 Sulayne Janayna Araujo Guimarães1,2†André Alvares Marques Vale1,2†
Sulayne Janayna Araujo Guimarães1,2†André Alvares Marques Vale1,2† Mirtes Castelo Branco Rocha2
Mirtes Castelo Branco Rocha2 Ana Luiza de Araújo Butarelli1,2
Ana Luiza de Araújo Butarelli1,2 Jenilson Mota da Silva1,3Amanda Jordão Silva de Deus1,3Leudivan Nogueira4Ronald Wagner Pereira Coelho4
Jenilson Mota da Silva1,3Amanda Jordão Silva de Deus1,3Leudivan Nogueira4Ronald Wagner Pereira Coelho4 Silma Regina Pereira3
Silma Regina Pereira3 Ana Paula Silva Azevedo-Santos2*
Ana Paula Silva Azevedo-Santos2*- 1Postgraduate Program in Health Science, Federal University of Maranhão, São Luís, Brazil
- 2Laboratory of Immunology Applied to Cancer, Department of Physiological Sciences, Biological and Health Sciences Center, Federal University of Maranhão, São Luís, MA, Brazil
- 3Laboratory of Genetics and Molecular Biology, Department of Biology, Biological and Health Sciences Center, Federal University of Maranhão, São Luís, MA, Brazil
- 4Maranhense Institute of Oncology Aldenora Bello, São Luís, Brazil
Penile squamous cell carcinoma (PSCC) is a largely neglected condition, predominantly affecting underdeveloped regions, and is associated with risk factors such as low socioeconomic status, phimosis, and human papillomavirus (HPV) infection. Unlike other urogenital cancers, its pathophysiology and therapeutic targets remain poorly understood, particularly regarding the immune response to the tumor microenvironment. This study aims to investigate immune cell infiltration profiles, dendritic cell maturation, and lymphocyte apoptosis in both HPV-positive and HPV-negative PSCC. Clinical and histopathological data, along with peripheral blood and tumor tissue samples, were collected from 30 patients (66.6% were HPV-positive and 33.3% HPV-negative), with an additional 19 healthy donors serving as controls. Tumor-infiltrating immune cells were analyzed following enzymatic digestion of tumor tissue, enabling detailed phenotypic characterization. A simulated tumor microenvironment was created using supernatants derived from primary cultures of HPV-positive PSCC tumors. Peripheral blood mononuclear cells were isolated and differentiated into dendritic cells (Mo-DCs) for further phenotyping and lymphoproliferation assays. Lymphocytes from healthy donors and patients were exposed to tumor culture supernatants to evaluate apoptosis induced by the tumor microenvironment. Results showed that HPV-positive tumors exhibited lower T lymphocyte frequencies compared to HPV-negative tumors. Additionally, patients infected with high-risk HPV demonstrated reduced maturation rates of Mo-DCs and decreased expression of co-stimulatory molecules on these cells compared to healthy donors. Furthermore, Mo-DCs from hrHPV-positive patients showed impaired lymphoproliferation capacity relative to controls, while HPV-negative patients exhibited a trend towards reduced lymphoproliferative ability. Regarding the simulated tumor microenvironment, lymphocytes from healthy donors underwent apoptosis, contrasting with patients' lymphocytes, which showed increased viability when cultured with tumor supernatants. These results underscore the impact of HPV infection on T lymphocyte infiltration, Mo-DC maturation, and lymphocyte survival in PSCC, offering critical insights for advancing our understanding of the tumor microenvironment and guiding the development of immunotherapy strategies.
1 Introduction
Penile squamous cell carcinoma (PSCC) is a rare disease in developed countries, however, it is more common in underdeveloped and developing countries (1). The Brazilian northeast region has the highest incidence of penile cancer (6,1/100,000 inhabitants) (2). Clinically, these patients are frequently characterized by late diagnosis in stage T2, grade II or III, with detection of HPV-DNA and usually undergo penectomy (2, 3). Therapy is complex and multi-layered. As it is a multifactorial disease, risk factors include phimosis, chronic inflammation, poor penile hygiene, smoking, immunosuppression, and human papillomavirus infection (1), in addition to the conditions of poverty and low access to health services, delaying diagnosis (2).
The mechanisms of carcinogenesis in PSCC have been described as HPV-dependent, through the integration of viral DNA into the host cell genome causing the overexpression of viral oncoproteins E6 and E7, host cell cycle deregulation and genomic instability of the host (3–7). Another mechanism is non-HPV related, defined by chronic inflammation and somatic genetic changes by reactive oxygen and reactive nitrogen intermediates associated with phimosis and lichen sclerosus (8–10). In both conditions, the immune system plays a fundamental role in the establishment and maintenance of the tumor microenvironment.
The dynamic interaction of neoplastic, immune, stromal, and vascular cells determines the survival of malignant cells in tissues and organs (11, 12). Chu et al. (13) reported immunophenotype similarities between PSCC HPV-positive and HPV-negative patients. However, the HPV-positive tumors showed higher density of intratumoral PD-1 (13). Furthermore, HPV infection reduces inflammation-associated signaling by decreasing the destruction of infected cells (14) and promoting the production of viral proteins E6 and E7 from high-risk HPV strains. These proteins inhibit key immune responses, suppressing interferon signaling pathways (15) and E7 downregulating TLR9 expression in phagocytes (16). This suppression impairs the mechanisms necessary for effective immune recognition and response, allowing the virus to evade the host’s immune system.
The HPV infection and the host immune response are integral components of the tumor microenvironment in PSCC and the Dendritic Cells (DCs) can be an interesting target as they orchestrate the immune response, serving as the primary antigen-presenting cells that determine lymphocyte activation and the profile of the adaptive immune response, whether effector or tolerogenic (17). So far, the studies on immune aspects associated with penile cancer have been based on theoretical approaches. This work proposes an experimental study using immune cells from penile cancer patients, evaluating the impact of HPV on the process of antigen activation and presentation, identifying mechanisms of immune evasion, and targets for immunotherapeutic strategies.
2 Materials and methods
2.1 Patient and sample collection
Patients’ blood and PSCC tissue samples were collected at Aldenora Bello Cancer Hospital, São Luis-Brazil, from 2017 to 2020. This study was approved by the Research Ethics Committee on Humans from the Federal University of Maranhão and by the National Research Ethics Commission (CONEP‐ Brazil, CAAE: 46371515.5.0000.5087).
2.2 Study design
This study employed experimental research techniques utilizing both in vitro and ex vivo methodologies. Peripheral blood was collected from patients (n=30) and healthy donor volunteers (n=19). The tumor tissue from patients was enzymatically digested to characterize the immune infiltrating cells and to establish ex vivo cultures. HPV genotyping was performed using nested-PCR, following the protocol provided by BioGenetics Molecular Technologies (Uberlândia, Minas Gerais, Brazil; patent number BR102017004615.0), to identify viral subtypes classified as high risk (16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 59, 66, 68, 73, and 82), intermediate risk (MM7, MM8, 26, 30, 34, 53, 54, 55, 61, 62, 64, 67, 69, 70, 71, 72, 74, and 81), and low risk (6, 11, 42, 43, 44, and 57). In addition, we performed DNA sequencing according to our previous protocol (3). The tumor cell culture supernatants were used as tumor microenvironment-like to perform the apoptosis lymphocyte assay. From the peripheral blood mononuclear cells (PBMCs), the monocytes were isolated for in vitro differentiation in mature Dendritic Cells (Mo-DCs) and the lymphocytes were used in proliferation and apoptosis assay. Mo-DCs activation was evaluated through the expression of costimulatory molecules and compared with patients’ groups and healthy donors (Supplementary Figure 1).
2.3 Analysis of tumor-infiltrating immune cells
Tumor tissues fragments were treated with Collagenase IV (0.056 mg/mL of RPMI 1640 medium) according to the manufacturer (Sigma-Aldrich, Saint Louis,USA) at 37°C for 2 hours (18) After enzymatic digestion, the cells were stained with 0.04% Trypan Blue dye and counted using a Neubauer chamber. Phenotypic analysis by flow cytometer was performed on 2x105 tumor cells, as described below. Additionally, 1x104 tumor cells were cultured in 12-well plates for 5 days. After 5 days, the supernatant was removed from the plate and centrifuged to remove non-adherent cells that remain in suspension in the medium. For the apoptosis assay, the tumor supernatant was added to the lymphocyte culture at a concentration of 10%. This assay was conducted over three days in 96-wells plates containing 100 µL of supplemented medium.
2.4 In vitro differentiation of Mo-DCs
Lymphocyte activation depends on the antigen presentation mechanism by Antigen-Presenting Cells (APCs), mainly represented by DCs. Thus, monocytes obtained from the peripheral blood of patients with HPV-positive and HPV-negative PSCCa were differentiated in vitro into dendritic cells. Buffy coat of peripheral blood (5-10mL) samples from patients and healthy blood donors were used for the isolation of peripheral blood mononuclear cells (PBMCs) and plasm. Dendritic Cells from both groups were produced by standard procedures (18). Briefly, PBMCs were isolated from peripheral blood buffy coats using Ficoll density gradient. After washing with phosphate buffered saline, 1x106 cells/mL were plated and kept in culture with cell culture medium (RPMI 1640, Gibco, New York, USA) supplemented with 10% of fetal bovine serum (FBS), 1% of Antibiotic-antimycotic (Gibco, New York, USA) overnight, to separate adherent cells (monocytes) from non-adherent cells (lymphocytes). The lymphocytes were separated for future proliferation and apoptosis assays. The monocytes were seeded in 12-well plates and cultured in RPMI 1640 medium with 10% FBS added 50 ng/mL−1 recombinant human GM-CSF (PeproTech, Cranbury, USA), and 50 ng/mL−1 IL-4 (PeproTech, Cranbury, USA) to generate immature DCs. On the fifth day of culture, 20 ng/mL−1 TNF-α (PeproTech, Cranbury, USA) was added to the culture to generate mature monocyte-derived dendritic cells (Mo-DCs). After 7 days of culture, the cells were harvested, washed once with PBS, and used for the experiments.
2.5 Immune cells phenotype
A total of 1x105 cells were incubated, at room temperature, for 30 min, with PBS and antibody: CD14 (BD Biosciences Cat# 555397, RRID: AB_395798), CD86 (BioLegend Cat# 305422, RRID: AB_2074981), HLA-DR (BD Biosciences Cat# 339194, RRID: AB_647443), CD3 (BioLegend Cat# 300312, RRID: AB_314048), CD8 (BioLegend Cat# 300906, RRID: AB_314110), CD4 (BD Biosciences Cat# 555347, RRID: AB_395752), CD19 (BioLegend Cat# 302210, RRID: AB_314240), and CD56 (BD Biosciences Cat# 555516, RRID: AB_395906) (Supplementary Table 1). Following incubation, cells were washed with PBS, and analyzed using an InCyte Guava flow cytometer (Luminex Corporation). Data analysis was performed using FlowJo software (v.10; RRID: SCR_008520).
2.6 Allogeneic lymphocytes proliferation index
To assess the ability of Mo-DCs to induce lymphocyte activation, the allogeneic lymphocytes obtained from healthy donors were labeled with 2.5 μM carboxyfluorescein succinimidyl ester (CFSE) (Sigma-Aldrich,Saint Louis, USA) and co-cultured with patients’ or healthy donors’ Mo-DCs for 3 days. Fluorescent signals were analyzed using a flow cytometer, following the manufacturer’s instructions. A minimum of 10,000 events were acquired using a FACSCalibur flow cytometer and analyzed with cytometer software (FlowJo Software, v.10; RRID: SCR_008520).
2.7 Analysis of lymphocyte apoptosis
To simulate the effect of the tumor microenvironment on lymphocyte viability, the ex vivo culture supernatant from the tumor tissue was used as treatment. The healthy donor and patient’s lymphocytes were incubated for three days in a 96-well plate containing 100 µL of RPMI medium supplemented with 10% tumor supernatant. Fluorescence-conjugated Annexin V and propidium iodide were utilized for apoptosis analysis (BD Pharmingen, Franklin Lakes, USA) following the manufacturer’s instructions. After the incubation, the lymphocytes isolated from patients and healthy donors were washed and incubated in Annexin V binding buffer at a 1:20 dilution for 15 minutes at room temperature. Subsequently, the cells were then washed, resuspended in a fresh Annexin V binding buffer, and promptly analyzed by flow cytometry, according to the manufacturer’s guidelines.
2.8 Statistical analysis
Statistical analyses were performed using GraphPad Prism software (v10). Data are presented as the mean ± standard error of the mean (SEM). The Shapiro-Wilk test was used to assess the normality of data distribution. Depending on the distribution (normal or non-normal), appropriate parametric or non-parametric tests were applied. For comparisons between two groups, the parametric t-test or the non-parametric Mann-Whitney U test (both two-tailed and unpaired) were used. For comparisons involving three or more groups, either the parametric one-way ANOVA followed by Tukey’s multiple comparison post hoc test or the non-parametric Kruskal-Wallis test followed by Dunn’s multiple comparison post hoc test was employed. A p-value < 0.05 was considered statistically significant, with significance levels indicated as *p < 0.05, **p < 0.005, and ***p < 0.0005. The sample size for each test and the specific test used in each analysis are detailed in the figure legends.
3 Results
3.1 Penile cancer patients profile
Most of the patients enrolled in the study have low levels of education. The main occupation was agriculture, and the majority were married. The average age of the patients was 60.18 (26-94 year) years old, and reported smoking and drinking habits (50% and 53.3%, respectively). Phimosis was present in 40%, but this information was not available for 40% of the patients. The most frequent site of the tumor was the glans (46.6%) or the glans associated with other areas (39.9%).
All tumors were squamous cell carcinoma with 56.5% HPV-related (Warty carcinoma: 33.3%; Basaloid: 6.6%; Warty-basaloid: 16.6), 26.6% non-HPV related, and 16.6% had no information. Histological grades I (16.6%) and II (16.6%) were the most frequent, however, this information was not available for 40% of tumors. The size of the tumor for most patients (70%) was 0.7 to 5.4 cm, and the tumors were defined as pT1 (13.3%), pT2 (20%), pT3 (23.3%), and pT4 (3.3%). However, 36.6% of this information was not available in the medical records. All tumor samples were tested for HPV DNA detection using nested-PCR, with 66.6% testing positive. Among the HPV-positive samples used in immunological assays, HPV16 was the most common genotype, present in 66.7% of cases, including one case of co-infection with HPV6. Additionally, 33.3% of the samples were positive for HPV18, defined as high-risk oncogenic HPV (hrHPV). Table 1 provides an overview of the sociodemographic, clinical, and pathological characteristics of the patients.
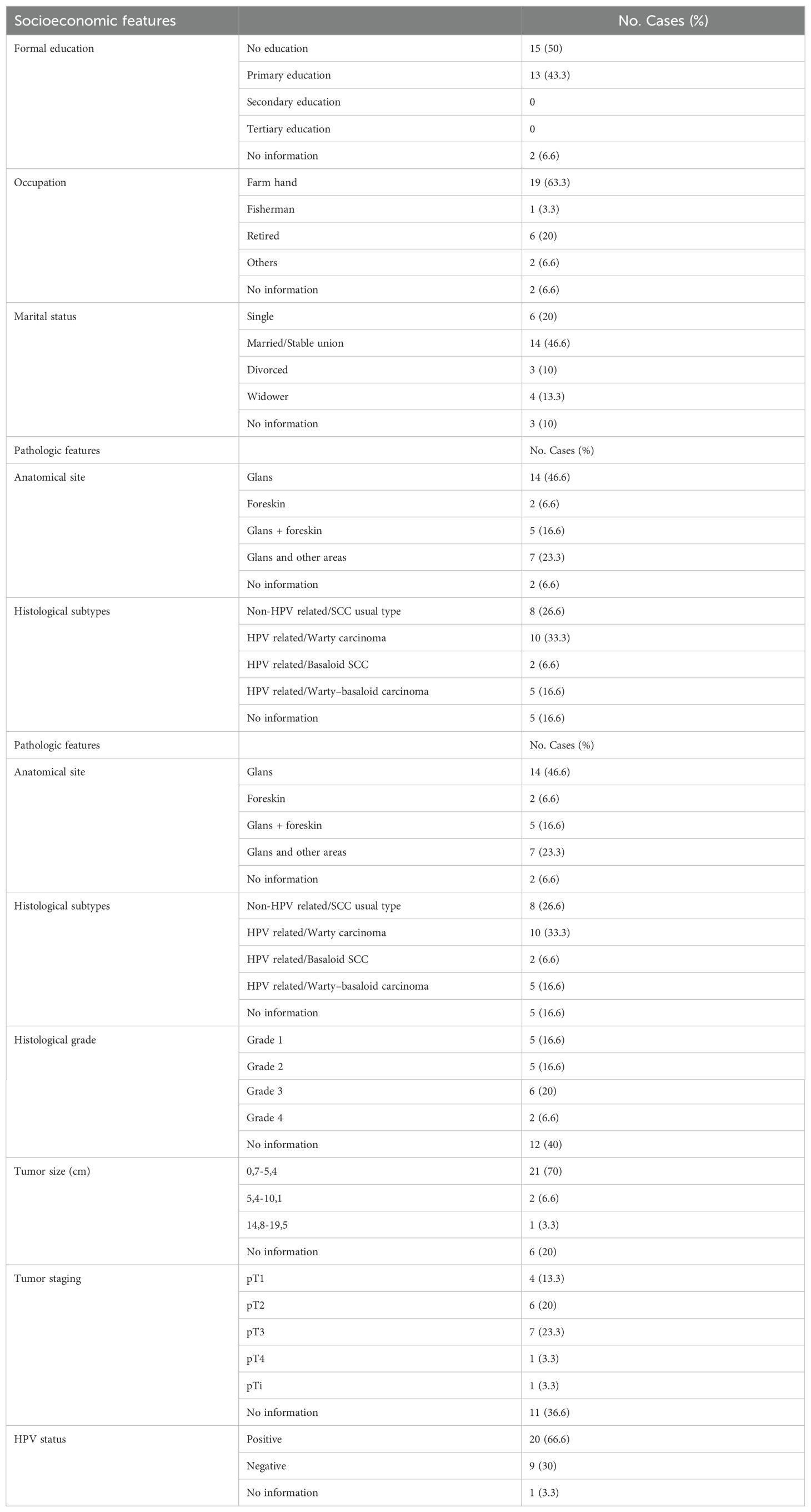
Table 1. Penile cancer socioeconomic, clinical, and pathologic profile.
3.2 Immune-infiltrated cells in hrHPV-positive and HPV-negative PSCC
The expression of specific markers CD3+ for T lymphocytes, CD19+ for B lymphocytes, CD56+ for NK cells, and CD14+ for monocytes/macrophages was used to assess immune cell infiltration within the tumor. The results revealed that the frequency of T lymphocytes was significantly lower in hrHPV-positive (37.96 ± 6.78) compared to HPV-negative PSCC (63.89 ± 5.12, p=0.0104). An inverse trend was observed in the frequency of NK cells, with a higher frequency in hrHPV-positive (3.44 ± 2.26) compared to HPV-negative cases (1.27 ± 0.54, p=0.2698). No significant differences (p≥0.05) were observed in the average frequencies of B lymphocytes and monocytes/macrophages between the two groups (Figure 1).
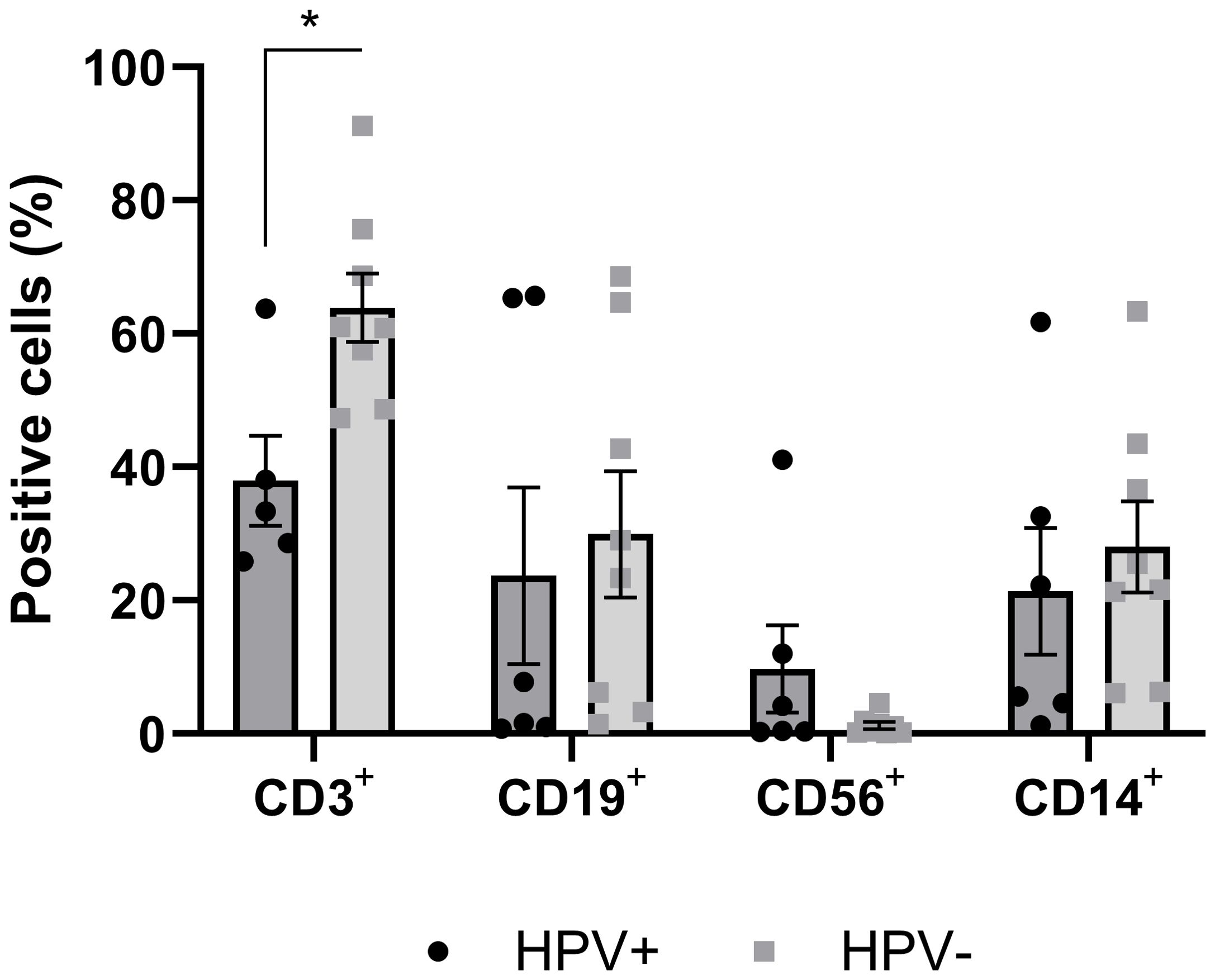
Figure 1. Frequency of cells positive for immune cell markers infiltrated in tumor tissue of hrHPV-positive and HPV-negative PSCC patients. Tumor tissue samples were enzymatically digested and labeled with surface markers CD3+, CD19+, CD56+, and CD14+, to identify T lymphocytes, B lymphocytes, Natural Killer (NK) cells, and Monocytes/Macrophages, respectively. Data were presented as mean ± standard error of the mean (SEM), and an unpaired t-test was performed for each marker, with statistical significance set at *p<0.05, comparing HPV-positive (n=8) and HPV-negative (n=8) carcinoma samples.
3.3 Dendritic cells phenotype and function associated to hrHPV-positive and HPV-negative PSCC
Comparing the frequency of Mo-DCs positive for CD86 between patients and healthy donors, it was observed that there was a significant reduction in cells obtained from hrHPV-positive patients (13.10 ± 3.03; healthy donors: 69.81 ± 9.10, p=0.0025). However, despite a trend towards reduction, the frequency was not significant in HPV-negative patients (49.36 ± 13.92, p= 0.3180). As for the frequency of Mo-DCs expressing co-stimulatory molecules, the results showed that hrHPV-positive patients have a lower proportion of positive cells compared to cells obtained from HPV-negative patients, but without significance (Figure 2A). The intensity of CD86 molecule expression on the membrane of Mo-DCs was lower in PSCC patients, both hrHPV-positive (101.8 ± 22.30; p=0.0293) and HPV-negative (145.2 ± 44.68; p=0.0490), compared to healthy donors (415.0 ± 103.4) (Figure 2B). The ability of Mo-DCs to induce lymphocyte proliferation was lower in cells from hrHPV-positive patients compared to healthy donors (Proliferation Index respectively: 1.33 ± 0.07; 2.08 ± 0.22, p=0.0275). No difference was observed in the proliferation index between HPV-negative patients and healthy donors. There was also no significant difference in the ability to induce lymphoproliferation between patient groups; however, the data showed a trend towards reduction in cells obtained from hrHPV-positive patients compared to HPV-negative patients (Proliferation Index respectively: 1.33 ± 0.07; 1.80 ± 0.21, p=0.1668) (Figure 2C). Interestingly, these findings correlate with the CD86 expression and frequency of positive cells.

Figure 2. Comparison of the phenotypic and functional profile of Mo-DCs obtained from patients with hrHPV-positive and HPV-negative PSCC patients, compared to healthy donors. (A) Frequency of Mo-DCs expressing the co-stimulatory molecule CD86 in patients with hrHPV-positive (n=6) and HPV-negative (n=6) PSCC, compared to healthy donors (n=9). (B) Mean Fluorescence Intensity (MFI) of CD86 on positive cells from patients with hrHPV-positive (n=6) and HPV-negative (n=6) PSCC, compared to healthy donors (n=9). (C) Proliferation index of healthy allogeneic lymphocytes co-cultured with Mo-DCs derived from hrHPV-positive (n=6) and HPV-negative (n=6) PSCC patients, compared to healthy donors (n=6). Data are presented as mean ± standard error of the mean (SEM). Statistical analysis was performed using One-way ANOVA followed by Tukey’s multiple comparison post hoc test, with significance set at p<0.05. Significance levels are denoted as *p < 0.05 and **p < 0.01.
3.4 Lymphocyte viability in the presence of primary culture supernatant from PSCC
Assessing the impact of the tumor microenvironment on the induction of apoptosis in lymphocytes, the results showed that lymphocytes from healthy donors (38,80 ± 0,6938) exhibited a lower initial apoptosis rate compared to lymphocytes from hrHPV-positive (42,07 ± 0,1687, p=0,0067) and HPV-negative (41,90 ± 0,6351, p=0,0229) patients when cultured in medium. However, when lymphocytes were cultured in the presence of tumor supernatant, lymphocytes from healthy donors showed a significant increase in apoptotic cells (42.53 ± 0.40, p= 0.0018), compared with healthy donors. Interestingly, lymphocytes from hrHPV-positive patients had a reduction in the frequency of apoptotic cells when cultured with tumor supernatant (38.37 ± 0.78), when compared to culture with medium only (42.07 ± 0.16, p=0.0019) (Figure 3A). The data remained consistent across groups in different treatments when evaluating late apoptosis (Figure 3B). On the other hand, the frequency of viable cells was higher in lymphocytes from healthy donors (47.10 ± 1.00) cultured in the medium compared to hrHPV-positive (43.70 ± 0.98, p=0.0392) and HPV-negative patients (42.20 ± 0.11, p=0.0496). However, there was a reduction in viability of lymphocytes from healthy donors (42.53 ± 0.40) when treated with tumor supernatant (47.10 ± 1.00, p=0.0392) (Figure 3C). The data on viability and initial apoptosis complement each other, both in lymphocytes from healthy donors and patients.
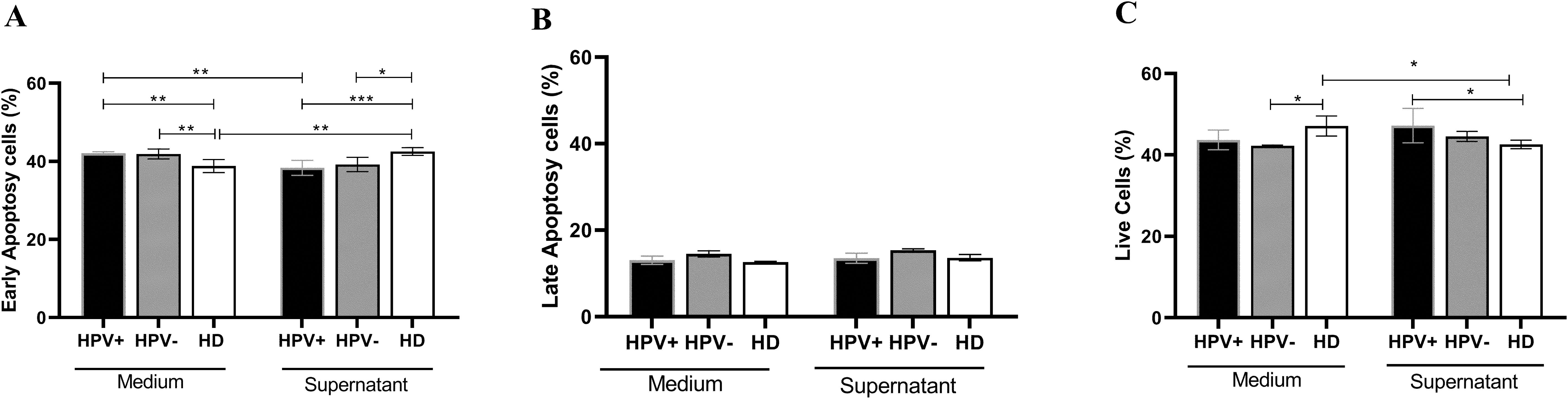
Figure 3. Viability of lymphocytes obtained from PSCC patients with hrHPV-positive and HPV-negative carcinoma and healthy donors, treated with supernatant obtained from ex vivo culture of tumor cells from penile carcinoma. (A) Frequency of lymphocytes in early apoptosis from patients with hrHPV-positive (n=6) and HPV-negative (n=6) PSCC, compared to healthy donors (n=5). (B) Frequency of lymphocytes in late apoptosis from hrHPV-positive (n=6) and HPV-negative (n=6) PSCC patients, compared to healthy donors (n=5). (C) Frequency of viable lymphocytes from hrHPV-positive (n=6) and HPV-negative (n=6) PSCC, compared to healthy donors (n=5). Data are presented as the mean ± standard error of the mean (SEM). Statistical analysis was conducted using One-way ANOVA followed by Tukey’s multiple comparison post hoc test and an unpaired t-test, with significance set at p<0.05. Statistical significance is denoted as *p < 0.05, **p < 0.01, and ***p < 0.001.
4 Discussion
In this study, the patients exhibited socioeconomic aspects related to poverty, including low economic status, low educational attainment, and rural occupation. The northeastern region of Brazil comprises states with the lowest development index in the country, being the region with the highest incidence of this cancer (9, 19, 20). Although the incidence and mortality rates of penile squamous cell carcinoma (PSCC) are predominantly concentrated in developing regions, recent studies have documented a rising trend in the incidence within developed countries (20). The data reinforce the importance of social policies aimed at education and improving the quality of life as a way to mitigate the disease.
PSCC was the most common among the patients studied, with the glans and prepuce being the most affected anatomical sites. In a study conducted at the Netherlands Cancer Institute, 84.5% of squamous cell carcinomas did not have the histological subtype identified (21). The classification of histological subtypes is important for the diagnosis of penile carcinoma, yet it is often neglected. Furthermore, upon reviewing the medical records, it was noted that some patients did not have documented data regarding HPV status at the time of diagnosis.
HPV infection is another significant risk factor in penile carcinogenesis. PSCC includes numerous histological subtypes, some of which are associated with HPV, and it is often an aggressive tumor (22–24). The data showed that both histopathological evaluation (56.5%) and Nested-PCR (66.6%) revealed that most patients had HPV infection. Macedo et al. (3) investigated HPV genotyping in patients from the same region, revealing a 96.4% frequency, with high-risk subtypes being predominant (88.6%), representing one of the highest rates ever recorded. This finding highlights the epigenetic changes induced by HPV in penile carcinogenesis (3), reinforcing the importance of high-risk HPV subtype infections in the development of this cancer.
The results of this study highlighted the importance of viral infection in modulating the immune response in the tumor microenvironment, underscoring the need to define the virus’s role in PSCC. Moreover, the data showed that some patients who tested negative for HPV by Nested-PCR showed histopathological evidence of cellular changes (koilocytosis), suggestive of HPV-associated carcinoma subtypes. HPV was identified in 42% of patients with PSCC, applying PCR assay (25). Despite being a standard technique for HPV diagnosis, this study reinforces other works that demonstrate false-negative results for HPV in PSCC samples. Considering this context, the importance of HPV in this type of tumor may be greater than previously indicated, making vaccination a powerful alternative in combating it. Moreover, this scenario underscores the significance of using additional techniques for the accurate detection of HPV infection.
Histological and immunohistochemical studies have evaluated the presence of immune cells infiltrating penile carcinoma, but the results are controversial as to density, immune profile, and the presence or absence of HPV (11–13, 26–28). On the other hand, a systematic review demonstrates that a higher presence of immune cells infiltrating penile carcinoma is associated with greater patient survival (29). In this work, flow cytometry was applied to a cell suspension obtained from tumor tissue. The data showed the presence of a phenotypic profile with both innate and adaptive immune cells in penile carcinoma. When analyzing the presence of HPV, hrHPV-positive carcinomas showed a lower frequency of T lymphocytes, and a trend for a higher frequency of NK cells compared to HPV-negative PSCC.
Viral infection induces the activation of NK cells, which can recognize and kill virus-infected cells that have down regulated surface MHC I molecules and are resistant to cytotoxic T lymphocyte-mediated death (30). In this study, the tumor-infiltrating immune cells showed that the HPV presence can modulate the immune response to an innate profile, reducing the antigen-specific response and, consequently, a better anti-tumor immune response. Thus, the data suggest that mechanisms associated with HPV can not only contribute to carcinogenesis through the infection of the virus in the host cell but also through immunoregulatory mechanisms that shape the tumor microenvironment.
In penile cancer, the overexpression of p16INK4a was associated with the presence of hrHPV and basaloid subtype tumors (31). Moreover, studies indicate that p16INK4a expression triggers a senescence-associated secretory phenotype in the tumor microenvironment, promoting the release of inflammatory cytokines. This cytokine release inhibits the activity of cytotoxic T lymphocytes and induces immunosuppression (32).
In HPV-infected epithelium, APCs also exhibit immature phenotypes characterized by the downregulation of surface MHC molecules and co-stimulatory molecules, reducing the capacity of DCs to stimulate antigen-specific T cells (33). To the best of our knowledge, this is the first study in which dendritic cells have been differentiated from monocytes of patients diagnosed with penile carcinoma. The presence of hrHPV was significant in determining an immature profile in DCs, reducing mature-DCs presence, and the capacity to express co-stimulatory molecules, suggesting viral modulation of tolerogenic immunity. However, the formation of the tumor microenvironment independent of HPV also reduced CD86 expression, indicating that there is a failure in antigen presentation as a tumor escape mechanism, which is a target for immunotherapeutic strategies. Induction of an immature profile of dendritic cells with a lower expression of co-stimulatory molecules has already been reported as a tumor escape mechanism for gastrointestinal, lung, breast, and other cancers (34–38), and the present study reinforces the participation of HPV in the formation of the microenvironment unfavorable to maturation of DCs.
The role of lymphocytes in the antitumor response has been studied and applied in immunotherapy (39, 40). Adoptive cell therapy (ACT) with tumor-infiltrating lymphocytes (TIL) initiated with the expansion of TILs from surgically resected human tumor samples has been applied (41). In this study, it was found that T lymphocytes from healthy donors are induced to undergo apoptosis when exposed to soluble factors produced by the tumor; on the other hand, T lymphocytes from patients showed greater viability. A study comparing the presence of cytotoxic T lymphocytes among HPV-positive and HPV-negative PSCC did not observe any differences (21, 41). Conversely, in usual subtype penile carcinoma, unrelated to HPV, the presence of infiltrating regulatory T cells was associated with poor survival and unfavorable prognosis (42). Thus, understanding the resistance of these lymphocytes to the tumor microenvironment simulation can lead to a better understanding of the mechanisms of tumor escape associated with HPV.
A review of patients with hrHPV-positive cervical cancer demonstrates that viral proteins increase the expression of Cyclooxygenase-2 (COX-2), leading to chronic inflammation. This, in turn, enhances the recruitment of Myeloid-Derived Suppressor Cells (MDSCs), shifts the immune response profile from Th1 to Th2, and promotes the differentiation of M2 macrophages and regulatory T cells (Tregs) (43). Similar findings have been observed in head and neck carcinomas (44). Data from this study indicate that the tumor microenvironment supports the viability of lymphocytes obtained from patients, particularly those with hrHPV-positive PSCC patients. On the other hand, it shows that the profile of DCs in these patients presents an immature phenotype and with a lower frequency of T lymphocytes infiltrating the tumor.
Thus, the presence of HPV appears to trigger an immune response that is insufficient to eliminate the tumor, thereby contributing to the progression of penile cancer. The data presented here deepen our understanding of HPV’s role in modulating the immune response in PSCC patients. This not only elucidates the pathophysiological mechanisms associated with the infection but also identifies potential therapeutic targets for immunotherapy as a promising strategy. Moreover, the immunological implications highlighted reinforce and expand the need for HPV vaccination as a means to prevent and/or mitigate both the local and systemic impacts of penile cancer.
Data availability statement
Original contributions presented in the study are included in the article and supplementary materials. Further inquiries can be directed to the corresponding author.
Ethics statement
The studies involving humans were approved by Research Ethics Committee on Humans from the Federal University of Maranhão and by the National Research Ethics Commission (CONEP- Brazil, CAAE: 46371515.5.0000.5087). The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study. Written informed consent was obtained from the individual(s) for the publication of any potentially identifiable images or data included in this article.
Author contributions
SG: Data curation, Formal analysis, Investigation, Methodology, Project administration, Writing – original draft. AV: Data curation, Formal analysis, Investigation, Methodology, Project administration, Writing – original draft. MR: Investigation, Writing – original draft, Data curation. AB: Investigation, Writing – original draft. JD: Investigation, Writing – original draft, Data curation, Methodology. AD: Investigation, Writing – original draft. LN: Investigation, Writing – original draft. RC: Writing – original draft, Investigation. SP: Funding acquisition, Resources, Writing – original draft, Writing – review & editing. AA: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This research was funded by Brazilian Federal Agency for Support and Evaluation of Graduate Education -CAPES (Finance Code 001), the Fundação de Amparo à Pesquisa do Estado do Maranhão (FAPEMA) Grant/Award Numbers 00971/17, 00744/19 and Federal University of Maranhão. AV is a post-doctoral research fellow and scholarship holder of CAPES, grant number 88887.691775/2022-00 (PDPG).
Acknowledgments
The authors are grateful to Aldenora Bello Hospital for the clinical assistance and mostly to their patients and volunteers without whom this study would not have been possible.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fonc.2024.1463445/full#supplementary-material
References
1. Thomas A, Necchi A, Muneer A, Tobias-Machado M, Tran ATH, Van Rompuy AS, et al. Penile cancer. Nat Rev Dis Primers. (2021) 7:1–24. doi: 10.1038/s41572-021-00246-5
2. Coelho RWP, Pinho JD, Moreno JS, Garbis DVEO, do Nascimento AMT, Larges JS, et al. Penile cancer in Maranhao, Northeast Brazil: the highest incidence globally? BMC Urol. (2018) 18:50. doi: 10.1186/s12894-018-0365-0
3. Macedo J, Silva E, Nogueira L, Coelho R, da Silva J, Dos Santos A, et al. Genomic profiling reveals the pivotal role of hrHPV driving copy number and gene expression alterations, including mRNA downregulation of TP53 and RB1 in penile cancer. Mol Carcinog. (2020) 59:604–17. doi: 10.1002/mc.23185
4. Leto M, Santos Júnior G, AM P, Tomimori J. Human papillomavirus infection: etiopathogenesis, molecular biology and clinical manifestations. Anais brasileiros dermatologia. (2011) 86:306–17. doi: 10.1590/S0365-05962011000200014
5. Steinestel J, Al Ghazal A, Arndt A, Schnoeller TJ, Schrader AJ, Moeller P, et al. The role of histologic subtype, p16INK4a expression, and presence of human papillomavirus DNA in penile squamous cell carcinoma. BMC cancer. (2015) 15:1–9. doi: 10.1186/s12885-015-1268-z
6. Spiess PE, Dhillon J, Baumgarten AS, Johnstone PA, Giuliano AR. Pathophysiological basis of human papillomavirus in penile cancer: Key to prevention and delivery of more effective therapies. CA: A Cancer J Clin. (2016) 66:481–95. doi: 10.3322/caac.21354
7. Canto L, da Silva JM, Castelo-Branco PV, Monteiro I, Nogueira L, Fonseca-Alves CE, et al. Mutational signature and integrative genomic analysis of human papillomavirus-associated penile squamous cell carcinomas from latin American patients. Cancers. (2022) 14:3514. doi: 10.3390/cancers14143514
8. Ornellas AA, Ornellas P. Should routine neonatal circumcision be a police to prevent penile cancer?| Opinion: Yes. Int Braz J urol. (2017) 43:7–9. doi: 10.1590/s1677-5538.ibju.2017.01.03
9. Vieira CB, Feitoza L, Pinho J, Teixeira-Júnior A, Lages J, Calixto J, et al. Profile of patients with penile cancer in the region with the highest worldwide incidence. Sci Rep. (2020) 10:1–7. doi: 10.1038/s41598-020-59831-5
10. Greenhough A, Smartt HJ, Moore AE, Roberts HR, Williams AC, Paraskeva C, et al. The COX-2/PGE 2 pathway: key roles in the hallmarks of cancer and adaptation to the tumour microenvironment. Carcinogenesis. (2009) 30:377–86. doi: 10.1093/carcin/bgp014
11. Angell H, Galon J. From the immune contexture to the Immunoscore: the role of prognostic and predictive immune markers in cancer. Curr Opin Immunol. (2013) 25:261–7. doi: 10.1016/j.coi.2013.03.004
12. Becht E, Giraldo NA, Dieu-Nosjean M-C, Sautès-Fridman C, Fridman WH. Cancer immune contexture and immunotherapy. Curr Opin Immunol. (2016) 39:7–13. doi: 10.1016/j.coi.2015.11.009
13. Chu C, Yao K, Lu J, Zhang Y, Chen K, Lu J, et al. Immunophenotypes based on the tumor immune microenvironment allow for unsupervised penile cancer patient stratification. Cancers. (2020) 12:1796. doi: 10.3390/cancers12071796
14. Pett MR, MT H, RD P, GS Y, MK S. Stanley MA: Selection of cervical keratinocytes containing integrated HPV16 associates with episome loss and an endogenous antiviral response. Proc Natl Acad Sci USA. (2006) 103:3822–7. doi: 10.1073/pnas.0600078103
15. Kanodia S, Fahey LM, Kast WM. Mechanisms used by human papillomaviruses to escape the host immune response. Curr Cancer Drug Targets. (2007) 7:79–89. doi: 10.2174/156800907780006869
16. Hasan UA, Bates E, Takeshita F, Biliato A, Accardi R, Bouvard V. TLR9 expression and function is abolished by the cervical cancer-associated human papillomavirus type 16. J Immunol. (2007) 178:3186–97. doi: 10.4049/jimmunol.178.5.3186
17. Miescher S, Whiteside TL, Carrel S, von Fliedner V. Functional properties of tumor-infiltrating and blood lymphocytes in patients with solid tumors: effects of tumor cells and their supernatants on proliferative responses of lymphocytes. J Immunol. (1986) 136:1899–907. doi: 10.4049/jimmunol.136.5.1899
18. Neves AR, Ensina LF, Anselmo LB, Leite KRM, Buzaid AC, Câmara-Lopes LH, et al. Dendritic cells derived from metastatic cancer patients vaccinated with allogeneic dendritic cell-autologous tumor cell hybrids express more CD86 and induce higher levels of interferon-gamma in mixed lymphocyte reactions. Cancer Immunol Immunother. (2005) 54:61–6. doi: 10.1007/s00262-004-0550-8
19. Koifman L, Vides AJ, Koifman N, Carvalho JP, Ornellas AA. Epidemiological aspects of penile cancer in Rio de Janeiro: evaluation of 230 cases. Int Braz J Urol. (2011) 37:231–40. doi: 10.1590/S1677-55382011000200010
20. Fu L, Tian T, Yao K, Chen XF, Luo G, Gao Y, et al. Global pattern and trends in penile cancer incidence: population-based study. JMIR Public Health surveillance. (2022) 8:e34874. doi: 10.2196/34874
21. Ottenhof SR, Djajadiningrat RS, Thygesen HH, Jakobs PJ, Jóźwiak K, Heeren AM, et al. The prognostic value of immune factors in the tumor microenvironment of penile squamous cell carcinoma. Front Immunol. (2018) 9:1253. doi: 10.3389/fimmu.2018.01253
22. Hakenberg OW, Dräger DL, Erbersdobler A, Naumann CM, Jünemann KP, Protzel C. The diagnosis and treatment of penile cancer. Deutsches Ärzteblatt Int. (2018) 115:646. doi: 10.3238/arztebl.2018.0646
23. Gross G, Pfister H. Role of human papillomavirus in penile cancer, penile intraepithelial squamous cell neoplasias and in genital warts. Med Microbiol Immunol. (2004) 193:35–44. doi: 10.1007/s00430-003-0181-2
24. Kidd LC, Chaing S, Chipollini J, Giuliano AR, Spiess PE, Sharma P. Relationship between human papillomavirus and penile cancer—implications for prevention and treatment. ” Trans Androl Urol. (2017) 6:791. doi: 10.21037/tau.2017.06.27
25. Rubin MA, Kleter B, Zhou M, Ayala G, Cubilla AL, Quint WGV, et al. Detection and typing of human papillomavirus DNA in penile carcinoma: Evidence for multiple independent pathways of penile carcinogenesis. Am J Pathol. (2001) 159:1211–8. doi: 10.1016/S0002-9440(10)62506-0
26. Comperat E. Pathology of penile cancer. Eur Urol Suppl. (2018) 17:132–7. doi: 10.1016/j.eursup.2017.08.005
27. Ahmed ME, Khalil M, Kamel MH, Karnes RJ, Spiess PE. Progress on management of penile cancer in 2020. Curr Treat options Oncol. (2021) 22:1–13. doi: 10.1007/s11864-020-00802-3
28. Mentrikoski MJ, Stelow EB, Culp S, Frierson HF, Cathro HP. Histologic and immunohistochemical assessment of penile carcinomas in a North American population. Am J Surg Pathol. (2014) 38:1340–8. doi: 10.1097/PAS.0000000000000124
29. Gooden MJM, De Bock GH, Leffers N, Daemen T, Nijman HW. The prognostic influence of tumour-infiltrating lymphocytes in cancer: a systematic review with meta-analysis. Br J Cancer. (2011) 105(1):93–103. doi: 10.1038/bjc.2011.189
30. Garcia-Iglesias T, Del Toro-Arreola A, Albarran-Somoza B, Del Toro-Arreola S, Sanchez-Hernandez PE, Ramirez-Duenas MG, et al. Low NKp30, NKp46 and NKG2D expression and reduced cytotoxic activity on NK cells in cervical cancer and precursor lesions. BMC Cancer. (2009) 9:186. doi: 10.1186/1471-2407-9-186
31. Sarier M. Association between human papillomavirus and urological cancers: an update. [Internet] In: Mol Mech Cancer. Budak M, editor. IntechOpen (2022), 177–90. doi: 10.5772/intechopen.101508
32. Reynolds LE, Maallin S, Haston S, Martinez-Barbera JP, Hodivala-Dilke KM, Pedrosa AR. Effects of senescence on the tumour microenvironment and response to therapy. FEBS J. (2024) 291:2306–19. doi: 10.1111/febs.v291.11
33. Bashaw AA, Leggatt GR, Chandra J, Tuong ZK, Frazer IH. Modulation of antigen presenting cell functions during chronic HPV infection. Papillomavirus Res. (2017) 4:58–65. doi: 10.1016/j.pvr.2017.08.002
34. Mortezaee K, Majidpoor J. (Im) maturity in tumor ecosystem. Front Oncol. (2022) 11:813897. doi: 10.3389/fonc.2021.813897
35. Wu Z, Shi H, Zhang L, Shi H, Miao X, Chen L, et al. Comparative analysis of monocyte-derived dendritic cell phenotype and T cell stimulatory function in patients with acute-on-chronic liver failure with different clinical parameters. Front Immunol. (2023) 14:1290445. doi: 10.3389/fimmu.2023.1290445
36. Morrissey ME, Byrne R, Nulty C, McCabe NH, Lynam-Lennon N, Butler CT, et al. The tumour microenvironment of the upper and lower gastrointestinal tract differentially influences dendritic cell maturation. BMC Cancer. (2020) 20:1–135. doi: 10.1186/s12885-020-07012-y
37. Perrot I, Blanchard D, Freymond N, Isaac S, Guibert B, Pacheco Y, et al. Dendritic cells infiltrating human non-small cell lung cancer are blocked at immature stage. J Immunol. (2007) 178:2763–9. doi: 10.4049/jimmunol.178.5.2763
38. Chen Xi, Shao Q, Hao S, Zhao Z, Wang Y, Guo X, et al. CTLA-4 positive breast cancer cells suppress dendritic cells maturation and function. Oncotarget. (2017) 8:13703. doi: 10.18632/oncotarget.14626
39. Tang Y, Hu X, Wu K, Li X. Immune landscape and immunotherapy for penile cancer. Front Immunol. (2022) 13:1055235. doi: 10.3389/fimmu.2022.1055235
40. Joshi VB, Spiess PE, Necchi A, Pettaway CA, Chahoud J. Immune-based therapies in penile cancer. Nat Rev Urol. (2022) 19:457–74. doi: 10.1038/s41585-022-00617-x
41. Aydin AM, Hall M, Bunch B, Branthoover H, Sannasardo Z, Mackay A, et al. Expansion of tumor-infiltrating lymphocytes (TIL) from penile cancer patients. Int Immunopharmacol. (2021) 94:107481. doi: 10.1016/j.intimp.2021.107481
42. Vassallo José, Freitas F, Hugo A, Rocha RM, da W, Zequi SC, et al. Pathologic and imunohistochemical characterization of tumoral inflammatory cell infiltrate in invasive penile squamous cell carcinomas: Fox-P3 expression is an independent predictor of recurrence. Tumor Biol. (2015) 36:2509–16. doi: 10.1007/s13277-014-2864-2
43. Chaberek K, Mrowiec M, Kaczmarek M, Dutsch-Wicherek M. The creation of the suppressive cancer microenvironment in patients with HPV-positive cervical cancer. Diagnostics. (2022) 12:1–12. doi: 10.3390/diagnostics12081906
Keywords: urological carcinoma, cancer immunomodulation, dendritic cells, HPV-related cancer, costimulatory molecules
Citation: Guimarães SJA, Vale AAM, Rocha MCB, Butarelli ALdA, da Silva JM, Deus AJSd, Nogueira L, Coelho RWP, Pereira SR and Azevedo-Santos APS (2024) Human papillomavirus infection affects the immune microenvironment and antigen presentation in penile cancer. Front. Oncol. 14:1463445. doi: 10.3389/fonc.2024.1463445
Received: 11 July 2024; Accepted: 30 September 2024;
Published: 18 October 2024.
Edited by:
Ana Luisa Teixeira, Portuguese Oncology Institute, PortugalReviewed by:
Mehmet Sarier, University of Istinye, TürkiyeMagaly Martinez-Ferrer, University of Puerto Rico, Puerto Rico
Copyright © 2024 Guimarães, Vale, Rocha, Butarelli, da Silva, Deus, Nogueira, Coelho, Pereira and Azevedo-Santos. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ana Paula Silva Azevedo-Santos, YW5hLmF6ZXZlZG9AdWZtYS5icg==
†These authors share first authorship