
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Oncol. , 01 November 2024
Sec. Thoracic Oncology
Volume 14 - 2024 | https://doi.org/10.3389/fonc.2024.1479454
This article is part of the Research Topic Editor's Challenge: Dr. Sara Pilotto - Modulating Tumor-Host Interplay through Lifestyle in Lung Cancer View all 3 articles
 Annamaria Mancini1,2
Annamaria Mancini1,2 Francesca Maria Orlandella1,2
Francesca Maria Orlandella1,2 Daniela Vitucci1,2
Daniela Vitucci1,2 Neila Luciano3
Neila Luciano3 Andreina Alfieri1,2
Andreina Alfieri1,2 Stefania Orrù1,2
Stefania Orrù1,2 Giuliana Salvatore1,2*
Giuliana Salvatore1,2* Pasqualina Buono1,2*
Pasqualina Buono1,2*Lung cancer is the major cause of cancer-related deaths worldwide with an estimated 1.8 million deaths and 2.4 million new cases in 2022. Poor cardiorespiratory fitness, dyspnea and fatigue are the common features in lung cancer patients, partially limiting the exercise prescription. Exercise improves cardiorespiratory and muscular fitness and reduces the risk of some types of cancer, including lung cancer. Recently, the American Society of Clinical Oncology has encouraged preoperative exercise for lung cancer patients. Nonetheless, only limited data, mostly obtained from mouse models of lung cancer, are available on the molecular effects of exercise in lung cancer. Thus, the present minireview aims to shed light on the molecular mechanisms induced by different type of exercise in lung cancer. In particular, the role of the exercise in tumor microenvironment remodeling, angiogenesis, gene expression, apoptosis and intermediate metabolism will be examined.
Globally, in 2022 the incidence of Lung Cancer (LC) is estimated of 2.4 million new cases representing, the first and the second most commonly diagnosed malignancy in men and women, respectively. Moreover, with 1.8 million estimated deaths in 2022, this tumor is the first cause of cancer death in men and the second cause in women (1, 2).
The most frequent form of LC is represented by the non-small cell lung cancer (NSCLC). NSCLC further encompass two major subtypes: lung adenocarcinoma (LUAD) and lung squamous cell carcinoma (LUSC). Small cell lung cancer (SCLC) includes approximately 15% of LC cases and is characterized by high proliferative rate, mutational burden and poor survival. The pathogenesis of LC, in particular the SCLC subtype, is mainly due to environmental factors as smoke (3, 4). The tumor is highly heterogenous from a genetic, metabolic and immunological point of view and both spatially and temporally. This heterogenicity has important implication in therapy resistance (5–8).
The therapeutic options for LC patients depend on the tumor stage, grade, histological subtype and the overall condition of the patient. Among the options surgery, chemotherapy and radiotherapy are included; however, LC patients treated with these standard procedures often relapse. Generally, almost all patients with SCLC will relapse, while for NSCLC the percentage of relapse is approximately of 30-50% (9). In the recent years progress in basic cancer research, allowing the identification of key genetic lesions, has improved the scenario therapy for LC patients (10–12). A range of inhibitors targeting key genes are currently available for LC treatment (13).
Immunotherapy is also being used for these patients. The use of antibodies targeting the programmed death receptor (PD-1), its ligand (PD-L1), and the cytotoxic T-lymphocyte-associated protein 4 (CTLA-4) receptor, has improved the survival of patients with NSCLC (14, 15).
Recently, the practice of exercise in cancer patients has gained increasingly attention with potential advantages beyond traditional treatments (16–18). In LC, the use of exercise is in part limited, mainly due to dyspnea being a major symptom. However, the role of exercise preoperatively has been described (19, 20). The American Society of Clinical Oncology has encouraged preoperative exercise in LC patients, for better recovery and minimizing complications after surgery (21–23). In addition, an active lifestyle should be recommended during cancer treatment to help patients not only maintain muscle mass and function but also to promote long-term health (24). As for the postoperative period, structured rehabilitation programs, supervised by professionals, play a key role in ensuring safe exercise after lung surgery, leading to a better recovery. Exercise not only could prevent complications like pneumonia but also could improve emotional health, reducing anxiety and depression. Additionally, it enhances muscle strength and oxygen efficiency, which reduces cardiovascular and pulmonary strain (25). Nevertheless, besides these strong evidences, precise prescription of exercise for people with LC is currently unknown and guidelines are lacking.
This minireview has the focus of giving an update insight on the molecular mechanisms induced by exercise in LC, contributing to current knowledge on the topic (26, 27). In particular, the role of exercise in tumor microenvironment, angiogenesis, gene expression, apoptosis and intermediate metabolism will be dissected. A summary of the main findings emerged from the literature is presented in Figure 1 and Table 1.

Figure 1. A graphical summary of the molecular mechanisms elicited by the exercise in LC.
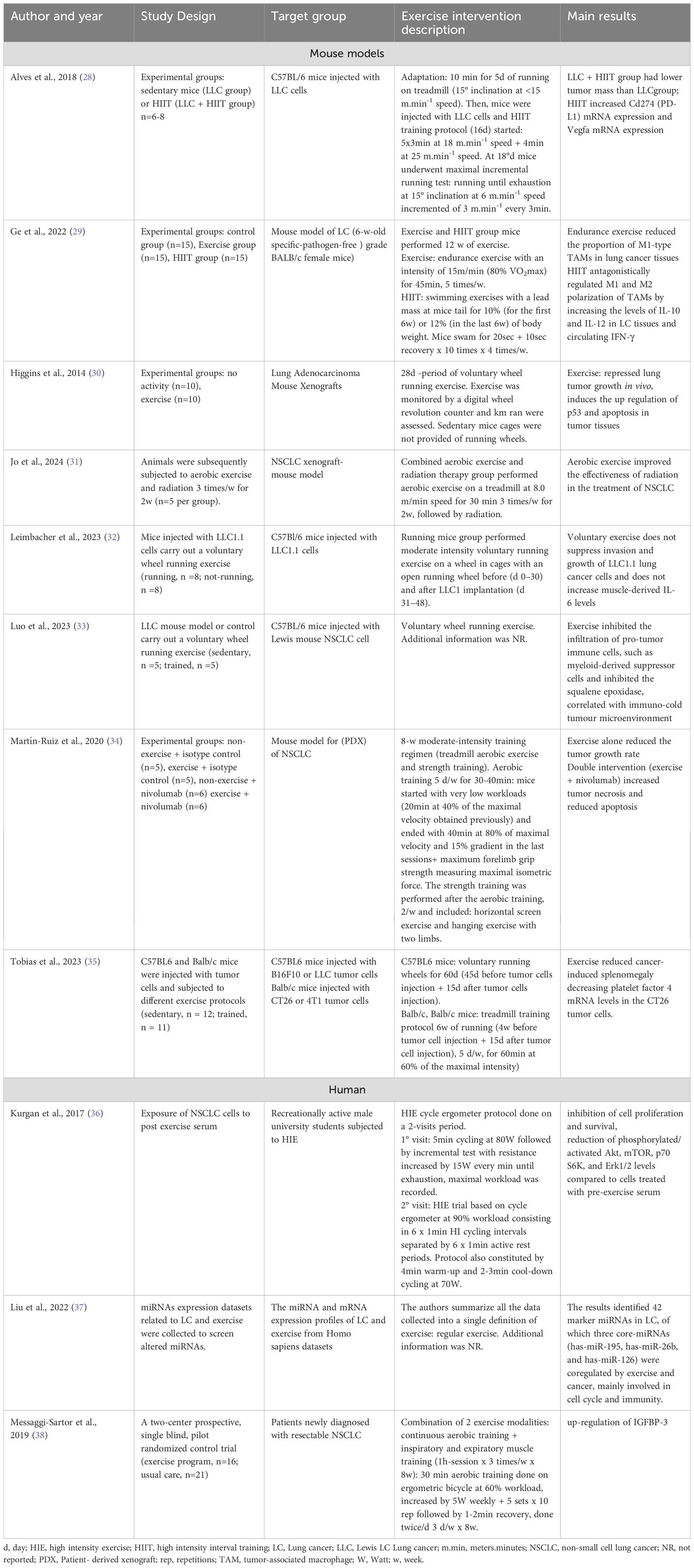
Table 1. Exercise-induced molecular changes in LC.
The tumor microenvironment is a complex and dynamic entity composed by a great variety of elements as immune and stromal cells, macrophages, extracellular matrix and blood vessels (39). To date, the relationship between exercise and tumor microenvironment has been elucidated in different types of cancer as breast (40), pancreatic (41), melanoma (42) and hepatocellular (43) carcinoma. Mechanistically, exercise is reported to be able to remodel the tumor microenvironment through the modulation of immune system (44). For example, in pancreatic cancer, following aerobic exercise an anti-tumor immune cells redistribution occurs characterized by an accumulation of interleukin (IL)-15 and CD8+ T cells in the tumor microenvironment, which are hurdle against tumor growth (41). Also in breast cancer, the potential anticancer effect of exercise is due to an accumulation of CD8+ T cells into tumor microenvironment (40).
However, despite the growing literature on the role of exercise in this context, if and how exercise could affect the tumor microenvironment in LC was only partially investigated. To our knowledge, the first evidence that the exercise intervention could modulate the tumor microenvironment in LC was highlighted by Martín-Ruiz and colleagues in 2020 investigating the effects of combination of exercise and immunotherapy in mice subcutaneously injected with cancer cells derived from NSCLC patient (34). After 8-week of moderate intensity exercise training, the authors evidenced that exercise alone reduced tumor volume and increased neutrophil tumor infiltrates. The combined treatment (exercise and therapy) increased tumor necrosis (34).
The benefits of exercise on tumor microenvironment were also reported in a recent preclinical study performed in a C57BL/6 mice injected with Lewis LC (LLC) mouse model (33). Mice after 20 days of voluntary wheel running, presented a reduction in tumor volume and a higher infiltration of CD8+ T cells, B cells and M1 macrophages compared to the group of non-exercise mice. Further in this LC mouse model, lower concentration of myeloid-derived suppressor cells was observed that, contrariwise, exert a protumor activity. Interestingly, the aerobic exercise training inhibited the squalene epoxidase, an enzyme involved in the reprogramming of cholesterol metabolism notably correlated with immuno-cold tumor microenvironment (33).
Also the link between the exercise and the polarization of tumor-associated macrophage (TAM) was analyzed in a mouse model of LC (29). The endurance exercise induced a reprogramming of M2 macrophages in M1 type, as witnessed by a reduction in the expression of CD86, Tumor necrosis factor (TNF)- α and nitric oxide synthase (iNOS) markers, delaying tumor growth. The authors also investigated the role of high-intensity interval training (HIIT), an anerobic exercise characterized by high-intensity peak (30 sec) interspersed by low-intensity (recovery) efforts (10 sec) (45). HIIT exerts an anti-inflammatory effect since occurred a reduction in cancer tissues of the expression of IL-6, TNF- α and iNOS and an increase in the level of IL-12 and of IL-10 in the HIIT mice group. Interestingly, in the blood of mice performing endurance or HIIT exercise, the circulating levels of interferon gamma (IFN-γ) were increased compared to the non-exercise mice group (29). Overall, these data suggested that benefits of exercise in LC could be also ascribed to the reprogramming of TAM and to the modulation of inflammatory markers cytokines expression.
In contrast, Leimbacher and colleagues asserted that tumor nodules from running mice had greater immune cell infiltration than those from non-running mice. Therefore, exercise improves the oxygenation in the lungs and tumor nodules by reducing anemia, but it did not influence either the lung invasion or the proliferation of LLC1 tumor cells (32).
The intra-tumoral hypoxia, through the activation of several genes expression, enhanced the aggressive behavior of cancer cells and hindered the delivery of drug contributing also to therapy resistance (46). Thus, the knowledge of the effects of exercise intervention on angiogenesis is an important step in the oncology field that could improve the therapeutic response.
Emerging data showed that the improvement of blood vessels could be obtained by acute and chronic aerobic exercise training contributing to a better perfusion and vascularization of tissue (47). However, the molecular pathways underlying the link between exercise and angiogenesis remain poorly understood. For example, it was reported that exercise training increased the recruitment of endothelial progenitor cells from the bone marrow in the blood inducing vascular normalization (48). Moreover, it has been evidenced that exercise could affects breast, prostate and hepatocellular tumor growth and the aggressiveness of these cancer cells by improving tumor angiogenesis through reduction of intra- tumoral hypoxia and enhancement of blood perfusion (49–51). Mechanistically, aerobic exercise is able to increase the expression of several angiogenetic factors, like Hypoxia inducible factor (HIF)-1, ANGIOPOIETINS, Vascular endothelial growth factor (VEGF), Platelet derived growth factor (PDGF), Fibroblast growth factor (FGF) that exert a pro-angiogenetic effect (52).
The relationship between exercise and angiogenesis is also supported in mice bearing hepatocellular carcinoma where the swimming intervention impaired hypoxia through the inhibition of HIF-1α and serine/threonine kinase 1 (AKT)/Glycogen synthase kinase 3 beta (GSK-3β)/β-CATENIN signaling pathways (51).
Besides these evidences in different types of cancer, molecular studies concerning the link between exercise and tumor angiogenesis in LC are lacking.
Recently, the potential therapeutic effects of exercise intervention in LC were suggested by Tobias and colleagues evidencing that aerobic exercise reduced the tumor growth and cancer-induced splenomegaly in Lewis LC bearing mice by decreasing the expression of platelet factor 4 (PF4), a protein correlated to vessel formation (35).
Also, higher levels of VEGF and of PD-L1 were found in mice bearing Lewis LC after HIIT, compared to sedentary tumor mice, that counteract tumor progression by angiogenesis promotion (28).
A growing body of evidence shows that regular endurance and resistance training can induce changes in gene expression profiles by epigenetic mechanisms (53–55). In addition, it has been found that exercise modulates the expression of microRNAs (miRNAs), their release into circulation and their target genes (17, 56–58).
Recent studies revealed alterations in miRNAs expression in LC patients in response to exercise. For instance, Liu and colleagues observed that exercise increased the expression of miR-195, miR-26b and miR-126, which are normally down-regulated in LC. These miRNAs affect biological processes like immune response modulation, tumor suppression and cell cycle regulation. Moreover, differential expression of circulating miR-195, miR-26b and miR-126 and their target genes have been linked to a worse prognosis in LC patients (37).
Recent research focused on unraveling the genes whose expression resulted modified by aerobic exercise in combination with radiotherapy in a xenograft mouse model of NSCLC. The authors found that the combination of exercise with radiation therapy reduced cell growth. Transcriptomic analysis was performed on tissues derived from the treated mice. Differentially expressed genes (DEGs) were analyzed using the Kyoto Encyclopedia of Genes and Genomes (KEGG) and gene ontology (GO), revealing a strong correlation among the different expression of most of DEGs and angiogenesis, vascular and miRNAs. Notably, four genes associated to the ROS pathways, i.e., Glutathione S-transferase mu 5 (GSTM5), mitochondrial permeability transition pore (MPTP), Glutathione S-transferase omega 1 (GSTO1) and solute carrier family 25 member 31 (SLC25A31), showed different expression between the radiation-alone and combination groups. This preclinical study suggests that combining aerobic exercise with radiation therapy may improve LC patients’ treatment outcomes (31).
In summary, these data highlight the potential of aerobic exercise associated to radiation therapy as an effective modulation of cell growth and gene expression in LC patients. However, the exact mechanisms by which exercise affects gene expression in LC patients should still being elucidated.
Autophagy and apoptosis are crucial processes in LC progression, promoting self-regulatory mechanisms in response to cellular stress and death signals (59, 60). The effects of the exercise on cancer cell apoptosis have been confirmed by numerous studies, supporting the beneficial effects as adjuvant therapy, promoting suppression of tumor growth and cancer cell apoptosis (24, 61, 62). Kurgan and colleagues show that serum from a post- high intensity exercise (12 intervals at 90% workload) is able to inhibit proliferation and survival of LC cells, in agreement with previous studies that demonstrated similar effects from a single session of moderate-intensity aerobic exercise (20 minutes at 50% or 40 minutes at 65% of VO2 max) on prostate cancer cells (63), as well as a single session of combined HIIT and resistance training (30 minutes warm-up, 60 minutes of resistance training, and 30 minutes of high-intensity interval spinning on stationary bicycles non è specificato intensità) on breast cancer cells (64). The authors also evidenced that the mechanism underlying this inhibition was due to the phosphorylation/activation of AKT, mechanistic target of rapamycin kinase (mTOR), P70 (ribosomal protein S6 kinase B1) S6K and mitogen-activated protein kinase (ERK)1/2 signaling pathways (36).
The tumor suppressor p53, that was found mutated in 50% of NSCLC, maintains genomic integrity by responding to cellular stress and DNA damage through promotion of cell cycle arrest and DNA repair or apoptosis (30, 65, 66). Higgins and colleagues demonstrated that p53 protein levels were strongly increased in LC of aerobic exercising mice compared to that of sedentary mice. In addition, levels of the pro-apoptotic proteins BCL2 associated X, apoptosis regulator (BAX) and BCL2 antagonist/killer 1 (BAK) were significantly higher in LC tissue from exercising than in sedentary mice tumor, indicating that p53-driven apoptosis occurs in exercise. Similarly, increased levels of the apoptotic intermediate, active CASPASE-3, were found in LC of aerobic exercising compared with sedentary mice tumor. These results suggest that aerobic exercise reduces the cancer growth through the increase of p53 expression and subsequent p53-driven apoptosis (30).
The BECLIN-1 protein is crucial in autophagy initiation and tumor suppression (67). The loss of BECLIN-1 slows-down the autophagy pathway and in turn potentially increases the carcinogenesis by preventing the degradation of harmful agents (59, 67). BECLIN-1, through the interaction with BCL-2 protein, also promotes the release of pro-apoptotic molecules, like BAX and BAK in some type of cancer cells. Additionally, the (ATG)5-ATG12-ATG16 complex, involved in the autophagy pathway activates the apoptosis through ATG12’s interaction with anti-apoptotic BCL-2 proteins, underscoring the complex interplay of autophagy and apoptosis in cancer development (60, 68, 69).
Finally, imbalance of autophagy also plays an important role in cancer cachexia (70). Cachexia occurs in about 50% of patients with lung cancer (71).
Recent research focused on the effects of the endurance and resistance exercise in the induction of proteins, involved in the autophagy pathway, like ATG5, p62, pULK1, in association to cancer cachexia (70, 72–74).
In conclusion, the interaction between exercise-induced apoptosis and LC results intricates yet crucial. Aerobic exercise induces the activation of p53-tumor suppression protein expression in LC mouse models; similarly, exercise increases the expression of BAX and BAK and CASPASE-3 proteins, and as suggested in other types of cancer, exercise could influence the expression of BECLIN-1, a bridge protein, linked to autophagy pathways and other proteins associated to the cachexia.
Energy balance is essential for maintaining cell survival and overall body stability. The AMP-activated protein kinase (AMPK) acts as cellular energy sensor, regulating different signals and metabolic pathways in response to different stimuli, such as obesity (75), aging (29), caloric restriction (76) and exercise (77). AMPK activation can influence different cellular processes, including cell proliferation, apoptosis and the response to oxidative stress (78). Dysregulation of AMPK signaling is a common finding in several cancer types, including LC (79). In recent years, AMPK activation has emerged as a promising therapeutic target for different types of cancer due to its role in cell proliferation and energetics. It is widely recognized that AMPK activation occurs in response to muscle contraction and exercise (80–82). Exercise, as well as other stressful factors such as fasting (83), determines changes in the expression levels of several hormones that activate AMPK and their intracellular signaling pathways to maintain cellular and systemic energetic homeostasis (84). Acute exercise induces AMPK activation (85), which in turn inhibits cancer cell growth and promote protective autophagy in cells activating liver kinase B1 (LKB1), the upstream activator of AMPK (86). Furthermore, AMPK activation improve tissue insulin sensitivity (85). Insulin and insulin-like growth factor (IGF)-1 play important role also in glucose metabolism and cell proliferation. Increased expression of IGF-1 has been described in association with increased risk of different type of cancer (87) although the molecular mechanisms are not completely elucidated. Lung fibroblasts have been shown to synthesize IGF-1 (88). IGF-1 plays a critical role in lung disease, like cancer and lung fibrosis (89); interestingly, the expression of IGF-1 in LC tissue was higher than in adjacent normal lung tissue (90). The IGF-1 receptor (IGF-1R) is a central component of LC signal transduction pathways (91). Overexpression of IGF-1R was reported in NSCLC and SCLC, by Long et al., that evidenced after intrasplenic injection in mice of LC cells and IGF-1R receptor, an increase in the metastatic activity (92). On the contrary, downregulation of IGF-1 and upregulation of IGFBP-1, achieved through diet and exercise, may have protective effects against cancer cell development, depending on the type, intensity, and duration of training (93). The first exercise intervention study involving surgically treated LC patients who had not undergone adjuvant chemotherapy or radiotherapy, resulted in up-regulation of IGFBP-3. The exercise intervention included an 8-week training aerobic exercise combined with inspiratory and expiratory muscle training improved the response to the treatment in the human (38). In conclusion, the exercise contributes to the reprogramming of intermediate metabolism in cancer cells/tissue through the activation of AMPK, IGF-1 and IGF-1R resulting in a potential nonpharmacological adjuvant in the management and treatment of some types of cancer including LC. However, the molecular mechanisms are not completely elucidated until now.
This minireview addressed some topics on molecular effects mediated by exercise in LC highlighting the evidence obtained in this field, mostly in mouse LC models, pointing-out the existing gap in knowledge on the molecular effects of exercise in human LC, which still requires further investigations.
Overall, these evidences support the positive impact of the exercise in LC and underline the poorly understood molecular mechanisms elicited in human.
The most relevant results discussed are summarized in Figure 1.
The focus of this minireview is to provide an updated overview of the impact of exercise on molecular mechanisms in LC. Due to the limited molecular data on the effects of exercise in LC patients, we primarily reported and discussed on animal models. This reliance on animal models limits the direct applicability of the findings to humans. Human clinical trials are essential to confirm the therapeutic potential of exercise and to validate the translatability of preclinical results. Furthermore, the heterogeneity in exercise protocols, including different types, frequency, intensity, time, volume, and progression, complicates the development of specific guidelines for clinical application. Personalized exercise prescription, adapted to the different clinical stages and specific to the type of oncological pathology, is needed to optimize and replicate therapeutic outcomes across different patient populations. Future human studies should focus on detailed mechanistic investigations, which could help in the development of targeted exercise interventions.
AM: Writing – original draft. FO: Writing – original draft. DV: Writing – original draft. NL: Writing – original draft. AA: Writing – review & editing. SO: Writing – review & editing. GS: Writing – review & editing. PB: Writing – review & editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This study was supported by Finanziato dall’Unione europea - Next Generation EU, Missione 4 Componente 1 CUP I53D23007060001 Codice progetto P2022P8JRJ and in part by MUR, DM 737 of 25 June 2021.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Bray F, Laversanne M, Sung H, Ferlay J, Siegel RL, Soerjomataram I, et al. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. (2024) 74:229–63. doi: 10.3322/caac.21834
2. Leiter A, Veluswamy RR, Wisnivesky JP. The global burden of lung cancer: current status and future trends. Nat Rev Clin Oncol. (2023) 20:624–39. doi: 10.1038/s41571-023-00798-3
3. Wang Q, Gümüş ZH, Colarossi C, Memeo L, Wang X, Kong CY, et al. SCLC: epidemiology, risk factors, genetic susceptibility, molecular pathology, screening, and early detection. J Thorac Oncol. (2023) 18:31–46. doi: 10.1016/j.jtho.2022.10.002
4. Zou K, Sun P, Huang H, Zhuo H, Qie R, Xie Y, et al. Etiology of lung cancer: Evidence from epidemiologic studies. J Natl Cancer Cent. (2022) 2:216–25. doi: 10.1016/j.jncc.2022.09.004
5. Lv X, Mao Z, Sun X, Liu B. Intratumoral heterogeneity in lung cancer. Cancers (Basel). (2023) 15:2709. doi: 10.3390/cancers15102709
6. de Sousa VML, Carvalho L. Heterogeneity in lung cancer. Pathobiology. (2018) 85:96–107. doi: 10.1159/000487440
7. Affinito O, Orlandella FM, Luciano N, Salvatore M, Salvatore G, Franzese M. Evolution of intra-tumoral heterogeneity across different pathological stages in papillary thyroid carcinoma. Cancer Cell Int. (2022) 22:263. doi: 10.1186/s12935-022-02680-1
8. Megyesfalvi Z, Gay CM, Popper H, Pirker R, Ostoros G, Heeke S, et al. Clinical insights into small cell lung cancer: Tumor heterogeneity, diagnosis, therapy, and future directions. CA Cancer J Clin. (2023) 73:620–52. doi: 10.3322/caac.21785
9. Uramoto H, Tanaka F. Recurrence after surgery in patients with NSCLC. Transl Lung Cancer Res. (2014) 3:242–9. doi: 10.3978/j.issn.2218-6751.2013.12.05
10. Herbst RS, Morgensztern D, Boshoff C. The biology and management of non-small cell lung cancer. Nature. (2018) 553:446–54. doi: 10.1038/nature25183
11. Planchard D, Popat S, Kerr K, Novello S, Smit EF, Faivre-Finn C, et al. Metastatic non-small cell lung cancer: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann Oncol. (2018) 29:iv192–237. doi: 10.1093/annonc/mdy275
12. Sabari JK, Lok BH, Laird JH, Poirier JT, Rudin CM. Unravelling the biology of SCLC: implications for therapy. Nat Rev Clin Oncol. (2017) 14:549–61. doi: 10.1038/nrclinonc.2017.71
13. Araghi M, Mannani R, Heidarnejad Maleki A, Hamidi A, Rostami S, Safa SH, et al. Recent advances in non-small cell lung cancer targeted therapy; an update review. Cancer Cell Int. (2023) 23:162. doi: 10.1186/s12935-023-02990-y
14. Reck M, Remon J, Hellmann MD. First-line immunotherapy for non-small-cell lung cancer. J Clin Oncol. (2022) 40:586–97. doi: 10.1200/JCO.21.01497
15. Zhang S, Cheng Y. Immunotherapy for extensive-stage small-cell lung cancer: current landscape and future perspectives. Front Oncol. (2023) 13:1142081. doi: 10.3389/fonc.2023.1142081
16. Hojman P, Gehl J, Christensen JF, Pedersen BK. Molecular mechanisms linking exercise to cancer prevention and treatment. Cell Metab. (2018) 27:10–21. doi: 10.1016/j.cmet.2017.09.015
17. Orlandella FM, De Stefano AE, Braile M, Luciano N, Mancini A, Franzese M, et al. Unveiling the miRNAs responsive to physical activity/exercise training in cancer: A systematic review. Crit Rev Oncol Hematol. (2022) 180:103844. doi: 10.1016/j.critrevonc.2022.103844
18. Orlandella FM, De Stefano AE, Iervolino PLC, Buono P, Soricelli A, Salvatore G. Dissecting the molecular pathways involved in the effects of physical activity on breast cancers cells: A narrative review. Life Sci. (2021) 265:118790. doi: 10.1016/j.lfs.2020.118790
19. Avancini A, Belluomini L, Milella M, Schena F, Novello S, Pilotto S. Drive the oncologists into exercise promotion in lung cancer. Lung Cancer. (2023) 176:1–3. doi: 10.1016/j.lungcan.2022.12.009
20. Avancini A, Cavallo A, Trestini I, Tregnago D, Belluomini L, Crisafulli E, et al. Exercise prehabilitation in lung cancer: Getting stronger to recover faster. Eur J Surg Oncol. (2021) 47:1847–55. doi: 10.1016/j.ejso.2021.03.231
21. Ligibel JA, Bohlke K, May AM, Clinton SK, Demark-Wahnefried W, Gilchrist SC, et al. Exercise, diet, and weight management during cancer treatment: ASCO guideline. J Clin Oncol. (2022) 40:2491–507. doi: 10.1200/JCO.22.00687
22. Billé A, Buxton J, Viviano A, Gammon D, Veres L, Routledge T, et al. Preoperative physical activity predicts surgical outcomes following lung cancer resection. Integr Cancer Ther. (2021) 20:1534735420975853. doi: 10.1177/1534735420975853
23. Steffens D, Beckenkamp PR, Hancock M, Solomon M, Young J. Preoperative exercise halves the postoperative complication rate in patients with lung cancer: a systematic review of the effect of exercise on complications, length of stay and quality of life in patients with cancer. Br J Sports Med. (2018) 52:344. doi: 10.1136/bjsports-2017-098032
24. Avancini A, Sartori G, Gkountakos A, Casali M, Trestini I, Tregnago D, et al. Physical activity and exercise in lung cancer care: will promises be fulfilled? Oncologist. (2020) 25:e555–69. doi: 10.1634/theoncologist.2019-0463
25. Toohey K, Mizrahi D, Hart NH, Singh B, Lopez P, Hunter M, et al. Exercise in cancer care for people with lung cancer: A narrative synthesis. J Sci Med Sport. (2024). doi: 10.1016/j.jsams.2024.08.002
26. Luo Z, Wan R, Liu S, Feng X, Peng Z, Wang Q, et al. Mechanisms of exercise in the treatment of lung cancer - a mini-review. Front Immunol. (2023) 14:1244764. doi: 10.3389/fimmu.2023.1244764
27. Messina G, Tartaglia N, Ambrosi A, Porro C, Campanozzi A, Valenzano A, et al. The beneficial effects of physical activity in lung cancer prevention and/or treatment. Life (Basel). (2022) 12:782. doi: 10.3390/life12060782
28. Alves CRR, das Neves W, Tobias GC, de Almeida NR, Barreto RF, Melo CM, et al. High-intensity interval training slows down tumor progression in mice bearing Lewis lung carcinoma. JCSM Rapid Commun. (2018) 1:1–10. doi: 10.1002/j.2617-1619.2018.tb00008.x
29. Ge Z, Wu S, Qi Z, Ding S. Exercise modulates polarization of TAMs and expression of related immune checkpoints in mice with lung cancer. J Cancer. (2022) 13:3297–307. doi: 10.7150/jca.76136
30. Higgins KA, Park D, Lee GY, Curran WJ, Deng X. Exercise-induced lung cancer regression: mechanistic findings from a mouse model. Cancer. (2014) 120:3302–10. doi: 10.1002/cncr.v120.21
31. Jo S, Jeon J, Park G, Do HK, Kang J, Ahn KJ, et al. Aerobic exercise improves radiation therapy efficacy in non-small cell lung cancer: preclinical study using a xenograft mouse model. Int J Mol Sci. (2024) 25:2757. doi: 10.3390/ijms25052757
32. Leimbacher AC, Villiger P, Desboeufs N, Aboouf MA, Nanni M, Armbruster J, et al. Voluntary exercise does not always suppress lung cancer progression. iScience. (2023) 26:107298. doi: 10.1016/j.isci.2023.107298
33. Luo ZW, Sun YY, Xia W, Xu JY, Xie DJ, Jiao CM, et al. Physical exercise reverses immuno-cold tumor microenvironment via inhibiting SQLE in non-small cell lung cancer. Mil Med Res. (2023) 10:39. doi: 10.1186/s40779-023-00474-8
34. Martín-Ruiz A, Fiuza-Luces C, Rincón-Castanedo C, Fernández-Moreno D, Gálvez BG, Martínez-Martínez E, et al. Benefits of exercise and immunotherapy in a murine model of human non-small-cell lung carcinoma. Exerc Immunol Rev. (2020) 26:100–15.
35. Tobias GC, Gomes JLP, Fernandes LG, Voltarelli VA, de Almeida NR, Jannig PR, et al. Aerobic exercise training mitigates tumor growth and cancer-induced splenomegaly through modulation of non-platelet platelet factor 4 expression. Sci Rep. (2023) 13:21970. doi: 10.1038/s41598-023-47217-2
36. Kurgan N, Tsakiridis E, Kouvelioti R, Moore J, Klentrou P, Tsiani E. Inhibition of Human Lung Cancer Cell Proliferation and Survival by Post-Exercise Serum Is Associated with the Inhibition of Akt, mTOR, p70 S6K, and Erk1/2. Cancers (Basel). (2017) 9:46. doi: 10.3390/cancers9050046
37. Liu Y, He L, Wang W. Systematic assessment of microRNAs associated with lung cancer and physical exercise. Front Oncol. (2022) 12:917667. doi: 10.3389/fonc.2022.917667
38. Messaggi-Sartor M, Marco E, Martínez-Téllez E, Rodriguez-Fuster A, Palomares C, Chiarella S, et al. Combined aerobic exercise and high-intensity respiratory muscle training in patients surgically treated for non-small cell lung cancer: a pilot randomized clinical trial. Eur J Phys Rehabil Med. (2019) 55:113–22. doi: 10.23736/S1973-9087.18.05156-0
39. Anderson NM, Simon MC. The tumor microenvironment. Curr Biol. (2020) 30:R921–5. doi: 10.1016/j.cub.2020.06.081
40. Gomes-Santos IL, Amoozgar Z, Kumar AS, Ho WW, Roh K, Talele NP, et al. Exercise training improves tumor control by increasing CD8+ T-cell infiltration via CXCR3 signaling and sensitizes breast cancer to immune checkpoint blockade. Cancer Immunol Res. (2021) 9:765–78. doi: 10.1158/2326-6066.CIR-20-0499
41. Kurz E, Hirsch CA, Dalton T, Shadaloey SA, Khodadadi-Jamayran A, Miller G, et al. Exercise-induced engagement of the IL-15/IL-15Rα axis promotes anti-tumor immunity in pancreatic cancer. Cancer Cell. (2022) 40:720–37.e5. doi: 10.1016/j.ccell.2022.05.006
42. Savage H, Pareek S, Lee J, Ballarò R, Conterno Minussi D, Hayek K, et al. Aerobic exercise alters the melanoma microenvironment and modulates ERK5 S496 phosphorylation. Cancer Immunol Res. (2023) 11:1168–83. doi: 10.1158/2326-6066.CIR-22-0465
43. Özdemir BH, Özdemir AA. How exercise affects the development and progression of hepatocellular carcinoma by changing the biomolecular status of the tumor microenvironment. Exp Clin Transplant. (2022). doi: 10.6002/ect.2021.0456
44. Zheng A, Zhang L, Yang J, Yin X, Zhang T, Wu X, et al. Physical activity prevents tumor metastasis through modulation of immune function. Front Pharmacol. (2022) 13:1034129. doi: 10.3389/fphar.2022.1034129
45. MacInnis MJ, Gibala MJ. Physiological adaptations to interval training and the role of exercise intensity. J Physiol. (2017) 595:2915–30. doi: 10.1113/tjp.2017.595.issue-9
46. Semenza GL. Targeting intratumoral hypoxia to enhance anti-tumor immunity. Semin Cancer Biol. (2023) 96:5–10. doi: 10.1016/j.semcancer.2023.09.002
47. Elming PB, Busk M, Wittenborn TR, Bussink J, Horsman MR, Lønbro S. The effect of single bout and prolonged aerobic exercise on tumor hypoxia in mice. J Appl Physiol (1985). (2023) 134:692–702. doi: 10.1152/japplphysiol.00561.2022
48. Chen S, Sigdel S, Sawant H, Bihl J, Wang J. Exercise-intervened endothelial progenitor cell exosomes protect N2a cells by improving mitochondrial function. Int J Mol Sci. (2024) 25:1148. doi: 10.3390/ijms25021148
49. Betof AS, Lascola CD, Weitzel D, Landon C, Scarbrough PM, Devi GR, et al. Modulation of murine breast tumor vascularity, hypoxia and chemotherapeutic response by exercise. J Natl Cancer Inst. (2015) 107:djv040. doi: 10.1093/jnci/djv040
50. McCullough DJ, Stabley JN, Siemann DW, Behnke BJ. Modulation of blood flow, hypoxia, and vascular function in orthotopic prostate tumors during exercise. J Natl Cancer Inst. (2014) 106:dju036. doi: 10.1093/jnci/dju036
51. Xiao CL, Zhong ZP, Lü C, Guo BJ, Chen JJ, Zhao T, et al. Physical exercise suppresses hepatocellular carcinoma progression by alleviating hypoxia and attenuating cancer stemness through the Akt/GSK-3β/β-catenin pathway. J Integr Med. (2023) 21:184–93. doi: 10.1016/j.joim.2023.01.002
52. Kwak SE, Lee JH, Zhang D, Song W. Angiogenesis: focusing on the effects of exercise in aging and cancer. J Exerc Nutr Biochem. (2018) 22:21–6. doi: 10.20463/jenb.2018.0020
53. Dimauro I, Paronetto MP, Caporossi D. Exercise, redox homeostasis and the epigenetic landscape. Redox Biol. (2020) 35:101477. doi: 10.1016/j.redox.2020.101477
54. Ingerslev LR, Donkin I, Fabre O, Versteyhe S, Mechta M, Pattamaprapanont P, et al. Endurance training remodels sperm-borne small RNA expression and methylation at neurological gene hotspots. Clin Epigenetics. (2018) 10:12. doi: 10.1186/s13148-018-0446-7
55. Davidsen PK, Gallagher IJ, Hartman JW, Tarnopolsky MA, Dela F, Helge JW, et al. High responders to resistance exercise training demonstrate differential regulation of skeletal muscle microRNA expression. J Appl Physiol (1985). (2011) 110:309–17. doi: 10.1152/japplphysiol.00901.2010
56. Dufresne S, Rébillard A, Muti P, Friedenreich CM, Brenner DR. A review of physical activity and circulating miRNA expression: implications in cancer risk and progression. Cancer Epidemiol Biomarkers Prev. (2018) 27:11–24. doi: 10.1158/1055-9965.EPI-16-0969
57. Xu T, Liu Q, Yao J, Dai Y, Wang H, Xiao J. Circulating microRNAs in response to exercise. Scand J Med Sci Sports. (2015) 25:e149–154. doi: 10.1111/sms.2015.25.issue-2
58. Mancini A, Vitucci D, Orlandella FM, Terracciano A, Mariniello RM, Imperlini E, et al. Regular football training down-regulates miR-1303 muscle expression in veterans. Eur J Appl Physiol. (2021) 121:2903–12. doi: 10.1007/s00421-021-04733-1
59. Chavez-Dominguez R, Perez-Medina M, Lopez-Gonzalez JS, Galicia-Velasco M, Aguilar-Cazares D. The double-edge sword of autophagy in cancer: from tumor suppression to pro-tumor activity. Front Oncol. (2020) 10:578418. doi: 10.3389/fonc.2020.578418
60. Galluzzi L, Vitale I, Aaronson SA, Abrams JM, Adam D, Agostinis P, et al. Molecular mechanisms of cell death: recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ. (2018) 25:486–541. doi: 10.1038/s41418-017-0012-4
61. Lopez Guerra JL, Gomez DR, Zhuang Y, Levy LB, Eapen G, Liu H, et al. Changes in pulmonary function after three-dimensional conformal radiotherapy, intensity-modulated radiotherapy, or proton beam therapy for non-small-cell lung cancer. Int J Radiat Oncol Biol Phys. (2012) 83:e537–543. doi: 10.1016/j.ijrobp.2012.01.019
62. Rivera MP, Detterbeck FC, Socinski MA, Moore DT, Edelman MJ, Jahan TM, et al. Impact of preoperative chemotherapy on pulmonary function tests in resectable early-stage non-small cell lung cancer. Chest. (2009) 135:1588–95. doi: 10.1378/chest.08-1430
63. Rundqvist H, Augsten M, Strömberg A, Rullman E, Mijwel S, Kharaziha P, et al. Effect of acute exercise on prostate cancer cell growth. PloS One. (2013) 8:e67579. doi: 10.1371/journal.pone.0067579
64. Dethlefsen C, Lillelund C, Midtgaard J, Andersen C, Pedersen BK, Christensen JF, et al. Exercise regulates breast cancer cell viability: systemic training adaptations versus acute exercise responses. Breast Cancer Res Treat. (2016) 159:469–79. doi: 10.1007/s10549-016-3970-1
65. Jin S, Levine AJ. The p53 functional circuit. J Cell Sci. (2001) 114:4139–40. doi: 10.1242/jcs.114.23.4139
66. Vogelstein B, Lane D, Levine AJ. Surfing the p53 network. Nature. (2000) 408:307–10. doi: 10.1038/35042675
67. Su M, Mei Y, Sinha S. Role of the crosstalk between autophagy and apoptosis in cancer. J Oncol. (2013) 2013:102735. doi: 10.1155/2013/102735
68. Pietrocola F, Izzo V, Niso-Santano M, Vacchelli E, Galluzzi L, Maiuri MC, et al. Regulation of autophagy by stress-responsive transcription factors. Semin Cancer Biol. (2013) 23:310–22. doi: 10.1016/j.semcancer.2013.05.008
69. Rubinstein AD, Eisenstein M, Ber Y, Bialik S, Kimchi A. The autophagy protein Atg12 associates with antiapoptotic Bcl-2 family members to promote mitochondrial apoptosis. Mol Cell. (2011) 44:698–709. doi: 10.1016/j.molcel.2011.10.014
70. Gunadi JW, Welliangan AS, Soetadji RS, Jasaputra DK, Lesmana R. The role of autophagy modulated by exercise in cancer cachexia. Life (Basel). (2021) 11:781. doi: 10.3390/life11080781
71. Teunissen SCCM, Wesker W, Kruitwagen C, de Haes HCJM, Voest EE, de Graeff A. Symptom prevalence in patients with incurable cancer: a systematic review. J Pain Symptom Manage. (2007) 34:94–104. doi: 10.1016/j.jpainsymman.2006.10.015
72. Møller AB, Lønbro S, Farup J, Voss TS, Rittig N, Wang J, et al. Molecular and cellular adaptations to exercise training in skeletal muscle from cancer patients treated with chemotherapy. J Cancer Res Clin Oncol. (2019) 145:1449–60. doi: 10.1007/s00432-019-02911-5
73. Fernandes LG, Tobias GC, Paixão AO, Dourado PM, Voltarelli VA, Brum PC. Exercise training delays cardiac remodeling in a mouse model of cancer cachexia. Life Sci. (2020) 260:118392. doi: 10.1016/j.lfs.2020.118392
74. Parry TL, Hayward R. Exercise protects against cancer-induced cardiac cachexia. Med Sci Sports Exerc. (2018) 50:1169–76. doi: 10.1249/MSS.0000000000001544
75. Canbolat E, Cakıroglu FP. The importance of AMPK in obesity and chronic diseases and the relationship of AMPK with nutrition: a literature review. Crit Rev Food Sci Nutr. (2023) 63:449–56. doi: 10.1080/10408398.2022.2087595
76. Weir HJ, Yao P, Huynh FK, Escoubas CC, Goncalves RL, Burkewitz K, et al. Dietary restriction and AMPK increase lifespan via mitochondrial network and peroxisome remodeling. Cell Metab. (2017) 26:884–96.e5. doi: 10.1016/j.cmet.2017.09.024
77. Richter EA, Ruderman NB. AMPK and the biochemistry of exercise: implications for human health and disease. Biochem J. (2009) 418:261–75. doi: 10.1042/BJ20082055
78. Hsu CC, Peng D, Cai Z, Lin HK. AMPK signaling and its targeting in cancer progression and treatment. Semin Cancer Biol. (2022) 85:52–68. doi: 10.1016/j.semcancer.2021.04.006
79. Ashrafizadeh M, Mirzaei S, Hushmandi K, Rahmanian V, Zabolian A, Raei M, et al. Therapeutic potential of AMPK signaling targeting in lung cancer: Advances, challenges and future prospects. Life Sci. (2021) 278:119649. doi: 10.1016/j.lfs.2021.119649
80. Campos JC, Marchesi Bozi LH, Krum B, Grassmann Bechara LR, Ferreira ND, Arini GS, et al. Exercise preserves physical fitness during aging through AMPK and mitochondrial dynamics. Proc Natl Acad Sci U.S.A. (2023) 120:e2204750120. doi: 10.1073/pnas.2204750120
81. Mancini A, Vitucci D, Labruna G, Imperlini E, Randers MB, Schmidt JF, et al. Effect of lifelong football training on the expression of muscle molecular markers involved in healthy longevity. Eur J Appl Physiol. (2017) 117:721–30. doi: 10.1007/s00421-017-3562-8
82. Spaulding HR, Yan Z. AMPK and the adaptation to exercise. Annu Rev Physiol. (2022) 84:209–27. doi: 10.1146/annurev-physiol-060721-095517
83. Townsend LK, Steinberg GR. AMPK and the endocrine control of metabolism. Endocr Rev. (2023) 44:910–33. doi: 10.1210/endrev/bnad012
84. Viollet B. The energy sensor AMPK: adaptations to exercise, nutritional and hormonal signals. In: Spiegelman B, editor. Hormones, Metabolism and the Benefits of Exercise. Springer, Cham (CH (2017). Available at: http://www.ncbi.nlm.nih.gov/books/NBK543786/.
85. Raun SH, Ali MS, Han X, Henríquez-Olguín C, Pham TCP, Meneses-Valdés R, et al. Adenosine monophosphate-activated protein kinase is elevated in human cachectic muscle and prevents cancer-induced metabolic dysfunction in mice. J Cachexia Sarcopenia Muscle. (2023) 14:1631–47. doi: 10.1002/jcsm.13238
86. Ge Y, Zhou M, Chen C, Wu X, Wang X. Role of AMPK mediated pathways in autophagy and aging. Biochimie. (2022) 195:100–13. doi: 10.1016/j.biochi.2021.11.008
87. Zhong W, Wang X, Wang Y, Sun G, Zhang J, Li Z. Obesity and endocrine-related cancer: The important role of IGF-1. Front Endocrinol (Lausanne). (2023) 14:1093257. doi: 10.3389/fendo.2023.1093257
88. Stiles AD, Moats-Staats BM. Production and action of insulin-like growth factor I/somatomedin C in primary cultures of fetal lung fibroblasts. Am J Respir Cell Mol Biol. (1989) 1:21–6. doi: 10.1165/ajrcmb/1.1.21
89. Jiang S, Xu Z, Shi Y, Liang S, Jiang X, Xiao M, et al. Circulating insulin-like growth factor-1 and risk of lung diseases: A Mendelian randomization analysis. Front Endocrinol (Lausanne). (2023) 14:1126397. doi: 10.3389/fendo.2023.1126397
90. Minuto F, Del Monte P, Barreca A, Fortini P, Cariola G, Catrambone G, et al. Evidence for an increased somatomedin-C/insulin-like growth factor I content in primary human lung tumors. Cancer Res. (1986) 46:985–8.
91. Hu Q, Zhou Y, Ying K, Ruan W. IGFBP, a novel target of lung cancer? Clin Chim Acta. (2017) 466:172–7. doi: 10.1016/j.cca.2017.01.017
92. Long L, Rubin R, Brodt P. Enhanced invasion and liver colonization by lung carcinoma cells overexpressing the type 1 insulin-like growth factor receptor. Exp Cell Res. (1998) 238:116–21. doi: 10.1006/excr.1997.3814
Keywords: exercise, lung cancer, tumor microenvironment, angiogenesis, gene expression, apoptosis, intermediate metabolism
Citation: Mancini A, Orlandella FM, Vitucci D, Luciano N, Alfieri A, Orrù S, Salvatore G and Buono P (2024) Exercise’s impact on lung cancer molecular mechanisms: a current overview. Front. Oncol. 14:1479454. doi: 10.3389/fonc.2024.1479454
Received: 12 August 2024; Accepted: 16 October 2024;
Published: 01 November 2024.
Edited by:
Sara Pilotto, University of Verona, ItalyReviewed by:
Valentina Natalucci, University of Urbino Carlo Bo, ItalyCopyright © 2024 Mancini, Orlandella, Vitucci, Luciano, Alfieri, Orrù, Salvatore and Buono. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Pasqualina Buono, YnVvbm9AdW5pcGFydGhlbm9wZS5pdA==; Giuliana Salvatore, Z2l1bGlhbmEuc2FsdmF0b3JlQHVuaXBhcnRoZW5vcGUuaXQ=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.