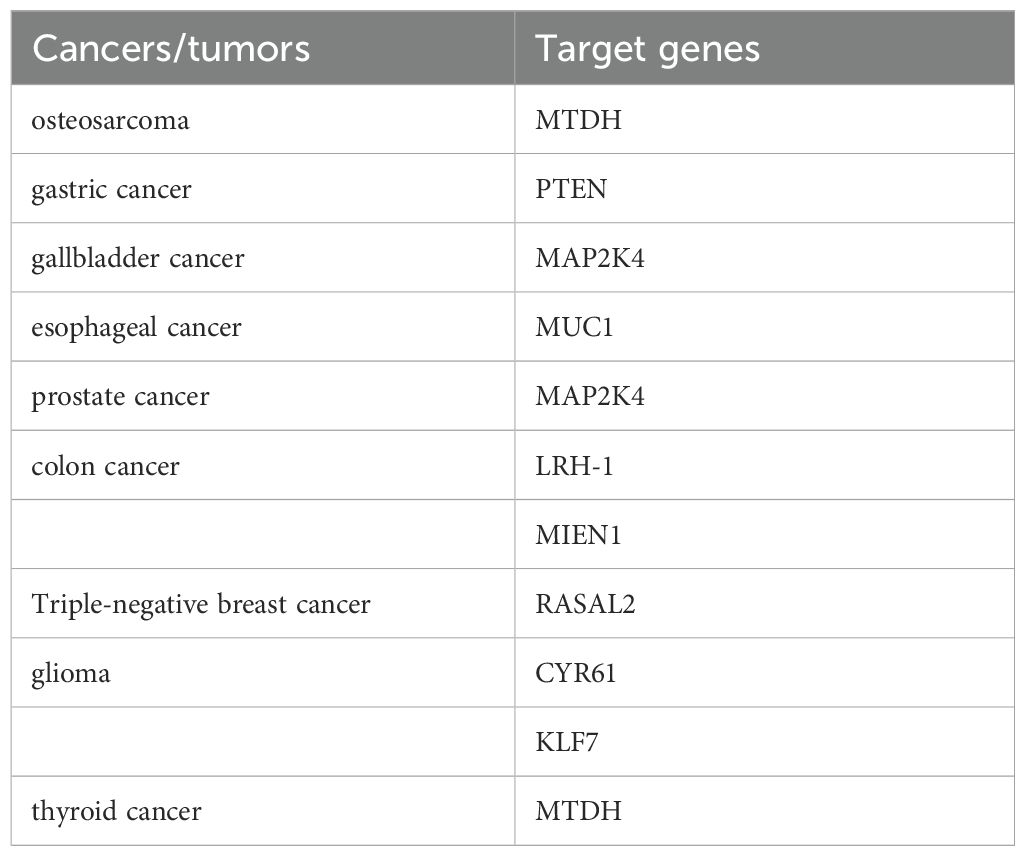
Table 1. miR-136 and the target genes in tumors.
Metastatic osteosarcoma (OS) is the third most common primary bone malignancy in children and adolescents (25), and commonly occurs in the proximal tibia, humerus, and metaphyseal region of the distal femur (26). The MTDH gene, located on human chromosome 8, facilitates tumor cells adhesion to distant blood vessels, playing a critical role in cancer spread and metastasis. MTDH is a involved in several oncogenic signaling pathways, such as ERK/mitogen-activated protein kinase, Wnt/β-catenin pathway, Ha-Ras and PI3K/Akt pathways, Aurora-A kinase signaling pathway, and nuclear factor-κB signaling pathway (27, 28). miR-136 inhibits the proliferation, invasion, and migration of osteosarcoma cells by negatively regulating MTDH.
Metastatic gastric cancer (GC) has the highest mortality rate among patients in China (29). Various pathogenic factors contribute to GC development and progression, including environmental factors, diet, infection, and genetic mutations, particularly the abnormal expression of proto-oncogenes or tumor suppressor genes (30). Compared with adjacent non-neoplastic gastric mucosal tissues and normal gastric epithelial cells, miR-136 expression was significantly increased while PTEN mRNA expression was decreased (19). Bioinformatics analysis has identified PTEN as a target of miR-136. Following miR-136 knockdown, PTEN mRNA and protein expression levels increase, whereas p-AKT protein levels decrease. PTEN expression was negatively correlated with miR-136 expression. PTEN negatively regulates various biological processes through the PI3K/AKT signaling pathway, including cell proliferation, migration, invasion, and apoptosis (31–34). Therefore, inhibiting miR-136 can suppress the proliferation, invasion, and metastasis of gastric cancer cells by modulating the PTEN/AKT/p-AKT signaling pathway.
Metastatic gallbladder cancer (GBC) is the most common malignancy of the biliary tract and the fifth most common tumor of the digestive tract (35). Compared with normal gallbladder epithelial cells, miR-136 is expressed at lower levels in gallbladder cancer cells (36). Mitogen-activated protein kinase 4 (MAP2K4) has been identified as a target gene of miR-136, which activates the JNK signaling pathway, a key mediator in tumorigenesis and apoptosis (36–38). In gallbladder cancer, miR-136 overexpression inhibits the MAP2K4-mediated JNK signaling pathway, thereby regulating the expression of downstream genes (39). Inhibition of the JNK signaling pathway decreases vascular endothelial growth factor (VEGF) expression, negatively regulating tumor growth and metastasis, as VEGF activates the angiogenesis signaling cascade and promotes tumor vascular endothelial cell proliferation, differentiation, and migration (40). Simultaneous inhibition of the JNK signaling pathway increases the ratio of c-caspase-3/t-caspase-3 and c-caspase-9/t-caspase-9, thereby promoting apoptosis (41, 42). Therefore, miR-136 overexpression inhibits angiogenesis and cell proliferation in gallbladder cancer while promoting apoptosis, suggesting a potential therapeutic role in gallbladder cancer treatment.
Metastatic esophageal cancer (EC) is the eighth most common cancer and the sixth leading cause of cancer-related deaths worldwide (43), EC comprises two main subtypes: esophageal adenocarcinoma (EAC), which has an increasing incidence, and esophageal squamous cell carcinoma (ESCC), prevalent in East Africa, Central Asia, and China (44). miR-136 expression is reduced in ESCC tissues, while MUC1 mRNA and protein expression levels are elevated compared to those in adjacent normal tissues. Bioinformatic analysis and luciferase activity assays confirm MUC1 as a miR-136 target, with an inverse correlation between their expression levels (45). miRNAs bind to the 3′ untranslated region (3′-UTR) of their target mRNAs, reducing their stability and post-transcriptional expression, thereby influencing biological processes such as cell growth, proliferation, differentiation, and death (46). Additionally, miR-136 upregulation reduces survival, inhibits colony formation, and induces apoptosis in ESCC cells under irradiation, whereas MUC1 upregulation reverses these effects. miRNAs can also influence cellular responses to precision drugs by interfering with DNA repair and drug targets (47).
Metastatic prostate cancer (PCa) is a common malignancy and the second most prevalent tumor of the urinary and reproductive systems (48). The occurrence and progression of PCa are regulated by miRNAs, and PCa-related miRNAs research provides novel biomarkers for diagnosis and treatment (49, 50). miR-136 expression is reduced in PCa tissues and cell lines, whereas its upregulation inhibits PCa cells. A luciferase reporter assay has confirmed that mitogen-activated protein kinase 4 (MAP2K4) is a miR-136 target gene. MAP2K4 is upregulated in PCa tissues, and its expression levels are inversely correlated with miR-136 levels. MAP2K4, located on chromosome 17, is involved in various tumorigenic and pathophysiological processes, including cell proliferation, invasion, metastasis, and apoptosis (51–53). In PCa, MAP2K4 overexpression promotes cell proliferation and metastasis while inhibiting G1-S phase arrest and apoptosis (39, 54).Thus, miR-136 may suppress PCa proliferation and invasion by targeting MAP2K4, making it a potential candidate for PCa therapy.
Metastatic colon cancer (CC) is one of the most prevalent malignancies and the fourth leading cause of cancer-related deaths worldwide (55). Hepatic receptor homolog-1 (LRH-1), a member of the nuclear receptor subfamily, is a recognized oncogene in many cancers (56), promoting the proliferation, invasion, and migration of cancer cells (57). LRH-1 plays a crucial role in various biological processes, including bile acid homeostasis, reverse cholesterol transport, steroid production, differentiation, and development (58). LRH-1 knockdown has been shown to inhibit colon cancer cell proliferation and induce G0/G1 cell cycle arrest (59). Additionally, LRH-1 promotes colon cancer cell growth by inhibiting the recruitment of p53 to the promoter of the cell cycle inhibitor p21 (60). Wnt signaling is aberrantly activated in approximately 80% of colon cancers (61), and its downstream genes, including cyclin D1, cyclin E1, and c-Myc, are implicated in the proliferation and metastasis of colon cancer cells (62). LRH-1 is a novel co-activator of Wnt signaling pathway transduction (59, 63), and can interact with transcription factor 4 and β-catenin to promote the expression of cyclin D1, cyclin E1, and c-Myc (63). miR-136 offers a novel pathway for the inhibition of Wnt signaling by significantly reducing the expression of cyclin D1, cyclin E1, and c-Myc in colon cancer through LRH-1 suppression (20).
MIEN1, located in the 17q12 region of the chromosome near the Her-2/neu locus (64), is frequently dysregulated in various cancers (65, 66). MIEN1 expression is elevated in colorectal cancer tissues and is closely associated with tumor serous invasion, lymph node metastasis, and advanced Dukes stage (67). miR-136 has been shown to inhibit colon cancer cell invasion, migration, and EMT progression by regulating the Akt/NF-κB signaling pathway through its target gene, MIEN1 (66).
Metastatic breast cancer (BC) is the most common cancer in women and a leading cause of cancer-related deaths (68, 69). Triple-negative breast cancer (TNBC) is a heterogeneous group of breast cancers characterized by the loss of estrogen receptor (ER), progesterone receptor (PR), and human epidermal growth factor receptor 2 (HER2) gene expression (70). The Ras pathway is one of the most commonly dysregulated pathways in cancer, with Ras protein mutations occurring at high frequency (71). Ras activity is negatively regulated by Ras GTPase-a activating proteins (RasGAPs), which catalyze the hydrolysis of Ras-GTP to Ras-GDP (72). Interestingly, RASAL2, a GAP, has been identified as an oncogene promoting tumor production and metastasis in various cancers (73–75), However, rather than suppressing tumors, RASAL2 facilitates mesenchymal invasion and metastasis (73, 76, 77). In TNBC, miR-136 has been shown to act as a tumor suppressor by directly targeting RASAL2. Through downregulation of RASAL2, miR-136 effectively inhibits tumor growth and metastasis, underscoring its therapeutic potential in TNBC.
Metastatic glioma is a common malignancy (78, 79). Bioinformatic studies suggest that miR-136 can function as either a tumor suppressor or an oncomiR, depending on the context. Overexpression of miR-136 has been shown to inhibit glioblastoma cell proliferation by targeting CYR61. Signal transduction via the mTOR pathway is activated alongside miR-136 expression and is dependent on the activities of AKT, ERK1/2, and mTORC1. miR-136 expression is reduced in glioma tissues compared to adjacent normal tissues. KLF7, a target gene of miR-136 (18), promotes polyamine biosynthesis and glioma progression by activating arginine succinate lyase (80). Overexpression of miR-136 has been shown to inhibit glioma cell growth and migration.
Metastatic thyroid cancer (TC) is a common endocrine neoplasm, accounting for approximately 3.1% of all human malignancies (81). Papillary thyroid cancer, the most prevalent pathological subtype of TC, constitutes approximately 80% of cases. Its incidence is higher than that of other subtypes, such as anaplastic, follicular, and medullary thyroid cancer. The incidence of TC has increased in many countries. Although the overall mortality rate of TC is relatively low, with a 5-year survival rate of 98%, the clinical outcomes of advanced TC remain poor. Nearly half of patients with distant TC metastases die within 5 years of diagnosis. Recurrence and lung metastasis remain the leading causes of mortality in TC patients. Studies have reported higher levels of miR-136 in papillary adenocarcinomas compared to benign nodular goiter (82). MTDH has been identified as a target gene in various cancers (13, 17), with its overexpression playing a key role in cancer development. A luciferase reporter assay has confirmed the targeted regulatory relationship between miR-136-5p and MTDH (15).
Chemotherapy is one of the most common clinical treatments for tumors; however, chemoresistance remains a widespread challenge (83, 84). Cisplatin, one of the most effective chemotherapeutic agents (85), inevitably encounters drug resistance, limiting the efficacy of other agents and leading to potential treatment failure. Abnormal miRNA expression can disrupt the regulation of chemotherapy drug target proteins, ultimately contributing to drug resistance. miR-136, one of the most extensively studied miRNAs, is abnormally expressed in various tumors. miR-136 overexpression reduces ROCK1 expression in cisplatin-treated tumor cells and attenuates the Akt/mTOR signaling pathway, leading to chemoresistance (86). Additionally, miR-136 overexpression promotes anlotinib resistance in non-small cell lung cancer by targeting PPP2R2A, thereby activating the Akt pathway. miR-136 can be transferred from anlotinib-resistant cells to anlotinib-sensitive cells via exosomes, inducing drug resistance and promoting cell proliferation (22). In gliocytomas, miR-136 overexpression enhances temozolomide cytotoxicity (24). In ovarian cancer, the miR-136-Notch3 signaling axis plays a crucial role in the development of chemoresistance (87). These findings suggest that miRNA-mediated chemoresistance and chemo sensitization can be modulated to enhance chemotherapy efficacy, offering new strategies for overcoming tumor drug resistance.
Immunotherapy is one of the most effective cancer treatment strategies. Among immune checkpoint molecules, miRNA-based PD-L1 regulation is the most widely studied. PD-L1, expressed on immune and cancer cell surfaces, inhibits T-cell proliferation by binding to its receptor, PD-1. In NSCLC, miR-34a directly binds to the 3’-UTR of PD-L1, inhibiting its expression. miR-140 functions as a PD-L1 modulator in osteosarcoma (88), whereas miR-15a and miR-15b exert antitumor effects by blocking PD-L1 in neuroblastoma (89). In breast cancer cells, PD-L1 activates PDCD4 via the PI3K/Akt pathway, a process significantly enhanced by miR-21 (90). However, the role of miR-136 in tumor immunotherapy remains unclear and requires further investigation.
miR-136 stands out for its involvement in epigenetic regulation (e.g., targeting EZH2) and its ability to enhance chemosensitivity in cancer cells. Like miR-34a and let-7, miR-136 acts as a tumor suppressor by targeting anti-apoptotic proteins and inhibiting proliferation. Similar to miR-21 and miR-155, miR-136 regulates the PI3K/AKT pathway, but it does so in a tumor-suppressive manner, unlike the pro-tumorigenic effects of miR-21 and miR-155. In conclusion, miR-136 shares some functional roles with other miRNAs in cancer, such as regulating apoptosis and proliferation, but its unique involvement in epigenetic regulation and chemosensitivity distinguishes it from other miRNAs. Its downregulation in cancer highlights its potential as a therapeutic target or biomarker.
MiRNAs regulate the expression of their corresponding target genes and exhibit direct or indirect carcinogenic or tumor-suppressive effects in various cancers. Many studies have identified miR-136 as a tumor suppressor gene. miR-136 is an anti-invasive miRNA that inhibits mesenchymal invasion and transfer in TNBC. The miR-136/RASAL2/MET axis functions as a repressor of TNBC metastasis. miR-136 also negatively regulates colon cancer progression by targeting LRH-1, preventing aberrant activation of Wnt signaling. In gliomas, miR-136 inhibits proliferation and induces apoptosis by regulating AEG-1 and BCL-2 gene expression. Inhibition of miR-136 expression upregulates its target AEG-1 gene and significantly improves the metastatic ability of hepatoma cells. Additionally, miR-136 suppresses lung cancer cell proliferation, invasion, and migration. In osteosarcoma, it negatively regulates MTDH, exerting tumor-suppressive effects. miR-136 also inhibits the MAP2K4-mediated JNK signaling pathway, thereby influencing downstream gene expression. These findings suggest that miR-136 has broad tumor-suppressive roles, making it a potential therapeutic target. Furthermore, miRNAs regulate multiple target genes, underscoring their functional diversity and importance in cancer treatment.
miR-136 plays a role in various signaling pathways in cancer and has been identified as a promising biomarker for cancer diagnosis and prognosis. Additionally, several cell-based and preclinical studies have shown that blocking or inhibiting miR-136 can lead to the regression of various cancer types, making it a strong candidate for cancer drug discovery. miR-136 is also actively involved in regulating drug resistance, and any effective miR-136 targeting strategy could help reduce cancer cell resistance and recurrence. Several small-molecule inhibitors of miR-136 have been reported; however, its functions in both cancerous and normal cells are not fully understood. Using advanced sequencing techniques and powerful bioinformatics tools to explore the regulatory function of miR-136 in complex oncogenic pathways will enhance our understanding and reveal new potential applications for miR-136 in cancer diagnosis and treatment. Beyond investigating miR-136 itself, exploring effective drug delivery methods is also essential. The emerging research focus on exosomes offers new avenues for drug delivery. Exosomes are messengers in cell-to-cell communication in the tumor microenvironment, and exosomal circRNAs have been reported to function as miRNA sponges, which are important in tumors. However, the role of miR-136 in human cancer requires further investigation. This review summarizes miR-136’s feasibility as a diagnostic or prognostic biomarker and provides new perspectives on cancer resistance and drug susceptibility research.
CW: Data curation, Writing – original draft. ZC: Methodology, Writing – original draft. WN: Visualization, Writing – original draft. JW: Supervision, Writing – review & editing. WZ: Supervision, Writing – review & editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This study was supported by the Natural Science Foundation of Hubei Province of China (No. 2021CFB521) and Huazhong University of Science and Technology Independent Innovation Research Fund (2018KFYYXJJ111). The funding bodies played no role in the design of the study and collection, analysis, and interpretation of data and in writing the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Lu TX, Rothenberg ME. MicroRNA. J Allergy Clin Immunol. (2018) 141(4):1202–7. doi: 10.1016/j.jaci.2017.08.034
2. Pillai RS, Bhattacharyya SN, Filipowicz W. Repression of protein synthesis by miRNAs: how many mechanisms? Trends Cell Biol. (2007) 17(3):118–26. doi: 10.1016/j.tcb.2006.12.007
3. Ambardekar VV, Wakaskar RR, Ye Z, Curran SM, McGuire TR, Coulter DW, et al. Complexation of chol-DsiRNA in place of chol-siRNA greatly increases the duration of mRNA suppression by polyplexes of PLL(30)-PEG(5K) in primary murine syngeneic breast tumors after i.v. administration. Int J Pharm. (2018) 543(1-2):130–8. doi: 10.1016/j.ijpharm.2018.03.045
4. Tan Y, Zhang B, Wu T, Skogerbø G, Zhu X, Guo X, et al. Transcriptional inhibiton of Hoxd4 expression by miRNA-10a in human breast cancer cells. BMC Mol Biol. (2009) 10:12. doi: 10.1186/1471-2199-10-12
5. Hawkins PG, Morris KV. RNA and transcriptional modulation of gene expression. Cell Cycle (Georgetown Tex). (2008) 7(5):602–7. doi: 10.4161/cc.7.5.5522
6. Baek D, Villén J, Shin C, Camargo FD, Gygi SP, Bartel DP. The impact of microRNAs on protein output. Nature. (2008) 455(7209):64–71. doi: 10.1038/nature07242
7. Yang Y, Chaerkady R, Kandasamy K, Huang TC, Selvan LD, Dwivedi SB, et al. Identifying targets of miR-143 using a SILAC-based proteomic approach. Mol Biosyst. (2010) 6(10):1873–82. doi: 10.1039/c004401f
8. Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. (2004) 116(2):281–97. doi: 10.1016/s0092-8674(04)00045-5
9. Calin GA, Croce CM. MicroRNA signatures in human cancers. Nat Rev Cancer. (2006) 6(11):857–66. doi: 10.1038/nrc1997
10. Hayes J, Peruzzi PP, Lawler S. MicroRNAs in cancer: biomarkers, functions and therapy. Trends Mol Med. (2014) 20(8):460–9. doi: 10.1016/j.molmed.2014.06.005
11. Calin GA, Dumitru CD, Shimizu M, Bichi R, Zupo S, Noch E, et al. Frequent deletions and down-regulation of micro- RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia. Proc Natl Acad Sci USA. (2002) 99(24):15524–9. doi: 10.1073/pnas.242606799
12. Jia H, Wang H, Yao Y, Wang C, Li P. miR-136 inhibits malignant progression of hepatocellular carcinoma cells by targeting cyclooxygenase 2. Oncol Res. (2018) 26(6):967–76. doi: 10.3727/096504018x15148192843443
13. Yu L, Zhou GQ, Li DC. MiR-136 triggers apoptosis in human gastric cancer cells by targeting AEG-1 and BCL2. Eur Rev Med Pharmacol Sci. (2018) 22(21):7251–6. doi: 10.26355/eurrev_201811_16259
14. Chu Y, Hu X, Wang G, Wang Z, Wang Y. Downregulation of miR-136 promotes the progression of osteosarcoma and is associated with the prognosis of patients with osteosarcoma. Oncol Letters. (2019) 17(6):5210–8. doi: 10.3892/ol.2019.10203
15. Gao RZ, Que Q, Lin P, Pang YY, Wu HY, Li XJ, et al. Clinical roles of miR-136-5p and its target metadherin in thyroid carcinoma. Am J Trans Res. (2019) 11(11):6754–74.
16. Yan M, Li X, Tong D, Han C, Zhao R, He Y, et al. miR-136 suppresses tumor invasion and metastasis by targeting RASAL2 in triple-negative breast cancer. Oncol Rep. (2016) 36(1):65–71. doi: 10.3892/or.2016.4767
17. Guo T, Pan G. MicroRNA-136 functions as a tumor suppressor in osteosarcoma via regulating metadherin. Cancer Biomarkers. (2018) 22(1):79–87. doi: 10.3233/cbm-170970
18. Xu Y. MicroRNA-136-3p inhibits glioma tumorigenesis in vitro and in vivo by targeting KLF7. World J Surg Oncol. (2020) 18(1):169. doi: 10.1186/s12957-020-01949-x
19. Chen X, Huang Z, Chen R. Microrna-136 promotes proliferation and invasion ingastric cancer cells through Pten/Akt/P-akt signaling pathway. Oncol Letters. (2018) 15(4):4683–9. doi: 10.3892/ol.2018.7848
20. Yuan Q, Cao G, Li J, Zhang Y, Yang W. MicroRNA-136 inhibits colon cancer cell proliferation and invasion through targeting liver receptor homolog-1/Wnt signaling. Gene. (2017) 628:48–55. doi: 10.1016/j.gene.2017.07.031
21. Zhao H, Liu S, Wang G, Wu X, Ding Y, Guo G, et al. Expression of miR-136 is associated with the primary cisplatin resistance of human epithelial ovarian cancer. Oncol Rep. (2015) 33(2):591–8. doi: 10.3892/or.2014.3640
22. Gu G, Hu C, Hui K, Zhang H, Chen T, Zhang X, et al. Exosomal miR-136-5p derived from anlotinib-resistant NSCLC cells confers anlotinib resistance in non-small cell lung cancer through targeting PPP2R2A. Int J Nanomedicine. (2021) 16:6329–43. doi: 10.2147/ijn.S321720
23. Chen W, Yang Y, Chen B, Lu P, Zhan L, Yu Q, et al. MiR-136 targets E2F1 to reverse cisplatin chemosensitivity in glioma cells. J Neuro-Oncology. (2014) 120(1):43–53. doi: 10.1007/s11060-014-1535-x
24. Wu H, Liu Q, Cai T, Chen YD, Liao F, Wang ZF. MiR-136 modulates glioma cell sensitivity to temozolomide by targeting astrocyte elevated gene-1. Diagn Pathology. (2014) 9:173. doi: 10.1186/s13000-014-0173-0
25. Kansara M, Teng MW, Smyth MJ, Thomas DM. Translational biology of osteosarcoma. Nat Rev Cancer. (2014) 14(11):722–35. doi: 10.1038/nrc3838
26. Shimizu T, Fuchimoto Y, Fukuda K, Okita H, Kitagawa Y, Kuroda T. The effect of immune checkpoint inhibitors on lung metastases of osteosarcoma. J Pediatr Surgery. (2017) 52(12):2047–50. doi: 10.1016/j.jpedsurg.2017.08.030
27. Yoo BK, Emdad L, Lee SG, Su ZZ, Santhekadur P, Chen D, et al. Astrocyte elevated gene-1 (AEG-1): A multifunctional regulator of normal and abnormal physiology. Pharmacol Ther. (2011) 130(1):1–8. doi: 10.1016/j.pharmthera.2011.01.008
28. Sarkar D, Emdad L, Lee SG, Yoo BK, Su ZZ, Fisher PB. Astrocyte elevated gene-1: far more than just a gene regulated in astrocytes. Cancer Res. (2009) 69(22):8529–35. doi: 10.1158/0008-5472.Can-09-1846
29. Xia C, Dong X, Li H, Cao M, Sun D, He S, et al. Cancer statistics in china and united states, 2022: profiles, trends, and determinants. Chin Med J. (2022) 135(5):584–90. doi: 10.1097/cm9.0000000000002108
30. Ueda T, Volinia S, Okumura H, Shimizu M, Taccioli C, Rossi S, et al. Relation between microRNA expression and progression and prognosis of gastric cancer: a microRNA expression analysis. Lancet Oncol. (2010) 11(2):136–46. doi: 10.1016/s1470-2045(09)70343-2
31. Mukhopadhyay UK, Mooney P, Jia L, Eves R, Raptis L, Mak AS. Doubles game: Src-Stat3 versus p53-PTEN in cellular migration and invasion. Mol Cell Biol. (2010) 30(21):4980–95. doi: 10.1128/mcb.00004-10
32. Waite KA, Eng C. Protean PTEN: form and function. Am J Hum Genet. (2002) 70(4):829–44. doi: 10.1086/340026
33. Stambolic V, Suzuki A, de la Pompa JL, Brothers GM, Mirtsos C, Sasaki T, et al. Negative regulation of PKB/Akt-dependenT cell survival by the tumor suppressor PTEN. Cell. (1998) 95(1):29–39. doi: 10.1016/s0092-8674(00)81780-8
34. Lee H, Choi SK, Ro JY. Overexpression of DJ-1 and HSP90α, and loss of PTEN associated with invasive urothelial carcinoma of urinary bladder: Possible prognostic markers. Oncol Letters. (2012) 3(3):507–12. doi: 10.3892/ol.2011.522
35. Roa JC, García P, Kapoor VK, Maithel SK, Javle M, Koshiol J. Gallbladder cancer. Nat Rev Dis Primers. (2022) 8(1):69. doi: 10.1038/s41572-022-00398-y
36. Niu J, Li Z, Li F. Overexpressed microRNA-136 works as a cancer suppressor in gallbladder cancer through suppression of JNK signaling pathway via inhibition of MAP2K4. Am J Physiol Gastrointestinal Liver Physiol. (2019) 317(5):G670–g81. doi: 10.1152/ajpgi.00055.2019
37. Hong H, He C, Zhu S, Zhang Y, Wang X, She F, et al. CCR7 mediates the TNF-α-induced lymphatic metastasis of gallbladder cancer through the "ERK1/2 - AP-1" and "JNK - AP-1" pathways. J Exp Clin Cancer Res. (2016) 35:51. doi: 10.1186/s13046-016-0318-y
38. Matsui Y, Kuwabara T, Eguchi T, Nakajima K, Kondo M. Acetylation regulates the MKK4-JNK pathway in T cell receptor signaling. Immunol Letters. (2018) 194:21–8. doi: 10.1016/j.imlet.2017.12.002
39. Pavese JM, Ogden IM, Voll EA, Huang X, Xu L, Jovanovic B, et al. Mitogen-activated protein kinase kinase 4 (MAP2K4) promotes human prostate cancer metastasis. PloS One. (2014) 9(7):e102289. doi: 10.1371/Journal.pone.0102289
40. Na HJ, Hwang JY, Lee KS, Choi YK, Choe J, Kim JY, et al. TRAIL negatively regulates VEGF-induced angiogenesis via caspase-8-mediated enzymatic and non-enzymatic functions. Angiogenesis. (2014) 17(1):179–94. doi: 10.1007/s10456-013-9387-0
41. Chen Y, Liang J, Liang X, Chen J, Wang Y, Cao J, et al. Limonin induces apoptosis of HL-60 cells by inhibiting NQO1 activity. Food Sci Nutr. (2021) 9(4):1860–9. doi: 10.1002/fsn3.2109
42. Jia J, Yin H, Chen C, Hu M, Zhong Q, Zheng S, et al. Design, synthesis, and evaluation of a novel series of mono-indolylbenzoquinones derivatives for the potential treatment of breast cancer. Eur J Med Chem. (2022) 237:114375. doi: 10.1016/j.ejmech.2022.114375
43. Okuda M, Inoue J, Fujiwara N, Kawano T, Inazawa J. Subcloning and characterization of highly metastatic cells derived from human esophageal squamous cell carcinoma KYSE150 cells by in vivo selection. Oncotarget. (2017) 8(21):34670–7. doi: 10.18632/oncotarget.16668
44. Kim JA, Shah PM. Screening and prevention strategies and endoscopic management of early esophageal cancer. Chin Clin Oncol. (2017) 6(5):50. doi: 10.21037/cco.2017.09.05
45. Huang HZ, Yin YF, Wan WJ, Xia D, Wang R, Shen XM. Up-regulation of microRNA-136 induces apoptosis and radiosensitivity of esophageal squamous cell carcinoma cells by inhibiting the expression of MUC1. Exp Mol Pathology. (2019) 110:104278. doi: 10.1016/j.yexmp.2019.104278
46. Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell. (2009) 136(2):215–33. doi: 10.1016/j.Cell.2009.01.002
47. Giovannetti E, Erozenci A, Smit J, Danesi R, Peters GJ. Molecular mechanisms underlying the role of microRNAs (miRNAs) in anticancer drug resistance and implications for clinical practice. Crit Rev Oncology/Hematology. (2012) 81(2):103–22. doi: 10.1016/j.critrevonc.2011.03.010
48. Peyromaure EM, Mao K, Sun Y, Xia S, Jiang N, Zhang S, et al. A comparative study of prostate cancer detection and management in china and in france. Can J Urology. (2009) 16(1):4472–7.
49. Sun Y, Jia X, Hou L, Liu X. Screening of differently expressed miRNA and mRNA in prostate cancer by integrated analysis of transcription data. Urology. (2016) 94:313. doi: 10.1016/j.urology.2016.04.041
50. Wang Z, Xu L, Hu Y, Huang Y, Zhang Y, Zheng X, et al. miRNA let-7b modulates macrophage polarization and enhances tumor-associated macrophages to promote angiogenesis and mobility in prostate cancer. Sci Rep. (2016) 6:25602. doi: 10.1038/srep25602
51. Tesser-Gamba F, Petrilli AS, de Seixas Alves MT, Filho RJ, Juliano Y, Toledo SR. MAPK7 and MAP2K4 as prognostic markers in osteosarcoma. Hum Pathology. (2012) 43(7):994–1002. doi: 10.1016/j.humpath.2011.08.003
52. Ishikawa M, Nakayama K, Rahman MT, Rahman M, Katagiri A, Iida K, et al. Functional and clinicopathological analysis of loss of MKK4 expression in endometrial cancer. Oncology. (2010) 79(3-4):238–46. doi: 10.1159/000322644
53. Wang L, Pan Y, Dai JL. Evidence of MKK4 pro-oncogenic activity in breast and pancreatic tumors. Oncogene. (2004) 23(35):5978–85. doi: 10.1038/sj.onc.1207802
54. Wan X, Huang W, Yang S, Zhang Y, Zhang P, Kong Z, et al. Androgen-induced miR-27A acted as a tumor suppressor by targeting MAP2K4 and mediated prostate cancer progression. Int J Biochem Cell Biol. (2016) 79:249–60. doi: 10.1016/j.biocel.2016.08.043
55. Patel SG, Karlitz JJ, Yen T, Lieu CH, Boland CR. The rising tide of early-onset colorectal cancer: a comprehensive review of epidemiology, clinical features, biology, risk factors, prevention, and early detection. Lancet Gastroenterol Hepatology. (2022) 7(3):262–74. doi: 10.1016/s2468-1253(21)00426-x
56. Nadolny C, Dong X. Liver receptor homolog-1 (LRH-1): a potential therapeutic target for cancer. Cancer Biol Ther. (2015) 16(7):997–1004. doi: 10.1080/15384047.2015.1045693
57. Chand AL, Herridge KA, Thompson EW, Clyne CD. The orphan nuclear receptor LRH-1 promotes breast cancer motility and invasion. Endocrine-Related Cancer. (2010) 17(4):965–75. doi: 10.1677/erc-10-0179
58. Fayard E, Auwerx J, Schoonjans K. LRH-1: an orphan nuclear receptor involved in development, metabolism and steroidogenesis. Trends Cell Biol. (2004) 14(5):250–60. doi: 10.1016/j.tcb.2004.03.008
59. Bayrer JR, Mukkamala S, Sablin EP, Webb P, Fletterick RJ. Silencing LRH-1 in colon cancer cell lines impairs proliferation and alters gene expression programs. Proc Natl Acad Sci USA. (2015) 112(8):2467–72. doi: 10.1073/pnas.1500978112
60. Kramer HB, Lai CF, Patel H, Periyasamy M, Lin ML, Feller SM, et al. LRH-1 drives colon cancer cell growth by repressing the expression of the CDKN1A gene in a p53-dependent manner. Nucleic Acids Res. (2016) 44(2):582–94. doi: 10.1093/nar/gkv948
61. Kolligs FT, Bommer G, Göke B. Wnt/beta-catenin/tcf signaling: a critical pathway in gastrointestinal tumorigenesis. Digestion. (2002) 66(3):131–44. doi: 10.1159/000066755
62. Ischenko I, Zhi J, Moll UM, Nemajerova A, Petrenko O. Direct reprogramming by oncogenic ras and myc. Proc Natl Acad Sci U S A. (2013) 110(10):3937–42. doi: 10.1073/pnas.1219592110
63. Botrugno OA, Fayard E, Annicotte JS, Haby C, Brennan T, Wendling O, et al. Synergy between LRH-1 and beta-catenin induces G1 cyclin-mediated cell proliferation. Mol Cell. (2004) 15(4):499–509. doi: 10.1016/j.molcel.2004.07.009
64. Dasgupta S, Wasson LM, Rauniyar N, Prokai L, Borejdo J, Vishwanatha JK. Novel gene C17orf37 in 17q12 amplicon promotes migration and invasion of prostate cancer cells. Oncogene. (2009) 28(32):2860–72. doi: 10.1038/onc.2009.145
65. Evans EE, Henn AD, Jonason A, Paris MJ, Schiffhauer LM, Borrello MA, et al. C35 (C17orf37) is a novel tumor biomarker abundantly expressed in breast cancer. Mol Cancer Ther. (2006) 5(11):2919–30. doi: 10.1158/1535-7163.Mct-06-0389
66. Hsu CH, Shen TL, Chang CF, Chang YY, Huang LY. Solution structure of the oncogenic MIEN1 protein reveals a thioredoxin-like fold with a redox-active motif. PloS One. (2012) 7(12):e52292. doi: 10.1371/Journal.pone.0052292
67. Dong X, Huang Y, Kong L, Li J, Kou J, Yin L, et al. C35 is overexpressed in colorectal cancer and is associated tumor invasion and metastasis. Bioscience Trends. (2015) 9(2):117–21. doi: 10.5582/bst.2015.01057
68. Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. (2021) 71(3):209–49. doi: 10.3322/caac.21660
69. Autier P, Boniol M, La Vecchia C, Vatten L, Gavin A, Héry C, et al. Disparities in breast cancer mortality trends between 30 european countries: retrospective trend analysis of WHO mortality database. BMJ (Clinical Res ed). (2010) 341:c3620. doi: 10.1136/bmj.c3620
70. Yin L, Duan JJ, Bian XW, Yu SC. Triple-negative breast cancer molecular subtyping and treatment progress. Breast Cancer Res. (2020) 22(1):61. doi: 10.1186/s13058-020-01296-5
71. Downward J. Targeting RAS signalling pathways in cancer therapy. Nat Rev Cancer. (2003) 3(1):11–22. doi: 10.1038/nrc969
72. Bos JL, Rehmann H, Wittinghofer A. GEFs and GAPs: critical elements in the control of small g proteins. Cell. (2007) 129(5):865–77. doi: 10.1016/j.Cell.2007.05.018
73. McLaughlin SK, Olsen SN, Dake B, De Raedt T, Lim E, Bronson RT, et al. The RasGAP gene, RASAL2, is a tumor and metastasis suppressor. Cancer Cell. (2013) 24(3):365–78. doi: 10.1016/j.ccr.2013.08.004
74. Xu Y, Deng Y, Ji Z, Liu H, Liu Y, Peng H, et al. Identification of thyroid carcinoma related genes with mRMR and shortest path approaches. PloS One. (2014) 9(4):e94022. doi: 10.1371/Journal.pone.0094022
75. Li N, Li S. RASAL2 promotes lung cancer metastasis through epithelial-mesenchymal transition. Biochem Biophys Res Commun. (2014) 455(3-4):358–62. doi: 10.1016/j.bbrc.2014.11.020
76. Feng M, Bao Y, Li Z, Li J, Gong M, Lam S, et al. RASAL2 activates RAC1 to promote triple-negative breast cancer progression. J Clin Invest. (2014) 124(12):5291–304. doi: 10.1172/jci76711
77. Marmolejo DH, Wong MYZ, Bajalica-Lagercrantz S, Tischkowitz M, Balmaña J. Overview of hereditary breast and ovarian cancer (HBOC) guidelines across europe. Eur J Med Genet. (2021) 64(12):104350. doi: 10.1016/j.ejmg.2021.104350
78. Visser O, Ardanaz E, Botta L, Sant M, Tavilla A, Minicozzi P. Survival of adults with primary malignant brain tumours in europe; results of the EUROCARE-5 study. Eur J Cancer (Oxford Engl 1990). (2015) 51(15):2231–41. doi: 10.1016/j.ejca.2015.07.032
79. Lapointe S, Perry A, Butowski NA. Primary brain tumours in adults. Lancet. (2018) 392(10145):432–46. doi: 10.1016/s0140-6736(18)30990-5
80. Guan F, Kang Z, Zhang JT, Xue NN, Yin H, Wang L, et al. KLF7 promotes polyamine biosynthesis and glioma development through transcriptionally activating ASL. Biochem Biophys Res Commun. (2019) 514(1):51–7. doi: 10.1016/j.bbrc.2019.04.120
81. Rusinek D, Chmielik E, Krajewska J, Jarzab M, Oczko-Wojciechowska M, Czarniecka A, et al. Current advances in thyroid cancer management. are we ready for the epidemic rise of diagnoses? Int J Mol Sci. (2017) 18(8):1817. doi: 10.3390/ijms18081817
82. Peng Y, Li C, Luo DC, Ding JW, Zhang W, Pan G. Expression profile and clinical significance of microRNAs in papillary thyroid carcinoma. Molecules (Basel Switzerland). (2014) 19(8):11586–99. doi: 10.3390/molecules190811586
83. Landrum LM, Java J, Mathews CA, Lanneau GS Jr., Copeland LJ, Armstrong DK, et al. Prognostic factors for stage III epithelial ovarian cancer treated with intraperitoneal chemotherapy: a gynecol oncol group study. Gynecol Oncol. (2013) 130(1):12–8. doi: 10.1016/j.ygyno.2013.04.001
84. Berek JS, Renz M, Kehoe S, Kumar L, Friedlander M. Cancer of the ovary, fallopian tube, and peritoneum: 2021 update. Int J Gynaecol Obstet. (2021) 155 Suppl 1(Suppl 1):61–85. doi: 10.1002/ijgo.13878
85. Ghosh S. Cisplatin: The first metal based anticancer drug. Bioorg Chem. (2019) 88:102925. doi: 10.1016/j.bioorg.2019.102925
86. Yang B, Zang J, Yuan W, Jiang X, Zhang F. The miR-136-5p/ROCK1 axis suppresses invasion and migration, and enhances cisplatin sensitivity in head and neck cancer cells. Exp Ther Med. (2021) 21(4):317. doi: 10.3892/etm.2021.9748
87. Jeong JY, Kang H, Kim TH, Kim G, Heo JH, Kwon AY, et al. MicroRNA-136 inhibits cancer stem cell activity and enhances the anti-tumor effect of paclitaxel against chemoresistant ovarian cancer cells by targeting Notch3. Cancer Lett. (2017) 386:168–78. doi: 10.1016/j.canlet.2016.11.017
88. Ji X, Wang E, Tian F. MicroRNA-140 suppresses osteosarcoma tumor growth by enhancing anti-tumor immune response and blocking mTOR signaling. Biochem Biophys Res Commun. (2018) 495(1):1342–8. doi: 10.1016/j.bbrc.2017.11.120
89. Pathania AS, Prathipati P, Olwenyi OA, Chava S, Smith OV, Gupta SC, et al. miR-15a and miR-15b modulate natural killer and CD8(+)T-cell activation and anti-tumor immune response by targeting PD-L1 in neuroblastoma. Mol Ther Oncol. (2022) 25:308–29. doi: 10.1016/j.omto.2022.03.010
Keywords: miRNA, miR-136, metastatic tumors, target, immunotherapy
Citation: Wang C, Chen Z, Ni W, Wang J and Zhou W (2025) Research and progress of microRNA-136 in metastatic tumors. Front. Oncol. 15:1555270. doi: 10.3389/fonc.2025.1555270
Received: 04 January 2025; Accepted: 11 February 2025;
Published: 04 March 2025.
Edited by:
Yingcheng Charles Wu, Fudan University, ChinaReviewed by:
Lubna Tariq, Baba Ghulam Shah Badshah University, IndiaCopyright © 2025 Wang, Chen, Ni, Wang and Zhou. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Wei Zhou, MjAwNWx5MDkwOUBodXN0LmVkdS5jbg==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.