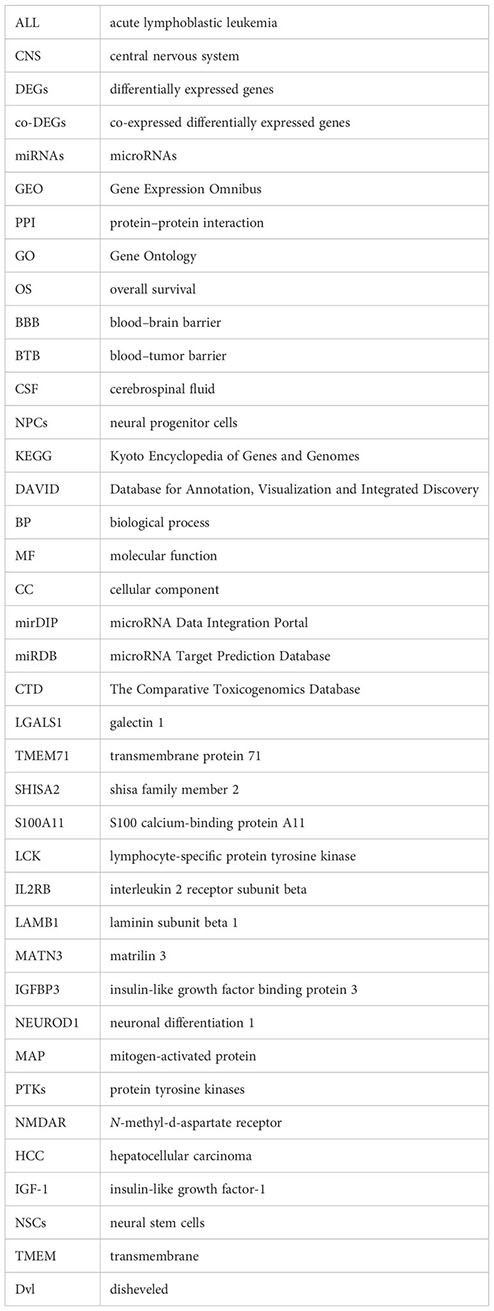
 Ziping Li1
Ziping Li1 Zhi Guo
Zhi Guo Wei Liu
Wei Liu Hao Zhou
Hao Zhou
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Oncol., 04 January 2024
Sec. Hematologic Malignancies
Volume 13 - 2023 | https://doi.org/10.3389/fonc.2023.1331802
This article is part of the Research TopicAcute Leukemias: Molecular Characterization, Leukemia-Initiating Cells, and Influence of the Microenvironment Vol. IIView all 9 articles
Background: Acute lymphoblastic leukemia (ALL) is prone to metastasize to the central nervous system (CNS), which is an important cause of poor treatment outcomes and unfavorable prognosis. However, the pathogenesis of CNS metastasis of ALL cells has not been fully illuminated. Recent reports have shed some light on the correlation between neural mechanisms and ALL CNS metastasis. These progressions prompt us to study the relationship between ALL central nervous system metastasis and neuronal development, exploring potential biomarkers and therapeutic targets of CNS metastasis.
Materials and methods: ALL central nervous system metastasis- and neuronal development-related differentially expressed genes (DEGs) were identified by analyzing gene expression datasets GSE60926 and GSE13715. Target prediction and network analysis methods were applied to assess protein–protein interaction networks. Gene Ontology (GO) terms and pathway enrichment for DEGs were assessed. Co-expressed differentially expressed genes (co-DEGs) coupled with corresponding predicted microRNAs (miRNAs) were studied as well. Reverse transcription–polymerase chain reaction (RT–PCR) and flow cytometry were employed for the validation of key co-DEGs in primary ALL cells. Furthermore, ALL cells were treated with a vascular endothelial growth factor (VEGF) inhibitor to block neuronal development and assess changes in the co-DEGs.
Results: We identified 216, 208, and 204 DEGs in ALL CNS metastasis specimens and neuronal development samples (GSE60926 and GSE13715). CD2, CD3G, CD3D, and LCK may be implicated in ALL CNS metastasis. LAMB1, MATN3, IGFBP3, LGALS1, and NEUROD1 may be associated with neuronal development. Specifically, four co-DEGs (LGALS1, TMEM71, SHISA2, and S100A11) may link ALL central nervous system metastasis and neuronal development process. The miRNAs for each co-DEG could be potential biomarkers or therapeutic targets for ALL central nervous system metastasis, especially hsa-miR-22-3p, hsa-miR-548t-5p, and hsa-miR-6134. Additionally, four co-DEGs (LGALS1, TMEM71, SHISA2, and S100A11) were validated in CNS-infiltrated ALL cells. The VEGF inhibitor demonstrated a suppressive effect on mRNA and protein expression of key co-DEGs.
Conclusion: The bioinformatic survey and key gene validation suggest a possible correlation between ALL CNS metastasis and the neuronal development process. Simulating the neuronal development process might be a possible strategy for CNS metastasis in ALL. LGALS1, TMEM71, SHISA2, and S100A11 genes are promising and novel biomarkers and targets in ALL CNS metastasis.
Acute lymphoblastic leukemia (ALL) is a malignancy that impacts the lymphoid lineage of hematopoietic cells and is distinguished by the excessive proliferation of immature lymphocytes (1). Specifically, ALL exhibits a notable tendency toward metastasizing to the central nervous system (CNS) (2). In spite of the advancements achieved in the management of ALL, individuals with unfavorable prognoses exhibit a higher incidence of CNS involvement. Moreover, the occurrence of late CNS relapse is often associated with a fatal outcome, thereby presenting a substantial obstacle in the attempt for a cure for these patients. The prognosis for adult patients who encounter CNS relapse is highly unfavorable, with a median overall survival (OS) of 6 months and a projected 5-year OS of zero (3). Additionally, even though routine diagnostic methods often fail to detect the presence of CNS involvement, early autopsy studies have already demonstrated that half of the patients suffer from submicroscopic CNS disease (4), underscoring the necessity for a suitable approach to investigate the entry of leukemia cells into the CNS.
At the time of diagnosis, the central nervous system is frequently affected as an extramedullary site in all cases. The CNS acts as a sanctuary site, contributing to the occurrence of relapse (5). The restricted permeability of the blood–brain barrier (BBB) and the existence of a blood–tumor barrier (BTB) limit the accessibility of conventional medications to the central nervous system, leading to resistance to treatment and a poor prognosis. Significant progress has been made in enhancing the survival rate of ALL patients through the implementation of intrathecal chemotherapy, high-dose systemic chemotherapy, and/or cranial radiotherapy, specifically targeting the central nervous system. Despite their lack of specificity, CNS-targeted therapies can lead to adverse effects such as seizures, leukoencephalopathy, and delayed neurodevelopmental effects (6).
The limitations of current diagnostic methods and CNS-directed therapy prompt us to uncover mechanisms by which ALL cells infiltrate the CNS. Recent reports have shed some light on the correlation between neural mechanisms and ALL CNS metastasis (2). In this study, we identified co-expressed differentially expressed genes (co-DEGs) of ALL central nervous system metastasis and neuronal development and elucidated molecular mechanisms and pathology of ALL central nervous system metastasis-related DEGs and neuronal development-related DEGs. Furthermore, we provided a bioinformatic analysis of DEGs and predicted microRNAs (miRNAs), which regulate the co-expression of DEGs. Finally, we validated four key DEGs in primary ALL cells and provided a potential and novel strategy against CNS metastasis.
After written informed consent was obtained, leukemic cells were gathered from ALL patients with CNS metastasis. By separating mononuclear cells on a Ficoll density gradient (Sigma-Aldrich, St. Louis, MO, USA), cell suspensions containing >80% leukemic cells were produced. The present study was approved by the Ethics Committee of Tongji Medical College, Huazhong University of Science and Technology, and performed in strict compliance with the Declaration of Helsinki.
We screened Gene Expression Omnibus (GEO) datasets GSE60926 and GSE13715 according to the following inclusion criteria: 1) Homo sapiens ALL central nervous system metastasis specimens, 2) expression profiling by array, 3) performed on the GPL570 platform ([HGU133_Plus_2] Affymetrix Human Genome U133 Plus 2.0 Array), and 4) ≥8 samples and downloaded from GEO (7). The GSE60926 dataset includes 20 specimens from bone marrow (BM) and eight specimens from cerebrospinal fluid (CSF) of ALL patients. The GSE13715 dataset contains four and three specimens from neural progenitor cells (NPCs) before differentiation and four and three specimens after differentiation cultured on 2D substrates and 3D porous polystyrene scaffolds, respectively. We used the identified two datasets to analyze the relationship between ALL central nervous system metastasis and neuronal development.
We identified DEGs between BM samples and CSF in ALL, with the DEGs about neuronal differentiation, from R using the packages “affy”, “affyPLM”, and “limma” (8). Gene expression values of the |log2 FC| > 2 and p-value <0.05 were significant. Additionally, we calculated and made Venn diagrams for co-DEGs for ALL central nervous system metastasis- and neuronal development-related DEGs.
Protein–protein interaction (PPI) networks of ALL central nervous system metastasis- and neuronal development-related DEGs were analyzed using the STRING software (9) and obtained the analytic results with a confidence score >0.7. Subsequently, the results were performed using the Cytoscape software (10) to visualize molecular interaction network node degrees.
Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analyses of ALL central nervous system metastasis- and neuronal development-related DEGs were carried out using the Database for Annotation, Visualization and Integrated Discovery (DAVID) (11) and Reactome databases (12). GO terms and KEGG pathways of biological functions were considered to be significantly enriched if p < 0.05. At the same time, the different bio functions of ALL central nervous system metastasis- and neuronal development-related DEGs in biological process (BP), molecular function (MF), and cellular component (CC) were presented using the DAVID database. Moreover, the AmiGO2 database (13) was used to analyze the GO terms for the selected co-DEGs to verify the accuracy and annotate bio-functions of identified co-DEGs.
We applied the online prediction tools microRNA Data Integration Portal (mirDIP) (14), microRNA Target Prediction Database (miRDB) (15), TargetScanHuman (16), and DIANA TOOLS (17) to predict potential microRNAs that could target the co-DEGs. We obtained the candidate microRNAs according to higher predicted scores for ≥3 prediction tools for each co-DEG.
The Comparative Toxicogenomics Database (CTD) (http://ctdbase.org/) (18) provides manually curated information about chemical–gene/protein interactions, chemical–disease, and gene–disease relationships, which was used to find gene–disease interactions to generate expanded networks and predict novel associations. These data were used to analyze relationships between the co-DEGs and hematological or nervous system diseases.
Primary ALL cells separated using density gradient centrifugation with Ficoll-Hypaque were cultured in 24-well plates at 2 × 106 cells/ml at 37°C with 5% CO2 in the following: RPMI 10 (control medium) or vascular endothelial growth factor (VEGF) inhibitor thalidomide (40 μM, Sigma-Aldrich).
All samples were detected using the flow cytometry analyzer BD FACSCanto II (Becton Dickinson, Franklin Lakes, NJ, USA). Measured data were analyzed using the BD CellQuest software (Becton Dickinson, USA). Anti-LGALS1 (ab228184) and anti-S100A11 (ab236123) for surface staining of leukemia cells were obtained from Abcam (Cambridge, UK). The procedure was as previously described (19).
For reverse transcription–polymerase chain reaction (RT–PCR) experiments, RNA was isolated using TRIzol reagent (Invitrogen, Carlsbad, CA, USA). cDNA was produced from 1 μg RNA using the First Strand cDNA synthesis kit (TaKaRa, Maebashi, Japan). PCR was then performed on 0.5 μl cDNA. The procedures were conducted according to the instructions by the manufacturers. The PCR primers (5′→3′) used were LGALS1 forward (Fw): CTGTGCCTGCACTTCAACC; TMEM71 Fw: GATGTCAACACCAGTAGCAAGT; SHISA2 Fw: GGAGACCATCCCCATGATCC; S100A11 Fw: CTGAGCGGTGCATCGAGTC; LGALS1 reverse (Rv): CATCTGGCAGCTTGACGGT; TMEM71 Rv: TGGTAACCATCCAGGGAATCA; SHISA2 Rv: AGCACAGAGAAATTCGTGGGC; S100A11 Rv: TGTGAAGGCAGCTAGTTCTGTA.
Statistical analyses were conducted using SPSS 20.0 (IBM Corp., Armonk, NY, USA) and GraphPad Prism 8.0.2 (GraphPad Software, San Diego, CA, USA). Student’s t-test and analysis of variance were applied to compare continuous variables between the two groups or among multiple groups. Categorical variables were analyzed using the chi-square test, continuity correction test, or Fisher’s exact test. Statistical significance was set at p-values <0.05.
We identified 216 DEGs in the GSE60926 dataset, and we identified a total of 208 and 204 DEGs in the 2D substrate specimens and 3D porous polystyrene scaffold specimens in the GSE13157 dataset, respectively. Furthermore, we defined 82 co-expressed DEGs in the 2D specimens and 3D specimens mentioned above as the neuronal development DEGs. We conducted a cluster analysis of ALL central nervous system metastasis-related DEGs in relation to immune response and cell signaling, endoderm formation, and T-cell co-stimulation for gene expression, and these data appear in Figure 1 and Supplementary Table S1. Both Figure 2 and Supplementary Table S2 show the gene expression value in relation to the neuronal cell body, axon, regulation of cell proliferation, and nervous system development above the neuronal development-related DEGs.
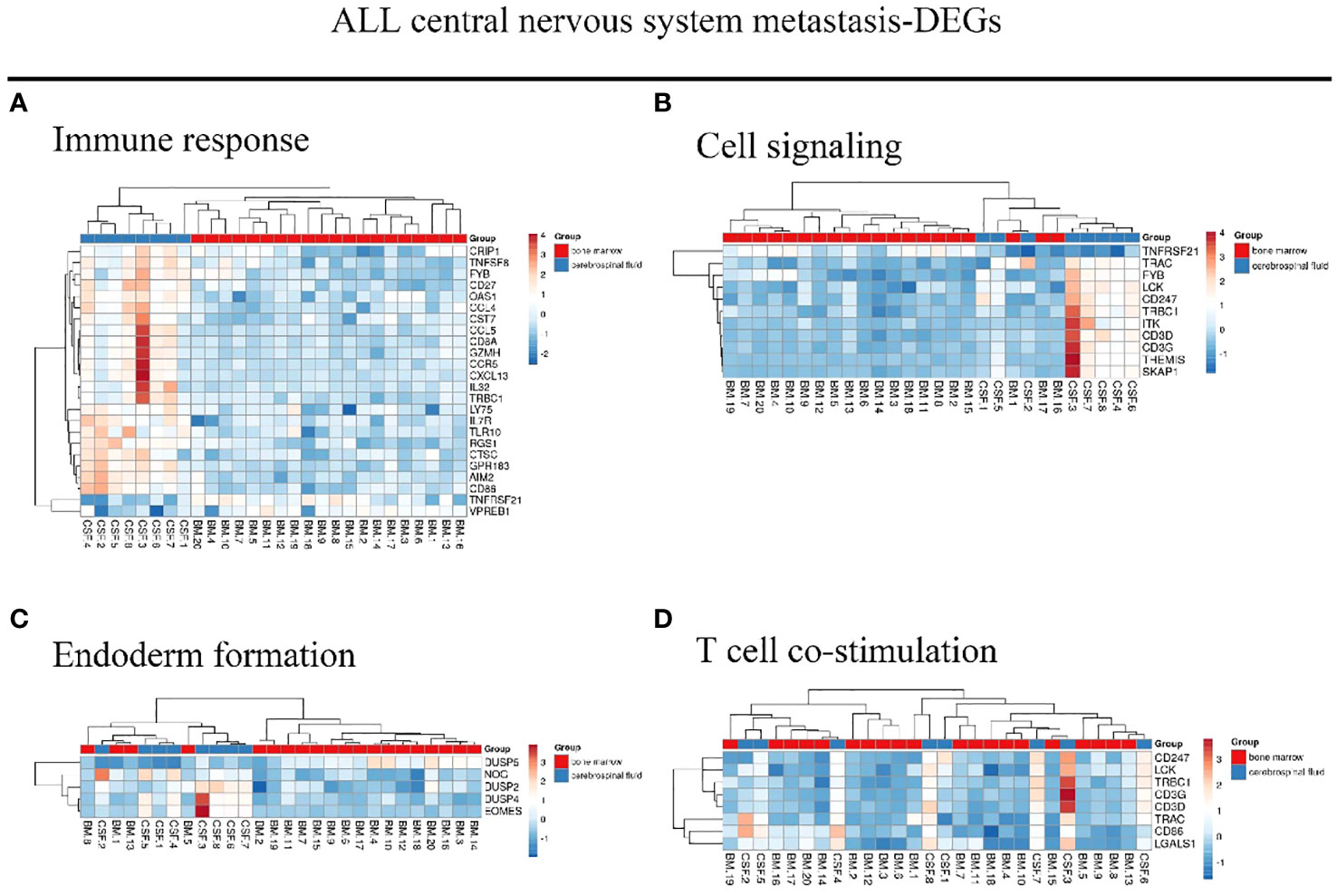
Figure 1 Hierarchical clustering analysis of ALL central nervous system metastasis-related differentially expressed genes. (A–D) Results of hierarchical clustering analysis for DEG expression in relation to immune response, cell signaling, endoderm formation, and T-cell co-stimulation. Red, greater expression. Blue, less expression. BM, bone marrow; CSF, cerebrospinal fluid; ALL, acute lymphoblastic leukemia; DEG, differentially expressed gene.
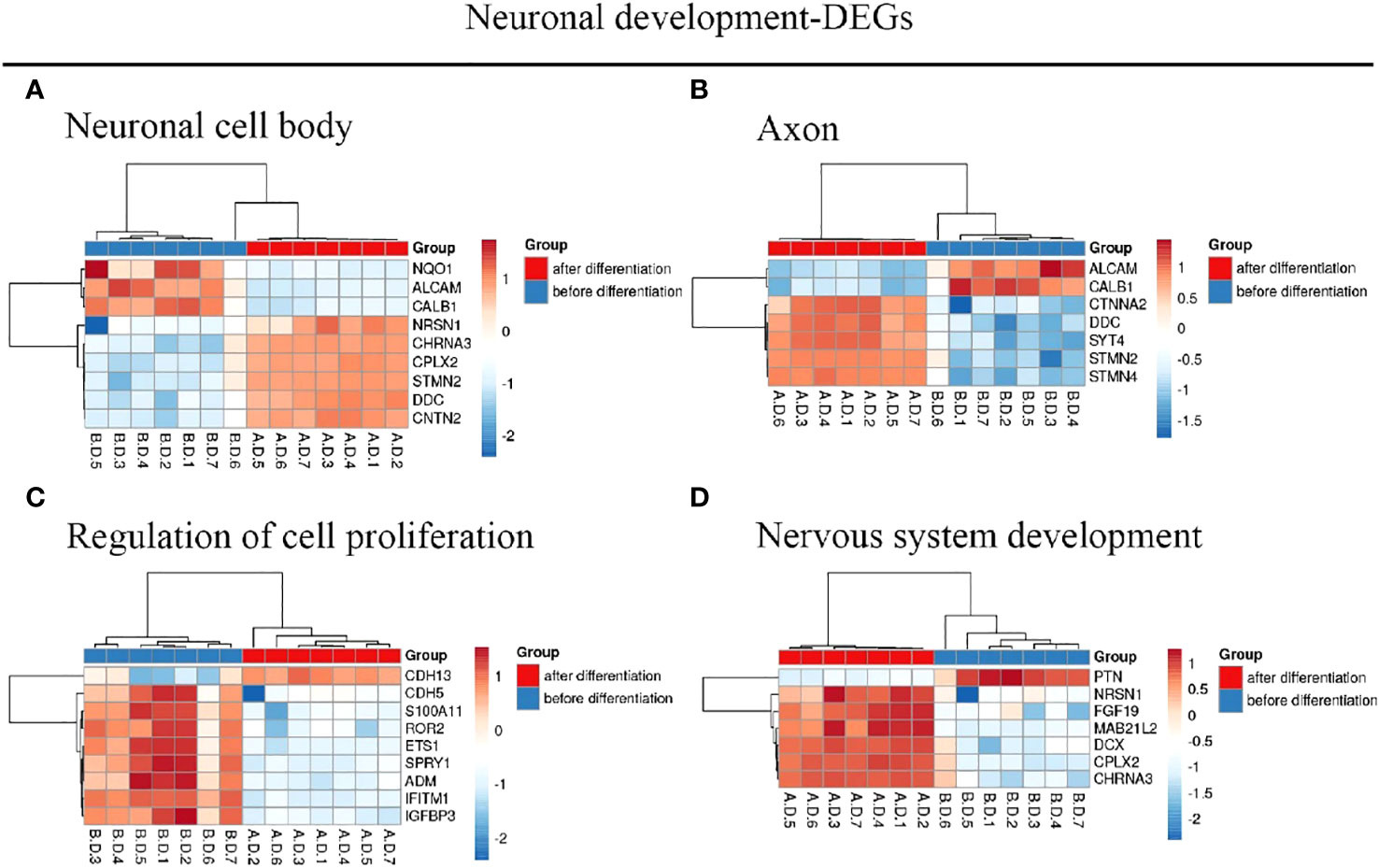
Figure 2 Hierarchical clustering analysis of neuronal development-related DEGs. (A–D) Results of hierarchical clustering analysis for DEG expression in relation to neuronal cell body, axon, regulation of cell proliferation, and nervous system development. Red, greater expression. Blue, less expression. AD, after differentiation; BD, before differentiation; DEGs, differentially expressed genes.
Figure 3C depicts the differentially expressed genes associated with both central nervous system metastasis and neuronal development, as well as the genes that are co-expressed with them. Interestingly, four co-expressed DEGs, including galectin 1 (LGALS1), transmembrane protein 71 (TMEM71), shisa family member 2 (SHISA2), and S100 calcium-binding protein A11 (S100A11), were observed. The AmiGO database was used to confirm GO term enrichment related to biological processes, molecular functions, and cellular components, and co-DEGs were associated with various processes as indicated in Table 1.
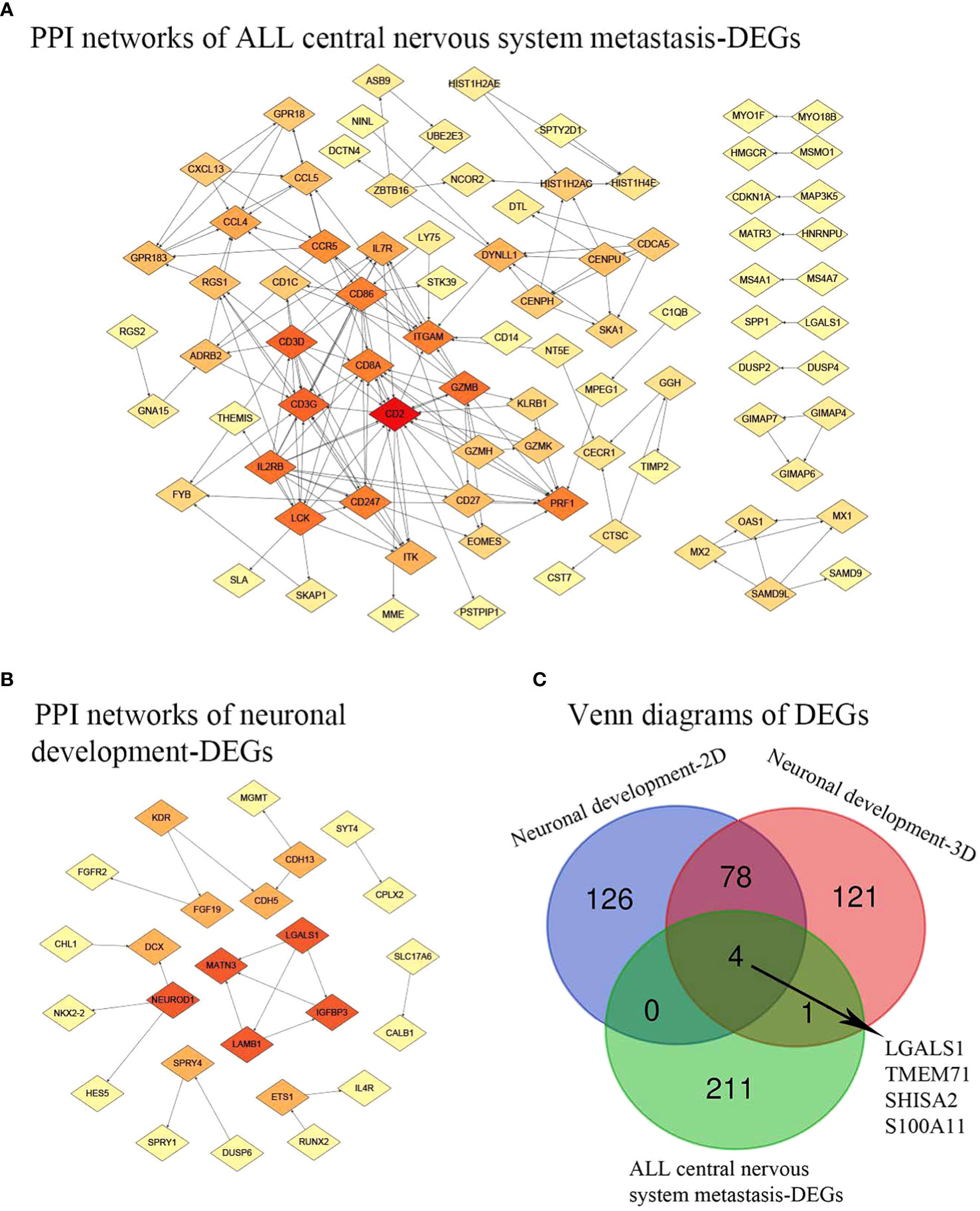
Figure 3 PPI network and Venn diagrams. (A) PPI network of ALL central nervous system metastasis-related DEGs. (B) PPI network of neuronal development-related DEGs. Red, greater degree. Yellow, lesser degree. (C) Venn diagrams of DEGs. Neuronal development-2D, differential genes found in specimens cultured on 2D substrates; Neuronal development-3D, differential genes found in specimens on 3D porous polystyrene scaffolds; PPI, protein–protein interaction; ALL, acute lymphoblastic leukemia; DEGs, differentially expressed genes.
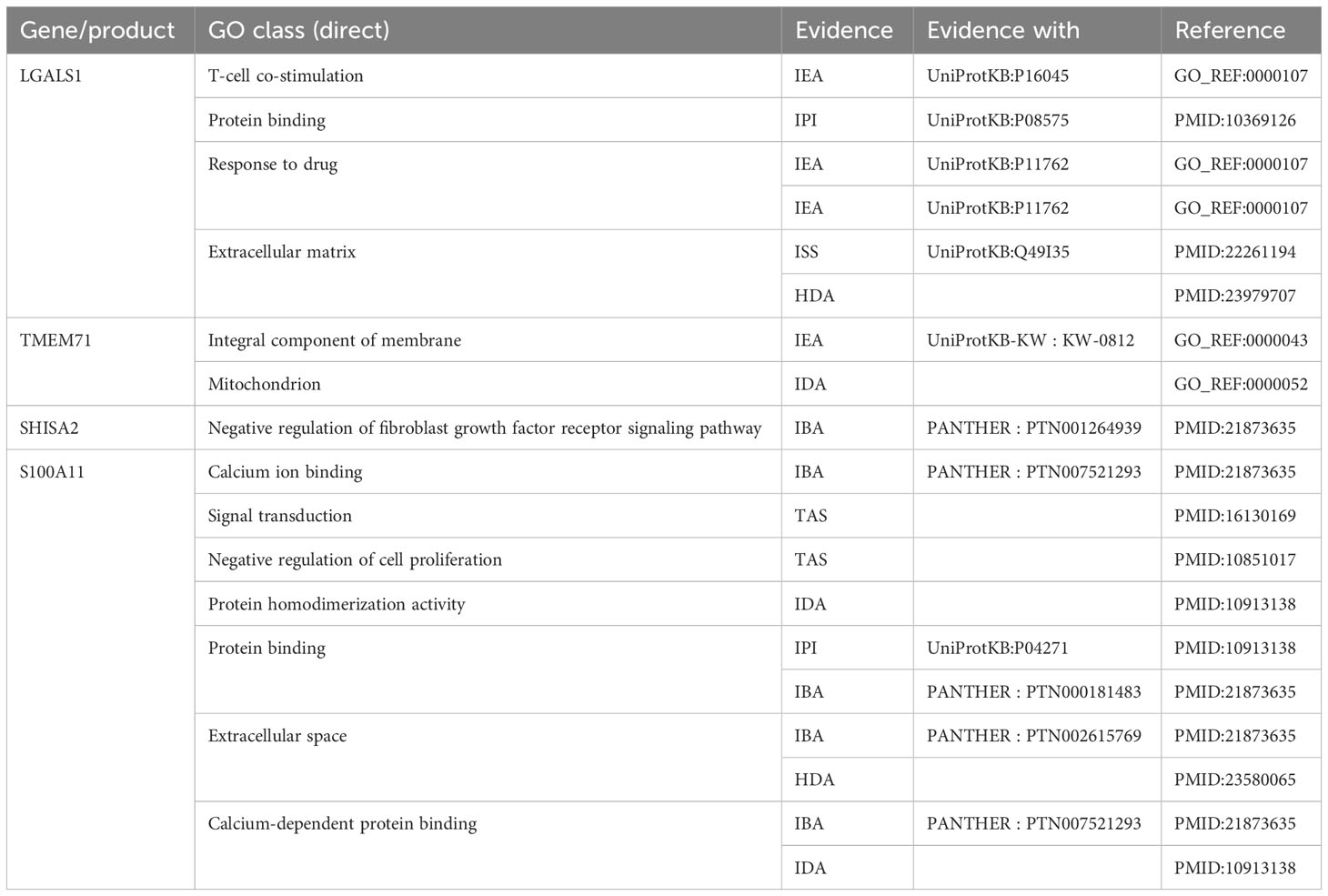
Table 1 The Gene Ontology (GO) term enrichment for the co-expressed genes of the acute lymphoblastic leukemia (ALL) central nervous system metastasis and neuronal development.
A total of 195 nodes were identified from the PPI network of DEGs associated with central nervous system metastasis in acute lymphoblastic leukemia. Additionally, 82 nodes were identified from the PPI network of DEGs associated with neuronal development. These findings are presented in Figure 3. In the context of central nervous system metastasis in ALL, the following five hub nodes were identified as significant: CD2 (degree = 19), CD3G (degree = 13), CD3D (degree = 13), lymphocyte-specific protein tyrosine kinase (LCK; degree = 12), and interleukin 2 receptor subunit beta (IL2RB; degree = 12). These genes are considered to play a crucial role as hub genes in the aforementioned condition. Additionally, the hub genes, involved in laminin subunit beta 1 (LAMB1; degree = 3), matrilin 3 (MATN3; degree = 3), insulin-like growth factor binding protein 3 (IGFBP3; degree = 3), galectin 1 (LGALS1; degree = 3), and neuronal differentiation 1 (NEUROD1; degree = 3), were demonstrated in neuronal development-related DEGs with a higher degree.
Using the DAVID database, the top 3 GO terms related biological processes among ALL central nervous system metastasis-related DEGs were primarily associated with immune response (Fold Enrichment: 5.60; p-value: 4.60E−11), T-cell receptor signaling pathway (Fold Enrichment: 7.30; p-value: 2.66E−06), and endoderm formation (Fold Enrichment: 40.92; p-value: 4.71E−06). There was a significant correlation in T-cell receptor complex (Fold Enrichment: 27.36; p-value: 2.76E−05), external side of the plasma membrane (Fold Enrichment: 4.62; p-value: 3.22E−04), and alpha–beta T-cell receptor complex (Fold Enrichment: 59.10; p-value: 9.94E−04) in relation to cellular components. In addition, the terms related to molecular functions were mainly involved in mitogen-activated protein (MAP) kinase tyrosine/serine/threonine phosphatase activity (Fold Enrichment: 31.10; p-value: 2.48E−04), protein homodimerization activity (Fold Enrichment: 2.35; p-value: 0.002), and proteoglycan binding (Fold Enrichment: 27.57; p-value: 0.004). With respect to neuronal development-related DEGs, the biological processes in terms of negative regulation of cell proliferation (Fold Enrichment: 4.96; p-value: 4.15E−04), regulation of fibroblast growth factor receptor signaling pathway (Fold Enrichment: 81.78; p-value: 5.56E−04), and single organismal cell–cell adhesion (Fold Enrichment: 10.80; p-value: 0.001) were significantly enriched. Additionally, the top 3 GO terms related to cellular components were associated with neuronal cell body (Fold Enrichment: 6.59; p-value: 5.98E−05), axon (Fold Enrichment: 7.27; p-value: 3.78E−04), and neuron projection (Fold Enrichment: 5.84; p-value: 0.003). Similarly, the terms of heparin binding-related (Fold Enrichment: 8.33; p-value: 7.27E−04), transmembrane receptor protein tyrosine kinase activity-related (Fold Enrichment: 17.54; p-value: 0.012), and calcium ion binding-related (Fold Enrichment: 2.79; p-value: 0.014) molecular functions were primarily enriched (as shown in Figures 4A, B; Supplementary Table S3).
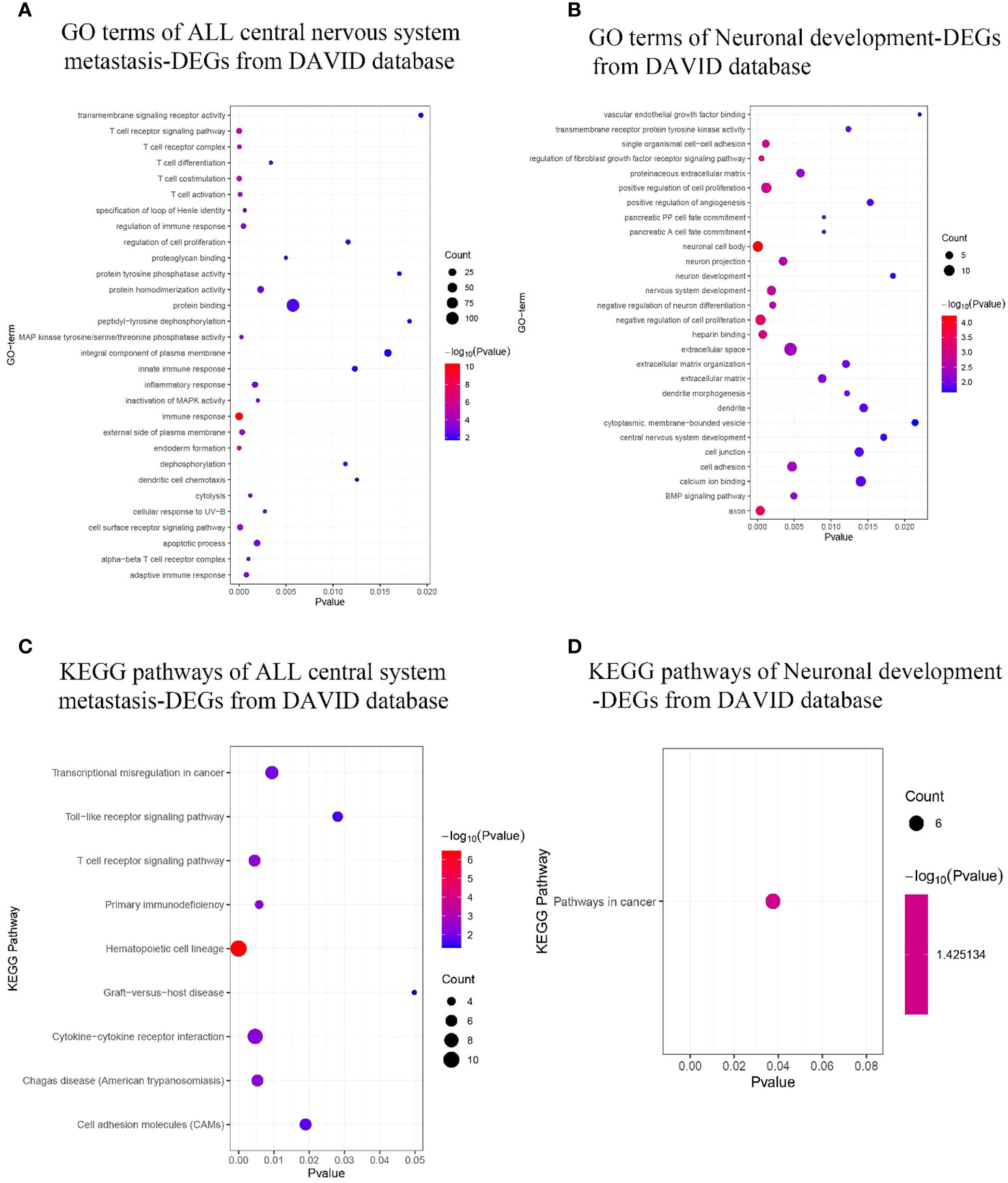
Figure 4 GO terms and KEGG pathway enrichment. (A, B) ALL central nervous system metastasis- and neuronal development-related GO term enrichment for DEGs. (C, D) KEGG pathway of ALL central nervous system metastasis- and neuronal development-related DEGs. Dot sizes represent counts of enriched DEGs, and dot colors represent negative Log10-p values. GO, Gene Ontology; KEGG, Kyoto Encyclopedia of Genes and Genomes; ALL, acute lymphoblastic leukemia; DEGs, differentially expressed genes.
KEGG pathway analysis data from the DAVID database are shown in Figures 4C, D. The results suggest that the ALL central nervous system metastasis-related DEGs were mainly enriched in pathways of hematopoietic cell lineage (p-value: 3.46E−07), T-cell receptor signaling pathway (p-value: 0.005), and cytokine–cytokine receptor interaction (p-value: 0.005). However, there is only one KEGG term pathway in cancer (p-value: 0.04) that was significantly enriched in neuronal development-related DEGs (as shown in Figure 4; Supplementary Table S4). Additionally, KEGG pathway enrichments from the Reactome database also appeared in Supplementary Table S4. The CTD showed that co-DEGs targeted several hematological diseases or nervous system diseases, and these data appear in Figure 5 and Supplementary Table S5.
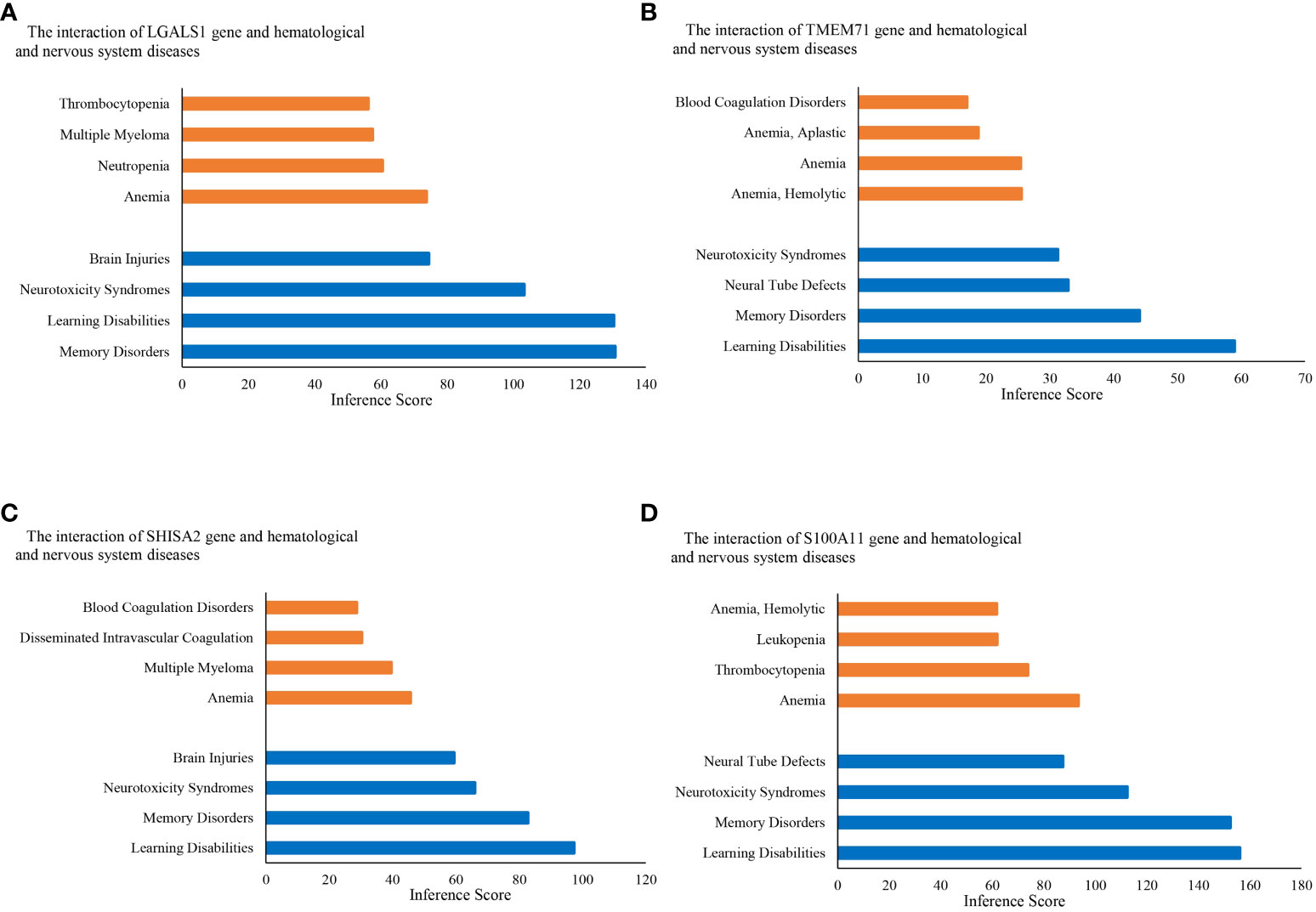
Figure 5 Relationship to hematological diseases and nervous system diseases related to co-expressed genes based on the CTD. Relationship between diseases and co-expressed genes (A–D). CTD, The Comparative Toxicogenomics Database.
The application of bioinformatic tools including mirDIP, miRDB, TargetScan, and DIANA enabled the identification of the top 5 miRNAs that target each co-DEG associated with central nervous system metastasis and neuronal development in ALL. The results of this prediction analysis are presented in Table 2. These data provide insights into the association between predicted miRNAs and central nervous system metastasis in relation to neuronal development in patients with ALL.
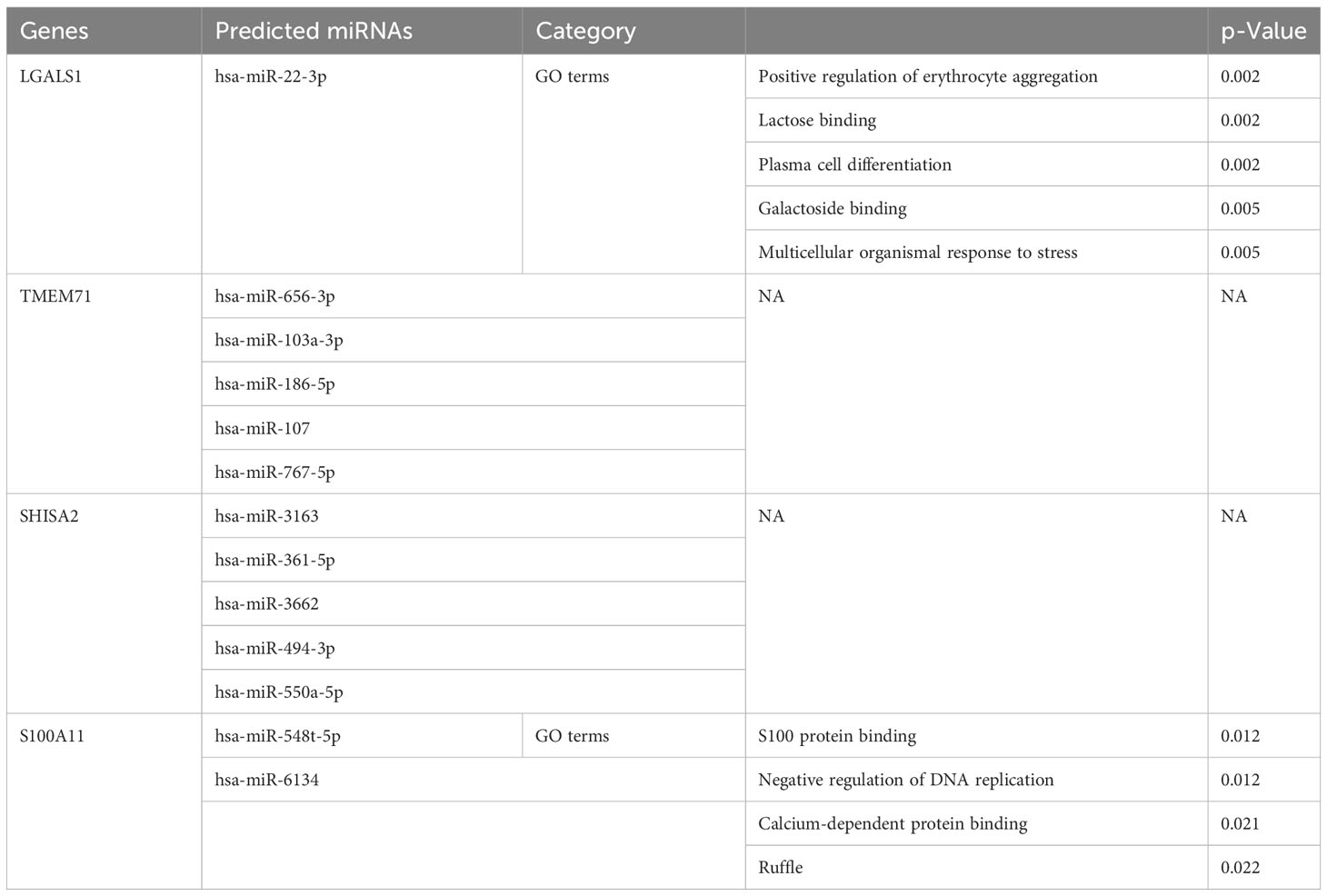
Table 2 The Gene Ontology (GO) terms among predicted microRNAs (miRNAs) and co-expressed differentially expressed genes (co-DEGs).
To validate the in silico finding, we applied RT–PCR to test the four co-DEGs (LGALS1, TMEM71, SHISA2, and S100A11) in CNS-metastasized ALL cells. The demographic and clinical characteristics of the ALL patients are presented in Supplementary Table 6. All four co-DEGs were significantly upregulated in CSF-derived ALL cells, compared with bone marrow-derived ALL cells (Figure 6A). To clarify the changes in protein level, we focused on two membrane-bound proteins (LGALS1 and S100A11) and assessed them using flow cytometry. LGALS1 and S100A11 proteins were increased in CSF-derived ALL cells, compared with bone marrow-derived ALL cells (Figures 6B, C). Furthermore, we used a VEGF inhibitor (thalidomide, 40 μM), which can hamper the neuronal development process (19), to treat primary cultured ALL cells. We found that LGALS1 and S100A11 were decreased after thalidomide treatment at both mRNA and protein levels (Figures 6D–F). The analysis of GEO data (GSE5820) showed that LGALS1 increased in poor clinical outcome patients (Supplementary Figure 1). These data verified four co-DEGs in CNS-metastasized ALL cells and implied a potential strategy for controlling CNS metastasis.
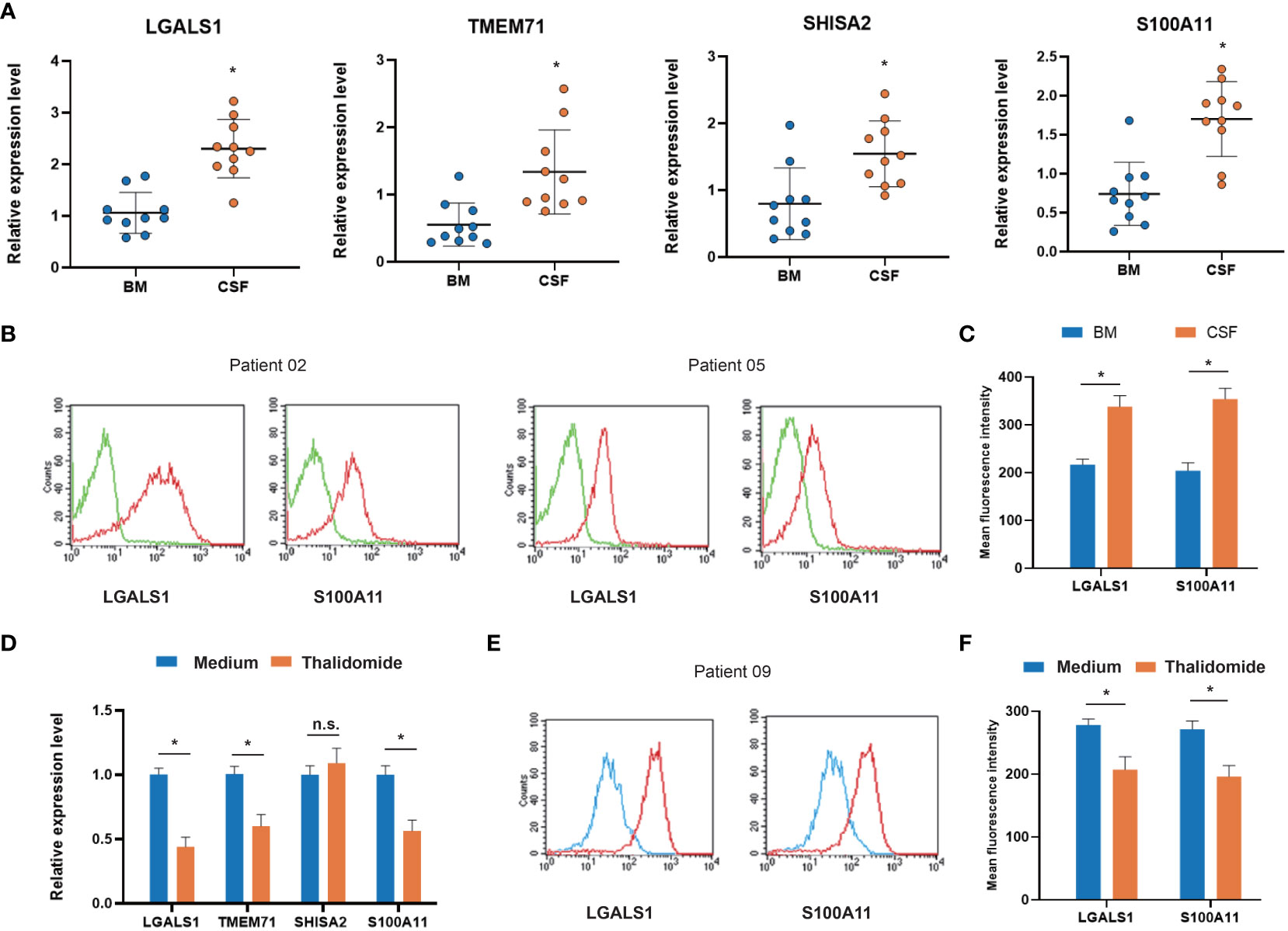
Figure 6 Expression of co-DEGs in primary ALL cells from CNS metastasis patients. (A) RT–PCR analysis of co-DEGs comparing bone marrow-derived and cerebrospinal fluid-derived ALL cells (n = 10). (B) Representative flow cytometry images of membrane-bound co-DEGs in bone marrow-derived and cerebrospinal fluid-derived ALL cells (n = 10; green, negative control). (C) Box-plot analysis of relative fluorescence intensity (RFI) of membrane-bound co-DEGs. (D) RT–PCR analysis of co-DEG changes in primary ALL cells treated with VEGF inhibitor thalidomide (n = 5). (E) Representative flow cytometry images of membrane-bound co-DEGs in primary ALL cells treated with VEGF inhibitor (n = 5; blue, thalidomide treated; red, medium control). (F) Box-plot analysis of RFI of co-DEGs after thalidomide treatment (* p < 0.05). BM, bone marrow; CSF, cerebrospinal fluid; VEGF, vascular endothelial growth factor; co-DEGs, co-expressed differentially expressed genes; ALL, acute lymphoblastic leukemia; CNS, central nervous system; RT–PCR, reverse transcription–polymerase chain reaction.
ALL has a marked propensity to metastasize to the central nervous system (19). The involvement of the central nervous system has long been recognized as a significant barrier to achieving successful long-term treatment outcomes and has been identified as a prominent factor contributing to mortality rates in the context of a more regulated systemic illness (20). Nevertheless, the mechanism by which leukemia cells invade the central nervous system remains uncertain. Various mechanisms have been proposed based on several lines of clinical or experimental evidence. Recent research has revealed that leukemia cells have the ability to migrate into the CNS by means of vessels, which are recognized for their role in guiding the pathfinding of neuronal progenitor cells within the CNS (19). These reports proposed a possible and functional relationship between neural mechanisms and ALL CNS metastasis. Based on these findings, we hypothesized that there might be a relationship between ALL CNS metastasis and neuronal development.
In the present study, we collected the gene expression profiles of ALL CNS metastasis GSE60926 and neuronal development GSE13715 from GEO. We identified a total of 216 genes differentially expressed between the primary site of bone marrow and the cerebrospinal fluid, as well as 82 differentially expressed genes before and after the differentiation of neurocytes. We identified four co-expression genes—LGALS1, TMEM71, SHISA2, and S100A11—between ALL CNS metastasis and neuronal development. Additionally, we constructed a PPI network and found hub genes between ALL central nervous system metastasis- and neuronal development-related DEGs. Subsequently, we performed GO term and KEGG pathway enrichment analyses for DEGs. Meanwhile, we predicted non-coding RNAs that may regulate the co-expression of DEGs. Finally, we identified hematologic and neurological diseases associated with co-expressed genes in the CTD.
The hub gene that related to neuronal development was found among ALL central nervous system metastasis-related DEGs in this study through the PPI network constructed. As we know, a variety of protein tyrosine kinases (PTKs) are present in the central nervous system of mammals, many of which play a role in the formation of neurons and the alteration of synapses (21). LCK is one of the five members of the Src family of non-receptor protein tyrosine kinases (22). Martinez’s group discovered that LCK can phosphorylate δ-catenin and bind to δ-catenin through its polyproline tract to regulate neurite growth (23). The N‐methyl‐d‐aspartate receptor (NMDAR), a ligand‐gated ion channel, plays a crucial role in controlling synaptic plasticity, brain development, and excitotoxicity within the central nervous system (24). Le’s study indicated that LCK may serve as an upstream to have impacts on NMDAR tyrosine phosphorylation, which mediated processes such as learning and memory and hippocampal neuron migration (25). Meanwhile, as a non-receptor tyrosine protein kinase, LCK is crucial for the selection and development of developing T cells, as well as the functioning of mature T cells. It has been documented that it is associated with the onset of acute lymphoblastic leukemia and appears to facilitate the metastatic characteristics of leukemic cells (26, 27). These findings imply that LCK may act as a bio-functional mediator to correlate neuronal development and ALL central nervous system metastasis.
Furthermore, we observed the presence of hub genes that are directly or indirectly involved in the regulation of hematologic malignant diseases within the differentially expressed genes associated with neuronal development. Laminins play a crucial role in a wide range of physiological and pathological processes, such as contributing to the formation of basement membranes and the growth of neurites and facilitating cell adhesion, migration, proliferation, and angiogenesis in cancer cells (28). The connection between cancer cells and laminins is essential for tumor spread and metastasis. The interaction between invading tumor cells and laminins leads to an increased potential for metastasis (29). Research has indicated that LAMB1 may be a viable biomarker for certain types of cancer (30, 31). Pan-cancer analysis showed the correlation between abnormal expression of LAMB1 in various cancers and clinical outcomes (32). LAMB1 plays a role in the movement and infiltration of cells into the adjacent extracellular matrix in prostate cancer (30). Moreover, hepatocellular carcinoma (HCC) exhibited an upregulation of LAMB1, which was associated with heightened tumor aggressiveness and unfavorable patient survival (33). Simultaneously, in the nervous system, it is believed that laminins play a role in several processes, including neurite outgrowth, neural survival, and synapse formation and function (34). Recent studies indicated that IGFBP3 could potentially inhibit tumor growth through induction of the tumor suppressor gene p53 (35). The subsequent occurrence of relapse in childhood leukemia was found to be inversely correlated with the rising levels of total IGFBP3 in a molecular mechanistic study (36). Likewise, multiple lines of evidence indicate that insulin-like growth factor-1 (IGF-1) is essential to normal brain growth and development (37). The regulatory action of IGF-1 is contingent upon the presence of IGFBP3, which has the ability to either impede or promote cellular proliferation. This outcome is determined by the expression of proteases that facilitate the release of IGF-1 from the IGF-1–IGFBP3 complex (38). Kalluri has demonstrated that IGFBP3 could regulate neural progenitor cell proliferation and reduce the amount of nestin in the neural progenitor cells, indicating its potential role in neuronal development (39). Galectins have been extensively linked to cancer, specifically controlling cell transformation, apoptosis, proliferation, migration, invasion, and angiogenesis (40), and LGALS1 has been previously documented to enhance the survival of hematological malignancies by directly targeting tumor cells (41). LGALS1 could promote the proliferation of adult neural stem cells (NSCs) found in several studies (42, 43). The role of hub genes LCK, LAMB1, IGFBP3, and LGALS1 in the pathogenesis and metastasis of acute lymphoblastic leukemia as well as in the development of the nervous system provides evidence to support our hypothesis that there may be a relationship between ALL central nervous system metastasis and neuronal development, which may be arise from locus mutations or gene variants.
The co-DEGs LGALS1, TMEM71, SHISA2, and S100A11 were identified in our study. LGALS1, a lectin that binds beta-galactoside and a wide array of complex carbohydrates, has a critical role in cell proliferation, cell differentiation, and ALL occurrence (41). TMEM71 is a member of an extensive gene family that encodes transmembrane (TMEM) proteins. In recent years, scholars have shown growing interest in the involvement of TMEMs in malignancies. For example, Wang et al. observed a significant correlation between elevated TMEM71 expression and reduced survival time among patients diagnosed with both glioma and glioblastoma (44). TMEM14A plays a role in the development and spread of ovarian cancer, TMEM88 has been documented to induce invasion and metastasis in breast cancer through its interaction with disheveled (Dvl) proteins, TMEM45A and TMEM71 facilitate the proliferation and invasion of glioma cells, and TMEM45B enhances the proliferation, migration, and invasion of gastric cancer cells through the JAK2/STAT3 signaling pathway (45). Nagano’s research demonstrated that SHISA2 is indispensable for the maturation of presomitic mesoderm cells (46). In a similar vein, there have been recent reports of its overexpression in high-grade prostate cancer cells and its involvement in the aggressive phenotype of prostate cancer (47). Research has demonstrated that S100A11 could be involved in movement, infiltration, and tubulin polymerization (48). Tumor metastasis has been linked to chromosomal rearrangements and modified gene expression (49). These co-DEGs play a role in both tumor metastasis and nervous system development, which further suggests that the occurrence of nervous system metastasis in acute lymphoblastic leukemia may be related to neuronal development.
MicroRNAs, a key component of the non-coding RNA family, are involved in multiple cellular functions. The development of miRNA delivery strategies for the treatment of cancer patients such as acute lymphoblastic leukemia was prompted by the emergence of the miRNA field. Cancer cells secrete miRNAs, and their abnormal expression is linked to tumor growth and advancement; these miRNAs are not only potential targets for treatment but also potential biomarkers for diagnosis and prognosis (50). We found that hsa-miR-22-3p, hsa-miR-548t-5p, and hsa-miR-6134 may be potential biomarkers of neuronal development-related ALL central nervous system metastasis. It is noteworthy that Saccomani’s team discovered that an increase in miR-22-3p expression in T-ALL cells can impede colony formation in a laboratory setting and the advancement of leukemia in a living organism (51). Li’s research showed that miR-22-3p had the ability to raise p-EGFR levels and lengthen neurites through the STAT3/GAP43 pathway (52). Additionally, miR-548t-5p and other non-coding RNAs could regulate cell proliferation and metastasis in multiple human cancers (53–55). As shown in Table 2, we revealed that miRNAs involved in hsa-miR-22-3p, hsa-miR-548t-5p, and hsa-miR-6134 are related to ALL central nervous system metastasis and neuronal development. These miRNAs may be considered potential biomarkers to develop novel diagnostic and therapeutic strategies in ALL central nervous system metastasis.
The CTD (https://ctdbase.org/) proposes to combine diverse data on chemical exposures and their biological effects. In the CTD, both the hematological diseases and nervous system diseases associated with the four co-DEGs are identified, which helps us develop hypotheses about mechanisms underlying ALL central nervous system metastasis related to neuronal development. In a translational view, we have applied a non-specific VEGF inhibitor (thalidomide) to evaluate whether interfering with the neuronal development process could hamper ALL cells from metastasizing to the CNS. The results showed a significant decrease in several neuronal development genes, which suggests a promising and novel therapy against CNS metastasis of ALL.
The present study provides several candidates for further research on mechanisms of ALL central nervous system metastasis pathogenesis. Notably, our results suggest that CNS infiltration in ALL may be caused by leukemia cells mimicking the process of neuronal development. Additionally, the co-DEG expression profile could be used to help diagnose, differentiate, predict, and design treatments for CNS metastasis in ALL. Additional research is necessary to validate our discoveries and clarify the mechanisms by which these potential biomarkers operate in the metastasis of the entire central nervous system.
The interpretation of our findings should be addressed with caution due to certain limitations. First, being largely limited to in silico evaluation, our findings should be verified and extended in laboratory experiments. Second, given that the microarray analysis was conducted using gene expression rather than protein expression, the biomarkers used in this study should be regarded as genes rather than proteins. We may be able to validate our results to some extent by conducting larger and more prospective clinical studies.
Hub genes CD2, CD3G, CD3D, and LCK may be associated with ALL central nervous system metastasis; LAMB1, MATN3, IGFBP3, LGALS1, and NEUROD1 may be associated with neuronal development. Additionally, co-DEGs of LGALS1, TMEM71, SHISA2, and S100A11 link ALL central nervous system metastasis and neuronal development. Additionally, the miRNAs for each co-DEG may be potential biomarkers or therapeutic targets for ALL central nervous system metastasis, especially hsa-miR-22-3p, hsa-miR-548t-5p, and hsa-miR-6134. Thus, we hypothesized that there may be an association between ALL central nervous system metastasis and neuronal development and that expression of LGALS1, TMEM71, SHISA2, and S100A11 genes that favor ALL central nervous system metastasis is related to neuronal development.
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
The studies involving humans were approved by Ethics Committee of Tongji Medical College, Huazhong University of Science and Technology. The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study. Written informed consent was obtained from the individual(s) for the publication of any potentially identifiable images or data included in this article.
ZL and ZG: data analysis and writing the manuscript. HX and XC: experiments execution and data analysis. HZ and WL: designing, project administration, supervision. All authors reviewed the manuscript.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by the Clinical Innovation Funds of Union Hospital (No. 20190125), Natural Science Foundation of Hubei Province of China (No. 2022CFB069), and National Natural Science Foundation of China (No. 82272182).
We thank all the patients and healthcare workers who consented to take part in this study. We thank Prof. Wanxin Chen for his critical comments.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fonc.2023.1331802/full#supplementary-material
1. Terwilliger T, Abdul-Hay M. Acute lymphoblastic leukemia: a comprehensive review and 2017 update. Blood Cancer J (2017) 7(6):e577. doi: 10.1038/bcj.2017.53
2. Lenk L, Alsadeq A, Schewe DM. Involvement of the central nervous system in acute lymphoblastic leukemia: opinions on molecular mechanisms and clinical implications based on recent data. Cancer metastasis Rev (2020) 39(1):173–87. doi: 10.1007/s10555-020-09848-z
3. Pui CH, Thiel E. Central nervous system disease in hematologic Malignancies: historical perspective and practical applications. Semin Oncol (2009) 36(4 Suppl 2):S2–s16. doi: 10.1053/j.seminoncol.2009.05.002
4. Frishman-Levy L, Izraeli S. Advances in understanding the pathogenesis of CNS acute lymphoblastic leukaemia and potential for therapy. Br J Haematol (2017) 176(2):157–67. doi: 10.1111/bjh.14411
5. Gaynes JS, Jonart LM, Zamora EA, Naumann JA, Gossai NP, Gordon PM. The central nervous system microenvironment influences the leukemia transcriptome and enhances leukemia chemo-resistance. Haematologica (2017) 102(4):e136–e9. doi: 10.3324/haematol.2016.152926
6. Jabbour E, Thomas D, Cortes J, Kantarjian HM, O’Brien S. Central nervous system prophylaxis in adults with acute lymphoblastic leukemia: current and emerging therapies. Cancer (2010) 116(10):2290–300. doi: 10.1002/cncr.25008
7. Barrett T, Wilhite SE, Ledoux P, Evangelista C, Kim IF, Tomashevsky M, et al. NCBI GEO: archive for functional genomics data sets–update. Nucleic Acids Res (2013) 41(Database issue):D991–5. doi: 10.1093/nar/gks1193
8. Chen Z, McGee M, Liu Q, Scheuermann RH. A distribution free summarization method for Affymetrix GeneChip arrays. Bioinf (Oxford England) (2007) 23(3):321–7. doi: 10.1093/bioinformatics/btl609
9. Szklarczyk D, Franceschini A, Wyder S, Forslund K, Heller D, Huerta-Cepas J, et al. STRING v10: protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res (2015) 43(Database issue):D447–52. doi: 10.1093/nar/gku1003
10. Koh GC, Porras P, Aranda B, Hermjakob H, Orchard SE. Analyzing protein-protein interaction networks. J Proteome Res (2012) 11(4):2014–31. doi: 10.1021/pr201211w
11. Huang da W, Sherman BT, Lempicki RA. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat Protoc (2009) 4(1):44–57. doi: 10.1038/nprot.2008.211
12. Croft D, O’Kelly G, Wu G, Haw R, Gillespie M, Matthews L, et al. Reactome: a database of reactions, pathways and biological processes. Nucleic Acids Res (2011) 39(Database issue):D691–7. doi: 10.1093/nar/gkq1018
13. Munoz-Torres M, Carbon S. Get GO! Retrieving GO data using amiGO, quickGO, API, files, and tools. Methods Mol Biol (Clifton NJ) (2017) 1446:149–60. doi: 10.1007/978-1-4939-3743-1_11
14. Pio G, Malerba D, D’Elia D, Ceci M. Integrating microRNA target predictions for the discovery of gene regulatory networks: a semi-supervised ensemble learning approach. BMC Bioinf (2014) 15 Suppl 1(Suppl 1):S4. doi: 10.1186/1471-2105-15-S1-S4
15. Wong N, Wang X. miRDB: an online resource for microRNA target prediction and functional annotations. Nucleic Acids Res (2015) 43(Database issue):D146–52. doi: 10.1093/nar/gku1104
16. Agarwal V, Bell GW, Nam JW, Bartel DP. Predicting effective microRNA target sites in mammalian mRNAs. eLife (2015) 4:e05005. doi: 10.7554/eLife.05005
17. Paraskevopoulou MD, Georgakilas G, Kostoulas N, Vlachos IS, Vergoulis T, Reczko M, et al. DIANA-microT web server v5.0: service integration into miRNA functional analysis workflows. Nucleic Acids Res (2013) 41(Web Server issue):W169–73. doi: 10.1093/nar/gkt393
18. Davis AP, Grondin CJ, Johnson RJ, Sciaky D, Wiegers J, Wiegers TC, et al. Comparative toxicogenomics database (CTD): update 2021. Nucleic Acids Res (2021) 49(D1):D1138–d43. doi: 10.1093/nar/gkaa891
19. Yao H, Price TT, Cantelli G, Ngo B, Warner MJ, Olivere L, et al. Leukaemia hijacks a neural mechanism to invade the central nervous system. Nature (2018) 560(7716):55–60. doi: 10.1038/s41586-018-0342-5
20. Del Principe MI, Buccisano F, Soddu S, Maurillo L, Cefalo M, Piciocchi A, et al. Involvement of central nervous system in adult patients with acute myeloid leukemia: Incidence and impact on outcome. Semin Hematol (2018) 55(4):209–14. doi: 10.1053/j.seminhematol.2018.02.006
21. Hubbard SR, Till JH. Protein tyrosine kinase structure and function. Annu Rev Biochem (2000) 69:373–98. doi: 10.1146/annurev.biochem.69.1.373
22. Scales TM, Derkinderen P, Leung KY, Byers HL, Ward MA, Price C, et al. Tyrosine phosphorylation of tau by the SRC family kinases lck and fyn. Mol neurodegeneration (2011) 6:12. doi: 10.1186/1750-1326-6-12
23. Kalia LV, Salter MW. Interactions between Src family protein tyrosine kinases and PSD-95. Neuropharmacology (2003) 45(6):720–8. doi: 10.1016/S0028-3908(03)00313-7
24. Newcomer JW, Farber NB, Olney JW. NMDA receptor function, memory, and brain aging. Dialogues Clin Neurosci (2000) 2(3):219–32. doi: 10.31887/DCNS.2000.2.3/jnewcomer
25. Le HT, Maksumova L, Wang J, Pallen CJ. Reduced NMDA receptor tyrosine phosphorylation in PTPalpha-deficient mouse synaptosomes is accompanied by inhibition of four src family kinases and Pyk2: an upstream role for PTPalpha in NMDA receptor regulation. J neurochem (2006) 98(6):1798–809. doi: 10.1111/j.1471-4159.2006.04075.x
26. Jiang G, Freywald T, Webster J, Kozan D, Geyer R, DeCoteau J, et al. In human leukemia cells ephrin-B-induced invasive activity is supported by Lck and is associated with reassembling of lipid raft signaling complexes. Mol Cancer research: MCR. (2008) 6(2):291–305. doi: 10.1158/1541-7786.MCR-07-0047
27. Wright DD, Sefton BM, Kamps MP. Oncogenic activation of the Lck protein accompanies translocation of the LCK gene in the human HSB2 T-cell leukemia. Mol Cell Biol (1994) 14(4):2429–37. doi: 10.1128/mcb.14.4.2429
28. Horwitz V, Davidson B, Reich R. Laminin-induced signaling in tumor cells. Cancer letters (2005) 223:1–10. doi: 10.1016/j.canlet.2004.08.030
29. Engbring JA, Kleinman HK. The basement membrane matrix in Malignancy. J Pathol (2003) 200(4):465–70. doi: 10.1002/path.1396
30. Alinezhad S, Väänänen RM, Mattsson J, Li Y, Tallgrén T, Tong Ochoa N, et al. Validation of novel biomarkers for prostate cancer progression by the combination of bioinformatics, clinical and functional studies. PloS One (2016) 11(5):e0155901. doi: 10.1371/journal.pone.0155901
31. Chen TY, Liu Y, Chen L, Luo J, Zhang C, Shen XF. Identification of the potential biomarkers in patients with glioma: a weighted gene co-expression network analysis. Carcinogenesis (2020) 41(6):743–50. doi: 10.1093/carcin/bgz194
32. Peng Y, Wu D, Li F, Zhang P, Feng Y, He A. Identification of key biomarkers associated with cell adhesion in multiple myeloma by integrated bioinformatics analysis. Cancer Cell Int (2020) 20(1):262. doi: 10.1186/s12935-020-01355-z
33. Petz M, Them NC, Huber H, Mikulits W. PDGF enhances IRES-mediated translation of Laminin B1 by cytoplasmic accumulation of La during epithelial to mesenchymal transition. Nucleic Acids Res (2012) 40(19):9738–49. doi: 10.1093/nar/gks760
34. Barros CS, Franco SJ, Müller U. Extracellular matrix: functions in the nervous system. Cold Spring Harbor Perspect Biol (2011) 3(1):a005108. doi: 10.1101/cshperspect.a005108
35. Jogie-Brahim S, Feldman D, Oh Y. Unraveling insulin-like growth factor binding protein-3 actions in human disease. Endocrine Rev (2009) 30(5):417–37. doi: 10.1210/er.2008-0028
36. Dawczynski K, Steinbach D, Wittig S, Pfaffendorf N, Kauf E, Zintl F. Expression of components of the IGF axis in childhood acute myelogenous leukemia. Pediatr Blood Cancer (2008) 50(1):24–8. doi: 10.1002/pbc.21294
37. Mir S, Cai W, Carlson SW, Saatman KE, Andres DA. IGF-1 mediated neurogenesis involves a novel RIT1/akt/sox2 cascade. Sci Rep (2017) 7(1):3283. doi: 10.1038/s41598-017-03641-9
38. Grimberg A, Cohen P. Role of insulin-like growth factors and their binding proteins in growth control and carcinogenesis. J Cell Physiol (2000) 183(1):1–9. doi: 10.1002/(SICI)1097-4652(200004)183:1<1::AID-JCP1>3.0.CO;2-J
39. Kalluri HS, Dempsey RJ. IGFBP-3 inhibits the proliferation of neural progenitor cells. Neurochemical Res (2011) 36(3):406–11. doi: 10.1007/s11064-010-0349-2
40. Liu FT, Rabinovich GA. Galectins as modulators of tumour progression. Nat Rev Cancer (2005) 5(1):29–41. doi: 10.1038/nrc1527
41. Paz H, Joo EJ, Chou C-H, Fei F, Mayo KH, Abdel-Azim H, et al. Treatment of B-cell precursor acute lymphoblastic leukemia with the Galectin-1 inhibitor PTX008. J Exp Clin Cancer Res (2018) 37(1):67. doi: 10.1186/s13046-018-0721-7
42. Kajitani K, Nomaru H, Ifuku M, Yutsudo N, Dan Y, Miura T, et al. Galectin-1 promotes basal and kainate-induced proliferation of neural progenitors in the dentate gyrus of adult mouse hippocampus. Cell Death Differ (2009) 16(3):417–27. doi: 10.1038/cdd.2008.162
43. Sakaguchi M, Imaizumi Y, Okano H. Expression and function of galectin-1 in adult neural stem cells. Cell Mol Life Sci (2007) 64(10):1254. doi: 10.1007/s00018-007-6476-5
44. Wang KY, Huang RY, Tong XZ, Zhang KN, Liu YW, Zeng F, et al. Molecular and clinical characterization of TMEM71 expression at the transcriptional level in glioma. CNS Neurosci Ther (2019) 25(9):965–75. doi: 10.1111/cns.13137
45. Schmit K, Michiels C. TMEM proteins in cancer: A review. Front Pharmacol (2018) 9:1345. doi: 10.3389/fphar.2018.01345
46. Nagano T, Takehara S, Takahashi M, Aizawa S, Yamamoto A. Shisa2 promotes the maturation of somitic precursors and transition to the segmental fate in Xenopus embryos. Development (2006) 133(23):4643–54. doi: 10.1242/dev.02657
47. Tamura K, Furihata M, Satake H, Hashida H, Kawada C, Osakabe H, et al. SHISA2 enhances the aggressive phenotype in prostate cancer through the regulation of WNT5A expression. Oncol letters (2017) 14(6):6650–8. doi: 10.3892/ol.2017.7099
48. Sakaguchi M, Sonegawa H, Murata H, Kitazoe M, Futami JI, Kataoka K, et al. S100A11, an dual mediator for growth regulation of human keratinocytes. Mol Biol Cell (2008) 19(1):78–85. doi: 10.1091/mbc.e07-07-0682
49. Mori M, Shimada H, Gunji Y, Matsubara H, Hayashi H, Nimura Y, et al. S100A11 gene identified by in-house cDNA microarray as an accurate predictor of lymph node metastases of gastric cancer. Oncol Rep (2004) 11(6):1287–93. doi: 10.3892/or.11.6.1287
50. Takahashi RU, Prieto-Vila M, Kohama I, Ochiya T. Development of miRNA-based therapeutic approaches for cancer patients. Cancer Sci (2019) 110(4):1140–7. doi: 10.1111/cas.13965
51. Saccomani V, Grassi A, Piovan E, Bongiovanni D, Martino LD, Minuzzo S, et al. miR-22-3p negatively affects tumor progression in T-cell acute lymphoblastic leukemia. Cells (2020) 9(7):1726. doi: 10.3390/cells9071726
52. Li B, Wang Z, Yu M, Wang X, Wang X, Chen C, et al. miR-22-3p enhances the intrinsic regenerative abilities of primary sensory neurons via the CBL/p-EGFR/p-STAT3/GAP43/p-GAP43 axis. J Cell Physiol (2020) 235(5):4605–17. doi: 10.1002/jcp.29338
53. Ge WL, Chen Q, Meng LD, Huang XM, Shi GD, Zong QQ, et al. The YY1/miR-548t-5p/CXCL11 signaling axis regulates cell proliferation and metastasis in human pancreatic cancer. Cell Death disease (2020) 11(4):294. doi: 10.1038/s41419-020-2475-3
54. Zhou H, Liu W, Zhou Y, Hong ZY, Ni J, Zhang XP, et al. Therapeutic inhibition of GAS6-AS1/YBX1/MYC axis suppresses cell propagation and disease progression of acute myeloid leukemia. J Exp Clin Cancer Res (2021) 40(1):353. doi: 10.1186/s13046-021-02145-9
Keywords: acute lymphoblastic leukemia (ALL), metastasis, central nervous system (CNS), neuronal development, differentially expressed genes
Citation: Li Z, Guo Z, Xiao H, Chen X, Liu W and Zhou H (2024) Simulating neuronal development: exploring potential mechanisms for central nervous system metastasis in acute lymphoblastic leukemia. Front. Oncol. 13:1331802. doi: 10.3389/fonc.2023.1331802
Received: 01 November 2023; Accepted: 07 December 2023;
Published: 04 January 2024.
Edited by:
Lokman Varisli, Dicle University, TürkiyeReviewed by:
Spiros Vlahopoulos, University of Athens, GreeceCopyright © 2024 Li, Guo, Xiao, Chen, Liu and Zhou. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hao Zhou, emhvdWhhb0BodXN0LmVkdS5jbg==; Wei Liu, MjAwNFhIMTAxMEBodXN0LmVkdS5jbg==
†ORCID: Hao Zhou, orcid.org/0000-0002-8411-1589
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.