
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Oncol. , 28 March 2025
Sec. Head and Neck Cancer
Volume 15 - 2025 | https://doi.org/10.3389/fonc.2025.1528822
 Dominika Gedeonová1,2†
Dominika Gedeonová1,2† Claretta Bianchi1,3†Jan Štembírek1,2,4Matouš Hrdinka1,3
Claretta Bianchi1,3†Jan Štembírek1,2,4Matouš Hrdinka1,3 Zuzana Chyra5,6
Zuzana Chyra5,6 Marcela Buchtová4,7
Marcela Buchtová4,7 Pavel Hurník8,9
Pavel Hurník8,9 Tomáš Blažek10
Tomáš Blažek10 Jana Režnarová1,3*
Jana Režnarová1,3*Oral squamous cell carcinoma (OSCC), a subset of head and neck cancers, primarily originates in the epithelial tissues of the oral cavity. Despite advancements in treatment, the mortality rate for OSCC remains around 50%, underscoring the urgent need for improved prognostic markers. This review explores the role of the BRCA1 and BRCA2 genes—traditionally associated with breast and ovarian cancers—in the context of OSCC. We discuss the molecular pathways involving BRCA genes, their potential as diagnostics and prognostic biomarkers, and their implications for personalized treatment strategies, including addressing chemotherapy resistance. Furthermore, this review emphasizes the significance of genome stability in cancer progression and examines both current and emerging methodologies for detecting BRCA mutations in OSCC patients. Despite limited prevalence of BRCA mutations in OSCC compared to other cancers, their role in DNA repair and therapeutic response underscores their potential as clinical biomarkers. However, standardized, multicenter studies are still needed to validate their utility in OSCC management. A better understanding of the role of BRCA genes in OSCC could pave the way for more effective therapeutic approaches and improved patient outcomes.
Oral squamous cell carcinoma (OSCC), a subtype of Head and Neck Squamous Cell Carcinoma (HNSCC), typically originates in the epithelial tissue of the gingiva, tongue, buccal mucosa, palate, and oral floor (1). While OSCC ranks as the sixteenth most common cancer globally, it is the second most widespread in certain high-risk regions (e.g., South Asia) (2), particularly due to the consumption of carcinogen-containing products (3). In contrast, the increasing incidence of oropharyngeal squamous cell carcinomas in Western countries, including the USA, has been linked to an increase in oropharyngeal human papillomavirus (HPV) infection (4). OSCC arises from multifactorial interactions between genetic mutations, environmental exposures, and immune dysregulation, making personalized treatment approaches particularly challenging (5) (Figures 1A–D). The progression to invasive OSCC involves a series of cellular changes, beginning with epithelial hyperplasia, progressing through various grades of dysplasia, and culminating in invasive carcinoma. These changes are driven by genomic alterations (6), which disrupt the balance between oncogenic and suppressor signaling pathways (5).
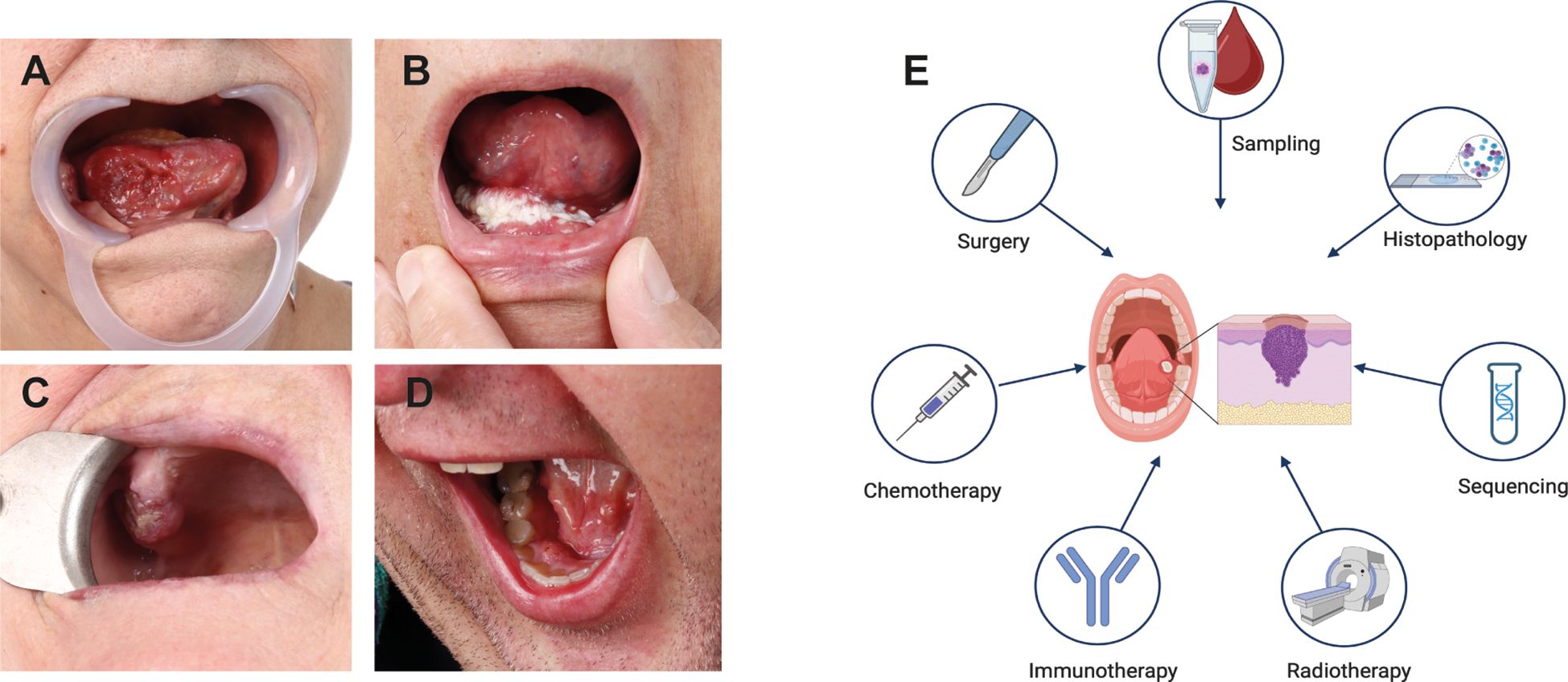
Figure 1. OSCC Tumor Location and Treatment Modalities. (A-D) Most common sites of OSCC occur in the oral cavity. (A) OSCC growing endophytically on the right side of the tongue. (B) OSCC of the floor of the oral cavity extending to the tongue and the mucosa of the alveolar ridge on the right. (C) OSCC affecting the alveolar process of the upper jaw on the right side. (D) OSCC located on the lingual side of the lower jaw’s alveolar process on the right side. (E) Schematic representation of commonly used OSCC treatment strategies. Created in https://BioRender.com.
Oncogenic pathways, such as EGFR, PI3K/AKT/mTOR, JAK/STAT, MET, Wnt/β-catenin, and RAS/RAF/MAPK, are often abnormally activated in OSCC, while tumor suppressor pathways like TP53/RB, p16/Cyclin D1/Rb, and NOTCH are frequently inactivated (5).
Among these genomic alterations, BRCA1 and BRCA2—genes traditionally associated with breast and ovarian cancers—are emerging as key players in OSCC due to their roles in maintaining genomic stability and regulating DNA damage repair. This review explores the potential of BRCA1 and BRCA2 as prognostic markers in OSCC by elucidating their involvement in molecular pathways and their diagnostic or therapeutic implications. We also aim to provide insights into their significance in OSCC management and patient outcomes.
Maintaining genome stability is crucial for cellular survival, necessitating an effective DNA damage response (DDR). DDR mechanisms repair double-strand breaks (DSBs) through two primary mechanisms: non-homologous end-joining (NHEJ) and homologous recombination (HR) (7) (Figure 2A).
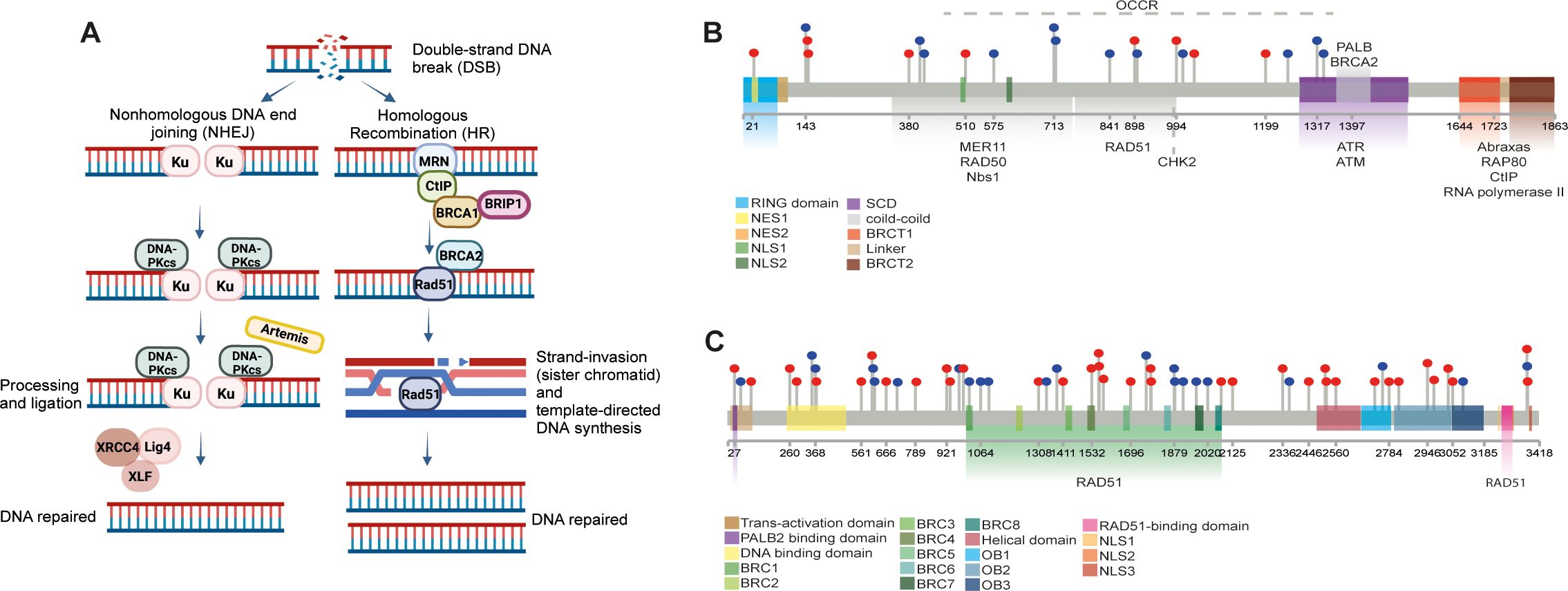
Figure 2. Overview of DNA Damage Response (DDR) and BRCA1/2 Functions in OSCC. (A) Schematic illustration of the DNA damage response (DDR) pathways, highlighting the roles of non-homologous (NHEJ) and homologous recombination (HR) in repairing double-strand breaks (DSBs). The involvement of BRCA1 and BRCA2 in HR-mediated repair of damaged DNA is depicted. Created at https://BioRender.com. (B, C) Lollipop schematic diagrams of BRCA1 and BRCA2 proteins, illustrating their structural domains and interactions with other HR-related proteins and enzymes necessary for DNA repair. Single-point mutations in BRCA1 and BRCA2 genes retrieved from online databases (COSMIC, cBioPortal, dbSNP, ClinVar), are indicated by lollipop markers.
The BRCA proteins–BRCA1 and BRCA2–play integral roles in HR-mediated DSB repair (8). The BRCA1 is characterized by two structural domains: the zinc-binding RING domain at the N-terminus and two phosphopeptide-binding BRCT domains at the C-terminus, facilitating complex protein interactions necessary for DNA repair (9). The RING domain, in combination with BRCA1-associated RING domain protein 1 (BARD1), forms a heterodimer that exhibits E3-ubiquitin ligase activity, essential for tagging damaged DNA and proteins for repair processes (10–12) (Figure 2B).
Similarly, BRCA2 contains several functional domains that interact with RAD51 recombinase, crucial for mediating the HR process. For instance, the N-terminal domain interacts with PALB2, a key scaffolding protein in the BRCA complex (13, 14); the central part of BRCA2 contains eight BRC repeats essential for binding RAD51 (15); and the C-terminus mediates DNA interactions via the DNA binding domain (DBD), which contributes to the initiation and stabilization of the repair process (16) (Figure 2C).
The HR pathway of DDR employs various proteins to accurately repair DNA without introducing mutations. Initial sensors recognize the break, and signaling mediators engage effectors to restore the damage (Figure 2A). Proteins such as ATM and ATR are the first to detect DNA disruption and initiate repair mechanisms. BRCA1 is recruited to DSBs through the Abraxas–RAP80 protein complex (17) and interacts with the MRN complex (MRE11-RAD50-NBS1), facilitating the recruitment of CtIP protein, which catalyzes DNA end resection alongside exonuclease EXOI (18).
BRCA1 also plays a central role in facilitating BRCA2 recruitment to DSBs, subsequently binds RAD51 and initiates HR on the single-stranded DNA (ssDNA) (19). The BRCA1-interacting protein 1 (BRIP1) binds to BRCA1 at the BRCT domain, unwinding DNA during HR and enabling the recruitment of RAD51 (20). This process begins with the formation of a D-loop DNA structure by RAD51, enabling template-directed DNA synthesis (21). Polymerase δ extends the invading strand, and DNA ligase seals the strand breaks, fully restoring DNA integrity (22).
In addition to DSB repair, genome stability is also compromised by interstrand crosslinks (ICLs). The Fanconi anemia (FA) pathway plays a crucial role in repairing ICLs. At least 19 proteins are involved in this pathway, including BRCA1 (FANCS), BRCA2 (FANCD1), RAD51 (FANCR) and BRIP1 (FANCJ). The FA pathway core complex recognizes ICLs and facilitates the monoubiquitination of Fanconi anemia group D2 protein (FANCD2), which is essential for activating the pathway. The monoubiquitinated FANCD2 interacts with BRCA1, allowing the recruitment of HR repair machinery, including BRCA2 and RAD51. BRIP1 is also recruited to unwind the DNA and support the repair process, resolving crosslinks and enabling the DNA repair proteins to access the damage (23).
In the context of OSCC, disruptions in BRCA1 and BRCA2 due to mutations or dysregulations significantly impair the HR and FA pathways, leading to genomic instability and contributing to carcinogenesis. This underscores the importance of BRCA1 and BRCA2 not only in hereditary cancers but also in the pathophysiology of OSCC, positioning them as potential therapeutic targets and valuable prognostic biomarkers (24).
The role of BRCA genes in maintaining genomic integrity suggests their potential significance in multiple cancers, including OSCC. Recent research on BRCA gene alterations in OSCC has employed various approaches, including proteomic, genomic, and differential gene expression studies. Immunohistochemistry (IHC) is commonly used to examine BRCA protein distribution in tumorous tissues, while quantitative PCR (qPCR) evaluates differential gene expression (25–27). However, more precise sequencing methods are needed to identify specific gene mutations, which could provide deeper insights into the genetic underpinnings of OSCC (28–30).
Several studies have investigated the role of BRCA proteins in the transformation of oral leukoplakia (OLK) to OSCC. Vora et al. (31) conducted a detailed evaluation of BRCA1 expression in specimens from 77 patients with early-stage and locally advanced SCC of the tongue and 18 patients with leukoplakia of the tongue, utilizing IHC techniques with a semi-quantitative staining intensity score ranging from negative (no staining) to 3+ (31). Their findings indicated that BRCA1 protein levels were higher in OLK compared to OSCC, suggesting potential downregulation during tumorigenesis. Among OLK samples with a staining intensity of 2+, hyperplastic tissues exhibited lower BRCA1 expression compared to dysplastic tissues. The distribution of BRCA1 in all evaluated tissues was predominantly cytoplasmic, and the protein was completely absent in 66% of OSCC samples, suggesting that the loss of BRCA1 function is relevant to neoplastic transformation. Vora et al. further noted that the observed cytoplasmic staining of BRCA1 may be attributed to naturally occurring alternatively spliced variants of BRCA1 that lack most of the exon 11 sequences and do not possess a nuclear localization signal. These cytoplasmic variants were prevalent in OSCC tissues. Interestingly, among patients with a family history of cancer, 63% expressed BRCA1 were with 1+ (80%) and 2+ (20%) staining intensities. In early-stage disease, BRCA1-positive patients exhibited reduced relapse-free survival compared to BRCA1-negative patients. However, no correlation was observed between clinicopathological parameters and BRCA1 expression. Additionally, a positive correlation between BRCA1 and c-myc expression, used as a predictor of unfavorable prognosis, was identified, further supporting the dynamic role of BRCA1 across different stages of OSCC development. These findings underscore the need for further research to validate BRCA1 as a prognostic marker.
BRCA1 and γH2AX were proposed as independent prognosis markers of OSCC by Oliveira-Costa et al. (2014), who evaluated protein and RNA levels (25). γH2AX,a phosphorylated histone protein correlated with DNA damage (32), was positively associated with poor overall survival. Interestingly, BRCA1 was detected predominantly in the cytoplasm, an unusual localization under conditions of cellular stress induced by DNA damage. These findings emphasize the need for further investigation into BRCA1 expression patterns and their clinical significance. Cytoplasmic localization of BRCA1/2, which typically reflects impaired functionality, has been documented in various cancers, including breast, prostate, gastric, colorectal, and pancreatic cancers (33–35). BRCA1 mutations in the BRCT domain are known to alter its nuclear localization, driving cytoplasmic retention and loss of nuclear repair functionality (36, 37). Chen et al. (38) demonstrated that BRCA1 was entirely mislocalized to the cytoplasm in breast cancer cells (39). Interestingly, cytoplasmic BRCA1/2 expression has been linked to better prognosis in breast and gastric cancers but is less explored in OSCC (33, 40).
A more recent study by Irani and Rafidazeh (2020) further explored the expression profiles of BRCA1 and BRCA2 in OSCC through retrospective analysis (26). Their findings indicated that 63.3% of intermediate and high-grade OSCC tissues exhibited moderate to strong cytoplasmic BRCA1 immunoreactivity, while only 28.3% displayed nuclear BRCA1 expression. Similarly, BRCA2 expression was entirely cytoplasmic, with 55.01% of intermediate and high-grade tissues showing moderate to strong cytoplasmic immunoreactivity. Only four low-grade samples (two for each protein) displayed strong cytoplasmic BRCA1/2 expression. Importantly, BRCA1 was also detected at the invasive front and in the detached tumor cells, suggesting its possible involvement in epithelial-mesenchymal transition (EMT) processes via regulation of E-cadherin and vimentin levels (41). These findings suggest that the aberrant subcellular localization of BRCA1/2, along with their expression at the invasive front, plays an essential role in OSCC pathogenesis and could serve as prognostic markers.
Despite these promising findings, variability across studies raises questions about methodological differences. Specifically, Vora et al. (31) used an antibody targeting the N-terminal region of BRCA1 (clone MS13), while Irani and Rafidazeh employed the MS110 antibody, whose immunogen was undisclosed (26, 31). Interestingly, Oliveira-Costa et al. used the same antibody as Vora et al. but reported findings more consistent with those of Irani and Rafidazeh. This suggests that factors beyond antibody selection, such as sample size, patient demographics, and study design, may also contribute to observed differences. Additionally, the choice of experimental endpoints, such as whether cytoplasmic versus nuclear localization is prioritized, may further influence conclusions across studies. Standardized methodologies and larger cohorts will be critical to reconciling these discrepancies.
Beyond their subcellular localization, genomic studies have provided further insights into BRCA1/2 mutations in OSCC pathogenesis. Exome sequencing of OLK and OSCC has identified BRCA1 and BRCA2 as key markers distinguishing progressive from non-progressive lesions (42). Using multivariate analysis, researchers demonstrated that BRCA1/2 expression decreases with lesion severity, from normal tissue to OSCC. Interestingly, these findings contrast with earlier studies, possibly due to differences in methodology.
High-resolution array-based comparative genomic hybridization (aCGH) has revealed frequent mutations in DDR pathway genes, including BRCA1, BRCA2, FANCD2, and FANCG, in over 25% of OSCC samples (43). Amplifications in BRCA1 and FANCG were observed in 33% and 29% of samples, respectively, while deletions in BRCA2 and FANCD2 occurred in 38% and 33% of samples. These findings highlight the critical role of the FA/BRCA pathway, which is central to DDR, in OSCC pathogenesis. BRCA1 (FANCS) and BRCA2 (FANCD1) are integral components of this pathway, which is responsible for repairing DNA interstrand crosslinks and maintaining genomic stability.
Disruptions in the FA/BRCA pathway, such as BRCA1/2 mutations, impair DNA damage repair processes, leading to increased genomic instability and heightened susceptibility to malignancies, including OSCC. Supporting this, studies on Fanconi anemia (FA)—a genetic disorder caused by mutations in one of 22 genes involved in the FA pathway—highlight a strong connection between FA gene dysfunction and OSCC. FA patients frequently develop OSCC, underscoring the importance of BRCA1/2 and other FA pathway genes in the disease’s pathogenesis (44). The high prevalence of OSCC in FA patients provides indirect evidence that alterations in BRCA1/2 contribute to tumorigenesis in non-FA populations as well. These findings reinforce the potential of BRCA genes as biomarkers for identifying genomic instability in OSCC and highlight their importance as therapeutic targets in this malignancy.
Oral squamous cell carcinoma (OSCC) develops through a complex interplay of genetic and environmental factors, contributing to its aggressive nature and genomic instability. Identifying metabolic, molecular, and immune characteristics that serve as predictive and prognostic markers remains a significant challenge. Despite advances in therapy and research, the overall survival rate for OSCC patients has stagnated at around 50%, primarily due to challenges such as late-stage diagnosis, limited availability of reliable biomarkers, and resistance to conventional therapies (25, 45–47). This highlights the urgent need for more effective biomarkers and therapeutic targets to improve early detection, prognostication, and treatment strategies.
BRCA1 and BRCA2 genes are well-known for their critical roles in DDR, particularly in HR, a highly accurate DNA repair mechanism (48). While mutations in these genes have been extensively studied in breast and ovarian cancers (49), their significance in other malignancies, including OSCC, is an emerging area of research. Alterations in BRCA gene expression, even in the absence of frequent mutations, have been implicated in OSCC tumorigenesis. Recent studies suggest that BRCA1 and BRCA2 expression patterns vary with tumor grade, potentially reflecting their roles in tumor progression (25–27). However, precise data on the frequency of BRCA mutations in OSCC remain limited. While somatic BRCA mutations in breast and ovarian cancers range between 5–20% (47, 50, 51), and pathogenic BRCA2 mutations occur in 2% of pancreatic cancers (52), mutations in OSCC or HNSCC are believed to be rare (25, 26, 28–31, 43, 53). The rarity of BRCA mutations in OSCC may reflect differences in tissue-specific genetic instability and environmental exposures that shape the mutational landscape (51).
Despite the low prevalence of BRCA mutations in OSCC, their altered expression and functional roles in DNA repair highlight their potential as biomarkers. Genomic profiling of OSCC samples has identified occasional BRCA1 and BRCA2 mutations, suggesting their contributions to tumor onset and progression (28–30).
Studies on oral leukoplakia (OLK) further suggest that BRCA1 and BRCA2 expression levels may distinguish high-risk dysplastic lesions that progress to carcinoma from those that remain benign (42, 54). Thus, assessing BRCA1/2 expression using standardized IHC scoring could help stratify patients based on their progression risk, enabling more targeted interventions. These findings underscore the potential utility of BRCA genes as early biomarkers in OPMD, offering an opportunity to identify patients at greatest risk of developing OSCC and enabling timely intervention.
The functions of BRCA1 and BRCA2 in DNA repair and cell cycle regulation are of particular interest because defects in these processes lead to genomic instability, a hallmark of carcinogenesis. Proper cell cycle regulation and DNA repair are essential for maintaining the integrity of the oral epithelium, which is highly susceptible to mutagenic damage from external factors such as tobacco, alcohol, and HPV infection, as well as internal cellular processes (5, 55). Studies suggest that DDR processes, including BRCA1 activity, are activated during the early stages of OSCC to mitigate genomic instability and support tumor survival. Increased expression of BRCA1 and other DNA repair proteins, such as p53, γ-H2AX, RAD51, and 53BP1, has been observed in early OSCC development, highlighting the potential of these molecules, particularly BRCA1, as prognostic biomarkers (25, 55, 56). The potential involvement of BRCA genes in treatment response has also gained attention. Standard care for locally advanced OSCC typically includes surgery followed by adjuvant radiotherapy, with or without chemotherapy (57) (Figure 1E). However, treatment resistance remains a significant obstacle (58). Molecular targeted therapies using specific antibodies against epidermal growth factor receptor (EGFR) and programmed cell death protein 1 (PD-1) have shown promise (59). BRCA1 expression has been implicated in modulating chemotherapy-induced DNA damage, influencing responses to chemotherapeutic agents such as cisplatin and paclitaxel (60, 61). Increased BRCA1 expression may enhance DNA repair, reducing sensitivity to DNA-damaging agents like cisplatin, while simultaneously increasing sensitivity to microtubule-targeting agents such as paclitaxel and docetaxel (62–64). Similarly, in tongue squamous cell carcinoma, the BRCA1-miR-593-5p-MFF axis regulates cisplatin sensitivity by modulating mitochondrial fission and apoptosis, further highlighting the complex role of BRCA1 in therapeutic responses (64).
Like BRCA1, BRCA2 plays a critical role in maintaining genomic stability (Figures 2B, C), and its mutations may impair DNA repair pathways, increasing susceptibility to oncogenic mutations in oral tissues (27). This has significant implications for targeted therapies, particularly Poly(ADP-ribose) polymerase (PARP) inhibitors(PARPi). PARPi block the activity of PARP enzymes, which are involved in repairing single-strand DNA breaks (65–67). In cells with BRCA mutations, where HR is dysfunctional, PARP inhibition leads to the accumulation of unrepaired DNA damage, resulting in cell death through synthetic lethality (68, 69). Given that OSCC often exhibits aberrant DNA repair pathways, including nucleotide excision repair (NER), base excision repair (BER), and double-strand break repair (DSBR) (70–73), PARPi represent a promising therapeutic strategy. However, emerging resistance mechanisms, such as secondary mutations restoring homologous recombination, highlight the need for combination strategies to enhance efficacy (74). Incorporating genetic analysis and molecular profiling into standard diagnostic protocols could identify OSCC patients most likely to benefit from PARPi and other targeted therapies, thereby improving outcomes.In summary, BRCA1 and BRCA2 are central to maintaining genomic stability by facilitating the repair of DNA double-strand breaks through homologous recombination. Their mutations or dysregulation in OSCC can disrupt DNA repair mechanisms, leading to genomic instability and oncogenic transformation. While their role in OSCC is not as well-defined as in breast and ovarian cancers, emerging evidence suggests that BRCA1 and BRCA2 contribute to tumor initiation, progression, and therapeutic resistance (25–27, 53, 75). Moreover, their involvement in the Fanconi anemia pathway and interactions with other DDR proteins underscore their significance in complex repair networks. Future studies focusing on the standardization of methodologies, larger cohorts, and detailed molecular analyses are essential to validate BRCA1 and BRCA2 as biomarkers and therapeutic targets in OSCC. Such advances could pave the way for more personalized and effective treatment strategies, ultimately improving patient outcomes.
DG: Conceptualization, Resources, Visualization, Writing – original draft, Writing – review & editing. CB: Conceptualization, Visualization, Writing – original draft, Writing – review & editing. JŠ: Funding acquisition, Supervision, Writing – review & editing. MH: Funding acquisition, Supervision, Visualization, Writing – review & editing. ZC: Supervision, Writing – review & editing. MB: Supervision, Writing – review & editing. PH: Writing – review & editing. TB: Writing – review & editing. JR: Conceptualization, Visualization, Writing – original draft, Writing – review & editing.
The author(s) declare that financial support was received for the research and/or publication of this article. This article has been produced with the financial support of the European Union under the LERCO (project number CZ.10.03.01/00/22_003/0000003) via the Operational Program Just Transition, the Institutional Supported by MH CZ-DRO-FNOs/2023.) and by the Ministry of Health, Czech Republic (NV19-08-00383).
We thank colleagues and members of our team for the stimulating discussions.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Gormley M, Creaney G, Schache A, Ingarfield K, Conway DI. Reviewing the epidemiology of head and neck cancer: definitions, trends and risk factors. Br Dent J. (2022) 233:780–6. doi: 10.1038/s41415-022-5166-x
2. WHO. International agency for research of cancer, Cancer today, Globocan 2020. (2020). Available at: https://gco.iarc.fr.
3. Hashibe M, Brennan P, Benhamou S, Castellsague X, Chen C, Curado MP, et al. Alcohol drinking in never users of tobacco, cigarette smoking in never drinkers, and the risk of head and neck cancer: pooled analysis in the international head and neck cancer epidemiology consortium. JNCI J Natl Cancer Instit. (2007) 99:777–89. doi: 10.1093/jnci/djk179
4. Mehanna H, Beech T, Nicholson T, El-Hariry I, McConkey C, Paleri V, et al. Prevalence of human papillomavirus in oropharyngeal and nonoropharyngeal head and neck cancer—systematic review and meta-analysis of trends by time and region. Head Neck. (2013) 35:747–55. doi: 10.1002/hed.22015
5. Tan Y, Wang Z, Xu M, Li B, Huang Z, Qin S, et al. Oral squamous cell carcinomas: state of the field and emerging directions. Int J Oral Sci. (2023) 15:44. doi: 10.1038/s41368-023-00249-w
6. Johnson DE, Burtness B, Leemans CR, Lui VWY, Bauman JE, Grandis JR. Head and neck squamous cell carcinoma. Nat Rev Dis Prime. (2020) 6:92. doi: 10.1038/s41572-020-00224-3
7. Gudmundsdottir K, Ashworth A. The roles of BRCA1 and BRCA2 and associated proteins in the maintenance of genomic stability. Oncogene. (2006) 25:5864–74. doi: 10.1038/sj.onc.1209874
8. Li M, Yu X. Function of BRCA1 in the DNA damage response is mediated by ADP-ribosylation. Cancer Cell. (2013) 23:693–704. doi: 10.1016/j.ccr.2013.03.025
10. Clark SL, Rodriguez AM, Snyder RR, Hankins GDV, Boehning D. STRUCTURE-FUNCTION OF THE TUMOR SUPPRESSOR BRCA1. Comput Struct Biotechnol J. (2012) 1:e201204005. doi: 10.5936/csbj.201204005
11. Wu LC, Wang ZW, Tsan JT, Spillman MA, Phung A, Xu XL, et al. Identification of a RING protein that can interact in vivo with the BRCA1 gene product. Nat Genet. (1996) 14:430–40. doi: 10.1038/ng1296-430
12. Roy R, Chun J, Powell SN. BRCA1 and BRCA2: different roles in a common pathway of genome protection. Nat Rev Cancer. (2012) 12:68–78. doi: 10.1038/nrc3181
13. Wu S, Zhou J, Zhang K, Chen H, Luo M, Lu Y, et al. Molecular mechanisms of PALB2 function and its role in breast cancer management. Front Oncol. (2020) 10:301. doi: 10.3389/fonc.2020.00301
14. Le HP, Heyer WD, Liu J. Guardians of the genome: BRCA2 and its partners. Genes. (2021) 12:1229. doi: 10.3390/genes12081229
15. Yoshida K, Miki Y. Role of BRCA1 and BRCA2 as regulators of DNA repair, transcription, and cell cycle in response to DNA damage. Cancer Sci. (2004) 95:866–71. doi: 10.1111/j.1349-7006.2004.tb02195.x
16. Kwon Y, Rösner H, Zhao W, Selemenakis P, He Z, Kawale AS, et al. DNA binding and RAD51 engagement by the BRCA2 C-terminus orchestrate DNA repair and replication fork preservation. Nat Commun. (2023) 14:432. doi: 10.1038/s41467-023-36211-x
17. Wang B, Matsuoka S, Ballif BA, Zhang D, Smogorzewska A, Gygi SP, et al. Abraxas and RAP80 form a BRCA1 protein complex required for the DNA damage response. Science. (2007) 316:1194–8. doi: 10.1126/science.1139476
18. McCarthy-Leo C, Darwiche F, Tainsky MA. DNA repair mechanisms, protein interactions and therapeutic targeting of the MRN complex. Cancers. (2022) 14:5278. doi: 10.3390/cancers14215278
19. Fradet-Turcotte A, Sitz J, Grapton D, Orthwein A. BRCA2 functions: from DNA repair to replication fork stabilization. Endocrine-Related Cancer. (2016) 23:T1–17. doi: 10.1530/ERC-16-0297
20. Li X, Heyer W-D. Homologous recombination in DNA repair and DNA damage tolerance. Cell Res. (2008) 18:99–113. doi: 10.1038/cr.2008.1
21. Wyman C, Ristic D, Kanaar R. Homologous recombination-mediated double-strand break repair. DNA Repair. (2004) 3:827–33. doi: 10.1016/j.dnarep.2004.03.037
22. Ranjha L, Howard SM, Cejka P. Main steps in DNA double-strand break repair: an introduction to homologous recombination and related processes. Chromosoma. (2018) 127:187–214. doi: 10.1007/s00412-017-0658-1
23. Ceccaldi R, Sarangi P, D’Andrea AD. The fanconi anaemia pathway: new players and new functions. Nat Rev Mol Cell Biol. (2016) 17:337–49. doi: 10.1038/nrm.2016.48
24. Prime SS, Darski P, Hunter KD, Cirillo N, Parkinson EK. A review of the repair of DNA double strand breaks in the development of oral cancer. IJMS. (2024) 25:4092. doi: 10.3390/ijms25074092
25. Oliveira-Costa JP, Oliveira LR, Zanetti JS, Silveira GGD, Buim MEC, Zucoloto S, et al. BRCA1 and γH2AX as independent prognostic markers in oral squamous cell carcinoma. Oncoscience. (2014) 1:383–91. doi: 10.18632/oncoscience.47
26. Irani S, Rafizadeh M. BRCA1/2 expression patterns in different grades of oral squamous cell carcinoma. Middle East J Cancer. (2020) 11:390–8. doi: 10.30476/mejc.2020.81282.0
27. Mistry NH, Gosavi SR, Dhobley A, Mishra S, Kherde P. Expression of breast cancer gene 2 in oral squamous cell carcinoma and its correlation with the metastatic potential: A retrospective study. J Precis Oncol. (2022) 2:120–4. doi: 10.4103/jpo.jpo_25_22
28. Vossen DM, Verhagen CVM, Verheij M, Wessels LFA, Vens C, Van Den Brekel MWM. Comparative genomic analysis of oral versus laryngeal and pharyngeal cancer. Oral Oncol. (2018) 81:35–44. doi: 10.1016/j.oraloncology.2018.04.006
29. Koo K, Mouradov D, Angel CM, Iseli TA, Wiesenfeld D, McCullough MJ, et al. Genomic signature of oral squamous cell carcinomas from non-smoking non-drinking patients. Cancers. (2021) 13:1029. doi: 10.3390/cancers13051029
30. Biswas NK, Das C, Das S, Maitra A, Nair S, Gupta T, et al. Lymph node metastasis in oral cancer is strongly associated with chromosomal instability and DNA repair defects. Intl J Cancer. (2019) 145:2568–79. doi: 10.1002/ijc.v145.9
31. Vora HH, Shah NG, Patel DD, Trivedi TI, Choksi TJ. BRCA1 expression in leukoplakia and carcinoma of the tongue. J Surg Oncol. (2003) 83:232–40. doi: 10.1002/jso.10213
32. Rogakou EP, Boon C, Redon C, Bonner WM. Megabase chromatin domains involved in DNA double-strand breaks in vivo. J Cell Biol. (1999) 146:905–16. doi: 10.1083/jcb.146.5.905
33. Santivasi WL, Wang H, Wang T, Yang Q, Mo X, Brogi E, et al. Association between cytosolic expression of BRCA1 and metastatic risk in breast cancer. Br J Cancer. (2015) 113:453–9. doi: 10.1038/bjc.2015.208
34. Thorgeirsson T, Jordahl KM, Flavin R, Epstein MM, Fiorentino M, Andersson S-O, et al. Intracellular location of BRCA2 protein expression and prostate cancer progression in the swedish watchful waiting cohort. Carcinogenesis. (2016) 37:262–8. doi: 10.1093/carcin/bgw001
35. Wang G-H, Zhao C-M, Huang Y, Wang W, Zhang S, Wang X. BRCA1 and BRCA2 expression patterns and prognostic significance in digestive system cancers. Hum Pathol. (2018) 71:135–44. doi: 10.1016/j.humpath.2017.10.032
36. Drikos I, Nounesis G, Vorgias CE. Characterization of cancer-linked BRCA1-BRCT missense variants and their interaction with phosphoprotein targets. Proteins. (2009) 77:464–76. doi: 10.1002/prot.22460
37. Drikos I, Boutou E, Kastritis PL, Vorgias CE. BRCA1-BRCT mutations alter the subcellular localization of BRCA1. In Vitro. Anticancer Res. (2021) 41:2953–62. doi: 10.21873/anticanres.15077
38. Chen Y, Chen CF, Riley DJ, Allred DC, Chen PL, Von Hoff D, et al. Aberrant subcellular localization of BRCA1 in breast cancer. Science. (1995) 270(5237):789–91. doi: 10.1126/science.270.5237.789
39. Krais JJ, Johnson N. BRCA1 mutations in cancer: Coordinating deficiencies in homologous recombination with tumorigenesis. Cancer Res. (2020) 80:4601–9. doi: 10.1158/0008-5472.CAN-20-1830
40. Rakha EA, El-Sheikh SE, Kandil MA, El-Sayed ME, Green AR, Ellis IO. Expression of BRCA1 protein in breast cancer and its prognostic significance. Hum Pathol. (2008) 39:857–65. doi: 10.1016/j.humpath.2007.10.011
41. Henneman L, Van Miltenburg MH, Michalak EM, Braumuller TM, Jaspers JE, Drenth AP, et al. Selective resistance to the PARP inhibitor olaparib in a mouse model for BRCA1-deficient metaplastic breast cancer. Proc Natl Acad Sci USA. (2015) 112:8409–14. doi: 10.1073/pnas.1500223112
42. Farah CS, Jessri M, Bennett NC, Dalley AJ, Shearston KD, Fox SA. Exome sequencing of oral leukoplakia and oral squamous cell carcinoma implicates DNA damage repair gene defects in Malignant transformation. Oral Oncol. (2019) 96:42–50. doi: 10.1016/j.oraloncology.2019.07.005
43. Sparano A, Quesnelle KM, Kumar MS, Wang Y, Sylvester AJ, Feldman M, et al. Genome-wide profiling of oral squamous cell carcinoma by array-based comparative genomic hybridization. Laryngos. (2006) 116:735–41. doi: 10.1097/01.mlg.0000205141.54471.7f
44. Amenábar JM, Torres-Pereira CC, Tang KD, Punyadeera C. Two enemies, one fight: An update of oral cancer in patients with Fanconi anemia. Cancer. (2019) 125:3936–46. doi: 10.1002/cncr.v125.22
45. Badwelan M, Muaddi H, Ahmed A, Lee KT, Tran SD. Oral squamous cell carcinoma and concomitant primary tumors, what do we know? A review of the literature. Curr Oncol. (2023) 30:3721–34. doi: 10.3390/curroncol30040283
46. Starzyńska A, Sobocki BK, Alterio D. Current challenges in head and neck cancer management. Cancers. (2022) 14:358. doi: 10.3390/cancers14020358
47. Armstrong N, Ryder S, Forbes C, Ross J, Quek RG. A systematic review of the international prevalence of BRCA mutation in breast cancer. CLEP. (2019) 11:543–61. doi: 10.2147/CLEP.S206949
48. Lieber MR. The mechanism of double-strand DNA break repair by the nonhomologous DNA end-joining pathway. Annu Rev Biochem. (2010) 79:181–211. doi: 10.1146/annurev.biochem.052308.093131
49. Petrucelli N, Daly MB, Feldman GL. Hereditary breast and ovarian cancer due to mutations in BRCA1 and BRCA2. Genet Med. (2010) 12:245–59. doi: 10.1097/GIM.0b013e3181d38f2f
50. Loboda AP, Adonin LS, Zvereva SD, Guschin DY, Korneenko TV, Telegina AV, et al. BRCA mutations—The achilles heel of breast, ovarian and other epithelial cancers. IJMS. (2023) 24:4982. doi: 10.3390/ijms24054982
51. Arun B, Couch FJ, Abraham J, Tung N, Fasching PA. BRCA-mutated breast cancer: the unmet need, challenges and therapeutic benefits of genetic testing. Br J Cancer. (2024) 131:1400–14. doi: 10.1038/s41416-024-02827-z
52. Luo G, Lu Y, Jin K, Cheng H, Guo M, Liu Z, et al. Pancreatic cancer: BRCA mutation and personalized treatment. Expert Rev Anticancer Ther. (2015) 15:1223–31. doi: 10.1586/14737140.2015.1086271
53. Hurník P, Režnarová J, Chyra Z, Motyka O, Putnová BM, Čermáková Z, et al. Enhancing oral squamous cell carcinoma prediction: the prognostic power of the worst pattern of invasion and the limited impact of molecular resection margins. Front Oncol. (2023) 13:1287650. doi: 10.3389/fonc.2023.1287650
54. Errazquin R, Carrasco E, Del Marro S, Suñol A, Peral J, Ortiz J, et al. Early diagnosis of oral cancer and lesions in fanconi anemia patients: A prospective and longitudinal study using saliva and plasma. Cancers. (2023) 15:1871. doi: 10.3390/cancers15061871
55. Toprani SM, Kelkar Mane V. A short review on DNA damage and repair effects in lip cancer. Hematology/Oncol Stem Cell Ther. (2021) 14:267–74. doi: 10.1016/j.hemonc.2021.01.007
56. Nikitakis NG, Rassidakis GZ, Tasoulas J, Gkouveris I, Kamperos G, Daskalopoulos A, et al. Alterations in the expression of DNA damage response-related molecules in potentially preneoplastic oral epithelial lesions. Oral Surge Oral Med Oral Pathol Oral Radiol. (2018) 125:637–49. doi: 10.1016/j.oooo.2018.03.006
57. Kende P, Mathur Y, Varte V, Tayal S, Patyal N, Landge J. The efficacy of neoadjuvant chemotherapy as compared to upfront surgery for the management of oral squamous cell carcinoma: a systematic review and meta-analysis. Int J Oral Maxillofac Surge. (2024) 53:1–10. doi: 10.1016/j.ijom.2023.03.007
58. Sa P, Singh P, Panda S, Swain RK, Dash R, Sahoo SK. Reversal of cisplatin resistance in oral squamous cell carcinoma by piperlongumine loaded smart nanoparticles through inhibition of Hippo-YAP signaling pathway. Trans Res. (2024) 268:63–78. doi: 10.1016/j.trsl.2024.03.004
59. Cao M, Shi E, Wang H, Mao L, Wu Q, Li X, et al. Personalized targeted therapeutic strategies against oral squamous cell carcinoma. An evidence-based review of literature. IJN. (2022) 17:4293–306. doi: 10.2147/IJN.S377816
60. Kennedy RD, Quinn JE, Mullan PB, Johnston PG, Harkin DP. The role of BRCA1 in the cellular response to chemotherapy. JNCI J Natl Cancer Instit. (2004) 96:1659–68. doi: 10.1093/jnci/djh312
61. Stordal B, Davey R. A systematic review of genes involved in the inverse resistance relationship between cisplatin and paclitaxel chemotherapy: role of BRCA1. CCDT. (2009) 9:354–65. doi: 10.2174/156800909788166592
62. Tassone P, Tagliaferri P, Perricelli A, Blotta S, Quaresima B, Martelli ML, et al. BRCA1 expression modulates chemosensitivity of BRCA1-defective HCC1937 human breast cancer cells. Br J Cancer. (2003) 88:1285–91. doi: 10.1038/sj.bjc.6600859
63. Quinn JE, Kennedy RD, Mullan PB, Gilmore PM, Carty M, Johnston PG, et al. BRCA1 functions as a differential modulator of chemotherapy-induced apoptosis. Cancer Res. (2003) 63:6221–8.
64. Fan S, Liu B, Sun L, Lv X, Lin Z, Chen W, et al. Mitochondrial fission determines cisplatin sensitivity in tongue squamous cell carcinoma through the BRCA1-miR-593-5p–MFF axis. Oncotarget. (2015) 6:14885–904. doi: 10.18632/oncotarget.3659
65. Wang F, Gouttia OG, Wang L, Peng A. PARP1 upregulation in recurrent oral cancer and treatment resistance. Front Cell Dev Biol. (2022) 9:804962. doi: 10.3389/fcell.2021.804962
66. Luong KV, Wang L, Roberts BJ, Wahl JK, Peng A. Cell fate determination in cisplatin resistance and chemosensitization. Oncotarget. (2016) 7:23383–94. doi: 10.18632/oncotarget.8110
67. Dulaney C, Marcrom S, Stanley J, Yang ES. Poly(ADP-ribose) polymerase activity and inhibition in cancer. Semin Cell Dev Biol. (2017) 63:144–53. doi: 10.1016/j.semcdb.2017.01.007
68. Nambiar KD, Mishra DP, Singh R. Targeting DNA repair for cancer treatment: Lessons from PARP inhibitor trials. Oncol Res. (2023) 31:405–21. doi: 10.32604/or.2023.028310
69. Chen A. PARP inhibitors: its role in treatment of cancer. Chin J Cancer. (2011) 30(7):463–71. doi: 10.5732/cjc.011.10111
70. Moutafi M, Economopoulou P, Rimm D, Psyrri A. PARP inhibitors in head and neck cancer: Molecular mechanisms, preclinical and clinical data. Oral Oncol. (2021) 117:105292. doi: 10.1016/j.oraloncology.2021.105292
71. Cheng L, Sturgis EM, Eicher SA, Spitz MR, Wei Q. Expression of nucleotide excision repair genes and the risk for squamous cell carcinoma of the head and neck. Cancer. (2002) 94:393–7. doi: 10.1002/cncr.10231
72. Chung CH, Guthrie VB, Masica DL, Tokheim C, Kang H, Richmon J, et al. Genomic alterations in head and neck squamous cell carcinoma determined by cancer gene-targeted sequencing. Ann Oncol. (2015) 26:1216–23. doi: 10.1093/annonc/mdv109
73. Li C, Hu Z, Lu J, Liu Z, Wang L, El-Naggar AK, et al. Genetic polymorphisms in DNA base-excision repair genes ADPRT, XRCC1, and APE1 and the risk of squamous cell carcinoma of the head and neck. Cancer. (2007) 110:867–75. doi: 10.1002/cncr.v110:4
74. Lim J, Tan DSP. Understanding resistance mechanisms and expanding the therapeutic utility of PARP inhibitors. Cancers. (2017) 9:109. doi: 10.3390/cancers9080109
Keywords: BRCA1, BRCA2, OSCC (oral squamous cell carcinoma), HNSCC (head and neck squamous cell carcinoma), gene alteration
Citation: Gedeonová D, Bianchi C, Štembírek J, Hrdinka M, Chyra Z, Buchtová M, Hurník P, Blažek T and Režnarová J (2025) BRCA1 and BRCA2 as prognostic markers in oral squamous cell carcinoma: a minireview. Front. Oncol. 15:1528822. doi: 10.3389/fonc.2025.1528822
Received: 15 November 2024; Accepted: 26 February 2025;
Published: 28 March 2025.
Edited by:
Omar Kujan, University of Western Australia, AustraliaReviewed by:
Faris Alabeed, University of Western Australia, AustraliaCopyright © 2025 Gedeonová, Bianchi, Štembírek, Hrdinka, Chyra, Buchtová, Hurník, Blažek and Režnarová. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jana Režnarová, amFuYS5yZXpuYXJvdmFAb3N1LmN6
†These authors share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.