 Shuo Wu1
Shuo Wu1 Xiaodong Jiang
Xiaodong Jiang
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Oncol., 17 June 2024
Sec. Cancer Molecular Targets and Therapeutics
Volume 14 - 2024 | https://doi.org/10.3389/fonc.2024.1408051
B7-H3 (CD276), an immune checkpoint molecule, is overexpressed in various types of cancer and their tumor vasculature, demonstrating significant associations with adverse clinical outcomes. In addition to its well-known immune functions, B7-H3 exhibits dual co-stimulatory/co-inhibitory roles in normal physiology and the tumor microenvironment. The non-immune functions of B7-H3 in tumor cells and the tumor vasculature, including promoting tumor cell anti-apoptosis, proliferation, invasion, migration, drug resistance, radioresistance, as well as affecting cellular metabolism and angiogenesis, have increasingly gained attention from researchers. Particularly, the co-expression of B7-H3 in both tumor cells and tumor endothelial cells highlights the higher potential and clinical utility of therapeutic strategies targeting B7-H3. This review aims to summarize the recent advances in understanding the non-immune functions of B7-H3 in tumors and provide insights into therapeutic approaches targeting B7-H3, focusing on its co-expression in tumor cells and endothelial cells. The aim is to establish a theoretical foundation and practical reference for the development and optimization of B7-H3-targeted therapies.
CD276, also known as B7 homolog 3 (B7-H3), is a type I transmembrane protein consisting of 316 amino acids. It is encoded by the gene located on chromosome 9 in mice and the human 15q24 region. The primary structure of B7-H3 includes an extracellular Ig-like domain, a transmembrane region, and a short cytoplasmic tail (1). Based on the number of extracellular Ig-like domains, membrane-bound B7-H3 can be classified into two isoforms: 2IgB7-H3 and 4IgB7-H3. The former contains one IgV (variable) and one IgC (constant) domain, while the latter possesses tandem IgV and IgC domains due to exon duplication (2).
B7-H3 is a cell surface tumor endothelial marker with up to 30% amino acid homology to other members of the B7 family. It is also expressed in tumor-associated endothelial cells and is often associated with advanced tumor staging (3). B7-H3 has been shown to differentiate pathological and physiological angiogenesis in both mice and humans (4). The high expression of B7-H3 protein is observed in various human cancers such as lung, breast, colon, endometrial, renal, and ovarian cancer tumor vasculature, while it is not expressed in normal ovarian vasculature (5).
Although initially characterized as a T-cell co-stimulatory protein, current research describes B7-H3 as a T-cell inhibitory molecule that promotes tumor invasion and proliferation. It mediates immune escape primarily by inhibiting T-cell infiltration and promoting CD8+ T-cell exhaustion, suggesting that B7-H3 may serve as an important immune target in cancer therapy (6). However, B7-H3 also functions as an independent protein outside of the immune system, participating in non-immune responses in tumors. It plays a crucial role in tumor cell anti-apoptosis, proliferation, invasion, migration, drug resistance, radioresistance, metabolism, and angiogenesis (7). The exact mechanisms through which B7-H3 promotes tumor development independently of the immune system are not fully understood. Nevertheless, extensive research suggests that B7-H3 may exert its effects upstream of signaling pathways, indicating that targeting these pathways could provide new approaches for cancer treatment (3). The non-immune functions of B7H3 promoting tumor progression are summarized in Figure 1.
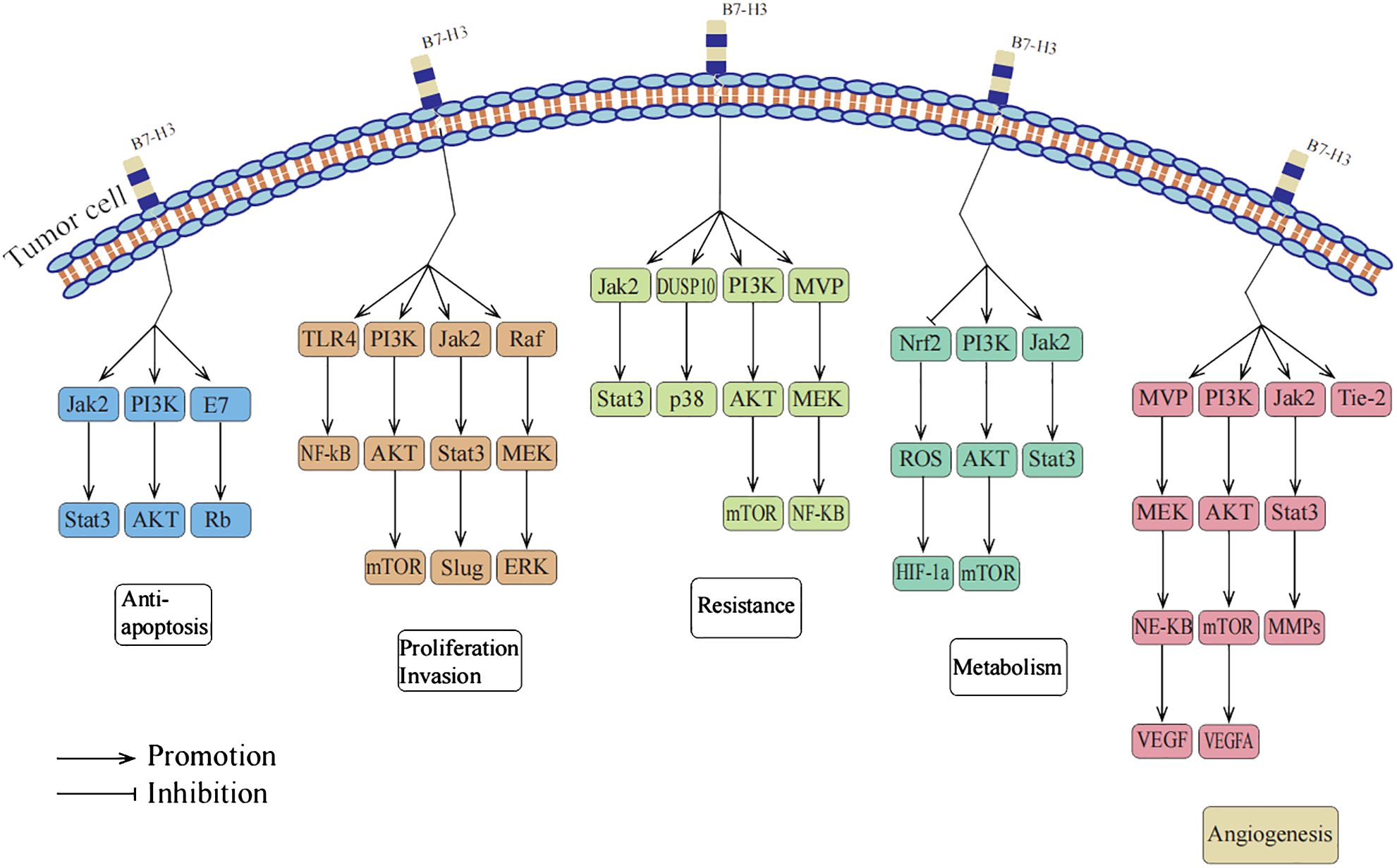
Figure 1 B7-H3 promotes tumor progression through non-immune functions.
Apoptosis is a genetically regulated, orderly process of cell death primarily responsible for eliminating aged or abnormal cells. Resistance to apoptosis has been widely recognized as a crucial hallmark of malignant tumors (8). B7-H3 promotes tumor development by suppressing apoptosis in tumor cells (9). In a multi-omic analysis and single-cell sequencing study, Zhou et al. found a significant increase in B7-H3 levels during the cellular senescence process in tumor cells, suggesting a critical role of B7-H3 in preventing tumor cell senescence (10). On the other hand, the role of B7-H3 in tumor growth has also garnered attention. Early studies on breast cancer cells demonstrated that silencing B7-H3 expression in the MCF-7 cell line led to an upregulation of vascular endothelial growth factor (VEGF) at both mRNA and protein levels, indicating that B7-H3 may inhibit tumor growth by suppressing VEGF expression (11). However, subsequent research revealed that the expression of B7-H3 actually facilitates tumor growth in other breast cancer cell lines, suggesting a complex and potentially contradictory role of B7-H3 in the pathogenesis of breast cancer (12). These findings indicate that B7-H3 may play a multifaceted role in the development of tumors, with specific effects likely dependent on the tumor type, underlying mechanisms, and biological context. Nonetheless, the majority of current research supports the view that B7-H3 promotes tumor growth through its anti-apoptotic effects. The summary of signaling pathways mediating the anti-apoptotic effects of B7-H3 is presented in Table 1.

Table 1 Signaling Pathways Mediating the Diverse Functions of B7-H3.
Recent studies have revealed the significant role of B7-H3 in promoting cancer cell proliferation, migration, and invasion (21, 35, 44, 45). In various cancers, including oral cancer, hepatocellular carcinoma, colorectal cancer, lung adenocarcinoma, hematologic malignancies, and gynecologic tumors, B7-H3 has been found to promote cancer cell proliferation by regulating different signaling pathways (46). B7-H3 can also enhance cancer cell migration by modulating the expression of E-cadherin, increasing their migratory capacity (20). Additionally, B7-H3 can augment cancer cell invasion into surrounding tissues by regulating the activity of matrix metalloproteinases (MMPs) (22). These findings provide us with new insights and suggest that targeting B7-H3 and its associated signaling pathways could be an effective strategy for the treatment of these cancers. The summary of signaling pathways mediating the proliferation, invasion, and migration effects of B7-H3 is presented in Table 1.
Drug resistance is a reflection of tumor evolution and is a major cause of cancer recurrence and patient mortality (47). Studies have shown that overexpression of B7-H3 often leads to drug resistance in cancer cells. Different mechanisms of resistance exist in different tumor cells. For example, the high expression of the B7-H3 immune checkpoint protein in neuroblastoma is involved in carcinogenic signaling, tumor cell plasticity, and the development of drug resistance (48). However, inhibiting the expression of B7-H3 or blocking its associated signaling pathways can reverse this resistance. One approach is to use a combination of paclitaxel and MEK inhibitors to block the B7-H3 signal, which has been demonstrated to effectively reverse chemotherapy resistance (16). Another approach is to use the specific inhibitor LY294002 to block the PI3K signaling pathway, which can also effectively inhibit this resistance (49). Another study found that bromodomain and extra-terminal domain inhibitors could be used to reduce the levels of B7-H3 protein and mRNA in pancreatic cancer cells, providing a new therapeutic strategy to overcome immune and chemotherapy resistance in pancreatic cancer (21). In related studies on traditional Chinese medicine, researchers found that artemisinin-mediated inhibition of B7-H3 may help enhance neuroblastoma cell sensitivity to doxorubicin (50). Additionally, Astragaloside IV enhanced the chemosensitivity to cisplatin by inhibiting B7-H3, suggesting that a combination therapy using Astragaloside IV and a B7-H3 inhibitor may be a potential treatment method for lung cancer patients (51). The summary of signaling pathways mediating the drug resistance effects of B7-H3 is presented in Table 1.
Radiation therapy is a common treatment modality for solid tumors; however, radiation resistance is a major factor contributing to treatment failure in cancer (52). A study by Ma et al. reported an interesting phenomenon: after X-ray irradiation, the expression of B7-H3 was upregulated in colorectal cancer cells, and this upregulation depended on the upregulation of KIF15 via the NF-κB signaling pathway. Further research revealed that B7-H3/KIF15, through activation of the ERK1/2 signaling pathway, promoted radiation resistance in colorectal cancer (31). Additionally, Zhou et al. found that B7-H3 increased the radiation resistance of gastric cancer cells by regulating baseline levels of cellular autophagy. In cells with high B7-H3 expression, increasing the baseline level of cellular autophagy using rapamycin enhanced their sensitivity to radiation (32). These studies all point to the important role of B7-H3 in regulating tumor cell resistance to radiation therapy. Therefore, finding effective ways to inhibit or modulate the expression and function of B7-H3 may provide new strategies to improve the effectiveness of radiation therapy.
The Warburg effect is one of the characteristic features of tumor cell metabolism, where cancer cells tend to produce energy through glycolysis even in the presence of abundant oxygen, which is significantly different from the metabolic mechanism of normal cells (53). Research by Lim et al. found that B7-H3 plays an important role in the metabolic reprogramming of cancer cells. In cells expressing B7-H3, glucose uptake and lactate production are increased (34). On the other hand, there is an interaction between the glycolytic enzyme ENO1 and B7-H3. Downregulation of B7-H3 in cervical cancer HeLa cells leads to decreased levels of ATP, lactate, c-Myc, and lactate dehydrogenase A (54). Furthermore, overexpression of B7-H3 effectively increases glucose consumption and lactate production rate, while knockout of B7-H3 has the opposite effect. Liu et al. demonstrated that using the anti-B7-H3 antibody 8H9 can shift cellular metabolism from glycolysis to oxidative phosphorylation, and they suggested that anti-B7-H3 blockade may alter tumor glucose metabolism through reactive oxygen species-mediated pathways by measuring cellular ROS levels in A549 cells treated with the anti-B7-H3 antibody using fluorescence probes (55). In addition to changes in glucose metabolism, cancer cells often possess altered fatty acid metabolism properties. In glioma cells, genetic risk features associated with fatty acid catabolism are significantly correlated with the immune checkpoint molecule B7-H3 (56). These research findings further emphasize the important role of B7-H3 in the metabolic regulation of cancer cells and may provide new therapeutic strategies for targeting this process. The summary of signaling pathways mediating the metabolism effects of B7-H3 is presented in Table 1.
In cancer development, angiogenesis plays a crucial role by providing oxygen and nutrients to tumors while allowing their spread to other parts of the body. Thus, inhibiting or reducing the blood supply in the tumor microenvironment to restrict tumor angiogenesis has become a key strategy against various solid tumors (57). B7-H3 is a highly expressed protein in many types of cancer, including lung, breast, colorectal, endometrial, renal, ovarian, hepatocellular carcinoma, and melanoma. However, this protein is not expressed in normal vascular tissues (4, 5). Furthermore, in hepatocellular carcinoma, colorectal cancer, renal cell carcinoma, and melanoma, the expression rate of B7-H3 in the tumor vasculature reaches 86% to 98% (58). In ovarian cancer, although the expression rate of B7-H3 is only 44%, it is undetectable in normal ovarian vascular tissues (5).
Under normal physiological conditions, the process of angiogenesis is strictly regulated by factors that promote and inhibit angiogenesis to maintain a stable internal environment and prevent uncontrolled blood vessel growth. However, B7-H3 protein can disrupt this balance by enhancing the activity of several pro-angiogenic factors, favoring tumor development (9). It mainly activates multiple signaling pathways, such as MMP-2, MMP-9, NF-κB, AKT1/mTOR/VEGFA, and Tie-2, thus promoting tumor angiogenesis and further facilitating tumor growth and spread (38–41). By inhibiting key molecules in these signaling pathways, we can effectively inhibit angiogenesis. For example, in neuroblastoma, overexpression of miR-29 can activate the JAK/STAT1 signaling pathway, weakening the regulatory effect of MYC-B7-H3 and inhibiting tumor angiogenesis (38). Additionally, Wang et al. found that recombinant VEGFA can eliminate the inhibitory effect of shB7-H3 on human umbilical vein endothelial cell angiogenesis in conditioned medium from colorectal cancer cells. Furthermore, siRNA against VEGFA or neutralizing antibodies against VEGFA can reverse the effects of B7-H3 in overexpressed colorectal cancer cell-conditioned medium on human umbilical vein endothelial cell angiogenesis (39). These studies further elucidate the potential role of B7-H3 in tumor angiogenesis and may serve as a crucial target for future anti-angiogenic therapies.
Moreover, B7-H3 also plays an important role in vasculogenic mimicry (VM). VM is an alternative microcirculation pattern independent of angiogenesis, where cancer cells form microvascular-like channels to provide nutrients and oxygen to tumors (59). Recent studies have found that B7-H3 expressed in tumors promotes VM formation in non-small cell lung cancer cells through the PI3K/AKT signaling pathway without affecting normal angiogenesis, providing a new strategy for future anticancer therapies (42). The summary of signaling pathways mediating B7-H3-induced tumor angiogenesis is presented in Table 1.
In numerous types of tumors, the widespread overexpression of B7-H3 on cancer cells and tumor-associated blood vessels makes it a potential ideal dual-target for therapy. CD276-ADC, utilizing pyrrolobenzodiazepine as a linker, demonstrates significant cytotoxicity against cancer cells while also exhibiting remarkable destructive effects on the tumor vascular system. This therapy shows great potential in eradicating large existing tumors and metastatic lesions, with the possibility of significantly improving long-term overall survival rates for patients (5). To target the overexpression of B7-H3 in tumor cells and tumor vasculature, along with its limited expression in normal tissue vasculature, researchers have employed an innovative strategy by conjugating the Fab fragment of anti-CD276 antibody with the photosensitizer IRDye700 to form a novel CD276-targeted agent. The findings show that combining photodynamic therapy using this agent with inhibition strategies targeting the PD-L1/PD-1 axis can induce strong local and systemic anti-tumor responses, effectively eliminating primary and metastatic tumors (60). Chen et al. reported the development of pH-responsive drug release using B7H3-targeted doxorubicin-conjugated gold nano-cages (B7H3/Dox@GNCs), which represents a selective, precise, and synergistic chemotherapy-photothermal therapy for NSCLC, capable of simultaneously destroying B7H3-positive tumor cells, tumor-associated vasculature, and cancer-associated fibroblasts (61). The expression of B7-H3 in the tumor vasculature can also be utilized to significantly improve the diagnostic accuracy of breast cancer, which is undoubtedly an important discovery (62).
Furthermore, as a promising immunotherapy target, B7-H3 has been applied in combination treatment strategies. When used as a key element in T cell co-stimulation and anti-angiogenesis therapy for hepatocellular carcinoma, the effect of B7-H3 is significantly superior to using these treatments alone. Injecting plasmids expressing B7-H3 into subcutaneous tumors in mice resulted in complete eradication of these tumors within 24 hours, which was not achieved when using angiostatin or B7-H3 treatment alone (63). This process revealed the ability of angiostatin gene transfer to inhibit tumor angiogenesis and enhance NK cell infiltration, as well as the critical role of B7-H3 in activating CD8+ tumor infiltration, increasing circulating IFN-c levels, and even completely regressing distant tumor nodules (63). However, although B7-H3 plays an important role in the aforementioned combination treatment strategies, its primary role in the broader field of immunotherapy is still as an immune checkpoint inhibitor. For example, in the treatment of triple-negative breast cancer, Cheng et al. found that blocking B7-H3 with anti-B7-H3 antibodies improved the abnormal state of the tumor vascular system, thereby enhancing the effects of chemotherapy and PD-1 treatment (64). These research findings further emphasize that B7-H3 can serve as an important therapeutic target in anticancer treatment strategies, whether used alone or in combination with other treatment modalities. More clinical trials targeting B7-H3 for cancer treatment are listed in Table 2.
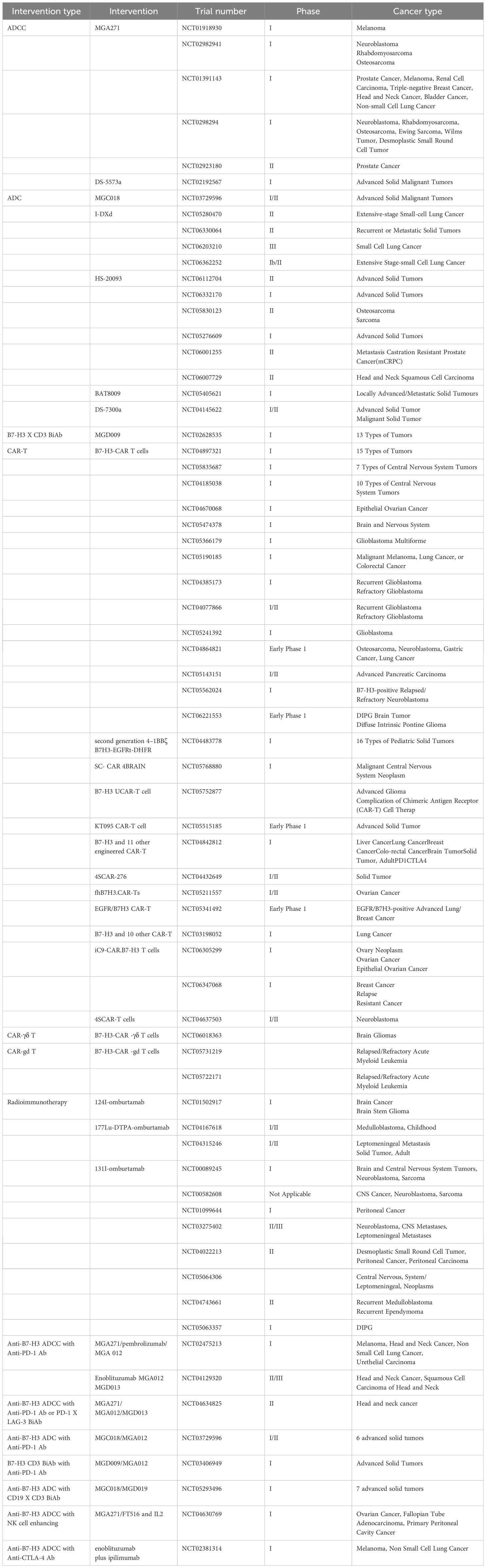
Table 2 Clinical Trials Targeting B7-H3.
Compared to other immune checkpoints, the uniqueness of B7-H3 lies in its ability to regulate both innate and adaptive immune responses. Furthermore, it promotes tumor development by stimulating various non-immune-related functions, making it an important prognostic marker for adverse outcomes. B7-H3 influences the life processes of tumor cells, including apoptosis, proliferation, invasion, migration, drug resistance, resistance to radiotherapy, metabolic processes, and abnormal angiogenesis, through the regulation of various signaling pathways. Therefore, inhibiting B7-H3 expression or blocking related signaling pathways may be an effective approach to prevent tumor progression.
Considering the high expression of B7-H3 in tumor cells and tumor vasculature, along with its low expression in normal vasculature, it becomes an ideal dual-target in cancer therapy. By considering B7-H3 as a bridge between tumor cells and the tumor vascular system, we can achieve dual therapeutic effects by targeting B7-H3. However, since the role and signaling pathways of B7-H3 may vary among different tumors and even within the vascular systems of different tumors, gaining a deeper understanding of the specific functions of B7-H3 in particular tumors will help us design more targeted treatment plans for patients, thus effectively controlling tumor development. This could be an important direction for future research in cancer therapy.
SW: Writing – original draft, Writing – review & editing. CH: Visualization, Writing – review & editing. KH: Visualization, Writing – review & editing. XJ: Conceptualization, Writing – review & editing.
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
The authors thank all members involved in this work.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Kontos F, Michelakos T, Kurokawa T, Sadagopan A, Schwab JH, Ferrone CR, et al. B7-H3: an attractive target for antibody-based immunotherapy. Clin Cancer Res. (2021) 27:1227–35. doi: 10.1158/1078-0432.CCR-20-2584
2. Kanchan RK, Doss D, Khan P, Nasser MW, Mahapatra S. To kill a cancer: Targeting the immune inhibitory checkpoint molecule, B7-H3. Biochim Biophys Acta (BBA) Rev Cancer. (2022) 1877:188783. doi: 10.1016/j.bbcan.2022.188783
3. Flem-Karlsen K, Fodstad Ø, Tan M, Nunes-Xavier CE. B7-H3 in cancer – beyond immune regulation. Trends Cancer. (2018) 4:401–4. doi: 10.1016/j.trecan.2018.03.010
4. Seaman S, Stevens J, Yang MY, Logsdon D, Graff-Cherry C, St. Croix B. Genes that distinguish physiological and pathological angiogenesis. Cancer Cell. (2007) 11:539–54. doi: 10.1016/j.ccr.2007.04.017
5. Seaman S, Zhu Z, Saha S, Zhang XM, Yang MY, Hilton MB, et al. Eradication of tumors through simultaneous ablation of CD276/B7-H3-positive tumor cells and tumor vasculature. Cancer Cell. (2017) 31:501–515.e8. doi: 10.1016/j.ccell.2017.03.005
6. Mortezaee K. B7-H3 immunoregulatory roles in cancer. BioMed Pharmacother. (2023) 163:114890. doi: 10.1016/j.biopha.2023.114890
7. Feng Y, Xie K, Yin Y, Li B, Pi C, Xu X, et al. A novel anti-B7-H3 × Anti-CD3 bispecific antibody with potent antitumor activity. Life (Basel). (2022) 12:157. doi: 10.3390/life12020157
8. Sun M, Xie J, Zhang D, Chen C, Lin S, Chen Y, et al. B7-H3 inhibits apoptosis of gastric cancer cell by interacting with Fibronectin. J Cancer. (2021) 12:7518–26. doi: 10.7150/jca.59263
9. Zhou X, Ouyang S, Li J, Huang X, Ai X, Zeng Y, et al. The novel non-immunological role and underlying mechanisms of B7-H3 in tumorigenesis. J Cell Physiol. (2019) 234:21785–95. doi: 10.1002/jcp.28936
10. Zhang Q, Tang Y, Hu G, Yuan Z, Zhang S, Sun Y, et al. Comprehensive pan-cancer analysis identifies cellular senescence as a new therapeutic target for cancer: multi-omics analysis and single-cell sequencing validation. Am J Cancer Res. (2022) 12:4103–19.
11. Sun J, Guo YD, Li XN, Zhang YQ, Gu L, Wu PP, et al. B7-H3 expression in breast cancer and upregulation of VEGF through gene silence. Onco Targets Ther. (2014) 7:1979–86. doi: 10.2147/OTT
12. Montoyo-Pujol YG, García-Escolano M, Ponce JJ, Delgado-García S, Martín TA, Ballester H, et al. Variable intrinsic expression of immunoregulatory biomarkers in breast cancer cell lines, mammospheres, and co-cultures. Int J Mol Sci. (2023) 24:4478. doi: 10.3390/ijms24054478
13. Zhang T, Jiang B, Zou ST, Liu F, Hua D. Overexpression of B7-H3 augments anti-apoptosis of colorectal cancer cells by Jak2-STAT3. World J Gastroenterol. (2015) 21:1804–13. doi: 10.3748/wjg.v21.i6.1804
14. Wang R, Sun L, Xia S, Wu H, Ma Y, Zhan S, et al. B7-H3 suppresses doxorubicin-induced senescence-like growth arrest in colorectal cancer through the AKT/TM4SF1/SIRT1 pathway. Cell Death Dis. (2021) 12:453. doi: 10.1038/s41419-021-03736-2
15. Han S, Shi X, Liu L, Zong L, Zhang J, Chen Q, et al. Roles of B7-H3 in cervical cancer and its prognostic value. J Cancer. (2018) 9:2612–24. doi: 10.7150/jca.24959
16. Liu Z, Zhang W, Phillips JB, Arora R, McClellan S, Li J, et al. Immunoregulatory protein B7-H3 regulates cancer stem cell enrichment and drug resistance through MVP-mediated MEK activation. Oncogene. (2019) 38:88–102. doi: 10.1038/s41388-018-0407-9
17. Wang C, Li Y, Jia L, Kim JK, Li J, Deng P, et al. CD276 expression enables squamous cell carcinoma stem cells to evade immune surveillance. Cell Stem Cell. (2021) 28:1597–1613.e7. doi: 10.1016/j.stem.2021.04.011
18. Zhang J, Liu L, Han S, Li Y, Qian Q, Zhang Q, et al. B7-H3 is related to tumor progression in ovarian cancer. Oncol Rep. (2017) 38:2426–34. doi: 10.3892/or.2017.5858
19. Li Y, Yang X, Wu Y, Zhao K, Ye Z, Zhu J, et al. B7-H3 promotes gastric cancer cell migration and invasion. Oncotarget. (2017) 8:71725–35. doi: 10.18632/oncotarget.v8i42
20. Zhong C, Tao B, Chen Y, Guo Z, Yang X, Peng L, et al. B7-H3 regulates glioma growth and cell invasion through a JAK2/STAT3/slug-dependent signaling pathway. Onco Targets Ther. (2020) 13:2215–24. doi: 10.2147/OTT
21. Zhao J, Meng Z, Xie C, Yang C, Liu Z, Wu S, et al. B7-H3 is regulated by BRD4 and promotes TLR4 expression in pancreatic ductal adenocarcinoma. Int J Biochem Cell Biol. (2019) 108:84–91. doi: 10.1016/j.biocel.2019.01.011
22. Kang FB, Wang L, Jia HC, Li D, Li HJ, Zhang YG, et al. B7-H3 promotes aggression and invasion of hepatocellular carcinoma by targeting epithelial-to-mesenchymal transition via JAK2/STAT3/Slug signaling pathway. Cancer Cell Int. (2015) 15:45. doi: 10.1186/s12935-015-0195-z
23. Liao H, Ding M, Zhou N, Yang Y, Chen L. B7−H3 promotes the epithelial−mesenchymal transition of NSCLC by targeting SIRT1 through the PI3K/AKT pathway. Mol Med Rep. (2022) 25:79. doi: 10.3892/mmr
24. Tekle C, Nygren MK, Chen YW, Dybsjord I, Nesland JM, Maelandsmo GM, et al. B7-H3 contributes to the metastatic capacity of melanoma cells by modulation of known metastasis-associated genes. Int J Cancer. (2012) 130:2282–90. doi: 10.1002/ijc.26238
25. Wang S, Zhang X, Ning H, Dong S, Wang G, Sun R. B7 homolog 3 induces lung metastasis of breast cancer through Raf/MEK/ERK axis. Breast Cancer Res Treat. (2022) 193:405–16. doi: 10.1007/s10549-022-06520-8
26. Xie J, Sun M, Zhang D, Chen C, Lin S, Zhang G. Fibronectin enhances tumor metastasis through B7-H3 in clear cell renal cell carcinoma. FEBS Open Bio. (2021) 11:2977–87. doi: 10.1002/2211-5463.13280
27. Liu H, Tekle C, Chen YW, Kristian A, Zhao Y, Zhou M, et al. B7-H3 silencing increases paclitaxel sensitivity by abrogating Jak2/Stat3 phosphorylation. Mol Cancer Ther. (2011) 10:960–71. doi: 10.1158/1535-7163.MCT-11-0072
28. Zhao X, Zhang GB, Gan WJ, Xiong F, Li Z, Zhao H, et al. Silencing of B7-H3 increases gemcitabine sensitivity by promoting apoptosis in pancreatic carcinoma. Oncol Lett. (2013) 5:805–12. doi: 10.3892/ol.2013.1118
29. Flem-Karlsen K, Tekle C, Øyjord T, Flørenes VA, Mælandsmo GM, Fodstad Ø, et al. p38 MAPK activation through B7-H3-mediated DUSP10 repression promotes chemoresistance. Sci Rep. (2019) 9:5839. doi: 10.1038/s41598-019-42303-w
30. Zhang P, Chen Z, Ning K, Jin J, Han X. Inhibition of B7-H3 reverses oxaliplatin resistance in human colorectal cancer cells. Biochem Biophys Res Commun. (2017) 490:1132–8. doi: 10.1016/j.bbrc.2017.07.001
31. Ma Y, Wang R, Lu H, Li X, Zhang G, Fu F, et al. B7-H3 promotes the cell cycle-mediated chemoresistance of colorectal cancer cells by regulating CDC25A. J Cancer. (2020) 11:2158–70. doi: 10.7150/jca.37255
32. Zhou L, Zhao Y. B7-H3 induces ovarian cancer drugs resistance through an PI3K/AKT/BCL-2 signaling pathway. Cancer Manag Res. (2019) 11:10205–14. doi: 10.2147/CMAR
33. Yue G, Tang J, Zhang L, Niu H, Li H, Luo S. CD276 suppresses CAR-T cell function by promoting tumor cell glycolysis in esophageal squamous cell carcinoma. J Gastrointest Oncol. (2021) 12:38–51. doi: 10.21037/jgo
34. Lim S, Liu H, Madeira da Silva L, Arora R, Liu Z, Phillips JB, et al. Immunoregulatory protein B7-H3 reprograms glucose metabolism in cancer cells by ROS-mediated stabilization of HIF1α. Cancer Res. (2016) 76:2231–42. doi: 10.1158/0008-5472.CAN-15-1538
35. Li Z, Liu J, Que L, Tang X. The immunoregulatory protein B7-H3 promotes aerobic glycolysis in oral squamous carcinoma via PI3K/Akt/mTOR pathway. J Cancer. (2019) 10:5770–84. doi: 10.7150/jca.29838
36. Shi T, Ma Y, Cao L, Zhan S, Xu Y, Fu F, et al. B7-H3 promotes aerobic glycolysis and chemoresistance in colorectal cancer cells by regulating HK2. Cell Death Dis. (2019) 10:308. doi: 10.1038/s41419-019-1549-6
37. Zhu X, Shi Y, Wang J. B7-H3 regulates glucose metabolism in neuroblastom via stat3/c-met pathway. Appl Biochem Biotechnol. (2023) 196:1386–98. doi: 10.1007/s12010-023-04606-7
38. Purvis IJ, Avilala J, Guda MR, Venkataraman S, Vibhakar R, Tsung AJ, et al. Role of MYC-miR-29-B7-H3 in medulloblastoma growth and angiogenesis. J Clin Med. (2019) 8:1158. doi: 10.3390/jcm8081158
39. Wang R, Ma Y, Zhan S, Zhang G, Cao L, Zhang X, et al. B7-H3 promotes colorectal cancer angiogenesis through activating the NF-κB pathway to induce VEGFA expression. Cell Death Dis. (2020) 11:55. doi: 10.1038/s41419-020-2252-3
40. Wu R, Zhang Y, Xu X, You Q, Yu C, Wang W, et al. Exosomal B7-H3 facilitates colorectal cancer angiogenesis and metastasis through AKT1/mTOR/VEGFA pathway. Cell Signal. (2023) 109:110737. doi: 10.1016/j.cellsig.2023.110737
41. Zhang X, Ji J, Zhang G, Fang C, Jiang F, Ma S, et al. Expression and significance of B7-H3 and Tie-2 in the tumor vasculature of clear cell renal carcinoma. Onco Targets Ther. (2017) 0:5417–24. doi: 10.2147/OTT
42. Fan X, Huang J, Hu B, Zhou J, Chen L. Tumor-expressed B7-H3 promotes vasculogenic mimicry formation rather than angiogenesis in non-small cell lung cancer. J Cancer Res Clin Oncol. (2023) 149:8729–41. doi: 10.1007/s00432-023-04790-3
43. Cheng R, Wang B, Cai XR, Chen ZS, Du Q, Zhou LY, et al. CD276 promotes vasculogenic mimicry formation in hepatocellular carcinoma via the PI3K/AKT/MMPs pathway. Onco Targets Ther. (2020) 13:11485–98. doi: 10.2147/OTT.S271891
44. Hu X, Xu M, Hu Y, Li N, Zhou L. B7-H3, negatively regulated by miR-128, promotes colorectal cancer cell proliferation and migration. Cell Biochem Biophys. (2021) 79:397–405. doi: 10.1007/s12013-021-00975-0
45. Li Y, Zhang J, Han S, Qian Q, Chen Q, Liu L, et al. B7-H3 promotes the proliferation, migration and invasiveness of cervical cancer cells and is an indicator of poor prognosis. Oncol Rep. (2017) 38:1043–50. doi: 10.3892/or.2017.5730
46. Tang XY, Luo ZL, Xiong YL, Yang J, Shi AP, Zheng KF, et al. The proliferative role of immune checkpoints in tumors: double regulation. Cancers (Basel). (2022) 14:5374. doi: 10.3390/cancers14215374
47. Shi Y, Xu Y, Xu Z, Wang H, Zhang J, Wu Y, et al. TKI resistant-based prognostic immune related gene signature in LUAD, in which FSCN1 contributes to tumor progression. Cancer Lett. (2022) 532:215583. doi: 10.1016/j.canlet.2022.215583
48. Pulido R, Nunes-Xavier CE. Hopes on immunotherapy targeting B7-H3 in neuroblastoma. Transl Oncol. (2023) 27:101580. doi: 10.1016/j.tranon.2022.101580
49. Jiang B, Liu F, Liu Z, Zhang T, Hua D. B7-H3 increases thymidylate synthase expression via the PI3k-Akt pathway. Tumour Biol. (2016) 37:9465–72. doi: 10.1007/s13277-015-4740-0
50. Tan WQ, Chen G, Ye M, Jia B. Artemether regulates chemosensitivity to doxorubicin via regulation of B7-H3 in human neuroblastoma cells. Med Sci Monit. (2017) 23:4252–9. doi: 10.12659/MSM.902068
51. Wang S, Mou J, Cui L, Wang X, Zhang Z. Astragaloside IV inhibits cell proliferation of colorectal cancer cell lines through down-regulation of B7-H3. BioMed Pharmacother. (2018) 102:1037–44. doi: 10.1016/j.biopha.2018.03.127
52. Wang XC, Yue X, Zhang RX, Liu TY, Pan ZZ, Yang MJ, et al. Genome-wide RNAi screening identifies RFC4 as a factor that mediates radioresistance in colorectal cancer by facilitating nonhomologous end joining repair. Clin Cancer Res. (2019) 25:4567–79. doi: 10.1158/1078-0432.CCR-18-3735
53. DeBerardinis RJ, Lum JJ, Hatzivassiliou G, Thompson CB. The biology of cancer: metabolic reprogramming fuels cell growth and proliferation. Cell Metab. (2008) 7:11–20. doi: 10.1016/j.cmet.2007.10.002
54. Zuo J, Wang B, Long M, Gao Z, Zhang Z, Wang H, et al. The type 1 transmembrane glycoprotein B7-H3 interacts with the glycolytic enzyme ENO1 to promote Malignancy and glycolysis in HeLa cells. FEBS Lett. (2018) 592:2476–88. doi: 10.1002/1873-3468.13164
55. Liu J, Yang S, Cao B, Zhou G, Zhang F, Wang Y, et al. Targeting B7-H3 via chimeric antigen receptor T cells and bispecific killer cell engagers augments antitumor response of cytotoxic lymphocytes. J Hematol Oncol. (2021) 14:21. doi: 10.1186/s13045-020-01024-8
56. Qi Y, Chen D, Lu Q, Yao Y, Ji C. Bioinformatic profiling identifies a fatty acid metabolism-related gene risk signature for Malignancy, prognosis, and immune phenotype of glioma. Dis Markers. (2019) 2019:3917040. doi: 10.1155/2019/3917040
57. Al-Ostoot FH, Salah S, Khamees HA, Khanum SA. Tumor angiogenesis: Current challenges and therapeutic opportunities. Cancer Treat Res Commun. (2021) 28:100422. doi: 10.1016/j.ctarc.2021.100422
58. Michelakos T, Kontos F, Barakat O, Maggs L, Schwab JH, Ferrone CR, et al. B7-H3 targeted antibody-based immunotherapy of Malignant diseases. Expert Opin Biol Ther. (2021) 21:587–602. doi: 10.1080/14712598.2021.1862791
59. de la Cruz ONH, López-González JS, García-Vázquez R, Salinas-Vera YM, Muñiz-Lino MA, Aguilar-Cazares D, et al. Regulation networks driving vasculogenic mimicry in solid tumors. Front Oncol. (2020) 9:1419. doi: 10.3389/fonc.2019.01419
60. Bao R, Wang Y, Lai J, Zhu H, Zhao Y, Li S, et al. Enhancing anti-PD-1/PD-L1 immune checkpoint inhibitory cancer therapy by CD276-targeted photodynamic ablation of tumor cells and tumor vasculature. Mol Pharm. (2019) 16:339–48. doi: 10.1021/acs.molpharmaceut.8b00997
61. Chen B, Zheng K, Fang S, Huang K, Chu C, Zhuang J, et al. B7H3 targeting gold nanocage pH-sensitive conjugates for precise and synergistic chemo-photothermal therapy against NSCLC. J Nanobiotechnol. (2023) 21:378. doi: 10.1186/s12951-023-02078-9
62. Kasten BB, Arend RC, Katre AA, Kim H, Fan J, Ferrone S, et al. B7-H3-targeted 212Pb radioimmunotherapy of ovarian cancer in preclinical models. Nucl Med Biol. (2017) 47:23–30. doi: 10.1016/j.nucmedbio.2017.01.003
63. Ma L, Luo L, Qiao H, Dong X, Pan S, Jiang H, et al. Complete eradication of hepatocellular carcinomas by combined vasostatin gene therapy and B7H3-mediated immunotherapy. J Hepatol. (2007) 46:98–106. doi: 10.1016/j.jhep.2006.07.031
Keywords: B7-H3, non-immune functions, tumor cells, tumor vasculature, cancer therapy
Citation: Wu S, Hu C, Hui K and Jiang X (2024) Non-immune functions of B7-H3: bridging tumor cells and the tumor vasculature. Front. Oncol. 14:1408051. doi: 10.3389/fonc.2024.1408051
Received: 27 March 2024; Accepted: 04 June 2024;
Published: 17 June 2024.
Edited by:
Nitesh Kumar, National Institute of Pharmaceutical Education and Research, IndiaCopyright © 2024 Wu, Hu, Hui and Jiang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xiaodong Jiang, anhkcGFwZXJAMTYzLmNvbQ==; Kaiyuan Hui, a3lodWkxOTg3QDE2My5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.