 Cui Wei
Cui Wei- Department of Emergency, The Third Hospital of Changsha, Changsha, China
Background: Though the matrix metalloproteinases (MMPs) are widely investigated in lung cancer (LC), however, almost no review systematically clarify their multi-faced roles in LC.
Methods: We investigated the expression of MMPs and their effects on survival of patients with LC, the resistance mechanisms of MMPs in anti-tumor therapy, the regulatory networks of MMPs involved, the function of MMPs inducing CSCLs, MMPs-related tumor immunity, and effects of MMP polymorphisms on risk of LC.
Results: High expression of MMPs was mainly related to poor survival, high clinical stages and cancer metastasis. Role of MMPs in LC are multi-faced. MMPs are involved in drug resistance, induced CSCLs, participated in tumor immunity. Besides, MMPs polymorphisms may increase risk of LC.
Conclusions: MMPs might be promising targets to restore the anti-tumor immune response and enhance the killing function of nature immune cells in LC.
Introduction
The matrix metalloproteinases (MMPs) are a group of zinc-containing endopeptidases which are widely involved in extracellular matrix (ECM) degradation, tumor cell proliferation and invasion, cell differentiation and apoptosis, cancer development, immune response, et al (1, 2). MMPs family contains 28 members, and at least 23 of them are expressed in human tissues (3). MMPs show highly homologous structures. Based on their structural domains and substrates, 23 MMPs expressed in human tissues are mainly classified into six subgroups: collagenases (MMP-1, MMP-8 and MMP-13); gelatinases (MMP-2 and MMP-9); stromelysins (MMP-3, MMP-10 and MMP-11); matrilysins (MMP-7 and MMP-26); membrane-type (MT) MMPs (MMP-14, MMP-15, MMP-16, MMP-17, MMP-24, and MMP-25); and other MMPs (MMP-12, MMP-19, MMP-20, MMP21, MMP-23, MMP-27, MMP-28) (Table 1) (3–5). MMPs exert various functions in both physiological and pathological conditions. Under physiological conditions, MMPs are mainly involved in cell apoptosis, embryogenesis, immune response, morphogenesis, tissue remodeling, tooth enamel formation, wound healing, angiogenesis, et al (Table 1) (3). Under pathological conditions, MMPs contributes to chronic venous disease, fibrotic disorder, inflammation, lung and liver disease, viral infection, cancer development, et al (Table 1) (3).
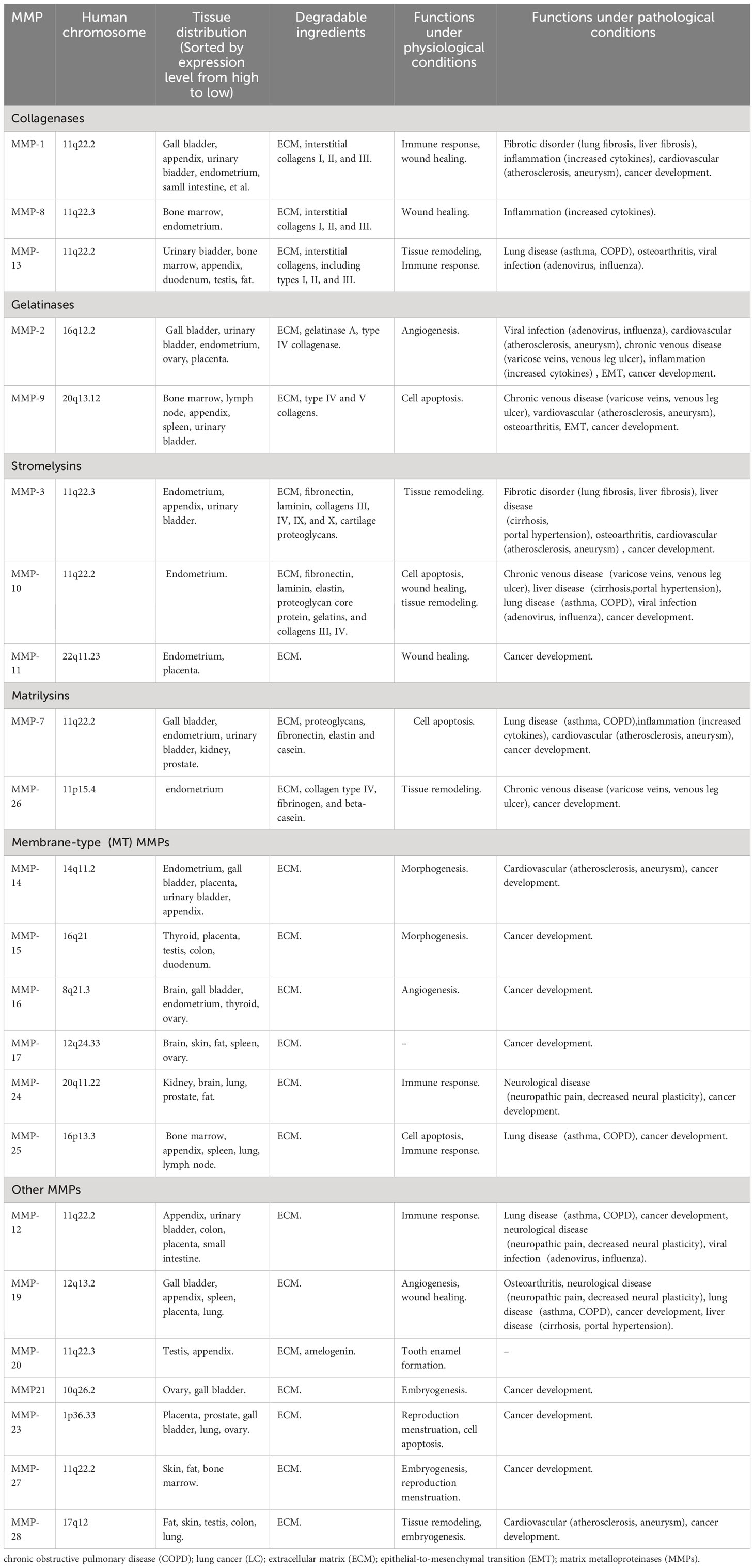
Table 1 Classification, tissue distribution and functions of MMPs.
Lung cancer(LC) is the most frequently new diagnosed cancer, and it’s estimated that LC ranked the top one among cancer deaths in 2020 (6). High activity and overexpression of MMPs in LC tissues contributed to LC invasion and metastasis, which induced to poor survival outcomes in cancer patients (7, 8). Study showed that MMP loss induced cell apoptosis in non-small cell lung cancer (NSCLC) (9). In A549 and H1299 LC cells, high expression of MMP-9 and MMP-2 contributed to epithelial-to-mesenchymal transition (EMT) (10). Together, high expression of MMPs induced cancer cell stemness, replication, inflammation, et al (11–13). In this review, we focus on the multi-faced roles of MMPs in LC, and try to discover potential application of targeting MMPs in immunotherapy in LC patients.
High MMPs are associated with cancer diagnosis, treatment and poor outcomes in patients with LC
Expression of MMPs were significantly correlated with TNM stage and poor survival outcomes in LC patients (14). A study has clearly investigated MMPs expression in lung adenocarcinoma (LUAD) and lung squamous cell carcinomas (LUSC) tissues (Figure 1). The results showed that MMP-1, 3, 9, 10, 11, 12, 13, and 17 were significantly upregulated in both LUAD and LUSC tissues (Figure 1). It’s reported that high expression of MMP-1 was related to male gender, smoking, and poorly differentiated tumor, besides, high serum MMP-1 showed a trend for short overall survival (14, 15) (Table 2). In addition, high expression of MMP-1 was also associated with tumor initiation, invasion, and metastasis in LC (34). MMP-2 was highly expressed in LC tissues, and adenocarcinomas showed higher expression of MMP-2 compared with squamous cell or large cell carcinomas (16, 17). Univariate and multivariate analyses showed that expression of MMP-2 showed significant prognostic value and predicated tumor recurrence in patients with NSCLC (16). Study also showed that MMP-2 may be implicated in early-stage tumor invasion, metastasis, and angiogenesis in NSCLC (35). Interestingly, in another study, MMP-2 expression didn’t show significant variation between metastatic and non-metastatic LC patients (36). The expression levels of MMP-3 differed significantly between patients with low and high N stage (P<0.001) (14). Higher MMP-3 expression levels were correlated with higher stages, therefore, MMP-3 was a potential marker associated with a high overall stage (P<0.001) (14). Study showed that expression of MMP-7 and MMP-9 were significantly higher in tumor tissue than in the adjacent tissues (37). Overexpression of MMP-7 played a role in cancer metastasis, and was associated with poor prognosis in NSCLC, besides, MMP-7 was involved in physiological processes including pathogenesis, invasion, and metastasis, contributing to predict the progression and prognosis of NSCLC (38, 39). However, another study showed that MMP-7 and MMP-9 may not be markers in early-stage tumor invasion, metastasis, and angiogenesis in NSCLC (35). MMP-9 expression in tissue was significantly higher in NSCLC than in small cell lung cancer (SCLC), and MMP-9 overexpression in NSCLC was related to the pathologic type and clinical stage of NSCLC, indicating its potential as a therapeutic target (19). MMP-9 expression differed significantly between cases with low and high N stage (P=0.025), and it was also correlated with overall stage, though without statistical significance (14). In addition, in vivo experiments showed that MMP-9 expressions of stage III and stage IV LC tissues were significantly higher than that in stage I and stage II LC tissues, indicating MMP-9 might be used as therapy and prognostic indicators for LC (20). Shorter survival time was found among positive MMP-9 expression in stage I NSCLC patients with negative lymph node (40). Interestingly, in this study, no difference in overall survival was observed with MMP-9 expression (41). In NSCLC tissues, the MMP-10 mRNA level was positively correlated to the MMP-10 protein level, however, there is no correlation in the adjacent tissues (42). MMP-10 plays an important role in the recurrence of stage IB LC patients, no matter what the histologic type is (43). However, univariate analysis showed that MMP-10 expression has no statistical significance with patients’ survival (P<0.289) (14). MMP-11, the highest upregulated MMP family member in LUAD cells, was also significantly increased in LUAD tissues (21). Besides, MMP-11was found to be upregulated in the recurred stage IB LC (43). In vitro study showed that MMP-11 depletion severely impaired cell proliferation, migration, and invasion of A549 LUAD cell, indicating MMP-11 is a key cancer driver gene in LUAD and is a potential target for cancer therapy (21). Expression of MMP12 protein was significantly increased in LUAD tissues compared with adjacent normal tissues (p= 0.019), and was closely correlated with the pathological stage and lymph node metastasis of LUAD patients (p= 0.01; p= 0.003), therefore, MMP-12 may be a promising therapeutic target for LUAD patients (22, 23). Immunostaining analyses showed that high MMP-13 index was found in most of the invasive LUAD lesions (24/27), but in none of the non-invasive tissues(0/4) (p=0.001) (44). Besides, MMP-13 positive expression accounts for larger proportion in grade II LC patients than grade I (24). MMP-14 was found to be upregulated in the recurred stage IB LC (43). In addition, high expression of MMP-14 is an unfavorable prognostic factor, and high levels of MMP-14 protein were positively correlated with advanced clinical stage, higher N classification, distant metastasis, and lower differentiated degree in patients with NSCLC (25, 26). Study showed that MMP-15(MT2-MMP)was also upregulated in the recurred stage IB LC (43). Besides, both mRNA and protein expression levels of MMP-15 were significantly upregulated in NSCLC tissues compared with adjacent normal tissues, in addition, MMP-15 might play an important role in promoting the tumor progression and angiogenesis in NSCLC (27). MMP-19 gene and protein expression were increased in LC tumors compared with adjacent normal tissues. The results derived from three independent datasets showed that increased MMP-19 gene expression conferred a poorer prognosis in NSCLC. Besides, in vitro experiments showed that overexpression of MMP-19 promotes EMT, migration, and invasiveness in several NSCLC cell lines (45). Study showed that the expression of MMP-20 in tumor tissues is very restricted, while in this study, expression of MMP-20 in LC was clearly observed and extremely low MMP-20 expression in normal tissues (28). Like most of MMP family members, MMP-24(MT5-MMP) was also overexpressed in LC tissues (29). Similarly, MMP-26 expression in LC tissues were also significantly higher than that in para-cancerous tissues, besides, high MMP-26 protein level was correlated to carcinogenesis, grade of cell differentiation, TNM stages, lymph node metastasis, and prognosis of NSCLC. Therefore, MMP-26 might be a potential tumor marker in monitoring progression and predicting prognosis of NSCLC (30–32). Study showed that MMP-27, MMP-28 were not significantly upregulated in LUSC and LUAD (46), suggesting that minor role of MMP-27, 28 played in NSCLC. Together, high expression of MMPs in LC tissues was significantly related with poor survival outcomes, advanced stages, larger tumor sizes, et al, indicating MMPs might be potential targets for LC therapy and useful markers for LC detecting.
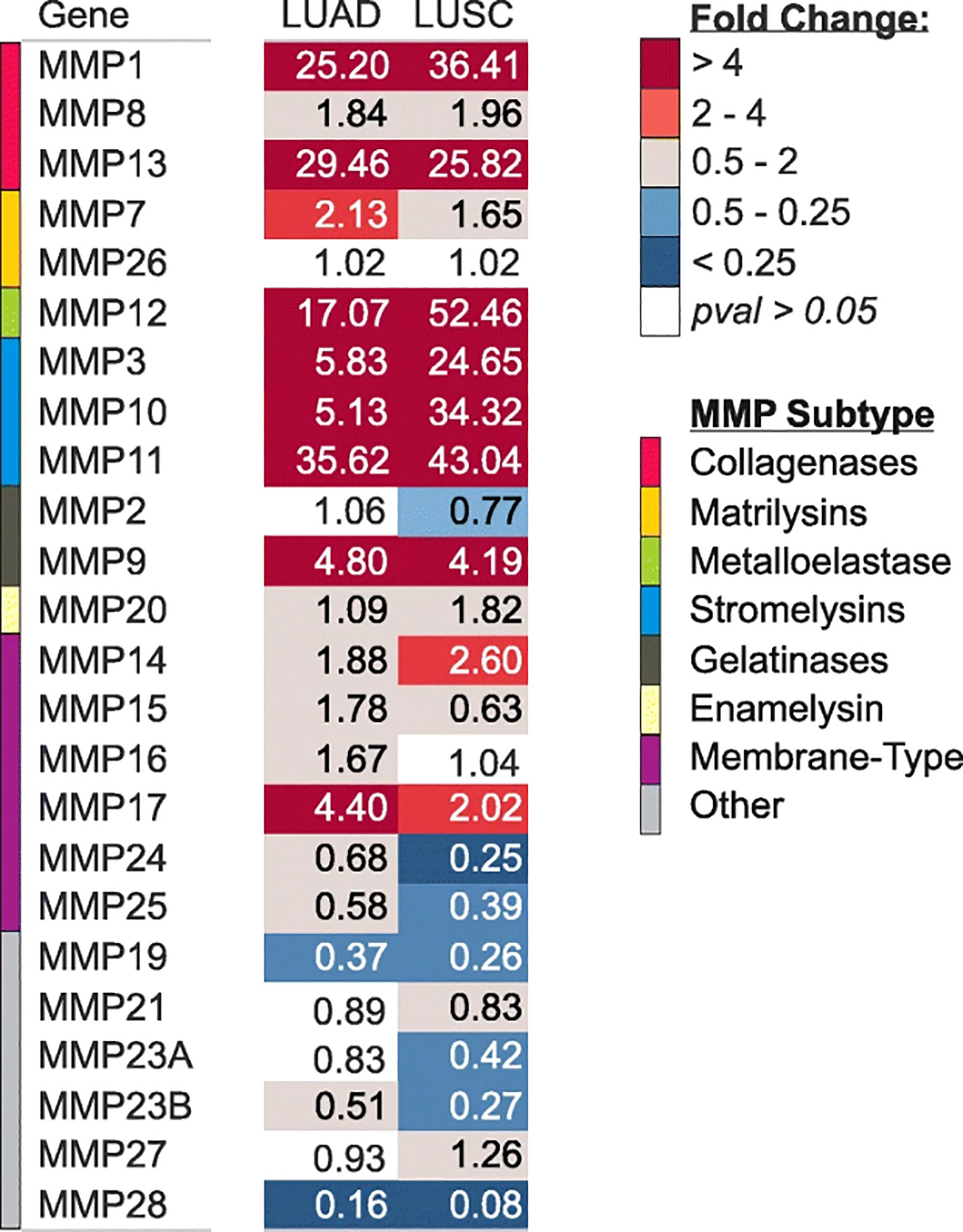
Figure 1 Gene expression of MMPs in LUAD and LUSC in TCGA. copyright 2009, with permission from Elsevier. Reprinted by permission from Springer Nature: Tumour Biol. Jiang J, Liu HL, Liu ZH, Tan SW, Wu (B) Identification of cystatin SN as a novel biomarker for pancreatic cancer. 2015;36 (5):3903–3910. Copyright 2015.
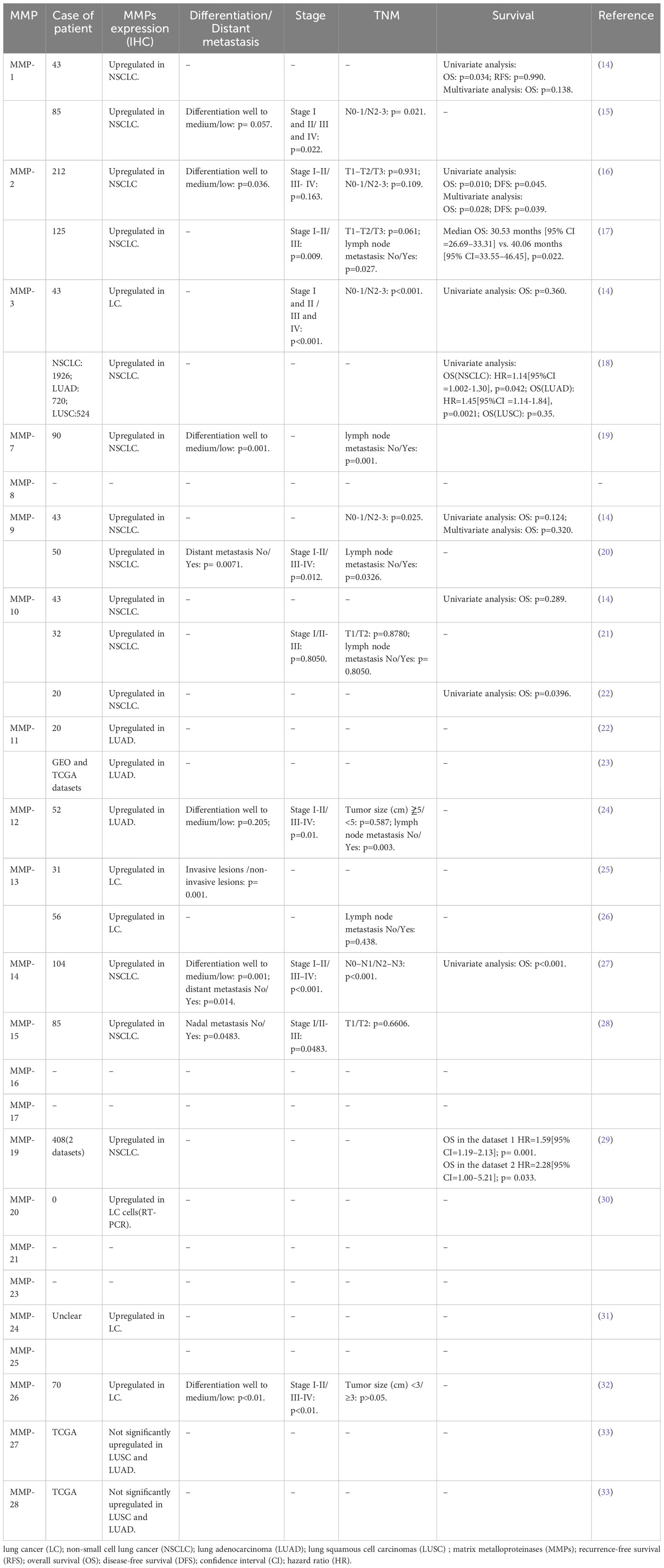
Table 2 Analyses of MMPs expression in lung cancer.
MMPs are involved in the resistance of LC to anti-tumor therapies
Accumulated evidence showed that MMPs were closely involved in anti-tumor treatment effects in patients with LC.
Overexpression of MMPs significantly contributed to EGFR-TKI-resistance in LC cells (Table 3). For example, upregulation of MMP-1, following the activation of mTOR signaling pathway, plays a significant role in migration and invasion of gefitinib- and erlotinib-resistant PC-9 cells, indicating that targeting MMP-1 and mTOR signaling pathways in EGFR-TKI-resistant LUAD might be potential therapy (47). In gefitinib-resistant NCI-H460 LC cells, MMP-2 is highly expressed, and MMP-2 suppression significantly inhibit cell adhesion, migration and invasion (33). In gefitinib-resistant NSCLC cells, MMP-12 was inhibited through suppression of the EGFR/ERK signaling pathway, thus inhibited cell proliferation, invasion, and migration (48). In sunitinib-resistant LC cells, upregulation of MMP-2/9 contributed to cell migration, invasion and metastasis (49). In the crizotinib-resistant A549 cells, increased expression of MMP-9 induced EMT, enhanced cell invasion and migration (50). Together, overexpression of MMP-1, 2, 9 induced EGFR-TKI-resistance in LC cells, inhibiting their expression might be useful to restore effects of EGFR-TKI on LC cells.
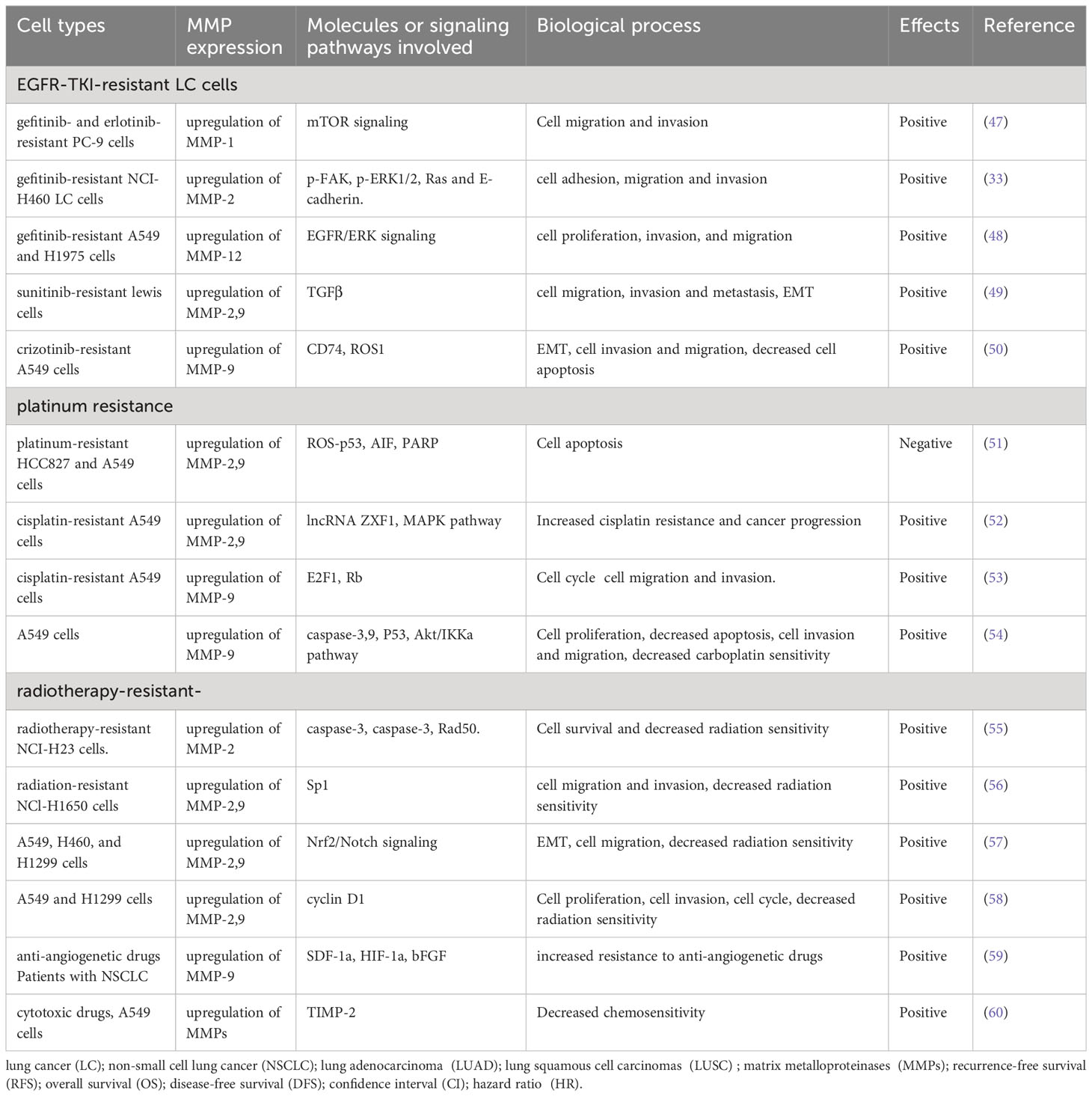
Table 3 MMPs are involved in the resistance of lung cancer to anti-tumor therapies.
MMPs are greatly involved in platinum resistance in LC cells (Table 3). Study showed that MMP-2/9 expression are significantly increased in HCC827 and A549 platinum-resistant LC cells, inhibition of the MMP-2/9 signaling pathways significantly induces cell apoptosis in cisplatin-resistant LC cells (51). In cisplatin-resistant LC cells, lncRNA ZXF1 contributes to cisplatin resistance and leads to the poor prognosis of LC patients by activating MAPK pathway and MMP 2/9 (52). Similarly, tyrphostin AG-1478, a selective EGFR-TKI, provide a potential therapeutic approach for cisplatin-resistant LC patients by inhibiting MMP-9 expression (53). In NSCLC cells, Curcumin(derived from the plant Curcuma longa) combined with carboplatin significantly inhibited tumor growth, cell migration, and invasion by effectively inhibiting MMP-2/9 expression, indicating that targeting MMP-2/9 and decreasing their expression and activities were potential effective way to enhance carboplatin sensitivity in patients with NSCLC (54). Together, inhibition of MMP-2, 7, 9 might be useful for restoring sensitivity of LC cells to platinum.
MMPs expression also affected radiation sensitivity (Table 3). MMP-2 is highly expressed in radiotherapy-resistant or radiotherapy-insensitive LC cells. Suppression of MMP-2 level significantly reduced, inhibited survival of cancer cells, and promoted radiosensitization (55). Overexpression of pecificity protein 1 (Sp1) lead to upregulation of MMP-2/9 in radiation-resistant LC cells. Celecoxib significantly restored radiation sensitivity and inhibited cell migration and invasion by inhibiting the expression and activity of Sp1, and MMP-2/9 (56). It’s reported that downregulation of MMP2/9 by inhibition of nuclear factor E2 related factor 2 (Nrf2) significantly reduced EMT, thus increased radiosensitivity of NSCLC cells (57). Interestingly, the proteasome inhibitor MG132, could regulated cell cycle and MMP 2/9 expression, which enhanced sensitivity of NSCLC cells to radiotherapy (58). In conclusion, reduced expression of MMP-2, 9 significantly enhanced sensitivity of LC cells to radiotherapy.
MMPs also affected other anti-tumor therapies (Table 3). Pre-clinical studies have showed that overexpression of MMP-9 contributed to resistance to anti-angiogenetic drugs (59). In A549 LC cells, TIMP-2 impeded tumor progression by inhibiting MMPs expression, which increased chemosensitivity of A549 cells to cytotoxic drugs, indicating that manipulating MMPs expression are potential approach to restore sensitivity of LC cells to cytotoxic drugs (60).
All in all, MMPs expression are closely associated with resistance of LC to anti-tumor therapies, targeting MMPs might be useful alternative to restore sensitivity of LC to anti-tumor therapies.
Regulatory network of MMPs in LC
MMPs were widely involved in various signaling network that are important to LC cells. Studies showed that transcription factors, growth factors, miRNAs and lncRNAs were related with expression and regulation of MMPs, which significantly affected LC development (Figure 2).
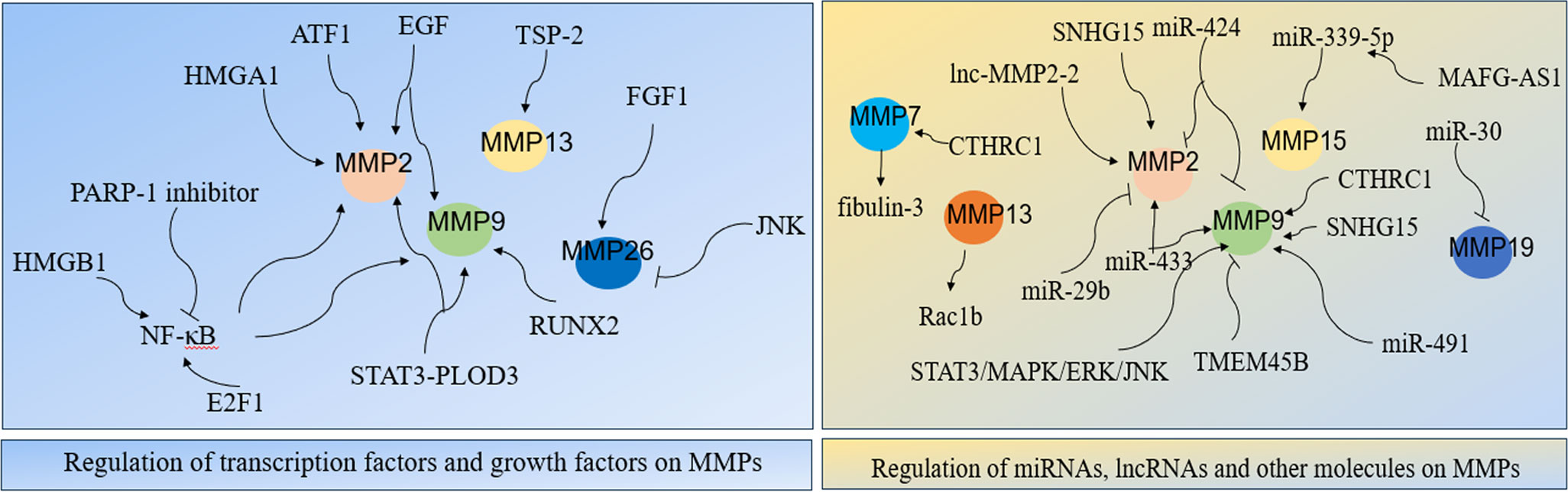
Figure 2 Regulatory network of MMPs in LC.
Transcription factors nuclear transcription factor-κB(NF-κB), E2F1, HMGA1, RUNX2, STAT3, and activating transcription factor 1 (ATF1) were important for expression and regulation of MMPs. NF-κB was firstly discovered in 1986, it regulates expression of its target genes and it plays crucial roles in cancer initiation and progression (61, 62). Studies have shown its role in MMPs in LC cells. Specifically, NF-κB stimulates the secretion of MMP-2/9, thus induced LC metastasis (63). In NSCLC cells, carbon ion (12C) radiotherapy combined with PARP-1 inhibitor significantly inhibited NF-kB expression, which followed by reduced MMP-2/9 expressions and significantly suppressed EMT and cancer metastasis (64). Study showed that suppressing MMP-9-dependent invasion pathway by regulating NF-κB activity was useful to inhibit ionizing radiation-induced LC metastasis (63). In LC A549 cells, curcumin suppressing cell migration and invasion by inhibiting adiponectin through NF-κB/MMP pathways, indicating that targeting NF-κB/MMP pathways might be an alternative for adjuvant therapy in LC patients (65). In SCLC cells, MMPs transcription could be directly enhanced by transcriptional activator E2F1 or be indirectly activated through enhanced NF-κB as a consequence of E2F1 activation (66). In LC, overexpression of TSP-2 leads to activation of integrin αvβ3/FAK/Akt/NF-κB/MMP-13 signaling pathway, which hence enhanced cell migration and invasion (67). The nucleoprotein HMGB1, promoted LC invasion and metastasis by upregulating the expression and activity of MMP-2 in an NF-κB-dependent manner (68). The architectural transcription factor HMGA1, contributed to transformation in undifferentiated large-cell LC by upregulating MMP-2 (69). RUNX2 is a Runt-related transcription factor, and it was aberrantly activated in cancer progression (70). In LC H1299 cells, inhibition of RUNX2 and its target gene MMP-9 by WW domain-containing oxidoreductase (WWOX) significantly suppressed cancer cell migration and invasion (71). Transcription factor STAT3 interacts with PLOD3, which hence lead to MMP-2/9 expression and contributed to LC metastasis (72). In LC cells, upregulation of activating transcription factor 1 (ATF1) increased MMP-2 expression, thereby causing cell invasion and migration (8). Together, MMPs expression largely depends on the regulation of transcription factors.
Growth factors are important proteins that regulate cell growth and proliferation, and they exert their functions through binding with specific receptors (73). Increasing evidences indicated that growth factors and their receptors are involved in MMPs regulation in LC. In NSCLC cells, four growth factors: insulin-like growth factor I/II (IGF I/II), hepatocyte growth factor (HGF), and epidermal growth factor (EGF) contributed to cell migration and invasion by increasing the expression and activity of MMP-2/9 (74). In A549 LC cells, suppressing p-EGFR lead to downregulation of MMP-2/9 expression, which hence inhibited cell proliferation, migration and invasion (75). Interestingly, overexpression of Derlin-1 in NSCLC contributed to cell invasion through EGFR-ERK-mediated up-regulation of MMP-2/9 (76). In A549 LC cells, fibroblast growth factor 1 (FGF1) -induced p-FGFR1 activated MMP26, which hence lead to cancer invasion, besides, inhibition of JNK significantly decreased the activation of MMP26 in response to FGF1 stimulation (31). All in all, growth factors and their receptors are closely involved in MMPs expression and activation, targeting or inhibiting combination of growth factors and their receptors might be potential effective ways to suppress MMP-induced LC invasion and metastasis.
More and more evidences indicated that miRNA interferes with MMPs expression. MiR-29b negatively regulated MMP-2, which might have potential usefulness for the treatment of NSCLC (77). MiR-30 isoforms (a microRNA family targeting MMP-19) are significantly down-regulated in human LC and regulate MMP-19 expression (45). Study showed that regulation of miR-339-5p/MMP15 axis by lncRNA MAFG-AS1promoted cell metastasis in NSCLC (78). In A549 cells, miR-424 inhibited MMP-2/9 expression, which thereby decreasing cell proliferation and migration (79). It’s reported that upregulation of miR-433 is closely related to decreased expression of MMP-2/9, which hence inhibited cell proliferation and invasion in NSCLC (80). In A549 cells, ginsenoside Rh2 (G-Rh2), derived from ginseng, exerts its anti-metastasis activity by repressing MMP-9 expression through miR-491, thereby inhibiting cancer metastasis (81). All in all, miRNAs are important for regulation of MMPs expression in LC.
LncRNA, is also significant for MMPs regulation in LC. In NSCLC, lncRNA MAFG-AS1 promoted cell metastasis by regulating miR-339-5p/MMP15 signaling pathway (78). TGF-β-mediated lnc-MMP2-2 regulates migration and invasion of LC cells by increasing MMP-2 expression (82). In A549 cells, knocking down lncRNA SNHG15 inhibited cell invasion and metastasis by suppressing the MMP-2/9 expression and EMT (83). In conclusion, lncRNA is greatly involved in expression and activation of MMPs.
Other molecules were also significant for MMPs network. Study showed that MMP3-Rac1b signaling axis is the important driver of tumor progression in LC (84). Downregulation of fibulin-3 contributes to LC invasion and metastasis by increasing MMP-7 expression (85). In NSCLC, overexpression of CTHRC1 contributes to cancer invasion and metastasis in a MMP7- and MMP9-dependent manner (86). In LC cells, inhibition of transmembrane protein 45B (TMEM45B) significantly inhibited cell invasion by regulating the expression of MMP-9 (18). Study showed that down-regulation of phosphorylation of JAK2/STAT3 significantly inhibited its downstream target genes MMP-2/9, thereby suppressing cancer metastasis and invasion in NSCLC (87). ERK activation in LC resulted to MMP-9 expression, thus led to cancer migration and development (88). In A549 cells, overexpression of spleen tyrosine kinase (Syk) significantly inhibited invasive ability of LC by inhibiting MMP-9 expression (89). In Lewis cells, suppressing the activities of MMP-2/9 through the modulation of STAT3/MAPK/ERK/JNK signaling pathway significantly inhibited cancer metastasis (90).
In conclusion, regulation of MMPs is complicated, and the regulatory network mainly included transcription factors, growth factors and their receptors, miRNA and lncRNAs, indicating that targeting or manipulating those regulators might be potential effective ways for LC therapy.
MMPs induced cancer stem cell-like cells
Cancer stem cells (CSC) are important factor of tumor recurrence, harboring CSC-like properties like self-renewal, aberrant differentiation, tumor recurrence, and acquired therapeutic resistance (91). Studies have shown that MMPs induced CSC-like cells in LC. Lung cancer stem-like cells (LCSLCs) from LUAD A549 cells carry the self-renewal potential, increased invasion, elevated tumorigenic activity, and high expression of stemness markers CD133, CD44, and aldehyde dehydrogenase 1 (ALDH1). Anti-tumor treatment suppressed properties of LCSLCs, inhibited expression of stemness markers, and also reduced MMP-9 activity (92). The CD133+ lung cancer stem cell (LCSC) is significantly correlated to the tumor metastasis and patients’ survival. Upregulation of MMP-9 induced by CD133+ LCSC contributed to cancer invasion and metastasis[96]. In NSCLC, cancer recurrence and metastasis are closely related to CSC, and these cells induced MMP-9 secretion (93). In LC cells, Aiolos overexpression promotes CSC-like properties by upregulating MMP-16 (94). Study showed that in the co-culture system of LUAD A549 cells, esophageal cancer cells, and mesenchymal stem cells (MSCs)-conditioned medium, MSCs induced cell apoptosis and downregulated MMP-2 in vitro, interestingly, this study also showed that MSCs enhanced tumor formation and growth in vivo (95). This result is really confusing, more vivo and vitro need to be done to figure out the phenomenon. In another study, cell fusion between LC cells and MSCs exerted increased metastatic capacity with upregulation of MMP-2/9 (96). Highly induced MMP-10 in lung bronchioalveolar stem cells (LBASCs) activated Kras, thus further contributed to tumor initiation, and maintained stem-like features (97). In conclusion, MMPs might be useful as diagnostic and prognostic biomarkers in LC, and may also represent a novel therapeutic approach to target LCSC.
MMPs are involved in tumor immunity
Studies showed that MMPs are involved in tumor immunity by regulating innate immunity through affecting proinflammatory cytokines, chemokines, other immune-related proteins, tumor microenvironment (TME) and several kinds of innate immune cells (98). Inhibition of MMPs has been showed to be effective in stimulation of immune system and inhibition of tumor growth (99).
Studies showed that MMPs functions in the tumor microenvironment (TME), and also affect immunotherapy effects. Accumulated evidence showed that MMPs promotes cancer immunosuppression, angiogenesis and inflammation in TME (100). MMP-9 is important in promoting the extravasation of tumor cells in TME (41). Myeloid-derived suppressor cells (MDSCs) are major components of the immune suppressive TME. MDSCs enhanced the pro-angiogenic, immune suppressive and pro-tumorigenic behavior of cancer cells by upregulating MMP-9 (101). Correspondingly, inhibition of MMP-9 by monoclonal antibody and TIMP-1 decreased MDSCs, which may inhibit tumor’s evasion of the immune response (102).
Complicated relationship existed between MMPs and macrophages(Mφ). Mφ are major sources of MMP-1, infection and inflammation increased MMP-1 secretion in lung tissues, indicating MMP-1 is an important driver for lung immunopathology (103). In co-cultured LC cells, G-Rh2 significantly inhibited MMP-2/9 expression, thereby converting tumor-associated macrophages (TAMs) from M2 to M1 and inhibited cancer migration (104). TAMs induced the remodeling of ECM through MMP-2/9 release, and contributed to angiogenesis and lymphangiogenesis through MMP-9 release (105). In NSCLC, TAM expressed high levels of MMP-9, which significantly increased cell migration and invasion, participated in vessel formation and sprouting, and thus lead to cancer progression (106, 107). Interestingly, in vivo experiments showed that MMP-9 lead to anti-tumor immune response by inducing neutrophil infiltration. Surprisingly, MMP-9 activated tumor-infiltrating macrophages into a tumor-inhibiting phenotype, inhibited tumor growth and angiogenesis, indicating MMP-9’s potential in regulating the innate immune response into anti-tumor action (108). TAMs were reported to be extremely important in delivering pro-MMP-9, thus induced angiogenesis in TME (109). Interestingly, MMP-9 in turn enhanced recruitment of Mφ during infection (110). During cancer metastasis, C-X-C chemokine receptor type 7 (CXCR7) recruits tumor-promoting macrophages (M2) to tumor tissues and upregulated MMP-2/9 expression (111).
Neutrophils played an important role in tumor immunity involving MMPs. TANs have been shown to both promote and inhibit tumor development. TANs promoted tumor by contributing to degradation of ECM via NE and MMP-9 (112). MMP-8, the neutrophil collagenase, is an important regulator of innate immunity that has onco-suppressive functions in cancers, and inhibition of MMP-8 might exert complicated effects on innate immunity (113). Neutrophil-derived MMP-8 is increased during pulmonary infection, indicating crucial role of MMP-8 played in the immunopathology of infectious disease (114). Similar to TAMs, tumor-associated neutrophils (TANs) are critically important for proMMP-9 delivering to TME, which mediating angiogenesis in tumor tissues (109). Interestingly, MMP-9 also plays a crucial role in the transmigration of neutrophils, lymphocytes, and eosinophils. MMP-9 inhibition suppressed inflammatory cell migration by suppressing IL-1beta, IL-4, and TNF-alpha expression (115). Study showed that TANs are major source of MMP-9 in human head and neck cancer and hepatocellular carcinoma, and neutrophil-derived MMP-9 has been involved in the angiogenic switch and tumor growth (116). It is worth noting that neutrophil-derived MMP-9 is secreted in a TIMP1-free manner, thus harboring powerful ability of angiogenesis (117).
T cells were also involved in MMP-related tumor immunity significantly. Membranes of activated T cells strongly induced the production of MMP-1/9 in lung tissue macrophages (LTM), however, unstimulated T cells failed to induce the secretion of MMPs. Meanwhile, IL-1 and TNFα are also closely involved in this process (118), indicating important roles of proinflammatory cytokines played on MMP-1/9 release. In vivo experiment showed that depletion of CD4(+) T lymphocytes lead to inhibition of antitumor activity and decreased antibodies against MMP-2, indicating that contribution of immune response against MMP-2 might be potential novel tumor methods for cancer therapy (119). Interestingly, Anti-MMP-9 treatment increases expression of T cell-stimulating factors, such as IL-12p70 and IL-18 (120). In vivo experiments showed that combination of anti-MMP-9 and anti-PDL1 induced TCR diversity, increased CD3+ T cells(including memory/effector CD4 and CD8 T cells), suggesting that inhibition of MMP-9 increased T-helper cell 1 type cytokines, induced delivery of effector/memory T cells to tumor tissues (120).
MMPs expression affected function of natural killer (NK) cells. In NCI−H23 human NSCLC cells, combination of ionizing radiation and MMP-2 inhibition promoted the killing function of NK−92 natural killer cells to cancer cells, suggesting that MMP-2 suppression is helpful for restoring host anti-tumor immune response (121). Interestingly, in cancer cells, IL-8/17 induced enhanced activity of MMP-2/9, thus promoted cancer metastasis (122). It has been investigated that inhibition of MMP25 by siRNA enhances the ADCC capacity of NK cells, emphasizing the important functional role of MMP25 in the regulation of ADCC activity, indicating that inhibition of MMP25 activity could improve ADCC efficacy of therapeutically administered NK cells (123). Interestingly, in vivo experiments showed that MMP-25-null mice exhibit a defective innate immune response, suggesting the possibility of activating innate immunity through induction of the activity of MMP-25 for the treatment of immune-related diseases (98).
In conclusion, MMPs are involved in tumor immunity by mutually action with various innate immune components, indicating modulating MMPs regulation and secretion might be potential for activating innate immune response in cancer therapy.
Effects of MMP polymorphisms on risk of LC
Polymorphisms in MMP genes were functional and may contribute to genetic susceptibility to cancers (124, 125).
Polymorphisms in the promoter region of MMP-1 significantly influenced the transcriptional activation of MMP1, as well as MMP-1 activity (126). A meta-analysis among diverse populations showed that MMP1-1607 2G/2G showed higher risk of LC than MMP1-1607 1G carriers (MMP1-1607 1G/1G and MMP1-1607 1G/2G), besides, MMP1-1607 2G carriers (MMP1-1607 2G/2G and MMP1-1607 1G/2G) showed significant higher risk of LC than MMP1-1607 1G/1G (34) (Table 4). In addition, a study showed that MMP1-1607 1G/2G showed higher risk of LC than MMP1-1607 1G/1G in never-smokers with odds ratio (OR) of 1.67 [95% confidence interval(CI)=1.02-2.76] (144), indicating 2G allele might be risk allele. However, a meta-analysis including 825 Han Chinese patients with LC showed no significant association between MMP1-1607 2G carriers and MMP1-1607 1G/1G, besides, another study showed that MMP1-1607 1G/2G and 2G/2G genotype (2G carriers) presented a lack of association with LC risk (OR = 1.04[95% CI = 0.68–1.58]and OR = 0.99[95% CI = 0.71–1.40], respectively), all of these results are consistent with the conclusion concluded by Xiao, X.Y etc. (34, 127, 139, 145)(Table 4). In addition, subgroup analyses by ethnicity identified significant association with LC risk in Asians, while ethnicity among Caucasian had no relationship with LC susceptibility (34) (Table 4), the results are consistent with the conclusion in another meta-analysis conducting on 24 studies with 10,099 LC cases and 9,395 controls (136). Possibly, genotypes varied between ethnic populations, therefore, there are differences of susceptibility to LC among various between ethnic populations. MMP1 2G/2G genotype enhances LC susceptibility especially in current and heavy smokers with OR=1.76 [95% CI= 1.26–2.39] and OR=2.55[95% CI=1.61–4.03], respectively, indicating prolonged cigarette exposure induced polymorphisms in MMP-1 (146) (Table 4). We speculated that smoking induced changes in tissues, which may build a microenvironment that is more suitable for tumorigenesis. Besides, microenvironment combined with increased MMP1 transcription might be important reason for that 2G/2G genotype have a higher risk of developing LC. A study showed that MMP1-1607 1G/2G polymorphism increased LC risk in Asians (131). While in another study, MMP-1 polymorphisms were found not associated with LC risk (135, 147). These findings are rather puzzling and contradictory, and we think they need to be further investigated in larger size of case–control studies. Based on the existing research results, we can speculate that Asians, especially Han Chinese, smoking and 2G carriers promote the risk of LC.
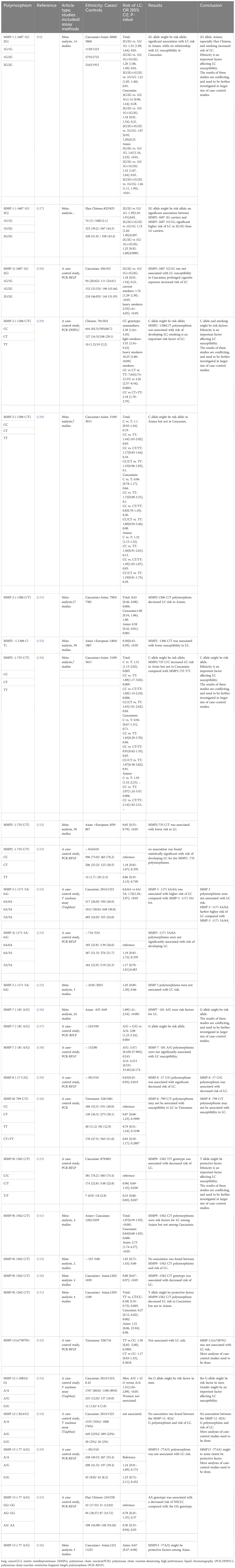
Table 4 Analysis of MMPs polymorphism in lung cancer.
It’s reported that MMP2 735C/T and 1306C/T polymorphisms are both associated with LC risk (128). The MMP2 -1306C/T polymorphism was significantly associated with risk of developing LC, and smoking is important risk factor of LC, indicating that MMP2 polymorphisms plays an important role in human carcinogenesis under smoking status. Interestingly, MMP2 -1306C/C polymorphisms showed higher risk than MMP2 -1306C/T and MMP2 -1306T/T, indicating that C allele might be risk allele in Han Chinese (129) (Table 4). Interestingly, a study showed that MMP2 -1306C/C and MMP-2 735C/C polymorphisms were significantly associated with higher LC risk in Asians but not in Caucasians compared with CT/TT, suggesting C allele’s risk feature varies among different ethnic population (130) (Table 4). However, it was found that MMP2-1306 C/T polymorphism decreased LC risk in Asians compared with patients in Caucasian (131) (Table 4). It is worth noting that a meta-analysis with more than 40,000 subjects showed that MMP2 -1306 C/T was associated with lower susceptibility to LC with OR= 0.50[95%CI=0.43-0.59], MMP2-735 C/T was associated with lower risk in LC (OR = 0.65[95%CI=0.53-0.79]) (132) (Table 4). Another meta-analysis even suggested that MMP2 -1306 C/T, MMP2 -735 C/T might be protective factors for LC (136). The results in this study also showed that MMP-2 1306C/T and -735C/T were significantly associated with protection against LC, with OR=0.53[95% CI=0.40-0.72] and OR=0.65[95% CI=0.53-0.79], respectively (135). Interestingly, in this case-control study, no association was found statistically significant with risk of developing LC for the MMP2 -735 polymorphisms (133) (Table 4).
MMP-2 gene expression was lower in homozygous -735CC LC patients than in those with CT or TT genotypes. Interestingly, the survival time was longer in patients with the MMP-2 -735T allele than in those with the CC genotype (148). The results are confusing because higher expression of MMP-2 usually indicates poor survival outcomes, therefore, the results showed be further investigated. A large cohort patients analysis showed that MMP-2-1306 CT/TT and CT genotypes showed significantly poor progression-free survival (PFS) in Caucasian patients with NSCLC, indicating MMP-2 polymorphisms might be potential prognostic markers in NSCLC (149). In addition, studies showed that MMP2 -735 T/T genotype was also proved a statistically significant independent prognosis factor associated with poor survival in NSCLC patients(hazard ratio (HR) = 1.79[95% CI=1.00-3.20]) (133, 141). However, a study showed that MMP-2 -1306C/T has no significant association with survival in stage I NSCLC patients and was not associated with risk of LC (145, 147). All in all, C allele and smoking might be risk factors of LC. Besides, ethnicity is also an important factor affecting LC susceptibility. Importantly, the results of these studies are conflicting, and need to be further investigated in larger size of case–control studies.
A study including 2014 Caucasian LC patients and 1323 healthy controls showed that MMP-3 -1171 6A/6A was associated with higher risk of LC in never smokers compared with MMP-3 -1171 5A/6A (OR=1.76[95%CI=1.04–2.97]) (134) (Table 4). However, in another study, MMP 3 polymorphisms were not associated with LC risk (135) (Table 4), and MMP3 -1171 5A/6A polymorphisms were also not significantly associated with risk of developing LC (133) (Table 4). A case-control study including 382 patients with stage I NSCLC showed that MMP-3 6A/5A was not significantly associated with recurrence-free survival(RFS) and overall survival(OS) (145). It’s evaluated that MMP3 -1171 5A/6A was also not associated with survival of LC patients (133). Together, MMP 3 polymorphisms were not associated with LC risk, and MMP-3 -1171 6A/6A harbor higher risk of LC compared with MMP-3 -1171 5A/6A.
A meta-analysis suggested that and MMP7 -181 A/G were risk factors for LC among Asian (136) (Table 4). A study including 243 NSCLC patients and 350 healthy controls showed that the frequency of the MMP-7 -181G allele in NSCLC patients was significantly higher than that in healthy controls, besides, the -181G allele (A/G + G/G) genotypes significantly increased susceptibility to NSCLC compared with the A/A genotype, with OR=2.00[95% CI = 1.23-3.24] for NSCLC (137) (Table 4).However, in another study, MMP-7 -181 A/G polymorphism was found not significantly associated with LC susceptibility (138) (Table 4). Stratification analysis in this study showed that smoking did not significantly influence the association between the MMP-7-181A/G and NSCLC, and MMP-7-181A/G polymorphism was not associated with lymphatic metastasis in NSCLC patients, indicating that the MMP-7-181A/G polymorphism might not predict lymphatic metastasis in NSCLC (137). In conclusion, G allele might be risk allele. Besides, the results of these studies are confusing, and need to be further investigated in larger size of case–control studies.
Multivariate analysis revealed that MMP8 polymorphisms are not independent prognostic factors for overall survival (139). The polymorphisms MMP-8 -799 C/T may not play a major role in susceptibility to LC in Taiwanese (140) (Table 4). However, the 17 C/G polymorphism in MMP8 was associated with a statistically significant decreased risk of developing LC (OR= 0.65[95%CI = 0.45-0.93]) (139) (Table 4).
A meta-analysis suggests that the MMP9 -1562 C/T polymorphisms were risk factors for LC among Asians (141) (Table 4). However, the MMP9 -1562 T/T genotype was associated with a statistically significant decreased risk of developing LC, but no association was found between MMP9 -1562 C/T polymorphisms and risk of LC (133, 135, 147) (Table 4). Subgroup analysis by smoking status showed no association between the MMP-9 1562 C/T polymorphism and the risk of NSCLC. Furthermore, the genotype distribution between NSCLC patients with and without lymphatic metastasis was not significantly different. Therefore, MMP-9 1562 C/T polymorphism may not be used as a useful marker to predicate susceptibility and lymphatic metastasis in NSCLC (150). However, a meta-analysis suggested that MMP9 -1562 C/T might be protective factors (136) (Table 4). Meanwhile, MMP9-1562 C/T polymorphism was proved decreasing LC risk in Caucasians (131) (Table 4). All in all, T allele might be protective factor, and ethnicity is an important factor affecting LC development. Importantly, the results of these studies are conflicting, and need to be further investigated in larger size of case–control studies.
The genotypes of MMP-11 play a minor role in determining LC risk in Taiwanese. MMP-11 rs738791 T allele did not confer LC risk compared with the C allele. Besides, there was no association between rs2267029, rs738792 or rs28382575 and LC risk (142) (Table 4).
A study conducted in 2014 Caucasian LC patients and 1323 healthy controls showed that the G allele of the MMP-12 1082 A/G polymorphism was associated with higher risk of LC in men (OR=1.51[95%CI=1.04–2.09]; A/G + G/G versus A/A), but not in women, indicating G allele was risk allele (134) (Table 4). No association was found between the MMP-12 -82A/G polymorphism and risk of LC (134) (Table 4). Patients with stage I NSCLC carrying the variant G allele of the MMP-12 1082A/G polymorphism have worse OS and RFS, while MMP-12 −82A/G was not significantly associated with survival (145). Together, the G allele might be risk factor in men, and gender might be an important factor affecting LC susceptibility. Importantly, more analyses of case-control studies need to be done to figure out the results.
The MMP-13 77 A/G polymorphisms AG and GG genotypes significantly increased in lung tumor DNA compared with healthy controls (138). However, studies showed that the MMP13 -77A/G polymorphism was not associated with LC risk (139, 141) (Table 4). Besides, a study showed that the AA genotype was associated with a decreased risk of NSCLC compared with the GG genotype (143) (Table 4). Interestingly, a meta-analysis even suggested that MMP13 -77A/G might be protective factors among Asian (136) (Table 4). Together, MMP13 -77A/G might to some extent be protective factor. However, more analyses of case-control studies need to be done to further investigate the results.
Discussion
In summary, roles of MMPs in LC are multi-faced. Most of MMPs are highly activated in cancers and contributed to cell proliferation and cancer development (8, 151–153). High expression of MMPs was mainly related to poor survival, high clinical stages and cancer metastasis. MMPs functions in a lot of physiological and pathological conditions. Expression of MMPs exerted significant prognostic value and predicated tumor recurrence in patients with LC. Besides, their expressions might be closely related with early-stage tumor invasion, metastasis, and angiogenesis, indicating their potential roles on monitoring progression and predicting prognosis of NSCLC. In addition, inhibition of MMPs might be useful for restoring sensitivity of LC cells to platinum, radiation, and immune therapy. Interestingly, MMPs polymorphisms also increase risk of LC. This review is the first to systematically clarify MMPs in LC. Although much work has been done, the reasons for increased MMP polymorphisms in LC, the mechanisms of MMPs induced CSCLs and the function of MMPs in tumor immunity should be further investigated. Accumulated evidences have shown that most of the MMP polymorphisms enhanced LC risks, but almost no study investigate the methods to decrease these risks, therefore, studies should be further explored to figure out the solutions. Nowadays, immunotherapy such as antibody‐mediated immune checkpoint blockade for LC has achieved great progress, however, LC remains the number one of cancer mortality (154, 155), indicating the necessities to uncover more new targets for immunotherapy. MMPs are closely involved in tumor immunity, significantly affected innate immune response. Targeting MMPs might be promising alternative to restore the host’s anti-tumor immune response and fight against LC cells.
Author contributions
CW: writing, editing, and revising the manuscript.
Conflict of interest
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Sternlicht MD, Werb Z. How matrix metalloproteinases regulate cell behavior. Annu Rev Cell Dev Biol (2001) 17:463–516. doi: 10.1146/annurev.cellbio.17.1.463
2. Visse R, Nagase H. Matrix metalloproteinases and tissue inhibitors of metalloproteinases: structure, function, and biochemistry. Circ Res (2003) 92(8):827–39. doi: 10.1161/01.RES.0000070112.80711.3D
3. Cui N, Hu M, Khalil RA. Biochemical and biological attributes of matrix metalloproteinases. Prog Mol Biol Trans science. (2017) 147:1–73. doi: 10.1016/bs.pmbts.2017.02.005
4. Kucukguven A, Khalil RA. Matrix metalloproteinases as potential targets in the venous dilation associated with varicose veins. Curr Drug Targets. (2013) 14(3):287–324.
5. English WR, Holtz B, Vogt G, Knäuper V, Murphy G. Characterization of the role of the "MT-loop": an eight-amino acid insertion specific to progelatinase A (MMP2) activating membrane-type matrix metalloproteinases. J Biol Chem (2001) 276(45):42018–26. doi: 10.1074/jbc.M107783200
6. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2020. CA: Cancer J Clin (2020) 70(1):7–30. doi: 10.3322/caac.21590
7. Gong L, Wu D, Zou J, Chen J, Chen L, Chen Y, et al. Prognostic impact of serum and tissue MMP-9 in non-small cell lung cancer: a systematic review and meta-analysis. Oncotarget. (2016) 7(14):18458–68. doi: 10.18632/oncotarget.7607
8. Cui J, Yin Z, Liu G, Chen X, Gao X, Lu H, et al. Activating transcription factor 1 promoted migration and invasion in lung cancer cells through regulating EGFR and MMP-2. Mol carcinogenesis. (2019) 58(10):1919–24. doi: 10.1002/mc.23086
9. Rasheduzzaman M, Jeong JK, Park SY. Resveratrol sensitizes lung cancer cell to TRAIL by p53 independent and suppression of Akt/NF-κB signaling. Life Sci (2018) 208:208–20. doi: 10.1016/j.lfs.2018.07.035
10. Baek SH, Ko JH, Lee JH, Kim C, Lee H, Nam D, et al. Ginkgolic acid inhibits invasion and migration and TGF-β-induced EMT of lung cancer cells through PI3K/akt/mTOR inactivation. J Cell Physiol (2017) 232(2):346–54. doi: 10.1002/jcp.25426
11. Hsiao YH, Hsieh MJ, Yang SF, Chen SP, Tsai WC, Chen PN. Phloretin suppresses metastasis by targeting protease and inhibits cancer stemness and angiogenesis in human cervical cancer cells. Phytomedicine Int J phytotherapy phytopharmacology. (2019) 62:152964. doi: 10.1016/j.phymed.2019.152964
12. Lou JS, Yan L, Bi CW, Chan GK, Wu QY, Liu YL, et al. Yu Ping Feng San reverses cisplatin-induced multi-drug resistance in lung cancer cells via regulating drug transporters and p62/TRAF6 signalling. Sci Rep (2016) 6:31926. doi: 10.1038/srep31926
13. Yan HQ, Zhang D, Shi YY, You X, Shi L, Li Q, et al. Ataxia-telangiectasia mutated activation mediates tumor necrosis factor-alpha induced MMP-13 up-regulation and metastasis in lung cancer cells. Oncotarget. (2016) 7(38):62070–83. doi: 10.18632/oncotarget.11386
14. Lim BJ, Jung SS, Choi SY, Lee CS. Expression of metastasis-associated molecules in non-small cell lung cancer and their prognostic significance. Mol Med Rep (2010) 3(1):43–9. doi: 10.3892/mmr_00000216
15. An HJ, Lee YJ, Hong SA, Kim JO, Lee KY, Kim YK, et al. The prognostic role of tissue and serum MMP-1 and TIMP-1 expression in patients with non-small cell lung cancer. Pathology Res practice. (2016) 212(5):357–64. doi: 10.1016/j.prp.2015.11.014
16. Leinonen T, Pirinen R, Böhm J, Johansson R, Kosma VM. Increased expression of matrix metalloproteinase-2 (MMP-2) predicts tumour recurrence and unfavourable outcome in non-small cell lung cancer. Histol Histopathol (2008) 23(6):693–700. doi: 10.14670/HH-23.693
17. Zhang L, Li N, Yan H, Jiang H, Fang X. Expression of novel CD44st and MMP2 in NSCLC tissues and their clinical significance. Oncol Res Treat (2017) 40(4):192–6. doi: 10.1159/000456669
18. Hu R, Hu F, Xie X, Wang L, Li G, Qiao T, et al. TMEM45B, up-regulated in human lung cancer, enhances tumorigenicity of lung cancer cells. Tumour Biol (2016) 37(9):12181–91. doi: 10.1007/s13277-016-5063-5
19. El-Badrawy MK, Yousef AM, Shaalan D, Elsamanoudy AZ. Matrix metalloproteinase-9 expression in lung cancer patients and its relation to serum mmp-9 activity, pathologic type, and prognosis. J bronchology interventional pulmonology. (2014) 21(4):327–34. doi: 10.1097/LBR.0000000000000094
20. Zhang DH, Zhang LY, Liu DJ, Yang F, Zhao JZ. Expression and significance of MMP-9 and MDM2 in the oncogenesis of lung cancer in rats. Asian Pacific J Trop Med (2014) 7(7):585–8. doi: 10.1016/S1995-7645(14)60099-7
21. Yang H, Jiang P, Liu D, Wang HQ, Deng Q, Niu X, et al. Matrix metalloproteinase 11 is a potential therapeutic target in lung adenocarcinoma. Mol Ther oncolytics. (2019) 14:82–93. doi: 10.1016/j.omto.2019.03.012
22. Wen Y, Cai L. Research progress of matrix metalloproteinase 12 in non-small cell lung cancer. Zhongguo Fei Ai Za Zhi (2014) 17(1):30–3. doi: 10.3779/j.issn.1009-3419.2014.01.05
23. Lv FZ, Wang JL, Wu Y, Chen HF, Shen XY. Knockdown of MMP12 inhibits the growth and invasion of lung adenocarcinoma cells. Int J immunopathol Pharmacol (2015) 28(1):77–84. doi: 10.1177/0394632015572557
24. Wang X, Yang J, Ma Y, Zheng H, Cheng B, Zheng H, et al. [Expression and significance of matrix metalloproteinases and their tissue inhibitors in the infiltration and metastasis of lung cancer]. Zhongguo Fei Ai Za Zhi. (2003) 6(4):278–82. doi: 10.3779/j.issn.1009-3419.2003.04.09
25. Wang YZ, Wu KP, Wu AB, Yang ZC, Li JM, Mo YL, et al. MMP-14 overexpression correlates with poor prognosis in non-small cell lung cancer. Tumour Biol (2014) 35(10):9815–21. doi: 10.1007/s13277-014-2237-x
26. Wang H, Zhang Y, Zhang Y, Liu W, Wang J. Cryptotanshinone inhibits lung cancer invasion via microRNA-133a/matrix metalloproteinase 14 regulation. Oncol Lett (2019) 18(3):2554–9. doi: 10.3892/ol.2019.10580
27. Chen L, Zhou Q, Xu B, Liu J, Shi L, Zhu D, et al. MT2-MMP expression associates with tumor progression and angiogenesis in human lung cancer. Int J Clin Exp Pathol (2014) 7(6):3469–77.
28. Kraus D, Reckenbeil J, Perner S, Winter J, Probstmeier R. Expression pattern of matrix metalloproteinase 20 (MMP20) in human tumors. Anticancer Res (2016) 36(6):2713–8.
29. Llano E, Pendás AM, Freije JP, Nakano A, Knäuper V, Murphy G, et al. Identification and characterization of human MT5-MMP, a new membrane-bound activator of progelatinase a overexpressed in brain tumors. Cancer Res (1999) 59(11):2570–6.
30. Li L, Mei TH, Zhou XD, Zhang XG. Expression and clinical significance of matrix metalloproteinase (MMP)-26 protein in non-small cell lung cancer. Ai Zheng. (2009) 28(1):60–3.
31. Zhao D, Lu Y, Yang C, Zhou X, Xu Z. Activation of FGF receptor signaling promotes invasion of non-small-cell lung cancer. Tumour Biol (2015) 36(5):3637–42. doi: 10.1007/s13277-014-3001-y
32. Zhang Y, Li C, Fang X, Sun M. Expression and clinical significance of matrix metalloproteinase-26 in non-small cell lung cancer. Zhongguo Fei Ai Za Zhi (2008) 11(3):420–4. doi: 10.3779/j.issn.1009-3419.2008.03.028
33. Huang AC, Yang MD, Hsiao YT, Lin TS, Ma YS, Peng SF, et al. Bufalin inhibits gefitinib resistant NCI-H460 human lung cancer cell migration and invasion in vitro. J ethnopharmacology (2016) 194:1043–50. doi: 10.1016/j.jep.2016.11.004
34. Xiao XY, Wang XD, Zang DY. MMP1-1607 1G/2G polymorphism and lung cancer risk: a meta-analysis. Tumour Biol (2012) 33(6):2385–92. doi: 10.1007/s13277-012-0502-4
35. Weng Y, Cai M, Zhu J, Geng J, Zhu K, Jin X, et al. Matrix metalloproteinase activity in early-stage lung cancer. Onkologie. (2013) 36(5):256–9. doi: 10.1159/000350304
36. Balla MM, Desai S, Purwar P, Kumar A, Bhandarkar P, Shejul YK, et al. Differential diagnosis of lung cancer, its metastasis and chronic obstructive pulmonary disease based on serum Vegf, Il-8 and MMP-9. Sci Rep (2016) 6:36065. doi: 10.1038/srep36065
37. Safranek J, Pesta M, Holubec L, Kulda V, Dreslerova J, Vrzalova J, et al. Expression of MMP-7, MMP-9, TIMP-1 and TIMP-2 mRNA in lung tissue of patients with non-small cell lung cancer (NSCLC) and benign pulmonary disease. Anticancer Res (2009) 29(7):2513–7.
38. Chen GL, Shen TC, Chang WS, Tsai CW, Li HT, Chuang CL, et al. The contribution of MMP-7 promoter polymorphisms to Taiwan lung cancer susceptibility. Anticancer Res (2018) 38(10):5671–7. doi: 10.21873/anticanres.12903
39. Xiao XY, Lang XP. Correlation between MMP-7 and bFGF expressions in non-small cell lung cancer tissue and clinicopathologic features. Cell Biochem biophysics. (2015) 73(2):427–32. doi: 10.1007/s12013-015-0656-y
40. Schveigert D, Cicenas S, Bruzas S, Samalavicius NE, Gudleviciene Z, Didziapetriene J. The value of MMP-9 for breast and non-small cell lung cancer patients' survival. Adv Med Sci (2013) 58(1):73–82. doi: 10.2478/v10039-012-0066-y
41. Ruiz-Morales JM, Dorantes-Heredia R, Arrieta O, Chávez-Tapia NC, Motola-Kuba D. Neutrophil gelatinase-associated lipocalin (NGAL) and matrix metalloproteinase-9 (MMP-9) prognostic value in lung adenocarcinoma. Tumour Biol J Int Soc Oncodevelopmental Biol Med (2015) 36(5):3601–10. doi: 10.1007/s13277-014-2997-3
42. Zhang X, Zhu S, Luo G, Zheng L, Wei J, Zhu J, et al. Expression of MMP-10 in lung cancer. Anticancer Res (2007) 27(4c):2791–5.
43. Cho NH, Hong KP, Hong SH, Kang S, Chung KY, Cho SH. MMP expression profiling in recurred stage IB lung cancer. Oncogene. (2004) 23(3):845–51. doi: 10.1038/sj.onc.1207140
44. Salaün M, Peng J, Hensley HH, Roder N, Flieder DB, Houlle-Crépin S, et al. MMP-13 in-vivo molecular imaging reveals early expression in lung adenocarcinoma. PloS One (2015) 10(7):e0132960. doi: 10.1371/journal.pone.0132960
45. Yu G, Herazo-Maya JD, Nukui T, Romkes M, Parwani A, Juan-Guardela BM, et al. Matrix metalloproteinase-19 promotes metastatic behavior in vitro and is associated with increased mortality in non-small cell lung cancer. Am J Respir Crit Care Med (2014) 190(7):780–90. doi: 10.1164/rccm.201310-1903OC
46. Gobin E, Bagwell K, Wagner J, Mysona D, Sandirasegarane S, Smith N, et al. A pan-cancer perspective of matrix metalloproteases (MMP) gene expression profile and their diagnostic/prognostic potential. BMC cancer. (2019) 19(1):581. doi: 10.1186/s12885-019-5768-0
47. Saito R, Miki Y, Ishida N, Inoue C, Kobayashi M, Hata S, et al. The significance of MMP-1 in EGFR-TKI-resistant lung adenocarcinoma: potential for therapeutic targeting. Int J Mol Sci (2018) 19(2):609. doi: 10.3390/ijms19020609
48. Xiao X, He Z, Cao W, Cai F, Zhang L, Huang Q, et al. Oridonin inhibits gefitinib-resistant lung cancer cells by suppressing EGFR/ERK/MMP-12 and CIP2A/Akt signaling pathways. Int J Oncol (2016) 48(6):2608–18. doi: 10.3892/ijo.2016.3488
49. Du Y, Liu JQ, Tang J, Ge J, Chen Y, Cheng K, et al. Acquired tumor cell resistance to sunitinib by increased invasion and epithelial-mesenchymal transition in LL/2 murine lung cancer. Oncotarget. (2017) 8(40):68270–9. doi: 10.18632/oncotarget.19295
50. Gou W, Zhou X, Liu Z, Wang L, Shen J, Xu X, et al. CD74-ROS1 G2032R mutation transcriptionally up-regulates Twist1 in non-small cell lung cancer cells leading to increased migration, invasion, and resistance to crizotinib. Cancer Lett (2018) 422:19–28. doi: 10.1016/j.canlet.2018.02.032
51. Zhu Z. Miltirone-induced apoptosis in cisplatin-resistant lung cancer cells through upregulation of p53 signaling pathways. Oncol Lett (2018) 15(6):8841–6. doi: 10.3892/ol.2018.8440
52. Yu T, Bai W, Su Y, Wang Y, Wang M, Ling C. Enhanced expression of lncRNA ZXF1 promotes cisplatin resistance in lung cancer cell via MAPK axis. Exp Mol pathology. (2020) 116:104484. doi: 10.1016/j.yexmp.2020.104484
53. Ma L, Yan H, Zhou Q. AG1478 inhibits the migration and invasion of cisplatin-resistant human lung adenocarcinoma cells via the cell cycle regulation by matrix metalloproteinase-9. Oncol Lett (2014) 8(2):921–7. doi: 10.3892/ol.2014.2224
54. Kang JH, Kang HS, Kim IK, Lee HY, Ha JH, Yeo CD, et al. Curcumin sensitizes human lung cancer cells to apoptosis and metastasis synergistically combined with carboplatin. Exp Biol Med (Maywood) (2015) 240(11):1416–25. doi: 10.1177/1535370215571881
55. Baruah TJ, Sharan RN, Kma L. Vicenin-2: a potential radiosensitizer of non-small cell lung cancer cells. Mol Biol Rep (2018) 45(5):1219–25. doi: 10.1007/s11033-018-4275-8
56. Liu R, Tan Q, Luo Q. Decreased expression level and DNA-binding activity of specificity protein 1 via cyclooxygenase-2 inhibition antagonizes radiation resistance, cell migration and invasion in radiation-resistant lung cancer cells. Oncol Lett (2018) 16(3):3029–37. doi: 10.3892/ol.2018.9035
57. Zhao Q, Mao A, Guo R, Zhang L, Yan J, Sun C, et al. Suppression of radiation-induced migration of non-small cell lung cancer through inhibition of Nrf2-Notch Axis. Oncotarget. (2017) 8(22):36603–13. doi: 10.18632/oncotarget.16622
58. Liu J, Shen W, Tang Y, Zhou J, Li M, Zhu W, et al. Proteasome inhibitor MG132 enhances the antigrowth and antimetastasis effects of radiation in human nonsmall cell lung cancer cells. Tumour Biol (2014) 35(8):7531–9. doi: 10.1007/s13277-014-2012-z
59. Dempke WCM, Heinemann V. Resistance to EGF-R (erbB-1) and VEGF-R modulating agents. Eur J Cancer (Oxford Engl 1990). (2009) 45(7):1117–28. doi: 10.1016/j.ejca.2008.11.038
60. Han H, Bourboulia D, Jensen-Taubman S, Isaac B, Wei B, Stetler-Stevenson WG. An endogenous inhibitor of angiogenesis inversely correlates with side population phenotype and function in human lung cancer cells. Oncogene. (2014) 33(9):1198–206. doi: 10.1038/onc.2013.61
61. Hoesel B, Schmid JA. The complexity of NF-κB signaling in inflammation and cancer. Mol Cancer. (2013) 12:86. doi: 10.1186/1476-4598-12-86
62. Sen R, Baltimore D. Multiple nuclear factors interact with the immunoglobulin enhancer sequences. Cell. (1986) 46(5):705–16. doi: 10.1016/0092-8674(86)90346-6
63. Jung CH, Han AR, Chung HJ, Ha IH, Um HD. Linarin inhibits radiation-induced cancer invasion by downregulating MMP-9 expression via the suppression of NF-κB activation in human non-small-cell lung cancer A549. Natural product Res (2019) 33(24):3582–6. doi: 10.1080/14786419.2018.1484460
64. Chowdhury P, Dey P, Ghosh S, Sarma A, Ghosh U. Reduction of metastatic potential by inhibiting EGFR/Akt/p38/ERK signaling pathway and epithelial-mesenchymal transition after carbon ion exposure is potentiated by PARP-1 inhibition in non-small-cell lung cancer. BMC Cancer. (2019) 19(1):829. doi: 10.1186/s12885-019-6015-4
65. Tsai JR, Liu PL, Chen YH, Chou SH, Cheng YJ, Hwang JJ, et al. Curcumin inhibits non-small cell lung cancer cells metastasis through the adiponectin/NF-κb/MMPs signaling pathway. PloS One (2015) 10(12):e0144462. doi: 10.1371/journal.pone.0144462
66. Li Z, Guo Y, Jiang H, Zhang T, Jin C, Young CY, et al. Differential regulation of MMPs by E2F1, Sp1 and NF-kappa B controls the small cell lung cancer invasive phenotype. BMC Cancer. (2014) 14:276. doi: 10.1186/1471-2407-14-276
67. Liu JF, Lee CW, Tsai MH, Tang CH, Chen PC, Lin LW, et al. Thrombospondin 2 promotes tumor metastasis by inducing matrix metalloproteinase-13 production in lung cancer cells. Biochem Pharmacol (2018) 155:537–46. doi: 10.1016/j.bcp.2018.07.024
68. Wu X, Wang W, Chen Y, Liu X, Wang J, Qin X, et al. High mobility group box protein 1 serves as a potential prognostic marker of lung cancer and promotes its invasion and metastasis by matrix metalloproteinase-2 in a nuclear factor-κB-dependent manner. BioMed Res Int (2018) 2018:3453706. doi: 10.1155/2018/3453706
69. Hillion J, Wood LJ, Mukherjee M, Bhattacharya R, Di Cello F, Kowalski J, et al. Upregulation of MMP-2 by HMGA1 promotes transformation in undifferentiated, large-cell lung cancer. Mol Cancer Res MCR. (2009) 7(11):1803–12. doi: 10.1158/1541-7786.MCR-08-0336
70. Manzotti G, Torricelli F, Donati B, Sancisi V, Gugnoni M, Ciarrocchi A. HDACs control RUNX2 expression in cancer cells through redundant and cell context-dependent mechanisms. J Exp Clin Cancer Res (2019) 38(1):346. doi: 10.1186/s13046-019-1350-5
71. Zheng QW, Zhou YL, You QJ, Shou F, Pang QF, Chen JL. WWOX inhibits the invasion of lung cancer cells by downregulating RUNX2. Cancer Gene Ther (2016) 23(12):433–8. doi: 10.1038/cgt.2016.59
72. Baek JH, Yun HS, Kwon GT, Kim JY, Lee CW, Song JY, et al. PLOD3 promotes lung metastasis via regulation of STAT3. Cell Death Dis (2018) 9(12):1138. doi: 10.1038/s41419-018-1186-5
73. Heldin CH. Dimerization of cell surface receptors in signal transduction. Cell. (1995) 80(2):213–23. doi: 10.1016/0092-8674(95)90404-2
74. Bredin CG, Liu Z, Klominek J. Growth factor-enhanced expression and activity of matrix metalloproteases in human non-small cell lung cancer cell lines. Anticancer Res (2003) 23(6c):4877–84.
75. Zhang H, Feng QQ, Gong JH, Ma JP. Anticancer effects of isofraxidin against A549 human lung cancer cells via the EGFR signaling pathway. Mol Med Rep (2018) 18(1):407–14. doi: 10.3892/mmr.2018.8950
76. Dong QZ, Wang Y, Tang ZP, Fu L, Li QC, Wang ED, et al. Derlin-1 is overexpressed in non-small cell lung cancer and promotes cancer cell invasion via EGFR-ERK-mediated up-regulation of MMP-2 and MMP-9. Am J pathology. (2013) 182(3):954–64. doi: 10.1016/j.ajpath.2012.11.019
77. Pastuszak-Lewandoska D, Kordiak J, Czarnecka KH, Migdalska-Sęk M, Nawrot E, Domańska-Senderowska D, et al. Expression analysis of three miRNAs, miR-26a, miR-29b and miR-519d, in relation to MMP-2 expression level in non-small cell lung cancer patients: a pilot study. Med Oncol (2016) 33(8):96. doi: 10.1007/s12032-016-0815-z
78. Jia YC, Wang JY, Liu YY, Li B, Guo H, Zang AM. LncRNA MAFG-AS1 facilitates the migration and invasion of NSCLC cell via sponging miR-339-5p from MMP15. Cell Biol Int (2019) 43(4):384–93. doi: 10.1002/cbin.11092
79. Li H, Lan H, Zhang M, An N, Yu R, He Y, et al. Effects of miR-424 on proliferation and migration abilities in non-small cell lung cancer A549 cells and its molecular mechanism. Zhongguo Fei Ai Za Zhi (2016) 19(9):571–6. doi: 10.3779/j.issn.1009-3419.2016.09.02
80. Li J, Chen M, Yu B. miR-433 suppresses tumor progression via Smad2 in non-small cell lung cancer. Pathology Res Practice (2019) 215(10):152591. doi: 10.1016/j.prp.2019.152591
81. Chen Y, Zhang Y, Song W, Zhang Y, Dong X, Tan M. Ginsenoside Rh2 Inhibits Migration of Lung Cancer Cells under Hypoxia via mir-491. Anticancer Agents Med Chem (2019) 19(13):1633–41. doi: 10.2174/1871520619666190704165205
82. Wu DM, Deng SH, Liu T, Han R, Zhang T, Xu Y. TGF-β-mediated exosomal lnc-MMP2-2 regulates migration and invasion of lung cancer cells to the vasculature by promoting MMP2 expression. Cancer Med (2018) 7(10):5118–29. doi: 10.1002/cam4.1758
83. Dong YZ, Meng XM, Li GS. Long non-coding RNA SNHG15 indicates poor prognosis of non-small cell lung cancer and promotes cell proliferation and invasion. Eur Rev Med Pharmacol Sci (2018) 22(9):2671–9. doi: 0.26355/eurrev_201805_14963
84. Mehner C, Miller E, Nassar A, Bamlet WR, Radisky ES, Radisky DC. Tumor cell expression of MMP3 as a prognostic factor for poor survival in pancreatic, pulmonary, and mammary carcinoma. Genes cancer. (2015) 6(11-12):480–9. doi: 10.18632/genesandcancer.90
85. Chen X, Meng J, Yue W, Yu J, Yang J, Yao Z, et al. Fibulin-3 suppresses Wnt/β-catenin signaling and lung cancer invasion. Carcinogenesis. (2014) 35(8):1707–16. doi: 10.1093/carcin/bgu023
86. He W, Zhang H, Wang Y, Zhou Y, Luo Y, Cui Y, et al. CTHRC1 induces non-small cell lung cancer (NSCLC) invasion through upregulating MMP-7/MMP-9. BMC Cancer (2018) 18(1):400. doi: 10.1186/s12885-018-4317-6
87. Su C, Zhang P, Liu J, Cao Y. Erianin inhibits indoleamine 2, 3-dioxygenase -induced tumor angiogenesis. BioMed Pharmacother (2017) 88:521–8. doi: 10.1016/j.biopha.2017.01.090
88. Wang XQ, Li H, Van Putten V, Winn RA, Heasley LE, Nemenoff RA. Oncogenic K-Ras regulates proliferation and cell junctions in lung epithelial cells through induction of cyclooxygenase-2 and activation of metalloproteinase-9. Mol Biol Cell (2009) 20(3):791–800. doi: 10.1091/mbc.e08-07-0732
89. Hao YT, Peng CL, Zhao YP, Sun QF, Zhao XG, Cong B. Effect of spleen tyrosine kinase on nonsmall cell lung cancer. J Cancer Res Ther (2018) 14(Supplement):S100–s4. doi: 10.4103/0973-1482.174170
90. Chen PC, Chen CC, Ker YB, Chang CH, Chyau CC, Hu ML. Anti-metastatic effects of antrodan with and without cisplatin on lewis lung carcinomas in a mouse xenograft model. Int J Mol Sci (2018) 19(6):1565–83. doi: 10.3390/ijms19061565
91. Eun K, Ham SW, Kim H. Cancer stem cell heterogeneity: origin and new perspectives on CSC targeting. BMB Rep (2017) 50(3):117–25. doi: 10.5483/BMBRep.2017.50.3.222
92. Liu F, Cao X, Liu Z, Guo H, Ren K, Quan M, et al. Casticin suppresses self-renewal and invasion of lung cancer stem-like cells from A549 cells through down-regulation of pAkt. Acta Biochim Biophys Sinica. (2014) 46(1):15–21. doi: 10.1093/abbs/gmt123
93. Roy S, Lu K, Nayak MK, Bhuniya A, Ghosh T, Kundu S, et al. Activation of D2 dopamine receptors in CD133+ve cancer stem cells in non-small cell lung carcinoma inhibits proliferation, clonogenic ability, and invasiveness of these cells. J Biol Chem (2017) 292(2):435–45. doi: 10.1074/jbc.M116.748970
94. Hung JJ, Kao YS, Huang CH, Hsu WH. Overexpression of Aiolos promotes epithelial-mesenchymal transition and cancer stem cell-like properties in lung cancer cells. Sci Rep (2019) 9(1):2991. doi: 10.1038/s41598-019-39545-z
95. Tian LL, Yue W, Zhu F, Li S, Li W. Human mesenchymal stem cells play a dual role on tumor cell growth in vitro and in vivo. J Cell Physiol (2011) 226(7):1860–7. doi: 10.1002/jcp.22511
96. Zhang LN, Kong CF, Zhao D, Cong XL, Wang SS, Ma L, et al. Fusion with mesenchymal stem cells differentially affects tumorigenic and metastatic abilities of lung cancer cells. J Cell Physiol (2019) 234(4):3570–82. doi: 10.1002/jcp.27011
97. Regala RP, Justilien V, Walsh MP, Weems C, Khoor A, Murray NR, et al. Matrix metalloproteinase-10 promotes Kras-mediated bronchio-alveolar stem cell expansion and lung cancer formation. PloS One (2011) 6(10):e26439. doi: 10.1371/journal.pone.0026439
98. Soria-Valles C, Gutiérrez-Fernández A, Osorio FG, Carrero D, Ferrando AA, Colado E, et al. MMP-25 metalloprotease regulates innate immune response through NF-κB signaling. J Immunol (Baltimore Md 1950). (2016) 197(1):296–302. doi: 10.4049/jimmunol.1600094
99. Luo KW, Yue GG, Ko CH, Lee JK, Gao S, Li LF, et al. In vivo and in vitro anti-tumor and anti-metastasis effects of Coriolus versicolor aqueous extract on mouse mammary 4T1 carcinoma. Phytomedicine Int J phytotherapy phytopharmacology. (2014) 21(8-9):1078–87. doi: 10.1016/j.phymed.2014.04.020
100. Guedez L, Jensen-Taubman S, Bourboulia D, Kwityn CJ, Wei B, Caterina J, et al. TIMP-2 targets tumor-associated myeloid suppressor cells with effects in cancer immune dysfunction and angiogenesis. J immunotherapy (Hagerstown Md 1997). (2012) 35(6):502–12. doi: 10.1097/CJI.0b013e3182619c8e
101. Mackert JR, Qu P, Min Y, Johnson PF, Yang L, Lin PC. Dual negative roles of C/EBPα in the expansion and pro-tumor functions of MDSCs. Sci Rep (2017) 7(1):14048. doi: 10.1038/s41598-017-12968-2
102. Shao L, Zhang B, Wang L, Wu L, Kan Q, Fan K. MMP-9-cleaved osteopontin isoform mediates tumor immune escape by inducing expansion of myeloid-derived suppressor cells. Biochem Biophys Res Commun (2017) 493(4):1478–84. doi: 10.1016/j.bbrc.2017.10.009
103. Elkington P, Shiomi T, Breen R, Nuttall R, Ugarte-Gil C, Walker N, et al. MMP-1 drives immunopathology in human tuberculosis and transgenic mice. J Clin Invest (2011) 121(5):1827–33. doi: 10.1172/JCI45666
104. Li H, Huang N, Zhu W, Wu J, Yang X, Teng W, et al. Modulation the crosstalk between tumor-associated macrophages and non-small cell lung cancer to inhibit tumor migration and invasion by ginsenoside Rh2. BMC Cancer. (2018) 18(1):579. doi: 10.1186/s12885-018-4299-4
105. Galdiero MR, Bonavita E, Barajon I, Garlanda C, Mantovani A, Jaillon S. Tumor associated macrophages and neutrophils in cancer. Immunobiology. (2013) 218(11):1402–10. doi: 10.1016/j.imbio.2013.06.003
106. Wang R, Zhang J, Chen S, Lu M, Luo X, Yao S, et al. Tumor-associated macrophages provide a suitable microenvironment for non-small lung cancer invasion and progression. Lung Cancer. (2011) 74(2):188–96. doi: 10.1016/j.lungcan.2011.04.009
107. Schoppmann SF, Birner P, Stöckl J, Kalt R, Ullrich R, Caucig C, et al. Tumor-associated macrophages express lymphatic endothelial growth factors and are related to peritumoral lymphangiogenesis. Am J pathology. (2002) 161(3):947–56. doi: 10.1016/S0002-9440(10)64255-1
108. Leifler KS, Svensson S, Abrahamsson A, Bendrik C, Robertson J, Gauldie J, et al. Inflammation induced by MMP-9 enhances tumor regression of experimental breast cancer. J Immunol (2013) 190(8):4420–30. doi: 10.4049/jimmunol.1202610
109. Deryugina EI, Zajac E, Juncker-Jensen A, Kupriyanova TA, Welter L, Quigley JP. Tissue-infiltrating neutrophils constitute the major in vivo source of angiogenesis-inducing MMP-9 in the tumor microenvironment. Neoplasia (New York NY). (2014) 16(10):771–88. doi: 10.1016/j.neo.2014.08.013
110. Volkman H, Pozos T, Zheng J, Davis J, Rawls J, Ramakrishnan L. Tuberculous granuloma induction via interaction of a bacterial secreted protein with host epithelium. Sci (New York NY). (2010) 327(5964):466–9. doi: 10.1126/science.1179663
111. Wani N, Nasser MW, Ahirwar DK, Zhao H, Miao Z, Shilo K, et al. C-X-C motif chemokine 12/C-X-C chemokine receptor type 7 signaling regulates breast cancer growth and metastasis by modulating the tumor microenvironment. Breast Cancer Res BCR. (2014) 16(3):R54. doi: 10.1186/bcr3665
112. Hurt B, Schulick R, Edil B, El Kasmi KC, Barnett C Jr.Cancer-promoting mechanisms of tumor-associated neutrophils. Am J surgery. (2017) 214(5):938–44 doi: 10.1016/j.amjsurg.2017.08.003
113. Decock J, Hendrickx W, Thirkettle S, Gutiérrez-Fernández A, Robinson SD, Edwards DR. Pleiotropic functions of the tumor- and metastasis-suppressing matrix metalloproteinase-8 in mammary cancer in MMTV-PyMT transgenic mice. Breast Cancer Res BCR. (2015) 17(1):38. doi: 10.1186/s13058-015-0545-8
114. Ong CW, Elkington PT, Brilha S, Ugarte-Gil C, Tome-Esteban MT, Tezera LB, et al. Neutrophil-derived MMP-8 drives AMPK-dependent matrix destruction in human pulmonary tuberculosis. PloS pathogens. (2015) 11(5):e1004917. doi: 10.1371/journal.ppat.1004917
115. Lee KS, Jin SM, Kim HJ, Lee YC. Matrix metalloproteinase inhibitor regulates inflammatory cell migration by reducing ICAM-1 and VCAM-1 expression in a murine model of toluene diisocyanate-induced asthma. J Allergy Clin Immunol (2003) 111(6):1278–84. doi: 10.1067/mai.2003.1501
116. Dumitru CA, Fechner MK, Hoffmann TK, Lang S, Brandau S. A novel p38-MAPK signaling axis modulates neutrophil biology in head and neck cancer. J leukocyte Biol (2012) 91(4):591–8. doi: 10.1189/jlb.0411193
117. Ardi VC, Kupriyanova TA, Deryugina EI, Quigley JP. Human neutrophils uniquely release TIMP-free MMP-9 to provide a potent catalytic stimulator of angiogenesis. Proc Natl Acad Sci USA (2007) 104(51):20262–7. doi: 10.1073/pnas.0706438104
118. Ferrari-Lacraz S, Nicod LP, Chicheportiche R, Welgus HG, Dayer JM. Human lung tissue macrophages, but not alveolar macrophages, express matrix metalloproteinases after direct contact with activated T lymphocytes. Am J Respir Cell Mol Biol (2001) 24(4):442–51. doi: 10.1165/ajrcmb.24.4.4008
119. Su JM, Wei YQ, Tian L, Zhao X, Yang L, He QM, et al. Active immunogene therapy of cancer with vaccine on the basis of chicken homologous matrix metalloproteinase-2. Cancer Res (2003) 63(3):600–7.
120. Juric V, O'Sullivan C, Stefanutti E, Kovalenko M, Greenstein A, Barry-Hamilton V, et al. MMP-9 inhibition promotes anti-tumor immunity through disruption of biochemical and physical barriers to T-cell trafficking to tumors. PloS One (2018) 13(11):e0207255. doi: 10.1371/journal.pone.0207255
121. Heo W, Lee YS, Son CH, Yang K, Park YS, Bae J. Radiation-induced matrix metalloproteinases limit natural killer cell-mediated anticancer immunity in NCI-H23 lung cancer cells. Mol Med Rep (2015) 11(3):1800–6. doi: 10.3892/mmr.2014.2918
122. Zarogoulidis P, Katsikogianni F, Tsiouda T, Sakkas A, Katsikogiannis N, Zarogoulidis K. Interleukin-8 and interleukin-17 for cancer. Cancer Invest (2014) 32(5):197–205. doi: 10.3109/07357907.2014.898156
123. Peruzzi G, Femnou L, Gil-Krzewska A, Borrego F, Weck J, Krzewski K, et al. Membrane-type 6 matrix metalloproteinase regulates the activation-induced downmodulation of CD16 in human primary NK cells. J Immunol (2013) 191(4):1883–94. doi: 10.4049/jimmunol.1300313
124. Zhang LF, Mi YY, Cao Q, Wang W, Qin C, Wei JF, et al. Update analysis of studies on the MMP-9 -1562 C>T polymorphism and cancer risk. Mol Biol Rep (2012) 39(4):3435–41. doi: 10.1007/s11033-011-1115-5
125. Sauter W, Rosenberger A, Beckmann L, Kropp S, Mittelstrass K, Timofeeva M, et al. Matrix metalloproteinase 1 (MMP1) is associated with early-onset lung cancer. Cancer epidemiology Biomarkers Prev Publ Am Assoc Cancer Research cosponsored by Am Soc Prev Oncol (2008) 17(5):1127–35. doi: 10.1158/1055-9965.EPI-07-2840
126. Liu B, Yi M, Tang Y, Liu Q, Qiu H, Zou Y, et al. MMP-1 promoter polymorphism is associated with risk of radiation-induced lung injury in lung cancer patients treated with radiotherapy. Oncotarget (2016) 7(43):70175–84. doi: 10.18632/oncotarget.12164
127. Liu L, Wu J, Wu C, Wang Y, Zhong R, Zhang X, et al. A functional polymorphism (-1607 1G→2G) in the matrix metalloproteinase-1 promoter is associated with development and progression of lung cancer. Cancer (2011) 117(22):5172–81. doi: 10.1002/cncr.26154
128. Wang J, Cai Y. Matrix metalloproteinase 2 polymorphisms and expression in lung cancer: a meta-analysis. Tumour Biol (2012) 33(6):1819–28. doi: 10.1007/s13277-012-0441-0
129. Yu C, Pan K, Xing D, Liang G, Tan W, Zhang L, et al. Correlation between a single nucleotide polymorphism in the matrix metalloproteinase-2 promoter and risk of lung cancer. Cancer Res (2002) 62(22):6430–3.
130. Guo XT, Wang JF, Zhang LY, Xu GQ. Quantitative assessment of the effects of MMP-2 polymorphisms on lung carcinoma risk. Asian Pacific J Cancer Prev APJCP. (2012) 13(6):2853–6. doi: 10.7314/APJCP.2012.13.6.2853
131. Hu C, Wang J, Xu Y, Li X, Chen H, Bunjhoo H, et al. Current evidence on the relationship between five polymorphisms in the matrix metalloproteinases (MMP) gene and lung cancer risk: a meta-analysis. Gene (2013) 517(1):65–71. doi: 10.1016/j.gene.2012.12.085
132. Peng B, Cao L, Ma X, Wang W, Wang D, Yu L. Meta-analysis of association between matrix metalloproteinases 2, 7 and 9 promoter polymorphisms and cancer risk. Mutagenesis. (2010) 25(4):371–9. doi: 10.1093/mutage/geq015
133. González-Arriaga P, Pascual T, García-Alvarez A, Fernández-Somoano A, López-Cima MF, Tardón A. Genetic polymorphisms in MMP 2, 9 and 3 genes modify lung cancer risk and survival. BMC Cancer. (2012) 12:121. doi: 10.1186/1471-2407-12-121
134. Su L, Zhou W, AsOmaning K, Lin X, Wain JC, Lynch TJ, et al. Genotypes and haplotypes of matrix metalloproteinase 1, 3 and 12 genes and the risk of lung cancer. Carcinogenesis. (2006) 27(5):1024–9. doi: 10.1093/carcin/bgi283
135. McColgan P, Sharma P. Polymorphisms of matrix metalloproteinases 1, 2, 3 and 9 and susceptibility to lung, breast and colorectal cancer in over 30,000 subjects. Int J Cancer (2009) 125(6):1473–8. doi: 10.1002/ijc.24441
136. Li X, Liu C, Ran R, Liu G, Yang Y, Zhao W, et al. Matrix metalloproteinase family gene polymorphisms and lung cancer susceptibility: an updated meta-analysis. J Thorac Dis (2020) 12(3):349–62. doi: 10.21037/jtd.2020.01.25
137. Zhang J, Jin X, Fang S, Wang R, Li Y, Wang N, et al. The functional polymorphism in the matrix metalloproteinase-7 promoter increases susceptibility to esophageal squamous cell carcinoma, gastric cardiac adenocarcinoma and non-small cell lung carcinoma. Carcinogenesis. (2005) 26(10):1748–53. doi: 10.1093/carcin/bgi144
138. Sanli M, Akar E, Pehlivan S, Bakır K, Tuncozgur B, Isik AF, et al. The relationship of metalloproteinase gene polymorphisms and lung cancer. J Surg Res (2013) 183(2):517–23. doi: 10.1016/j.jss.2013.01.045
139. González-Arriaga P, López-Cima MF, Fernández-Somoano A, Pascual T, Marrón MG, Puente XS, et al. Polymorphism +17 C/G in matrix metalloprotease MMP8 decreases lung cancer risk. BMC Cancer. (2008) 8:378. doi: 10.1186/1471-2407-8-378
140. Shen TC, Hsia TC, Chao CY, Chen WC, Chen CY, Chen WC, et al. The contribution of MMP-8 promoter polymorphisms in lung cancer. Anticancer Res (2017) 37(7):3563–7. doi: 10.21873/anticanres.11726
141. Li H, Liang X, Qin X, Cai S, Yu S. Association of matrix metalloproteinase family gene polymorphisms with lung cancer risk: logistic regression and generalized odds of published data. Sci Rep (2015) 5:10056. doi: 10.1038/srep10056
142. Chen GL, Wang SC, Huang WC, Chang WS, Tsai CW, Li HT, et al. The association of MMP-11 promoter polymorphisms with susceptibility to lung cancer in Taiwan. Anticancer Res (2019) 39(10):5375–80. doi: 10.21873/anticanres.13731
143. Li W, Jia MX, Wang JH, Lu JL, Deng J, Tang JX, et al. Association of MMP9-1562C/T and MMP13-77A/G polymorphisms with non-small cell lung cancer in southern chinese population. Biomolecules (2019) 9(3):10056–67. doi: 10.3390/biom9030107
144. Su L, Zhou W, Park S, Wain JC, Lynch TJ, Liu G, et al. Matrix metalloproteinase-1 promoter polymorphism and lung cancer risk. Cancer epidemiology Biomarkers Prev Publ Am Assoc Cancer Research cosponsored by Am Soc Prev Oncol (2005) 14(3):567–70. doi: 10.1158/1055-9965.EPI-04-0482
145. Heist RS, Marshall AL, Liu G, Zhou W, Su L, Neuberg D, et al. Matrix metalloproteinase polymorphisms and survival in stage I non-small cell lung cancer. Clin Cancer Res (2006) 12(18):5448–53. doi: 10.1158/1078-0432.CCR-06-0262
146. Zhu Y, Spitz MR, Lei L, Mills GB, Wu X. A single nucleotide polymorphism in the matrix metalloproteinase-1 promoter enhances lung cancer susceptibility. Cancer Res (2001) 61(21):7825–9.
147. Lei Z, Liu R, Chen J, Zhou Q, Zhang H. Meta Analysis of Association between Polymorphisms in Promoter Region of MMPs gene and Risk of Lung Cancer. Zhongguo Fei Ai Za Zhi (2009) 12(5):381–6. doi: 10.3779/j.issn.1009-3419.2009.05.017
148. Rollin J, Régina S, Vourc'h P, Iochmann S, Bléchet C, Reverdiau P, et al. Influence of MMP-2 and MMP-9 promoter polymorphisms on gene expression and clinical outcome of non-small cell lung cancer. Lung Cancer. (2007) 56(2):273–80. doi: 10.1016/j.lungcan.2006.11.021
149. Butkiewicz D, Krześniak M, Drosik A, Giglok M, Gdowicz-Kłosok A, Kosarewicz A, et al. The VEGFR2, COX-2 and MMP-2 polymorphisms are associated with clinical outcome of patients with inoperable non-small cell lung cancer. Int J cancer. (2015) 137(10):2332–42. doi: 10.1002/ijc.29605
150. Wang Y, Fang S, Wei L, Wang R, Jin X, Wen D, et al. No association between the C-1562T polymorphism in the promoter of matrix metalloproteinase-9 gene and non-small cell lung carcinoma. Lung Cancer (2005) 49(2):155–61. doi: 10.1016/j.lungcan.2005.04.006
151. Ci Y, Zhang Y, Liu Y, Lu S, Cao J, Li H, et al. Myricetin suppresses breast cancer metastasis through down-regulating the activity of matrix metalloproteinase (MMP)-2/9. Phytotherapy Res PTR. (2018) 32(7):1373–81. doi: 10.1002/ptr.6071
152. Knapinska AM, Estrada CA, Fields GB. The roles of matrix metalloproteinases in pancreatic cancer. Prog Mol Biol Trans science. (2017) 148:339–54. doi: 10.1016/bs.pmbts.2017.03.004
153. Ren H, Wang Z, Zhang S, Ma H, Wang Y, Jia L, et al. IL-17A promotes the migration and invasiveness of colorectal cancer cells through NF-κB-mediated MMP expression. Oncol Res (2016) 23(5):249–56. doi: 10.3727/096504016X14562725373716
154. Steven A, Fisher SA, Robinson BW. Immunotherapy for lung cancer. Respirology (Carlton Vic). (2016) 21(5):821–33. doi: 10.1111/resp.12789
Keywords: MMP, lung cancer, tumor immunity, drug resistance, MMP polymorphisms
Citation: Wei C (2023) The multifaceted roles of matrix metalloproteinases in lung cancer. Front. Oncol. 13:1195426. doi: 10.3389/fonc.2023.1195426
Received: 28 March 2023; Accepted: 08 August 2023;
Published: 12 September 2023.
Edited by:
Sridhar Muthusami, Karpagam Academy of Higher Education, IndiaReviewed by:
Elena Gershtein, Russian Cancer Research Center NN Blokhin, RussiaAruljothi Muralidharan, United States Department of Veterans Affairs, United States
Copyright © 2023 Wei. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Cui Wei, d2VpY3VpMjAyMzAzQDE2My5jb20=